
Universidade Federal de Goiás
Instituto de Ciências Biológicas
Lília Cristina de Souza Barbosa
MORFO-ANATOMIA E FITOQUÍMICA DE
Cymbopogon densiflorus (Steud.) Stapf e Cymbopogon nardus (L.)
Rendle (Poaceae: Panicoideae)
Goiânia

2007Lília Cristina de Souza Barbosa
MORFO-ANATOMIA E FITOQUÍMICA DE Cymbopogon densiflorus (Steud.)
Stapf e Cymbopogon nardus (L.) Rendle (Poaceae: Panicoideae)
Dissertação apresentada ao Programa de Pós-
graduação em Biologia – Mestrado – do Instituto
de Ciências Biológicas, da Universidade Federal
de Goiás, como requisito necessário à obtenção do
título de Mestre em Biologia.
Orientadora: Profa. Dra. Maria Helena Rezende
Co-orientador: Prof. Dr. José Realino de Paula

Goiânia
2007


Dados Internacionais de Catalogação-na-Publicação (CIP)
(GPT/BC/UFG)
Barbosa, Lília Cristina de Souza.B238m Morfo-anatomia e fitoquímica de Cymbopogon densiflorus
(Steud.) Stapf e Cymbopogon nardus (L.) Rendle (Poaceae: Panicoideae) [manuscrito] / Lília Cristina de Souza Barbosa. – 2007.
113 f. : il. ; figs., tabs.
Orientadora: Profa. Dra. Maria Helena Rezende. Co-Orientador: Prof. Dr. José Realino de Paula.
Dissertação (Mestrado) – Universidade Federal de Goiás, Instituto de Ciências Biológicas, 2007.
Bibliografia. Inclui lista de ilustrações, tabelas e gráficos.
1. Plantas medicinais 2. Essências e óleos essenciais 3. Cymbopogon densiflorus (Steud.) Stapf 4. Cymbopogon nardus (L.) Rendle 5. Fitoterapia I. Rezende, Maria Helena II. Paula, José Realino III. Universidade Federal de Goiás, Instituto de Ciências Biológicas. IV. Título.
CDU: 615.89:633.2

DEDICO...
A Deus, que me tornou uma pessoa privilegiada e capacitada para adquirir
aprendizados.
OFEREÇO...
À todas as pessoas que me ajudaram a tornar possível a realização deste trabalho.
HOMENAGEIO...

Aos meus pais e meu irmão, que sempre me incentivaram, me dando apoio, amor e
carinho sem cessar.
AGRADECIMENTOS
À Professora Dra. Maria Helena Rezende, pela paciência e dedicação, em
orientar essa dissertação.
Ao Professor Dr. José Realino de Paula, pela co-orientação e grandes
contribuições fornecidas a este trabalho.
À Professora Dra. Dalva Graciano-Ribeiro, da Universidade de Brasília (UnB)
pelas preciosas dicas e materiais literários, participação, atenção, colaboração com essa
dissertação.
Ao Professor Dr. Tarciso Sousa Filgueiras, pesquisador da Reserva Ecológica do
IBGE – RECOR, Brasília-DF, especialista em Poaceae, pela contribuição ao identificar
as espécies vegetais estudadas neste trabalho e por disponibilizar materiais literários que
enriqueceram e deram subsídios a esta pesquisa.
À Professora Dra. Simone Maria Teixeira Sabóia-Morais, do Laboratório de
Comportamento Celular/Laboratório de Neurobiologia, do Instituto de Ciências
Biológicas/UFG e à técnica Rosana Falcão, da Embrapa Recursos Genéticos e
Biotecnologia - CENARGEN, Brasília-DF - pela importante colaboração na
metodologia que foram importantes para realizar a Microscopia Eletrônica de
Varredura.
Ao professor Dr. Pedro Henrique Ferri e seus estagiários, do Laboratório de
Química de Produtos Naturais, do Instituto de Química/UFG, pela colaboração na
Cromatografia Gasosa e Espectrometria de Massa dos óleos essenciais.
Ao professor MSc. Heleno Dias Ferreira, do Departamento de Biologia
Geral/UFG, pelo auxílio na descrição morfológica e sugestões importantes fornecidas a
este trabalho.
À Professora Dra. Alessandra Feijó M. Viu, do Centro de Ciências Agrárias e
Biológicas, Campus Avançado de Jataí, UFG, pela atenção e empréstimo de recursos
que permitiram o desenvolvimento deste trabalho.
À Professora Dra. Maria Bernadete Gonçalves Martins, da UNESP de São
Vicente-SP, pela atenção e material literário que enriqueceu esta pesquisa.

Aos colegas Maria Tereza Faria, Leonice Faustino Manrique Tresvenzol, Joelma
Abadia Marciano de Paula, Darlene Ana de Paula Vieira, Fernanda Pimenta Simon
Ferreira, Alexandre Pereira Santos, Daniel Teles Zatta e Edson Ferreira Duarte e às
estagiárias Letícia Domingos e Dayana Abdalla, pelos grandes auxílios na metodologia
desta pesquisa e pela amizade.
Aos técnicos Eli Noronha, do Laboratório de Anatomia Vegetal, da
Universidade de Brasília, pelas preciosas dicas que contribuíram para a execução deste
trabalho.
À colega Sílvia Fernandes pela boa vontade, paciência e importante auxílio na
metodologia, que foi imprescindível para a realização da Microscopia Eletrônica de
Varredura, na Universidade de Brasília.
À MSc. Carina Verna, Tecnóloga Oftálmica da Escola Paulista de Medicina, São
Paulo-SP, pelo auxílio na edição das fotos, nas análises estatísticas e também, pela
enorme amizade.
À Coordenadoria de Apoio Pessoal ao Ensino Superior - CAPES - pelo apoio
financeiro concedido por meio da bolsa.
A todos que de forma direta e indiretamente contribuíram para tornar possível a
execução deste trabalho.

“Ninguém é tão ignorante que não tenha algo a ensinar; e ninguém é tão sábio que não tenha algo a aprender.”
Blaise Pascal (Filósofo, físico, matemático e escritor francês)
“O que importa na vida não é o ponto de partida, mas a caminhada. Caminhando e semeando, no fim terás o que colher!”

Cora Coralina (Poetisa goiana)

SUMÁRIO
RESUMO ................................................................................................................................. 11ABSTRACT .............................................................................................................................. 12
INTRODUÇÃO GERAL1 INTRODUÇÃO ................................................................................................................... 141.1 Referências Bibliográficas ................................................................................................. 182 OBJETIVOS .......................................................................................................................... 222.1 Objetivo Geral .................................................................................................................... 222.2 Objetivos Específicos ........................................................................................................ 22
CAPÍTULO 1 – Morfo-anatomia das Folhas e Colmos de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)1 INTRODUÇÃO ................................................................................................................... 252 MATERIAL E MÉTODOS ............................................................................................... 272.1 Material Botânico ............................................................................................................... 272.2 Caracterização Morfológica .............................................................................................. 272.3 Caracterização Anatômica ................................................................................................ 272.3.1 Microscopia Óptica ......................................................................................................... 272.3.2 Microscopia Eletrônica de Varredura .......................................................................... 293 RESULTADOS ..................................................................................................................... 313.1 Caracterização Morfológica .............................................................................................. 313.1.1 Cymbopogon densiflorus (Steud.) Stapf .............................................................................. 313.1.2 Cymbopogon nardus (L.) Rendle ........................................................................................ 31

3.2 Caracterização Anatômica ................................................................................................ 323.2.1 Cymbopogon densiflorus (Steud.) Stapf .............................................................................. 323.2.1.1 Lâmina Foliar ............................................................................................................... 323.2.1.2 Bainha Foliar ................................................................................................................ 343.2.1.3 Colmo ............................................................................................................................
343.2.2 Cymbopogon nardus (L.) Rendle ........................................................................................ 363.2.2.1 Lâmina Foliar ............................................................................................................... 363.2.2.2 Bainha Foliar ................................................................................................................ 383.2.2.3 Colmo ............................................................................................................................
383.3 Densidade Estomática ....................................................................................................... 414 DISCUSSÃO ......................................................................................................................... 425 CONCLUSÃO ..................................................................................................................... 476 REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................
48 7 ILUSTRAÇÕES ................................................................................................................... 54
CAPÍTULO 2 – Determinação dos Teores de Umidade e Cinzas e Prospecção Fitoquímica das Folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)1 INTRODUÇÃO ................................................................................................................... 67 2 MATERIAL E MÉTODOS ............................................................................................... 69

2.1 Material Botânico ............................................................................................................... 692.2 Determinação do Teor de Umidade ................................................................................ 692.3 Determinação do Teor de Cinzas .................................................................................... 702.3.1 Determinação do Teor de Cinzas Totais ..................................................................... 702.3.2 Determinação do Teor de Cinzas Insolúveis em Ácido Clorídrico (HCl) .............
702.4 Prospecção Fitoquímica .................................................................................................... 712.4.1 Pesquisa de Heterosídeos Antraquinônicos ...............................................................
712.4.2 Pesquisa de Heterosídeos Cardioativos ................................................................... 722.4.3 Pesquisa de Heterosídeos Flavonóides ....................................................................... 732.4.4 Pesquisa de Heterosídeos Saponínicos ........................................................................ 742.4.5 Pesquisa de Taninos ....................................................................................................... 752.4.6 Pesquisa de Alcalóides .................................................................................................... 762.4.7 Pesquisa de Cumarinas ................................................................................................... 773 RESULTADOS ..................................................................................................................... 783.1 Teores de Umidade, Cinzas Totais e Cinzas Insolúveis em Ácido Clorídrico(HCl) .......................................................................................................................................... 78 3.2 Prospecção Fitoquímica ................................................................................................... 794 DISCUSSÃO ........................................................................................................................ 815 CONCLUSÃO ....................................................................................................................
85REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................ 86
CAPÍTULO 3 – Análise do Óleo Essencial Extraído das Folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)1 INTRODUÇÃO ................................................................................................................... 92

2 MATERIAL E MÉTODOS ............................................................................................... 942.1 Material Botânico ............................................................................................................... 942.2 Extração dos Óleos Essenciais ........................................................................................ 942.3 Análise da Composição Química dos Óleos Essenciais ..............................................
953 RESULTADOS .................................................................................................................... 963.1 Rendimento dos Óleos Essenciais .................................................................................. 963.2 Análise da Composição Química dos Óleos Essenciais .......................................... 973.2.1 Análise da Composição Química dos Óleos Essenciais de Cymbopogondensiflorus (Steud.) Stapf ............................................................................................................ 973.2.2 Análise da Composição Química dos Óleos Essenciais de Cymbopogon nardus (L.) Rendle .................................................................................................................................
101
4 DISCUSSÃO ......................................................................................................................... 1045 CONCLUSÃO ...................................................................................................................... 108REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................ 109
LISTA DE ILUSTRAÇÕES
CAPÍTULO 1 – Estudo Morfo-anatômico das Folhas e Colmos de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)Figura 1 Aspectos morfológicos de C. densiflorus ............................................................... 55

Figura 2 Aspectos morfológicos de C. nardus ................................................................... 56Figuras 3-8 C. densiflorus - lâmina foliar ............................................................................... 57Figuras 9-12 C. densiflorus - lâmina foliar, em microscopia eletrônica de varredura .....
58Figuras 13-18 C. densiflorus - lâmina foliar ............................................................................ 59Figuras 19-23 C. densiflorus - bainha foliar ........................................................................... 60Figuras 24-28 C. densiflorus - colmo ...................................................................................... 61Figuras 29-34 C. nardus - lâmina foliar ................................................................................ 62Figuras 35-38 C. nardus - lâmina foliar ............................................................................... 63Figuras 39-42 C. nardus - bainha foliar ................................................................................ 64Figuras 43-48 C. nardus - colmo ........................................................................................... 65
CAPÍTULO 3 – Análise do Óleo Essencial Extraído das Folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)Figura 1 Cromatograma do óleo essencial da amostra 1 de C. densiflorus .......................
100Figura 2 Cromatograma do óleo essencial da amostra 2 de C. densiflorus ......................
100Figura 3 Cromatograma do óleo essencial da amostra 1 de C. nardus ............................
103Figura 4 Cromatograma do óleo essencial da amostra 2 de C. nardus ............................
103

LISTA DE TABELAS E GRÁFICOS
CAPÍTULO 1 – Morfo-anatomia das Folhas e Colmos de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)Tabela 1 Caracteres anatômicos diferenciais entre C. densiflorus e C. nardus ................... 40Tabela 2 Média do número de estômatos por mm² da lâmina foliardas amostras 1 e 2 de C. densiflorus ........................................................................................ 41Tabela 3 Média do número de estômatos por mm² da lâmina foliardas amostras 1 e 2 de C. nardus ............................................................................................... 41
CAPÍTULO 2 – Determinação dos Teores de Umidade e Cinzas e Prospecção Fitoquímica das Folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)Tabela 1 Teores de umidade, cinzas totais e cinzas insolúveis em ácido clorídricodas folhas de C. densiflorus e C. nardus, expressos em porcentagem (p/p) ....................... 78Tabela 2 Principais classes de metabólitos secundários detectados nas folhas de C. densiflorus...................................................................................................................................... 79Tabela 3 Principais classes de metabólitos secundários detectados nas folhas de C. nardus........................................................................................................................................... 79
CAPÍTULO 3 – Análise do Óleo Essencial Extraído das Folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae: Panicoideae)Tabela 1 Teor de óleo essencial em porcentagem (V/p) das folhas deC. densiflorus coletadas em duas localidades diferentes de Jataí-GO .................................. 96Tabela 2 Teor de óleo essencial em porcentagem (V/p) das folhas de C. narduscoletadas na Reserva Ecológica do IBGE-RECOR, Brasília-DF (amostra 1) eHorto de Plantas Medicinais da Faculdade de Farmácia - UFG, Goiânia-GO

(amostra 2) .................................................................................................................................
96Tabela 3 Componentes do óleo essencial das folhas de C. densiflorus ............................. 98Gráfico 1 Classes dos constituintes do óleo essencial das folhas deC. densiflorus (Steud.) Stapf, em porcentagem ....................................................................... 99Tabela 4 Constituintes do óleo essencial de C. densiflorus e suasrespectivas classes .................................................................................................................... 99Tabela 5 Componentes do óleo essencial das folhas de C. nardus ..................................
101Gráfico 2 Classes dos constituintes do óleo essencial das folhas de C. nardus,em porcentagem .......................................................................................................................
102Tabela 6. Constituintes do óleo essencial de C. nardus (L.) Rendle e suas respectivas classes ..................................................................................................................... 102

RESUMO
O gênero Cymbopogon Sprengel pertence à família Poaceae e compreende 40 espécies, distribuídas pela África Tropical e Subtropical, Ásia e Austrália, embora algumas foram introduzidas na América. Diversas espécies deste gênero são cultivadas para a extração de óleos essenciais, a partir de suas folhas, sendo então de grande importância medicinal, alimentar e industrial. As espécies em estudo Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle, são originárias da África e Ásia, respectivamente. Esta pesquisa teve por objetivo, ampliar o conhecimento sobre as espécies C. densiflorus e C. nardus, por meio do estudo morfo-anatômico das folhas e colmo, prospecção fitoquímica e análise dos óleos essenciais das folhas. Estudos anatômicos têm sido de relevante importância para pesquisas farmacognósticas; principalmente para a identificação de diversas matérias-prima vegetal, que muitas vezes são conhecidas pelo mesmo nome popular ou então, comercializadas com contaminantes ou de outras partes da mesma espécie. Por meio das análises anatômicas verificaram-se caracteres comuns entre ambas as espécies, tais como lâminas e bainhas foliares anfiestomáticas, estômatos com células-guarda halteriformes e células subsidiárias em forma de cúpula, raros na face adaxial e abundantes na face abaxial predominando nas regiões intercostais. As faces adaxial e abaxial apresentam células longas e células curtas: suberosas e silicificadas halteriforme e cruciforme, sendo as últimas localizadas nas regiões costais; macrotricomas e microtricomas predominam na face abaxial. Nas lâminas foliares, as células buliformes estão presentes na face adaxial da epiderme, intercaladas por fibras na região costal e o mesofilo é homogêneo disposto radialmente aos feixes vasculares, com distância intervenal de uma a três células, caracterizando anatomia Kranz. Os feixes vasculares colaterais, de 1ª, 2ª e 3ª ordens com bainha de feixe única. A região do bordo é formada por fibras e a epiderme apresenta células silicificadas. Entretanto, ocorrem características anatômicas distintas entre as duas espécies, como formato da nervura central, nas lâminas foliares; C. densiflorus apresenta células parenquimáticas incolores, na bainha foliar, que estão ausentes em C. nardus e o número de elementos de metaxilema em cada lado dos elementos de protoxilema: 1, em C. densiflorus e 2 a 3, em C. nardus e presença de cilindro esclerenquimático e medula fistulosa em C. nardus, caracteres ausentes em C. densiflorus. A análise fitoquímica preliminar das folhas de C. densiflorus e C. nardus evidenciou flavonóides, saponinas, cumarinas e traços de heterosídeos cardioativos. Na análise dos óleos essenciais, as folhas de C. densiflorus apresentaram trans-p-menta-1(7),8-dien-2-ol, trans-p-menta-2-8-dien-1-ol, cis-carveol e cis-p-menta-2-8-dien-1-ol, como constituintes majoritários; enquanto que em C. nardus foram geraniol, citronelol e citronelal. Os caracteres anatômicos observados podem ser importantes para as determinações taxonômicas das espécies estudadas. Por meio dos resultados encontrados, verifica-se o potencial fitoterápico de ambas as espécies. Futuras pesquisas em isolamento e purificação dos metabólitos secundários, análises farmacológicas e toxicológicas dos extratos e dos óleos essenciais, serão importantes para assegurar a eficácia terapêutica destas.
Palavras-chave: plantas medicinais, morfo-anatomia; flavonóides, óleos essenciais, fitoterápicos.

ABSTRACT
The genus Cymbopogon Sprengel belong to the Poaceae family and it has 40 species distributed in Tropical and Subtropical Africa, Asia and Australia, although some species went introduced in America. Many species of this genus are cultivated for the extraction of essential oil, from their leaves, with large medicinal, food and industrial importance. The species in focus, Cymbopogon densiflorus (Steud.) Stapf and C. nardus (L.) Rendle are originated from Africa and Asia, respectively. This research had as objective, to broaden the knowledge about the species C. densiflorus and C. nardus, by the morphological and anatomy studies from leaves and culms, phytochemical analysis and essential oil analysis from the leaves. Anatomical studies have been of relevant importance to the pharmacognosy researches, mainly for the identification of many vegetal raw materials. Several times, these raw materials are known by the same popular name or then, they are commercialized with contaminated agents or with other parts of the specie. Through of anatomical analysis, it was checked commons characters, such as leaf lamina and sheath amphistomata, stomatas with guard cells dumbbell and subsidiary cells dome-shape, rares in adaxial surface and abundant in abaxial surface, predominated in intercostal zones. The adaxial and abaxial surfaces had long cells and short cells: cork and dumb-bell and cross-shaped silica cells, these last it is placed in costal zones; macro-hairs and micro-hairs abundant in abaxial surface. In the leaf lamina, bulliforms cells are presents in adaxial surface, they were alternated with fibers in the costal zones and the mesophyll is homogeneous with chlorenchyma radiated to the bundle sheaths and arm cells with walls invaginated that they determined the intervenal distance by one or three cells, characterized Kranz anatomy. Bundle sheaths collateral, of 1st, 2nd and 3rd orders with single vascular bundle sheaths. The cap region is constituted by sclerenchyma and the epidermis has silica cells. However, both species had different anatomical features, as the form of midrib, in the leaf laminas; C. densiflorus showed colourless parenchyma cells in the mesophyll of leaf sheaths, that they do not exist in C. nardus. In the culms, numbers of metaxylem vessels in the each side of protoxylem vessels in vascular bundles: 1, in C. densiflorus, 2 and 3, in C. nardus; and the presence of sclerenchyma cylinder and fistula in C. nardus, absent characters in C. densiflorus. Moreover, in C. densiflorus, while C. nardus showed these characters. The preliminary phytochemistry analysis C. densiflorus and C. nardus leaves evidencied flavonoids, saponins, coumarins and traces of cardioactive glycosides. In the essential oil analysis, C. densiflorus leaves showed trans-p-mentha-1(7),8-dien-2-ol, trans-p-mentha-2-8-dien-1-ol, cis-carveol and cis-p-mentha-2-8-dien-1-ol as majority constituents; while C. nardus leaves had geraniol, citronellol and citronellal. The anatomical characters observed can be important to the taxonomic determinations of species studied, in the genus. Through the results found, it verifies the phytotherapics potential of both species. Future researches in isolation and purify of the secondary metabolites, pharmacologics and toxicologics analysis of extracts and of the essential oil, it will be important to assure the therapeutic efficiency of these.
Keywords: medicinal plants, morphology and anatomy, flavonoids, essential oil, phytotherapics.
14

____________________________________________________________
INTRODUÇÃO GERAL
15

1 INTRODUÇÃO
O gênero Cymbopogon Sprengel vem do grego kymbe (bote, navio) e pogon
(barba), em referência ao formato das inflorescências. Pertence à família Poaceae
(subfamília Panicoideae) que apresenta cerca de 700 gêneros e 12.000 espécies
(SOENARKO, 1977; WATSON e DALLWITZ, 2005), sendo uma das principais
famílias de Angiospermas, do ponto de vista econômico, em número de espécies
utilizadas e importantes ao homem (JOLY, 2002; SOUZA & LORENZI, 2005). Esse
gênero está representado com aproximadamente 40 espécies, distribuídas pela África
Tropical e Subtropical, Ásia e Austrália. Compreende espécies perenes ou raramente
anuais, cespitosas e aromáticas. Colmos frequentemente eretos, que medem de 30 cm a
3 metros de altura. Lâminas foliares lineares (largas a filiformes), lígula membranosa,
cartácea ou membrano-cartácea, glabra. Inflorescências em espiguetas densas dispostas
em panículas. Diversas espécies são cultivadas para a extração de óleos essenciais a
partir de suas folhas, de grande importância medicinal, alimentar e industrial
(SOENARKO, 1977; NATH et al., 2002; WATSON e DALLWITZ, 2005; CHEN e
PHILLIPS, 2006).
As espécies do gênero Cymbopogon encontradas no Brasil foram introduzidas e
aclimatadas durante o período colonial, quanto posteriormente, devido às diversas
imigrações ocorridas ao decorrer da história brasileira (FILGUEIRAS, 2005). Dentre as
espécies que foram introduzidas no Brasil, pode-se citar: C. citratus (DC) Stapf., C.
densiflorus (Steud.) Stapf e C. nardus (L.) Rendle, popularmente conhecidas como
capim-caboclo e capim-citronela, respectivamente. Segundo MATOS (2002), no Brasil,
C. citratus produz raras flores e estéreis e desta forma se reproduz por perfilhação.
As espécies em estudo, Cymbopogon densiflorus encontrada no Zaire, Zâmbia,
Rodésia, Gabão foi introduzida e aclimatada no Brasil (SOENARKO, 1977;
RENVOIZE, 1984; SILVA, 2000), enquanto que C. nardus encontrada na Índia e no
Ceilão e foi introduzida nos trópicos (SOENARKO, 1977).
Segundo SOENARKO (1977), C. densiflorus é usada como um tônico e
estíptico pelos nativos na África e o óleo de C. nardus é utilizado na indústria para a
fabricação de sabões, desinfetantes e repelentes de insetos. De acordo com MARTINS
et al. (1995), C. densiflorus é indicada para tosse, bronquite, rouquidão e afecções
16

catarrais, sendo usada popularmente em banhos e defumações, enquanto que C. nardus
se assemelha ao C. citratus, mas com aroma que lembra o Eucalyptus citriodora Hook.
(eucalipto citriodora), em razão do alto teor de geraniol e citronelal em seu óleo
essencial, sendo utilizada na fabricação de perfumes e cosméticos, e também como
repelente de insetos. TAKAISI-KIKUNI et al. (1996) reportam que essa espécie é
popularmente usada no Congo contra várias doenças, tais como: asma, febre, resfriado,
epilepsia, câimbras e dores abdominais, bem como na culinária e fabricação de
perfumes. TAKAISI-KIKUNI et al. (2000) relataram que o óleo essencial de C.
densiflorus mostrou um amplo espectro de atividade contra bactérias Gram-negativas e
Gram-positivas, apresentando maior sensibilidade nestas últimas. KANKO et al. (2004)
estudaram o óleo essencial das folhas de C. nardus e verificaram a presença de
geranial, mirceno e limoneno. SILVA (2007) verificou em um levantamento
etnobotânico, que as folhas de C. densiflorus são popularmente utilizadas para o
tratamento de constipação intestinal e também, em banhos de descarrego contra a inveja
e mau-olhado em rituais de Umbanda.
Os vegetais fazem parte da vida do homem desde seus primórdios como fonte
de alimentos, de materiais para o vestuário, habitação, utilidades domésticas, defesa e
ataque, na produção de meios de transporte, como utensílios para manifestações
artísticas, culturais e religiosas e como meio restaurador da saúde (SCHENKEL et al.,
2003). Historicamente, o homem utiliza recursos naturais, para diversos fins,
principalmente alimentício e medicinal (VILA VERDE et al., 2003). O uso dos vegetais
como alimento, medicamento ou cosmético, se perde na história do homem na face da
Terra (TESKE e TRENTINI, 2001) e tem-se difundido largamente nos últimos anos
(ALMEIDA et al., 2002).
As plantas têm tido uma longa história de utilização na medicina popular
(BARNES e PRASAIN, 2005), sendo o resultado do acúmulo secular de
conhecimentos empíricos sobre a ação dos vegetais, por diversos grupos étnicos
(SIMÕES et al., 1989). No Brasil, a utilização de plantas no tratamento de doenças
apresenta, fundamentalmente, influências da cultura indígena, africana e, naturalmente,
européia (MARTINS et al., 1995), ocasionando em uma diversidade de conhecimentos
populares a respeito do uso medicinal das plantas.
Os recursos da biodiversidade são fundamentais para o desenvolvimento
econômico, social e cultural das sociedades humanas (FONSECA-KRUEL e
PEIXOTO, 2004) e a partir de uma perspectiva histórica, a produção de medicamentos
17

e o tratamento farmacológico de doenças começaram com o uso de plantas medicinais
(SCHULZ et al., 2002).
As plantas medicinais têm sido utilizadas nos países em desenvolvimento como
tratamentos alternativos para problemas de saúde (DUARTE et al., 2005). A fitoterapia
vem sendo aprimorada em países em desenvolvimento não somente como um caminho
para resgatar tradições antigas, mas também como uma solução alternativa para
problemas de saúde (MARTÍNEZ et al., 1996). Em comparação com as preparações
convencionais, os produtos fitoterápicos utilizados na medicina popular podem
apresentar alguns problemas relacionados ao aspecto qualidade. Isso ocorre por causa
da natureza das plantas, formadas por misturas complexas de compostos químicos que
podem variar consideravelmente dependendo dos fatores ambientais e genéticos
(NEWALL et al., 2002). Os metabólitos secundários das plantas são influenciados por
três fatores principais: hereditários (composição genética), ontogênicos (estádio de
desenvolvimento) e ambientais (ROBBERS et al., 1997).
O conhecimento sobre plantas medicinais simboliza muitas vezes o único
recurso terapêutico de muitas comunidades e grupos étnicos. Nas regiões pobres do
Brasil e até mesmo nas grandes cidades brasileiras, plantas medicinais são
comercializadas em feiras livres, mercados populares e cultivadas em quintais
residenciais (MACIEL et al., 2002).
Em diferentes países, a medicina popular através de plantas é amplamente
praticada por raizeiros e pequenos ervanários, e apresenta-se em franca expansão. No
outro extremo, encontra-se o usuário que se adapta ao mercado de acordo com sua
situação sócio-econômica, mas com interesse em solucionar suas necessidades primárias
de saúde (NUNES et al., 2003). Muitas destas plantas são comercializadas por raizeiros
que desconhecem quaisquer procedimentos que mantenham as propriedades
farmacológicas das plantas. Assim, percebe-se a importância da realização de estudos
que visam o controle da qualidade de tais plantas. Entende-se por qualidade o conjunto
de critérios que caracterizam a matéria-prima para o uso ao qual se destina. Portanto, a
qualidade da matéria-prima vegetal é a determinante inicial da qualidade do fitoterápico
(FARIAS, 2003). A eficácia e a segurança terapêutica de espécies vegetais dependem da
qualidade, sofrendo influência de diversos fatores extrínsecos e intrínsecos, exigindo a
obediência às condições ideais de cultura, colheita, secagem, estabilização, manufatura,
conservação e armazenamento (AMARAL et al., 2003).
18

Dentro desta perspectiva, neste trabalho elegeram-se as espécies C. densiflorus
(Steud.) Stapf e C. nardus (L.) Rendle como objeto de estudo. Visou-se a pesquisa
morfo-anatômica e fitoquímica para fins de controle de qualidade de amostras de folhas
e colmos dessas espécies.
19

1.1 REFERÊNCIAS BIBLIOGRÁFICAS(*)
ALMEIDA, M.M.B.; LOPES, M.F.G.; NOGUEIRA, C.M.D.; MAGALHÃES, C.E.C.;
MORAIS, N.M.T. Determinação de nutrientes minerais em plantas medicinais. Ciência
em Tecnologia de Alimentos. 22(1): 94-97. 2002.
AMARAL, F.M.F.; COUTINHO, D.F.; RIBEIRO, M.N.S.; OLIVEIRA, M.A.
Avaliação da qualidade de drogas vegetais comercializadas em São Luís/Maranhão.
Revista Brasileira de Farmacognosia. 13(supl.): 27-30. 2003.
BARNES, S; PRASAIN, J. Current Progress in the use of traditional medicines and
nutraceuticals. Current Opinion in Plant Biology. 8: 324-328. 2005.
CHEN, S.; PHILLIPS, S. M. Cymbopogon Sprengel. Flora of China. 22: 624-631.
2006.
DUARTE, M.C.T.; FIGUEIRA, G.M.; SARTORATTO, A. REHDER, V.G.;
DELARMELINA, C. Anti-Candida activity of Brazilian medicinal plants. Journal of
Ethnopharmacology. 97: 305-311. 2005.
FARIAS, M.R. Avaliação da qualidade de matérias-primas vegetais. In: SIMÕES,
C.M.O.; SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.;
PETROVICK, P. R. (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS. p. 263-288. 2003.
FILGUEIRAS, T.S. Asiáticas no Brasil: Gramíneas (Poaceae) Introduzidas da Ásia.
Eugeniana. 28: 3-18. 2005.
FONSECA-KRUEL, V.S.; PEIXOTO, A.L. Etnobotânica na Reserva Extrativista
Marinha de Arraial do Cabo, RJ, Brasil. Acta Botânica Brasílica. 18(1): 177-190.
2004.
JOLY, A.B. Botânica: introdução à taxonomia vegetal. 13ª ed. São Paulo:
Companhia Editora Nacional. 2002.
______________________________________________________________________

(*) Referências bibliográficas conforme ABNT/NBR-6023: 2002.

