efeito do Ácido acÉtico nas enzimas xilose...
TRANSCRIPT

GOVERNO DO ESTADO DE SÃO PAULO SECRETARIA DA CIÊNCIA, TECNOLOGIA E DESENVOLVIMENTO ECONÔMICO FACULDADE DE ENGENHARIA QUÍMICA DE LORENA . DEPARTAMENTO DE BIOTECNOLOGIA
Dissertação de Mestrado
EFEITO DO ÁCIDO ACÉTICO NAS ENZIMAS
XILOSE REDUTASE E XILITOL DESIDROGENASE
DE Candida guilliermondii
Luanne Helena Augusto Lima i..~"""""""~==~~=ca-·.,.='~7zc=""''"•"'"~
em i1 r.»: n~ 202/04
Tranferidc da Biblioteca di:i
LORENA - SP - BRASIL
2002

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA
DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
EFEITO DO ÁCIDO ACÉTICO NAS ENZIMAS
XILOSE REDUTASE E XILITOL DESIDROGENASE
DE Candida guilliermondii
Dissertação de mestrado apresentada como parte das exigências para a obtenção do título de Mestre em Biotecnologia Industrial
Banca examinadora:
Ora. Maria das Graças de Almeida Felipe (presidente) Dr. Michele Vitelo Ora. Adriane Maria Ferreira Milagres
Estudante:
Luanne Helena Augusto Lima
Lorena - SP - Brasil
2002·

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA
DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
EFEITO DO ÁCIDO ACÉTICO NAS ENZIMAS XILOSE REDUTASE E XILITOL DESIDROGENASE
DE Candida guilliermondii
Este exemplar corresponde à versão final da Dissertação de Mestrado aprovada pela banca examinadora.
Dra. Maria dar:l:!!!:e Almeida Felipe
Presidente da Banca Examinadora
Lorena - SP - Brasil
2002

AGRADECIMENTOS
Agradeço primeiramente a Profa. Ora. Maria das Graças de Almeida
Felipe pela amizade e pela confiança, além da paciência e dedicação com que
me orientou durante todo o mestrado.
À mamãe pelo incomensurável apoio em todo e qualquer momento e a
toda a minha família pelo incentivo sempre necessário.
Ao Christian Niel Berlinck pelos novos horizontes, pelos momentos de
escape e pelo apoio logístico na finalização deste trabalho.
Aos amigos do laboratório, principalmente Gilvane, Talita. Ely, Dani, Zéa
e aos "companeros" por compartilhar conhecimento, equipamentos,
micropipetas e por tornar a rotina mais agradável.
Ao Eduardo Gomes Gonçalves pelo incentivo, principalmente nos
momentos decisivos.
Às parceiras de república: Priscila, pela mão estendida na hora certa e
pela catálise de diversas amizades e Karlia, pelas várias gargalhadas e por ter
me mostrado que a FAENQUIL fica muito mais perto do que eu imaginava.
Aos técnicos do DEBIQ, principalmente a Rita, Paulinho, Nica e Zé
Cobrinha pelo auxílio e pela amizade.
Aos meus amigos de Brasília, que mesmo longe fisicamente sempre
estiveram presentes.
Às leveduras que se ofereceram em sacrifício para que este trabalho
pudesse ser realizado.

RESUMO
Efeito do Ácido Acético nas Enzimas Xilose Redutase e Xilitol Desidrogenase de Candida guilliermondii. Luanne Helena Augusto Lima. Dissertação de Mestrado. Programa de Pós-Graduação em Biotecnologia Industrial. Departamento de Biotecnologia, Faculdade de Engenharia Q~::r.:ca de Lorena. Orientadora: Maria das Graças de Almeida Felipe (Departamento de Biotecnologia, FAENQUIL, C.P.116, 12600-970, Lorena, SP, Brasil). Banca Examinadora: Dr. Michele Vitolo e Ora. Adriane Maria Ferreira Milagres. Abril de 2002.
O xilitol é um produto de grande interesse econormco pelas suas características como poder adoçante, anticariogenicidade e metabolismo independente de insulina. Estas características são exploradas na formulação de produtos alimentícios, odontológicos e farmacêuticos, sendo a utilização de xilitol também permitida para pessoas diabéticas, obesas ou com anemia hemolítica. A etapa inicial do metabolismo de xilose é a formação de xilitol a partir da redução de xilose, catalisada pela enzima xilose redutase (EC 1.1.1.21) na presença de cofatores reduzidos NADPH e/ou NADH. A seguir o xilitol é oxidado a xilulose pela enzima xilitol desidrogenase (EC 1.1.1.9), com participação de NADP+ e/ou NAD+. Assim como na via química, os materiais lignocelulósicos constituem a matéria-prima para a obtenção de xilitol pela via biotecnológica. O bagaço de cana de açúcar consiste em um resíduo promissor para este bioprocesso, apesar de ser utilizado para geração de energia nas próprias usinas. Embora, o hidrolisado de bagaço apresente elevado teor de xilose, ele também possui compostos tóxicos resultantes da hidrólise ácida da estrutura lignocelulósica. Dentre estes compostos, o ácido acético destaca-se pelo seu efeito inibitório na fermentação e na produtividade em xilitol. Desta forma. o objetivo deste projeto foi avaliar o efeito deste ácido nas atividades das enzimas intracelulares xilose redutase (XR) e xilitol desidrogenase (XDH) e consequentemente na formação de xilitol. Para a realização deste estudo, empregou-se Candida guil/iermondii cultivada em meio elaborado à base de hidrolisado ácido de bagaço de cana concentrado e tratado, acrescido de ácido acético em concentrações superiores à inibitória (3,0 g/L). Experimentos também foram realizados em meio sintético contendo os mesmos teores de açúcares do hidrolisado. Para o estudo do efeito na atividade das enzimas, acetato de sódio foi utilizado no ensaio enzimático em diferentes concentrações. Observou-se que os valores de atividade específica para XR e XDH obtidas de células cultivadas no hidrolisado foram menores que em meio sintético, evidenciando que o efeito tóxico do ácido acético na expressão das enzimas foi potencializado pelos outros compostos presentes no hidrolisado. Foi observado aumento na atividade específica das enzimas (58% e 60%, para XR e XDH respectivamente), com o aumento da concentração de acetato de sódio no ensaio, sugerindo que o ácido acético presente no meio de cultura pode aumentar a atividade destas enzimas. Não houve aumento de produtividade do xilitol durante as fermentações do meio com ácido acético, porque embora a formação de xilitol tenha sido favorecida pelo aumento da atividade de XR, seu consumo também foi aumentado por causa da atividade de XDH.
iv

ABSTRACT
Effect of the acetic acid on the enzymes xylose reductase and xylitol dehydrogenase of Candida guilliermondii
Xylitol is a produr+ of great economic interest due !:- its characteristics, such as sweetening power, anticariogenicity and insulin-independent metabolism. These qualities have been exploited in the production of foodstuffs, odontological products and pharmaceuticals. Besides, xylitol can be ingested by people with diabetes, obesity and hemolytic anemia. This sweetener is commercialy produced by chemical process, but it can also be obtained biotecnologically. ln the biotecnological process, the initial stage of the xylose metabolism is the formation of xylitol from the reduction of xylose catalyzed by the xylose reductase enzyme (EC 1.1.1.21) when the reduced cofactors NADH are present. Next, xylitol is oxidized to xylulose by the xylitol dehydrogenase enzyme (EC 1.1.1.9), with the participation of NADP+ and/or NAD+. Like the chemical path, the biotechnological process also utilizes lignocellulosics, not only because they have high contents of xylan, which can be hydrolysed to xylose, but also because they are low-cost renewable resources. A good example of lignocellulosic materiais is sugar cane bagasse. ln general bagasse is utilized to produce electrical power for the mill itself. Another good use however, would be the generation of a product of high aggregate value, like xylitol. Although bagasse hydrolysate has a high xylose content, it also has toxic compounds coming from acid hydrolysis of the lignocellulosic structure. One of such compounds is acetic acid, which inhibitis fermentation, reducing the xylitol productivity as a consequence. The aim of this project was to evaluate the effects of this acid on the activities of xylose reductase (XR) and xylitol dehidrogenase (XDH) enzymes and therefore on xylitol formation. ln this study Candida guilliermondii yeast was used for fermentation of concentrated and treated hydrolysate medium with acetic acid concentration considered as inhibitory (higher than 3.0 g/1). Experiments were also performed with syntetic medium containing the sarne amounts of sugar as the hydrolysate. To study the effect of the acid on the enzyme activity, sodium acetate was used at different concentrations. The XR and XDH specific activities in the hydrolysate were lower than in the synthetic medium, which provided the evidence that the toxic effect of the acetic acid on these enzyme was potentiated by the other compounds found in the hydrolysate. lncreases in the XR and XDH specific activities (58% and 60% respectively) were observed together with an increase in sodium acetate in the reaction medium, suggesting that the acetic acid present in the culture medium enhanced the activities of these enzymes. No xylitol productivity increase was observed during the fermentation of medium containing acetic acid, because although the xylitol formation was favored by the enhancement of XR activity, the xylitol consumption also íncreased as a result of the XDH activity.

CONTEÚDO
RESUMO
..... asTRAcr
LISTA DE TABELAS
LISTA DE FIGURAS
iv
V
viii
X
1. INTRODUÇÃO 1
2. REVISÃO BIBLIOGRÁFICA 3
2.1 Xilitol 2.1.1 Propriedades e Aplicações 2.1.2 Tolerância e Toxicidade
3 3 5
2.2 Obtenção de Xilitol 6
2.3 Bioconversão de Xilose em Xilitol 2.3.1 Aspectos Bioquímicos 2.3.2 Fatores que influenciam o metabolismo
8 8
10
2.4 Matéria-Prima 2.4.1 Bagaço de Cana de Açúcar
15 16
2.5 Efeito Tóxico dos Hidrolisados Hemicelulósicos 2.5.1 Efeito do Ácido Acético
17 19
3. OBJETIVOS 22
3.1 Geral 22
3.2 Específicos 22
4. MATERIAL E MÉTODOS 23 4.1 Microrganismo e Preparo do lnóculo 23
4.2 Preparo do Hidrolisado de Bagaço de Cana de Açúcar 23
4.3 Concentração e Tratamento 24
4.4 Meios e Condições de Fermentação 24
4.5 Métodos Analíticos 4.5.1 Viabilidade e Pureza da Cultura 4.5.2 Determinação da Concentração Celular 4.5.3 Determinação de pH 4.5.4 Determinação da Concentração de Açúcares e Ácido Acético 4.5.5 Determinação da Concentração de Furfural e Hidroximetilfuriural 4.5.6 Determinação da Concentração de Fenóis
25 25 26 26 26 26 27
vi

4.6 Rompimento Celular 27
4.7 Determinação de Proteína Solúvel 28
4.8 Medida das Atividades Enzimáticas 28
4.9 Efeito do Acetato de Sódio nas Atividades Enzimáticas 29
4.10 Determinação dos Parâmetros Fermentativos 29
5. RESULTADOS E DISCUSSÃO 31
5.1 Efeito do Ácido Acético na Fermentação em Meio Sintético 31 5.1.1 Efeito do Ácido Acético no Consumo de Açúcares 31 5.1.2 Consumo de Ácido Acético Durante a Fermentação 34 5.1.3 Formação de Células na Presença de Ácido Acético 37 5.1.4 Efeito do Ácido Acético na Formação de Xilitol 38 5.1.5 Efeito do Ácido Acético nas Enzimas Xilose Redutase e Xilitol Desidrogenase 40
5.2 Efeito do Ácido Acético na Fermentação em Hidrolisado 42 5.2.1 Caracterização do Hidrolisado 42 5.2.2 Efeito do Ácido Acético no Consumo de Açúcares 44 5.2.3 Consumo de Ácido Acético Durante a Fermentação 46 5.2.4 Formação de Células na Presença de Ácido Acético 48 5.2.5 Efeito do Ácido Acético na Formação de Xilitol 49 5.2.6 Efeito do Ácido Acético nas Enzimas Xilose Redutase e Xilitol Desidrogenase 50
5.3 Efeito do Acetato de Sódio na Atividade das Enzimas Xilose Redutase e Xilitol Desidrogenase 52
6. CONCLUSÕES 54
7. RECOMENDAÇÕES 55
8. REFERÊNCIAS BIBLIOGRÁFICAS 56
vii

LISTA DE TABELAS
TABELA 1 - Leveduras identificadas como produtoras de xilitol a partir de
xilose 7
TABELA li - Características dos meios de cultura após a adição de ácido
acético 30
TABELA li 1 - Concentrações de arabinose durante cultivo de C. guilliermondii
em meio sintético contendo ácido acético em diferentes
concentrações 32
TABELA IV - Ácido acético consumido durante cultivo de C. guilliermondii em
meio sintético contendo diferentes concentrações de ácido acético 33
TABELA V - Valores de pH durante cultivo de C. guilliermondii em meio
sintético contendo ácido acético em diferentes concentrações 35
TABELA VI - Concentrações celulares de C. guil/iermondii durante as
fermentações em meio sintético contendo ácido acético em diferentes
concentrações 36
TABELA VII - Proporções de células inviáveis durante cultivo de
C. guilliermondii em meio sintético contendo ácido acético em diferentes
concentrações 37
TABELA VIII - Produtividade (Op), rendimento (Yp1s) e eficiência de conversão
(ri) de xilitol após 58 horas de cultivo de C. guilliermondii em meio
sintético contendo ácido acético em diferentes concentrações 39
TABELA IX - Atividades específicas de xilose redutase durante cultivo de
C. guilliermondii em meio sintético contendo ácido acético em diferentes
concentrações 39
TABELA X - Atividades específicas de xilitol desidrogenase durante cultivo de
C. guilliermondii em meio sintético contendo ácido acético em diferentes
concentrações 40
TABELA XI - Características do hidrolisado hemicelulósico de bagaço de cana
de açúcar original, após a concentração e depois do
tratamento 41
TABELA XI 1 - Características dos meios de cultura preparados com hidrolisado
contendo ácido acético em diferentes concentrações .42
viii

TABELA XIII - Concentrações de arabinose durante cultivo de C. guilliermondii
em meio formulado a partir de hidrolisado hemicelulósico de bagaço de
cana, contendo ácido acético em diferentes concentrações 44
TABELA XIV - Ácido acético consumido durante cultivo de C. guilliermondii em
meio formulado él partir de hidrolisado hera .icelulósico de bagaço de cana,
contendo diferentes concentrações de ácido acético .45
TABELA XV - Valores de pH durante cultivo de C. guilliermondii em meio
formulado a partir de hidrolisado hemicelulósico de bagaço de cana,
contendo ácido acético em diferentes concentrações .46
TABELA XVI - Concentrações celulares de C. guilliermondii durante as
fermentações em meio formulado a partir de hidrolisado hemicelulósico de
bagaço de cana, contendo ácido acético em diferentes
concentrações 47
TABELA XVII - Proporções de células inviáveis durante cultivo de
C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico
de bagaço de cana, contendo ácido acético em diferentes
concentrações 48
TABELA XVIII - Produtividade (Qp), rendimento (Ypts) e eficiência de conversão
(11) de xilitol após 58 horas de cultivo de C. guilliermondii em meio
formulado a partir de hidrolisado hemicelulósico de bagaço de cana,
contendo diferentes concentrações de ácido acético .49
TABELA XIX - Atividades específicas de xilose redutase durante cultivo de
C. guil/iermondii em meio formulado a partir de hidrolisado hemicelulósico
de bagaço de cana, contendo diferentes concentrações de ácido
acético 50
TABELA XX - Atividades específicas de xilitol desidrogenase durante cultivo de
C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico
de bagaço de cana. contendo diferentes concentrações de ácido
acético 50
ix

LISTA DE FIGURAS
FIGURA 1 - Esquema do metabolismo de xilose e regeneração de
cofatores 9
FIGURA 2 - Formação de hidroximet.furfural ~LfMF) e furfural a partir da
desidratação de hexoses e pentases respectivamente 18
FIGURA 3 - Mecanismo de acidificação do citoplasma pela forma não
dissociada do ácido acético e transporte de prótons através da membrana
pela enzima H+ -A TPase 20
FIGURA 4 - Concentrações de xilose durante cultivo de C. guilliermondíí em
meio sintético simulando a constituição de hidrolisado hemicelulósico de
bagaço de cana, com diferentes concentrações de ácido acético
(A- 5 g/L; 8- 6 g/L; C - 8 g/L; D - 10 g/L) 31
FIGURA 5 - Concentrações de ácido acético durante cultivo de C. guil/iermondii
em meio sintético simulando a constituição de hidrolisado hemicelulósico de
bagaço de cana, com diferentes concentrações de ácido acético
(A - 5 g/L; B - 6 g/L; C - 8 g/L; D - 1 O g/L) 34
FIGURA 6 - Proposta de metabolismo de acetato pela formação de
Acetil-CoA 36
FIGURA 7 - Concentrações de xilitol durante durante cultivo de C. guilliermondii
em meio sintético simulando a constituição de hidrolisado hemicelulósico de
bagaço de cana, com diferentes concentrações de ácido acético
(A - 5 g/L; B - 6 g/L; C - 8 g/L; D - 1 O g/L) 38
FIGURA 8 - Concentrações de xilose durante cultivo de C. guillíermondii em
meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana,
com diferentes concentrações de ácido acético (A - 3 g/L; B - 4 g/L;
C - 6 g/L; D - 8 g/L) 43
FIGURA 9 - Concentrações de ácido acético durante cultivo de C. guil/iermondii
em meio formulado a partir de hidrolisado hemicelulósico de bagaço de
cana, com diferentes concentrações de ácido acético (A - 3 g/L;
B - 4 g/L; C - 6 g/L; D - 8 g/L) .46
X

FIGURA 1 O - Concentrações de xilitol durante as fermentações em meio com
hidrolisado hemicelulósico de bagaço de cana, com diferentes
concentrações de ácido acético (A - 3 g/L; B - 4 g/L; C - 6 g/L;
D- 8 g/L) 48
FICURA 11 - Atividade específica de xilose redutase por concentração de
acetato de sódio no meio de reação 51
FIGURA 12 - Atividade específica de xilitol desidrogenase por concentração de
acetato de sódio no meio de reação 52
xi

1. INTRODUÇÃO
Xilitol é um poliol com propriedades adoçante e anticariogênica, que
pode ser utilizado em substituição à sacarose por pessoas diabéticas, obesas
ou com anerr= hemolítica. Além diste o xilitol possui diversas aplicações
clínicas, como prevenção de otite média aguda, proteção contra osteoporose e
prevenção de infecção pulmonar. Este já é empregado em vários países,
inclusive no Brasil, como insumo de produtos como pastilhas, creme dental e
gomas de mascar, porém o seu uso é limitado pelo seu elevado custo.
O xilitol é comercialmente produzido a partir da catálise química da
xilose, obtida de resíduos lignocelulósicos ricos em xilana. Este processo
possui a desvantagem de requerer várias etapas de purificação para a
obtenção de uma solução inicial de xilose de elevada pureza, visto que a
presença de impurezas, como resíduos de lignina, interfere na etapa de
redução catalítica. É ainda necessário purificar o produto para remoção de
resíduos tóxicos do catalisador e de subprodutos gerados.
Pesquisas vêm sendo desenvolvidas para se estabelecer um processo
biotecnológico alternativo à produção convencional de xilitol. Pesquisadores do
Grupo de Processos Fermentativos do Departamento de Biotecnologia
(DEBIQ) da Faculdade de Engenharia Química de Lorena (FAENQUIL), vem
há anos trabalhando para obtenção de xilitol a partir de microrganismos
fermentadores da xilose obtida por hidrólise ácida de materiais lignocelulósicos.
Estes materiais constituem recursos renováveis, e são representados pelos
resíduos agroindustriais, como bagaço de cana de açúcar, palha de arroz e
palha de trigo. Neste sentido, espera-se que, além da redução do custo de
produção do xilitol, o processo microbiológico possa contribuir para a
diminuição do impacto ambiental causado por estes resíduos.
Os trabalhos buscam estabelecer condições ideais para este
bioprocesso, como a hidrólise da fração hemicelulósica da biomassa, o
tratamento do hidrolisado, a suplementação nutricional e as condições ótimas
de fermentação para a levedura Candida gui/Jiermondii FTI 20037, uma das
espécies selecionadas mais promissoras para esta bioconversão. Tem sido
também avaliados aspectos quanto à via metabólica envolvida na conversão de
xilose em xilitol, a partir do estudo do comportamento das enzimas

2
intracelulares xilose redutase e xilitol desidrogenase responsáveis pelos passos
iniciais desta via metabólica.
As pesquisas têm evidenciado que apesar do tratamento prévio dos
hidrolisados para a redução da concentração de compostos tóxicos oriundos da
hidrólise .;c;Ja como o ácido acético, este ai, ida permanece no meio em
concentrações consideradas inibitórias à conversão de xilose em xilitol. Dados
obtidos de fermentação em meio sintético revelaram que na presença deste
ácido em concentrações superiores a 3,0 g/L, ocorreu redução na
produtividade do xilitol, enquanto 1,0 g/L deste favoreceu sua formação. No
entanto, na concentração presente nos hidrolisados concentrados e tratados
(- 4 g/L) o seu efeito tóxico é potencializado pela presença de outros
compostos tóxicos à célula como furfural, hidroximetilfurfural e fenóis.
A fim de contribuir para o estabelecimento de uma biotecnologia de
produção de xilitol a partir de bagaço de cana de açúcar, este trabalho teve
como principal objetivo estudar o efeito do ácido acético nas atividades das
enzimas xilose redutase e xilitol desidrogenase de C. guillierrnondii cultivada
tanto em meio sintético contendo xilose como principal fonte de carbono,
quanto em hidrolisado hemicelulósico de bagaço de cana de açúcar.

