
UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
MARCELO VENTURA RUBIO
EFEITO DE N-GLICANAS SOBRE PROPRIEDADES
FUNCIONAIS DE GLICOSIL HIDROLASES
N-GLYCANS EFFECT ON FUNCTIONAL PROPERTIES OF
GLYCOSYL HYDROLASES
CAMPINAS
2018

MARCELO VENTURA RUBIO
EFEITO DE N-GLICANAS SOBRE PROPRIEDADES FUNCIONAIS DE GLICOSIL HIDROLASES
N-GLYCANS EFFECT ON FUNCTIONAL PROPERTIES OF GLYCOSYL HYDROLASES
Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do Título de Doutor em Ciências, na área de concentração em Fármacos, Medicamentos e Insumos para Saúde
Thesis presented to the Institute of Biology of the University of Campinas in partial fulfillment of the requirements for the degree of PhD in Sciences, in the field of concentration Pharmaceuticals, Medicines and Health Supplies
Orientador: ANDRÉ RICARDO DE LIMA DAMÁSIO Co-Orientador: FABIO MARCIO SQUINA
CAMPINAS
2018
ESTE ARQUIVO DIGITAL CORRESPONDE À
VERSÃO FINAL DA TESE DEFENDIDA PELO
ALUNO MARCELO VENTURA RUBIO E
ORIENTADA PELO ANDRÉ RICARDO DE LIMA
DAMÁSIO


Campinas, 12 de julho de 2018.
COMISSÃO EXAMINADORA
Profa. Dra. Maria de Lourdes Teixeira de Moraes Polizeli
Prof. Dr. Marcos Silveira Buckeridge
Dr. Fausto Bruno dos Reis Almeida
Dra. Thamy Lívia Ribeiro Corrêa
Prof. Dr. André Ricardo de Lima Damásio (Orientador)
Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no processo de vida acadêmica do aluno.

Dedicatória
Aos meus pais e irmã
A toda minha família
A todos os meus amigos
“Education is the most powerful weapon which you can use to change the world”
“Always seems impossible until it’s done”
(Nelson Mandela)

Agradecimento
Ao longo dessa jornada, chamada doutorado, muito tenho a agradecer a todos que
fizeram parte dessa etapa da minha vida. Diretamente ou indiretamente todos vocês
são responsáveis e o motivo de chegar aonde cheguei e ser o que eu sou.
Agradeço a minha família por estar sempre ao meu lado, tanto nos momentos fáceis
quanto nos difíceis.
Agradeço a minha namorada, Aline, pela paciência, compreensão e companheirismo
ao longo desses anos.
Agradeço aos meus amigos que tornaram essa jornada mais fácil e muito mais
divertida. Pela ajuda no dia a dia, dentro e fora de um laboratório.
Agradeço a Universidade Estadual de Campinas (Unicamp) e a Fundação de Amparo
a Pesquisa do Estado de São Paulo (Fapesp; processo n° 2013/24988-5) pelo suporte
concedido ao longo de minha vida acadêmica.
Agradeço ao meu orientador, Prof. Dr. André Damásio, por todos esses anos de
ensinamentos passados e pela amizade criada nesses anos.
Muito obrigado a todos!
“We must find time to stop and thank the people who make a difference in our lives”
(John F. Kennedy)