KANKO, C.; SAWALIHO, B.E.H.; KONE, S.; KONKOUA, G.; GUESSAN, Y.T.N.
Étude dês proprietés physico-chimiques des huiles essentielles de Lippia multiflora,
Cymbopogon citratus, Cymbopogon nardus e Cymbopogon giganteus. Comptes
Rendus Chimie. 7: 1039-1042. 2004.
MACIEL, M.A.M.; PINTO, A.C.; VEIGA JR, V.F.; GRYNBERG, N. F.;
ECHEVARRIA, A. Plantas medicinais: a necessidade de estudos multidisciplinares.
Química Nova. 25(3): 429-438. 2002.
MARTÍNEZ, M.J.; BETANCOURT, J. ALONZO-GONZÁLEZ, N.; JAUREGUI, A.
Screening of some Cuban medicinal plants for antimicrobial activity. Journal of
Ethnopharmacology. 52: 171-174. 1996.
MARTINS, E.R.; CASTRO, D.M.; CASTELLANI, D.C.; DIAS, J.E. Plantas
medicinais. 1ª ed (Reimpressão). Viçosa: UFV. 1995.
MATOS, F. J. A. Farmácias Vivas. Fortaleza: UFC. 4ª ed. (Revisada e atualizada).
2002.
NATH, S.C.; SARMA, K.K.; VAJEZIKOVA, I.; LECLERCQ, P.A. Comparison of
volatile inflorescence oils and taxonomy of certain Cymbopogon taxa described as
Cymbopogon flexuosus (Ness ex Steud.) Wats. Biochemical Systematics and Ecology.
30: 151-162. 2002.
NEWALL, C.A.; ANDERSON, L.A. & PHILLIPSON, J.D. Plantas Medicinais: guia
para profissional de saúde. São Paulo: Editorial Premier. 2002.
NUNES, G.P.; SILVA, M.F.; RESENDE, U. M.; SIQUEIRA, J.M. Plantas medicinais
comercializadas por raizeiros no Centro de Campo Grande, Mato Grosso do Sul.
Revista Brasileira de Farmacognosia. 13(2): 83-92. 2003.
RENVOIZE, S. A. The grasses of Bahia. 1ª ed. Royal Botanic Gardens Kew. 1984.

ROBBERS, J.E.; SPEEDIE, M.K. & TYLER, V.E. Farmacognosia e
Farmacobiotecnologia. São Paulo: Editorial Premier. 1997.
SCHENKEL, E. P.; GOSMANN, G.; PETROVICK, P. R. Produtos de Origem Vegetal
e o Desenvolvimento de Medicamentos. In: SIMÕES, C.M.O.; SCHENKEL, E.P.;
GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.; PETROVICK, P. R. (Org.)
Farmacognosia: da planta ao medicamento. 5ª ed. Porto Alegre/Florianópolis:
UFRGS. p. 371-400. 2003.
SCHULZ, V.; HÄNSEL, R.; TYLER, V.E. Fitoterapia Racional: Um guia de
Fitoterapia para as Ciências da Saúde. 4ª ed. (1ª ed. Brasileira). São Paulo: Manole,
2002.
SILVA, C.S.P. As plantas medicinais no município de Ouro Verde de Goiás, GO,
Brasil: Uma abordagem etnobotânica. Dissertação (Mestrado em Botânica) –
Universidade de Brasília, Brasília-DF. 2007.
SILVA, R.R. Gramíneas (Poaceae) da Área de Relevante Interesse Ecológico
(ARIE) Santuário de Vida Silvestre do Riacho Fundo, DF-Brasil. Dissertação
(Mestrado em Botânica) – Universidade de Brasília, Brasília-DF. 2000.
SIMÕES, C.M.O.; MENTZ, L.A.; SCHENKEL, E.P.; IRGANG, B. E. Plantas da
Medicina Popular do Rio Grande do Sul. Porto Alegre: UFRGS. 1989.
SOENARKO, S. The Genus Cymbopogon Sprengel (Gramineae). Reinwardtia. Vol. 9
(3): 225-375. 1977.
SOUZA, V.C.; LORENZI, H. Botânica Sistemática (Guia ilustrado para identificação
das famílias das Angiospermas da flora brasileira, baseado em APG II). Nova Odessa,
SP: Instituto Plantarum, 2005.
TAKAISI-KIKUNI, N.B.; KRUGER, D.; GNANN, W.; WECKE, J. Microcalorimetric
and eletron microscopic investigation on the effects of essential oil from Cymbopogon
densiflorus on Staphylococcus aureus. Microbios. 88(354): 55-62. 1996.

TAKAISI-KIKUNI, N.B.; TSHILANDIA, D.; BABADY, B. Antibacterial activity of
the essential oil of Cymbopogon densiflorus. Fitoterapia. 71: 69-71. 2000.
TESKE, M.; TRENTINI, A.M.M. Compêndio de Fitoterapia. 4ª ed. Curitiba:
Herbarium Lab. Bot. Ltda., 2001.
VILA VERDE, G.M.; PAULA, J.R.; CARNEIRO, D.M. Levantamento Etnobotânico
das Plantas Medicinais do Cerrado Utilizadas pela População de Mossâmedes (GO).
Revista Brasileira de Farmacognosia. 13(supl.): 64-66. 2003.
WATSON, L.; DALLWITZ, M.J. The grass genera of the world: Cymbopogon
Spreng. Version 28th November 2005. Disponível em: <http://delta-inkey.com>. Acesso
em: 30/01/2007.
2 OBJETIVOS
2.1 Objetivo Geral

Ampliar o conhecimento sobre as espécies Cymbopogon densiflorus (Steud.)
Stapf. e C. nardus (L.) Rendle, realizando o estudo morfo-anatômico das folhas e
colmos, prospecção fitoquímica e análise dos óleos essenciais das folhas destas
espécies.
2.2 Objetivos Específicos
Realizar a análise comparativa da morfo-anatomia das folhas e colmos de
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle;
Realizar a prospecção fitoquímica das folhas destas espécies, verificando as
classes de metabólitos secundários presentes em ambas;
Extrair e analisar a composição química dos óleos essenciais das folhas das
espécies em estudo;
Estabelecer parâmetros de qualidade da matéria-prima vegetal;
Fornecer subsídios para pesquisas taxonômicas e áreas afins.

______________________________________________________________
CAPÍTULO 1
MORFO-ANATOMIA DAS FOLHAS E DOS COLMOS DE Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle (Poaceae:
Panicoideae)

1 INTRODUÇÃO
A família Poaceae (Gramineae) trata-se de uma das maiores entre as
Angiospermas, sendo possivelmente a de importância econômica mais relevante para o
homem (JOLY, 2002). Possui aproximadamente 700 gêneros e 12.000 espécies e estão
amplamente distribuídas geograficamente, isto é, são cosmopolitas (WATSON e
DALLWITZ, 1992).
A grande variação nos caracteres morfo-anatômicos das folhas e dos caules
deve-se a um conjunto de fatores adaptativos que ocorreram ao longo da evolução das
plantas, para que a planta obtivesse sucesso ao meio, fornecendo importantes dados
sobre a ecologia das espécies (FAHN e CUTLER, 1992), além de serem importantes na
descrição Sistemática das espécies vegetais (DICKINSON, 2000).
Segundo JUDD et al. (1999), a importância econômica e ecológica dessa família
tem motivado estudos aplicados à Taxonomia. Características anatômicas úteis na
sistemática desta família têm sido demonstradas por diversos autores (FREIER, 1959;
METCALFE, 1960; SENDULSKY e LABOURIAU, 1966; CAVALCANTE, 1968;
CAMPOS e LABOURIAU, 1969; SÖNDAHL e LABOURIAU, 1970; TEIXEIRA DA
SILVA e LABOURIAU, 1970; FIGUEIREDO e HANDRO, 1971; ELLIS, 1976 e
1979; KRISHNAN et al., 2000; PRYCHID, 2004, entre outros).
Estudos anatômicos em pesquisas farmacognósticas apresentam papel relevante
no diagnóstico taxonômico e no controle de qualidade da matéria-prima vegetal.
DUARTE e BARDAL (2002) concluíram que aproximadamente 3,3% das amostras
comercializadas tinham erros de identidade, pois diferentes espécies são popularmente
conhecidas pelo mesmo nome.
Alguns estudos anatômicos com espécies do gênero Cymbopogon, visando
fornecer subsídios para análises farmacognósticas foram realizados por LEWINSOHN
et al. (1998) e MARTINS et al. (2004). AMARAL et al. (2003) analisando diversas
amostras vegetais comercializadas, dentre elas Cymbopogon citratus (DC.) Stapf,
verificaram que tais amostras, além de conter elementos estranhos acima do limite
permitido na literatura especializada, continham também outros órgãos da própria planta
ou de uma outra espécie vegetal qualquer.
Nas monografias apresentadas pela Farmacopéia Brasileira percebe-se a
importância da Morfologia Externa e da Anatomia Vegetal para a identificação de
grande parte de matérias-prima vegetais. OLIVEIRA et al. (1991) ressaltam que,

geralmente as matérias-prima vegetais são comercializadas por raizeiros ou ervanários
sob a forma de pó, assim, macroscopicamente poucas avaliações podem ser realizadas.
A análise microscópica de amostras pulverizadas permite identificar características
histológicas que auxiliam na diagnose da matéria-prima vegetal, estabelecendo meios
para o controle de qualidade das mesmas (ALICE et al., 1995; COSTA, 2001).
Esta pesquisa teve por objetivo, apresentar caracteres anatômicos e úteis para a
determinação de parâmetros de qualidade da matéria-prima vegetal, bem como fornecer
dados anatômicos inéditos que contribuam à Sistemática destas espécies dentro do
gênero, da família e comparação entre as duas espécies estudadas.

2 MATERIAL E MÉTODOS
2.1 Material Botânico
Para o estudo morfo-anatômico foram utilizados exemplares cultivados de C.
densiflorus (Steud.) Stapf, coletados em duas localidades diferentes no município de
Jataí-GO - localidade 1 (17°52’14,0”S 051°43’14,9” W) e localidade 2 (17°52’07,3’’ S
e 51°43”18,8’’ W) e C. nardus (L.) Rendle, coletados na Reserva Ecológica do IBGE-
RECOR, Brasília-DF – localidade 1 (15°56’41’’ S e 47°56’07’’ W) e no Horto de
Plantas Medicinais da Faculdade de Farmácia/UFG, Goiânia-GO – localidade 2
(16°48’33,3’’S 49°14’39,5’’ W).
O material botânico foi identificado pelo Professor Dr. Tarciso S. Filgueiras,
especialista em Poaceae e pesquisador da Reserva Ecológica do IBGE-RECOR,
Brasília-DF. As exsicatas das espécies encontram-se depositadas no Herbário UFG,
Universidade Federal de Goiás, sob os números UFG-30.334 (C. densiflorus, localidade
1), UFG-30.335 (C. densiflorus, localidade 2), UFG-30.333 (C. nardus, localidade 1) e
UFG-30.336 (C. nardus, localidade 2).
2.2 Caracterização Morfológica
A caracterização macroscópica das folhas foi realizada à vista desarmada,
seguindo os parâmetros de CHASE e SENDULSKY (1991), OLIVEIRA et al. (1991) e
OLIVEIRA e AKISUE (1993).
As espécies foram desenhadas por Haroldo de Araújo e as escalas que
acompanham as ilustrações foram obtidas a partir do tamanho original.
2.3 Caracterização Anatômica
2.3.1 Microscopia óptica
Para a análise anatômica, ambas as espécies encontravam-se em fase reprodutiva
e foram utilizadas a quarta e quinta folhas completamente expandidas, a partir da folha-
cartucho. Destas folhas, foram retirados fragmentos da região mediana da lâmina e

bainha. Da lâmina foliar foram retiradas amostras da nervura central, entrenervuras e
bordo. Nos colmos foram retirados, o terceiro e quarto entrenós. As folhas e os colmos
foram preservados em álcool 70%.
A confecção das lâminas histológicas foi realizada a partir de secções
paradérmicas e transversais das lâminas e bainhas foliares e colmo, obtidas à mão livre.
As secções foram clarificadas em hipoclorito de sódio 12%, lavadas em água destilada,
e a seguir submetidas à dupla coloração com azul de astra 0,3% e fucsina básica 0,1%.
Posteriormente, desidratadas em série alcoólica crescente (30%, 50%, 70%, 90% e
100%) e pós-desidratadas em xilol e álcool (1:1) e xilol puro (JOHANSEN, 1940). As
secções foram montadas entre lâminas e lamínulas utilizando-se resina Verniz Vitral
incolor 500®, da marca Acrilex (PAIVA et al., 2006).
A dissociação da epiderme foi realizada segundo GHOUSE e YUNUS (1972),
consistindo na fervura de fragmentos de amostras da lâmina e bainha foliar em solução
de ácido nítrico a 50%, e em seguida, os fragmentos foram neutralizados com hidróxido
de sódio a 1%. Posteriormente, foram lavados em água destilada e submetidos à
coloração com azul de metileno 1% (JOHANSEN, 1940) com borato de sódio 1%, para
a identificação das células silicificadas.
Para os testes microquímicos, as secções foram submetidas ao reagente de
Steinmetz (COSTA, 1970), para identificação de amido, celulose, lignina, suberina,
lipídeos diversos, compostos fenólicos, cutina e látex; ao floroglucinol acidificado para
identificação de lignina (JOHANSEN 1940; KNEEBONE, 1962) e ao reagente sudan
IV para evidenciar compostos lipídicos (GERLACH, 1984).
O cálculo da densidade estomática que é o número de estômatos por mm² de
superfície foliar (BORGHEZAN et al., 2003), foi realizado utilizando-se 5 folhas
completamente expandidas e escolhidas ao acaso, nos mesmos exemplares. Dessas, foi
retirado um fragmento dos terços basal, mediano e apical da lâmina foliar (superfícies
adaxial e abaxial).
A confecção das lâminas, para o cálculo da densidade estomática, foi realizada a
partir da técnica de impressão da superfície foliar em lâmina de microscópio, contendo
uma gota de cola de secagem rápida à base de éster de cianoacrilato (comercialmente
conhecida como ‘Super-Bonder’), de acordo com a metodologia descrita por
SEGATTO et al. (2004).
A contagem dos estômatos foi feita em fotomicroscópio ZEISS Axioskop
equipado com câmara clara. Para cada fragmento, foram contados os estômatos em 3

campos aleatórios, totalizando 90 campos para cada uma das amostras. Segundo a
metodologia de LABOURIAU et al. (1961), com auxílio de lâmina graduada em
micrômetros (µm), foi desenhado um campo de 230 µm x 230 µm, utilizando-se
objetiva de 40x e ocular de 10x. Após a contagem, foram calculados a média do número
de estômatos por mm², desvio-padrão e coeficiente de variação. As médias do número
total de estômatos por mm² foram analisadas pelo teste t de Student, verificando se as
amostras apresentavam médias significativamente diferentes. Utilizou-se o programa
Prism 4.0 para essas análises e foi adotado nível de significância de 5%.
As fotomicrografias foram realizadas em fotomicroscópio ZEISS Axioskop,
utilizando-se filme Kodacolor ASA 100. Os desenhos foram feitos utilizando-se
fotomicroscópico ZEISS Axioskop equipado com câmara clara. As escalas que
acompanham as ilustrações foram obtidas nas mesmas condições ópticas.
2.3.2 Microscopia Eletrônica de Varredura
Fragmentos da porção mediana das folhas de Cymbopogon densiflorus e C.
nardus foram fixados em solução de Karnovsky modificada, composta por glutaraldeído
2%, paraformaldeído 2% em tampão cacodilato de sódio pH 7,2 0,05 M por 24 horas.
Após esse tempo, os fragmentos foram submetidos a três lavagens em tampão
cacodilato 0,05 M, com duração de 10 minutos cada, deixando-os em tampão,
acondicionados em eppendorf devidamente identificado. Foram armazenados em
refrigerador, fazendo trocas temporárias quando o tampão se apresentasse escuro.
Após, os fragmentos foram submetidos a tampão cacodilato de sódio pH 7,2
0,05 M e tetróxido de ósmio (1:1) por 1 hora, a seguir lavados em água destilada de 2 a
3 vezes e posteriormente, foram desidratados em série etanólica crescente (30%, 50%,
70% e 90%) com duração de 15 minutos cada e 3 vezes em etanol 100%, 10 minutos
cada (BOZZOLA e RUSSEL, 1992). Em seguida, os fragmentos foram levados à
secagem ao ponto crítico de CO2, utilizando-se o Aparelho de Secagem ao Ponto Crítico
da Balzers CPD30, no Laboratório de Microscopia Eletrônica, da Universidade de
Brasília (UnB). Posteriormente, foram fixados utilizando fita adesiva com dupla face
em pequenos ‘stubs’ e metalizados em ouro, com camada de cobertura de
aproximadamente 40 nm por 2 minutos, em Aparelho Metalizador Zeiss DSM-962 no

laboratório de Microscopia Eletrônica da Embrapa Recursos Genéticos e
Biotecnologia / CENARGEN, em Brasília-DF.
A análise foi realizada em Microscópio de Varredura JEOL-JSM 840A, no
laboratório de Microscopia Eletrônica da UnB. As eletromicrografias foram obtidas em
filme FUJI NEOPAN Asa 100. As escalas e feixes de elétrons encontram-se registradas
nas fotos.

3 RESULTADOS
3.1 CARACTERIZAÇÃO MORFOLÓGICA
3.1.1 Cymbopogon densiflorus (Steud.) Stapf
Os colmos dos espécimes de C. densiflorus medem 2 a 2,5 m de altura e
apresentam-se eretos, glabros e lisos, sendo pêndulos no ápice devido às densas
inflorescências (Fig. 1a). As lâminas foliares são inteiras, linear–lanceoladas, com
coloração verde-escura. Medem de 40 a 60 cm de comprimento por aproximadamente
3,8 cm de largura. Apresentam-se glabras e lisas em ambas as superfícies e um pouco
áspera nas margens. As folhas são paralelinérveas e invaginantes. Ápice lanceolado que
se estreita abruptamente e a base larga, levemente arredondada em contato com a
bainha. Bainha foliar glabra e lisa, medindo aproximadamente 13 cm, com coloração
verde-clara e grande quantidade de cêra de cor branca. Lígula membranosa, de tamanho
reduzido (Fig. 1b e 1c) e de coloração marrom-esverdeada. Folhas aromáticas,
apresentando cheiro característico de limão.
3.1.2 Cymbopogon nardus (L.) Rendle
Os colmos dos espécimes de C. nardus medem aproximadamente 1 m de altura e
apresentam-se eretos, glabros e lisos, sendo levemente pêndulos no ápice devido às
inflorescências (Fig. 2a). As lâminas foliares são inteiras, linear-lanceoladas, com
coloração verde-clara. Medem 120 a 150 cm de comprimento por 3 cm de largura.
Apresentam-se lisas na superfície adaxial e levemente áspera na superfície abaxial e ao
longo das margens. As folhas são paralelinérveas e invaginantes. Ápice lanceolado que
se estreita aos poucos e a base é estreita com nervura principal muito proeminente.
Bainha foliar glabra e lisa, medindo aproximadamente 38 cm, com coloração verde-
clara a ligeiramente rosada. Lígula membranosa-ciliada (Fig. 2b e 2c), saliente e de
coloração marrom. Folhas aromáticas, apresentando cheiro característico de eucalipto.

3.2 CARACTERIZAÇÃO ANATÔMICA
3.2.1 Cymbopogon densiflorus (Steud.) Stapf
3.2.1.1 Lâmina Foliar
Em vista frontal, verificam-se células epidérmicas longas com paredes
anticlinais levemente sinuosas (Fig. 6-8), intercaladas por uma a duas células curtas
com paredes delgadas ou espessamento de suberina nas paredes, caracterizando as
células suberosas. Ambas as faces apresentam nas regiões costais, células silicificadas
cruciformes, nodulares e halteriformes, sendo as últimas predominantes (Fig. 6, 8 e 11-
face adaxial e Fig. 14-face abaxial).
Os estômatos encontram-se presentes em ambas as faces e se localizam no
mesmo nível das células epidérmicas comuns. Em vista frontal, as células-guarda são
halteriformes e as células subsidiárias são tipo cúpula (Fig. 7, 9 e 12-face adaxial e Fig.
13-face abaxial). Conforme Tabela 2, pode-se observar que a epiderme abaxial
apresentou maior densidade estomática, caracterizando folha anfihipostomática.
Em secção transversal observa-se que a epiderme adaxial é unisseriada com
cutícula delgada (Fig. 3-5). Nas regiões intercostais foram observadas células
buliformes volumosas com paredes anticlinais delgadas e retas (Fig. 3-5). Essas células
mostram dimensões variadas, fazendo com que a superfície da lâmina foliar seja um
pouco ondulada. Nas regiões próximas à nervura central, abaixo das células buliformes,
ocorre uma camada de células parenquimáticas incolores de formato circular. O número
dessas células diminui gradativamente à medida que se afasta da nervura central,
apresentando apenas uma a duas células isoladas (Fig. 3 e 4).
Em secção transversal, a epiderme abaxial é unisseriada com cutícula delgada e
as células epidérmicas comuns são papilosas (Fig. 3 e 4).
Macrotricomas em forma de gancho (Fig. 9 e 10) e microtricomas ocorrem tanto
na face adaxial como na abaxial.
O parênquima clorofiliano é homogêneo e junto aos feixes vasculares se dispõe
radialmente aos mesmos, caracterizando a estrutura Kranz (Fig. 3 e 4).
Na região da nervura central, em feixes de 1ª e 2ª ordens, essas células
parenquimáticas radiadas são interrompidas por fibras esclerenquimáticas que se
estendem até a face abaxial (Fig. 15-17). Nos feixes de 2ª ordem localizados na região

compreendida entre a nervura central e o bordo, as fibras se estendem em direção às
duas faces da lâmina foliar (Fig. 3 e 4).
A região de nervura central é constituída por três feixes de 1ª ordem, sendo o
central maior e entre os mesmos, ocorrem feixes de 2ª e 3ª ordens. Essa região apresenta
face adaxial côncava e quilha pouco proeminente na face abaxial (Fig. 15). Os feixes de
1ª ordem apresentam forma circular e a extensão de bainha esclerenquimática possui
algumas fibras gelatinosas. Essas fibras se caracterizam por apresentar a parte interna da
parede celular não-lignificada, mostrando coloração azul quando submetidas à dupla
coloração com fucsina básica e azul de astra (Fig. 16) e também não apresentou
coloração vermelha quando tratada com floroglucinol acidificado (Fig. 17). Adjacente à
epiderme adaxial ocorrem três grupos de cordões esclerenquimáticos intercalados por
células buliformes, os quais se localizam em direção oposta aos feixes de 1ª ordem (Fig.
15). Na região compreendida entre os cordões esclerenquimáticos e os feixes vasculares
ocorrem células parenquimáticas isodiamétricas de tamanhos variados, com paredes
delgadas e destituídas de cloroplastos (Fig. 15).
Os feixes vasculares são colaterais e o floema é circundado por células
parenquimáticas pequenas com paredes lignificadas. Nos feixes de 1ª ordem, o xilema
possui lacuna do protoxilema e apresenta um elemento de metaxilema circular em cada
lado do protoxilema (Fig. 15-17). Esses feixes vasculares são envoltos por bainha dupla,
sendo a bainha externa parenquimática e a interna esclerenquimática (Fig. 16 e 17).
Os feixes de 2ª e 3ª ordens são completamente envoltos por bainha
parenquimática única, formada por células relativamente grandes contendo cloroplastos.
Essa bainha é circundada pelo parênquima clorofiliano radiado. A distância intervenal é
determinada por uma a três células parenquimáticas (Fig. 4) com paredes providas de
pequenas invaginações.
A região do bordo é constituída por fibras esclerenquimáticas. As células
epidérmicas possuem paredes periclinais externas lignificadas, ocorrendo também
células silicificadas (Fig. 18), retangulares e lume reduzido. Quanto ao formato, o bordo
é classificado em estreito projetado.
3.2.1.2 Bainha Foliar

Em corte transversal, a bainha foliar apresenta-se convoluta (Fig. 19). A
superfície adaxial é lisa, enquanto que a abaxial é levemente ondulada. A epiderme
adaxial e abaxial é unisseriada, recoberta por cutícula delgada. As células da epiderme
adaxial são alongadas periclinalmente e maiores do que as da abaxial. Em vista frontal,
as células epidérmicas apresentam paredes anticlinais levemente sinuosas. Os estômatos
ocorrem no mesmo nível que as células epidérmicas comuns em ambas as faces, sendo
raros na epiderme adaxial e abundantes na abaxial. Na epiderme abaxial, observam-se
macrotricomas (Fig. 20). Células suberosas e células silicificadas em forma de cruz e
sela presentes na região costal (Fig. 21).
O mesofilo é heterogêneo, constituído por cerca de dez camadas de células
parenquimáticas isodiamétricas de tamanhos variados, com paredes delgadas e
desprovidas de cloroplastos, junto à face adaxial e parênquima clorofiliano cujas células
se dispõem radialmente aos feixes vasculares que estão próximos à face abaxial (Fig. 22
e 23).
Os feixes vasculares são colaterais, sendo de 1ª, 2ª e 3ª ordens (Fig. 22 e 23) e
esses feixes estão envoltos por bainha única. Os feixes vasculares de 1ª ordem
apresentam forma elítpica, floema circundado por células parenquimáticas com paredes
lignificadas, presença de um elemento de metaxilema em cada lado dos elementos de
protoxilema e da lacuna (Fig. 22 e 23). Grupos de células esclerenquimáticas com
paredes pouco lignificadas ocorrem em lado oposto aos feixes vasculares de 1ª ordem,
imediatamente abaixo à epiderme adaxial (Fig. 22 e 23). Intercaladas aos feixes
vasculares foram observadas células parenquimáticas incolores com paredes delgadas
que se encontram isoladas ou em grupos de três a cinco células (Fig. 22 e 23).
3.2.1.3 Colmo
Em vista frontal, a epiderme é constituída por células epidérmicas comuns
longas com paredes espessas e levemente onduladas, intercaladas por células curtas
(Fig. 24). Ocorrem estômatos com células-guarda halterifornes e células subisidiárias
em forma de cúpula (Fig. 24).
Em secção transversal, a epiderme é unisseriada, constituída por células
predominantemente retangulares (Fig. 26 e 27), com paredes periclinais externas
espessadas, recobertas por cutícula delgada. Ocorrem células silicificadas cubóides e
alongadas; os estômatos estão situados no mesmo nível das células epidérmicas

comuns, no entanto as células subsidiárias se projetam na superfície do colmo (Fig. 26 e
27). Essas células são alongadas anticlinalmente e apresentam paredes periclinais
externas e anticlinais espessadas, enquanto que as periclinais internas são delgadas (Fig.
27). Ocorre uma a duas camadas de células subepidérmicas com paredes esclerificadas e
sob os estômatos, essas células possuem paredes celulósicas delgadas (Fig. 27). Logo
abaixo, ocorre uma a duas camadas de células parenquimáticas celulósicas (Fig. 26 e
27).
Os feixes vasculares envoltos por células esclerenquimáticas encontram-se
dispersos no tecido parenquimático (Fig. 25). Na parte mais externa do colmo, esse
tecido é constituído predominantemente, por células de paredes espessadas. Entre essas
células, são observadas faixas parenquimáticas cujas células possuem paredes delgadas
(Fig. 25). Nessa região, há duas ou três fileiras de feixes vasculares menores em relação
àqueles localizados na região mais interna do colmo, que é constituída por células
parenquimáticas maiores, com paredes delgadas, delimitando pequenos espaços
intercelulares triangulares (Fig. 25). Na parte central ocorre rompimento dessas células,
formando uma medula fistulosa. Os feixes vasculares são colaterais de 1ª ordem, o
xilema possui lacuna do protoxilema e apresenta um elemento de metaxilema em cada
lado do protoxilema (Fig. 28).