3
2. REVISÃO BIBLIOGRÁFICA
2.1 XILITOL
2.1.1 PROPRIEDADES E APLICAÇÕES
O xilitol é um poliol edulcorante com fórmula empírica CsH120s, que
ocorre na natureza em diversos vegetais, líquens e algas, aparecendo também
como um intermediário no metabolismo de carboidratos em animais (PEPPER
& OLINGER, 1988; WINKELHAUSEN et ai., 1996).
O poder adoçante do xilitol é comparável ao da sacarose e superior ao
de outros polióis como manitol e sorbitol. Possui um terço a menos de calorias
que a sacarose, ou seja 2,4 calorias por grama (BAR, 1991). Devido à doçura e
ao baixo valor calórico, o xilitol vem sendo utilizado como adoçante desde os
anos 60, o que, além de outras características que serão apresentadas a
seguir, proporciona aumento no seu valor econômico.
O xilitol tem a propriedade cariostática por não ser metabolizado por
microrganismos da microbiota bucal, principalmente Streptococcus mutans, o
que impossibilita a proliferação de bactérias e conseqüentemente impede
produção de ácidos que atacam o esmalte dos dentes. Além disto, também
pode ser classificado como anticariogênico, por promover a estimulação da
produção de saliva que possui capacidade tamponante, que juntamente com o
aumento na concentração de íons cálcio e fosfato, induz a remineralização,
revertendo lesões de cáries recém formadas (BIRKHED, 1994; MAKINEN et
ai., 1998). Estudos demonstraram que o consumo diário de chicletes contendo
xilitol por crianças de 1 O anos reduziu em 53,5% a incidência de cáries
(ALANEN et ai., 2000). Os efeitos cariostático e anticariogênico impulsionam o
uso de xilitol em cremes dentais, pastilhas, gomas de mascar e outros produtos
para controle e prevenção às cáries.
O xilitol é apropriado para o uso em produtos alimentícios processados
em temperaturas elevadas, quando reações de Maillard não são desejadas.
Estas reações podem ocorrer quando proteínas são aquecidas na presença de
açúcares redutores, resultando na glicosilação do aminoácido terminal ou de
resíduos de lisina. O xilitol, devido à ausência de grupos aldeídicos ou

4
cetônicos em sua molécula. não participa destas reações de escurecimento,
sendo esta mais uma característica que o torna potencialmente aplicável na
indústria alimentícia (MANZ et ai., 1973; BAR 1991).
Outra vantagem do xilitol é não ser fermentado por muitos
microrganismos, permitindo d.,;:,,m seu uso em xaropes e refrescos sem a
necessidade de pasteurização e da adição de conservantes, quando o produto
é estocado por 4 ou 5 meses em frascos fechados (MANZ et ai., 1973).
A sensação de vaporização nas cavidades oral e nasal é outra
característica importante do xilitol. Esta sensação é proporcionada pelo valor
negativo do calor de dissolução e é explorada em produtos farmacêuticos
(vitaminas e expectorantes), confeitos (pastilhas e balas), sendo este
usualmente combinado a manitol, sorbitol ou ácido cítrico, nestes produtos
(PARAJÓ et ai., 1998a).
Em alguns países europeus o xilitol já vem sendo utilizado como
adoçante, tendo a vantagem de não apresentar sensação desagradável após o
uso (EMODI, 1978). No Brasil sua utilização se limita à composição de
produtos como creme dental, gomas de mascar e pastilhas.
Clinicamente, o xilitol é indicado para obesos por ser menos calórico e
diminuir o nível de ácidos graxos livres no sangue (MANZ et ai., 1973). Pode
ser empregado no tratamento de desordens metabólicas, como em casos de
anemia hemolítica que ocorre em indivíduos com deficiência na enzima glicose-
6-fosfato desidrogenase (van EYS et ai., 1974). É também indicado para
consumo por diabéticos em substituição a outros. açúcares, uma vez que seu
metabolismo não ocorre por vias insulino-dependentes (PEPPER & OLINGER,
1988).
A utilização de xilitol também em nutrição parenteral vem sendo
amplamente aceita no Japão e em alguns países da Europa. Estudos
mostraram que a administração hipocalórica de xilitol juntamente com
aminoácidos preserva as proteínas do paciente mais eficientemente que a
infusão apenas de aminoácidos ou em combinação com glicose (WAITZBERG,
1995).
Novas descobertas tem sido feitas quanto às aplicações do xilitol,
principalmente em uso clínico. Foi demonstrado que o xilitol pode ser utilizado
para a prevenção de otite média aguda, por inibir o crescimento e a adesão de

5
espécies de Pneumococcus e a adesão de Haemophilus influenzae em células
da nasofaringe. O xilitol foi testado na forma de xarope, de goma de mascar e
de pastilha; diminuindo em 40% a ocorrência de otite em crianças (UHARI et
ai., 1998). Outra aplicação do xilitol é a proteção contra osteoporose, uma vez
que ratos submetidos a ova: iéctemia, em uma dieta suplementada com 10% de
xilitol, apresentaram maior densidade óssea, além de manter a constituição
mineral dos ossos. Estes resultados protegem contra a perda das propriedades .
biomecânicas dos ossos, causada pela osteopenia (redução da calcificação ou
da densidade óssea) devido à falta de estrógenos, situação similar a que
acontece na menopausa em mulheres (MA TIILA et ai., 1998). Além disto, o
xilitol em aerossol pode também prevenir infecções pulmonares, inclusive em
pacientes com fibrose cística. Ele reduz a concentração salina do líquido que
cobre as células do revestimento interno dos pulmões, aumentando a atividade
antibiótica corpórea natural contra as bactérias (ZABNER et ai., 2000).
2.1.2 TOLERÂNCIA E TOXICIDADE
Em organismos superiores o metabolismo do xilitol ocorre principalmente
no fígado, onde pode ser transformado em glicose a uma taxa entre 20 e 80%,
dependendo da necessidade. Sua absorção é lenta e portanto também pode
ser metabolizado indiretamente pela biata intestinal. Além disto é bem tolerado
pelo corpo humano em uma dieta contendo acima de 200g por dia (ALAIS &
LINDEN, 1991), e em alguns casos pode apresentar efeito laxativo devido ao
desbalanço osmótico causado no intestino grosso.
Quanto à segurança do consumo e utilização por humanos, o xilitol já é
aceito pela European Economic Community - EEC desde 1984, enquanto a
U.S. Food and Drug Administration - FDA o classifica como "geralmente
reconhecido como seguro" ( General/y Recognise as Safe - GRAS) desde 1986
e "seguro para os dentes" (Safe for Teeth) desde 1994. A Joint Expert Comittee
on Food Additives (JECFA), uma divisão prestigiada da Word Health
Organization confirmou os estudos de toxicidade de xilitol e o classificou como
aceitável para consumo diário (Acceptable Daily lntake - ADI). Atualmente, o
xilitol já é utilizado em produtos alimentícios, farmacêuticos ou de saúde oral
em mais de 35 países (CCC - Calorie Control Council, 2001 ). No Brasil, o xilitol

6
foi aprovado como produto dietético pelo Ministério da Saúde em 1980
(AGUIAR et ai., 1999).
2.2 OBTENÇÃO DE XILITOL
O xilitol pode ser recuperado de fontes naturais como vegetais, fungos e
líquens por extração sólido-líquido, porém como ele está presente em pequena
proporção - menos de 900mg/1 OOg, este processo se torna inviável
economicamente (PARAJÓ et ai., 1998a). Sua obtenção convencional ocorre
por via química, porém a via microbiológica vem se destacando nos últimos
anos.
A produção de xilitol em larga escala ocorre pelo processo químico, que
consiste na redução de xilose, derivada principalmente de hidrolisados de
materiais lignocelulósicos ricos em xilana. O processo convencional inclui
4 etapas básicas: hidrólise ácida do material vegetal, purificação do hidrolisado
até a obtenção de xilose pura, hidrogenação catalítica da xilose em xilitol e
finalmente cristalização do xilitol. Este processo foi patenteado em 1977 por
MELAJA & HAMALAINEN.
O rendimento do processo químico, bem como a qualidade do xilitol são
dependentes da pureza da solução inicial de xilose, uma vez que a presença
de impurezas interfere na redução catalítica. São necessárias operações de
troca-iônica, descoloração e fracionamento cromatográfico para obtenção de
uma solução de xilose de elevada pureza. Além disto, após a remoção do
catalisador por filtração e troca-iônica, a solução de xilitol é concentrada,
fracionada por cromatografia com o emprego de resinas catiônicas e é
cristalizada para obtenção do produto puro (AGUIAR et ai., 1999). Estas
diversas etapas de purificação para remoção de resíduos resultam no aumento
de tempo de processamento e encarecimento do produto (PARAJÓ et ai.,
1998a).
O xilitol também pode ser obtido microbiologicamente a partir da
fermentação de hidrolisados ricos em xilose obtido de materiais
lignocelulósicos, sem a necessidade de purificação prévia da xilose (ONISHI &
SUZUKI, 1971; SILVA et ai., 1996; FELIPE et ai., 1997b; PARAJÓ et ai.,
1998a, b e c). Vários microrganismos já foram identificados como

7
fermentadores de xilose em xilitol. Dentre os fungos filamentosos pode-se citar
espécies dos gêneros Penicil/ium, Aspergi/Jus, Rhizoous, Glicocladium.
Byssoch/amys, Myrothecium, Neurospora, Rhodotorula, Torulopsis (CHIANG &
KNIGHT, 1960); Mucor e Fusarium (PARAJO et ai., 1998a), além de
Peaomyces a/bertensis identificado por DAH({A (1991 ). Poucas bactérias,
como Corynebacterium sp., Enterobacter liquefaciens e Mycobacterium
smegmatis formam xilitol, geralmente as bactérias convertem xilose
diretamente em xilulose sem os passos de oxirredução (WINKELHAUSEN &
KUZMANOVA, 1998).
Quanto às leveduras, várias espécies foram identificadas como
produtoras de xilitol, destacando-se as do gênero Candida pela maior eficiência
de conversão (Tabela 1).
TABELA 1 - Leveduras identificadas como produtoras de xilitol a partir de xilose, adaptado de WINKELHAUSEN & KUZMANOVA (1998).
Levedura Xilitol (g/L) Candida boídinii NRRL Y-17213 C. guillíermondií FTl-20037 C. íntermedia RJ-248 e. mogii ATCC 18364 C. parapsilosis ATCC 34078 C. pseudotropicalís IZ-431 e. tropical is e. tropicalis HXP 2 C. tropicalis 1004 C. tropicalis ATCC 7349 e. tropicalis ATCC 20240 e. utilis ATCC 22023 C. utilis C-40 Debaryomyces hansenii C-98 M-21 Hansenula anomala IZ-1420 Kluyveromyces fragilís FTl-20066 K. marxianus IZ-1821 Píchía (Hansenu/a) anomala NRRL Y-366 Pachysolen tannophilus NRRL Y-2460 Saccharomyces SC-13 Saccharomyces SC-37 Schízosaccharomyces pombe 16979
2,9 37,0 5,7
31,0 20,0 4,3 2,1 4,8 17,0 20,0 5,5 1,8 3,0 0,8 6, 1 4,6 6, 1 2,0 2,2 0,7 2,3 0,2

8
2.3 BIOCONVERSÃO DE XILOSE EM XILITOL
2.3.1 ASPECTOS BIOQUÍMICOS
Em microrganismos, o xilitol é um intermediário do metabolismo da
xilose. porém na maioria das bactérias e em algumas leveduras que assimilam
xílose, ela é convertida diretamente a xilulose pela enzima xilose isomerase
(EC 5.3.1.5) sem a formação deste poliol (YOKOYAMA et ai., 1995).
A maioria das leveduras metabolizam xilose pela redução em xilitol
catalisada pela enzima xilose redutase (EC 1.1.1.21) na presença de cofatores
reduzidos NADPH e/ou NADH. O xilitol formado pode ser excretado para o
meio extracelular ou oxidado a xilulose, sendo esta reação catalisada pela
enzima xilitol desidrogenase (EC 1.1.1.9) dependente de cofatores oxidados
NADP+ e/ou NAD+ (JEFFRIES, 1983; SLININGER et ai., 1987). A xilulose é
então fosforilada no carbono 5, pela xilulose quinase e metabolizada pela via
pentase fosfato. A xilulose também pode sofrer isomerização a xilose e reiniciar
a seqüência de reações (DAHIYA, 1991). A Figura 1 apresenta o esquema do
metabolismo inicial de xilose em leveduras.
As enzimas xilose redutase e xilitol desidrogenase podem ter diferentes
especificidades, dependendo da levedura estudada. Em Candida utilis, a
enzima xilose redutase requer como cofator NADPH, enquanto xilitol
desidrogenase depende de NAD+ (BRUINENBERG et ai., 1984). Já em
C. guilliermondii FTI 20037 observa-se que a enzima xilose redutase é NADPH
dependente e xilitol desidrogenase é NAD+ ou NADP+ dependente (SILVA et
ai., 1996). Nas leveduras Pichia stipitis, Candida shehatae e Pachysolen
tannophilus a enzima xilose redutase utiliza tanto NADPH como NADH, e a
xilitol desidrogenase NADP+ ou NAD+ (HAHN-HAGERDAL et ai., 1994).
VERDUYN et ai. (1985a e b) demonstraram que P. stipitis possui somente uma
xilose redutase, enquanto P. tannophilus possui duas formas de xilose
redutase, com diferentes especificidades por cofatores. YOKOYAMA et ai.
(1995) sugerem que microrganismos que apresentam a enzima xilose redutase
dependente de NADH são melhores produtores de etanol e ao contrário,
aqueles que apresentam xilose redutase dependente de NADPH, acumulam
xilitol ao invés de produzir etanol. Em C. intermedia foram isoladas duas formas

9
de xilose redutase, uma com especificidade apenas por NADPH e outra que
pode usar NADH ou NADPH como cofator, porém a razão do cofator disponível
no citossol deve determinar qual das duas formas será utilizada para conversão
de xilose em xilitol (MAYR et ai., 2000).
L D·Xll .. OSE _J Red::: e :p~2:) ViáPentÔ$e(FÔ~fâtô
Xilose lsomerase
FIGURA 1 - Esquema do metabolismo de xilose e regeneração de cofatores, adaptado de BARBOSA et ai. (1988) e DAHIYA (1991).
--~"= te ::.:x:: º ~d~'~pj- l_º x_.L_U_L_O_S_E~J
Xilulose lCATP Quinase ADP
j XILULOSE·SP I L..___ -~
LEE (1998) comparando as seqüências de aminoácidos de xilose
redutase de diferentes leveduras observou que os resíduos (Asp43, Tyr48,
Lys77, His110, Lys262) que participam diretamente da catálise ou da ligação
do cofator são conservados em todas elas.
Segundo BRUINENBERG et ai. (1984), a produção de xilitol por
leveduras está intimamente relacionada com a regeneração de cofatores
reduzidos. Pela via pentase fosfato, 1 mol de glicose-6-fosfato é
completamente oxidado a H20 e C02, gerando 12 moles de NADPH a partir de
NADP+. A fim de manter o balanço de cofatores, o NADPH produzido é usado
para reduzir xilose em xilitol, sendo assim o xilitol é catabolizado para produzir
glicose-6-fosfato e regenerar NADP+ suficiente para manter o ciclo
(BARBOSA et ai., 1988).
íl \J Via Pentase Fosfato

10
SILVA et ai. (1996) explicam que em C. guilliermondii FTI 20037, o
acúmulo de xilitol em relação a xilulose ocorre devido às constantes cinéticas
das enzimas, uma vez que o KM da xilose redutase (O, 18 M) é 4 vezes menor
do que o da xilitol desidrogenase (0,75 M), enquanto a Vrnax para xilose
redutase (243 ü/,nl) é maior do que a Vmax da xilitol desidroqenase (168 U/ml).
GIRIO et ai. (1996) sugerem que os valores de KM da enzima xilitol
desidrogenase para o substrato e o cofator podem mostrar qual levedura
acumularia maior concentração de xilitol. Comparada ao de outras leveduras, a
xilitol desidrogenase de D. hansenii apresenta KM na mesma ordem de
magnitude para xilitol e de 2 a 5 vezes maior para NAD+. Assim sendo,
diminuindo a concentração do cofator, a oxidação de xilítol cessaria, havendo
acúmulo deste intermediário no meio.
2.3.2 FATORES QUE INFLUENCIAM O METABOLISMO
A eficiência de conversão de xilose em xilitol, assim como a
produtividade, dependem do microrganismo e das condições empregadas na
fermentação. A produção de xilitol está relacionada principalmente às
atividades das enzimas xilose redutase e xilitol desidrogenase e, por
conseguinte, a indução ou repressão destas enzimas influencia diretamente o
processo.
• Efeito da Presença de Outros Glicídeos
A composição de hidrolisados hemicelulósicos utilizados como meio de
cultura para a produção biotecnológica de xilitol varia de acordo com o tipo de
material lignocelulósico utilizado, porém estão sempre presentes nos
hidrolisados outros açúcares além da xilose, normalmente glicose, arabinose,
galactose e manose, em menores concentrações. WALTHER et ai. (2001),
utilizando Candida tropicalis, observaram que glicose (20 g/L) em meio
contendo 60 g/L de xilose reprimiu fortemente (41 %) a formação de xilitol,
enquanto manose (20 g/L), e galactose (20 g/L) causaram moderada inibição
na produção de xilitol (11 % e 22% respectivamente). Além disto, xilose,

11
manose e galactose foram simultaneamente consumidas após o esgotamento
de glicose, enquanto não se verificou consumo de arabinose.
A presença de hexases, como a glicose, no meio de fermentação é um
dos fatores que regula a produção de xilitol por leveduras. Em fermentações
com C. shehatae e P. stipitis, en, meio de cultura contendo xilose e glicose, foi
verificado o consumo preferencial de glicose em relação a xilose, havendo um
período lag para o consumo desta pentase, necessário para a síntese de
enzimas do metabolismo de xilose (KILIAN & van UDEN, 1988). Contudo, este
período pode ser reduzido com o crescimento do inóculo em meio contendo
xilose como fonte de carbono (SILVA et ai., 1998).
Em C. tropicalis, no entanto, foi observado que quando o meio é
suplementado com glicose, a produção de xilitol é melhorada, inclusive com
menor consumo de xilose. Este efeito, à primeira vista contraditório, foi
explicado pelo uso de glicose como fonte de carbono preferencial pela
levedura, iniciando seu crescimento no meio de cultura mais rapidamente do
que na presença de xilose (YAHASHI et ai., 1996) Além disto a glicose permite
a regeneração de NADPH, cofator da enzima xilose redutase, pela via pentase
fosfato (PARAJÓ et ai., 1998b). ROSA et ai. (1998) constataram o
favorecimento da formação de xilitol por C. · guilliermondii, em meio sintético
contendo 30 g/L de xilose e 5,0 g/L glicose. TAVARES et ai. (2000) também
observaram com D. hansenii ,que a adição de 1,0% de glicose em meio
contendo 60 g/L xilose, a produtividade de xilitol aumentou 30%. O mesmo
efeito não foi observado com a adição de galactose.
Segundo LUCAS & van UDEN (1986), o transporte de moléculas para o
interior da célula interfere na eficiência e na produtividade do xilitol. Estes
autores estudaram o transporte de monômeros de hemicelulose em
C. shehatae e constataram que tanto a glicose como a xilose são transportadas
por sistema de difusão facilitada e também por simporte de prótons. Quando a
levedura cresce sob condições de supressão de fontes de carbono ela produz
um sistema de difusão facilitada que reconhece glicose, D-manose e D-xilose e
não reconhece D-galactose e L-arabinose. Os açúcares glicose e xilose são
mutuamente inibidores competitivos, indicando que as duas atividades nos
sistemas de afinidade correspondem a dois simportes distintos com KS e
Vmax próprios (KS ± 2 mM, Vmax ± 2,3 mmoles/g.h para glicose e

12
KS ± 125 mM, Vmax ± 22,5 mmoles/g.h para xilose). Para P. stipitis, também
foi observado dois simportes distintos para o transporte de glicose e xilose,
porém com comportamento de inibição não competitiva entre eles (KILIAN &
van UDEN, 1988). LEE et ai. (1996) venncaram que em C. guiffiermondii, D-xilose e
L-arabinose são os melhores glicídeos indutores, enquanto glicose é um
repressor catabólico das enzimas xilose redutase e xilitol desidrogenase.
KERN et ai. (1997) também observaram, para xilose redutase de C. tenuis, o
mesmo comportamento de indução por D-xilose e L-arabinose, além de
D-lixose e D-arabinose. Então, pela análise das moléculas indutoras, foi
sugerido que o efeito pode estar relacionado com a posição equatorial dos
grupos hidroxila nos carbonos da posição 3 e 4.
• Concentração Inicial e Idade do lnóculo
CAO et ai. (1994) observaram, em uma linhagem de Candida sp., que
quando a concentração do inóculo foi aumentada de 3,8 a 26 g/L, houve um
aumento de 80% na produção de xilitol em meio com 260 g/L de xilose. Em
hidrolisado de bagaço de cana de açúcar e com C. guilliermondii FTI 20037, a
bioconversão de xilose em xilitol foi favorecida utilizando inóculo de O, 1 a 3,0
g/L, sendo seu aumento limitado pela concentração de substrato e aeração do
meio (FELIPE et ai., 1997a).
PFEIRER et ai. (1996) observaram durante fermentações em meio
sintético, que a utilização de inóculos de C. guil/iermondii de 16 a 24 horas
resultou em maior produtividade do xilitol, dados corroborados por FELIPE et
ai. (1997a). Estes autores constataram um aumento de 36% neste parâmetro
com emprego de inóculo de 24 horas em relação ao de 48 horas durante
fermentações em hidrolisado de bagaço de cana.
• Temperatura e pH
A temperatura é um fator importante pois interfere tanto no crescimento
da levedura, como na atividade das enzimas envolvidas na via microbiológica
de obtenção de xilitol. Pesquisas com C. guilliermondii demonstraram que a

13
máxima concentração de xilitol acumulado ocorreu tanto em 30 ºC, como em
35 ºC. Nestas temperaturas a taxa de crescimento foi alta e a xilose foi
consumida rapidamente (BARBOSA et ai., 1988). SOUZA et ai. (1993)
verificaram, durante o cultivo de C. gui/liermondii em hidrolisado hemicelulósico
de palha de arroz, em i:ernperaturas acima de 40 ºC, um decréscimo da
produção de xilitol, enquanto que abaixo de 20 ºC não foi observada formação
deste palio!. Em hidrolisado hemicelulósico de bagaço, o maior rendimento de
xilitol e as atividades máximas para as enzimas xilose redutase e xilitol
desidrogenase de C. guifliennondii foram encontrados com a temperatura de
fermentação igual a 35 ºC (SENE et ai., 2000a).
O pH é outro parâmetro que interfere no crescimento celular e nas
atividades enzimáticas, sendo este efeito dependente do tipo de meio de
cultura. FELIPE et ai. (1997b) constataram que em fermentações de hidrolisado
hemicelulósico de bagaço de cana, a produção de xilitol por C. guilliermondii
foi fortemente inibida em pH abaixo de 4,5, o mesmo não foi observado por
SILVA et ai. (1994) em meio sintético. MORITA et ai. (2000a), também
utilizando hidrolisado de bagaço, encontraram melhores resultados para
consumo de xilose, produtividade e rendimento de xilitol em pH 7,0.
SENE et ai. (2000a) encontraram as máximas atividades de xilose redutase e
xilitol desidrogenase de C. guifliermondii foram em pH 5,5 e 8,5
respectivamente. Em hidrolisado de bagaço de cana, a atividade máxima para
xilose redutase ocorreu quando o pH do meio de cultura foi igual a 6,0,
condição na qual foram também encontrados os máximos valores dos
parâmetros fermentativos da produção de xilitol.
WEBB & LEE (1990) relacionaram o efeito do pH nas atividades
enzimáticas aos resíduos de histidina e cisteína, que podem estar envolvidos
na ligação do cofator à enzima xilose redutase de P. stipitis. A histidina é um
resíduo de aminoácido que é frequentemente encontrado em sítios
enzimáticos, seu anel imidazólico varia entre os estados sem carga e com
carga positiva para catalisar a formação e ruptura de ligações. Estes estados
são influenciados pelo pH do meio e interferem na atividade enzimática.

14
• Disponibilidade de Oxigênio
Para a formação de xilitol, outro fator crítico é a disponibilidade de
oxigênio durante a fermentação. Em condições anaeróbicas a xilose não pode
ser assimilada pelo fato de que o cofator reduzido NADH não pode ser
regenerado através da fosforilação oxidativa (NOLLEAU et ai., 1993), além do
que a atividade de transhidrogenase não foi detectada em leveduras e
aparentemente o balanço não pode ser restabelecido por esta via (HAHN-
HAGERDAL et ai., 1994). VANDESKA et ai. (1996) concluíram que a taxa de
consumo de oxigênio determina quando a xilose é utilizada em processos
respiratórios ou fermentativos.
Segundo GIRIO et ai. (1990), condições de forte aeração favorecem o
aumento de biomassa e prejudicam a produção de xilitol. Na cadeia
transportadora de elétrons, o oxigênio oxida o NADH em NAD+ e a alta
proporção de NAD+ /NADH favorece a oxidação de xilitol em xilulose. A xilulose
seria então degradada através da via da pentase fosfato e pela via glicolítica
para formar piruvato, que é o substrato do ciclo dos ácidos tricarboxílicos,
resultando no aumento de massa celular (FURLAN et ai. 1991). Para favorecer
a bioconversão de xilose em xilitol o meio deve ser mantido sob condições de
baixa aeração, pois ocorre uma elevação da concentração de NADH,
paralisando as etapas posteriores do catabolismo do xilitol e consequente
acúmulo do mesmo (NOLLEAU et ai., 1993; HAHN-HAGERDAL et ai., 1994).
BRUINENBERG et ai. (1984) demonstraram que a taxa de fermentação
anaeróbica de xilose por P. tannophilus e P. stipitis está relacionada à atividade
de xilose redutase NADH dependente, que é a chave para a fermentação
aeróbica em leveduras porque depende da cadeia respiratória para repor o
cofator reduzido. PREEZ et ai. (1989) sugerem que o grau de aeração pode
modular a razão entre xilose redutase NADH dependente e xilose redutase
NADPH dependente em P. tannophi/us. Foi observado para Candida boidinii,
que as atividades de xilose redutase NADH dependente e xilitol desidrogenase
NAD+ dependente foram diminuídas com o aumento da disponibilidade de
oxigênio (VANDESKA et ai., 1995). O mesmo foi observado para
C. guilliermondii, porém esta só possui uma forma da enzima xilose redutase,
que é NADPH dependente (SENE, 2000b).