Resumo
O mercado de enzimas industriais abrange uma ampla variedade de aplicações, bem
como: cuidados pessoais, indústria alimentícia, biocombustíveis, biopolímeros, entre
outros. O principal desafio para o uso de coquetéis enzimáticos em larga escala é seu
custo elevado, aumentando o valor final dos bioprodutos. Portanto, o custo dos
bioprodutos pode ser reduzido significativamente, através do aumento do rendimento
da produção de enzimas em cepas fúngicas por técnicas de biologia molecular e/ou
melhorando a eficiência enzimática por engenharia de proteínas. Os fungos
filamentosos são os principais produtores de enzimas industriais devido ao vasto
repertório enzimático em seu genoma e elevada secreção. O gênero Aspergillus inclui
microrganismos que naturalmente degradam a biomassa lignocelulósica secretando
grandes quantidades de enzimas ativas em carboidratos (CAZymes). A capacidade
de Aspergillus de realizar modificações pós-traducionais, tais como: clivagem
proteolítica, ligações dissulfeto e glicosilação, proporciona uma vantagem adicional à
sua utilização para a produção de enzimas heterólogas. Contudo, a super-expressão
de proteínas sobrecarrega a via de N-glicosilação e os mecanismos de enovelamento,
resultando no acúmulo de proteínas mal ou não enoveladas. Proteínas mal
enoveladas são direcionadas à degradação, consequentemente reduzindo o
rendimento de sua secreção. Além disso, a posição e o número de N-glicanas ligados
às proteínas podem influenciar sua secreção e propriedades funcionais. Com o
objetivo de minimizar o custo de produção de enzimas, Aspergillus nidulans foi
utilizado como organismo modelo para estudar o efeito da N-glicosilação na secreção
de enzimas industriais. Utilizando abordagem proteômica identificou-se 265 N-
glicoproteínas secretadas por A. nidulans quando cultivado em xilano e bagaço de
cana-de-açúcar pré-tratado. As CAZymes corresponderam a mais de 50% do
secretoma e 182 sítios de N-glicosilação foram validados por LC-MS/MS. A fim de
investigar a influência das N-glicosilações na secreção de proteínas em A. nidulans, a
β-xilosidase (BxlB) da família glicosil hidrolase 3 foi selecionada como alvo devido ao
seu elevado nível de secreção durante o crescimento em xilano. As β-xilosidases são
hidrolases glicosídicas que auxiliam na degradação da biomassa vegetal, liberando
xilose a partir de xilooligossacarídeos e/ou xilobiose. Sete sítios de N-glicosilação
foram preditos em BxlB e cinco deles foram validados por LC-MS/MS. Glicomutantes
foram desenhados para investigar a influência da glicosilação na secreção e função

de BxlB. O mutante deglicosilado (BxlBDeglyc) apresentou secreção e atividade
enzimática semelhantes com a proteína selvagem (BxlBwt). Interessantemente, o
mutante parcialmente glicosilado (BxlBN1;5;7) apresentou níveis aumentados de
atividade e secreção. Por outro lado, o mutante BxlBCC, no qual o contexto de N-
glicosilação foi alterado, foi expresso, mas não secretado em A. nidulans. BxlBwt,
BxlBDeglyc e BxlBN1;5;7 mostraram estrutura secundária semelhante, embora os
mutantes tivessem menor temperatura de fusão em comparação com o tipo selvagem.
Além disso, um novo glicomutante mantendo apenas dois sítios de N-glicosilação
(BxlBN5;7) mostrou uma melhor eficiência catalítica. Este estudo mostra a influência da
N-glicosilação na função e produção de BxlB em A. nidulans, reforçando que a
glicoengenharia de proteínas é uma ferramenta promissora para aumentar a
estabilidade térmica, secreção e atividade enzimática. Esse trabalho, também, poderá
servir de base para modificações de N-glicosilação em CAZymes para aplicações
biotecnológicas.

Abstract
The industrial enzymes market covers a wide variety of applications such as personal
care, food industries, biofuels, biopolymers, among others. The main bottleneck for
using enzymatic cocktails at large scale is the high-cost, which increases the
bioproducts final value. Thus the bioproducts cost can be significantly reduced by
improving the yield of enzymes production by molecular biology of fungal strains and/or
by improving enzymes efficiency by protein engineering. Filamentous fungi are the
main producers of industrial enzymes due to the great enzymatic repertoire and the
high levels of protein secretion. The genus Aspergillus includes microorganisms that
naturally degrade lignocellulosic biomass by secreting large amounts of carbohydrate-
active enzymes (CAZymes). The capacity of Aspergillus to perform post-translational
modifications such as proteolytic cleavage, disulfide bond formation and glycosylation,
provides an additional advantage to their use as hosts for heterologous protein
production. However, the overexpression of target proteins overloads the N-
glycosylation pathway and folding mechanisms resulting in the accumulation of
unfolded or misfolded proteins. Misfolded proteins are directed to degradation,
consequently reducing the secretion yield. Furthermore, the position and the number
of N-glycans attached to proteins can influence their secretion and functional
properties. Aiming to minimize the cost of enzymes production, Aspergillus nidulans
was used as a model organism to study the effect of N-glycosylation in the secretion
of industrial enzymes. A proteomics approach identified 265 N-glycoproteins secreted
by A. nidulans grown on xylan and pretreated sugarcane bagasse. CAZymes
corresponded to more than 50% of the secretome and 182 N-glycosylated sites were
validated by LC-MS/MS. In order to investigate the influence of N-glycosylation in
protein secretion by A. nidulans, a β-xylosidase (BxlB) was selected as target protein
due to its high secretion level during growth on xylan. β-xylosidases are glycoside
hydrolases that assist plant biomass degradation by releasing xylose from
xylooligosaccharides and/or xylobiose. Seven N-glycosylation sites were predicted in
the BxlB and five were validated by LC-MS/MS. Glycomutants were designed in order
to investigate the influence of glycosylation on β-xylosidase secretion and function.
The deglycosylated mutant (BxlBDeglyc) showed similar results regarding enzyme
secretion and activity compared to the wild-type (BxlBwt). Interestingly, a partially
glycosylated mutant (BxlBN1;5;7) showed increased activity and secretion levels. On the