3.2.1. Cymbopogon nardus (L.) Rendle
3.2.2.1 Lâmina Foliar
Em vista frontal (Fig. 32), na epiderme adaxial, ocorrem células epidérmicas
comuns longas com paredes anticlinais levemente sinuosas, intercaladas por uma a duas
células curtas, com paredes delgadas ou com espessamento de suberina nas paredes,
caracterizando células suberosas. Em ambas as faces, ocorrem células silicificadas
cruciformes, nodulares e halteriformes, sendo as últimas predominantes.
Os estômatos encontram-se presentes em ambas as faces e se localizam no
mesmo nível das células epidérmicas comuns. Em vista frontal, as células-guarda são
halteriformes e as células subsidiárias são tipo cúpula (Fig. 32). Conforme Tabela 3,
observa-se que a epiderme abaxial apresentou maior densidade estomática,
caracterizando folha anfihipostomática.
Em secção transversal (Fig. 29-31) observa-se que a epiderme adaxial é
unisseriada com cutícula delgada. Nas regiões intercostais foram observadas células
buliformes volumosas com paredes anticlinais delgadas e retas (Fig. 29-31). Nas regiões
de lâmina foliar próximas à nervura central, abaixo das células buliformes, ocorre uma
camada de células parenquimáticas incolores de formato circular (Fig. 29 e 30). O
número dessas células diminui gradativamente à medida que se afasta da nervura
central, apresentando apenas uma a duas células isoladas.
Em secção transversal, a epiderme abaxial apresenta-se unisseriada, com
cutícula delgada e as células epidérmicas são papilosas. Cordões de esclerênquima
ocorrem nas regiões costais, formando sinuosidades (Fig. 29 e 30).
Macrotricomas em forma de espinho e gancho e microtricomas ocorrem em
ambas as superfícies, predominando na superfície abaxial (Fig. 33 e 34).
O parênquima clorofiliano é homogêneo e junto aos feixes vasculares se dispõe
radialmente aos mesmos, caracterizando estrutura Kranz (Fig. 29 e 30).
Na região da nervura central, em feixes de 1ª e 2ª ordens, essas células
parenquimáticas radiadas são interrompidas por fibras esclerenquimáticas que se
estendem até a face abaxial (Fig. 36 e 37). Nos feixes de 2ª ordem localizados na região
compreendida entre a nervura central e o bordo, as fibras se estendem em direção às
duas faces da lâmina foliar e as células parenquimáticas radiadas também são
interrompidas pelas fibras (Fig. 29 e 30).

Os feixes vasculares são colaterais e o floema é circundado por células
parenquimáticas pequenas com parede lignificada. Nos feixes de 1ª ordem, o xilema
apresenta lacuna do protoxilema e possui de um a dois elementos de metaxilema
circular em cada lado do protoxilema (Fig. 36 e 37). Esses feixes vasculares são
envoltos por bainha dupla, sendo a bainha externa parenquimática e a interna
esclerenquimática (Fig. 36 e 37).
Os feixes de 2ª e 3ª ordens são completamente envolvidos por bainha
parenquimática única, formada por células relativamente grandes contendo cloroplastos
volumosos. Essa bainha é circundada pelo parênquima radiado. A distância intervenal é
determinada por uma a três células parenquimáticas (Fig. 29 e 30), com paredes
apresentando pequenas invaginações.
A região da nervura central é constituída por três feixes de 1ª ordem, sendo o
central maior e entre os mesmos, ocorrem feixes de 2ª e 3ª ordens. Essa região apresenta
face adaxial plana e quilha proeminente (Fig. 35) na face abaxial. Os feixes de 1ª ordem
apresentam forma elíptica, a extensão de bainha esclerenquimática possui algumas
fibras gelatinosas. Essas fibras apresentam a parte interna da parede celular não-
lignificada, mostrando coloração azul quando submetidas à dupla coloração com fucsina
básica e azul de astra (Fig. 36) e também, não apresenta coloração vermelha quando
tratada com floroglucinol acidificado (Fig. 37). Próximo à epiderme adaxial, ocorrem
três grupos de cordões esclerenquimáticos intercalados por células parenquimáticas
isodiamétricas, os quais se localizam em direção oposta aos feixes de 1ª ordem (Fig.
35). Na região compreendida entre os cordões esclerenquimáticos e os feixes vasculares
ocorrem células parenquimáticas isodiamétricas de tamanhos variados, com paredes
delgadas e destituídas de cloroplastos (Fig. 35).
A região do bordo é constituída por fibras esclerenquimáticas. As células
epidérmicas possuem paredes periclinais externas lignificadas, ocorrendo também
células silicificadas (Fig. 38) retangulares e lume reduzido. Quanto ao formato, o bordo
é classificado em pontado (“cap pointed”).
3.2.2.2 Bainha Foliar

Em corte transversal, a bainha foliar apresenta-se involuta (Fig. 39). A superfície
adaxial é lisa, enquanto que a abaxial é levemente sinuosa. A epiderme adaxial e abaxial
é unisseriada e recoberta por cutícula delgada. As células da epiderme adaxial são
alongadas periclinalmente e mais estreitas. Em vista frontal, as células epidérmicas
apresentam paredes anticlinais levemente sinuosas. Os estômatos ocorrem no mesmo
nível que as células epidérmicas comuns em ambas as faces, sendo raros na epiderme
adaxial e abundantes na abaxial. Na epiderme abaxial, observam-se macrotricomas e
células silicificadas em forma de sela na região costal (Fig. 40).
O mesofilo é homogêneo, constituído por cerca de dez camadas de células
parenquimáticas isodiamétricas de tamanhos variados, com paredes delgadas e
desprovidas de cloroplastos (Fig. 41). Células parenquimáticas radiadas circundando os
feixes vasculares não foram observadas (Fig. 41 e 42).
Os feixes vasculares são colaterais, sendo de 1ª e 3ª ordens e esses feixes estão
envoltos por bainha dupla, sendo a externa parenquimática e a interna
esclerenquimática. Os feixes de 1ª ordem apresentam forma elíptica, o floema é
circundado por células com paredes fortemente lignificadas. Presença de um a dois
elementos de metaxilema em cada lado dos elementos de protoxilema e da lacuna (Fig.
41 e 42). Observou-se feixes de 1ª ordem com dois tamanhos, o maior se localiza mais
internamente e o menor encontra-se próximo à epiderme abaxial, com extensões de
bainha esclerenquimática. Grupos de células esclerenquimáticas com paredes pouco
lignificadas ocorrem em lado oposto aos feixes vasculares de 1ª ordem maiores,
imediatamente abaixo da epiderme adaxial (Fig. 41 e 42). As células parenquimáticas
incolores não foram observadas.
3.2.2.3 Colmo
Em vista frontal (Fig. 43 e 44), a epiderme é constituída por células longas com
paredes delgadas e retas, intercaladas por células curtas. Observaram-se células
silicificadas cubóides e alongadas.
Em secção transversal, a epiderme é unisseriada, constituída por células
predominantemente quadradas (Fig. 46 e 47), com paredes periclinais externas
espessadas, recobertas por cutícula delgada. Os estômatos apresentam células
subsidiárias em forma de cúpula e células-guarda halteriformes, estão situados no
mesmo nível das células epidérmicas (Fig. 43, 44 e 47) e as células subsidiárias não se

projetam na superfície do colmo. As células-guarda apresentam paredes periclinais
externa e interna espessada, e as células subsidiárias possuem base larga que se estreita
a medida que chega a superfície apresentando paredes periclinais externas e interna e
anticlinais delgadas (Fig. 47).
Ocorre uma a duas camadas de células subepidérmicas com paredes
esclerificadas e são intercaladas por grupos de células parenquimáticas que apresentam
paredes celulósicas delgadas (Fig. 46 e 47). Logo abaixo, ocorrem camadas de células
parenquimáticas com paredes lignificadas (Fig. 46), formando a região externa do
colmo. Nessa região localizam feixes vasculares, com bainha esclerenquimática densa
(Fig. 45 e 46).
A região interna do colmo é constituída por células parenquimáticas
isodiamétricas, com paredes delgadas limitando pequenos espaços intercelulares
triangulares (Fig. 45 e 46). Na parte central, ocorre medula fistulosa.
Os feixes vasculares são colaterais e de 1ª ordem e com lacunas do protoxilema,
ocorrendo 1 elemento de metaxilema a cada lado dos elementos de protoxilema (Fig.
48). Esses feixes encontram-se dispersos pelo parênquima, envoltos por densa bainha
esclerenquimática (Fig. 45 e 46).
Chave de identificação baseada em caracteres anatômicos
1. Presença de células buliformes na nervura central da lâmina foliar (Fig. 15) e
bordo estreito projetado (Fig. 18); bainha foliar convoluta, em secção transversal (Fig.
19) e presença de células incolores (Fig. 23); presença de células epidérmicas
retangulares e células subsidiárias que se projetam na superfície em secção transversal
do colmo (Fig. 27) ....................................................................... C. densiflorus
2. Ausência de células buliformes na nervura central da lâmina foliar (Fig. 35) e
bordo pontado (Fig. 38); bainha foliar involuta, em secção transversal (Fig. 39) e
ausência de células incolores (Fig. 42); presença de células epidérmicas quadradas e
células subsidiárias que não se projetam na superfície em secção transversal do colmo
(Fig. 47) ................................................................................................................... C.
nardus
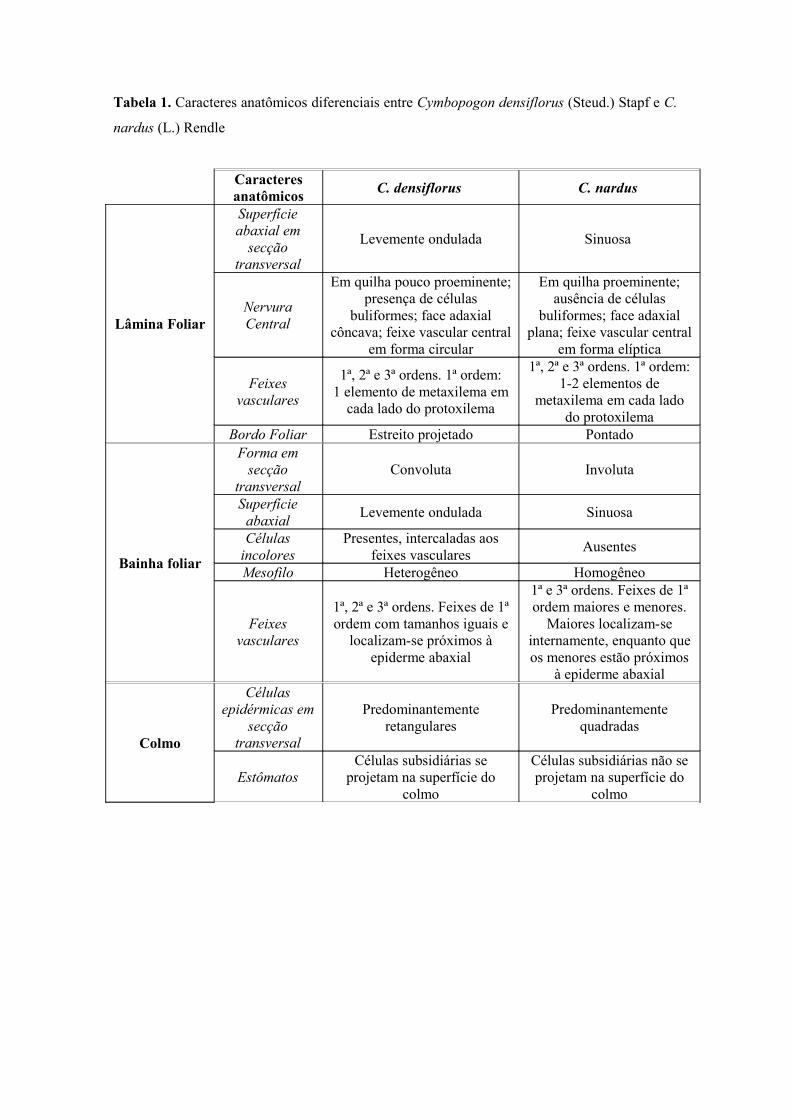
Tabela 1. Caracteres anatômicos diferenciais entre Cymbopogon densiflorus (Steud.) Stapf e C.
nardus (L.) Rendle
Caracteres anatômicos C. densiflorus C. nardus
Lâmina Foliar
Superfície abaxial em
secção transversal
Levemente ondulada Sinuosa
Nervura Central
Em quilha pouco proeminente; presença de células
buliformes; face adaxial côncava; feixe vascular central
em forma circular
Em quilha proeminente; ausência de células
buliformes; face adaxial plana; feixe vascular central
em forma elíptica
Feixes vasculares
1ª, 2ª e 3ª ordens. 1ª ordem: 1 elemento de metaxilema em
cada lado do protoxilema
1ª, 2ª e 3ª ordens. 1ª ordem: 1-2 elementos de
metaxilema em cada lado do protoxilema
Bordo Foliar Estreito projetado Pontado
Bainha foliar
Forma em secção
transversalConvoluta Involuta
Superfície abaxial Levemente ondulada Sinuosa
Células incolores
Presentes, intercaladas aos feixes vasculares Ausentes
Mesofilo Heterogêneo Homogêneo
Feixes vasculares
1ª, 2ª e 3ª ordens. Feixes de 1ª ordem com tamanhos iguais e
localizam-se próximos à epiderme abaxial
1ª e 3ª ordens. Feixes de 1ª ordem maiores e menores.
Maiores localizam-se internamente, enquanto que os menores estão próximos
à epiderme abaxial
Colmo
Células epidérmicas em
secção transversal
Predominantemente retangulares
Predominantemente quadradas
EstômatosCélulas subsidiárias se
projetam na superfície do colmo
Células subsidiárias não se projetam na superfície do
colmo
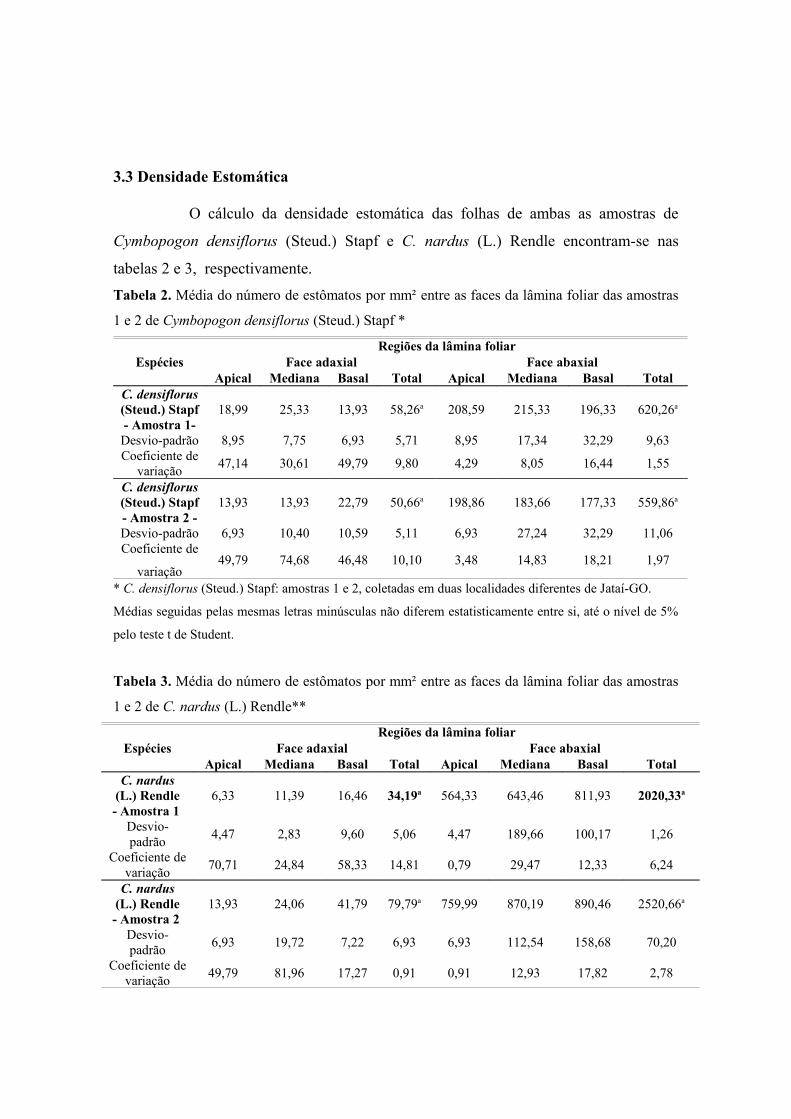
3.3 Densidade Estomática
O cálculo da densidade estomática das folhas de ambas as amostras de
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle encontram-se nas
tabelas 2 e 3, respectivamente.
Tabela 2. Média do número de estômatos por mm² entre as faces da lâmina foliar das amostras
1 e 2 de Cymbopogon densiflorus (Steud.) Stapf *
EspéciesRegiões da lâmina foliar
Face adaxial Face abaxialApical Mediana Basal Total Apical Mediana Basal Total
C. densiflorus (Steud.) Stapf - Amostra 1-
18,99 25,33 13,93 58,26a 208,59 215,33 196,33 620,26a
Desvio-padrão 8,95 7,75 6,93 5,71 8,95 17,34 32,29 9,63Coeficiente de
variação 47,14 30,61 49,79 9,80 4,29 8,05 16,44 1,55
C. densiflorus (Steud.) Stapf- Amostra 2 -
13,93 13,93 22,79 50,66a 198,86 183,66 177,33 559,86a
Desvio-padrão 6,93 10,40 10,59 5,11 6,93 27,24 32,29 11,06Coeficiente de
variação49,79 74,68 46,48 10,10 3,48 14,83 18,21 1,97
* C. densiflorus (Steud.) Stapf: amostras 1 e 2, coletadas em duas localidades diferentes de Jataí-GO.
Médias seguidas pelas mesmas letras minúsculas não diferem estatisticamente entre si, até o nível de 5%
pelo teste t de Student.
Tabela 3. Média do número de estômatos por mm² entre as faces da lâmina foliar das amostras
1 e 2 de C. nardus (L.) Rendle**
EspéciesRegiões da lâmina foliar
Face adaxial Face abaxialApical Mediana Basal Total Apical Mediana Basal Total
C. nardus (L.) Rendle- Amostra 1
6,33 11,39 16,46 34,19a 564,33 643,46 811,93 2020,33a
Desvio-padrão 4,47 2,83 9,60 5,06 4,47 189,66 100,17 1,26
Coeficiente de variação 70,71 24,84 58,33 14,81 0,79 29,47 12,33 6,24
C. nardus (L.) Rendle- Amostra 2
13,93 24,06 41,79 79,79a 759,99 870,19 890,46 2520,66a
Desvio-padrão 6,93 19,72 7,22 6,93 6,93 112,54 158,68 70,20
Coeficiente de variação 49,79 81,96 17,27 0,91 0,91 12,93 17,82 2,78

** C. nardus (L.) Rendle: amostra 1 – coletada na Reserva Ecológica do IBGE, RECOR, Brasília-DF;
amostra 2 – coletada no Horto de Plantas Medicinais, Faculdade de Farmácia/UFG, Goiânia-GO.
Médias seguidas pelas mesmas letras minúsculas não diferem estatisticamente entre si, até o nível de 5%
pelo teste t de Student.
4 DISCUSSÃO
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle apresentaram
caracteres anatômicos semelhantes e também, distintos entre si, nas lâminas e bainhas
foliares e colmos.
Na lâmina foliar, ambas as espécies apresentam epiderme adaxial e abaxial com
cutícula delgada; células epidérmicas longas e curtas: células suberosas e células
silicificadas; macrotricomas em forma de ganchos ou espinhos e microtricomas e
estômatos com células subsidiárias em forma de cúpula.
As espécies apresentam estômatos com células-guarda halteriformes e conforme
METCALFE (1960), as células subsidiárias são do tipo cúpula. FERRO et al. (1996) e
DUARTE e ZANETTI (2004) também verificaram estômatos com células-guarda
halteriformes em C. citratus (DC.) Stapf. Entretanto, FERRO et al. (1996), DUARTE e
ZANETTI (2004) e MARTINS et al. (2004) em estudos realizados com C. citratus, não
determinaram o tipo dos estômatos.
De acordo com as tabelas 2 e 3, o número de estômatos por mm² em ambas as
faces das folhas de C. densiflorus e C. nardus não apresentaram diferenças significantes
entre as amostras das mesmas espécies. No entanto, a face abaxial destas espécies
apresentam maior número de estômatos por mm². DUARTE e ZANETTI (2004)
ressaltam o maior número de estômatos na face abaxial em C. citratus.
As células buliformes em C. densiflorus e em C. nardus apresentam-se
arredondadas. De acordo com METCALFE (1960), as células buliformes, quando
examinadas em cortes transversais são grupos de células incolores que formam parte da
epiderme, mas diferem de suas vizinhas por serem mais largas e infladas. Atribuem a
essas células, uma função motora em que pela expansão e retração, devido à mudança
de turgor, as mesmas controlam o enrolamento das folhas.
Na região da nervura central de C. densiflorus, a epiderme adaxial apresenta
células buliformes, sendo que essa característica não foi registrada em C. nardus.
MONTEIRO e PACE (1984) também verificaram a presença dessas células na
epiderme adaxial na região da nervura central em Axonopus compressus (SW.) Beav.

As células silicificadas encontradas em C. densiflorus e C. nardus foram
analisadas de acordo com a classificação de METCALFE (1960) e ELLIS (1976-1979),
assim em vista frontal, foram registrados os tipos cruciformes, nodulares e
halteriformes, sendo os últimos predominantes. De acordo com BRANDENBURG et al.
(1985), EPSTEIN (1994), KRISHNAN et al. (2000) e PRYCHID et al. (2004), devido
às diferentes formas que apresentam, as células silicificadas possuem valor taxonômico
na identificação de capins, especialmente ao nível de subfamília e tribo.
CAVALCANTE (1968) analisou as células silicificadas das folhas de diversas
gramíneas amazônicas pertencentes à subfamília Panicoideae e verificou que as células
silicificadas halteriformes são as mais freqüentes. METCALFE (1960) destaca as
células silicificadas halteriformes e cruciformes como predominantes na subfamília
Panicoideae.
MOTOMURA et al. (2002) ressaltam que os capins absorvem do solo, a sílica
na forma de ácido silícico, durante a absorção de água, e a maior parte da sílica é
acumulada na parte aérea como sílica amorfa hidratada. Segundo MOTOMURA et al.
(2004), a deposição de sílica é uma das mais importantes características das plantas da
família Poaceae. Muitos pesquisadores têm examinado a distribuição, deposição e
funções da sílica nesta família, em que diversos mecanismos têm sido propostos para
explicar o acúmulo de sílica nas Poaceae. KAUFMAN et al. (1971) e EPSTEIN (1994),
reportam que o acúmulo de sílica (I) resulta da transpiração da superfície de partes
aéreas, (II) atua no controle das plantas para aumentar a estabilidade de seus tecidos,
(III) confere proteção contra microrganismos e herbívoros e (IV) protege epiderme
contra perda excessiva de água. De acordo com MCNAUGHTON e TARRANTS
(1983), a deposição de sílica nas folhas de capins foi um processo evolutivo importante
na defesa dessas plantas em relação aos herbívoros.
O mesofilo em C. densiflorus e C. nardus é homogêneo e junto aos feixes
vasculares se dispõem radialmente aos mesmos. Considerando a disposição ao redor dos
feixes, o parênquima radiado é tabular conforme ELLIS (1976-1979) e segundo
FREIER (1959), as células desse parênquima são raquimorfas, pois apresentam formato
que se assemelham aos ossos da coluna vertebral. Segundo FREIER, esse tipo celular é
característico de gramíneas pertencentes à subfamília Panicoideae. CAROLIN et al.
(1973) ressaltam que mesofilo arranjado radialmente aos feixes vasculares, é uma
característica de gramíneas da subfamília Panicoideae. O mesofilo homogêneo
constituído por clorênquima apresentando células pequenas e isodiamétricas foi

registrado na lâmina foliar de C. citratus (FERRO et al., 1996, DUARTE e ZANETTI,
2004 e MARTINS et al., 2004).
Nas espécies em estudo, a distância intervenal é determinada por uma a três
células parenquimáticas com paredes providas de curtas invaginações. Essas células são
denominadas por ELLIS (1976-1979) de células braciformes.
Em ambas as espécies, os feixes de 2ª e 3ª ordens apresentam bainha
parenquimática contendo cloroplastos. De acordo com JOHNSON e BROWN (1973),
essa é uma das características de plantas com metabolismo C4, dentre outras
características citológicas, anatômicas e fisiológicas.
C. densiflorus possui feixes de 1ª ordem em forma circular e C. nardus
apresentam feixes de 1ª ordem em forma elíptica, conforme classificação proposta por
METCALFE (1960) e ELLIS (1976-1979).
De acordo com METCALFE (1960), SHAW e SMEINS (1981) e ALVAREZ et
al. (2005), presença de microtricomas bicelulares com diversas formas; células
silicificadas halteriformes, nodulares e cruciformes dispostas em longas fileiras;
estômatos com células subsidiárias em forma de cúpula e triangular; células curtas
isoladas ou em pares nas regiões intercostais, nervura central com muitos feixes
vasculares de variados tamanhos; bainha de feixe vascular dupla: presença de bainha
interna esclerenquimática e bainha externa parenquimática e parênquima radiado em
torno dos feixes vasculares, são características típicas das Poaceae pertencentes à
subfamília Panicoideae.
Segundo a classificação proposta por METCALFE (1969) e ELLIS (1976-1979),
C. densiflorus possui bordo estreito com ponta projetada e C. nardus apresenta bordo
pontada.
A epiderme da bainha foliar de C. densiflorus e C. nardus apresentam células
epidérmicas longas e curtas: células suberosas e silicificadas e estômatos com células-
guarda halteriformes e células subsidiárias em cúpula. Estômatos raros na epiderme
adaxial e em maior número na epiderme abaxial. VIEIRA e MANTOVANI (1995)
verificaram que na bainha foliar de Deschampsia antarctica Desv., os estômatos
ocorrem predominantemente na epiderme abaxial.
C. densiflorus e C. nardus apresentam células silicificadas em forma de sela
(ELLIS, 1976-1979).
Em secção transversal, a bainha foliar não apresentou quilha. BRITO e
RODELLA (2002) verificaram que as bainhas de Brachiaria brizantha (Hochst. ex A.