15
• Concentração Inicial de Xilose
O efeito da concentração inicial de xilose na produção de xilitol foi
estudado em diferentes microrganismos, demonstrando que a produção é
favorecida com o aumento da concentração de xilose inicial (PARAJÓ et ai..
1998b). O aumento na produção é observado até o ponto em que a
concentração de xilose prejudica o crescimento celular. A queda no rendimento
pode estar associada à alta pressão osmótica no meio de cultura ou a inibição
das enzimas pelo substrato (DAHIYA, 1991; DOMINGUEZ et ai., 1997;
PARAJÓ et ai., 1998b).
SILVA et ai. (1996) verificaram que a levedura C. guilliermondii cultivada
em meio sintético apresentou maior produtividade de xilitol com a concentração
de xilose inicial de 70 g/L e maior rendimento com concentração igual a 90 g/L,
porém seus resultados demonstraram que a melhor produção de xilitol e as
maiores atividades de xilose redutase e xilitol desidrogenase ocorrem na faixa
de 70 a 170 g/L de xilose inicial. Segundo ROSA et ai. (1998), os maiores
valores de atividades para xilose redutase e xilitol desidrogenase de
C. guilliermondii cultivada em meio sintético foram obtidos em meio contendo
15 g/L de xilose, entretanto, quando esta concentração foi aumentada até
609/L observou-se uma redução em cerca de 50% nas atividades enzimáticas.
Em pesquisas com C. tropicalis, WAL THER et ai. (2001) observaram
que a xilose na concentração de 156,5 g/L não causou estresse osmótico sob
baixa taxa de aeração (condições semi-aeróbicas), contudo, com o aumento da
aeração a produtividade de xilitol foi prejudicada em 50% quando a
concentração de xilose foi de 120 g/L.
2.4 MATÉRIA-PRIMA
Um dos objetivos do desenvolvimento de tecnologia para produção
biotecnológica de xilitol é apresentar uma alternativa mais barata possível à via
química, forma atualmente empregada para obtenção deste adoçante, a fim de
possibilitar a produção em larga escala com custos mais baixos. Seguindo este
objetivo, busca-se como fonte de xilose, materiais lignocelulósicos por serem
uma fonte abundante e de grande potencial em recursos renováveis.

16
Lignocelulose é o material orgânico mais abundante na biosfera,
compondo cerca de 50% da biomassa no mundo. Sua produção anual tem
sido estimada em 10 a 50x109 toneladas (KUHAD, 1993). A lignocelulose é
composta de 20 a 35% de celulose, 20 a 39% de hemicelulose e 20 a 30% de
, lignina, considerando 0 cceo seco . de diferentes í-i .ateria is. Os componentes
hemicelulósicos das paredes de células vegetais incluem uma variedade de
polissacarídeos lineares ou ramificados compostos de açúcares tais como
D-xilose, D-galactose, D-manose, D-glicose e L-arabinose (FERREIRA FILHO,
1998). J
Em madeiras e em diversos resíduos agroindustriais a porção
hemicelulósica contém alto teor de xilanas, que são polímeros compostos
principalmente por unidades xilanopiranosídicas. Diferentes métodos de
hidrólise são utilizados para romper a ligação entre as unidades monoméricas e
consequente liberação de sacarídeos, entre eles, principalmente a xilose. Entre
os métodos empregados podem ser citados: extração com soluções alcalinas
(TOIT et ai., 1984), "steam explosion" (MARCHAL et ai., 1986), hidrólise
enzimática (TSAO, 1986) e hidrólise ácida (PESSOA JR., 1997).
Entre os diversos resíduos agroindustriais, pesquisas confirmam que a
xilose presente nos hidrolisados hemicelulósicos de palha de arroz
(ROBERTO et ai., 1994), eucalipto (FELIPE et ai., 1996), bagaço de cana de
açúcar (FELIPE et et., 1997a e b) e palha de trigo (CANILHA, 2002) pode ser
convertida em xilitol por C. guilliennondii FTI 20037. Dentre estes materiais, o
bagaço de cana de açúcar tem se destacado pela sua disponibilidade,
principalmente no estado de São Paulo, o que o torna alvo de inúmeras
pesquisas para o seu aproveitamento no processo microbiológico de obtenção
de xilitol.
'-. ,__
2.4.1 BAGAÇO DE CANA DE AÇÚCAR
O bagaço de cana de açúcar é um resíduo gerado pela indústria
sucroalcooleira após o esmagamento da cana de açúcar para extração do suco
contendo sacarose. Este resíduo é normalmente utilizado pelas próprias
indústrias como combustível para as caldeiras, no entanto este pode ser

17
empregado na geração de produtos de alto valor agregado, como no caso do
xilitol. O bagaço de cana de açúcar possui aproximadamente 50% de celulose,
25% de hemicelulose, 20% de lignina e 3% de cinzas. Comparado a outros
resíduos lignoceluló5;c;:;:;, como palha de arroz, casca de a, .oz e palha de trigo,
entre outros, o bagaço de cana é o que apresenta maior proporção de xilose
(23,3%) em relação aos carboidratos totais (LEE, 1997). Além disto pode ser
considerado uma reserva abundante de energia solar pela sua alta
produtividade (cerca de 80 toneladas/hectare) e capacidade de regeneração
anual (PANDEY et ai., 2000). Estas características têm impulsionado pesquisas
em tecnologias de aproveitamento do bagaço de cana, principalmente em
processos biotecnológicos utilizando microrganismos fermentadores de xilose.
2.5 EFEITO TÓXICO DOS HIDROLISADOS HEMICELULÓSICOS
Para que os hidrolisados hemicelulósicos sejam utilizados como meios
de cultivo para obtenção de xilitol, alguns problemas devem ser contornados,
como a presença de compostos tóxicos inerentes ao processo de hidrólise da
biomassa. Diversos destes compostos já foram identificados, tais como
minerais e metais contidos no material lignocelulósico ou resultantes do
processo de corrosão de equipamentos; produtos da hidrólise de hemicelulose
como hidroximetilfurfural,. furfural e ácido acético; produtos derivados da
degradação de lignina como compostos fenólicos, ácidos aromáticos e
aldeídos; e ainda compostos derivados de extrativos (PARAJÓ et ai., 1998c).
Para minimizar os efeitos destes compostos, diversas técnicas já foram
empregadas, como adaptação de células das leveduras, adição de substâncias
redutoras, neutralização e "overliming", evaporação e "steam stripping",
extração por solvente e adsorção por carvão ativo (CONVERTI et ai., 1999).
O Furfural e o hidroximetilfurfural (HMF) são compostos quimicamente
relacionados, ambos possuem um anel furano e um grupo aldeídico e são
, formados a partir da quebra de carboidratos (Figura 2). Foi demonstrado que
tanto o furfural, como o HMF, exercem efeito inibitório no crescimento de
C. guilliermondii; furfural em concentrações maiores que 1,0 g/L, e HMF em
concentrações acima de 1,5 g/L (SANCHES & BATISTA, 1988).

18
o o Hexases
Desidratação
HMF
'( ? o. .. ._//·,r/-..... .. :;'.'·º
o Pentases
o ,..o . ....__ ... li \_( Desidratação
Furfural
FIGURA 2 - Formação de hidroximetilfurfural (HMF) e furfural a partir da desidratação de hexases e pentases, respectivamente (adaptado de TAHERZADEH et ai., 2000).
O consumo de furfural é mais rápido em relação ao HMF. Quando os
dois estão presentes no meio, HMF é convertido lentamente até que todo o
furfural seja metabolizado. Esta característica é importante porque a presença
de HMF é prolongada durante o processo. O metabolismo destes compostos
pode ocorrer pela enzima piruvato descarboxilase, que catalisa a formação do
grupo acilo, como na reação entre piruvato e acetaldeído. Assim HMF e furfural
competem com a reação com acetaldeído, interferindo na via glicolítica e
prejudicando o crescimento do microrganismo. Além disto, foi demonstrado que
a redução de HMF consome NADH, sendo necessária uma adicional formação
deste cofator para manter o balanço na célula (TAHERZADEH et ai., 2000).
O efeito inibitório dos compostos fenólicos é devido à capacidade de se
associarem a membranas biológicas, causando perda de integridade, afetando
assim sua habilidade de servir como barreira seletiva e matriz de enzimas
transmembrânicas. Os compostos fenólicos comumente encontrados em
hidrolisados hemicelulósicos são ácido 4-hidroxibenzóico, vanilina e catecol
(PALMQVIST et ai., 2000).
O efeito de metais foi pesquisado por WATSON et ai. (1984), que
observaram, para P. tannophilus, que o crescimento em meio sintético não foi
alterado na presença de íons como cobre e cromo em baixas concentrações,
inferiores a 0,05 g/L, enquanto na presença de O, 1 g/L de níquel, observou-se
redução rio crescimento.
O efeito dos compostos tóxicos sobre o metabolismo de xilose é
bastante complexo, havendo algumas evidências de que a inibição da
fermentação é causada pelo efeito sinergístico entre eles

19
(LOHMEIER-VOGEL et ai., 1998). Dentre os compostos presentes no
hidrolisado hemicelulósico de bagaço de cana de açúcar considerados como
potencialmente tóxicos à C. guilliermondii, destaca-se o ácido acético,
encontrado em concentrações superiores a 3,0 g/L (RODRIGUES, 1999).
2.5.1 EFEITO DO ÁCIDO ACÉTICO
O ácido acético, proveniente dos grupos acetil das cadeias laterais da
hemicelulose, é considerado o principal inibidor do metabolismo de xilose
(ZYL et ai., 1988; FERRARI et ai. 1992; CONVERTI et ai., 2000). Sua
toxicidade está relacionada com o pH do meio de cultura (NODA et ai., 1982;
FERRARI et ai., 1992), a disponibilidade de oxigênio (ZYL et ai. 1988), a
temperatura (PINTO et ai., 1989), a relação xilose/ácido acético (PREEZ et ai.,
1991 ), a concentração do ácido (FELIPE et el., 1995) e o microrganismo
empregado (FELIPE et ai., 1997b).
O efeito tóxico do ácido acético é aumentado em pH ácido, pois nestas
condições predomina a sua forma não dissociada no meio de cultura (PREEZ,
1991 ). Isto pode explicar a inibição do crescimento de C. guilliermondii em
hidrolisado com pH inferior a 4,5, onde o ácido acético, que possui pKa igual a
4,74 está, em sua maioria, não dissociado (NODA et ai., 1982). Quando este
ácido é transportado para dentro da célula, a molécula se dissocia em íon
acetato mais próton, devido ao pH do citossol ser próximo a neutralidade. Com
o aumento da concentração de prótons, o pH interno diminui. Para a
manutenção homeostática do pH, as células dependem da enzima
transmenbrânica H+ -ATPase que transporta íons H+ para fora das células
consumindo ATP (HOLYOAK et ai., 1996). Como resultado da restauração da
neutralidade, ocorre o desacoplamento da produção de energia e do transporte
de nutrientes, resultando na diminuição do crescimento e do metabolismo do
microrganismo (HERRERO et ai., 1985; lYL et ai. 1988; LOHMEIER-VOGEL
et ai., 1998). HERRERO et ai. (1985) atribuíram o decréscimo no
desenvolvimento celular de C/ostridium thermocellum na presença de ácido
acético ao fato de grande quantidade de ATP ser consumido via H+ -ATPase, a
fim de manter o fluxo de prótons para restaurar o pH intracelular, resultando em
menor energia para a biossíntese. Segundo NARENDRANATH et ai. (2001),

20
estima-se que o consumo de ATP pela atividade de A TPase é de 10-15%
durante o crescimento celular, tendo uma relação estequiométrica de uma
molécula de ATP hidrolisada por próton expulso da célula.
O processo de acidificação do citossol pela entrada de ácido acético
pode também mcdificar o controle da via glic~lítica (P/\MPULHJ\ & LOUREIRO-
DIAS, 1990). A Figura 3 apresenta o mecanismo de acidificação do citoplasma
pela dissociação da molécula de ácido acético e o transporte de prótons para o
meio extracelular via enzima H+ -ATPase.
Meio Extracelular Meia Intracelular
FIGURA 3 - Mecanismo de acidificação do citoplasma pela forma não dissociada do ácido acético e transporte de prótons através da membrana pela enzima H+-ATPase (adaptado de LAWFORD & ROUSSEAU, 1993).
FELIPE et ai. (1995) demonstraram que o efeito inibitório do ácido
acético no metabolismo de xilose por C. guil/iermondii é dependente da
concentração deste no meio de cultura. Em meios com baixa concentração do
ácido, até 1,0 g/L, houve aumento de 20% no rendimento e de 33% na
produtividade de xilitol em relação ao meio ausente de ácido. Foi demonstrado
que esta levedura foi capaz de metabolizar ácido acético em meio sintético e,
segundo os autores, este efeito pode ser devido à entrada deste ácido
diretamente no ciclo dos ácidos tricarboxílicos via acetil-CoA. A assimilação de
ácido acético por C. guilliermondii também é constatada quando a levedura é
cultivada em hidrolisado hemicelulósico de bagaço de cana, sendo favorecida
com o esgotamento de xilose (FELIPE et ai., 1997a). Melhor crescimento
celular foi observado em condições aeróbicas, com aeração correspondente a
um kLa igual a 35 h-1 e pH inicial igual a 7,0. A capacidade de metabolizar ácido
acético pela levedura, sugere a possibilidade de detoxificação do hidrolisado
utilizando condições ótimas de pH e aeração (MORITA et ai., 2000b).

21
O metabolismo de acetato por leveduras ainda não foi completamente
elucidado. Sabe-se que o acetato pode ser utilizado como fonte de carbono,
sendo convertido em acetil-CoA pela enzima acetato quinase, com consumo de
ATP. SOUSA et ai. (1998) observaram que para Zygosaccharomyces bailii,
nem o ciclo =~ glioxilato, nem a glk,,:meogêr..Jse estão envolvidos no
metabolismo de ácido acético, mas sim o ciclo de Krebs. LEE et ai. (1996)
verificaram que o nível de mRNA e a atividade da enzima acetil-CoA hidrolase
são aumentados em meio contendo acetato, o que indica que a expressão
desta enzima pode estar envolvida na utilização de ácido acético. Acetil-CoA
hidrolase é uma das enzimas que controlam o nível de acetil-CoA e CoASH no
citossol.

22
3. OBJETIVOS
3.1 GERAL
Contribuir para o desenvolvimento de uma tecnologia de obtenção de
xilitol po: via biotecnológica a partir de bagaço de cana de açúce..
3.2 ESPECÍFICOS
• Avaliar a influência da adição de diferentes concentrações de ácido
acético na atividade das enzimas intracelulares xilose redutase (XR) e xilitol
desidrogenase (XDH) de C. guillíerrnondíí cultivada em hidrolisado
hemicelulósico de bagaço de cana de açúcar.
• Avaliar a influência da adição de diferentes concentrações de ácido
acético na atividade das enzimas XR e XDH de C. guí/Jíerrnondií cultivada em
meio sintético, simulando a composição de açúcares do hidrolisado de bagaço.
• Avaliar o efeito da adição de diferentes concentrações de acetato de
sódio nas atividades de XR e XDH.
• Verificar a existência da relação entre a concentração de ácido
acético nas atividades de XR e XDH e o rendimento e produtividade em xilitol.

23
4. MATERIAL E MÉTODOS
4.1 MICRORGANISMO E PREPARO DO INÓCULO
Os ensaios foram conduzidos com a levedura Candida gui/liermondii
FTI 20037, selecionada por BARBOSA et ai. (1988) para produção de xilitol.
Foi utilizada uma cultura estoque do DEBIQ/FAENQUIL, mantida em ágar
extrato de malte 5% a 4 ºC.
Para o preparo de inóculo, o cultivo da levedura foi realizado em frascos
Erlenmeyer de 125 ml, contendo 50 mL de meio composto de xilose (30 g/L),
suplementado com sulfato de amônia (2 g/L), cloreto de cálcio dihidratado
(O, 1 g/L), solução de extrato de farelo de arroz (20 g/L), conforme tem sido
empregado nos trabalhos do grupo (FELIPE et ai., 1997a; ALVES et ai., 1998;
RODRIGUES, 1999; SILVA, 2001). O cultivo foi feito em incubadora de
movimento rotatório (New Brunswick, Scientific Co.) com agitação de 200 rpm,
a 30 ºC, por 24 horas.
As células foram recuperadas por centrifugação a 2000xg (Cu-5000 -
Damon/lEC Division) e lavadas com água destilada estéril. Após a segunda
centrifugação as células foram utilizadas para o preparo da suspensão que foi
empregada como inóculo, com concentração igual a 1,0 g/L.
4.2 PREPARO DO HIDROLISADO DE BAGAÇO DE CANA DE AÇÚCAR
O bagaço de cana foi obtido na Usina Guarani, em Olímpia-SP.
Primeiramente o bagaço teve seu teor de umidade determinado através da
secagem de amostras em estufa a 100 ºC até peso constante. Em seguida foi
hidrolisado na planta de hidrólise ácida da FAENQUIL, em reator de aço inox
AISI 316, com capacidade volumétrica de 250 litros, com aquecimento indireto
por resistência elétrica em camisa de óleo térmico. A hidrólise foi realizada
conforme metodologia estabelecida por PESSOA JR. (1997), empregando as
seguintes condições: temperatura de 121 ºC, por 10 minutos e
100 mg de ácido sulfúrico por grama de matéria seca, para uma relação
sólido-líquido de 1: 1 O. O hidrolisado obtido foi centrifugado para remoção de

24
massa residual de sólidos (celulose e lignina) e armazenado a 4 ºCem câmara
fria.
4.3 CONCENTRAÇÃO E TRATAMENTO
O hidrolisado obtido foi submetido ao processo de concentração, com a
finalidade de aumentar em 3 vezes o teor inicial de xilose e reduzir o teor de
compostos tóxicos voláteis. A concentração foi realizada em um evaporador
rotativo modelo MA 175, com aquecimento por fluído térmico (água destilada)
em banho termostático controlado em aproximadamente 70 ºC.
O tratamento foi feito segundo metodologia estabelecida por ALVES et
ai. (1998), que consistiu em adicionar ao hidrolisado concentrado (pH 2,2),
óxido de cálcio comercial até pH 7,0, seguido de redução de pH até 5,5 com
ácido fosfórico. Ao hidrolisado foi adicionado carvão ativo (2,4% p/v), mantido
sob agitação de 200 rpm, por 1 hora, a 30 ºC. A cada alteração de pH e após o
tratamento com carvão o hidrolisado foi filtrado em papel de filtro, a vácuo em
filtro de porcelana, sendo descartado o precipitado.
Os hidrolisados original, concentrado e tratado foram caracterizados
quanto ao pH, à concentração de açúcares (xilose, glicose e arabinose) e
também quanto à concentração de compostos tóxicos (furfural,
hidroximetilfurfural, ácido acético e compostos fenólicos).
4.4 MEIOS E CONDIÇÕES DE FERMENTAÇÃO
Os experimentos para avaliação do efeito do ácido acético foram
realizados inicialmente em meio de composição semelhante a do hidrolisado
quanto às concentrações de xilose, glicose e arabinose, suplementado com os
mesmos nutrientes utilizados para o cultivo do inóculo, acrescentado de 5, 6, 8
e 1 O g/L de ácido acético. O pH de cada meio foi corrigido para 5,5 com
hidróxido de sódio 1 O M. Como controle foi utilizado meio sem adição de ácido.
Para avaliação do efeito do ácido acético em meio formulado com
hidrolisado, este foi tratado conforme descrito no item 3.3, e suplementado com
os mesmos nutrientes empregados para os meios anteriores. Ao meio de
cultura preparado foi adicionado ácido acético concentrado para as

25
concentrações finais iguais a 5, 6, 8 e 10 g/L, acertando-se o pH para 5,5 com
hidróxido de sódio 1 O M.
O hidrolisado e as soluções de glicose, arabinose e xilose foram
autoclavados a 111 ºC por 15 minutos. As soluções estoque de sulfato de
amónio e cloreto de cálcio di-hidratadato foram preparadas nas concent, ações
de 250 e 50 g/L, respectivamente, e esterilizadas a 121°C por 20 minutos.
A solução de extrato de farelo do arroz foi preparada conforme procedimento
anteriormente estabelecido, uma solução de 250 g/L foi autoclavada a 111 ºC,
por 15 minutos, seguida de centrifugação para remoção de sólidos (FELIPE et
ai., 1997a; ALVES et ai., 1998; RODRIGUES, 1999).
Os ensaios foram conduzidos em frascos Erlenmeyer de 125 ml com
50 ml de meio, em triplicata, a 30 ºC, sob agitação 200 rpm em incubadora de
movimento rotatório (New Brunswick, Scientific Co). Para o acompanhamento
da fermentação, as amostras representadas pelo volume total de cada frasco
Erlenmeyer, foram retiradas em intervalos de 12 horas (12, 24, 36, 48 e 58),
esta última amostra foi analisada neste tempo para evitar o esgotamento da
xilose no meio de cultura, prevenindo o consumo de xilitol.
Após a determinação da concentração celular e avaliação da viabilidade
e pureza da cultura. as células foram separadas por centrifugação a 2000xg
(Cu-5000 - Damon/lEC Division) e lavadas com água destilada estéril. O pH do
sobrenadante foi determinado e este foi congelado para posterior quantificação
da concentração de xilose, glicose, arabinose, ácido acético e xilitol. As células
recuperadas foram ressuspensas em tampão fosfato de potássio O, 1 M, pH 7,2
e congeladas para o rompimento celular.
4.5 MÉTODOS ANALÍTICOS
4.5.1 VIABILIDADE E PUREZA DA CULTURA
A viabilidade da cultura foi verificada por análise em microscópio óptico
(Leitz) de lâminas preparadas a fresco, coradas com azul de metileno
(0,01% p/v), dissolvido em citrato de sódio (2% p/v) (ODUMERO et ai., 1992),
enquanto a pureza foi avaliada a partir de lâminas fixadas e coradas com
fucsina.