other hand, the mutant BxlBCC, in which the glycosylation context was changed by the
design of four new N-glycosylation sites, was expressed but not secreted in A.
nidulans. BxlBwt, BxlBDeglyc and BxlBN1;5;7 showed similar secondary structure, although
the mutants had lower melting temperature compared to the wild-type. Moreover, an
additional BxlB glycomutant maintaining only two N-glycosylated sites (BxlBN5;7)
showed improved catalytic efficiency. This study showed the influence of N-
glycosylation on BxlB function and production in A. nidulans, reinforcing that proteins
glycoengineering is a promising tool to enhance thermal stability, secretion and
enzymatic activity. Our report may also support N-glycosylation modification in
CAZymes to biotechnological applications.

Sumário
Capítulo 1. Revisão bibliográfica ...................................................................................... 12
Produção de enzimas de interesse por Aspergillus spp. .................................................. 16
Secreção de proteínas recombinantes em fungos filamentosos ....................................... 19
Obstáculos para a produção de proteínas recombinantes em fungos filamentosos.......... 20
Estratégias para superar os principais “gargalos” ............................................................. 26
Glicosilação de proteínas ................................................................................................. 31
N-glicosilação e biotecnologia .......................................................................................... 38
Estratégias empregadas para estudos de N-glicosilação ................................................. 38
Capítulo 2. Glicoproteômica e Glicômica de Aspergillus nidulans ................................ 43
Introdução ........................................................................................................................ 43
Mapping N-Linked Glycosylation of Carbohydrate-Active Enzymes in the Secretome of
Aspergillus nidulans Grown on Lignocellulose .................................................................. 44
Abstract ............................................................................................................................ 45
Background ...................................................................................................................... 46
Results ............................................................................................................................. 48
Discussion ........................................................................................................................ 62
Conclusion ....................................................................................................................... 71
Material and methods ....................................................................................................... 72
Additional files .................................................................................................................. 77
Capítulo 3 – Influência da N-glicosilação na produção e função de uma β-xilosidase de
A. nidulans ....................................................................................................................... 110
Introdução ...................................................................................................................... 110
Redesigning N-glycosylation sites in a GH3 β-xylosidase improves enzyme efficiency in
Aspergillus nidulans ....................................................................................................... 111
Abstract .......................................................................................................................... 112
Introduction .................................................................................................................... 112
Results ........................................................................................................................... 114
Discussion ...................................................................................................................... 122
Experimental procedure ................................................................................................. 126
Capítulo 4. Considerações finais .................................................................................... 142
Referências ...................................................................................................................... 144
Anexos.............................................................................................................................. 175
Termo de aprovação da pesquisa pela Comissão de Biossegurança ............................. 175
Declaração referente aos direitos autorais ..................................................................... 176