Rich.) Stapf e B. humidicola (Rendle) Schweick, também não apresentaram quilha. No
mesofilo, C. densiflorus apresenta células de parênquima clorofiliano rodeando os
feixes vasculares, enquanto que C. nardus não as possuem. Em C. densiflorus,
observou-se uma a cinco células incolores com paredes delgadas, intercaladas aos feixes
vasculares, essas células não foram observadas em C. nardus. METCALFE (1960) e
ELLIS (1976-1979) denominam-as de células incolores e citam a ocorrência destas no
mesofilo das folhas de capins, em que sua presença e freqüência variam de acordo com
a espécie em questão, porém nada citam sobre a ocorrência de tais células no mesofilo
da bainha.
FERRO et al. (1996), DUARTE e ZANETTI (2004) e MARTINS et al. (2004)
em estudos realizados com C. citratus, não forneceram dados anatômicos sobre a bainha
foliar.
METCALFE (1960) ressalta os caracteres microscópicos da lâmina foliar como
os mais importantes em estudos taxonômicos, pois apresenta mais estruturas
especializadas que a bainha, que se torna madura posteriormente.
Os colmos de C. densiflorus e C. nardus apresentam variações anatômicas. Em
vista frontal, a epiderme do colmo de C. densiflorus apresenta células silicificadas
cubóides e alongadas, enquanto que C. nardus possui células silicificadas cubóides e
alongadas, segundo ELLIS (1976-1979).
Em ambas as espécies, os estômatos são do tipo cúpula, quanto à forma das
células subsidiárias (METCALFE, 1960).
Em corte transversal, pode ser observado que ambas as espécies apresentam
epiderme unisseriada, com cutícula delgada e presença de estômatos. C. densiflorus
apresenta células epidérmicas com paredes periclinais externas espessadas, estômatos
em maior tamanho que C. nardus. Ambas as espécies apresentam células subsidiárias
com formato arredondado, no entanto, em C. densiflorus estas células se projetam para
a superfície, o que não occore em C. nardus. ESAU (1976) relata a presença de
estômatos na epiderme dos colmos, em gramíneas.
C. densiflorus e C. nardus apresentam camadas subepidérmicas com células
parenquimáticas esclerificadas. METCALFE (1960) cita que em colmos de gramíneas,
o parênquima periférico geralmente apresenta paredes espessadas ou então, pode ser
substituído por esclerênquima periférico.
Os feixes vasculares em C. densiflorus e C. nardus são todos de 1ª ordem, com um
elemento de metaxilema grande em cada lado dos elementos de protoxilema e com

lacuna de protoxilema. Segundo METCALFE (1960), esses feixes apresentam formato
denominado como básico, constituindo o tipo mais comum em espécies de Gramineae.
A distribuição dos feixes vasculares no colmo de C. densiflorus está de acordo
com o padrão apresentado por METCALFE (1960) e ESAU (1976) em que os feixes
vasculares estão amplamente dispersos, sendo o estelo classificado em atactostelo.
ZIMMERMANN e TOMLINSON (1972) esclarecem que, os colmos de
monocotiledôneas usualmente possuem feixes vasculares individuais dispersos por todo
o parênquima; entretanto o termo “atactostelo” é incorreto, uma vez que esses
pesquisadores observaram que a distribuição dos feixes vasculares em colmos de
monocotiledôneas resulta de um processo de desenvolvimento ordenado e o termo mais
indicado seria “um estelo sem ordem”.

5 CONCLUSÃO
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle apresentaram
características anatômicas comuns, entretanto alguns caracteres são diferenciais como
formato da nervura central, presença ou ausência de células incolores, na bainha foliar,
formato das células epidérmicas e células subsidiárias no colmo, dentre outros, o que
permite distinguir as duas espécies, contribuindo para a taxonomia do gênero. Esses
caracteres podem assim, fornecer parâmetros a diagnose da matéria-prima vegetal, em
pesquisas farmacognósticas e no controle da qualidade de fitoterápicos.
As características anatômicas observadas como disposição radiada do parênquima
clorofiliano, presença de bainha de feixe parenquimática contendo cloroplastos
volumosos e pequena distância intervenal permitem concluir que as espécies estudadas
apresentam caracteres de plantas com metabolismo C4.

6 REFERENCIAS BIBLIOGRÁFICAS(*)
ALICE, C.B.; SIQUEIRA, N.C.S.; MENTZ, L.A.; SILVA, G.A.A.B. JOSÉ, K.F.D.J.
Plantas medicinais de uso popular: atlas farmacognóstico. Canoas: Ed. ULBRA.
1995.
ALVAREZ, J.M.; ROCHA, J.F.; MACHADO, S.R. Estrutura foliar de Loudetiopsis
chrysothrix (Nees) Conert e Tristachya leiostachya Nees (Poaceae). Revista Brasileira
de Botânica. 28(1): 23-37. 2005.
AMARAL, F.M.F.; COUTINHO, D.F.; RIBEIRO, M.N.S.; OLIVEIRA, M.A.
Avaliação da qualidade de drogas vegetais comercializadas em São Luís/Maranhão.
Revista Brasileira de Farmacognosia. 13(supl.): 27-30. 2003.
BORGHEZAN, M; MORAES, L.K.A; MOREIRA, F.M.; SILVA, A.L. Propagação in
vitro e avaliação de parâmetros morfofisiológicos de porta-enxertos de videira.
Pesquisa Agropecuária Brasileira. 7: 783-789. 2003.
BOZZOLA, J.J.; RUSSEL, L.D. Electron Microscopy: principles and techniques for
biologists. Boston: Jones and Bartlett Publishers. 1992.
BRANDENBURG, D.M.; RUSSELL, S.D.; ESTES, J.R.; CHISSOE, W.F.
Backscattered electron imaging as a technique for visualizing silica bodies in grasses.
Scanning Electron Microscopy. 4: 1509-1517. 1985.
CAMPOS, A. C.; LABOURIAU, L.G. Corpos silicosos de gramíneas dos cerrados-II.
Pesquisa Agropecuária Brasileira. 4: 143-151. 1969.
CAROLIN, R.C.; JACOBS, S.W.L.; VESK, M. The structure of the cells of mesophyll
and parenchymatous bundle sheath of the Gramineae. Botanical Journal of Linnean
Society. 66: 259-275. 1973.
______________________________________________________________________
(*) Referências Bibliográficas conforme ABNT/NBR-6023: 2002.

CAVALCANTE, P.B. Contribuição ao estudo dos corpos silicosos das gramíneas
amazônicas. Boletim do Museu Paraense Emílio Goeldi. 30: 1-39. 1968.
CHASE, A.; SENDULSKY, T. Primeiro livro de gramíneas: noções sobre a
estrutura com exemplos da flora brasileira. São Paulo: Instituto de Botânica. 1991.
COSTA, A.F. Farmacognosia. 3 v. 3ª ed. Lisboa: Fundação Calouste Gulbenkian.
1970.
DICKINSON, W.C. Integrative Plant Anatomy. Harcourt Academic Press/Elsevier:
2000.
DUARTE, M.R.; BARDAL, D. Qualidade de amostras vegetais comercializadas em
Curitiba-PR. Visão Acadêmica. 3(2): 65-68. 2002.
DUARTE, M.R.; ZANETTI, C.C. Estudo farmacobotânico de folhas de capim-limão:
Cymbopogon citratus (DC.) Stapf, Poaceae. Visão Acadêmica. 5(2): 117-124. 2004.
ELLIS, R.P. A procedure for standardizing comparative leaf anatomy in the Poaceae.
Bothaelia. 12(1): 65-109. 1976.
ELLIS, R.P. A procedure for standardizing comparative leaf anatomy in the Poaceae-II.
The epidermis as seen in surface view. Bothaelia. 12(4): 641-671. 1979.
EPSTEIN, E. Anomaly of silicon in plant biology. Proceedings of the National
Academy of Sciences USA. 91: 11-17. 1994.
ESAU, K. Anatomia das plantas com sementes. Reimpressão. São Paulo: Edgard
Blücher. 1976.
FAHN, A.; CUTLER, D.F. Xerophytes. Berlin: Gerbrüder Borntraeger. 1992.
FARMACÓPEIA BRASILEIRA. Parte 1. 4ª ed. São Paulo: Atheneu. 1988.

FERRO, V.C.; OLIVEIRA, I.; JORGE, L.I.F. Diagnose comparativa de três espécies
vegetais comercializadas como “ervas cidreiras”: Lippia alba (Mill) N.E.Br ex. Britt &
Wilson, Cymbopogon citratus (DC.) Stapf e Melissa officinalis L. Lecta – USF. 14(2):
53-63. 1996.
FIGUEIREDO, R.C.L; HANDRO, W. Corpos silicosos de gramíneas dos cerrados-V.
III Simpósio sobre o Cerrado. São Paulo: EDUSP – Editora da Universidade de São
Paulo. 1971.
FREIER, F. Las células clorenquimáticas del mesófilo de las gramíneas. Revista
Argentina de Agronomia. 1-2: 1-16. 1959.
GERLACH, D. Botanische mikrotechnik. Struttgart, Georg Thieme Verlag. 1984.
GHOUSE, A.K.; YUNUS, M. Preparation of epidermal peels from leaves of
Gymnosperm by treatment with hot, 60° GL HNO3. Stain Technology. 47(6): 322-325.
1972.
JOHANSEN, D.A. Plant microtechnique. New York : McGraw-Hill. 1940.
JOLY, A.B. Botânica: introdução à taxonomia vegetal. 13ª ed. São Paulo:
Companhia Editora Nacional. 2002.
JOHNSON, S.C.; BROWN, W.V. Grass leaf ultrastructural variations. American
Journal of Botany. 60(8): 727-735. 1973.
JUDD, W.S.; CAMPBELL, C.S.; KELLOGG, E.A.; STEVENS, P.F. Plant
Systematics: a phylogenetic approach. Sunderland, Massachusetts: Sinauer Associats.
1999.
KAUFMAN, P.B.; BIGELOW, W.C.; SCHMID, R.; GHOSHEH, N.S. Electron
microprobe analysis of silica in epidermal cells of equisetum. American Journal of
Botany. 58: 309-316. 1971.

KNEEBONE, W.R. A simple rapid, precise staining procedure for identification of
lignified tissue in grasses. Crop Science. 2: 268-269. 1962.
KRISHNAN, S.; SAMSON, N.P.; RAVICHANDRAN, P.; NARASIMHAN, D.;
DAYANANDAN, P. Phytoliths of Indian grasses and their potential use in
identification. Botanical Journal of the Linnean Society. 132: 241-252. 2000.
LABOURIAU, L.G.; OLIVEIRA, J.G.; LABOURIAU, M.L.S.; Transpiração de
Schizolobium parahyba (Vell.) Toledo. Anais da Academia Brasileira de Ciências.
33(2): 248-253. 1961.
LEWINSOHN, E.; DUDAI, N.; TADMOR, Y.; KATZIR, I.; RAVID, U.;
PUTIEVSKY, E.; JOEL, D.M. Histochemical localization of citral accumulation in
lemongrass leaves (Cymbopogon citratus (D.C.) Stapf., Poaceae). Annals of Botany.
81: 35-39. 1998.
MARTINS, M.B.G.; MARTINS, A.R.; TELASCRÊA, M.; CAVALHEIRO, A.J.
Caracterização anatômica de Cymbopogon citratus e perfil químico do óleo essencial.
Revista Brasileira de Plantas Medicinais. 6(3): 20-29. 2004.
MCNAUGHTON, S.J.; TARRANTS, J.L. Grass leaf silification: natural seletion for na
inducible defense against herbivores. Proceedings of the National Academy of
Science USA. 80: 790-791. 1983.
METCALFE, C.R. Anatomy of the monocotyledons. I-Gramineae. Oxford: Clarendon
Press. 1960.
MONTEIRO, M.C.C.; PACE, L.B. Anatomia foliar de Axonopus compressus (SW.)
Beauv. Atas da Sociedade Botânica do Brasil (Secção Rio de Janeiro). 2(5): 37-43.
1984.

MOTOMURA, H.; NITA, N.; SUZUKI, M. Silica accumulation in long-lived leaves of
Sasa veitchii (Carrière) Rehder (Poaceae-Bambusoideae). Annals of Botany. 90: 149-
152. 2002.
MOTOMURA, H.; FUJI, T.; SUZUKI, M. Silica deposition in relation to ageing of leaf
tissues in Sasa veitchii (Carrière) Rehder (Poaceae: Bambusoideae). Annals of Botany.
93: 235-248. 2004.
OLIVEIRA, F.; AKISUE, G. Fundamentos de Farmacobotânica. 1ª reimpressão. São
Paulo: Atheneu. 1993.
OLIVEIRA, F.; AKISSUE, G.; AKISUE, M.K. Farmacognosia. São Paulo: Atheneu.
1991.
PAIVA, J.G.A.; FANK-DE-CARVALHO, S.M.; MAGALHÃES, M.P; GRACIANO-
RIBEIRO, D. Verniz Vitral incolor 500®: uma alternativa de meio de montagem
economicamente viável. Acta Botanica Brasílica. 20(2): 257-264. 2006.
PRYCHID, C.J.; RUDALL, P.J.; GREGORY, M. Systematics and Biology of Silica
Bodies in Monocotyledons. The Botanical Review. 69(4): 377-440. 2004.
SEGATTO, F.B.; BISOGNIN, D.A.; BENEDETII, M.; COSTA, L.C.;
RAMPELOTTO, M.V.; NICOLOSO, F.T. Técnica para o estudo da anatomia da
epiderme foliar de batata. Ciência Rural. 34(5): 1597-1601. 2004.
SENDULSKY, T.; LABOURIAU, L.G. Corpos silicosos de gramíneas dos cerrados-I.
Anais da Academia Brasileira de Ciências. 38(supl.): 159-178. 1966.
SHAW, R.B.; SMEINS, F.E. Some anatomical and morphological characteristics of the
North American species of Eriochloa (Poaceae: Paniceae). Botanical Gazette. 142(4):
534-544. 1981.
SÖNDAHL, M.R.; LABOURIAU, L.G. Corpos silicosos de gramíneas dos cerrados-IV.
Pesquisa Agropecuária Brasileira. 5: 167-182. 1970.

TEIXEIRA DA SILVA, S.; LABOURIAU, L.G. Corpos silicosos de gramíneas dos
cerrados-III. Pesquisa Agropecuária Brasileira. 5: 167-182. 1970.
VIEIRA, R.C.; MANTOVANI, A. Anatomia foliar de Deschampsia antarctica Desv.
(Gramineae). Revista Brasileira de Botânica. 18(2): 207-220. 1995.
WATSON, L.; DALLWITZ, M.J. The families of flowering plants: descriptions,
illustrations, identification and information retrieval. Disponível em: <http://delta-
intkey.com>. Acesso em: 30/01/2007. 1992.
ZIMMERMANN, M.H.; TOMLINSON, P.B. The vascular system of
monocotyledonous stems. Botanical Gazette. 133(2): 141-155. 1972.

7 ILUSTRAÇÕES

Figura 1. Aspectos morfológicos de Cymbopogon densiflorus (Steud.) Stapf. A. Aspecto geral de um ramo, demonstrando inflorescências, disposição e formato das folhas. B. Lígula membranosa, com tamanho reduzido. C. Detalhe da lígula.
54

Figura 2. Aspectos morfológicos de Cymbopogon nardus (L.) Rendle. A. Aspecto geral de um ramo, demonstrando inflorescências, disposição e formato das folhas. B. Lígula membranoso-ciliada. C. Detalhe da lígula.
55

Figuras 3-8. Cymbopogon densiflorus (Steud.) Stapf - lâmina foliar. Fig. 3-5: Secções transversais. Fig. 3. Aspecto geral. Fig. 4. Detalhe mostrando células buliformes, parênquima clorofiliano radiado e feixes vasculares de 2ª e 3ª ordem. Fig. 5. Detalhe da epiderme adaxial, teste com reagente de Steinmetz, para evidenciar cutina. Fig. 6-8: Secções paradérmicas. Fig. 6. Aspecto geral da epiderme adaxial. Fig. 7. Detalhe da epiderme adaxial, demonstrando células longas, células suberosas e estômato. Fig. 8. Detalhe da epiderme adaxial, visualizando células silicificadas, células longas e células suberosas. BF – bainha de feixe; CB – células buliformes; CL – célula longa; CS – célula silicificada; CSub – célula suberosa; CUT – cutícula; EP AB – epiderme abaxial; EP AD – epiderme adaxial; EST – estômato; FV2a – feixe vascular de 2ª ordem; FV3a – feixe vascular de 3ª ordem; seta – distância intervenal; Asterisco: célula parenquimática incolor.
56
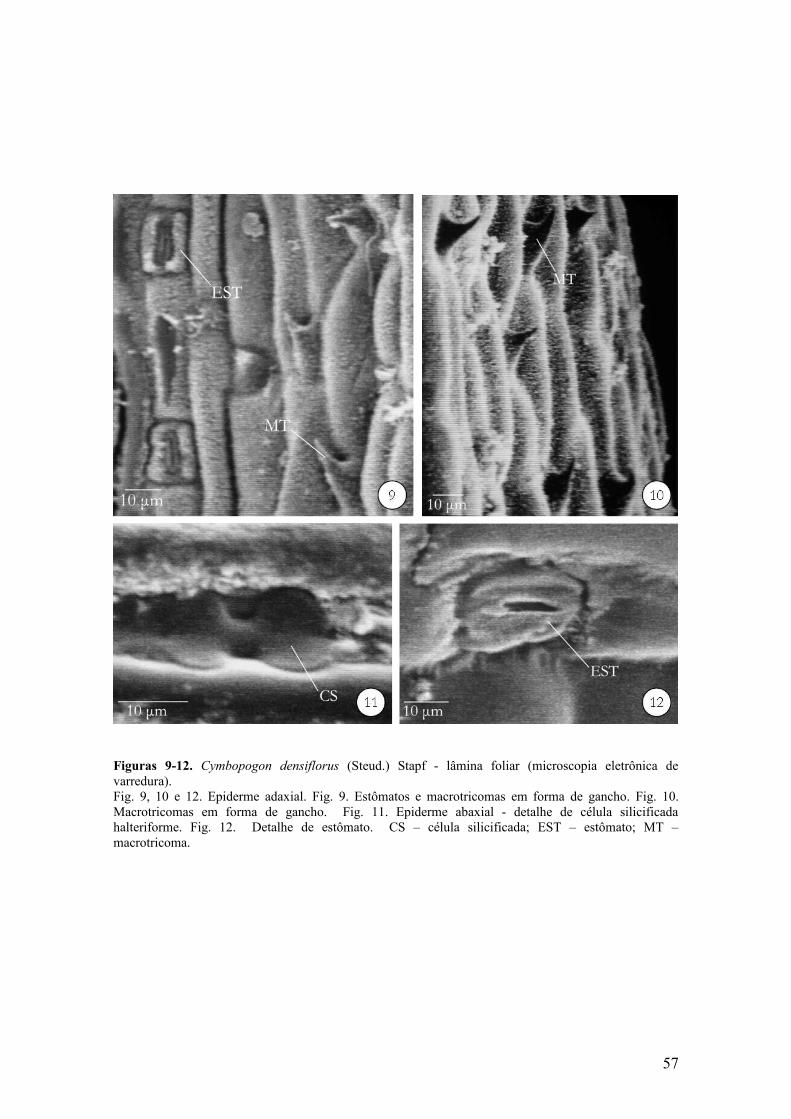
Figuras 9-12. Cymbopogon densiflorus (Steud.) Stapf - lâmina foliar (microscopia eletrônica de varredura).Fig. 9, 10 e 12. Epiderme adaxial. Fig. 9. Estômatos e macrotricomas em forma de gancho. Fig. 10. Macrotricomas em forma de gancho. Fig. 11. Epiderme abaxial - detalhe de célula silicificada halteriforme. Fig. 12. Detalhe de estômato. CS – célula silicificada; EST – estômato; MT – macrotricoma.
57
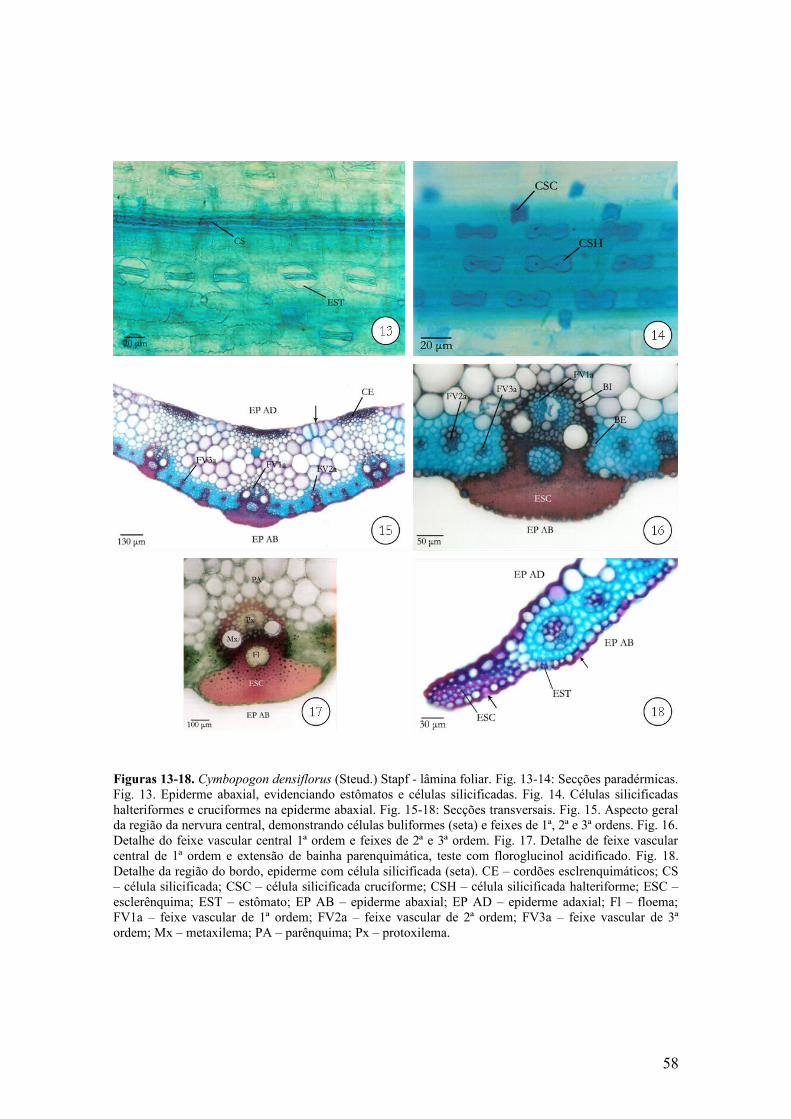
Figuras 13-18. Cymbopogon densiflorus (Steud.) Stapf - lâmina foliar. Fig. 13-14: Secções paradérmicas. Fig. 13. Epiderme abaxial, evidenciando estômatos e células silicificadas. Fig. 14. Células silicificadas halteriformes e cruciformes na epiderme abaxial. Fig. 15-18: Secções transversais. Fig. 15. Aspecto geral da região da nervura central, demonstrando células buliformes (seta) e feixes de 1ª, 2ª e 3ª ordens. Fig. 16. Detalhe do feixe vascular central 1ª ordem e feixes de 2ª e 3ª ordem. Fig. 17. Detalhe de feixe vascular central de 1ª ordem e extensão de bainha parenquimática, teste com floroglucinol acidificado. Fig. 18. Detalhe da região do bordo, epiderme com célula silicificada (seta). CE – cordões esclrenquimáticos; CS – célula silicificada; CSC – célula silicificada cruciforme; CSH – célula silicificada halteriforme; ESC – esclerênquima; EST – estômato; EP AB – epiderme abaxial; EP AD – epiderme adaxial; Fl – floema; FV1a – feixe vascular de 1ª ordem; FV2a – feixe vascular de 2ª ordem; FV3a – feixe vascular de 3ª ordem; Mx – metaxilema; PA – parênquima; Px – protoxilema.
58

Figuras 19-23. Cymbopogon densiflorus (Steud.) Stapf - bainha foliar. Fig. 19: Diagrama da bainha foliar, em secção transversal – aspecto geral. Fig. 20-21: Secções paradérmicas, epiderme abaxial. Fig. 20. detalhe de macrotricomas. Fig. 21. Detalhe de células silicificadas e células suberosas. Fig. 22-23: Secções transversais. Fig. 22. Aspecto geral. Fig. 23. Detalhe da área delimitada na Fig. 22, mostrando feixes vasculares de 1ª, 2ª e 3ª ordem e grupos de células esclerenquimáticas próximos à epiderme adaxial da bainha foliar (seta). CI – células incolores; EP AD – epiderme adaxial; EP AB – epiderme abaxial; FV1a – feixe vascular de 1ª ordem; FV2a – feixe vascular de 2ª ordem; FV3a – feixe vascular de 3ª ordem; MT – macrotricomas; PAR – parênquima; PRad – parênquima radiado; Px – protoxilema. Seta preta: grupo de células esclerenquimáticas com paredes pouco lignificadas.
59
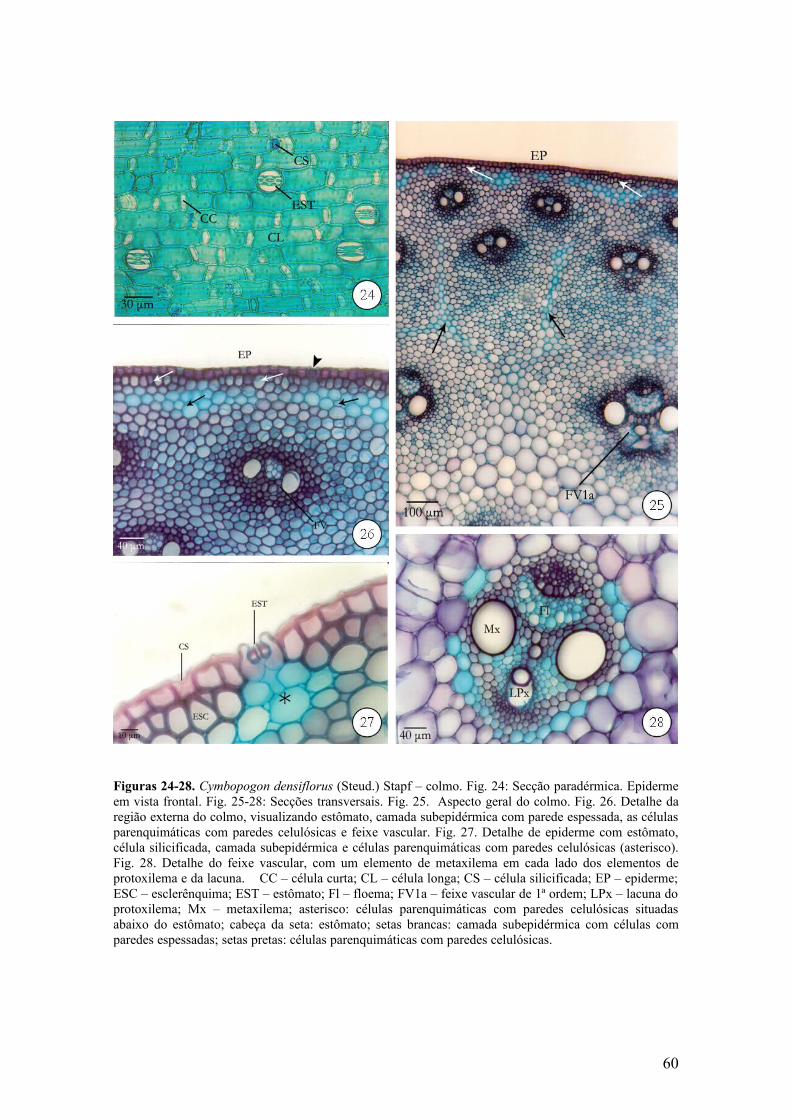
Figuras 24-28. Cymbopogon densiflorus (Steud.) Stapf – colmo. Fig. 24: Secção paradérmica. Epiderme em vista frontal. Fig. 25-28: Secções transversais. Fig. 25. Aspecto geral do colmo. Fig. 26. Detalhe da região externa do colmo, visualizando estômato, camada subepidérmica com parede espessada, as células parenquimáticas com paredes celulósicas e feixe vascular. Fig. 27. Detalhe de epiderme com estômato, célula silicificada, camada subepidérmica e células parenquimáticas com paredes celulósicas (asterisco). Fig. 28. Detalhe do feixe vascular, com um elemento de metaxilema em cada lado dos elementos de protoxilema e da lacuna. CC – célula curta; CL – célula longa; CS – célula silicificada; EP – epiderme; ESC – esclerênquima; EST – estômato; Fl – floema; FV1a – feixe vascular de 1ª ordem; LPx – lacuna do protoxilema; Mx – metaxilema; asterisco: células parenquimáticas com paredes celulósicas situadas abaixo do estômato; cabeça da seta: estômato; setas brancas: camada subepidérmica com células com paredes espessadas; setas pretas: células parenquimáticas com paredes celulósicas.
60