26
4.5.2 DETERMINAÇÃO DA CONCENTRAÇÃO CELULAR
A concentração celular foi determinada por espectrometria a 600 nm, no
caso do preparo do inóculo, e por contagem de células em Câmara de
Neubauer (1/400 mm2 x 1/10 mm) para avaliação do crescimento celular nos
diferentes tempos de fermentação.
4.5.3 DETERMINAÇÃO DE PH
O pH das amostras foi determinado em pHmetro modelo 8474
(Micronal).
4.5.4 DETERMINAÇÃO DA CONCENTRAÇÃO DE AÇÚCARES E ÁCIDO ACÉTICO
As concentrações de glicose, xilose, arabinose, xilitol e ácido acético
foram determinadas em Cromatógrafo Líquido de Alta Eficiência (Shimadzu -
LC-10 AD), empregando-se as seguintes condições: coluna 810 RAD Aminex
HPX-87H (300 X 7,8 mm); temperatura da coluna 45°C, detector de índice de
refração RID-6\ eluente solução de ácido sulfúrico 0,01 N, fluxo de 0,6 ml/min;
volume da amostra injetada 20 µL.
As amostras, após devidamente diluídas, foram filtradas em filtro
Sep Pak C18 (MILLIPORE). O eluente, antes do uso, foi filtrado a vácuo em
membrana em éster de celulose, 0,45 µm de poro, 47 mm de diâmetro
(MILLIPORE) e simultaneamente foi desgaseificado em banho ultra-som
(THORTON) por 25 minutos.
4.5.5 DETERMINAÇÃO DA CONCENTRAÇÃO DE FURFURAL E HIDROXIMETILFURFURAL
As concentrações de furfural e hidroximetilfurfural foram determinadas
em Cromatógrafo Líquido de Alta Eficiência (Shimadzu - LC-1 O AD),
empregando-se as seguintes condições: coluna Hewlett-Packard RP 18
(200 mm); temperatura da coluna 25 ºC, detector de ultravioleta SPD-10A UV-
VIS no comprimento de onda de 276 nm, fluxo de 0,8 mllmin; eluente solução
de acetonitrila/água (1 :8) com 1 % de ácido acético; volume da amostra injetada
20 µL.

27
As amostras foram devidamente diluídas e filtradas em filtros Swennex,
com membrana HA em éster de celulose, 0,45 µm de poro, 0,47 mm de
diâmetro (MILLIPORE). Na composição do eluente, a água bidestilada foi
filtrada a vácuo empregando-se membrana HA em éster de celulose, 0,45µm
de poro, 0,47 mm de diâmetro ,ivilLLIPORE) e os outros componentes, como
ácido acético e acetonitrila, foram nas proporções adequadas, adicionados à água devidamente filtrada. Em seguida o eluente foi desgaseificado em banho
de ultra-som (THORTON) por 15 minutos.
4.5.6 DETERMINAÇÃO DA CONCENTRAÇÃO DE FENÓIS
Os compostos fenólicos foram determinados utilizando o método
descrito por KIM & YOO (1996), que consiste na adição de O, 1 ml de
K3Fe(CN)6 8 mM a 1,5 ml de amostra, seguida de adição imediata de O, 1 ml
de uma solução de FeC'3.6H20 O, 1 M em HCI O, 1 M. A absorbância da solução
resultante foi lida após 5 minutos, em uma cubeta de quartzo de 1 cm de
percurso ótico, a 700 nm, em espectrofotômetro computadorizado BECKMAN®
DU 640 8 e comparada com uma curva de calibração feita usando vanilina
como padrão.
4.6 ROMPIMENTO CELULAR
Para o rompimento celular e obtenção das enzimas intracelulares xilose
redutase e xilitol desidrogenase, as células foram descongeladas,
centrifugadas e ressuspensas em tampão fosfato de potássio 0,1 M, pH 7,2 a
uma concentração de 15 a 20 g/L. Foi empregado um volume de 5 ml da
suspensão em um tubo cônico, o qual foi imerso em um banho refrigerado a
-10 ºC para prevenir o aquecimento decorrente das vibrações ultra-sônicas
(Vibra Cell 1 OOW Sonics & Materiais). Ao tubo foi introduzida uma sonda de
13 mm, que emite vibrações a uma frequência de 20 KHz, programada para
vibrações em pulso de 1 s com intervalos de 1 s, durante 35 min. Os restos
celulares foram separados em centrífuga (Jouan NRL 1812) a 6708xg por
1 O min, a 4 ºC, recolhendo-se o sob renadante para determinação das

28
atividades enzimáticas de xilose redutase e xilitol desidrogenase e proteína
solúvel.
4.7 DETERMINAÇÃO DE PROTEÍNA SOLÚVEL
A concentração de proteína no extrato livre de células foi determinada
conforme método BRADFORD (1976), empregando-se albumina sérica bovina
(BSA) como padrão e reagente de Bradford (Azul-Brilhante de Coomassie
G-250) como cromóforo. As leituras de absorbância foram feitas em
espectrofotômetro a 550 nm.
4.8 MEDIDA DAS ATIVIDADES ENZIMÁTICAS
As atividades enzimáticas foram determinadas em extratos celulares
preparados a partir da suspensão de células, realizadas segundo método
utilizado por SENE (2000b). Os valores de absorbância foram determinados em
espectrofotômetro BECKMAN® OU 640 B.
O meio padrão de reação para xilose redutase foi composto por 200 µL
de extrato celular; 570 µL de água; 150 µL de P-mercaptoetanol O, 1 M; 80 µL
de tampão fosfato de potássio (KH2P04-K2HP04) 1,0 M, pH 7,2; 100 µL de
NADPH 1,2 mM e 200 µL de xilose 0,5 M; correspondendo a um volume total
de 1,3 ml. A atividade foi determinada pela diminuição da absorbância a
340 nm, decorrente da oxidação do NADPH.
Para xilitol desidrogenase, o meio padrão de reação foi constituído de
100 µL de extrato celular; 450 µL de água; 200 µL de p-mercaptoetanol 0,1 M;
150 µL de tampão TRIS 0,5 M, pH 8,6; 150 µL de NAD+ 1,26 mM e 150 µL de
xilitol 0,5 M; correspondendo a um volume total de 1,20 ml. A atividade foi
determinada pelo aumento da absorbância a 340 nm, decorrente da redução do NAD+.
Uma unidade internacional de atividade enzimática (UI) foi definida como
a quantidade de enzima que catalisa a formação de um micromol de NADPH
por minuto, no caso de xilose redutase, ou de NAD+ para xilitol desidrogenase,
à temperatura ambiente, utilizando-se o coeficiente de extinção molar para os
cofatores de 6220 M-1.cm·1 (SENE 2000b).

29
4.9 EFEITO DO ACETATO DE SÓDIO NAS ATIVIDADES ENZIMÁTICAS
Para avaliação na atividade in vitro de xilose redutase e xilitol
desidrogenase em presença de acetato de sódio, foi utilizado o ensaio
enzimático descrito no item anterior, porém adicionando-se uma solução de
acetato até as concentrações finais iguais a 0,25; 0,5; 1,0; 2,0 e 4,0 g/L,
mantendo-se o volume final pela diminuição da quantidade de água. A solução
estoque de acetato de sódio teve seu pH corrigido com hidróxido de sódio 1 M,
para o pH ótimo de atividade enzimática (7,2 para xilose redutase e 8,6 para
xilitol desidrogenase).
4.10 DETERMINAÇÃO DOS PARÂMETROS FERMENTATIVOS
• Fator de rendimento de xilitol em relação a xilose consumida (Yp/s)
O fator de rendimento expressa a razão de massa de xilitol produzido
por massa de xilose consumida, em gramas, calculado pela equação:
_ M PJ-Pi Yp1s- --=----
1:lS Sf-Si
Onde:
Pi e P, correspondem às concentrações inicial e final de xilitol (g/L)
Si e St correspondem às concentrações inicial e final de xilose (g/L)
• Produtividade volumétrica de xilitol (Qp)
A produtividade de xilitol expressa a concentração de xilitol produzida
(g/L) pelo tempo (h), calculada pela equação:
Op = M _ Pf-Pi t t
Onde:
Pi e Pt correspondem às concentrações inicial e final de xilitol (g/L)
t: tempo de fermentação (h)

30
• Eficiência de conversão (rl)
Este parâmetro expresso em porcentagem, representa a razão entre o
fator de rendimento obtido experimentalmente e o fator de rendimento teórico
de 0,917 g/g calculado segundo BARBOSA et ai. (1988).
l1 = Yp i s • 100 0,917
• Velocidade de Consumo
Este parâmetro representa a velocidade de consumo de um composto,
calculado pela variação da sua concentração (g/L) dividida pelo tempo (h) de
fermentação.
ÍlC Ci-Cf V= =--
t t
Onde:
Ci e Cf correspondem às concentrações inicial e final do composto
analisado (g/L)
t: tempo de fermentação (h)
31
5. RESULTADOS E DISCUSSÃO
5.1 EFEITO DO ÁCIDO ACÉTICO NA FERMENTAÇÃO EM MEIO SINTÉTICO
A avaliação do efeito do ácido na fermentação foi realizada em meio
sintético simulando a composição do hidrolisado quanto às concentrações de
glicídeos (xilose, glicose e arabinose) contendo 5, 6, 8 e 1 O g/L de ácido
acético, concentrações superiores à considerada inibitória (3,0 g/L) para
formação de xilitol, por C. guilliermondii, segundo FELIPE et ai. (1995). A
Tabela li apresenta as características destes meios.
TABELA li - Características dos meios de cultura após a adição de ácido acético.
Meios de Cultura Características
li Ili IV
Ácido Acético (g/L) 5,330 6,204 7,974 10,22 Xilose (g/L) 46,01 47,49 42,28 46,49
Glicose (g/L) 2,474 2,591 2,625 2,635 Arabinose (g/L) 4,878 5,006 5,120 5,347 ~ - Proporção xilose:ácido acético 8,632 7,655 5,302 4,549
pH 5,87 5,26 5,77 5,69
Os meios sintéticos preparados possuíram constituição similar quanto
aos açúcares e pH variando próximo a 5,5, valor ótimo para produção de xilitol
segundo SENE et ai. (1998). As concentrações finais de ácido acético (5,3; 6,2;
8,0 e 10,2 g/L) foram compatíveis com a esperada para avaliação do efeito
deste ácido na fermentação.
5.1.1 EFEITO DO ÁCIDO ACÉTICO NO CONSUMO DE AÇÚCARES
Analisando os gráficos apresentados na Figura 4, observa-se que o
consumo de xilose não foi influenciado pelas concentrações de ácido acético
avaliadas. Em 58 horas o consumo de xilose foi igual a 99, 95, 97 e 95%, para
os meios com 5, 6, 8 e 1 O g/L de ácido acético, respectivamente.
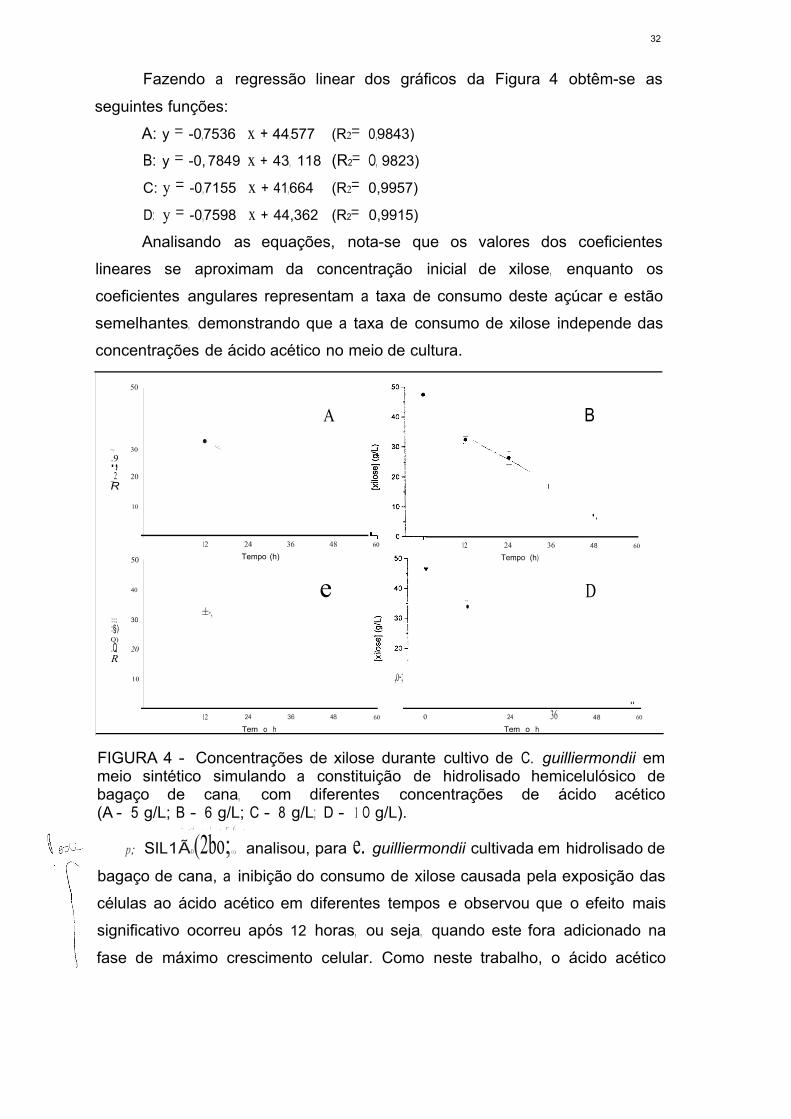
32
Fazendo a regressão linear dos gráficos da Figura 4 obtêm-se as
seguintes funções:
A: y = -0,7536 x + 44,577 (R2= 0,9843)
B: y = -0, 7849 x + 43, 118 (R2= O, 9823)
C: y = -0,7155 x + 41,664 (R2= 0,9957)
D: y = -0,7598 x + 44,362 (R2= 0,9915)
Analisando as equações, nota-se que os valores dos coeficientes
lineares se aproximam da concentração inicial de xilose, enquanto os
coeficientes angulares representam a taxa de consumo deste açúcar e estão
semelhantes, demonstrando que a taxa de consumo de xilose independe das
concentrações de ácido acético no meio de cultura.
50
A B • '<, ~ 30
.9 ., _2 20 R I
10 ·- 12 24 36 48
Tempo (h) 60 12 24 36
Tempo (h) 48 60
-- •
50
e D 40
::; 30 :§)
±-,
Q) .Q 20 R
10 ,o-; ..
24 36 Tem o h
60 12 24 36
Tem o h o 48 48 60
FIGURA 4 - Concentrações de xilose durante cultivo de C. guilliermondii em meio sintético simulando a constituição de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 5 g/L; B - 6 g/L; C - 8 g/L; D - 1 O g/L).
·-· .,.-i , \ , r (. ,
p; SIL1Ã0(2bo;0) analisou, para e. guilliermondii cultivada em hidrolisado de
bagaço de cana, a inibição do consumo de xilose causada pela exposição das
células ao ácido acético em diferentes tempos e observou que o efeito mais
significativo ocorreu após 12 horas, ou seja, quando este fora adicionado na
fase de máximo crescimento celular. Como neste trabalho, o ácido acético
33
quando adicionado no tempo inicial de fermentação, a inibição do consumo de
xilose pelo aumento do teor de ácido não foi significativa. Isto ocorre
possivelmente devido à capacidade de adaptação da levedura à presença
deste ácido.
Verifica-se pe:c Tabela Ili que o consurr., de arat..nose se iniciou após
24 horas de fermentação. Isto ocorre devido ao esgotamento inicial de glicose
e a diminuição da concentração de xilose em torno de 50%, levando a levedura
a recorrer a outras fontes de carbono para seu metabolismo, ou ainda por estar
diminuindo a concentração de monossacarídeos concorrentes ao transporte de
arabinose. Observa-se que o consumo de arabinose foi igual a 32, 26, 21 e
33% para os meios com 5, 6, 8 e 1 O g/L de ácido acético. respectivamente.
TABELA Ili - Concentrações de arabinose durante cultivo de C. guilliermondii em meio sintético contendo ácido acético em diferentes concentrações. [Ácido Acético] O g/L 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) [Arabinose] g/L o 5,838 4,878 5,006 5,120 5,347 12 4,871 4,587 4,963 4,878 5,096 24 4,913 5,432 5,561 5,497 5,264 36 4,084 4,781 5,175 4,770 5,082 48 3,679 4,301 4,466 4,361 4,503 58 1,826 3,316 3,728 3,547 3,609
Quanto à glicose, esta foi totalmente consumida nas primeiras 12 horas
de cultivo nos quatro meios de cultura com diferentes concentrações de ácido
acético. Já na primeira amostragem, após 12 horas de fermentação, este
açúcar não foi detectado, possivelmente pelo fato de que a glicose, além de ser
a fonte de carbono preferencial, também está presente em baixas
concentrações (-2,6 g/L).
Como neste trabalho, em experimentos realizados anteriormente por
FELIPE et ai. (1995), a levedura C. guilliermondii consumiu glicose
independente da concentração de ácido acético no meio de cultura. ROSA et
ai. (1998) constataram favorecimento na formação de xilitol por
C. guilliermondii, em meio sintético, na presença de glicose até 5,0 g/L. Para
34
Oebaryomyces hansenii, a adição de 10% de glicose em meio contendo 60 g/L
de xilose, resultou em aumento 30% na produtividade de xilitol (TAVARES et
ai., 2000). De acordo com os resultados, foi verificado que a presença de ácido
acético, xilose e arabinose não afetam o consumo de: glicose por
C. guillermondii. Tal comportamento se deve ao fato de que o transporte de
glicose pela membrana celular é independente dos demais açúcares (LUCAS &
van UDEN, 1986) além do que as enzimas utilizadas no metabolismo de glicose
são constitutivas (WALKER, 1999).
5.1.2 CONSUMO DE ÁCIDO ACÉTICO DURANTE A FERMENTAÇÃO
TABELA IV - Ácido acético consumido durante cultivo de C. guilliermondii em meio sintético, contendo diferentes concentrações de ácido acético. [Ácido Acético] 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) Ácido Acético Consumido (g/L) 12 1,942 1,566 1,483 1,568 24 2, 101 1,893 1,708 2,538 36 2,498 2,412 2,387 2,861 48 2,884 2,869 2,819 3,241 58 3,080 3, 101 3,040 3,683
O ácido acético foi consumido durante toda a fermentação (Tabela IV),
sendo que nas primeiras 12 horas a velocidade foi maior. como também pode
ser observado na Figura 5. Estes resultados sugerem que este ácido é
metabolizado concomitantemente à glicose, como já observado para
Zygosaccharomyces bailii por SOUSA et ai. (1998).
Observa-se que o consumo do ácido acético é favorecido com o
aumento da concentração deste no meio de cultura, porque houve aumento
das velocidades nas quatro condições estudadas, 0,053; 0,053; 0,052 e
0,064 g/L.h, para os meios com 5, 6, 8 e 1 O g/L de ácido acético,
respectivamente.
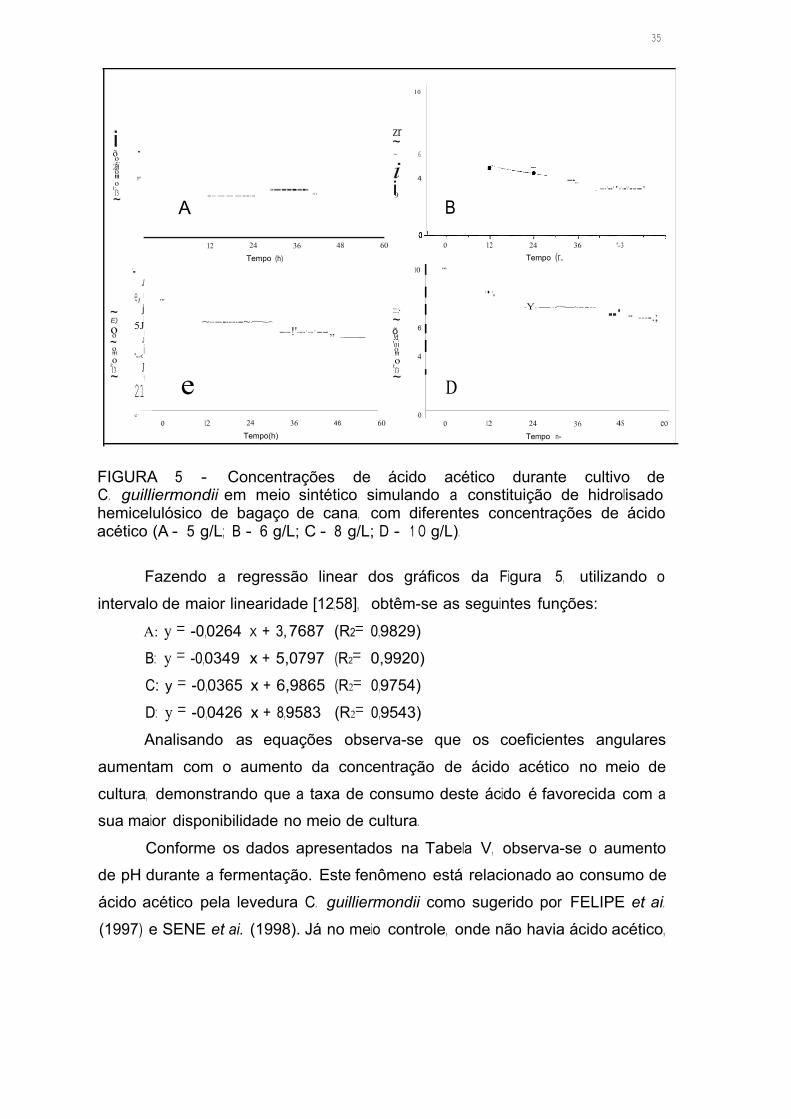
10
B
zr ~ ~ 6
i 4
--------- 9 ---------- .... i
i õ - o
:ãi o '" ,. o i:, ·13 ~
--.. - --·--··-·--·-··---·
A
12 24 36 48 60 o 12 24 36 '-3
Tempo (h) Tempo (r., ·- 10 ...
J .. 1 ···- e, ...
-Y---~----- ~ j ::::;- --· E) 1 ~-----------~~- ~ -- ----.; o 5J õ 6
1 ---!'---·--·--.,, ___ o J .!d ~ i 'oi o o "' '--< "' 4 o J o i:, i:, ·13 ·13 ~ 1 ~ 21 e D
e· o o 12 24 36 46 60 o 12 24 36 4S eo
Tempo(h) Tempo n-
FIGURA 5 - Concentrações de ácido acético durante cultivo de C. guilliermondii em meio sintético simulando a constituição de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 5 g/L; B - 6 g/L; C - 8 g/L; D - 1 O g/L).
Fazendo a regressão linear dos gráficos da Figura 5, utilizando o
intervalo de maior linearidade [12,58], obtêm-se as seguintes funções:
A: y = -0,0264 X + 3, 7687 (R2= 0,9829)
B: y = -0,0349 x + 5,0797 (R2= 0,9920)
C: y = -0,0365 x + 6,9865 (R2= 0,9754)
D: y = -0,0426 x + 8,9583 (R2= 0,9543)
Analisando as equações observa-se que os coeficientes angulares
aumentam com o aumento da concentração de ácido acético no meio de
cultura, demonstrando que a taxa de consumo deste ácido é favorecida com a
sua maior disponibilidade no meio de cultura.
Conforme os dados apresentados na Tabela V, observa-se o aumento
de pH durante a fermentação. Este fenômeno está relacionado ao consumo de
ácido acético pela levedura C. guilliermondii como sugerido por FELIPE et ai.
(1997) e SENE et ai. (1998). Já no meio controle, onde não havia ácido acético,
35
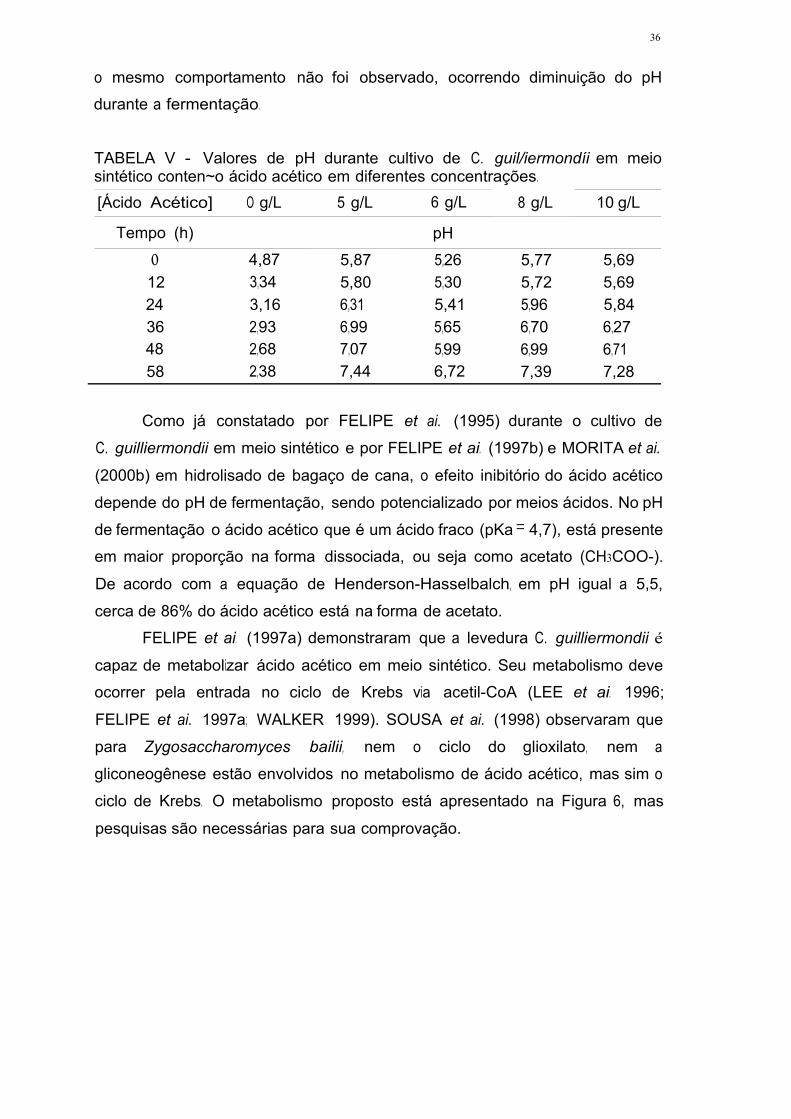
36
o mesmo comportamento não foi observado, ocorrendo diminuição do pH
durante a fermentação.
TABELA V - Valores de pH durante cultivo de C. guil/iermondíi em meio sintético conten~o ácido acético em diferentes concentrações. [Ácido Acético] O g/L 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) pH o 4,87 5,87 5,26 5,77 5,69 12 3,34 5,80 5,30 5,72 5,69 24 3,16 6,31 5,41 5,96 5,84 36 2,93 6,99 5,65 6,70 6,27 48 2,68 7,07 5,99 6,99 6,71 58 2,38 7,44 6,72 7,39 7,28
Como já constatado por FELIPE et ai. (1995) durante o cultivo de
C. guilliermondii em meio sintético e por FELIPE et ai. (1997b) e MORITA et ai.
(2000b) em hidrolisado de bagaço de cana, o efeito inibitório do ácido acético
depende do pH de fermentação, sendo potencializado por meios ácidos. No pH
de fermentação o ácido acético que é um ácido fraco (pKa = 4,7), está presente
em maior proporção na forma dissociada, ou seja como acetato (CH3COO-).
De acordo com a equação de Henderson-Hasselbalch, em pH igual a 5,5,
cerca de 86% do ácido acético está na forma de acetato.
FELIPE et ai. (1997a) demonstraram que a levedura C. guilliermondii é
capaz de metabolizar ácido acético em meio sintético. Seu metabolismo deve
ocorrer pela entrada no ciclo de Krebs via acetil-CoA (LEE et ai. 1996;
FELIPE et ai. 1997a; WALKER 1999). SOUSA et ai. (1998) observaram que
para Zygosaccharomyces bailii, nem o ciclo do glioxilato, nem a
gliconeogênese estão envolvidos no metabolismo de ácido acético, mas sim o
ciclo de Krebs. O metabolismo proposto está apresentado na Figura 6, mas
pesquisas são necessárias para sua comprovação.
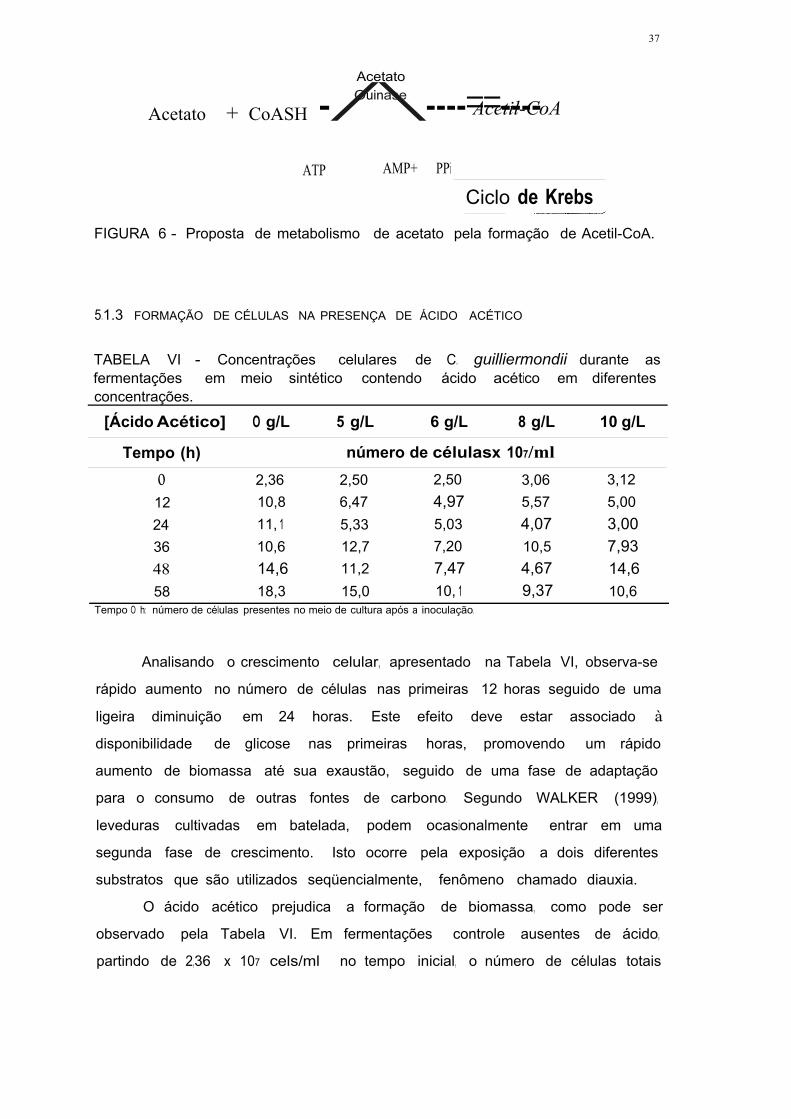
37
Acetato + CoASH -/\----==---- Acetato Ouinase
Acetil-CoA
ATP AMP+ PPi
Ciclo de Krebs FIGURA 6 - Proposta de metabolismo de acetato pela formação de Acetil-CoA.
5.1.3 FORMAÇÃO DE CÉLULAS NA PRESENÇA DE ÁCIDO ACÉTICO
TABELA VI - Concentrações celulares de C. guilliermondii durante as fermentações em meio sintético contendo ácido acético em diferentes concentrações.
[Ácido Acético] O g/L 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) número de células x 107/ml o 12 24 36 48 58
2,36 10,8 11, 1 10,6 14,6 18,3
2,50 6,47 5,33 12,7 11,2 15,0
2,50 4,97 5,03 7,20 7,47 10, 1
3,06 5,57 4,07 10,5 4,67 9,37
3,12 5,00 3,00 7,93 14,6 10,6
Tempo O h: número de células presentes no meio de cultura após a inoculação.
Analisando o crescimento celular, apresentado na Tabela VI, observa-se
rápido aumento no número de células nas primeiras 12 horas seguido de uma
ligeira diminuição em 24 horas. Este efeito deve estar associado à
disponibilidade de glicose nas primeiras horas, promovendo um rápido
aumento de biomassa até sua exaustão, seguido de uma fase de adaptação
para o consumo de outras fontes de carbono. Segundo WALKER (1999),
leveduras cultivadas em batelada, podem ocasionalmente entrar em uma
segunda fase de crescimento. Isto ocorre pela exposição a dois diferentes
substratos que são utilizados seqüencialmente, fenômeno chamado diauxia.
O ácido acético prejudica a formação de biomassa, como pode ser
observado pela Tabela VI. Em fermentações controle ausentes de ácido,
partindo de 2,36 x 107 cels/ml no tempo inicial, o número de células totais