12
Capítulo 1. Revisão bibliográfica
Desde os tempos antigos, os microrganismos têm desempenhado um papel central
em diversos processos na fabricação de alimentos, tais como na produção de queijo,
cerveja e vinho, e na fabricação de produtos como couro e linho. Além disso, diversas
características do metabolismo dos fungos vêm sendo exploradas para produção e
obtenção de compostos bioativos e antibióticos (Magaña-Ortíz et al., 2013). Com o
passar dos anos descobriu-se que o papel dos microrganismos nesses processos
produtivos estava diretamente relacionado à produção de enzimas (Kirk, Borchert, &
Fuglsang, 2002).
Atualmente, as enzimas microbianas têm sido aplicadas em diversos setores da
indústria, os quais são classificados como: cuidados pessoais; alimentício;
bioenergético; agricultura e ração; e técnico e farmacêutico (Fleiβner & Dersch, 2010;
Maloy & Schaechter, 2006; Owen P. Ward, 2012). Nesses casos, as enzimas podem
ser parte componente ou então serem aplicadas na forma de coquetéis para a
obtenção de produtos de limpeza, detergentes, produção de xaropes, etanol de
primeira e segunda geração; ração animal; produção de tecidos, papel,
medicamentos, entre outros.
A produção de enzimas em escala industrial tem sido reportada desde 1874, quando
Christian Hansen fabricou queijo, usando renina (quimosina) obtido a partir de extratos
de estômagos de bezerros (Sani & Krishnaraj, 2017). Esta enzima é produzida
atualmente usando a técnica do DNA recombinante com o gene expresso em
Escherichia coli K-12, sendo a primeira enzima aprovada pela US Food and Drug
Administration (FDA) para uso em alimentos (Flamm, 1991). Na década de 1930, as
pectinases passaram a ser utilizadas para clarificação de suco; e no início dos anos
1940s a invertase começou a ser empregada na hidrólise de sacarose para produção
de xarope de açúcar invertido, aplicação pioneira de enzimas imobilizadas. A
aplicação de enzimas em larga escala começou na década de 1960, quando a
glicosidase foi utilizada para hidrólise de amido na produção xaropes de glicose
(Fernandes, 2010). Este processo substituiu a hidrólise ácida devido às suas muitas
vantagens, isto é, maiores rendimentos do produto, maior grau de pureza,
cristalização, menor geração de resíduos, entre outras.

13
A procura por produtos inovadores como tendência para um mercado sustentável
inspirou o desenvolvimento tecnológico, estimulando, por sua vez, a criação de novas
aplicações para enzimas em diferentes setores industriais nos últimos anos. Os
problemas ambientais contemporâneos aumentaram a importância e há, atualmente,
um crescente interesse pelo uso eficiente de vários resíduos agroindustriais. Esta
preocupação resultou em uma fonte importante para a produção de novos materiais,
produtos químicos e energia (Rosa et al., 2011). Como resultado, uma extensa gama
de produtos com maior valor agregado pode ser obtida a partir do que antes era
considerado “lixo”. Diversas pesquisas têm mostrado o enorme potencial econômico
da reutilização de resíduos (Figura 1) (Singh Nee Nigam & Pandey, 2009; White,
2015).
Figura 1. Representação simplificada do conceito de uma biorefinaria e exemplos de
possíveis produtos. Adaptado de (White, 2015).
A biomassa vegetal é o recurso orgânico renovável mais abundante, sendo produzida
a partir da fotossíntese das plantas diretamente de luz, gás carbônico e água. A
lignocelulose é composta por celulose, hemicelulose, lignina, pectina e outras
substâncias em menores quantidades (Ghaffar & Fan, 2013; Kumar, Barrett, Delwiche,
& Stroeve, 2009). Quando submetidos à degradação enzimática, os polissacarídeos
celulose e hemicelulose são convertidos em glicose e outros açúcares