Figuras 29-34. Cymbopogon nardus (L.) Rendle - lâmina foliar. Fig. 29-30: Secções transversais. Fig. 29. Aspecto geral. Fig. 30. Detalhe de feixes vasculares de 2ª e 3ª ordem, células buliformes e parênquima clorofiliano radiado. Fig. 31. Detalhe da epiderme adaxial e cutícula revestindo-a, teste com Steinmetz. Fig. 32: Detalhe da epiderme abaxial, mostrando estômatos em secção paradérmica. Fig. 33-34: Microscopia eletrônica de varredura – epiderme abaxial. Fig. 33. Detalhe de macrotricoma em forma de gancho. Fig. 34. Detalhe de macrotricomas e estômato. BF – bainha de feixe vascular; CB – células buliformes; CE – cordões esclerenquimáticos; EP AB – epiderme abaxial; EP AD – epiderme adaxial; EST – estômatos; FV1a – feixe vascular de 1ª ordem; FV2a – feixe vascular de 2ª ordem; FV3a – feixe vascular de 3ª ordem; MT – macrotricoma; PRad – parênquima radiado.
61
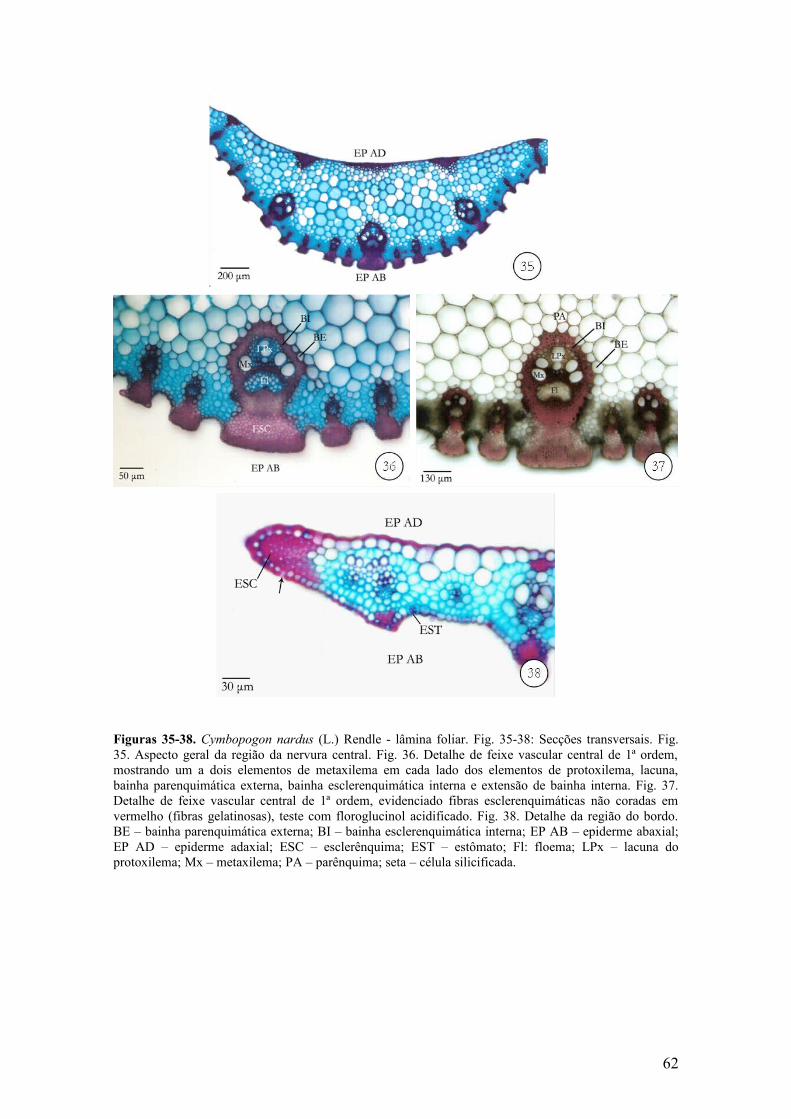
Figuras 35-38. Cymbopogon nardus (L.) Rendle - lâmina foliar. Fig. 35-38: Secções transversais. Fig. 35. Aspecto geral da região da nervura central. Fig. 36. Detalhe de feixe vascular central de 1ª ordem, mostrando um a dois elementos de metaxilema em cada lado dos elementos de protoxilema, lacuna, bainha parenquimática externa, bainha esclerenquimática interna e extensão de bainha interna. Fig. 37. Detalhe de feixe vascular central de 1ª ordem, evidenciado fibras esclerenquimáticas não coradas em vermelho (fibras gelatinosas), teste com floroglucinol acidificado. Fig. 38. Detalhe da região do bordo. BE – bainha parenquimática externa; BI – bainha esclerenquimática interna; EP AB – epiderme abaxial; EP AD – epiderme adaxial; ESC – esclerênquima; EST – estômato; Fl: floema; LPx – lacuna do protoxilema; Mx – metaxilema; PA – parênquima; seta – célula silicificada.
62

Figuras 39-42. Cymbopogon nardus (L.) Rendle - bainha foliar. Fig. 39: Diagrama da bainha foliar em secção transversal – aspecto geral. Fig. 40: Detalhe da epiderme abaxial, demonstrando macrotricomas e células silicificadas. Fig. 41-42: Secções transversais. Fig. 41. Aspecto geral do mesofilo. Fig. 42. Detalhe da área delimitada na Fig. 39, evidenciando feixe de 1ª e 3ª ordem. CS – célula silicificada; EP AD – epiderme adaxial; EP AB – epiderme abaxial; FV1a – feixe de 1ª ordem; FV3a – feixe de 3ª ordem; MT – macrotricoma; PA – parênquima.
63
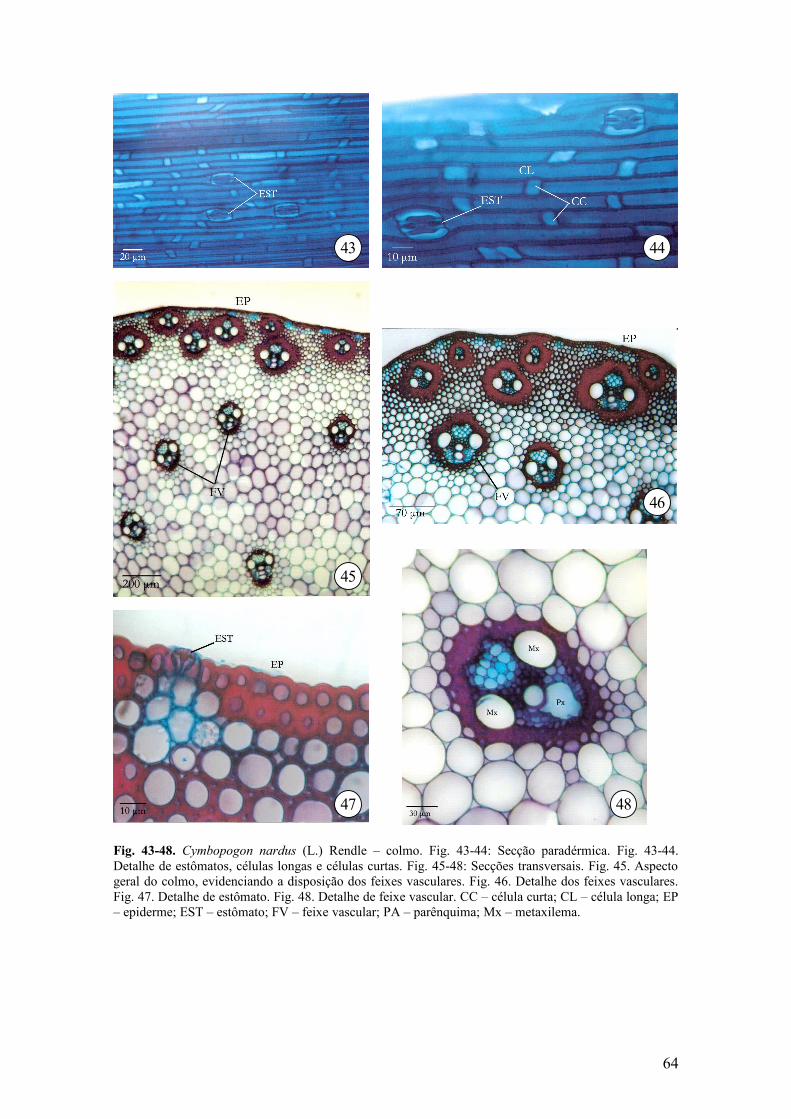
43 44
45
46
47 48
43 44
45
46
47 48
Fig. 43-48. Cymbopogon nardus (L.) Rendle – colmo. Fig. 43-44: Secção paradérmica. Fig. 43-44. Detalhe de estômatos, células longas e células curtas. Fig. 45-48: Secções transversais. Fig. 45. Aspecto geral do colmo, evidenciando a disposição dos feixes vasculares. Fig. 46. Detalhe dos feixes vasculares. Fig. 47. Detalhe de estômato. Fig. 48. Detalhe de feixe vascular. CC – célula curta; CL – célula longa; EP – epiderme; EST – estômato; FV – feixe vascular; PA – parênquima; Mx – metaxilema.
64

____________________________________________________________
CAPÍTULO 2
ESTUDOS FITOQUÍMICOS E ENSAIOS DE PUREZA DAS
FOLHAS DE Cymbopogon densiflorus (Steud.) Stapf e C.
nardus (L.) Rendle (Poaceae: Panicoideae)
____________________________________________________________
65

1 INTRODUÇÃO
Em um contexto histórico, o homem se utiliza de recursos da natureza para
muitos fins. Dentre esses recursos, as plantas têm um amplo uso, principalmente
alimentício e medicinal e com a constante interação entre o homem e as plantas, fez-se
necessário delinear a medicina popular regional, ou seja, a abordagem etnobotânica
(VILA VERDE et al., 2003). A medicina popular baseada no uso de plantas medicinais
como fitoterápicos continua tendo um papel importante na cura de enfermidades
(BUENO et al., 2005).
A etnobotânica e a etnofarmacologia tem contribuído para a descoberta de novos
agentes terapêuticos importantes, derivados a partir de metabólitos secundários das
plantas (KATEWA, 2004; KLOUCEK et al., 2005). A definição original ao termo
“etnobotânica” era aplicada ao estudo da relação de uso entre os homens e as plantas;
entretanto, verifica-se que além do uso, outras relações também devem ser consideradas,
tais como: a ecologia, a história, a economia da sociedade em questão que se beneficia
de determinados vegetais. O termo “etnofarmacologia” é recém-criado e define-se como
uma área multidisciplinar de pesquisa que se ocupa da observação, descrição e
investigações experimentais de fitoterápicos e suas atividades biológicas, utilizadas por
uma determinada sociedade (SOEJARTO et al., 2005).
A obtenção de fitoterápicos a partir de metabólitos secundários de origem
vegetal, bem como o desenvolvimento de fármacos, vem despertando grande interesse
não só por parte dos pesquisadores de produtos naturais, mas principalmente de
pequenas e grandes indústrias farmacêuticas (SEIDL, 2002; MONTANHER et al.,
2003).
Nos países em desenvolvimento e comunidades rurais, o uso de plantas
medicinais é muitas vezes o único e valioso recurso. Devido o difícil acesso e alto custo
dos medicamentos industrializados, essas comunidades optam pela medicina popular
para a cura de muitas enfermidades (MACIEL et al., 2002; ADAMU et al., 2005;
MACÍA et al., 2005). Conseqüentemente, em decorrência desses fatores, o comércio de
plantas medicinais e produtos fitoterápicos encontra-se em expansão por todo o mundo
(BRANDÃO et al., 1998).
Em geral, parte da população faz uso das plantas medicinais por indicação dos
raizeiros, que desenvolvem o extrativismo e comercialização, em pequena escala, das

plantas nativas da região (VILA VERDE et al., 2003). Geralmente os raizeiros fazem a
comercialização informal de plantas medicinais em locais cujas condições físicas são
inadequadas, muitas vezes próximos aos pedestres e ao tráfego de veículos que
proporcionam vários tipos de poluições. Muitos materiais vegetais ficam
completamente expostos ao ar, sendo vulneráveis aos ataques de insetos e contaminação
por microorganismos, bem como aos efeitos de luz, gases e temperatura (STAFFORD
et al., 2005).
A qualidade do material vegetal é um determinante imprescindível para a
qualidade do fitoterápico. A eficácia é fornecida a partir de ensaios farmacológicos pré-
clínicos e clínicos das atividades biológicas recomendadas para tais fitoterápicos, bem
como a segurança, que por meio de ensaios, determina a ausência de efeitos tóxicos e
também a inexistência de agentes contaminantes nocivos à saúde, como os agrotóxicos,
metais pesados e microorganismos (FARIAS, 2004).
Como a eficácia e a segurança terapêutica das plantas dependem da qualidade,
que pode sofrer influências de diversos fatores, há um risco real ao consumidor da
matéria-prima vegetal comercializada por raizeiros. A livre venda de material vegetal
inadequado para o consumo, deixa evidente a necessidade de que autoridades de
vigilância sanitária estejam ativas na fiscalização e controle de qualidade destes
materiais vegetais comercializados e usados para fins terapêuticos (AMARAL et al.,
2003).
NUNES et al. (2003) ressaltam que, além do grande extrativismo e dos fatores
que diminuem a qualidade das plantas medicinais comercializadas por raizeiros e
consequentemente, reprovem o seu consumo, há ainda a automedicação em que as
pessoas consomem espécies não inteiramente avaliadas do ponto de vista farmacológico
e toxicológico, podendo acarretar em sérios efeitos colaterais às pessoas.
Em relação às espécies de interesse nesta pesquisa, Cymbopogon densiflorus
(Steud.) Stapf. e C. nardus (L.) Rendle, não foram encontrados dados na literatura que
tenham fornecido parâmetros de qualidade destas espécies, reforçando a necessidade de
se realizar a prospecção fitoquímica, a fim divulgar dados valiosos que possam
contribuir para o controle de qualidade das mesmas.

2 MATERIAL E MÉTODOS
2.1 Material botânico
Para a realização desta pesquisa foram coletadas folhas de exemplares de
Cymbopogon densiflorus (Steud.) Stapf, em duas localidades do município de Jataí-GO
(17°52’14,0”S 051°43’14,9” W) e (17°52’07,3’’ S e 51°43”18,8’’ W) – denominadas C.
densiflorus amostras 1 e 2, respectivamente e C. nardus (L.) Rendle, na Reserva Ecológica do
IBGE-RECOR, Brasília-DF (15°56’41’’ S e 47°56’07’’ W) e do Horto de Plantas Medicinais
da Faculdade de Farmácia - UFG, Goiânia-GO (16°48’33,3’’S 49°14’39,5’’ W) –
denominadas C. nardus amostras 1 e 2 . Em todos os indivíduos, as amostras foram coletadas
durante a época em que as inflorescências encontravam-se maduras, no período de junho a
setembro de 2006. As exsicatas das espécies estão depositadas no Herbário UFG, da
Universidade Federal de Goiás sob os números 30.334 (C. densiflorus, amostra 1), 30.335 (C.
densiflorus, amostra 2), 30.333 (C. nardus, amostra 1) e 30.336 (C. nardus, amostra 2).
A fim de realizar a prospecção fitoquímica e também, determinar os teores de
umidade, cinzas totais e cinzas insolúveis em ácido clorídrico, as folhas coletadas
encontravam-se completamente expandidas e expostas ao sol. Para o preparo das amostras, as
folhas foram dessecadas ao ar, e em seguida, transportadas para o Laboratório de
Farmacognosia, da Faculdade de Farmácia, da Universidade Federal de Goiás.
As folhas dessecadas foram trituradas em moinho de facas. O pó assim obtido foi
devidamente acondicionado, identificado e armazenado até a sua utilização nos ensaios.
2.2 Determinação do teor de umidade
Para a determinação do teor de umidade, os ensaios foram realizados em triplicata
conforme FARMACOPÉIA BRASILEIRA IV (1988). Pesou-se, em balança analítica,
aproximadamente 2 g do material botânico pulverizado e transferidos para um béquer
previamente pesado e dessecado a 100-105o C por 1 hora. Em seguida, a amostra foi
dessecada em estufa a 100-105o C por 5 horas, resfriada em dessecador e pesada. Prosseguiu-
se a dessecação pesando o material em intervalos de 1 hora. O ensaio foi considerado
concluído quando duas

pesagens sucessivas não diferiram entre si por mais de 5 mg. A porcentagem de água foi
calculada em relação à amostra seca ao ar utilizando a equação:
Equação 1: Cálculo do teor de umidade em porcentagem
% teor de umidadePxN100=
Onde:N – perda de peso da amostra em gramas;P – quantidade de amostra em gramas.
2.3 Determinação do teor de cinzas
2.3.1 Determinação do teor de cinzas totais
Para a determinação do teor de cinzas totais, os ensaios foram realizados em triplicata,
conforme a FARMACOPÉIA BRASILEIRA IV (1988). Pesou-se em balança analítica,
aproximadamente 3 g da amostra pulverizada, os quais foram transferidos para um cadinho de
porcelana previamente calcinado, resfriado e pesado. A amostra foi distribuída de forma
uniforme e incinerada em mufla, aumentando-se a temperatura até 500o C, até a obtenção de
cinzas brancas. Em seguida, a amostra foi resfriada em dessecador e pesada. A porcentagem
de cinzas totais foi calculada em relação à amostra seca ao ar conforme equação abaixo:
Equação 2: Cálculo do teor de cinzas em porcentagem
% teor de cinzas PxN100=
Onde:N – quantidade de cinzas totais da amostra em gramas;P – quantidade de amostra em gramas.
2.3.2 Determinação do teor de cinzas insolúveis em ácido clorídrico (HCl)
Para a determinação do teor de cinzas insolúveis em ácido clorídrico, os resíduos
obtidos na determinação do teor de cinzas totais, em triplicata, foram fervidos durante 5
minutos com 12,5 ml de HCl SR, preparado conforme FARMACOPÉIA BRASILEIRA IV
(1988) em cadinhos cobertos com vidro de relógio. O resíduo foi filtrado em papel de filtro
quantitativo, os cadinhos e os vidros de relógio foram lavados com água quente. O papel de
filtro contendo o resíduo foi lavado com água quente até o filtrado tornar-se neutro.

O papel de filtro contendo o resíduo foi transferido para o cadinho original e em
seguida, transferidos para uma mufla pré-aquecida a 500o C, incinerando-se o resíduo por 5
horas ou até formação de cinzas brancas. Os cadinhos foram resfriados em dessecador e
pesados.
A porcentagem de cinzas insolúveis em ácido clorídrico foi calculada em relação à
amostra inicial (COSTA, 2001).
Equação 3: Cálculo do teor de cinzas insolúveis em ácido clorídrico em
porcentagem
% teor de cinzas insolúveis em ácido clorídrico PxN100=
Onde:N – quantidade de cinzas insolúveis da amostra em gramas;P – quantidade da amostra em gramas.
2.4 Prospecção fitoquímica
A análise qualitativa das principais classes de metabólitos secundários presentes nas
folhas de C. densiflorus (Steud.) Stapf e C. nardus (L.) Rendle foi realizada na amostra
pulverizada.
As pesquisas qualitativas foram realizadas utilizando metodologias adaptadas de
MATOS (1988) e COSTA (2001), descritas a seguir.
2.4.1 Pesquisa de Heterosídeos Antraquinônicos
Extração:
Para a extração dos possíveis heterosídeos antraquinônicos presentes na amostra
pulverizada pesou-se, em balança semi-analítica, 1 g da amostra pulverizada e acrescentados
30 mL de etanol a 75% (v/v). A mistura foi aquecida durante 3 minutos em chapa aquecedora,
e, em seguida, filtrada ainda quente, utilizando-se papel de filtro.
Caracterização:
Para a caracterização dessa classe de metabólitos secundários transferiu-se 10 mL do
filtrado para um béquer de 40 mL que foi designado (I) e 10 mL para outro béquer que foi
designado (II). O conteúdo do béquer (I) foi acidificado com 0,5 mL de HCl a 10% (v/v) e

levado à fervura por 2 minutos em chapa aquecedora, sendo que com o conteúdo do béquer
(II) fez-se o mesmo procedimento, exceto a acidificação.
Os líquidos foram transferidos para tubos de ensaio designados de (I) e (II),
respectivamente, após o resfriamento, foram adicionados, a cada um, 10 mL de éter etílico P.
A., agitando-os levemente. Em seguida, separou-se 5 mL da fase etérea dos tubos (I) e (II), e
acrescentou-se 4 mL de amônia 50% (v/v) em cada um, deixando-os em repouso por 5
minutos. A coloração rósea a vermelha na fase amoniacal indica reação positiva para
heterosídeos antraquinônicos.
2.4.2 Pesquisa de Heterosídeos Cardioativos
Extração:
Para a extração dos possíveis heterosídeos cardioativos presentes na amostra
pulverizada foram pesados, em balança semi-analítica, 2,5 g da amostra pulverizada,
adicionou-se 25 mL de etanol a 50% (v/v) e 10 mL de solução de acetato de chumbo a 10%
(p/v) e levou-se à fervura por 4 minutos. Após o resfriamento, o volume foi completado para
25 mL com etanol a 50% (v/v) e filtrado. Ao filtrado, foram adicionados 15 mL de
clorofórmio P. A. (por duas vezes), separando-se em seguida, a fase clorofórmica.
Caracterização:
A caracterização dessa classe de metabólitos secundários foi realizada a partir das
seguintes reações:
2.4.2.1. Reação de Liebermann-Burchard (reação de caracterização do núcleo
esteróide e triterpenos): Foram transferidos 3 mL da fração clorofórmica para um tubo de
ensaio e levou-se à secura em banho-maria. Ao resíduo do tubo, foi adicionado 1 mL do
reagente de Liebermann-Burchard, recém preparado (1 mL de clorofórmio P. A., 1mL de
anidrido acético P. A. e 3 – 4 gotas de ácido sulfúrico concentrado). Deixou-se o tubo em
repouso por 5 minutos. O desenvolvimento de coloração acastanhada a esverdeada indica
reação positiva para heterosídeos cardioativos.
2.4.2.2. Reação de Keller-Kiliani (reação que detecta desoxi-açúcares): Foram
evaporados, até a secura, 5 mL da fração clorofórmica num tubo de ensaio em banho-maria.
Ao resíduo do tubo, foi adicionado um reagente recém-preparado que contém ácido acético
glacial P. A. e cloreto férrico 9% (p/v) na proporção de 3:0,1. Homogeneizou-se o conteúdo
do tubo e foi lentamente vertido para outro tubo de ensaio contendo 2 mL de ácido sulfúrico
concentrado. Foi

observado se houve desenvolvimento de um anel de coloração castanho-avermelhada na zona
de contato, assim como se houve aparecimento de coloração azul-esverdeada na camada
acética, que indica reação positiva para heterosídeos cardioativos.
2.4.2.3. Reação de caracterização do núcleo esteróide: Foram evaporados 3 mL da
fração clorofórmica até à secura numa cápsula de porcelana em chapa aquecedora. Foram
acrescentadas, ao conteúdo da cápsula, após resfriamento, 3 – 6 gotas de ácido fosfórico
concentrado. Observou-se o conteúdo, sob luz ultravioleta com comprimento de onda 365 nm,
onde o aparecimento de fluorescência indicou reação positiva para heterosídeos cardioativos.
2.4.2.4. Reação de Kedde (reação específica para o anel lactônico): Foram transferidos
6 mL da fração clorofórmica para um tubo de ensaio e levados à secura em banho-maria. Ao
resíduo do tubo acrescentou-se 2 mL de etanol 50% (v/v), 2 mL de água, 2 mL de reagente
ácido 3-5 dinitrobenzóico a 1% (p/v) recém-preparado em etanol a 96% (v/v) e 2 mL de
hidróxido de potássio a 10%. Após repouso de 5 minutos foi observado se houve o
desenvolvimento de uma coloração castanho-avermelhada a vermelho-violeta, que indica
reação positiva para heterosídeos cardioativos.
2.4.3 Pesquisa de Heterosídeos Flavonóides
Extração:
Para a extração dos possíveis heterosídeos flavonóides presentes na amostra
pulverizada, foram pesados em balança semi-analítica, 7 g da amostra e adicionou-se 60 mL
de etanol a 70% (v/v). Essa mistura foi fervida durante 5 minutos e filtrada em papel de filtro
umedecido com etanol a 70% (v/v).
Caracterização:
A partir do filtrado obtido, realizaram-se as seguintes reações a fim de caracterizar
essa classe de metabólitos secundários:
2.4.3.1. Reação de Shinoda: Foram transferidos 3 mL do filtrado para um tubo de
ensaio. Adicionou-se cerca de 1 cm de fita de magnésio fina e acrescentou-se cuidadosamente
1 mL de HCl concentrado. O aparecimento de coloração vermelha indica reação positiva para
os heterosídeos flavonóides (COSTA, 2001).
2.4.3.2. Reação com Ácido Sulfúrico Concentrado: Foram adicionados 3 mL da
solução extrativa numa cápsula de porcelana e deixou-se evaporar até a semi-secura. Juntou-
se 0,5 mL de ácido sulfúrico concentrado e observou-se sob luz ultravioleta com comprimento
de onda 365 nm, em que o aparecimento de fluorescência verde-amarelada indica reação

positiva para heterosídeos flavonóides (COSTA, 2001).
2.4.3.3. Reação com Hidróxidos Alcalinos: Foram transferidos 3 mL da solução
extrativa para um tubo de ensaio. Adicionou-se 1 mL de NaOH a 20% (p/v) e agitou-se o
tubo. De acordo com COSTA (2001), na reação com hidróxidos alcalinos, o desenvolvimento
de coloração amarela indica reações positivas para heterosídeos flavonóides.
2.4.3.4. Reação com Cloreto de Alumínio: Foram transferidos cerca de 5 mL da
solução extrativa para um béquer. Concentrou-se à metade e transferiu-se para um pedaço de
papel de filtro espalhando sobre toda a superfície. A seguir, umedeceu-se uma das regiões do
papel com solução de cloreto de alumínio a 5% (p/v) e observou-se sob luz ultravioleta com
comprimento de onda 365 nm quanto ao aparecimento de fluorescência. A ocorrência de
fluorescência que vai do amarelo ao azul-esverdeado, sob luz ultravioleta, indica reação
positiva (COSTA, 2001).
2.4.3.5. Reação com Cloreto Férrico: Foram transferidos 3 mL da solução extrativa
para um tubo de ensaio. Juntou-se 2 gotas de cloreto férrico a 4,5% (p/v) e observou-se o
aparecimento de coloração. O desenvolvimento de coloração azul, verde, marrom ou
vermelha indicam reações positivas para heterosídeos flavonóides (COSTA, 2001).
2.4.4 Pesquisa de Heterosídeos Saponínicos
Extração:
Para a extração dos possíveis heterosídeos saponínicos presentes na amostra
pulverizada, pesou-se em balança semi-analítica, 1 g da amostra em pó que foi adicionado a
um béquer contendo 100 mL de água destilada. Essa mistura foi levada à fervura em chapa
aquecedora por 5 minutos, adicionando durante a decocção, carbonato de sódio em solução
até a neutralização, que foi observada utilizando-se papel de tornassol como indicador. Em
seguida, a mistura foi filtrada em algodão e ao filtrado acrescentou-se água destilada até
completar um volume de 100 mL.
Caracterização:
Posteriormente, para a caracterização dessa classe de metabólitos secundários foi
montada uma bateria contendo 10 tubos de ensaio de tamanhos e diâmetros iguais. Os tubos
de ensaio foram marcados em duas graduações diferentes: a primeira correspondente a 10 mL
e a segunda 1 cm acima desta. Em seguida, foram adicionados ao primeiro tubo 1 mL de
solução
extrativa e 9 mL de água destilada. Ao segundo tubo, foram adicionados 2 mL de solução

extrativa e 8 mL de água destilada. Ao terceiro tubo foram adicionados 3 mL de solução
extrativa e 7 mL de água destilada e assim por diante. Ao último tubo foi adicionada apenas
solução extrativa (10 mL). Após esse procedimento, cada tubo foi vigorosamente agitado por
20 segundos e deixado em repouso por 10 minutos. Observou-se o desenvolvimento de
espuma persistente. Calculou-se o índice de espuma, que é a maior diluição, referente a 1g de
amostra, que nas condições do ensaio forma 1 cm de espuma, de acordo com COSTA (2001).
2.4.5 Pesquisa de Taninos
Extração:
Para a extração dos possíveis taninos presentes na amostra pulverizada, foram pesados
em balança semi-analítica 2 g da amostra e adicionados 50 mL de água destilada. A mistura
foi fervura durante 5 minutos em chapa aquecedora. Em seguida, a mistura ainda quente foi
filtrada em papel de filtro. Completou-se o volume do filtrado obtido para 100 mL e
procedeu-se a pesquisa de taninos.
Caracterização:
Foi montada uma bateria contendo 6 tubos de ensaio. A cada tubo, foram adicionados 5
mL da solução extrativa e em seguida, realizou-se as seguintes reações:
2.4.5.1. Reação com gelatina: Foram adicionadas ao primeiro tubo, 5 gotas de solução
de gelatina a 2,5% (p/v) em solução de cloreto de sódio a 5% (p/v). Segundo COSTA (2001),
o aparecimento de precipitado branco, indica reação positiva para taninos.
2.4.5.2. Reação com alcalóides: Foram adicionadas ao segundo tubo, 5 gotas de
solução de sulfato de quinino a 1% (p/v) em ácido sulfúrico a 5% (p/v). Ao terceiro tubo,
foram adicionadas 5 gotas de solução de brucina a 1% (p/v) em ácido sulfúrico 5% (p/v).
Segundo COSTA (2001), a presença de precipitado branco indica reação positiva para
taninos.
2.4.5.3. Reação com sais metálicos: Ao quarto tubo foram adicionadas 5 gotas de
acetato de cobre a 4% (p/v). Ao quinto tubo, foram acrescentadas 2 gotas de cloreto férrico a
2% (p/v). Segundo Costa (2001), o aparecimento de precipitado indica reação positiva para
fenóis.
2.4.5.4. Reação com Hidróxidos Alcalinos: Ao sexto tubo foram adicionadas 5 gotas
de solução de hidróxido de sódio ou potássio a 20% (p/v). De acordo com COSTA (2001), o
escurecimento da solução extrativa indica reação positiva para fenóis.
Para cada uma dessas reações, paralelamente, foi preparado um tubo-controle contendo