38
após 58 horas foi de 18,3 x 107 cels/ml. Observa-se para as condições com
ácido acético que os valores de células totais são inferiores ao do controle,
tendendo a diminuir com o aumento na concentração de ácido acético. Este
efeito inibitório já foi demonstrado por FELIPE et ai. (1995) e SILVA (2001 ).
TABELA VII - Proporções de células inviáveis durante cultivo de C. guilliermondií em meio sintético contendo ácido acético em diferentes concentrações.
[Ácido Acético] 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) Células inviáveis (%)
12 3,09 6,71 4,19 6,67 24 5,00 2,65 1.64 2,22 36 11,0 3,70 12,66 2,52 48 17,8 14,29 17.14 14,61 58 19,8 22,70 21.35 23,27
Tempo O h: número de células inviáveis no meio de cultura após a inoculação.
Analisando a proporção de células inviáveis, apresentada na Tabela VII,
observa-se que com o aumento da concentração de ácido acético o número de
células inviáveis tende a aumentar, demonstrando que além de inibir o
crescimento celular, este ácido acelera a mortalidade das leveduras.
5.1.4 EFEITO DO ÁCIDO ACÉTICO NA FORMAÇÃO DE XILITOL
A formação de xilitol foi independente da concentração de ácido acético,
o que pode ser verificado pela Figura 7. Depois de 58 horas, a concentração de
xilitol no meio foi de 23,50; 20,77; 22,61 e 21,98 g/L para os meios com 5, 6, 8
e 10 g/L de ácido acético, respectivamente. Pode-se notar que entre 12 e
48 horas há uma maior linearidade dos gráficos. Isto ocorre porque nas
primeiras 12 horas a presença de glicose interfere na formação de xilitol e
depois de 48 horas, com o esgotamento de xilose, o xilitol começa a ser
consumido mais rapidamente.
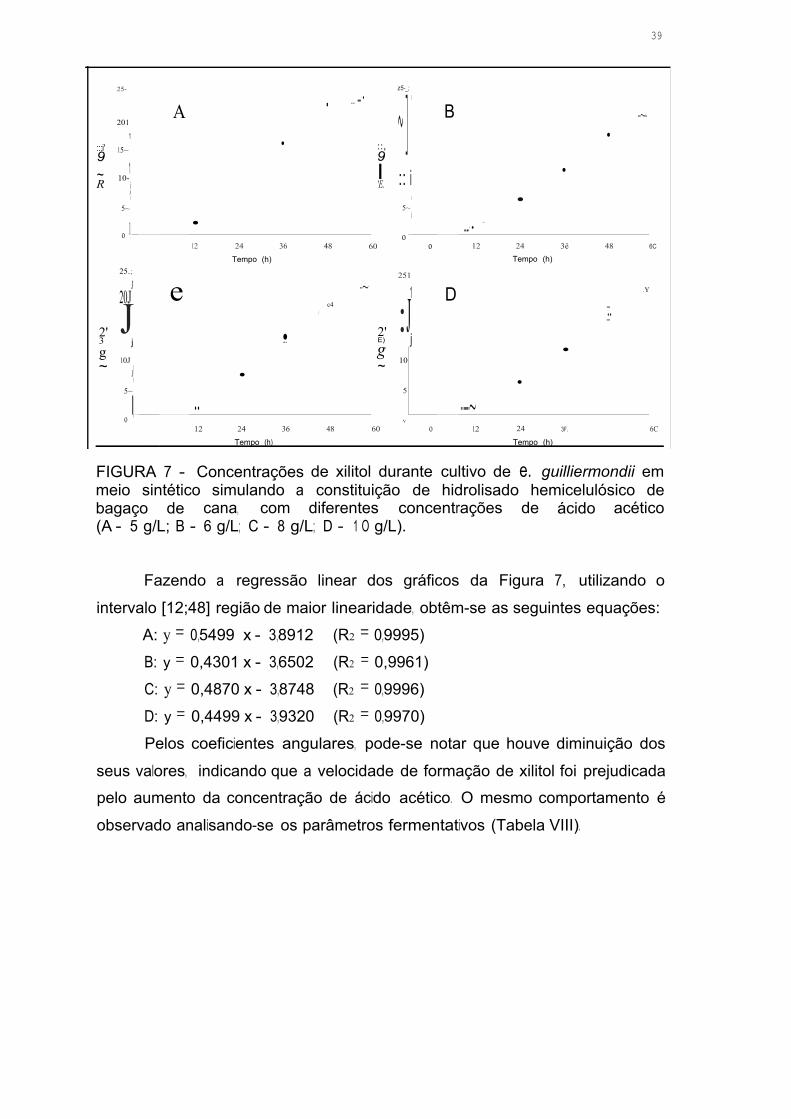
25- z5-_;
-- -·· i
A . B 201 ~] --~-
1 • :::i' 15~ • ::, 9
1 9
~ I :: i • 10- R i 'E.
J 1 1 •
5~ 5~
1 i
-- • --·· o . o 12 24 36 48 60 o 12 24 3ê 48 6C
Tempo (h) Tempo (h) 25..; 251
J --~ 20J e 1 D .Y
J c4 :J - / .. -
2' 2' 3 j • E) j -- g
10J g •
10 ~ 1 ~ J • 1 • 5~ 5
l .. --~ o ' V
12 24 36 48 60 o 12 24 3F. 6C
Tempo (h) Tempo (h)
FIGURA 7 - Concentrações de xilitol durante cultivo de e. guilliermondii em meio sintético simulando a constituição de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 5 g/L; B - 6 g/L; C - 8 g/L; D - 1 O g/L).
Fazendo a regressão linear dos gráficos da Figura 7, utilizando o
intervalo [12;48] região de maior linearidade, obtêm-se as seguintes equações:
A: y = 0,5499 x - 3,8912 (R2 = 0,9995)
B: y = 0,4301 x - 3,6502 (R2 = 0,9961)
C: y = 0,4870 x - 3,8748 (R2 = 0,9996)
D: y = 0,4499 x - 3,9320 (R2 = 0,9970)
Pelos coeficientes angulares, pode-se notar que houve diminuição dos
seus valores, indicando que a velocidade de formação de xilitol foi prejudicada
pelo aumento da concentração de ácido acético. O mesmo comportamento é
observado analisando-se os parâmetros fermentativos (Tabela VIII).
39
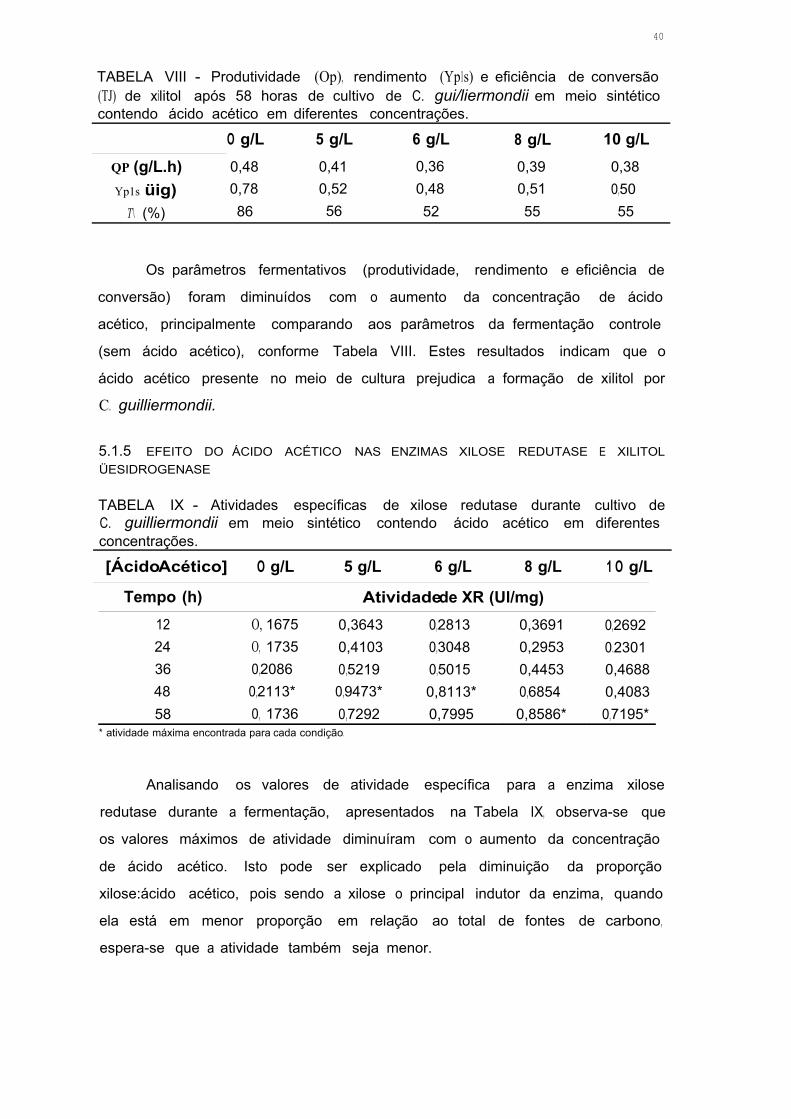
40
TABELA VIII - Produtividade (Op), rendimento (Yp1s) e eficiência de conversão (TJ) de xilitol após 58 horas de cultivo de C. gui/liermondii em meio sintético contendo ácido acético em diferentes concentrações.
O g/L 5 g/L 6 g/L 8 g/L 10 g/L QP (g/L.h) 0,48 0,41 0,36 0,39 0,38 Yp1s üig) 0,78 0,52 0,48 0,51 0,50
T\ (%) 86 56 52 55 55
Os parâmetros fermentativos (produtividade, rendimento e eficiência de
conversão) foram diminuídos com o aumento da concentração de ácido
acético, principalmente comparando aos parâmetros da fermentação controle
(sem ácido acético), conforme Tabela VIII. Estes resultados indicam que o
ácido acético presente no meio de cultura prejudica a formação de xilitol por
C. guilliermondii.
5.1.5 EFEITO DO ÁCIDO ACÉTICO NAS ENZIMAS XILOSE REDUTASE E XILITOL ÜESIDROGENASE
TABELA IX - Atividades específicas de xilose redutase durante cultivo de C. guilliermondii em meio sintético contendo ácido acético em diferentes concentrações.
[Ácido Acético] O g/L 5 g/L 6 g/L 8 g/L 1 O g/L
Tempo (h) Atividade de XR (Ul/mg) 12 O, 1675 0,3643 0,2813 0,3691 0,2692 24 O, 1735 0,4103 0,3048 0,2953 0.2301 36 0,2086 0,5219 0,5015 0,4453 0,4688 48 0,2113* 0,9473* 0,8113* 0,6854 0,4083 58 0, 1736 0,7292 0,7995 0,8586* 0,7195*
* atividade máxima encontrada para cada condição.
Analisando os valores de atividade específica para a enzima xilose
redutase durante a fermentação, apresentados na Tabela IX, observa-se que
os valores máximos de atividade diminuíram com o aumento da concentração
de ácido acético. Isto pode ser explicado pela diminuição da proporção
xilose:ácido acético, pois sendo a xilose o principal indutor da enzima, quando
ela está em menor proporção em relação ao total de fontes de carbono,
espera-se que a atividade também seja menor.
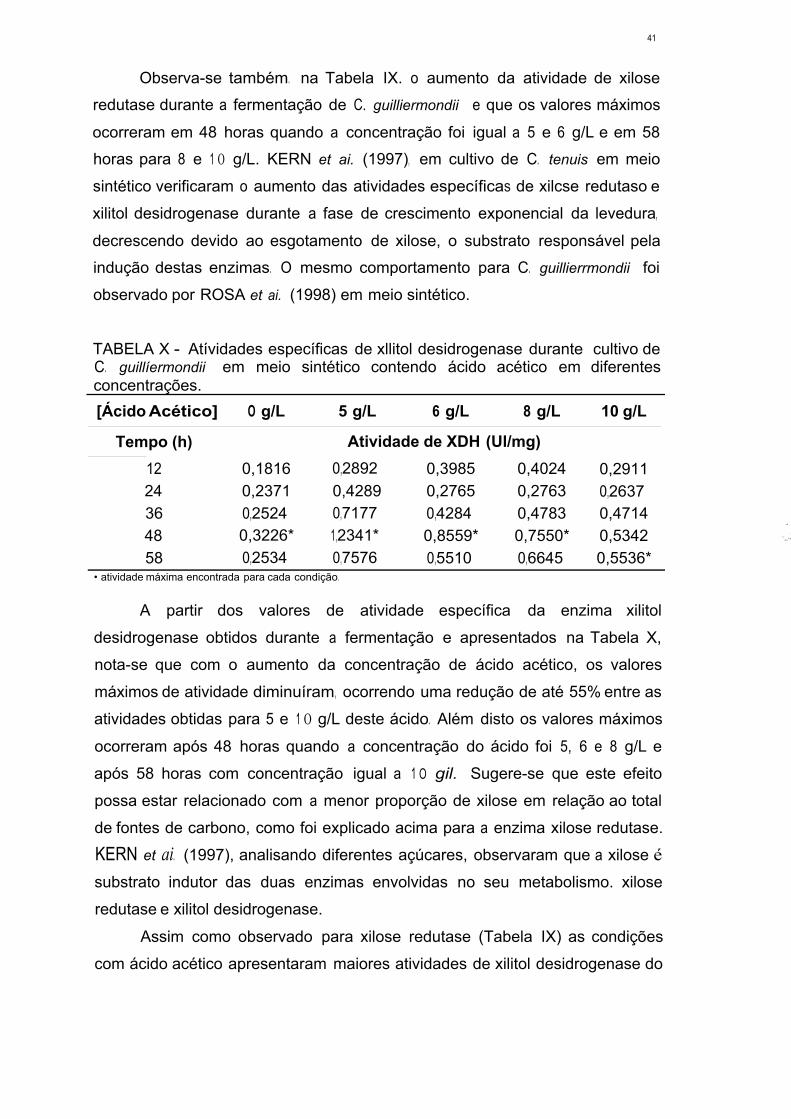
41
Observa-se também. na Tabela IX. o aumento da atividade de xilose
redutase durante a fermentação de C. guilliermondii e que os valores máximos
ocorreram em 48 horas quando a concentração foi igual a 5 e 6 g/L e em 58
horas para 8 e 1 O g/L. KERN et ai. (1997), em cultivo de C. tenuis em meio
sintético verificaram o aumento das atividades específicas de xilcse redutaso e
xilitol desidrogenase durante a fase de crescimento exponencial da levedura,
decrescendo devido ao esgotamento de xilose, o substrato responsável pela
indução destas enzimas. O mesmo comportamento para C. guillierrmondii foi
observado por ROSA et ai. (1998) em meio sintético.
TABELA X - Atívidades específicas de xllitol desidrogenase durante cultivo de C. guillíermondii em meio sintético contendo ácido acético em diferentes concentrações. [Ácido Acético] O g/L 5 g/L 6 g/L 8 g/L 10 g/L
Tempo (h) Atividade de XDH (Ul/mg) 12 0,1816 0,2892 0,3985 0,4024 0,2911 24 0,2371 0,4289 0,2765 0,2763 0,2637 36 0,2524 0,7177 0,4284 0,4783 0,4714
-- 48 0,3226* 1,2341* 0,8559* 0,7550* 0,5342 - ~_.,.,
58 0,2534 0,7576 0,5510 0,6645 0,5536* • atividade máxima encontrada para cada condição.
A partir dos valores de atividade específica da enzima xilitol
desidrogenase obtidos durante a fermentação e apresentados na Tabela X,
nota-se que com o aumento da concentração de ácido acético, os valores
máximos de atividade diminuíram, ocorrendo uma redução de até 55% entre as
atividades obtidas para 5 e 1 O g/L deste ácido. Além disto os valores máximos
ocorreram após 48 horas quando a concentração do ácido foi 5, 6 e 8 g/L e
após 58 horas com concentração igual a 1 O gil. Sugere-se que este efeito
possa estar relacionado com a menor proporção de xilose em relação ao total
de fontes de carbono, como foi explicado acima para a enzima xilose redutase.
KERN et ai. (1997), analisando diferentes açúcares, observaram que a xilose é substrato indutor das duas enzimas envolvidas no seu metabolismo. xilose
redutase e xilitol desidrogenase.
Assim como observado para xilose redutase (Tabela IX) as condições
com ácido acético apresentaram maiores atividades de xilitol desidrogenase do