14
fermentescíveis, os quais poderão ser convertido a combustíveis líquidos e diversos
outros produtos de valor agregado (Kamm, Kamm, Schmidt, Hirth, & Schulze, 2006).
Em geral, a conversão de materiais lignocelulósicos a açúcares fermentescíveis
envolve as etapas de pré-tratamento e degradação enzimática. O pré-tratamento
(químico, físico e/ou biológico) auxilia a ação enzimática na celulose, ao alterar ou
remover a hemicelulose e/ou a lignina, aumentar a área superficial e diminuir o grau
de polimerização e cristalinidade da celulose (Canilha et al., 2013).
Durante a etapa de degradação enzimática da celulose, três tipos de enzimas
hidrolíticas constituem o complexo celulolítico: (I) exo-1,4-β-D-glicanases que
hidrolisam a cadeia celulósica a partir de suas extremidades liberando celobiose, (II)
endo-1,4-β-D- glicanases que hidrolisam a cadeia celulósica internamente de maneira
aleatória, e (III) 1,4-β-D-glicosidases que promovem a hidrólise da celobiose em
glicose e podem também liberar unidades glicosídicas a partir de
celooligossacarídeos. Essas enzimas atuam em sinergia na degradação da celulose,
criando sítios acessíveis umas para as outras e aliviando problemas de inibição por
produtos (Figura 2) (Canilha et al., 2013; Glass, Schmoll, Cate, & Coradetti, 2013).
Além destas, hemicelulases podem ser adicionadas à biomassa durante a etapa
enzimática, sendo que estas enzimas constituem um grupo de caráter bastante
diverso, o que está diretamente relacionado caráter heterogêneo dos polímeros que
compõem a classe das hemiceluloses: (I) endo-1,4-β-D- xilanases que hidrolisam
ligações glicosídicas internas aleatoriamente na cadeia de xilana, (II) 1,4-β-D-
xilosidases que atuam em xilobiose e/ou xilooligossacarídeos a partir da extremidade
não redutora liberando xilose, (III) endo-1,4-β-D-mananases que clivam ligações
internas na cadeia de manana, (IV) 1,4-β-D-manosidases que clivam
manooligossacarídeos em manose, e (V) enzimas acessórias que removem os grupos
substituintes laterais (ramificações), como α-D-galactosidases, α-L-
arabinofuranosidases, α-D- glicuronidases, acetil xilana esterases e feruloil esterases
(Figura 2) (Canilha et al., 2013; Decker, Siika-Aho, & Viikari, 2009; Scheller & Ulvskov,
2010). Essas enzimas envolvidas na clivagem de carboidratos complexos, bem como
aquelas relacionadas à sua biossíntese, são atualmente conhecidas como enzimas
ativas em carboidratos (CAZymes) (Cantarel et al., 2009; Levasseur, Drula, Lombard,
Coutinho, & Henrissat, 2013; Lombard, Golaconda Ramulu, Drula, Coutinho, &
Henrissat, 2014)
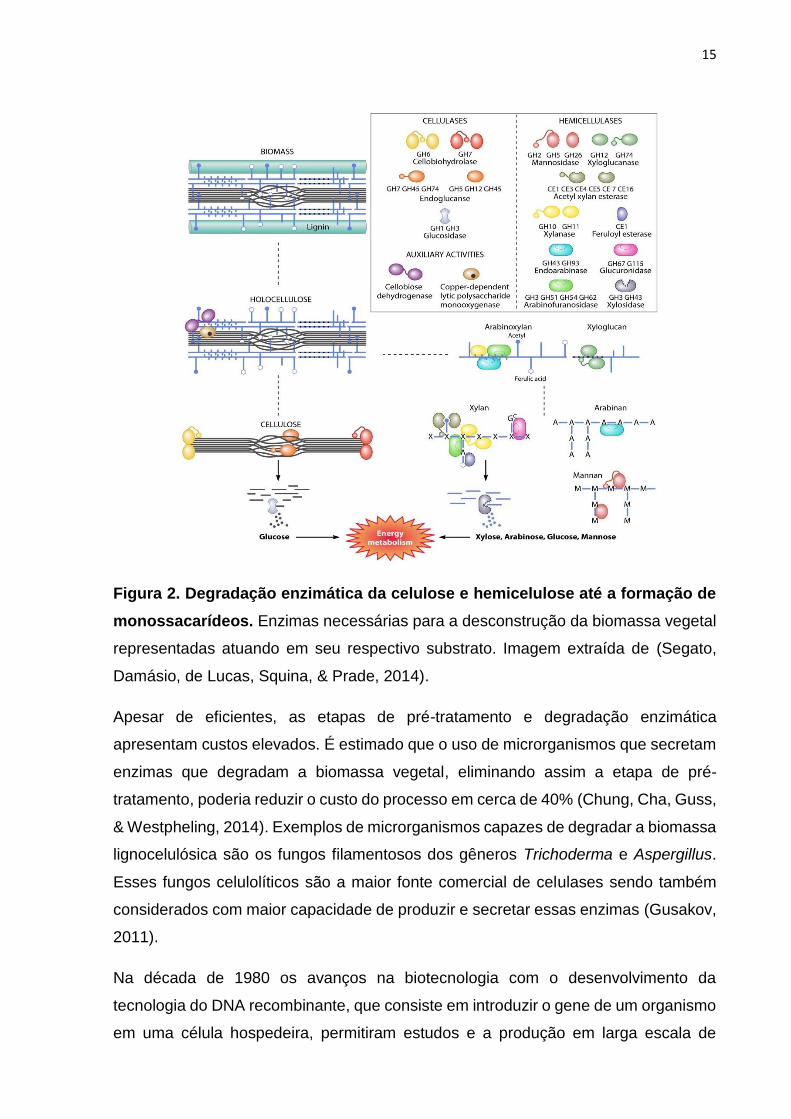
15
Figura 2. Degradação enzimática da celulose e hemicelulose até a formação de
monossacarídeos. Enzimas necessárias para a desconstrução da biomassa vegetal
representadas atuando em seu respectivo substrato. Imagem extraída de (Segato,
Damásio, de Lucas, Squina, & Prade, 2014).
Apesar de eficientes, as etapas de pré-tratamento e degradação enzimática
apresentam custos elevados. É estimado que o uso de microrganismos que secretam
enzimas que degradam a biomassa vegetal, eliminando assim a etapa de pré-
tratamento, poderia reduzir o custo do processo em cerca de 40% (Chung, Cha, Guss,
& Westpheling, 2014). Exemplos de microrganismos capazes de degradar a biomassa
lignocelulósica são os fungos filamentosos dos gêneros Trichoderma e Aspergillus.
Esses fungos celulolíticos são a maior fonte comercial de celulases sendo também
considerados com maior capacidade de produzir e secretar essas enzimas (Gusakov,
2011).
Na década de 1980 os avanços na biotecnologia com o desenvolvimento da
tecnologia do DNA recombinante, que consiste em introduzir o gene de um organismo
em uma célula hospedeira, permitiram estudos e a produção em larga escala de