5 mL de ácido tânico 0,5% (p/v) e os reagentes da reação correspondente, a fim de comparar
o tubo-teste com o tubo-controle.
2.4.6 Pesquisa de Alcalóides
Extração:
Para a extração dos possíveis alcalóides presentes na amostra pulverizada, foram
pesado em balança semi-analítica 2 g da amostra e adicionados 20 mL de ácido sulfúrico a
5% (v/v). Essa mistura foi levada à fervura por 3 minutos em chapa aquecedora, em seguida,
filtrada em papel de filtro e resfriada.
Caracterização:
O filtrado foi submetido à pesquisa de alcalóides utilizando os reagentes gerais para
alcalóides de acordo com metodologias adaptadas de COSTA (2001). Abaixo estão descritos
os reagentes e suas respectivas técnicas de preparo:
2.4.6.1. Reativo de Mayer: Dissolver em água 2,71 g de cloreto de mercúrio e 10 g de
iodeto de potássio e, em seguida, completar o volume para 200 mL com água destilada. Agitar
e filtrar.
2.4.6.2. Reativo de Dragendorff: Dissolver 8 g de subnitrato de bismuto em 20 mL de
ácido nítrico a 30% (p/v). Dissolver, em separado, 22,8 g de iodeto de potássio em um volume
mínimo de água destilada. Adicionar a primeira solução, pouco a pouco, sobre a segunda.
Deixar em repouso durante alguns minutos e filtrar. Completar o volume com água destilada
para 100 mL.
2.4.6.3. Reativo de Bouchardat: Dissolver 4 g de iodeto de potássio e 2 g de iodo em
100 mL de água destilada.
2.4.6.4. Reativo de Bertrand: Dissolver 5 g de ácido sílico-túngstico em 100 mL de
água destilada.
2.4.6.5. Reativo de Hager: Dissolver 2 g de ácido pícrico em 100 mL de água
destilada.
2.4.6.6. Ácido Tânico: Dissolver 1 g de ácido tânico em 100 mL de água destilada.
A solução extrativa foi distribuída igualmente em 6 tubos de ensaio, sendo que em cada
tubo, respectivamente, foram acrescentadas 3 – 9 gotas dos reativos gerais para alcalóides
citados acima. Foi montada, em paralelo, uma outra bateria de 6 tubos de ensaio, contendo 3
mL de solução padrão de sulfato de quinina 1% (p/v). Em cada um destes tubos foram

adicionadas 3 – 9 gotas dos respectivos reativos gerais para alcalóides a fim de servirem de
padrão para comparação com a primeira bateria de tubos. Segundo COSTA (2001), o
aparecimento de precipitados nas amostras submetidas aos reagentes, indica reação positiva
para alcalóides.
2.4.7 Pesquisa de Cumarinas
Extração:
Para extração das possíveis cumarinas presentes na amostra pulverizada, foram
pesados 2 g da amostra em balança semi-analítica e adicionados a 30 mL de água quente.
Essa mistura foi filtrada, e ao filtrado adicionou-se 1 mL de HCl 1N (até pH 1). Em seguida,
procedeu-se a extração das cumarinas em funil de separação, com 10 mL de éter etílico P.A.
A fase etérea foi concentrada até a metade de seu volume.
Caracterização:
Para caracterização dessa classe de metabólitos secundários foram aplicadas gotas da
fase etérea obtida anteriormente sobre duas regiões de um papel de filtro. Em uma das
manchas formadas foi adicionada uma gota de NaOH 1N. O papel foi observado sob luz
ultravioleta com comprimento de onda 365 nm, o desenvolvimento de fluorescência azul-
brilhante ou verde, na região do papel contendo apenas a gota da fase etérea e fluorescência
amarelada, na gota da fase etérea com NaOH adicionado, indicam reação positiva para
cumarinas.

3 RESULTADOS
3.1 Teores de umidade, cinzas totais e cinzas insolúveis em ácido clorídrico
Os teores de umidade, cinzas totais e cinzas insolúveis em ácido clorídrico para as
folhas de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle estão apresentados
na Tabela 1.
Tabela 1. Teores de umidade, cinzas totais e cinzas insolúveis em ácido clorídrico das folhas
de C. densiflorus (Steud.) Stapf e C. nardus (L.) Rendle, expressos em porcentagem (p/p).
Espécie Umidade* Cinzas totais* Cinzas insolúveis em HCl*C. densiflorus (Steud.)
StapfAmostra 1 9,91%a 9,07%a 0,12%a
Amostra 2 9,81%a 8,17%a 0,12%a
C. nardus (L.) RendleAmostra 1 11,84%a 7,23%a 0,11%a
Amostra 2 12,31%b 6,78%b 0,11%a
* Médias obtidas a partir das triplicatas.
Médias seguidas pelas mesmas letras minúsculas não diferem estatisticamente entre si, até o
nível de 5% pelo teste t de Student.
3.2 Prospecção fitoquímica
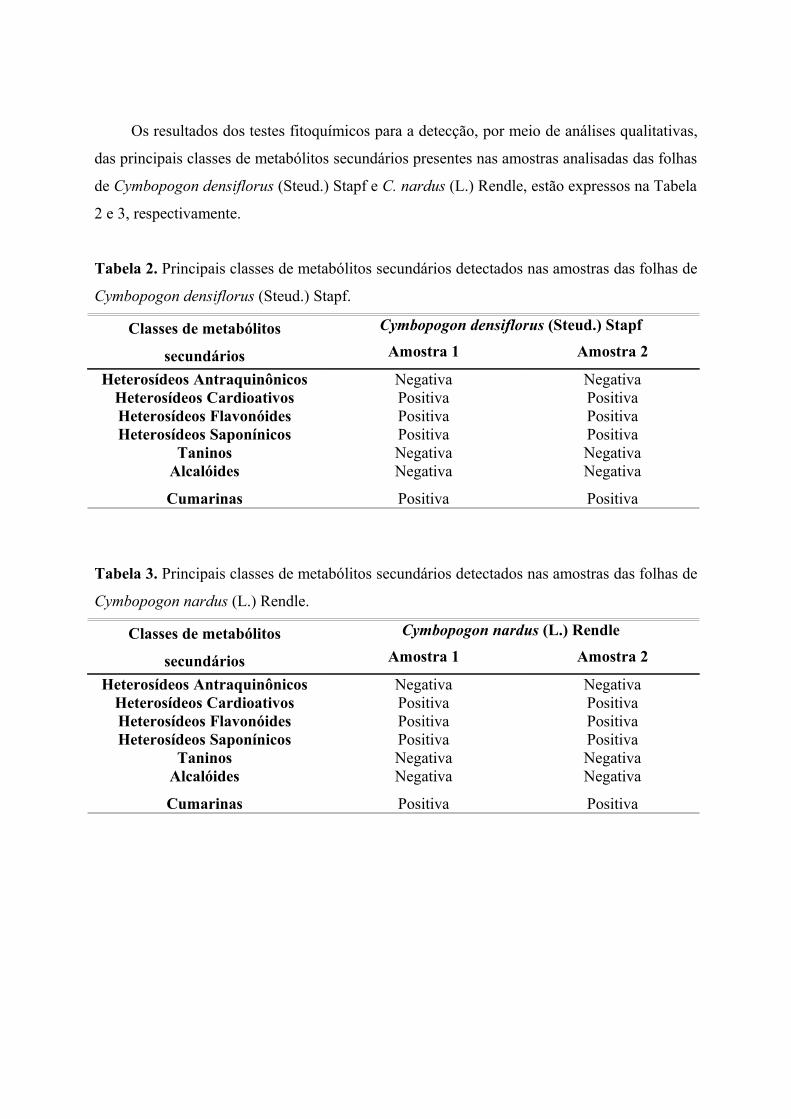
Os resultados dos testes fitoquímicos para a detecção, por meio de análises qualitativas,
das principais classes de metabólitos secundários presentes nas amostras analisadas das folhas
de Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle, estão expressos na Tabela
2 e 3, respectivamente.
Tabela 2. Principais classes de metabólitos secundários detectados nas amostras das folhas de
Cymbopogon densiflorus (Steud.) Stapf.
Classes de metabólitos
secundários
Cymbopogon densiflorus (Steud.) StapfAmostra 1 Amostra 2
Heterosídeos Antraquinônicos Negativa NegativaHeterosídeos Cardioativos Positiva PositivaHeterosídeos Flavonóides Positiva PositivaHeterosídeos Saponínicos Positiva Positiva
Taninos Negativa NegativaAlcalóides
Cumarinas
Negativa
Positiva
Negativa
Positiva
Tabela 3. Principais classes de metabólitos secundários detectados nas amostras das folhas de
Cymbopogon nardus (L.) Rendle.
Classes de metabólitos
secundários
Cymbopogon nardus (L.) RendleAmostra 1 Amostra 2
Heterosídeos Antraquinônicos Negativa NegativaHeterosídeos Cardioativos Positiva PositivaHeterosídeos Flavonóides Positiva PositivaHeterosídeos Saponínicos Positiva Positiva
Taninos Negativa NegativaAlcalóides
Cumarinas
Negativa
Positiva
Negativa
Positiva

As Tabelas 2 e 3 resumem os resultados observados e descritos a seguir. As
amostras de ambas as espécies de Cymbopogon densiflorus e C. nardus demonstraram
presença de heterosídeos cardioativos e presença de heterosídeos flavonóides,
heterosídeos saponínicos e cumarinas nos testes qualitativos.
Na pesquisa de heterosídeos cardioativos, em ambas as espécies, foi observado
que na reação de Liebermann-Burchard a solução extrativa corou-se em castanho. Na
reação de Keller-Kiliani, teve formação de um anel de coloração castanha na zona de
contato e a camada acética corou-se ligeiramente esverdeada. Na reação de
caracterização do núcleo esteróide se formou fluorescência verde, sob comprimento de
onda 365 nm. Na reação de Kedde, a solução extrativa de ambas as espécies apresentou
coloração avermelhada. Devido as colorações observadas nas reações descritas, pôde-se
concluir que há presença de heterosídeos cardioativos nas amostras de ambas as
espécies.
Na pesquisa de heterosídeos flavonóides, foi observado que em ambas as
espécies, na reação de Shinoda, a solução extrativa das amostras apresentou coloração
vermelho-tijolo. Na reação com ácido sulfúrico concentrado, forma-se fluorescência
verde sob comprimento de onda 365 nm. Na reação com os hidróxidos alcalinos, a
solução extrativa apresenta coloração amarelo-acastanhada. Na reação com cloreto de
alumínio, apresentou-se fluorescência amarela sob comprimento de onda 365 nm. Na
reação com cloreto férrico, a solução extrativa apresentou coloração verde marrom.
Na pesquisa de heterosideos saponínicos, em ambas as amostras de C.
densiflorus, a formação de espuma persistente ocorreu a partir de 6 mL de solução
extrativa, o índice de espuma calculado foi de 166,6 mL/g, enquanto que nas amostras
de C. nardus, a formação de espuma persistente se deu a partir de 5 mL de solução
extrativa e o índice de espuma calculado foi de 200 mL/g. De acordo com COSTA
(2001), a formação de espuma persistente indica a presença de saponinas na amostra.
Na pesquisa de cumarinas, foi observado que, em ambas as amostras de C.
densiflorus, na região do papel de filtro contendo a fase etérea e uma gota de NaOH 1N
formou-se fluorescência amarela sob comprimento de onda 365 nm, enquanto que em
C. nardus, verificou-se fluorescência amarelo-esverdeado sob comprimento de onda
365 nm. Segundo COSTA (2001), o desenvolvimento de fluorescência azul-brilhante ou
verde sob luz ultravioleta em comprimento de onda igual a 360 nm, indica reação
positiva para cumarinas.

4 DISCUSSÃO
Conforme descrito na tabela 1, as folhas de Cymbopogon densiflorus (Steud.)
Stapf. coletadas em indivíduos de Jataí-GO, as amostras 1 e 2, apresentaram 9,91%
(p/p) e 9,81% (p/p) de teor de umidade, respectivamente. As amostras de C. nardus (L.)
Rendle coletadas em indíviduo de Brasília-DF (amostra 1) e de Goiânia-GO (amostra
2), apresentaram 11,84% (p/p) e 12,31% (p/p) de teor de umidade, respectivamente.
A matéria-prima vegetal deve possuir o mínimo de umidade para que apresente
uma boa conservação. A presença excessiva de água em materiais vegetais propicia o
desenvolvimento de enzimas, acarretando na degradação de constituintes químicos,
além de possibilitar o desenvolvimento de microorganismos e insetos
(FARMACOPÉIA BRASILEIRA IV, 1988; OLIVEIRA et al., 1991; BACCHI, 1996;
FARIAS, 2004). A FARMACOPÉIA BRASILEIRA IV (1988) estabelece que os
limites de umidade para as drogas vegetais, em geral, deve encontrar-se na faixa de 8 a
14%. A metodologia adotada para a secagem das amostras analisadas neste trabalho foi
adequada e propiciou um teor de umidade para as folhas de C. densiflorus e C. nardus
dentro da faixa estabelecida pela literatura, permitindo uma boa conservação da matéria-
prima vegetal.
De acordo com a tabela 1, as amostras 1 e 2 de C. densiflorus coletadas em
indivíduos de Jataí-GO, apresentaram 9,07% e 8,17% de teor de cinzas totais,
respectivamente e 0,12 % de teor de cinzas insolúveis em ácido clorídrico em ambas
as amostras, enquanto que as amostras de C. nardus coletadas em indíviduo de Brasília-
DF (amostra 1) e de Goiânia-GO (amostra 2), apresentaram 7,23% (p/p) e 6,78% (p/p)
de teor de cinzas totais e 0,11 % (p/p) de teor de cinzas insolúveis em ácido clorídrico
em ambas as amostras.
Segundo a FARMACOPÉIA BRASILEIRA IV (1988), os valores encontrados
que fornecem parâmetros para o teor de cinzas totais para as amostras analisadas,
indicam em porcentagem, a quantidade de material residual derivada de tecidos vegetais
(cinzas fisiológicas) e de materiais estranhos, especialmente areia e terra que se
encontravam aderentes à superfície do material vegetal analisado (cinzas não-
fisiológicas). A determinação do teor de cinzas destina-se estabelecer a quantidade de
impurezas não-voláteis resultante do processo de incineração e que podem estar
presentes como agentes contaminantes (FARMACOPÉIA BRASILEIRA IV, 1988;
FARIAS, 2004).

Verifica-se que esses teores de umidade e teores de cinzas totais obtidos das
amostras de C. nardus coletadas em duas localidades distintas (Brasília-DF e Goiânia-
GO) tiveram diferenças significativas em 5%, no teste de Student. Dessa forma, tais
diferenças podem ser resultantes das distintas concentrações de sílica presentes no solo,
nas localidades em que essas amostras foram coletadas.
As cinzas insolúveis em ácido clorídrico permitem verificar, por exemplo, a
presença de agentes contaminantes como resíduos de terra ou areia, além de determinar
sílicas e constituintes silicosos da droga (FARMACOPÉIA BRASILEIRA IV, 1988;
FARIAS, 2004).
Muitas amostras comercializadas por raizeiros e ervanários apresentam-se
desqualificadas, com excesso de matéria orgânica desconhecida, geralmente
constituindo de outros órgãos do próprio vegetal, diferentes daquele que constitui a
matéria-prima. Tais impurezas podem indicar tanto falta de cuidados no preparo da
matéria-prima, bem como fraude intencional (DUARTE e BARDAL, 2002). Sendo
assim, os diferentes resultados nos teores de cinzas totais e cinzas insolúveis em ácido
clorídrico obtidos a partir das folhas de C. densiflorus e C. nardus analisadas, podem
estar relacionados com as diferentes localidades em que as amostras se encontravam.
Entretanto, tais resultados fornecem subsídios para o estabelecimento de um limite para
este parâmetro de qualidade (PAULA, 2006).
As análises fitoquímicas realizadas foram apenas qualitativas e são, portanto,
preliminares. Ambas as espécies estudadas, C. densiflorus e C. nardus apresentaram as
mesmas classes de metabólitos secundários, nas duas amostras coletadas em localidades
geográficas diferentes.
Os metabólitos secundários, por definição, são aqueles não essenciais para o
crescimento e desenvolvimento de uma planta. Por muito tempo, acreditava-se que
esses metabólitos eram produtos de excreção do vegetal. Atualmente, sabe-se que os
metabólitos secundários têm importante função nos mecanismos de interação e
adaptação do vegetal às condições ambientais. Tais componentes químicos são
amplamente determinados por fatores genéticos e ambientais (MILLS e BONE, 2001;
SCHULZ et al., 2002; FOLEY e MOORE, 2005; KUTCHAN e DIXON, 2005). No
entanto, alguns componentes químicos são muito característicos para um determinado
vegetal, podendo então, servir como parâmetro para sua caracterização e identificação
(MIGLIATO et al., 2007).

Os heterosídeos cardioativos são esteróides e têm este nome porque agem sobre
o músculo cardíaco, sendo então recomendados para o tratamento de insuficiência
cardíaca. Heterosídeos cardioativos geralmente são tóxicos devido ao seu baixo índice
terapêutico, pois a concentração suficiente para causar efeitos tóxicos é duas vezes
maior que a concentração terapêutica. Na medicina popular, os heterosídeos
cardioativos foram
utilizados por muitos anos, como diurético, laxativo, emético, anti-malárico,
antieplilético e também para picadas de insetos. Atualmente há controvérsias a respeito
das propriedades terapêuticas dos heterosídeos cardioativos, pois a toxicidade em
conjunto com a dificuldade
em controlar a dosagem, fez o uso popular ser totalmente desaconselhado (MILLS e
BONE, 2001; RATES e BRIDI, 2004).
Os flavonóides formam um grupo de pigmentos vegetais de ampla distribuição e
diversidade na natureza e sua presença nas plantas relaciona-se com funções de defesa e
de atração de polinizadores (SANTOS, 2004). São heterosídeos possuindo um açúcar
diferente da glicose ligado a uma fração aglicona, que corresponde a um pigmento. As
ações farmacológicas dos flavonóides são: antiinflamatória, antiespasmódica,
antitumoral, anti-hemorrágicos, anitmicrobianos, antioxidante, antiviral, entre outras
(ALVES e SILVA, 2002; ZUANAZZI e MONTANHA, 2004).
As saponinas são glicosídeos de esteróides e formam soluções coloidais em
água, por possuir uma parte lipofílica (triterpeno ou esteróide) e outra parte hidrofílica
(açúcar), que determinam a propriedade de redução da tensão superficial. Em solução
aquosa, as saponinas quando agitadas criam espumas persistente e abundante,
apresentam sabor amargo e acre, e os medicamentos que as contêm, geralmente são
esternutatórios e irritantes para as mucosas (ROBBERS et al., 1997; MILLS e BONE,
2001). Drogas vegetais que contêm saponinas são tradicionalmente utilizadas devido à
ação mucolítica, expectorante, diurética, anti-séptica, antimicrobiana e antiinflamatória
deste grupo de metabólitos (ALVES e SILVA, 2002). Por apresentar comportamento
anfifílico e formar complexos com esteróides, proteínas e fosfolipídeos, as saponinas
agem com as membranas celulares, alterando a permeabilidade e destruindo-a
(SCHENKEL et al., 2004).
A palavra ‘cumarina’ origina-se de uma árvore denominada vulgarmente de
cumaru (Dypterix odorata (Aublet) Willdenow), da família Fabaceae. Encontrada
amplamente na natureza, ocorre sob a forma de cristais prismáticos solúvel em álcool,

com odor fragrante e sabor ardente, aromático e amargo. A cumarina e seus derivados
são utilizados por suas propriedades anticoagulantes, vasodilatadora, antiespasmódica e
antifúngica. Há estudos que também atribuem atividades antiinflamatória,
anticonvulsivante, antileucêmica e antidepressiva às cumarinas (ROBBERS et al., 1997;
KUSTER e ROCHA, 2004).
Na medicina popular, as folhas de C. densiflorus são utilizadas para o tratamento
de diversas afecções como: asma, febre, resfriado, epilepsia, câimbras e dores
abdominais (TAKAISI-KIKUNI, 1996). Segundo FILGUEIRAS (2005), as folhas de C.
nardus apresentam usos medicinais, no entanto, as recomendações não são esclarecidas
pelo autor. De acordo com CAVALINI et al. (2005), C. nardus é empregada para o
tratamento de dores musculares e no trato gastrointestinal, neuralgia, distúrbios
nervosos e condições de exaustão.
A eficácia destas espécies para tais tratamentos ainda não é comprovada
cientificamente e futuros ensaior quantitativos, farmacológicos e toxicológicos deverão
ser realizados para confirmar o potencial terapêutico de ambas.

5 CONCLUSÕES
Os teores de umidade e de cinzas totais e insolúveis, registrados para as espécies
estudadas constituem parâmetros para o controle de qualidade da matéria-prima vegetal.
Os testes qualitativos da prospecção fitoquímica demonstraram que
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle apresentam potencial
fitoterápico que merece investigações futuras mais aprofundadas. Análises
quantitativas, testes farmacológicos e toxicológicos serão importantes para comprovar
cientificamente a eficácia dessas espécies.
6 REFERENCIAS BIBLIOGRÁFICAS (*)

ADAMU, H.M.; ABAYED, O.J.; AGHO, M.O.; ABDULLAHI, A.L.; UBÁ, A.;
DUKKU, H.U.; WUFEM, B.M. An ethnobotanical survey of Bauchi State herbal plants
and their antimicrobial activity. Journal of Ethnopharmacology. 99: 1-4. 2005.
ALVES, D.L.; SILVA, C.R. Fitohormônios: Abordagem natural da terapia
hormonal. São Paulo: Atheneu. 2002.
AMARAL, F.M.F.; COUTINHO, D.F.; RIBEIRO, M.N.S.; OLIVEIRA, M.A.
Avaliação da qualidade de drogas vegetais comercializadas em São Luís/Maranhão.
Revista Brasileira de Farmacognosia. 13(supl.): 27-30. 2003.
BACCHI, E.M. Controle de qualidade de fitoterápicos. In: DI STASI, L.C. (Org.).
Plantas medicinais: arte e ciência. Um guia de estudo interdisciplinar. São Paulo:
Editora da Universidade Estadual Paulista, p 169-186. 1996.
BRANDÃO, M.G.L.; FREIRE, N.; VIANNA-SOARES, C.D. Vigilância de
fitoterápicos em Minas Gerais. Verificação da qualidade de diferentes amostras
comerciais de camomila. Cadernos de Saúde Pública. 14(3): 613-616. 1998.
BUENO, N.R.; CASTILHO, R.O.; COSTA, R.B.; POTT, A.; POTT, V.J.; SCHEIDT,
G.N.; BATISTA, M.S. Medicinal plants used by the Kaiowá and Guarani indigenous
populations in the Caarapó Reserve, Mato Grosso do Sul, Brazil. Acta Botanica
Brasílica. 19(1): 39-44. 2005.
CAVALINI, M.; FOLIS, G.P.; RESENER, M.; ALEXANDRE, R.F.; ZANNIN, M.;
SIMÕES, C.M.O. Serviço de informações sobre plantas medicinais e medicamentos
fitoterápicos. Revista Eletrônica de Extensão (Extensio). 2:1-11. 2005.
COSTA, A.F. Farmacognosia. 3 v. 3ª ed. Lisboa: Calouste Gulbenkian. 2001.
______________________________________________________________________
(*) Referências bibliográficas conforme ABNT/NBR-6023: 2002.
DUARTE, M.R.; BARDAL, D. Qualidade de amostras vegetais comercializadas em
Curitiba-PR. Visão Acadêmica. 3 (2): 65-68. 2002.

FARIAS, M.R. Avaliação da qualidade de matérias-primas vegetais. In: SIMÕES,
C.M.O.; SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.;
PETROVICK, P.R (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS, p 263-288. 2004.
FARMACOPÉIA BRASILEIRA. Parte 1. 4.ed. São Paulo: Atheneu. 1988.
FOLEY, W.J.; MOORE, B.D. Plant secondary metabolites and vertebrate herbivores –
from physiological regulation to ecosystem function. Current Opinion in Plant
Biology. 8: 430-435. 2005.
KATEWA, S.S.; CHAUDHARY, B.L.; JAIN, A. Folk herbal medicines from tribal area
of Rajasthan, India. Journal of Ethnopharmacology. 92: 41-46. 2004.
KLOUCEK, P.; POLESNY, Z.; SVOBODOVA, B.; VLKOVA, E.; KOKOSKA, L.
Antibacterial screening of some Peruvian medicinal plants used in Callería District.
Journal of Ethnopharmacology. 99: 309-312. 2005.
KUSTER, R.M.; ROCHA, L.M. Cumarinas. In: SIMÕES, C.M.O.; SCHENKEL, E.P.;
GOSMANN, G. MELLO, J.C.P.; MENTZ, L.A.; PETROVICK, P.R. (Org.).
Farmacognosia: da planta ao medicamento. 5ª ed. Porto Alegre/Florianópolis:
UFRGS, p. 538-575. 2004.
KUTCHAN, T.; DIXON, R.A. Secondary metabolism: nature’s chemical reservoir
under deconvolution. Current Opinion in Plant Biology. 8: 227-229. 2005.
MACÍA, M.J.; GARCÍA, E.; VIDAURRE, P.J. An ethnobotanical survey of medicinal
plants commercialized in the markets of La Paz and El Alto, Bolívia. Journal of
Ethnopharmacology. 97: 337-350. 2005.
MACIEL, M.A.M.; PINTO, A.C.; VEIGA JR., V.F.; GRYNBERG, N.F.;
ECHEVARRIA, A. Plantas medicinais: a necessidade de estudos multidisciplinares.
Química Nova. 25(3): 429-438. 2002.

MATOS, F.J.A. Introdução à Fitoquímica Experimental. Fortaleza: UFC. 1988.
MIGLIATO, K.F.; MOREIRA, R.R.D.; MELLO, J.C.P.; SACRAMENTO, L.V.S.;
CORRÊA, M.A.; SALGADO, H.R.N. Controle da qualidade do fruto de Syzygium
cumini (L.) Skeels. Revista Brasileira de Farmacognosia. 17 (1): 94-101. 2007.
MILLS, S..; BONE, K. Principles and Practice of Phytotherapy. 1ª reimpressão.
Churchill Livingstone. 2001.
MONTANHER, A.B.P.; PIZZOLATTI, M.G.; BRIGHENTE, I.M.C. Monitoramento
dos extratos brutos de espécies de Polygala (Polygalaceae) utilizando Artemia salina.
Revista Brasileira de Farmacognosia. 13(supl.): 27-30. 2003.
NUNES, G.P.; HOLETZ, F.B.; SANCHES, N.R.; CORTEZ, D.A.G.; DIAS FILHO,
B.P.; NAKAMURA, C.V. Plantas medicinais comercializadas por raizeiros no centro de
Campo Grande, Mato Grosso do Sul. Revista Brasileira de Farmacognosia. 13(2): 83-
92. 2003.
OLIVEIRA, F.; AKISUE, G.; AKISUE, M.K. Farmacognosia. São Paulo: Atheneu.
1991.
PAULA, J.A.M. Estudo farmacognóstico e avaliação antimicrobiana das folhas de
Pimenta pseudocaryophyllus (Gomes) L.R. Landrum – Myrtaceae. Dissertação
(Mestrado em Biologia Celular e Molecular) – Universidade Federal de Goiás, Goiânia-
GO. 2006.
RATES, S.M.K.; BRIDI, R. Heterosídeos cardioativos In: SIMÕES, C.M.O.;
SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.; PETROVICK,
P.R. (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS, p.685-710. 2004.