42
que no meio controle, sugerindo que a presença do ácido acético no meio de
cultura pode aumentar o metabolismo de xilose. Este aumento pode estar
associado à regeneração dos ATPs gastos para o transporte de prótons do
meio intracelular para o meio extracelular, a fim de se restabelecer a
neutralidade do pH intraceluls., acidificado pelo ácido acético (Figura 3).
5.2 EFEITO DO ÁCIDO ACÉTICO NA FERMENTAÇÃO EM HIDROLISADO
5.2.1 CARACTERIZAÇÃO DO HIDROLISADO
O hidrolisado hemicelulósico obtido do processo de hidrólise ácida de
bagaço de cana de açúcar foi concentrado a vácuo até 3 vezes e depois
submetido ao tratamento descrito na metodologia. As características do
hidrolisado após cada etapa estão apresentadas na Tabela XI. A constituição
foi semelhante às apresentadas por ALVES et a/.(1998), RODRIGUES (1999),
MORITA et ai. (2000a), SENE (2000b) e SILVA (2001).
TABELA XI - Características dos hidrolisados hemicelulósicos de bagaço de cana de açúcar original, após a concentração e depois do tratamento. Características Original Concentrado Tratado*
........ ·- ·-·-··········--~--- -
Ácido Acético (g/L) 2,54 2,16 4,72 Xilose (g/L) 14,85 45,03 47,17
Glicose (g/L) 0,39 2,41 2,37 Arabinose (g/L) 1,50 4,41 4,78
Furfural (g/L) 0,050 0,038 Nd Hidroximetilfurfural (g/L) 0,004 0,010 Nd
Fenóis Totais (g/L) 1,023 3.032 1,089 pH 1,27 0,67 5,40
nd: não detectado por cromatografia liquida. * tratamento após concentração
A glicose presente em baixas concentrações favorece o processo
fermentativo, uma vez que esta é a fonte de carbono preferencial da levedura
no início do seu crescimento no meio de cultura (YAHASHI et ai., 1996).
Segundo KIM et ai. (1999), o consumo inicial de glicose pela levedura e

43
consequente aumento de massa celular favorece a utilização de xilose para a
produção de xilitol.
O monossacarídeo arabinose, também presente no hidrolisado, pode
aumentar a eficiência de conversão de xilose em xilitol, uma vez que este é
indutor das enzimas xilose redutase e xititol desidrogenase (LEE et ai., í SJG;
KERN et ai., 1997).
Verifica-se também na Tabela XI, além do ácido acético, a presença de
inibidores do metabolismo microbiano, como furfural, hidroximetilfurfural e
fenóis originados durante o processo de hidrólise da biomassa lignocelulósica
(PARAJÓ et ai., 1998c).
O ácido acético e o furfural, por serem compostos voláteis, tiveram suas
concentrações diminuídas após a etapa de concentração a vácuo. O
tratamento foi eficiente para a remoção de furfural e hidroximetilfurfural e
diminuição de compostos fenólicos. A concentração de fenóis foi menor do que
a considerada inibitória para o crescimento de Sacharomyces cerevisiae, ou
seja, menor que 2 g/L (PALMIQVIST et ai., 2000). Mesmo após o tratamento, a
concentração de ácido acético permaneceu acima da considerada como
inibitória (3,0 g/L) ao crescimento e produção de xilitol por C. guil/iermondii
(FELIPE et ai., 1995).
TABELA XII - Características dos meios de cultura preparados com hidrolisado contendo ácido acético em diferentes concentrações. ~~--''--~~~~~~~~~~-
Meios de Cultura Características
li Ili IV
Ácido Acético (g/L) 3,303 4,029 6,122 8,273 Xilose (g/L) 36,99 38,41 38,56 37,46
Glicose (g/L) 1,690 1,734 1,886 1,726 Arabinose (g/L) 4,601 4,689 4,367 4,684
Proporção xilose:ácido acético 11,20 9,533 6,299 4,528 pH 5,19 5,55 5,67 5,64
Para avaliar o efeito de diferentes concentrações de ácido acético na
bioconversão de xilose em xilitol, este foi adicionado ao hidrolisado, já
contendo 4,72 g/L de ácido. a fim de atingir as concentrações iguais a 6, 8 e
1 O g/L. Os meios preparados apresentaram como concentrações finais deste
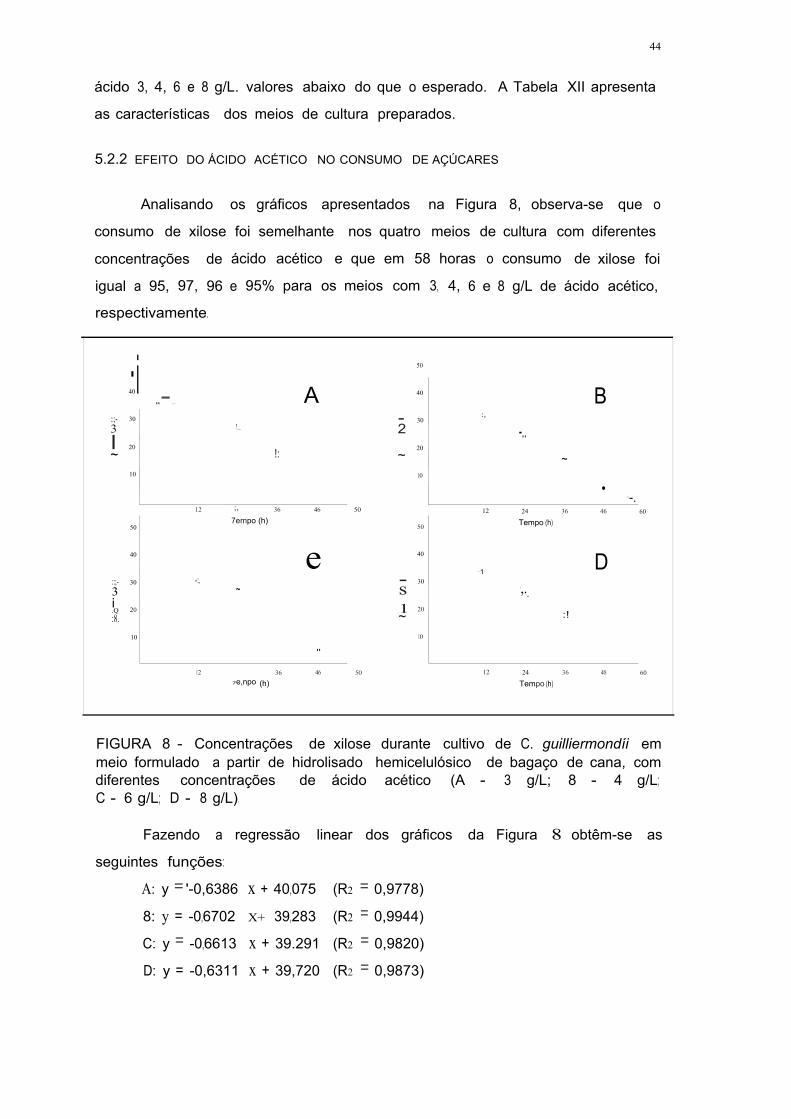
ácido 3, 4, 6 e 8 g/L. valores abaixo do que o esperado. A Tabela XII apresenta
as características dos meios de cultura preparados.
5.2.2 EFEITO DO ÁCIDO ACÉTICO NO CONSUMO DE AÇÚCARES
Analisando os gráficos apresentados na Figura 8, observa-se que o
consumo de xilose foi semelhante nos quatro meios de cultura com diferentes
concentrações de ácido acético e que em 58 horas o consumo de xilose foi
igual a 95, 97, 96 e 95% para os meios com 3, 4, 6 e 8 g/L de ácido acético,
respectivamente.
·i 50
40 A 40 B .. - ... ::;- 30 - 30
·:,
3 !_ 2 I -..
20 20 ~ !: ~ ~
10 10
•· ··-. 12 ;, 36 46 50 12 24 36 46 60
50 7empo (h)
50 Tempo (h)
40 e 40 D ···1
::;- 30 -:. - 30
3 ~ s ·,._ i
20 1 20 .Q :! :8. ~ 10 10 ..
12 36 46 50 12 24 36 48 60 7e,npo (h) Tempo (h)
FIGURA 8 - Concentrações de xilose durante cultivo de C. guilliermondíi em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 3 g/L; 8 - 4 g/L; C - 6 g/L; D - 8 g/L).
Fazendo a regressão linear dos gráficos da Figura 8 obtêm-se as
seguintes funções:
A: y = '-0,6386 x + 40,075 (R2 = 0,9778)
8: y = -0,6702 X+ 39,283 (R2 = 0,9944)
C: y = -0,6613 x + 39.291 (R2 = 0,9820)
D: y = -0,6311 x + 39,720 (R2 = 0,9873)
44

45
Analisando as equações, nota-se que os valores dos coeficientes
lineares se aproximam da concentração inicial de xilose, enquanto os
coeficientes angulares representam a taxa de consumo deste açúcar. Como
estes valores são semelhantes, pode-se inferir que a taxa de consumo de
xilose indccende ds concentração de ácido acético no meio de cultura. Esse
comportamento se deve ao fato da levedura poder se adaptar à presença de
ácido acético, inclusive utilizando-o para seu metabolismo. Porém os
coeficientes angulares foram menores do que os obtidos para meio sintético,
sugerindo que em hidrolisado o consumo de xilose ocorre mais lentamente,
possivelmente pela presença de outros compostos inibidores, além do ácido
acético.
TABELA XIII - Concentrações de arabinose durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo ácido acético em diferentes concentrações.
[Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) [Arabinose] g/L o 4,601 4,689 4,367 4,684 12 3,904 4,795 4,828 4,612 24 3,645 3,497 3,308 3,495 36 3,120 3,486 3,370 3,765 48 2,819 2,488 2,492 2,847 58 2,375 2,165 2, 195 2,417
A partir dos dados apresentados na Tabela XIII, verifica-se que o
consumo de arabinose é iniciado após 12 horas de fermentação para as
condições de 4, 6 e 8 g/L de ácido acético, enquanto para o meio com 3 g/L o
consumo é simultâneo ao dos demais açúcares, como a xilose (Figura 8) e a
glicose (dados não apresentados). Observa-se o consumo de 52. 46, 50 e 52%
para os meios com 3, 4, 6 e 8 g/L de ácido acético, respectivamente,
demonstrando que, como já observado para meio sintético, a concentração de
ácido acético não interfere no consumo de arabinose.
Baixas taxas de assimilação de arabinose foram observadas durante
fermentações em hidrolisado de bagaço de cana (SENE, 2000b), como
também em hidrolisado de eucalipto (FERRARI et ai., 1992) e de palha de arroz
(ROBERTO et ai., 1995). Em meio sintético (Tabela Ili), o consumo, além de
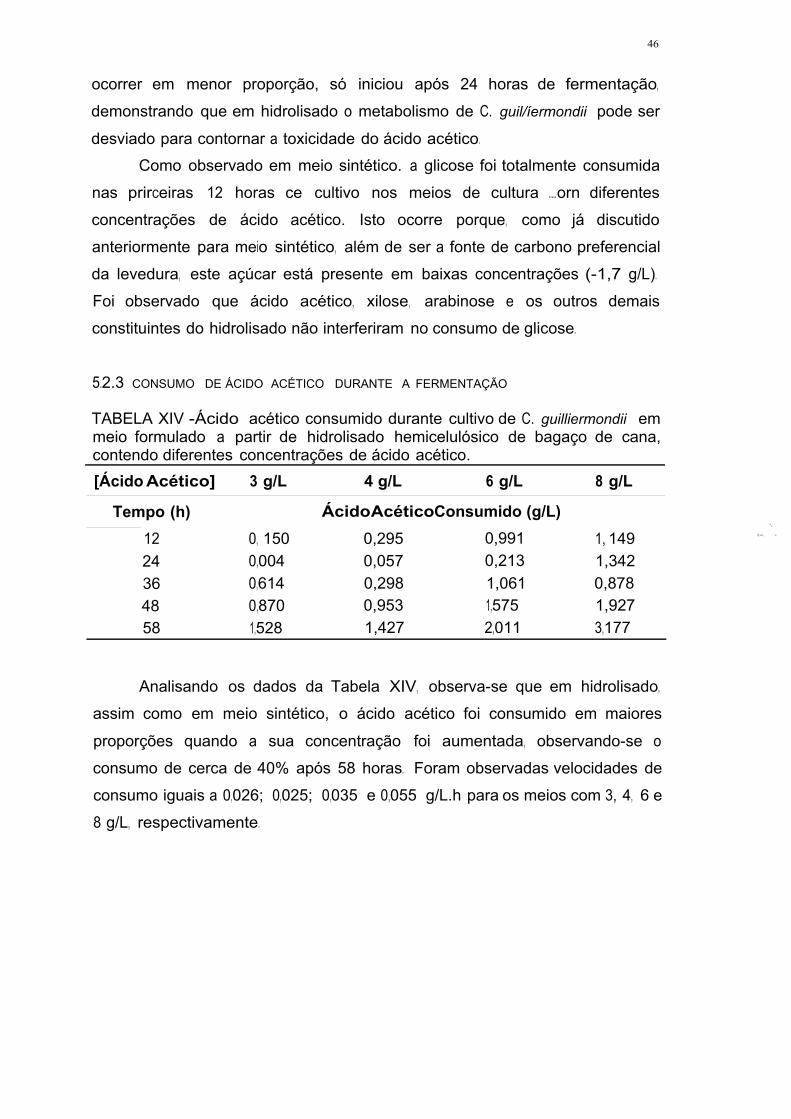
46
ocorrer em menor proporção, só iniciou após 24 horas de fermentação,
demonstrando que em hidrolisado o metabolismo de C. guil/íermondii pode ser
desviado para contornar a toxicidade do ácido acético.
Como observado em meio sintético. a glicose foi totalmente consumida
nas prirceiras 12 horas ce cultivo nos meios de cultura ..... orn diferentes
concentrações de ácido acético. Isto ocorre porque, como já discutido
anteriormente para meio sintético, além de ser a fonte de carbono preferencial
da levedura, este açúcar está presente em baixas concentrações (-1,7 g/L).
Foi observado que ácido acético, xilose, arabinose e os outros demais
constituintes do hidrolisado não interferiram no consumo de glicose.
5.2.3 CONSUMO DE ÁCIDO ACÉTICO DURANTE A FERMENTAÇÃO
TABELA XIV -Ácido acético consumido durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo diferentes concentrações de ácido acético. [Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) Ácido Acético Consumido (g/L) -.
12 0, 150 0,295 0,991 1, 149 e-. -
24 0,004 0,057 0,213 1,342 36 0,614 0,298 1,061 0,878 48 0,870 0,953 1,575 1,927 58 1,528 1,427 2,011 3,177
Analisando os dados da Tabela XIV, observa-se que em hidrolisado,
assim como em meio sintético, o ácido acético foi consumido em maiores
proporções quando a sua concentração foi aumentada, observando-se o
consumo de cerca de 40% após 58 horas. Foram observadas velocidades de
consumo iguais a 0,026; 0,025; 0,035 e 0,055 g/L.h para os meios com 3, 4, 6 e
8 g/L, respectivamente.
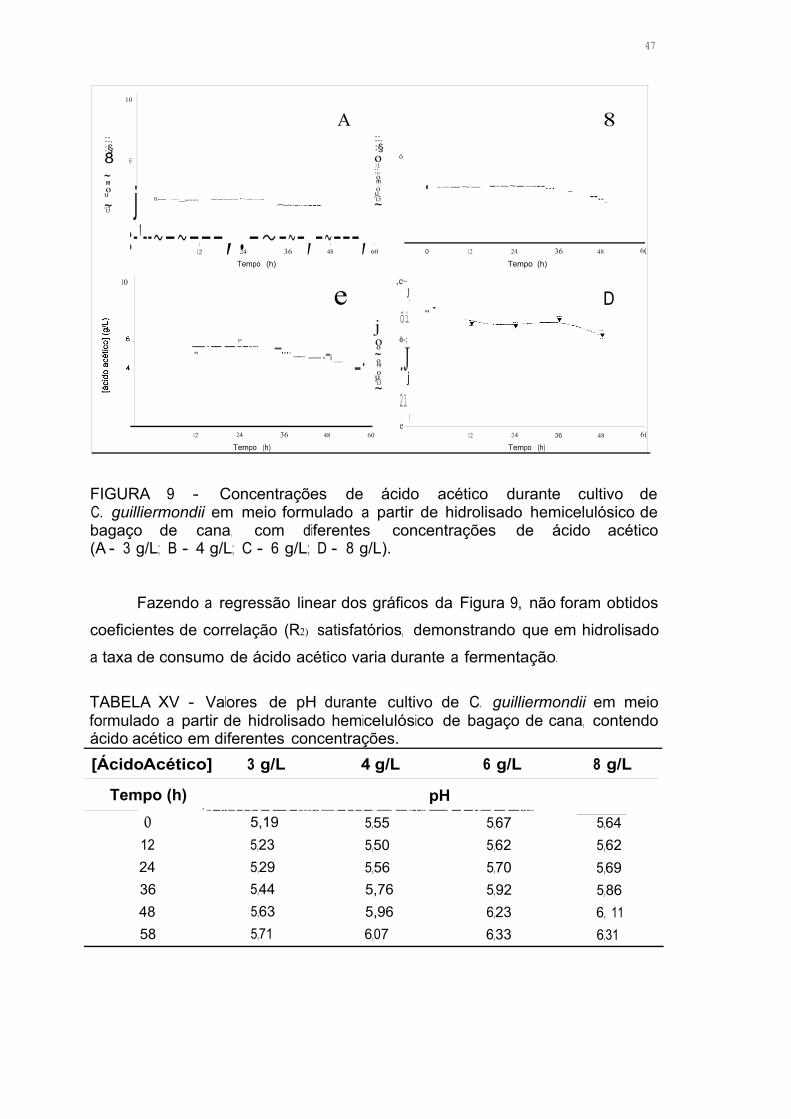
47
10
. -·-----~-· ---~-------... _
A 8 ::::, :§ o ô .\1 :;. o "' o u ºi3 ~
::::, :§ 8 6
~ "' o u
ºi3 j ~
:-'--~-~---,.-~-~-,-~---, 11------ ---__,,....... -----·-·----- ~---------
--..._
1 1 1 12 24 36 48 60 o
Tempo (h)
,e~
e J 1
ôi .. - j ,. o ê-; --------- o .J .. - ..... ___ ~ --""i __ o -·· "' o j u ºi3 ~ 21
! e 12 24 36 48 60
Tempo (h)
24 48 6( 36 12
Tempo (h)
10
D
36 48 6( 12 24
Tempo (h)
FIGURA 9 - Concentrações de ácido acético durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 3 g/L; B - 4 g/L; C - 6 g/L; D - 8 g/L).
Fazendo a regressão linear dos gráficos da Figura 9, não foram obtidos
coeficientes de correlação (R2) satisfatórios, demonstrando que em hidrolisado
a taxa de consumo de ácido acético varia durante a fermentação.
TABELA XV - Valores de pH durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo ácido acético em diferentes concentrações. [Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) pH ·--------·--------------------------------- o 5,19 5,55 5,67 5,64
12 5,23 5,50 5,62 5,62 24 5,29 5,56 5,70 5,69 36 5,44 5,76 5,92 5,86 48 5,63 5,96 6,23 6, 11 58 5,71 6,07 6,33 6,31
48
Observa-se pela Tabela XV, que o pH aumenta durante a fermentação.
Como sugerido anteriormente, este fenômeno pode estar relacionado ao
consumo de ácido acético pela levedura C. guilliermondii. Nota-se que o
aumento do pH foi menor do que em meio sintético, isto porque menos ácido
acét.co foi consumido cm hidrolisado.
O consumo de ácido acético por C. guilliermondii cultivada em
hidrolisado hemicelulósico já havia sido observado por FELIPE et ai. (1997b) e
MORITA et ai., (2000b). Isto sugere que esta levedura pode ser utilizada para
detoxificação do hidrolisado otimizando-se as condições de pH e aeração.
5.2.4 FORMAÇÃO DE CÉLULAS NA PRESENÇA DE ÁCIDO ACÉTICO
TABELA XVI - Concentrações celulares de C. guilliermondii durante as fermentações em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo ácido acético em diferentes concentrações. [Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) o 2,88 12 5,50 24 6,08 36 6,50 48 7,08 58 11,5
número de células x 107/ml 2,42 6,50 6,67 6,83 9,08 13,3
2,04 1,98 7,42 7,17 8,25 7,33 8,33 7,42 8,42 7,50 10,3 11,3
Tempo O h: número de células presentes no meio de cultura após a inoculação
Analisando o crescimento celular (Tabela XVI), observa-se que o
aumento na concentração de ácido acético não prejudica a formação de
biomassa, diferente do encontrado utilizando meio sintético. Quanto à proporção de células inviáveis, apresentada na Tabela XVII, não é observada
influência da concentração de ácido acético na mortalidade das leveduras,
indicando que em hidrolisado, C. guilliermondii tem capacidade de se adaptar e
manter seu crescimento.
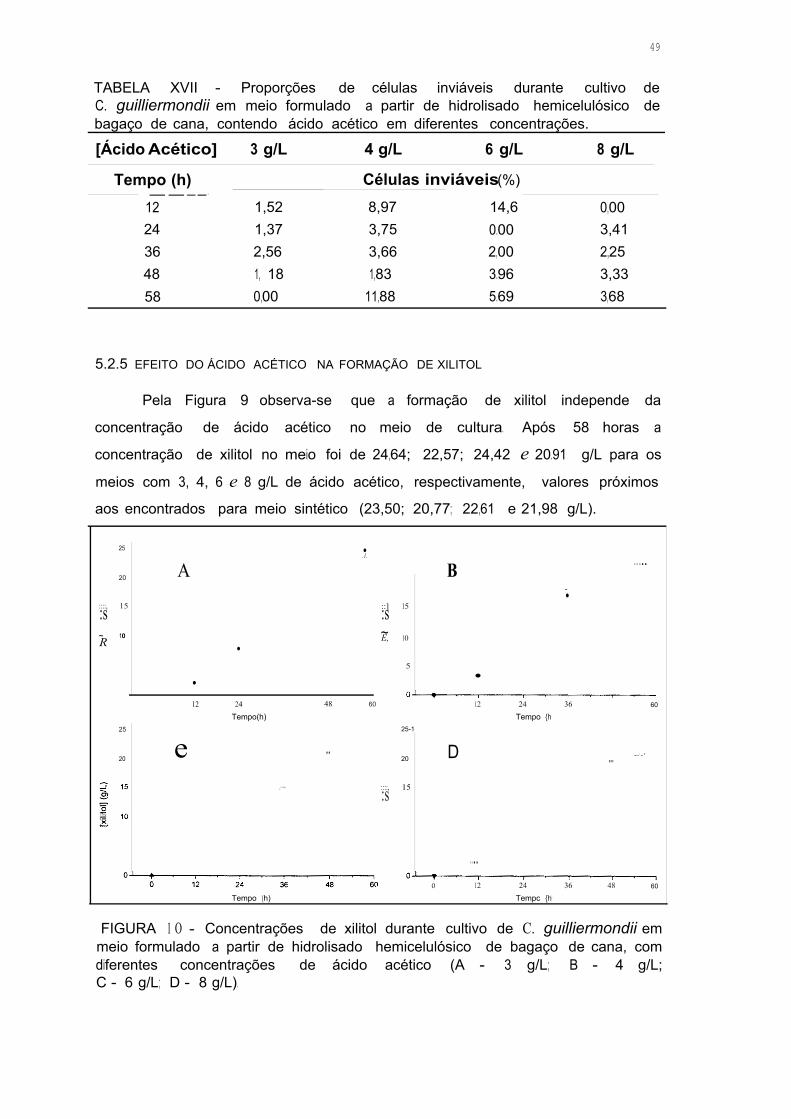
49
TABELA XVII - Proporções de células inviáveis durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo ácido acético em diferentes concentrações.
[Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) Células inviáveis (%) ----· 12 1,52 8,97 14,6 0,00 24 1,37 3,75 0.00 3,41 36 2,56 3,66 2,00 2,25 48 1, 18 1,83 3.96 3,33 58 0,00 11,88 5.69 3,68
5.2.5 EFEITO DO ÁCIDO ACÉTICO NA FORMAÇÃO DE XILITOL
Pela Figura 9 observa-se que a formação de xilitol independe da
concentração de ácido acético no meio de cultura. Após 58 horas a
concentração de xilitol no meio foi de 24,64; 22,57; 24,42 e 20.91 g/L para os
meios com 3, 4, 6 e 8 g/L de ácido acético, respectivamente, valores próximos
aos encontrados para meio sintético (23,50; 20,77; 22,61 e 21,98 g/L).
::::, 15 :s ~ 10 R
25
20
25
20
• .l.
A
•
::] 15 :s ~ E, 10
5
B .......
•
- •
• 12 24
Tempo {h 36 60 12 48 60
25-1
20
....
24 Tempo(h)
e .. D ... ---·-·
_..., ::::, 15 :s
o 12 24 Tempc {h'·
36 48 60
Tempo (h)
FIGURA 1 O - Concentrações de xilitol durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, com diferentes concentrações de ácido acético (A - 3 g/L; B - 4 g/L; C - 6 g/L; D - 8 g/L).