ROBBERS, J.E.; SPEEDIE, M.K.; TYLER, V.E. Farmacognosia e
Farmacobiotecnologia. São Paulo: Editorial Premier. 1997.
SANTOS, R.I. Metabolismo básico e origem dos metabólitos secundários In: SIMÕES,
C.M.O.; SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.;
PETROVICK, P.R. (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS, p.403-434. 2004.
SCHENKEL, E.P.; GOSMANN, G.; ATHAYDE, M.L. Saponinas In: SIMÕES,
C.M.O.; SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.;
PETROVICK, P.R. (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS, p.711-740. 2004.
SCHULZ, V.; HÄNSEL, R.; TYLER, V.E. Fitoterapia racional: Um guia de
fitoterapia para as ciências da saúde. 4ª ed. (1ª ed. brasileira). São Paulo: Manole.
2002.
SEIDL, P.R. Pharmaceuticals from natural products: current trends. Anais da
Academia Brasileira de Ciências. 74(1): 145-150. 2002.
SOEJARTO, D.D.; FONG, H.H.S.; TAN, G.T.; ZHANG, H.J.; MA, C.Y.;
FRANZBLAU, S.G.; GYLLENHAAL, C.; RILEY, M.C.; KADUSHIN, M.R.;
PEZZUTO, J.M.; XUAN, L.T.; HIEP, N.T.; HUNG, N.V.; VU, B.M.; LOC, P.K.;
DAC, L.X.; BINH, L.T.; CHIEN, N.Q.; HAI, N.V.; BICH, T.Q.; CUONG, N.M.;
SOUTHAVONG, B.; SYDARA, K.; BOUAMANIVONG, S.; LY, H.M.; THUY, T.V.;
ROSE, W.C.; DIETZMAN, G.R. Ethnobotany/ethnopharmacology and mass
bioprospecting: Issues on intellectual property and benefit-sharing. Journal of
Ethnopharmacology. 100: 15-22. 2005.
STAFFORD, G.I.; JÄGER, A.K.; STADEN, J.V. Effect of storage on the chemical
composition and biological activity of several Popular South African medicinal plants.
Journal of Ethnopharmacology. 97: 107-115. 2005.

VILA VERDE, G.M.; PAULA, J.R.; CARNEIRO, D.M. Levantamento etnobotânico
das plantas medicinais do cerrado utilizadas pela população de Mossâmedes (GO).
Revista Brasileira de Farmacognosia. 13(supl.): 64-66. 2003.
ZUANAZZI, J.A.S.; MONTANHA, J.A. Flavonóides. In: SIMÕES, C.M.O.;
SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.; PETROVICK,
P.R. (Org.). Farmacognosia: da planta ao medicamento. 5ª ed. Porto
Alegre/Florianópolis: UFRGS, p.578-614. 2004.

____________________________________________________________
CAPÍTULO 3
ANÁLISE DO ÓLEO ESSENCIAL EXTRAÍDO DAS FOLHAS DE
Cymbopogon densiflorus (Steud.) Stapf e C. nardus (L.) Rendle
(Poaceae: Panicoideae)
____________________________________________________________
1 INTRODUÇÃO
Os óleos essenciais, conhecidos como óleos aromáticos, óleos perfumados, óleos
etéreos e óleos voláteis, são extensivamente utilizados na indústria farmacêutica, como

aromatizantes, cosmética e em aromaterapia. São extraídos de diferentes partes das
plantas aromáticas e suas utilizações estão aplicadas às diferentes atividades biológicas
reconhecidas (CIMANGA et al., 2002; KANKO et al., 2004; RAO et al., 2005). Além
disso, os óleos essenciais apresentam atividade antimicrobiana contra diversas bactérias,
incluindo as espécies resistentes aos antibióticos e fungos (PRASHAR et al., 2003).
Óleos voláteis ou essenciais evaporam quando expostos ao ar em temperatura
ambiente e são compostos de aroma forte, geralmente agradável, oriundos do
metabolismo secundário. São insolúveis em água, porém solúveis em solventes
orgânicos, sendo então extraídos por técnicas como o arraste a vapor, por exemplo
(ROBBERS et al., 1997). Quanto às funções biológicas, os óleos essenciais encontrados
nas plantas estão relacionados com seu alto grau de volatilidade, que por meio desse
fator, atua como sinal de comunicação química planta-animal, atuando como forma de
defesa ou atração. Assim, pode-se citar: inibição da germinação, proteção contra
predadores, atração de polinizadores, entre outros (FAHN, 1979; SIMÕES e SPITZER,
2004).
A importância dos óleos essenciais para a indústria farmacêutica consiste nas
atividades farmacológicas que estes compostos possuem, tais como: antisséptica,
antiinflamatória, anti-helmíntica, analgésica, anestésica, sedativa, expectorante,
estimulante e estomáquica, que são muito utilizadas na medicina popular e na
fabricação de medicamentos (ALVES e SILVA, 2002). Diante dessas importâncias,
inúmeras pesquisas com óleos essenciais vêm sendo realizadas, seja em técnicas de
extração bem como em seus constituintes (SOUZA et al., 2005; EHLERT et al., 2006).
Dentre várias famílias, com suas espécies produtoras de substâncias aromáticas,
destacam-se as do gênero Cymbopogon Sprengel. As espécies do gênero Cymbopogon
são comumente conhecidas como capins aromáticos e compreende espécies altamente
tolerantes ao estresse, bem como adaptadas às diversas condições climáticas e edáficas,
ocorrendo amplamente nos trópicos e subtrópicos. Dentre as 55 espécies que existem no
gênero, sabe-se que aproximadamente cinco são cultivadas comercialmente para a
extração de óleos: Cymbopogon. nardus (L.) Rendle, C. winteranus Jowitt, C. flexuosus
(Nees ex Steud.) Wats., C. citratus (DC.) Stapf e C. martinii (Roxb.) Wats.
(SOENARKO, 1977; SANGWAN
et al., 2001), sendo que C. citratus é a espécie mais estudada em diversos aspectos, bem
como a de uso medicinal e industrial mais conhecida.

Cymbopogon citratus apresenta óleo essencial amplamente utilizado na
medicina e na indústria, e o principal constituinte é o citral (47-85%), composto por
uma mistura dos isômeros neral e geranial. Aplica-se ao citral grande parte das
atividades atribuídas ao óleo, como anti-bacteriano, anti-fungicida e anti-mutagênica,
além do uso farmacológico, síntese de vitamina A e de compostos químicos
denominados iononas, que são utilizados na perfumaria (DUARTE e ZANETI, 2004;
MARTINS et al., 2004; RAUBER et al., 2005; PAVIANI et al., 2006; QUIALA et al.,
2006; MARTINAZZO et al, 2007).
Nessa perspectiva, esta pesquisa teve como objetivo, analisar o teor e a
composição química do óleo essencial das folhas de Cymbopogon densiflorus (Steud.)
Stapf. e C. nardus (L.) Rendle.

2 MATERIAL E MÉTODOS
2.1 Material botânico
Para a análise dos óleos essenciais de folhas de Cymbopogon densiflorus
(Steud.) Stapf, e C. nardus (L.) Rendle, foram coletadas duas amostras de cada espécie.
As amostras de C. densiflorus foram coletadas em duas localidades diferentes do
município de Jataí-GO, (17°52’14,0”S e 051°43’14,9” W) (17°52’07,3’’ S e
51°43”18,8’’ W) – denominadas C. densiflorus amostras 1 e 2, respectivamente e C.
nardus (L.) Rendle, na Reserva Ecológica do IBGE-RECOR, Brasília-DF (15°56’41’’ S
e 47°56’07’’ W) e do Horto de Plantas Medicinais da Faculdade de Farmácia - UFG,
Goiânia-GO (16°48’33,3’’S 49°14’39,5’’ W) – denominadas C. nardus amostras 1 e 2 .
Em todos os indivíduos, as amostras foram coletadas durante a época em que as
inflorescências encontravam-se maduras, no período de junho a setembro de 2006. As
exsicatas das espécies estão depositadas no Herbário UFG, da Universidade Federal de
Goiás sob os números 30.334 (C. densiflorus amostra 1), 30.335 (C. densiflorus amostra
2), 30.333 (C. nardus amostra 1) e 30.336 (C. nardus amostra 2).
Após coletadas, as folhas foram dessecadas ao ar livre e, em seguida
transportadas para o Laboratório de Farmacognosia, da Faculdade de Farmácia, da
Universidade Federal de Goiás e trituradas em moinho de facas. O pó assim obtido foi
identificado, acondicionado e armazenado até ser utilizado na extração do óleo
essencial.
2.2 Extração dos óleos essenciais
Para a extração do óleo essencial, 50 g de cada amostra pulverizada foram
submetidos à hidrodestilação em aparelho tipo Clevenger por 2 horas. O volume foi
medido no tubo graduado do próprio aparelho e o rendimento, em porcentagem, foi
calculado em relação à quantidade inicial de amostra pulverizada utilizada na extração.
Os óleos essenciais assim extraídos foram acondicionados em pequenos frascos de
vidro, livres de impurezas, hermeticamente fechados, identificados e estocados em
refrigerador até a utilização.

2.3 Análise da composição química dos óleos essenciais
O óleo essencial de cada espécie vegetal foi submetido à análise cromatográfica,
em fase gasosa, acoplada à espectrometria de massas (GC/MS) em aparelho
SHIMADZU QP5050A, no Laboratório de Produtos Naturais, do Instituto de Química,
Universidade Federal de Goiás. Foi utilizada uma coluna capilar de sílica fundida (CBP
– 5; 30 m x 0,25 mm x 0,25 µm), manteve um fluxo de 1 mL/min de Hélio, como gás de
arraste, aquecimentos com temperatura programada (60°C/2 min; 3°C min-1/240°C;
10°C min-1/ 280°C; 280°C/10 min), e energia de ionização de 70 eV. O volume de
injeção foi 1 µL de cada amostra diluída em CH2Cl2 na proporção de 1:5.
A partir dos cromatogramas obtidos e tempos de retenção, os índices de retenção
puderam ser calculados através da conjeção de uma mistura de hidrocarbonetos, C9 –
C28, e utilizando-se a equação de VAN DEN DOOL e KRATZ (1963).
Equação 1. Equação de Van Den Dool e Kratz
IR =100.N [(tx-tn-1)/(tn-tn-1)] + 100.Cn-1
Onde:
N= Cn – Cn-1
Cn = número de carbonos do n-alcano que elui após a substância analisada
Cn-1 = número de carbonos do n-alcano que elui antes da substância analisada
tx = tempo de retenção da substância analisada
tn = tempo de retenção do n-alcano que elui após a substância analisada
tn-1 = tempo de retenção do n-alcano que elui antes da substância analisada
Os componentes químicos dos óleos essenciais foram identificados por
comparação dos espectros de massas e índices de retenção com os relatados na literatura
para os componentes mais comuns de óleos essenciais (ADAMS, 2007).

3 RESULTADOS
3.1 RENDIMENTO DOS ÓLEOS ESSENCIAIS
Os
rendimentos dos óleos essenciais, em porcentagem (V/p), calculados a partir das
amostras 1 e 2 de C. densiflorus e amostras 1 e 2 de C. nardus, empregadas na extração
encontram-se expressos nas Tabelas 1 e 2, respectivamente.
Tabela 1. Rendimento de óleo essencial em porcentagem (V/p) das amostras de
Cymbopogon densiflorus (Steud.) Stapf coletadas em duas localidades diferentes de
Jataí-GO.
Amostras
Cymbopogon densiflorusRendimento de óleo essencial
Amostra 1 0,4Amostra 2 0,5
Tabela 2. Rendimento de óleo essencial em porcentagem (V/p) das amostras de
Cymbopogon nardus (L.) Rendle coletadas na Reserva Ecológica do IBGE-RECOR,
Brasília-DF (amostra 1) e Horto de Plantas Medicinais da Faculdade de Farmácia -
UFG, Goiânia-GO (amostra 2).
Amostras
Cymbopogon nardusRendimento de óleo essencial
Amostra 1 0,5Amostra 2 0,5

3.2 ANÁLISE DA COMPOSIÇÃO QUÍMICA DOS ÓLEOS ESSENCIAIS
3.2.1 Análise da composição química dos óleos essenciais de Cymbopogon
densiflorus (Steud.) Stapf.
A Cromatografia Gasosa acoplada à Espectrometria de Massa (GC/MS),
permitiu detectar 29 constituintes no óleo essencial da amostra 1 de C. densiflorus (Fig.
1), destes 24 constituintes foram identificados (98,56%) e na amostra 2, foram
detectados 28 constituintes (Fig. 2), destes 23 foram identificados (98,78%). Os
constituintes identificados em ambas as amostras e as suas classes estão presentes nas
Tabelas 3 e 4, respectivamente.
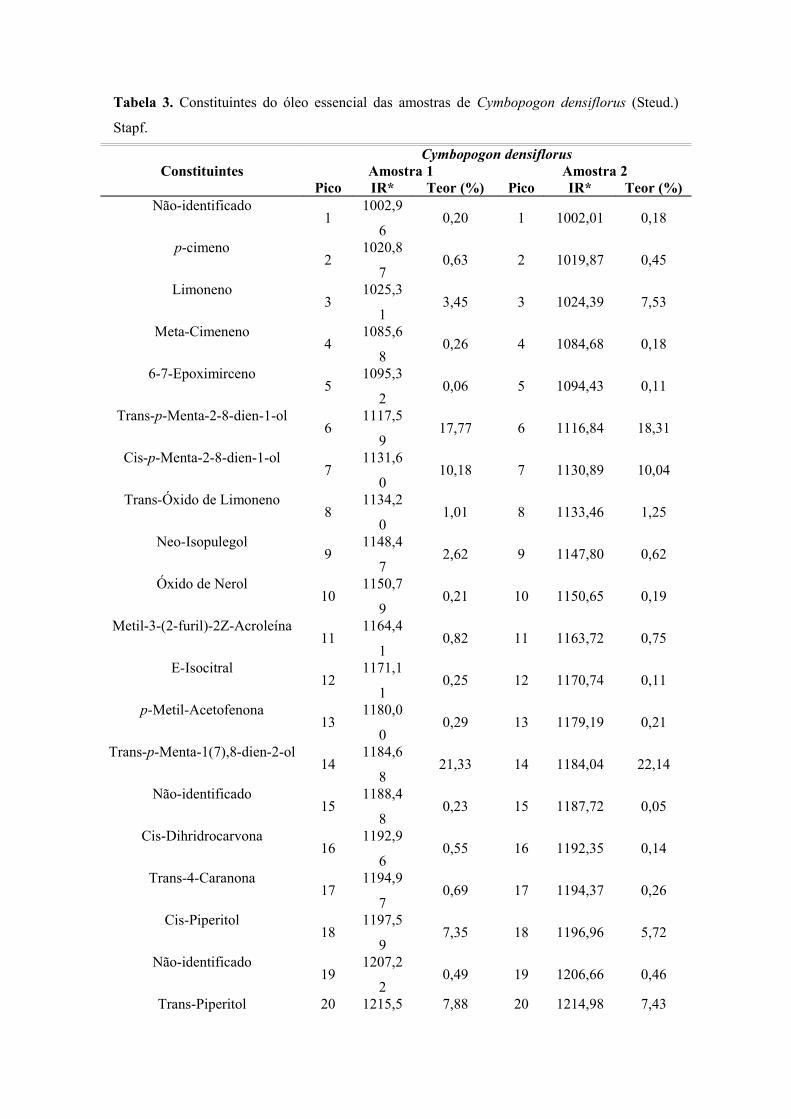
Tabela 3. Constituintes do óleo essencial das amostras de Cymbopogon densiflorus (Steud.)
Stapf.
ConstituintesCymbopogon densiflorus
Amostra 1 Amostra 2Pico IR* Teor (%) Pico IR* Teor (%)
Não-identificado1
1002,9
60,20 1 1002,01 0,18
p-cimeno2
1020,8
70,63 2 1019,87 0,45
Limoneno3
1025,3
13,45 3 1024,39 7,53
Meta-Cimeneno4
1085,6
80,26 4 1084,68 0,18
6-7-Epoximirceno5
1095,3
20,06 5 1094,43 0,11
Trans-p-Menta-2-8-dien-1-ol6
1117,5
917,77 6 1116,84 18,31
Cis-p-Menta-2-8-dien-1-ol7
1131,6
010,18 7 1130,89 10,04
Trans-Óxido de Limoneno8
1134,2
01,01 8 1133,46 1,25
Neo-Isopulegol9
1148,4
72,62 9 1147,80 0,62
Óxido de Nerol10
1150,7
90,21 10 1150,65 0,19
Metil-3-(2-furil)-2Z-Acroleína11
1164,4
10,82 11 1163,72 0,75
E-Isocitral12
1171,1
10,25 12 1170,74 0,11
p-Metil-Acetofenona13
1180,0
00,29 13 1179,19 0,21
Trans-p-Menta-1(7),8-dien-2-ol14
1184,6
821,33 14 1184,04 22,14
Não-identificado15
1188,4
80,23 15 1187,72 0,05
Cis-Dihridrocarvona16
1192,9
60,55 16 1192,35 0,14
Trans-4-Caranona17
1194,9
70,69 17 1194,37 0,26
Cis-Piperitol18
1197,5
97,35 18 1196,96 5,72
Não-identificado19
1207,2
20,49 19 1206,66 0,46
Trans-Piperitol 20 1215,5 7,88 20 1214,98 7,43
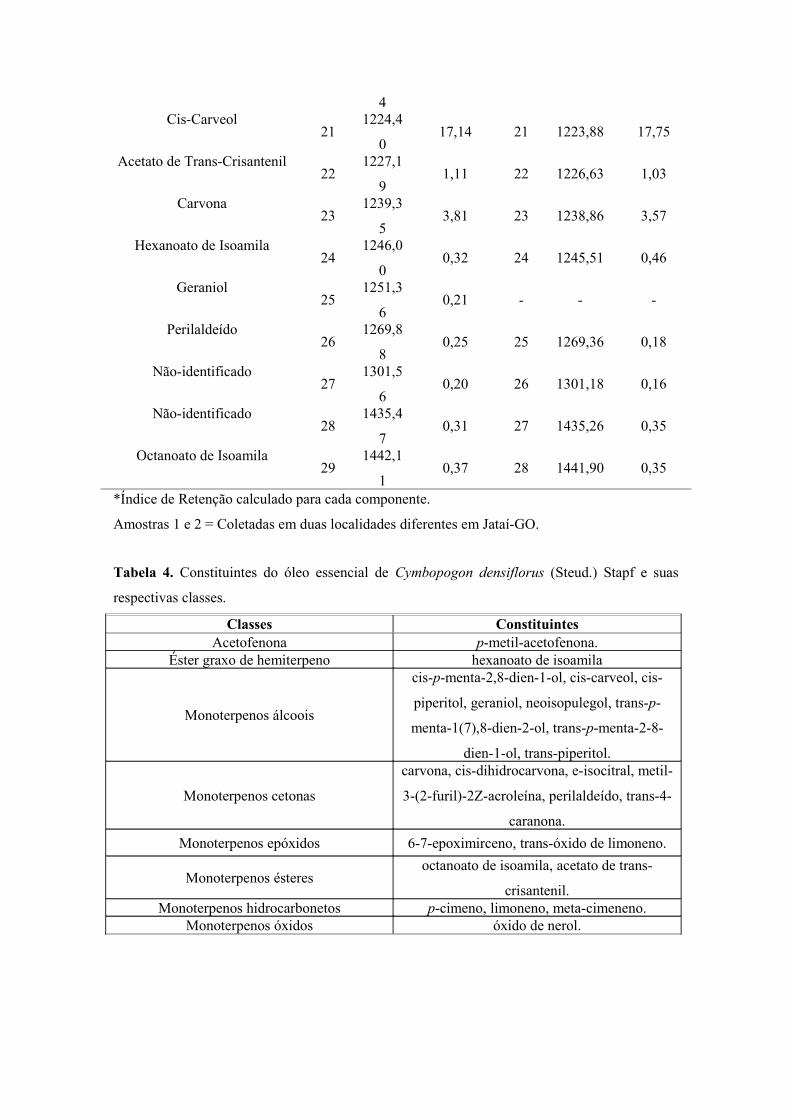
4Cis-Carveol
211224,4
017,14 21 1223,88 17,75
Acetato de Trans-Crisantenil22
1227,1
91,11 22 1226,63 1,03
Carvona23
1239,3
53,81 23 1238,86 3,57
Hexanoato de Isoamila24
1246,0
00,32 24 1245,51 0,46
Geraniol25
1251,3
60,21 - - -
Perilaldeído26
1269,8
80,25 25 1269,36 0,18
Não-identificado27
1301,5
60,20 26 1301,18 0,16
Não-identificado28
1435,4
70,31 27 1435,26 0,35
Octanoato de Isoamila29
1442,1
10,37 28 1441,90 0,35
*Índice de Retenção calculado para cada componente.
Amostras 1 e 2 = Coletadas em duas localidades diferentes em Jataí-GO.
Tabela 4. Constituintes do óleo essencial de Cymbopogon densiflorus (Steud.) Stapf e suas
respectivas classes.
Classes ConstituintesAcetofenona p-metil-acetofenona.
Éster graxo de hemiterpeno hexanoato de isoamila
Monoterpenos álcoois
cis-p-menta-2,8-dien-1-ol, cis-carveol, cis-
piperitol, geraniol, neoisopulegol, trans-p-
menta-1(7),8-dien-2-ol, trans-p-menta-2-8-
dien-1-ol, trans-piperitol.
Monoterpenos cetonas
carvona, cis-dihidrocarvona, e-isocitral, metil-
3-(2-furil)-2Z-acroleína, perilaldeído, trans-4-
caranona.Monoterpenos epóxidos 6-7-epoximirceno, trans-óxido de limoneno.
Monoterpenos ésteresoctanoato de isoamila, acetato de trans-
crisantenil.Monoterpenos hidrocarbonetos p-cimeno, limoneno, meta-cimeneno.
Monoterpenos óxidos óxido de nerol.
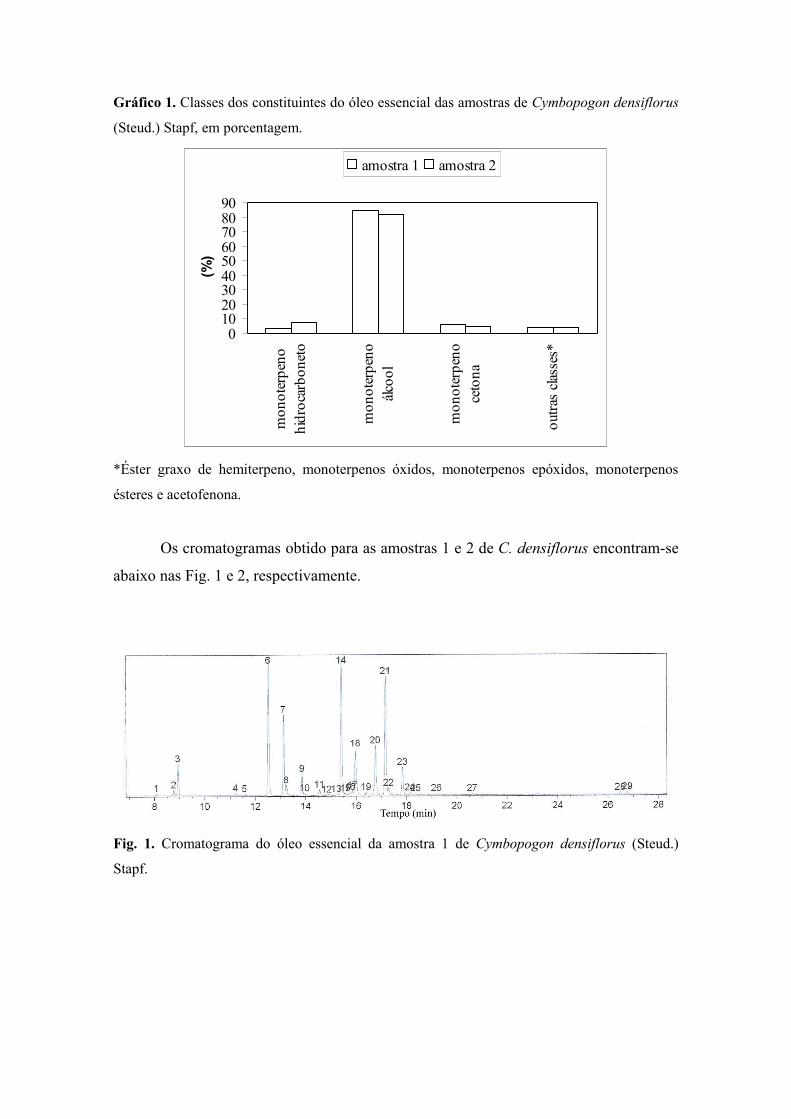
Gráfico 1. Classes dos constituintes do óleo essencial das amostras de Cymbopogon densiflorus
(Steud.) Stapf, em porcentagem.
0102030405060708090
mon
oter
peno
hidr
ocar
bone
to
mon
oter
peno
álcoo
l
mon
oter
peno
ceto
na
outra
s cla
sses
*
(%)
amostra 1 amostra 2
*Éster graxo de hemiterpeno, monoterpenos óxidos, monoterpenos epóxidos, monoterpenos
ésteres e acetofenona.
Os cromatogramas obtido para as amostras 1 e 2 de C. densiflorus encontram-se
abaixo nas Fig. 1 e 2, respectivamente.
Fig. 1. Cromatograma do óleo essencial da amostra 1 de Cymbopogon densiflorus (Steud.)
Stapf.