50
Fazendo a regressão linear dos gráficos da Figura 1 O. utilizando o
intervalo [12:48] obtêm-se as seguintes funções:
A: y = -0,4963 x - 3,871 (R2 = 0,9995)
8: y = -0,5065 x - 2,435 (R2 = 0,9885)
C: y = -0,496~ x - 2,867 (R2 = 0,9997)
D: y = -0,4934 x - 3,829 (R2 = 0,9949)
Analisando a variação dos coeficientes angulares das equações obtidas,
pode-se inferir que a formação de xilitol ocorre com a mesma velocidade e é
independente da concentração de ácido acético no meio de cultura
TABELA XVIII - Produtividade (Qp), rendimento (Ypts) e eficiência de conversão (ri) de xilitol após 58 horas de cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo diferentes concentrações de ácido acético.
3 g/L 4 g/L 6 g/L 8 g/L Qp (g/L.h) 0,42 0,39 0,42 0,36 Ypts (g/g) 0,70 0,60 0,66 0,59
ri (%) 77 66 72 64
Foi observado que a produtividade em xilitol foi similar entre as
condições estudadas, com diminuição no meio com 8 g/L de ácido acético. Os
parâmetros rendimento e eficiência de conversão foram infiuenciados pela
concentração de ácido acético. Desta forma quanto menor a proporção
xilose/ácido acético, menor o rendimento e a eficiência de conversão.
Comparando com os parâmetros em meio sintético a produtividade foi
semelhante, contudo o rendimento e a eficiência foram maiores em hidrolisado
hemicelulósico. Porém todos os parâmetros foram menores do que em
fermentações controle (na ausência de ácido acético), que apresentou
rendimento igual a 0,48 g/L.h, rendimento de 0,78 g/g e eficiência de 86%.
5.2.6 EFEITO DO ÁCIDO ACÉTICO NAS ENZIMAS XILOSE REDUTASE E XILITOL ÜESIDROGENASE
De acordo com os valores de atividade específica para a enzima xilose
redutase apresentados na Tabela XIX, nota-se que a concentração crescente
de ácido acético pouco influenciou nos valores máximos de atividades. A
atividade foi reduzida em 24% em hidrolisado em relação ao meio sintético.
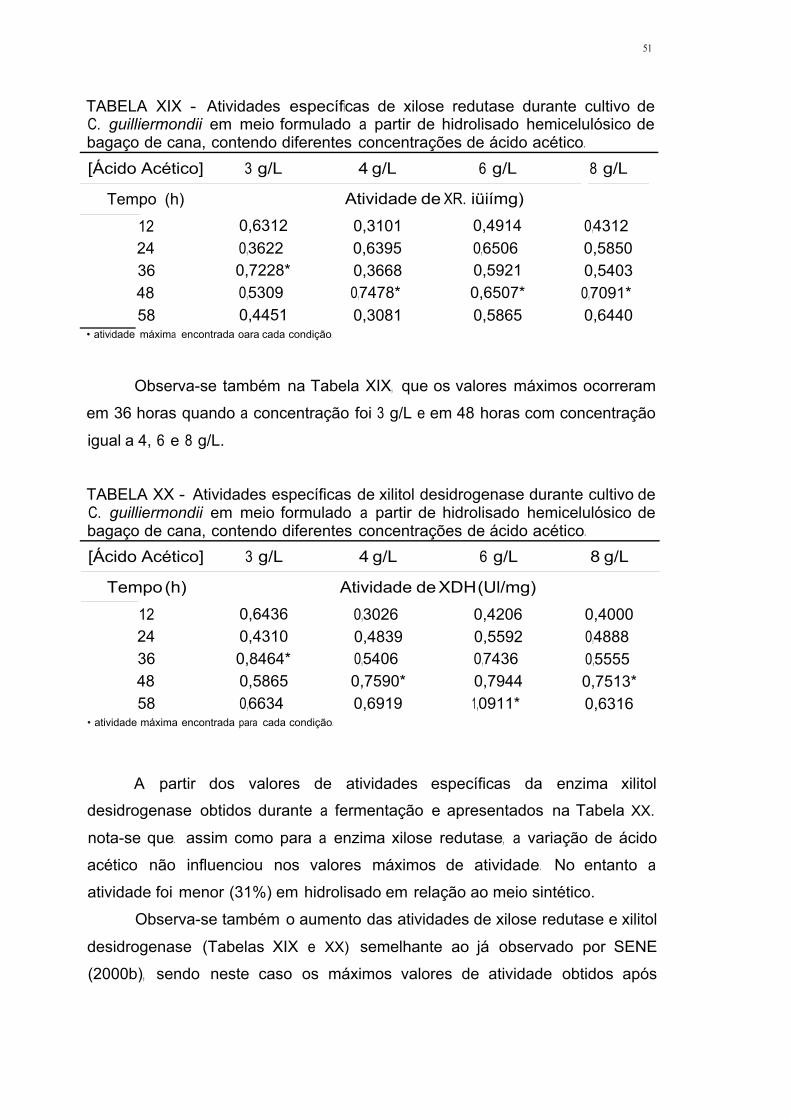
51
TABELA XIX - Atividades específicas de xilose redutase durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo diferentes concentrações de ácido acético. [Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) Atividade de XR. iüiímg)
12 0,6312 0,3101 0,4914 0,4312 24 0,3622 0,6395 0,6506 0,5850 36 0,7228* 0,3668 0,5921 0,5403 48 0,5309 0,7478* 0,6507* 0,7091* 58 0,4451 0,3081 0,5865 0,6440
• atividade máxima encontrada oara cada condição.
Observa-se também na Tabela XIX, que os valores máximos ocorreram
em 36 horas quando a concentração foi 3 g/L e em 48 horas com concentração
igual a 4, 6 e 8 g/L.
TABELA XX - Atividades específicas de xilitol desidrogenase durante cultivo de C. guilliermondii em meio formulado a partir de hidrolisado hemicelulósico de bagaço de cana, contendo diferentes concentrações de ácido acético. [Ácido Acético] 3 g/L 4 g/L 6 g/L 8 g/L
Tempo (h) Atividade de XDH (Ul/mg)
12 0,6436 0,3026 0,4206 0,4000 24 0,4310 0,4839 0,5592 0,4888 36 0,8464* 0,5406 0,7436 0,5555 48 0,5865 0,7590* 0,7944 0,7513* 58 0,6634 0,6919 1,0911* 0,6316
• atividade máxima encontrada para cada condição.
A partir dos valores de atividades específicas da enzima xilitol
desidrogenase obtidos durante a fermentação e apresentados na Tabela XX.
nota-se que. assim como para a enzima xilose redutase, a variação de ácido
acético não influenciou nos valores máximos de atividade. No entanto a
atividade foi menor (31%) em hidrolisado em relação ao meio sintético.
Observa-se também o aumento das atividades de xilose redutase e xilitol
desidrogenase (Tabelas XIX e XX) semelhante ao já observado por SENE
(2000b), sendo neste caso os máximos valores de atividade obtidos após
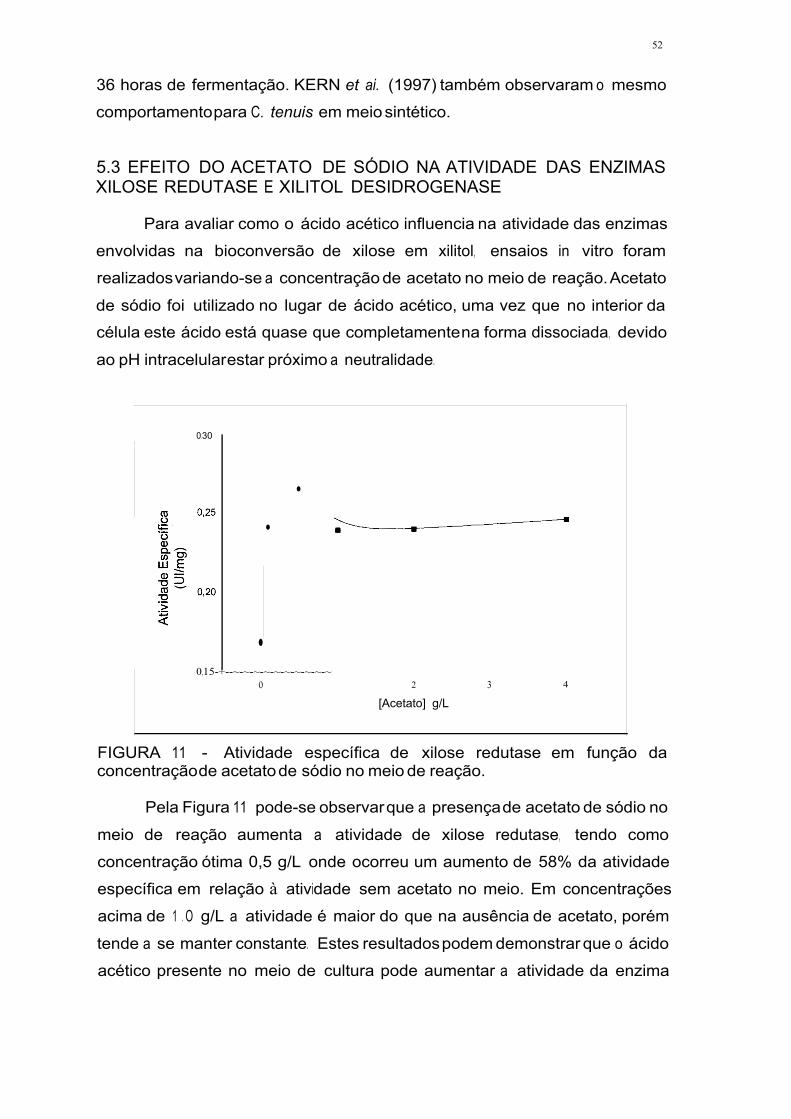
52
36 horas de fermentação. KERN et ai. (1997) também observaram o mesmo
comportamento para C. tenuis em meio sintético.
5.3 EFEITO DO ACETATO DE SÓDIO NA ATIVIDADE DAS ENZIMAS XILOSE REDUTASE E XILITOL DESIDROGENASE
Para avaliar como o ácido acético influencia na atividade das enzimas
envolvidas na bioconversão de xilose em xilitol, ensaios in vitro foram
realizados variando-se a concentração de acetato no meio de reação. Acetato
de sódio foi utilizado no lugar de ácido acético, uma vez que no interior da
célula este ácido está quase que completamente na forma dissociada, devido
ao pH intracelular estar próximo a neutralidade.
o 2
[Acetato] g/L 3 4
0.30
•
•
• 0,15-+--~-~-~-~-~-~-~-~-~-~
FIGURA 11 - Atividade específica de xilose redutase em função da concentração de acetato de sódio no meio de reação.
Pela Figura 11 pode-se observar que a presença de acetato de sódio no
meio de reação aumenta a atividade de xilose redutase, tendo como
concentração ótima 0,5 g/L onde ocorreu um aumento de 58% da atividade
específica em relação à atividade sem acetato no meio. Em concentrações
acima de 1 . O g/L a atividade é maior do que na ausência de acetato, porém
tende a se manter constante. Estes resultados podem demonstrar que o ácido
acético presente no meio de cultura pode aumentar a atividade da enzima
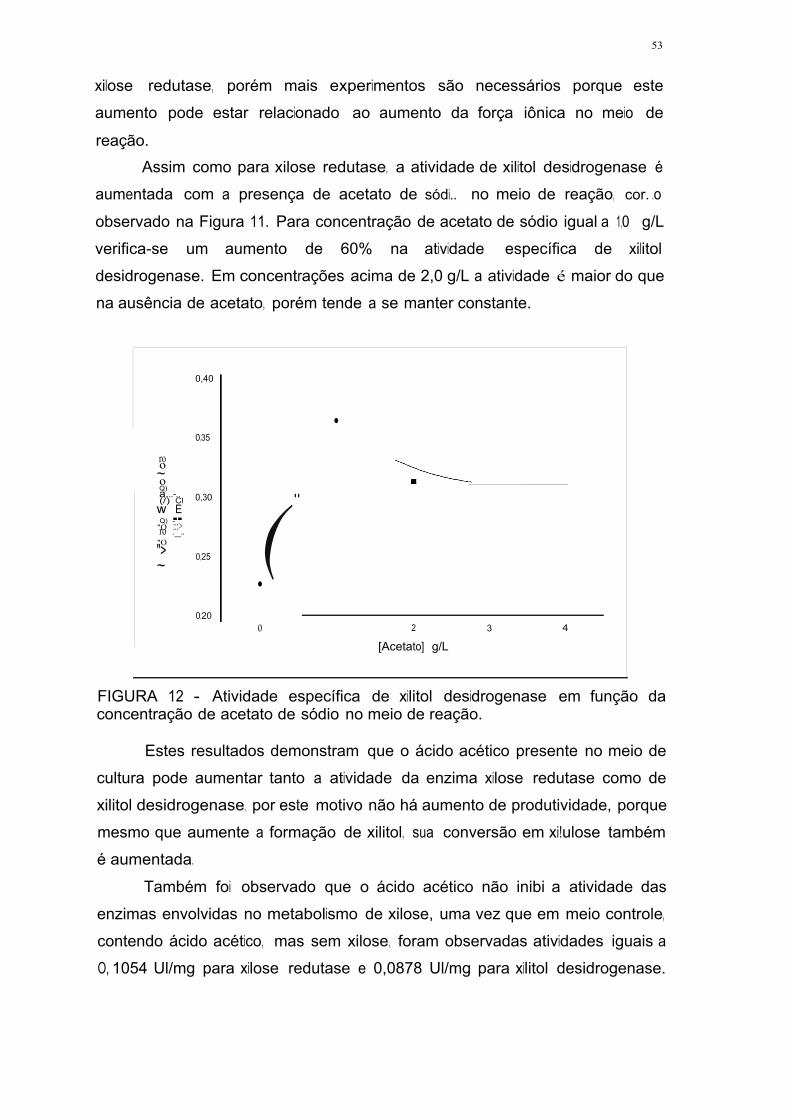
53
xilose redutase, porém mais experimentos são necessários porque este
aumento pode estar relacionado ao aumento da força iônica no meio de
reação. Assim como para xilose redutase, a atividade de xilitol desidrogenase é
aumentada com a presença de acetato de sódi.. no meio de reação, cor. .o
observado na Figura 11. Para concentração de acetato de sódio igual a 1,0 g/L
verifica-se um aumento de 60% na atividade específica de xilitol
desidrogenase. Em concentrações acima de 2,0 g/L a atividade é maior do que
na ausência de acetato, porém tende a se manter constante.
0,40
• 0,35
ro o ~ o Q) .. a...-.. 0,30 (/) Cl
( w E -- Q) - "O :::> ro .__.. "O "> 0,25 ~
• 0,20
o 2 3 4
[Acetato] g/L
FIGURA 12 - Atividade específica de xilitol desidrogenase em função da concentração de acetato de sódio no meio de reação.
Estes resultados demonstram que o ácido acético presente no meio de
cultura pode aumentar tanto a atividade da enzima xilose redutase como de
xilitol desidrogenase, por este motivo não há aumento de produtividade, porque
mesmo que aumente a formação de xilitol, sua conversão em xi!ulose também
é aumentada.
Também foi observado que o ácido acético não inibi a atividade das
enzimas envolvidas no metabolismo de xilose, uma vez que em meio controle,
contendo ácido acético, mas sem xilose, foram observadas atividades iguais a
O, 1054 Ul/mg para xilose redutase e 0,0878 Ul/mg para xilitol desidrogenase.

54
6. CONCLUSÕES
Com base nos resultados obtidos é possível concluir que:
• O fato de que no meio com hicirnlisado, o consumo de é'."'ido
acético e as atividades das enzimas xilose redutase e xilitol
desidrogenase terem sido menores do que em meio sintético, pode
ser explicado pelo efeito inibitório do ácido acético ter sido
potencializado por outros compostos presentes no hidrolisado.
• O favorecimento das atividades específicas de xilose redutase e de
xilitol desidrogenase pelo ácido acético presente no meio de
cultura, pode ser comprovado pelo aumento destas in vitro
verificado na presença de acetato de sódio.
• O favorecimento das atividades de xilose redutase e de xilitol
desidrogenase pelo ácido acético poderia estar relacionado com o
fato de que a produtividade não foi aumentada, possivelmente
porque a maior formação de xilitol coincidiu com sua maior
conversão em xilulose.
• O ácido acético prejudicou a formação de células e de xilitol, tanto
em meio sintético como em hidrolisado, diminuindo o rendimento, a
produtividade e a eficiência de conversão de xilitol.

55
7. RECOMENDAÇÕES
Sugestões para trabalhos futuros:
• Estudar o metabolismo de ácido acético em Candida guilliermondii,
utilizando substrato marcado com carbono radioativo. Inclusive
medindo a atividade da enzima H+ -ATPase, para comprovar se
ocorre desvio de energia para o transporte extracelular de prótons
e a atividade de citrato liase para comprovar que o metabolismo
não ocorre via glioxilato.
• Utilizar técnicas de engenharia metabólica, como repressão por
RNA antisense, superexpressão, e disrupção de genes visando o
melhoramento genético de Candida guil/iermondii a fim de se obter
maior rendimento e produtividade de xilitol.
• Avaliar a regulação e expressão das enzimas intracelulares pelo
estudo dos promotores dos seus genes.

56
8. REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, C. L.; OETTERER, M.; MENEZES, T. J. B. Caracterização e Aplicações do xilitol na indústria alimentícia. Boletim SBCTA, v.33, n.2, p.184-193, 1999
ALAIS, C.; LINDEN, G. Food Biochemistry., New York: Ellis Horwood Limited, 1991
ALANEN, P.; ISOKANGAS, P.; GUTMANN K. Xylitol Candies in Caries Prevention: Results of a Field Study in Estonian Children. Community Dentistry and Oral Epidemiology, v.28,n.3, p.218-224, 2000
ALVES, L. A.; FELIPE, M. G. A.; SILVA, J. B. A.; SILVAS. S.; PRATA A. M. R. Pretreatment of Sugar Gane Bagasse Hemicellulose Hydrolysate for Xylitol Production by Candida guilliermondii. Applied Microbiology and Biotechnology, v. 70-72, p.89-98, 1998
BAR, A. Xylitol. in: NABORS L. B.; GELARDI, R.C. Alternative Sweeteners. 2 ed. New York: Marcel Dekker, 1991
BARBOSA, M. F. S.; MEDEIROS, M. B.; MANCILHA, 1. M.; SCHNEIDER, H.; LEE, H. Screening of Yeasts for Production of Xylitol from D-Xylose and Some Factors which Affect Xylitol Yield in Candida guilliermondii. Journal oflndustrial Microbiology, v.3, p.241-251, 1988
BIRKHED, D. Cariologic aspects of xylitol and its use in chewing gum: A review. Acta Odontologica Scandinavica, v.52, n.2, p.116-127, 1994
BRADFORD, M. M. A Rapid and Sensitive Method for the Quantification of Microgram Quantities of Protein Utilizing the Principie of Protein-Dry Binding. Analytical Biochemistry, v.72, p.248-254, 1976
BRUINENBERG, P. M.; de BOT, P. H.; van DIJKEN, J. P.; SCHEFFERS, S. A. NADH-linked Aldose Reductase: The key to Anaerobic Alcoholic Fermentation of Xylose by Yeasts. Applied Microbiology and Biotechnology, v.19, n.4, p.256-260, 1984
CANILHA, L. Aproveitamento do hidrolisado de palha de trigo para obtenção de xilitol por via microbiana. Lorena, FAENQUIL - Departamento de Biotecnologia, 2002. (Dissertação de Mestrado)
CAO, N. J.; TANG, R.; GONG, C. S.; CHEN, L. F. The Effect of Cell-Density on the Production of Xylitol from D- Xylose by Yeast. Applied Biochemistry and Biotechnology, v.45, p. 515-519, 1994

57
CCC - Calorie Contrai Council. Reduced-Calorie Sweeteners: Xylitol. [online] Disponível na Internet via WWW. URL: http://www.caloriecontrol.org/xylitol.html. Arquivo capturado em 03 de fevereiro de 2001
CHIANG, C.; KNIGHT, S.S. Metabolism of D-Xylose by Moulds. Nature, v.188, r..4744, p.78-81, 1960
CONVERTI, A.; PEREGO, P.; DOMINGUEZ, J.M. Xylitol production from hardwood hemicellulose hydrolysates by Pachysolen tannophilus, Oebaryomyces hansenii, and Candida guilliermondii. Applied Biochemistry and Biotechnology, v. 82, n. 2, p. 141-151, 1999
CONVERTI, A.; PEREGO, P.; TORRE P.; DA SILVA S.S. Mixed inhibitions by methanol, furfural and acetic acid on xylitol production by Candida guilliermondiil. Biotechnology Letters, v.22, n.23, p. 1861-1865, 2000
DAHIYA, J. S. Xylitol Production by Petromyces albertensis Grown Medium Containing D-Xylose. Canadian Journal of Microbiology, v.37 p. 14-18, 1991
DOMINGUEZ, J. M.; GONG C. S.; TSAO G. T. Production of Xylitol from D-Xylose by Debaryomyces hansenii. Applied Microbiology and Biotechnology. v.63-65, p.117-127, 1997
EMODI, A. Xylitol: lts Preoperties and Food Aplications. Food Technology, Jan, p. 28-32, 1978
FELIPE, M.G.A.; VEIRA, M.V.; VITOLO, M.; MANCILHA, I.M.; ROBERTO, C.I.; SILVA, S.S. Effect of acetic acid on xylose fermentation to xylitol by Candida guilliermondii. Journal Basic Microbiology, v. 35, n.3, p.171- 177, 1995
FELIPE, M.G.A.; ALVES, LA.; SILVA, S.S.; ROBERTO, I.C.; MANCILHA, I.M.; ALMEIDA SILVA, J.B. Fermentation of Eucaliptus Hemicellulosic Hydrolysate to Xylitol by Candida guillliermondii. Bioresource Technology, v. 56, p.281-283, 1996
FELIPE, M.G.A., VITOLO, M.; MANCILHA, 1. M.; SILVA, S. S. Environmental Parameters Affecting Xylitol Production from Sugar Cane Bagasse Hemicellulosic Hydrolysate by Candida guil/iermondii. Journal of Industrial Microbiology and Biotechnology, v.18, p.251-254, 1997a
FELIPE, M.G.A.; VITOLO, M.; MANCILHA, 1. M.; SILVA, S. S. Fermentation of Sugar Cane Bagasse Hemicellulosic Hydrolysate for Xylrto! Production: Effect of pH. Biomass and Bioenergy, v. 13, n 1-2, p.11-14, 1997b
FERRARI, M. D.; NEIROTTI, E.; ALBORNOZ, C.; SAUCEDO, E. Ethanol Production from Eucalyptus Wood Hemicelulose Hydrolysate by Pichia stiptis. Biotechnology and Bioengineering, v. 40, p.753-759, 1992

58
FERREIRA FILHO, E. X. Hemicellulases and Biotechnology Recent Research Development in Microbiology. v.2, p.165-176, 1998
FURLAN, S. A.; BOUILLOUD, P.; STREHAINO, P.; RISA, J. P. Study of Xylitol Formation from Xylose under Oxigen Limiting Conditions. Biotechnology Letters, v.13, n.3, p.203-206, 19G~
GIRIO, F. M.; PEITO, M. A.; AMARAL-COLLAÇO, M. T. Xylitol Production by Fungi 1 - An Enzymatic Test for Screening Good Xylitol Producing Fungi. ln: GRASSI, G.; GOSSE, G.; SANTOS, G. eds. Biomass for Energy and lndustry 5th E. C. Conference. Elsevier Apllied Science, v.2, p.1010-1014, 1990
GIRIO, M. F.; PELICA, F.; AMARAL-COLLAÇO, M. T. Characterization of xylitol dehidrogenase from Debaryomyces hansenii. Applied Biochemistry and Biotechnology, v. 56, p. 79-88, 1996
HAHN-HAGERDAL, B.; JEPPSSON, H.; SKOOG, K.; PRIOR, B. A. Biochemistry and Physiology of Xylose Fermentation by Yeasts. Enzyme Microbial Technology, v.16, p.933-943, 1994
HERRERO, A. A.; GOMES, R. F., SNEDECOR, B.; TOLMAN, C. J.; ROBERTS, M. F.; Growth lnibition of Clostridium thermocellum by Carboxilic Acids: A mechanism Based on Uncoupling by Weak Acids. Applied Microbiology and Biotechnology, v.22, p. 53-62, 1985
HOL YOAK, e.o.; STRATFORD, S.; MCMULLIN, Z.; COLE M.B.; CRIMMINS, K.; BROWN, A.J.P.; COOTE P.J. Activity of the plasma membrane H+ ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak acid preservative sorbic acid. Applied and Environmental Microbiology v.62 p.3158-3164, 1996
JEFFRIES, T. W. Utilization of Xylose by Bacteria, Yeasts and Fungi. Advances in Biochemical Engineering, v.27, p.1-32, 1983
KERN, M.; HALTRICH, D.; NIDETZKY, 8. KULBE, K. D. lnduction of Aldose Reductase and Xylitol Dehydrogenase Activities in Candida tenuis CBS 4435. FEMS Microbiology Letters, v.149, p.31-37, 1997
KILIAN, S. G.; van UDEN, N. Transport of xylose and glucose in the xylose-fermenting yeast Pichia stipitis. Applied Microbiology and Biotechnology, v. 27, p. 545-548, 1988
KIM, Y.; YOO, Y. Peroxidase production form carrot hairy root cell culture. Enzyme and Microbial Technology, v.18, p.535-553, 1996

59
KIM J-H, RYU Y-M. SEO J-H. Analysis and optimization of a two-substrate fermentation for xylitol production using Candida tropica/is. Journal of Industrial Microbiology and Biotechnology, v. 22, p. 181-186, 1999
KUHAD, R. C.; SINGH, A. Lignocellulose Biotechnology: Current and Future Prospects.Critical Reviews in Biotechnology, v.13, n.2, p.151-172, 1993
LAWFORD. H. G.; ROSSEAU, J. D. Effects of pH Acetíc Acíd on Glucose and Xylose Metabolísm by a Genitically Engineered Ethanolegeníc Escherichia coli. Applied Biochemistry and Biotechnology, v.39/40, p.301-332, 1993
LEE, F-J. S.: LIN, L.; SMITH J. A. Acetyl-Coa hydrolase involved in acetate utílizatíon ín Saccharomyces cerevisiae. Biochimica et Biophisica Acta, v.1297, p.105-109, 1996
LEE, H.; SOPHER, C. R., YAU, K. Y. F.; lnduction of Xylose Reductase and Xylitol Dehydrogenase Activities on Mixed Sugars in Candida guilliermondii. Journal of Chemistry Technology and Biotechnology, v.66, p.375-379, 1996
LEE, H. The structure and function of yeast xylose (aldose) reductases. Yeast, v.14, n.11, p. 977-984, 1998
LEE, J. Biological conversion of lígnocellulosic biomass to ethanol. Journal of Biotechnology, v. 56, p. 1-24, 1997
LINDEN T.; HAHN-HAGERDAL B. Fermentation of Lignocellulose Hydrolísates with Yeasts and Xylose lsomerase, Enzyme Microbiology Technology, v.11, n.9, p. 583-589, 1989
LOHMEIER-VOGEL, E. M.; SOPHER, C. R.; LEE, H. lntracellular Acidifícation as a Mechanism for the lnhibitors of Xylose Fermentatíon by Yeast. Journal of Industrial Microbiology and Biotechnology, v.20, p.75-81, 1998
LUCAS, C.; van UDEN, N. Transport of Hemicellulose Monomers in the Xylose- Fermeting Yeast Candida shehatae. Applied Microbiology and Biotechnology, v. 23 p. 491- 495, 1986
MAKINEN, K. K.; CHIEGO, D. J.; ALLEN, P.; BENNETT, C.; ISOTUPA, K. P.; TIEKSO. J.; MAKINEN, P. L. Physical, chemical and histologíc changes in dentin caries lesíons of primary teeth induced by regular use of polyol chewing gums. Acta Odontologica Scandinavica, v. 56, n. 3, p. 148-156. 1998
MANZ, U.; VANNINEN, E.; VOIROL, F. Xylitol - lts Properties and Use as a Sugar Substitute in Foods. ln: FOOD R.A. SYMP. SUGAR ANO SUGAR REPLACEMENTS. oct. 1973

60
MARCHAL, R.; ROPARS, M.; VANDECASTEELE, J. P. Conversion into Acetone and Butano! of Lignocellulosic Substrates Pretreated by Steam Explosion. Biotechnology Letters, v. 8, n. 5, p 365-370, 1986
MATIILA, P; SVANBERG, J.; PôKKA P.; KNUUTTILA, M. Dietary Xylitol Protects Against Weakening of Bone Biomechanical Properties in Ovariectomized Rats. The Joü;-;1.al of Nutrition, v.128, n.1G, p.1811-1816, 1998
MAYR, P.; BRUGGLER, K.; KULBE, K. D.; NIDETZKY,B. D-Xylose Metabolism by Candida intermedia: isolation and characterisation of two forms of aldose reductase with different coenzyme specificities. Journal of Chromatography B, v. 737, p. 195-202, 2000
MELAJA, A. J.; HAMALAINEN, L. Process for Making Xylitol. US 4.008.285, 15 fev. 1977
MORITA, T.A.; SILVA, S.S.; FELIPE, M.G.A. Effects of initial pH on biological synthesis of xylitol using xylose-rich hydrolysate. Applied Microbiology and Biotechnology, v. 84-86, p. 751-759, 2000a
MORITA, T.A.; SILVA, S.S. lnhibition of Microbial Xylitol Production by Acetic Acid and lts Relation with Fermentative Parameters. Applied Microbiology and Biotechnology, v.84-86, p.801-808, 2000b
NARENDRANATH, N. V.; THOMAS, K. C.; INGLEDEW, W. N. Effects of acetic acid and lactic acid on the growth of Saccharomyces cerevisiae in a minimal medium. Journal of Industrial Microbiology and Biotechnology, v. 26, n.3, p. 171-177, 2001
NODA, F.; HAYASHI, K.; MIZUNUMA, T.lnfluence of pH on lnhibitory Activity of Acetic Acid on Osmophilic Yeast Used in Brine Fermentation of Soy Sauce. Applied and Environmental microbiology, v.43, p.245-246, 1982
NOLLEAU, V.; PREZIOSI-BELLOY, L.; DELGENES, J. P.; DELGENES, J. M. Xylitol Production from Xylose by Two Yeasts Strains: Sugar Tolerance. Current Microbiology, v.27, p. 191-197, 1993
ODUMERO, J.A.; D'AMORE, T.; RUSSELL, I.; STEWART, G. Effects of Heat Shock and Ethanol Stress on the Viability of a Sacharomyces ovarum (carisbergensisi) Brewing Yeast Strain During Fermentation of High Gravity Wort. Journal of Industrial Microbiology, v.10, p.111-116, 1992
ONISHI, H.; SUZUKI, T. Process for Producing Xylitol by Fermentation. US 3.619.369. 09 nov. 1971
PALMQVIST, E.; HAHN-HAGERDAL, B. Fermentation of lignocellulosic hydrolysates. li: inhibitors and mechanism of inhibition. Bioresource Technology, v. 74, p. 25-33, 2000

61
PAMPULHA, M. E.: LOUREIRO-DIAS, M. C.; Activity of Glicolitic Emzymes of Saccharomyces cerevisae in presence of Acetic Acid. Applied Microbiology and Biotechnology, v. 34, p. 375-380, 1990
PANDEY, A.; SOCCOL, C.R.; NIGAM, P.; SOCCOL, V.T. Biotechnological potential of agro-industrial residues 1: sugarcane bagasse. Bioresource Technology, v. 74, n.1, p. GC 80, 2000
PARAJÓ, J. C.; DOMINGUEZ, H. DOMINGUEZ, J. M. Biotechnological Production of Xylitol. Part 1: lnterest of Xylitol and fundamentais of its biosynthesis. Bioresource Technology, v. 65, p.191-201, 1998a
PARAJÓ, J. C.; DOMINGUEZ, H. DOMINGUEZ, J. M. Biotechnological Production of Xylitol. Part 2: Operation in Culture Media made with Commercial Sugars. Bioresource Technology, v. 65, p.203-212, 1998b
PARAJÓ, J. C.; DOMINGUEZ, H. DOMINGUEZ, J. M. Biotechnological Production of Xylitol. Part 3: Operation in Culture Media made from Lignocellulose Hydrolysates. Bioresource Technology, v. 66, p.25-40, 1998c
PESSOA JR., A. Acid Hidrolysis of Hemicellulose from Sugarcane Bagasse. Brazilian Journal of Chemical Engineering, v.14, n.03, p. 291-297, 1997
PEPPER, T.; OLINGER, P.M. Xylitol in Sugar - Free Confections. Food Technology, v.42, n.1 O, 1988
PFEIRER, M. J.; SILVAS. S.; FELIPE, M. G. A. ROBERTO, 1. C. MANCILHA, 1. M. Effect of Culture Condictions on Xylitol Production by Candida guilliermondii FTI 20037. Applied Biochemistry and Biotechnology, v.57/58, p.423-430, 1996
PINTO, J.; CARDOSO, H.; LEÃO, C.; van UDEN, N. High Enthalpy na Low Enthalph Death in Saccharomyces cerevisae lnduced by Acetic Acid. Biotechnology and Bioengineering, v.33, p. 1350-1352, 1989
PREEZ J.C.; van DRIESSEL, B.; PRIOR, B. A. Effect of Aerobiosis on Fermentation and Key Enzyme Leveis during Growth of Pichia stipitis, Candida shehatae and Candida tenuis on D-Xylose. Archives of Microbiology, v. 152, p. 143-147, 1989
PREEZ J. C.; MEYER P. S.; KILIAN S. G. The effect of mixtures of acetic acid and D-xylose on the growth rate of Candida blankii. Biotechnology Letters, v. 13, n. 11, p.827-832, 1991
ROBERTO, I.C.; MANCILHA, I.M.; SOUZA, C.M.A. ; FELIPE, M.G.A.; SATO, S.;CASTRO, H.F. Evaluation of Rice Straw Hemicellulose Hydrolysate in the Production of Xylitol by Candida gui/1/iermondii, Biotechnology Letters, v.16, n.11, p. 1211-1216, 1994

62
ROBERTO. 1.C.; FELIPE M.G.A.: MANCILHA, I.M.; VITOLO M.; SATO, S.; SILVA S.S. Xylitol production by Candida guil/iermondii as an approach for the utilization of agroindustrial residues, Bioresource Technology, v. 51, p. 255-257, 1995
RODRIGUES, R. C. L. B. Avaliação das Condições de Concentração a Vácuo do Hidrolisado Hemicelulósico de Bagaço de Cana para a Produção do Xilitol. Lorena, FAENQUIL - Departamento de Biotecnologia e Química, 1999. 109 p. (Dissertação de Mestrado)
ROSA, S.M.A.; FELIPE, M.G.A.; SILVA, S.S.; VITOLO, M. Xylose Reductase Production by Candida guil/iermondii. Applied Biochemistry and Biotechnology, v. 70-72, p.127-135, 1998
SANCHES, B.; BATISTA, J. Effects of Furfural and 5-Hidroxymethylfurfural on the Fermentation of Saccharomyces cerevisae and Biomass Production from Candida guilliermondii. Enzyme Microbial and Technology, v.1 O, p. 315-318, 1988
SENE, L.; VITOLO, M.; FELIPE, M.G.A.; SILVA, S.S.; MANCILHA, I.M. Adaptation and reutilization of Candida guilliermondii Cells for Xylitol Production in Bagasse Hydrolysate. Journal of Basic Microbiology, v.38, p.61-69, 1998
SENE, L.; VITOLO, M.; FELIPE M.G.A.; SILVA, S. S. Effect of Environmental Conditions on Xylose Reductase and Xylitol Dehydriogenase Production by Candida guil/iermondii. Applied Microbiology and Biotechnology, V. 84-86, p.371-380, 2000a
SENE, L. Obtenção e cinética da xilose redutase e xilitol desidrogenase de Candida guilliermondii cultivada em hidrolisado hemicelulósico de bagaço de cana-de-açúcar. São Paulo, USP - Faculdade de Ciências Farmacêuticas, 2000b. 107 p. (Tese de Doutorado)
SILVA, D. D. V. Bioconversão de Xilose em Xilitol por Candida guilliermondii em Hidrolisado de Bagaço de Cana: Estudo do Efeito do Ácido Acético. Lorena, FAENQUIL - Departamento de Biotecnologia, 2001. 73 p. (Dissertação de Mestrado)
SILVA, S. S.; MANCILHA, 1. M.; QUEIROZ, M. A.; FELIPE M. G. A.: ROBERTO, 1. C.; VITOLO, M. Xylitol Formation by Candida guilliermondii in Media Containing Different Nitrogen Source. Journal Basic Microbiology, v. 34, p. 12-17, 1994
SILVA, S. S.; VITOLO, M.; PESSOA JUNIOR, A.; FELIPE, M. G. A. Xylose Reductase and Xylitol Dehydroqenase Activities by Xylose Fermeting Candida guilliermondii. Journal Basic Microbiology, v. 36, n.3, 1996

63
SILVA, S.S.; FELIPE M.G.A.; MANCILHA I.M. Factors that affect the biosynthesis of xylitol by xylose-fermenting yeasts - A review. Applied Biochemistry and Biotechnology, v. 70-2, p. 331-339, 1998
SLININGER, P.J.; BOLEN, O. L.; KURTZMAN, C.P. Pachysolen tannophilus: Properties and Process Consideration for Ethanol Production form D-Xylc:;e. Enzyme and Microbial Technology,v. ~. i:i. 5-15, 1987
SOUSA, M. J.; RODRIGUES, F.; CORTE-REAL, M.; LEAO, C. Mechanism underlying the transport and intracellular metabolism of acetic acid in the presence of glucose in the yeast Zygosaccharomyces bailii. Microbiology, V. 144, p. 665-670, 1998.
SOUZA, C.M.A.; ROBERTO, 1. C.; FELIPE, M. G. A.; SILVA, S. S. Influência da Temperatura na produção de Xilitol por C. guil/iermondii em Hidrolisado Hemicelulósico de Palha de Arroz. ln: CONGRESSO LA TINO- AMERICANO DE MICROBIOLOGIA, 17° SIMPÓSIO NACIONAL DE FERMENTAÇÃO, Sociedade Brasileira de Microbiologia, 1993, p.84
TAHERZADEH, M. J.; GUSTAFSSON, L.; NIKLASON, C.; LIDÉN G. Physiological effects of 5-hydroxymethyl-furfural (HMF) on Saccharomyces cerevisiae. Applied Microbiology and Biotechnology, v. 53, n. 6, p. 701- 708, 2000
TAVARES, J.M.; DUARTE, L. C.; AMARAL-COLLACO, M.T.; GIRIO, F.M. The influence of hexoses addition on the fermentation of D-xylose in Debaryomyces hansenii under continuous cultivation. Enzyme and Microbial Technology, v. 26, n. 9-10, p.743-747, 2000
TSAO, G.T. Converson of Cellulosic. Part 1. Structures of Cellulosic Material and their Hydrolisis by Enzynes. ln: ALANI, D. I.; MOO-YOUNG, M. Biotechnology and Applied Microbiology. Elsevier Applied Science Publischers, p. 205-212, 1986
TOIT, P. J.; OLIVIER, S. P.; van BILJON, P. L. Sugar Cane Bagasse as a Possible Source of Fermentable Carbohydrates. 1. Characterization of Bagasse with Regard to monosaccharide. Hemicellulose, and Amino Acid Composition. Biotechnology and Bioengineering, v. 26, p. 1071-1078, 1984
UHARI, M.; KONTIOKARI, T.; NIEMELA M. a novel Use of Xylitol Sugar in Preventin Acute Otitis Media. Pediatrics, v.102, n.4, p.879-884, 1998
van EYS, J.; WANG, Y.; CHAN, S.; TANPHAICHITR, V.S. Xylitol as a Therapeutic Agent on Glucose-6-Phosphate Dehydrogenase Deficiency. ln: Sipple, H.L., McNutt, K.W. Sugar in Nutrition, Academic Press, p.613, 1974

64
VANDESKA. E.; KUZMANOVA, S.; JEFFRIES, T. W. Xylitol Formation and Key Enzyme Activities in Candida boidinii under different Oxygen Transfer Rates, Journal of Fermentation and Bioengineering, v.89, n. 8, p. 513- 516, 1995.
VANDESKA E.; AMARTEY, S.; KUZMANOVA, S; JEFFRIES, T. W. Fed-Batch Culture for Xylitol Production by Candida boidinii, ?rocess Biochemistry, v.31, n.3, p. 265-270, 1996
VERDUYN, C.; FRANK, J.; van DIJKEN, J. P.; SCHEFFERS, A. Multiple forms of Xylose Reductase in Pachysolen tannophilus CBS4044, FEMS Microbiology, v.30, p. 313-317, 1985a
VERDUYN, C.; VAN KLEEF, R.; FRANK, J.; SCHEREUDER, H.; van DIJKEN, J. P.; SCHEFFERS, W. A. Properties of the NAD(P)H-Dependent Xylose Reductase from the Xylose-Fermeting Yeast Pichia stipitis. Biochemical Journal, v.226, p. 667-669, 1985b
WAITZBERG, D. L. Nutrição enteral e parenteral na prática clínica. 2ed. Rio de Janeiro: Editora Atheneu, 1995
WALKER G. M. Yeast Phisiology and Biotechnology. Chichester: John Wiley & Sons Ltd, 1999
WALTHER, T.; HENSIRISAK P.; AGBLEVOR, F. A. The influence of aeration and hemicellulosic sugars on xylitol production by Candida tropicalis. Bioresource Technology, v.76, p. 213-220, 2001
WATSON, N. E., PRIOR, B. A., du PREEZ, J. C.; LATEGAN, P. M. Oxigen Requeriments for the Xylose Fermentation to Ethanol and Polyol by Pachysolen tannophilus. Enzyme and Microbial Technology, v. 6, p. 447- 450, 1984
WEBB, S. R.; LEE, H. Regulation of Xylose Utilization by Hexases in Pentose- Fermeting Yeast. Biotechnology Advances, v.8, p. 685-697, 1990
WINKELHAUSEN, E.; PITIMAN, P.; KUZMANOVA, S.; JEFFRIES, T.W. Xylitol Formation by Candida boidinii in Oxygen Limited Chemostat Culture. Biotechnology Letters, v.18, n.7, p.753-758, 1996
WINKELHAUSEN, E.; KUZMANOVA S. Microbial Conversion of D- Xylose to Xylitol. Journal of Fermentation and Bioengineering, v.86, n.1, p. 1-14, 1998
YAHASHI, Y.; HATSU, M.; HORITSU, H.; KAWAI, K.; SUZUKI, T.; TAKAMIZAWA. K. D-Glucose Feeding for lmprovement of Xylitol Productivity from D-Xylose Using Candida tropicalis lmmobilized on a Non- Woven Fabric Biotechnology Letters, v.18, p.1395-1400, 1996

65
YOKOYAMA, S.; SUZUKI, T.; KAWAI, K.; HORITSU, H.; TAKAMIZAWA, K. Purification, Characterization and Structure Analysis of NADPH-Dependent D-Xylose Reductases from Candida tropicalis. Journal of fermentation and Bioengineering, v. 79, p. 217-223,1995
ZABNER, J.; SEILER M.P.; LAUNSPACH J.L.; KARP P.H.; KEARNEY W.R.; LOOK D.C., SMITH J.J.; WELSH M.J. The osrnolyte xylitol reduces the salt concentration of airway surface liquid and may enhance bacterial killing. Proceedings of The National Academy of Sciences of The USA, v.97, n.21, p. 11614-11619, 2000
ZYL, C. van, PRIOR, B.A. du PREEZ, J. C. Production of Ethanol from Sugar Cane Bagasse Hemicellulose Hydrolysate by Pichia stipitis. Applied Biochemistry and Biotechnology, v.17, p. 357-369, 1988