Fig. 2. Cromatograma do óleo essencial da amostra 2 de Cymbopogon densiflorus (Steud.)
Stapf.
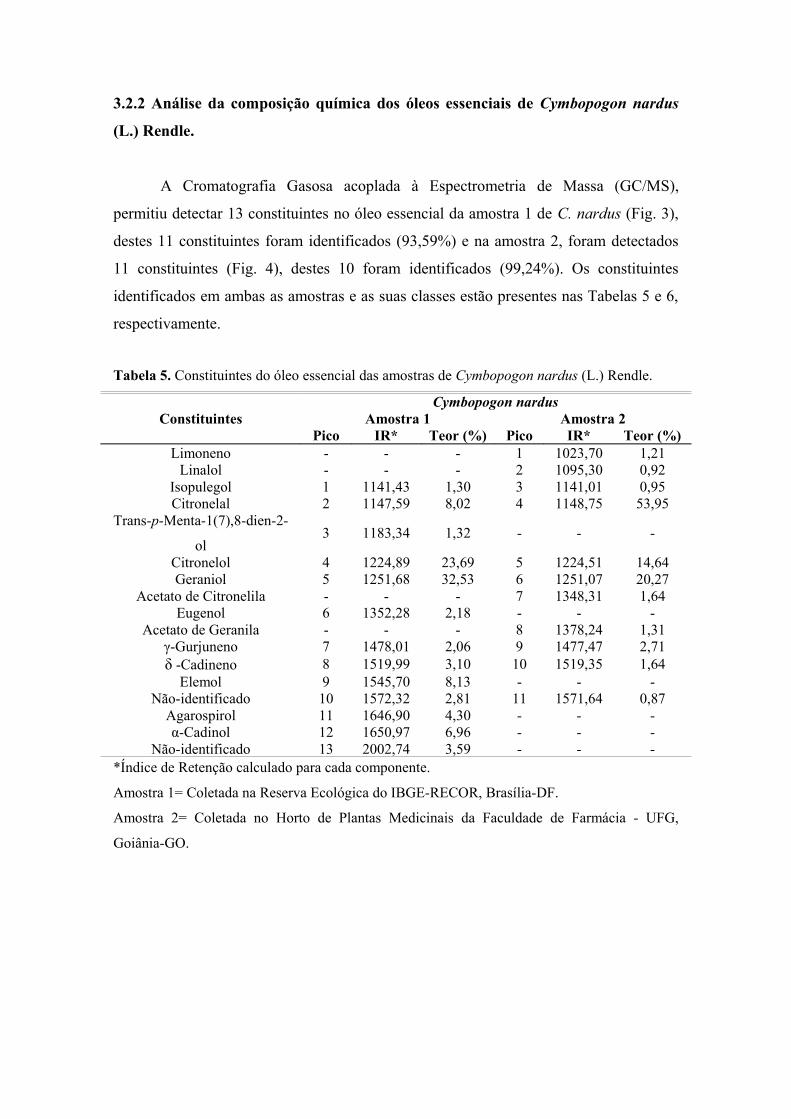
3.2.2 Análise da composição química dos óleos essenciais de Cymbopogon nardus
(L.) Rendle.
A Cromatografia Gasosa acoplada à Espectrometria de Massa (GC/MS),
permitiu detectar 13 constituintes no óleo essencial da amostra 1 de C. nardus (Fig. 3),
destes 11 constituintes foram identificados (93,59%) e na amostra 2, foram detectados
11 constituintes (Fig. 4), destes 10 foram identificados (99,24%). Os constituintes
identificados em ambas as amostras e as suas classes estão presentes nas Tabelas 5 e 6,
respectivamente.
Tabela 5. Constituintes do óleo essencial das amostras de Cymbopogon nardus (L.) Rendle.
ConstituintesCymbopogon nardus
Amostra 1 Amostra 2Pico IR* Teor (%) Pico IR* Teor (%)
Limoneno - - - 1 1023,70 1,21Linalol - - - 2 1095,30 0,92
Isopulegol 1 1141,43 1,30 3 1141,01 0,95Citronelal 2 1147,59 8,02 4 1148,75 53,95
Trans-p-Menta-1(7),8-dien-2-
ol3 1183,34 1,32 - - -
Citronelol 4 1224,89 23,69 5 1224,51 14,64Geraniol 5 1251,68 32,53 6 1251,07 20,27
Acetato de Citronelila - - - 7 1348,31 1,64Eugenol 6 1352,28 2,18 - - -
Acetato de Geranila - - - 8 1378,24 1,31γ-Gurjuneno 7 1478,01 2,06 9 1477,47 2,71δ -Cadineno 8 1519,99 3,10 10 1519,35 1,64
Elemol 9 1545,70 8,13 - - -Não-identificado 10 1572,32 2,81 11 1571,64 0,87
Agarospirol 11 1646,90 4,30 - - -α-Cadinol 12 1650,97 6,96 - - -
Não-identificado 13 2002,74 3,59 - - -*Índice de Retenção calculado para cada componente.
Amostra 1= Coletada na Reserva Ecológica do IBGE-RECOR, Brasília-DF.
Amostra 2= Coletada no Horto de Plantas Medicinais da Faculdade de Farmácia - UFG,
Goiânia-GO.
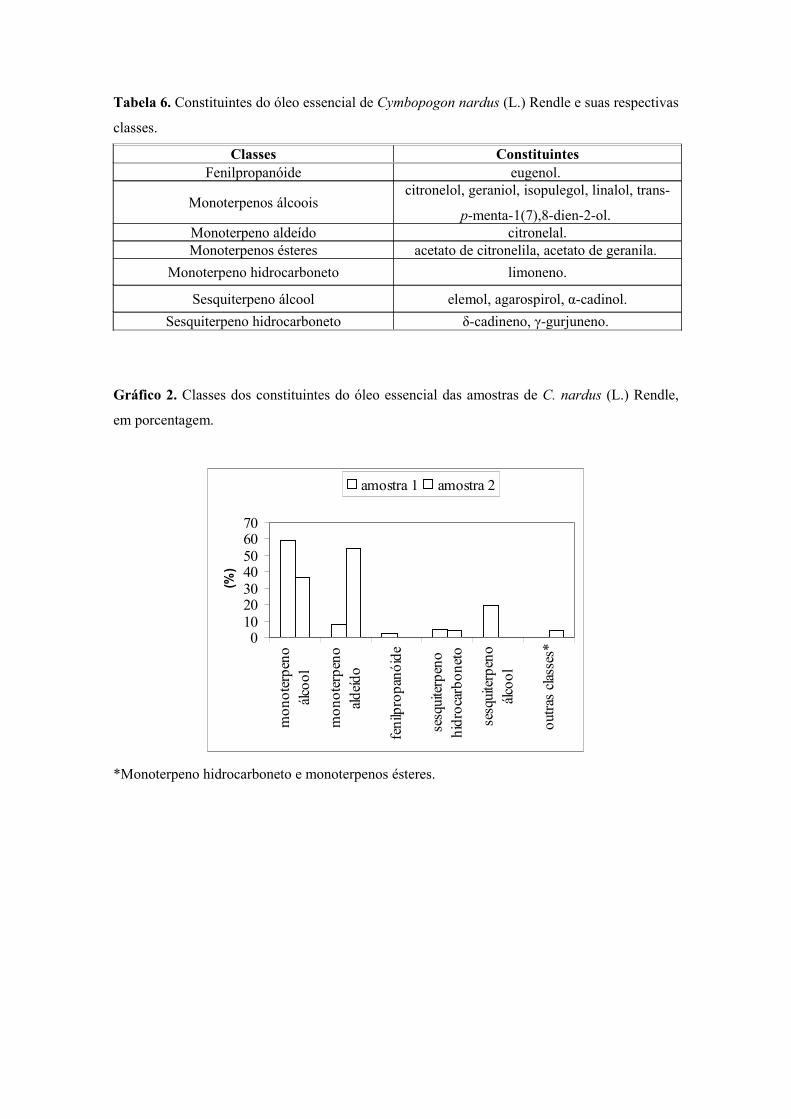
Tabela 6. Constituintes do óleo essencial de Cymbopogon nardus (L.) Rendle e suas respectivas
classes.
Classes ConstituintesFenilpropanóide eugenol.
Monoterpenos álcooiscitronelol, geraniol, isopulegol, linalol, trans-
p-menta-1(7),8-dien-2-ol. Monoterpeno aldeído citronelal.Monoterpenos ésteres acetato de citronelila, acetato de geranila.
Monoterpeno hidrocarboneto limoneno.
Sesquiterpeno álcool elemol, agarospirol, α-cadinol.Sesquiterpeno hidrocarboneto δ-cadineno, γ-gurjuneno.
Gráfico 2. Classes dos constituintes do óleo essencial das amostras de C. nardus (L.) Rendle,
em porcentagem.
010203040506070
mon
oter
peno
álcoo
l
mon
oter
peno
aldeíd
o
feni
lpro
panó
ide
sesq
uite
rpen
ohi
droc
arbo
neto
sesq
uite
rpen
oálc
ool
outra
s cla
sses
*
(%)
amostra 1 amostra 2
*Monoterpeno hidrocarboneto e monoterpenos ésteres.
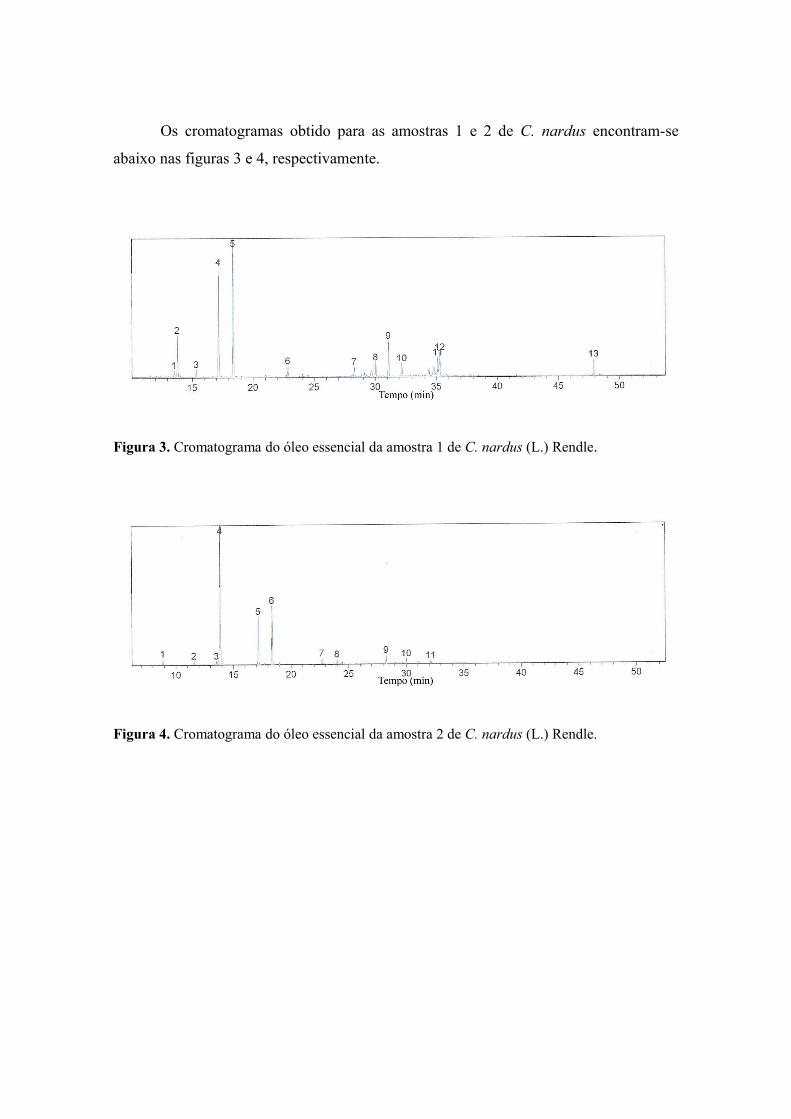
Os cromatogramas obtido para as amostras 1 e 2 de C. nardus encontram-se
abaixo nas figuras 3 e 4, respectivamente.
Figura 3. Cromatograma do óleo essencial da amostra 1 de C. nardus (L.) Rendle.
Figura 4. Cromatograma do óleo essencial da amostra 2 de C. nardus (L.) Rendle.

4 DISCUSSÃO
As folhas de C. densiflorus, coletadas em duas localidades diferentes do
município de Jataí-GO, apresentaram rendimento de óleo essencial de 0,4% e 0,5%,
para as amostras 1 e 2, respectivamente (Tabela 1). As folhas de C. nardus coletadas na
Reserva Ecológica do IBGE-RECOR, Brasília-DF (amostra 1) e na Horto de Plantas
Medicinais da Faculdade de Farmácia (amostra 2), tiveram rendimento de óleo essencial
de 0,5% cada uma (Tabela 2).
Possivelmente, os rendimentos obtidos entre as amostras estudadas não variaram
uma vez que as coletas foram realizadas na mesma estação do ano (período da seca:
junho a setembro de 2006). Além disso, deve-se levar em consideração, que os
indivíduos coletados localizavam-se em locais geográficos distintos, porém
relativamente próximos entre si. FIGUEIREDO et al. (2006) e KOSHIMA et al. (2006),
detectaram melhor rendimento do teor de óleo e conteúdo de citral nas folhas de C.
citratus (DC.) Stapf quando a colheita era realizada na época seca do ano (outono e
inverno).
Os rendimentos obtidos de óleo essencial de 0,5% e 0,4% nas amostras
analisadas de C. densiflorus corroboram os resultados obtidos por TAKAISI-KIKUNI et
al. (2000), em estudos realizados com essa espécie.
Verifica-se que os rendimentos obtidos, nas amostras analisadas das duas
espécies, coincidem com os apresentados na monografia de C. citratus, onde se
estabelece que as folhas dessecadas dessa espécie devem conter, no mínimo, 0,5% de
óleo essencial (FARMACOPÉIA BRASILEIRA IV, 2003), assim como ao rendimento
de 0,5% de óleo essencial de C. giganteus (Hochst.) Chiov (KANKO et al., 2004).
Entretanto, os rendimentos verificados para as duas amostras analisadas de C.
densiflorus e C. nardus estão relativamente baixos, quando comparados aos obtidos de
outras espécies pertencentes a este gênero. GALVÃO (2004) obteve um rendimento de
1,70% de óleo essencial de C. winteranus Jowitt. KANKO et al. (2004) registraram
rendimento de óleo essencial de 1,2% e 0,7%, para C. nardus e C. citratus,
respectivamente. RAO et al. (2004) verificaram rendimento de 0,75% na extração do
óleo essencial de C. martinii (Roxb.) Wats.
O rendimento de óleo essencial é avaliado a partir da matéria seca, podendo ser
muito variável, dependendo de diversos fatores internos e externos, como por exemplo:
época e horário de colheita (SILVA, 2005). Dentre outros fatores que influenciam na

variação do rendimento do óleo essencial estão os fatores abióticos: tipo de clima e solo
e bióticos: interação planta-planta, planta-microorganismos e planta-herbívoros
(ROBBERS et al., 1997). Tais fatores resultam em pressões ambientais que ocasionam
mudanças quanto à constituição genética e também, nas atividades fisiológicas, que
levam as plantas a se adaptarem aos diversos ambientes (KULKARNI e RAMESH,
1987; SANGWAN et al., 2001).
De acordo com KOSHIMA et al., 2006, apesar das espécies do gênero
Cymbopogon serem plantas rústicas que se adaptam e toleram todos os tipos de solos,
há uma preocupação em otimizar a produção do óleo essencial, devido ao uso industrial
e popular. A otimização do rendimento dos óleos essenciais nestas plantas, varia
conforme as condições do solo (fertilidade e drenagem) e das condições climáticas.
Nas folhas de C. densiflorus foram detectados 29 constituintes no óleo essencial
na amostra 1, destes, 24 constituintes foram identificados, correspondendo a 98,56%
(Fig. 1 e Tabela 3). Os constituintes majoritários presentes foram trans-p-menta-1(7),8-
dien-2-ol (21,33%), trans-p-menta-2-8-dien-1-ol (17,77%), cis-carveol (17,14%) e cis-
p-menta-2-8-dien-1-ol (10,18%). No óleo essencial da amostra 2 de C. densiflorus,
foram detectados 28 constituintes, destes, 23 foram identificados, o que corresponde a
98,78% (Fig. 2 e Tabela 3). Os constituintes majoritários foram trans-p-menta-1(7),8-
dien-2-ol (22,14%), trans-p-menta-2-8-dien-1-ol (18,31%), cis-carveol (17,75%) e cis-
p-menta-2-8-dien-1-ol (10,04%).
C. densiflorus apresentou óleo essencial constituído majoritária por
monoterpenos (Gráfico 1), percebendo que os monoterpenos álcoois estão em maiores
porcentagens: (84,48%) na amostra 1 e (82,05%) na amostra 2. Todos os constituintes
majoritários encontrados em ambas as amostras de C. densiflorus, são monoterpenos
álcoois (Tabela 4). Os ésteres graxos de hemiterpenos, monoterpenos óxidos,
monoterpenos epóxidos, monoterpenos ésteres e acetofenonas estão presentes em baixas
porcentagens e estão, portanto, reunidos (em outras classes) no Gráfico 1.
Nas folhas de C. nardus, foram detectados 13 constituintes no óleo essencial da
amostra 1, destes, 11 foram identificados, correspondendo a 93,59% (Fig. 3 e Tabela 5).
Os constituintes majoritários presentes foram geraniol (32,53%), citronelol (23,69%),
elemol (8,13%) e citronelal (8,02%). No óleo essencial da amostra 2 de C. nardus foram
detectados 11 constituintes, destes, 10 foram identificados o que corresponde a 99,24%
(Fig. 4 e Tabela 5). Os constituintes majoritários presentes foram geraniol (20,27%),

citronelol (14,64%) e citronelal (53,95%), entretanto, o elemol não foi encontrado nessa
amostra.
C. nardus contém óleo essencial composto por monoterpenos e sesquiterpenos
(Gráfico 2). Os monoterpenos álcoois estão em 58,84% (amostra 1) e 36,78% (amostra
2), os monoterpenos aldeídos em 8,02% (amostra 1) e 53,95% (amostra 2) e os
sesquiterpenos álcoois em 19,39% apenas na amostra 2. Dentre os componentes
majoritários verificados em ambas as amostras de C. nardus, citronelol e geraniol são
monoterpenos álcoois, citronelal é monoterpeno aldeído e elemol, sesquiterpeno álcool
(Tabela 6). Os monoterpenos hidrocarbonetos e monoterpenos ésteres apresentaram-se
em baixas porcentagens e estão reunidos (em outras classes) no Gráfico 2.
De acordo com GOMES e NEGRELLE (2003) e SIMÕES e SPITZER (2003),
os monoterpenos, às vezes constituem acima de 90% do óleo essencial, sendo que estas
porcentagens e ausência de alguns constituintes podem variar conforme o solo de
cultivo da planta, época de colheita, método de extração, dentre outras variáveis.
Observou-se que as duas amostras analisadas de C. densiflorus apresentaram os
mesmos constituintes químicos entre si (Tabela 3), enquanto que as amostras de C.
nardus demonstraram uma maior variação nos constituintes entre si (Tabela 5). Esta
variabilidade pode ser devida à influências bióticas e abióticas, como a diversidade
genética, o habitat, e os tratos culturais, que interferem na composição química dos
óleos essenciais (GOMES e NEGRELLE, 2003).
Os óleos essenciais presentes nas amostras estudadas de C. densiflorus
apresentaram os mesmos constituintes majoritários verificados por TAKAISI-KIKUNI
et al. (2000), em C. densiflorus; KANKO et al. (2004), em C. nervatus (Hochst.) Chiov.
e EL-KAMALI et al. (2005), em C. giganteus (Hochst.) Chiov. TAKAISI-KIKUNI et
al. (2000) verificaram que os constituintes principais do óleo essencial de C. densiflorus
são limoneno, cimeneno, p-cimeno, cis- e trans-carveol, cariona, iso-piperitenona, p-
menta-2, 8-dien-1-ol e p-menta-1, 7-dien-ol, em contraste aos óleos de outras espécies
do gênero Cymbopogon que, segundo a literatura, apresentam mirceno e citral como
constituintes principais.
Os resultados obtidos, para os constituintes majoritários do óleo essencial das
amostras de C. nardus, foram semelhantes aos encontrados por GALVÃO (2004), no
óleo essencial de C. winteranus Jowitt que apresentou citronelal (74,88%), citronelol
(8,23%), geraniol (6,48%). Essa espécie é morfologicamente semelhante à C. nardus e

as folhas apresentam aroma característico de eucalipto. De acordo com SOENARKO
(1977), C. winteranus é também conhecida por capim-citronela ou Citronela de Java.
Os constituintes acetato de geranila, citronelal, citronelol, δ-cadineno, elemol,
geraniol, isopulegol e limoneno presentes no óleo essencial das amostras analisadas de
C. nardus, também foram registrados em C. afronardus Stapf (BASER et al., 2005), C.
citratus (DC.) Stapf (MARTINS et al., 2004 ; QUIALA et al., 2006), C. martinii
(Roxb.) Wats. (DUARTE et al., 2007), C. nardus (L.) Rendle (KANKO et al., 2004 ;
SAMARASEKERA et al., 2006).
De acordo com os constituintes verificados no óleo essencial de C. densiflorus e
C. nardus, pôde-se perceber que algumas espécies se relacionam mais quimicamente
entre si, dentro do gênero. Essas análises da composição dos óleos essenciais podem
ser importantes como subsídios para a quimiotaxonomia, uma vez que este gênero
apresenta espécies diferentes que se assemelham morfologicamente.
Além da importância medicinal dos óleos essenciais de C. densiflorus e C.
nardus, essas espécies apresentam constituintes nos óleos que as fazem ser
industrialmente importante. Geraniol, nerol e muitos de seus derivados, devido ao
agradável odor floral, são utilizados na perfumaria e produtos cosméticos, por exemplo,
perfumes, loções, sabões, desodorantes e como aromatizantes em alimentos, tais como
bebidas não-alcoólicas, sorvetes e doces (JIROVETZ et al., 2007). SILVA SANTOS et
al. (2006), em trabalho de revisão, realizaram levantamento dos principais constituintes
de óleos essenciais, utilizados industrialmente e citam que o citral, citronelol, citronelal,
geraniol e linalol são utilizados para fragrâncias.

5 CONCLUSÃO
Apesar das espécies estudadas pertencerem ao mesmo gênero, Cymbopogon
densiflorus (Steud.) Stapf apresentou mais constituintes nos óleos essenciais do que C.
nardus (L.) Rendle. Além disso, essas duas espécies se demonstraram quimicamente
bem distintas, compartilhando poucos constituintes entre si.
Quanto aos constituintes verificados, C. densiflorus se assemelha com C.
nervatus e C. giganteus, enquanto que C. nardus está mais próximo à C. afronardus, C.
citratus, C. martinii e, principalmente, à C. winteranus. Tais análises podem fornecer
subsídios para estudos taxonômicos dentro do gênero Cymbopogon.
As espécies demonstram-se relevantes para os fins medicinais aos quais são
popularmente utilizados e devido a este potencial fitoterápico, merecem estudos mais
aprofundados, ou seja, experimentos de isolamento, purificação e determinação
estrutural dos constituintes detectados. Além de testes farmacológicos e toxicológicos
que forneçam embasamento científico para o uso seguro dos óleos essenciais destas
espécies.

6 REFERÊNCIAS BIBLIOGRÁFICAS(*)
ADAMS, R.P. Identification of essential oil components by gas
chromatography/quadrupole mass spectroscopy. 4ª ed. Carol Stream, IL: Allured
Publishing Corporation. 2007.
ALVES, D.L.; SILVA, C.R. Fitohormônios: Abordagem natural da terapia
hormonal. São Paulo: Atheneu. 2002.
BASER, K.H.C.; OZEK, T.; DEMIRCI, B. Composition of the essential oil of
Cymbopogon afronardus Stapf from Uganda. Journal of Essential Oil Research. 17:
139-140. 2005.
CIMANGA, K.; KAMBU, K.; TONA, L.; APERS, S.; DE BRUYNE, T.; HERMANS,
N.; TOTTÉ, J.; PIETERS, L.; VLIETINCK, A.J. Correlation between chemical
composition and antibacterial activity of essential oils of some aromatic medicinal
plants growning in the Democratic Republic of Congo. Journal of
Ethnopharmacology. 79: 213-220. 2002.
DUARTE, M.C.T.; LEME, E.E.; DELARMELINA, C.; SOARES, A.A.; FIGUEIRA,
G.M.; SARTORATTO, A. Activity of essential oils from Brazilian medicinal plants on
Escherichia coli. Journal of Ethnopharmacology. 111: 197-201. 2007.

DUARTE, M.R.; ZANETI, C.C. Estudo farmacobotânico das folhas de capim-limão:
Cymbopogon citratus (DC.) Stapf., Poaceae. Visão Acadêmica. 5(2): 117-124. 2004.
EHLERT, P.A.D.; BLANK, A.F.; ARRIGONI-BLANK, M.F.; PAULA, J.W.A.;
CAMPOS, D.A.; ALVIANO, C.S. Tempo de hidrosdestilação de óleo essencial de sete
espécies medicinais. Revista Brasileira de Plantas Medicinais. 8 (2): 79-80. 2006.
EL-KAMALI, H.J.; HAMZA, M.A.; EL-AMIR, M.Y. Antibacterial activity of the
essential oil from Cymbopogon nervatus inflorescence. Fitoterapia. 76: 446-449. 2005.
FAHN, A. Secretory tissues in plants. Academic Press: New York/London. 1979.
______________________________________________________________________
(*) Referências bibliográficas conforme ABNT/NBR
FARMACOPÉIA BRASILEIRA. 4ª ed. Parte II – Fascículo 4. São Paulo: Atheneu.
2003.
FIGUEIREDO, R.O.; DELACHIAVE, M.E.A.; MING, L.C. Reguladores vegetais na
produção de biomassa e teor de óleos essenciais em Cymbopogon citratus (DC.) Stapf.,
em diferentes épocas do ano. Revista Brasileira de Plantas Medicinais. 8 (3): 31-35.
2006.
GALVÃO, E.L. Extração do óleo essencial de Cymbopogon winteranus (Jowitt) com
CO2 pressurizado. Dissertação (Mestrado em Engenharia Química) – Universidade
Federal do Rio Grande do Norte, Natal-RN. 2004.
GOMES, E.C.; NEGRELLE, R.R.B. Cymbopogon citratus (DC.) Stapf.: Aspectos
Botânicos e Econômicos. Visão Acadêmica. 4(2): 137-144. 2003.
JIROVETZ, L.; BUCHBAUER, G. SCHMIDT, E.; STOYANOVA, A.S.; DENKOVA,
Z.; NIKOLOVA, R.; GEISSLER, M. Purity, Antimicrobial activities and olfactoric
evaluations of Geraniol/Nerol and various of their derivatives. Journal of Essential Oil
Research. 19: 288-291. 2007.
KANKO, C.; SAWALIHO, B.E.H.; KONE, S.; KOUKOUA, G.; N’GUESSAN, Y.T.
Étude des propriétés physico-chimiques des huiles essentielles de Lippia multiflora,

Cymbopogon citratus, Cymbopogon nardus, Cymbopogon giganteus. Chimie. 7: 1039-
1042. 2004.
KOSHIMA, F.A.T.; MING, L.C.; MARQUES, M.O.M. Produção de biomassa,
rendimento de óleo essencial e de citral em capim-limão, Cymbopogon citratus (DC.)
Stapf., com cobertura morta nas estações do ano. Revista Brasileira de Plantas
Medicinais. 8 (4): 112-116. 2006.
KULKARNI, R.N.; RAMESH, S. Hereditability estimates of citral content in East
Indian lemongrass. Theoretical and Applied Genetics. 73: 476-477. 1987.
MARTINAZZO, A.P.; CORRÊA, P.C.; MELO, E.C.; BARBOSA, F.F. Difusividade
efetiva em folhas de Cymbopogon citratus (DC.) Stapf submetidas à secagem com
diferentes comprimentos de corte e temperaturas do ar. Revista Brasileira de Plantas
Medicinais. 9(1): 68-72. 2007.
MARTINS, M.B.G.; MARTINS, A.R.; TELASCRÊA, M.; CAVALHEIRO, A.J.
Caracterização Anatômica da folha de Cymbopogon citratus (DC.) Stapf. (Poaceae) e
perfil químico do óleo essencial. Revista Brasileira de Plantas Medicinais. 6(3): 20-
29. 2004.
MILLS, S.; BONE, K. Principles and Practice of Phytotherapy. 1ª reimpressão.
Churchil Livingstone. 2001.
PAVIANI, L.; PERGHER, S.B.C.; DARIVA, C. Aplication of molecular sieves in the
fractionation of lemongrass oil from high-pressure carbon dioxide extraction. Brazilian
Journal of Chemical Engineering. 23(2): 219-225. 2006.

PRASHAR, A.; HILI, P.; VENESS, R.G.; EVANS, C.S. Antimicrobial action of
palmarosa oil (Cymbopogon martini) on Saccharomyces cerevisiae. Phytochemistry.
63? 569-575. 2003.
QUIALA, E.; BARBÓN, R.; JIMÉNEZ, E.; DE FERIA, M.; CHÁVEZ, M.; CAPOTE,
A.; PÉREZ, N. Biomass production of Cymbopogon citratus (DC.) Stapf., a medicinal
plant in temporary immersion systems. In Vitro Cellular and Developmental Biology
– Plant. 42: 298-300. 2006.
RAO, B.R. RAJESWARA; KAUL, P.N.; SYAMASUNDAR, K.V.; RAMESH, S.
Chemical profiles of primary and secondary essential oils of palmarosa (Cymbopogon
martinii (Roxb.) Wats var. motia Burk.). Industrial Crops and Products. 21: 121-127.
2004.
RAUBER, C.S.; GUTERRES, S.S.; SCHAPOVAL, E.E.S. LC determination of citral in
Cymbopogon citratus volatile oil. Journal of Pharmaceutical and Biomedical
Analysis. 37: 597-601. 2005.
ROBBERS, J.E.; SPEEDIE, M.K.; TYLER, V.E. Farmacognosia e
Farmacobiotecnologia. São Paulo: Editorial Premier. 1997.
SAMARASEKERA, R.; KALHARI, K.; WEERASINGHE, I.S. Insecticidal activity of
essential oils of Ceylon Cinnamomum and Cymbopogon species against Musca
domestica. Journal of Essential Oil Research. 18: 352-354. 2006.
SANGWAN, N.S.; YADAV, U.; SANGWAN, R.S. Molecular analysis of genetic
diversity in elite Indian cultivars of essential oil trade types of aromatic grasses
(Cymbopogon species). Plant Cell Reports. 20: 437-444. 2001.
SILVA, F. Avaliação do teor e da composição química do óleo essencial de plantas
medicinais submetidas a processos de secagem e armazenamento. Tese (Doutorado
em Engenharia Agrícola) – Universidade Estadual de Campinas, Campinas-SP. 2005.

SILVA SANTOS, A; BIZZO, H.R.; ANTUNES, A.M.S.; D’AVILA, I.A. A proteção
patentária na utilização de óleos essenciais e compostos terpênicos para o
desenvolvimento tecnológico e industrial. Revista Brasileira de Plantas Medicinais.
8(4): 14-22. 2006.
SIMÕES, C.M.O.; SPITZER, V. Óleos Essenciais. In: SIMÕES, C.M.O.; SCHENKEL,
E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.; PETROVICK, P.R. (Org.).
Farmacognosia: da planta ao medicamento. 5ª ed. Porto Alegre/Florianópolis:
UFRGS, p. 467-495. 2003.
SOENARKO, S. The Genus Cymbopogon Sprengel (Gramineae). Reinwardtia. Vol. 9
(3): 225-375. 1977.
SOUZA, E.M.B.D.; CÂMARA, A.P.C.; COSTA, W.A.; COSTA, A.C.; OLIVEIRA,
H.N. M.; GALVÃO, E.L.; MARQUES, M.M.O. Evaluation of process of the essential
oil from Cymbopogon schoenanthus with pressurized carbon dioxide. Brazilian
Archives of Biology and Technology. 48: 231-236. 2005.
TAKAISI-KIKUNI, N.B. TSHILANDA, D.; BABADY, B. Antibacterial activity of the
essential oil of Cymbopogon densiflorus. Fitoterapia. 71: 69-71. 2000.
VAN DEN DOOL, H; KRATZ, P.D.J.A. Generalization of the retention index system
including linear temperature programmed gas – liquid partition chromatography.
Journal of Chromatography. 11: 463-471. 1963.