16
enzimas de interesse (Alvarez-Leefmans & Delpire, 2010; Pasternak, 2005).
Atualmente, a biotecnologia aprimorou a manipulação genética permitindo a produção
de proteínas recombinantes em diversos microrganismos e tipos celulares. As
aplicações envolvendo enzimas aumentaram em quantidade e aplicabilidade
seguindo a demanda do mercado global (Fleiβner & Dersch, 2010; Owen P. Ward,
2012). De acordo com a empresa Ameri Research Inc., o mercado global de enzimas
está em constante crescimento, sendo estimada a movimentação de US$ 11,1 bilhões
em 2018 e podendo atingir US$16,9 bilhões em 2024.
É possível realizar a produção de enzimas recombinantes em sistemas procarióticos
e eucarióticos, os quais apresentam suas especificidades conforme mostrado na
Tabela 1. No geral, os sistemas eucarióticos têm como vantagem a capacidade de
secretar proteínas ao meio externo e realizar modificações pós-traducionais (MPTs).
Dentre os sistemas eucarióticos destacam-se os fungos por serem capazes de
produzir elevados níveis de proteínas com baixo custo, apresentando assim melhor
custo-benefício. Apesar das similaridades entre os fungos filamentosos e
leveduriformes, este último leva desvantagem por produzir proteínas hiper-
glicosiladas o que pode afetar negativamente a atividade das proteínas produzidas
(Fleiβner & Dersch, 2010; Tang et al., 2016). Além disso, os fungos filamentosos
apresentam maior capacidade de secreção do que os demais sistemas de expressão
eucarióticos (Fleiβner & Dersch, 2010).
Produção de enzimas de interesse por Aspergillus spp.
Dentre os microrganismos utilizados em processos industriais, os fungos filamentosos
do gênero Aspergillus são de grande destaque. Este gênero compreende cerca de
350 espécies as quais podem viver numa grande variedade de ambientes, como no
solo e parasitando plantas e animais, resultado da diversidade metabólica e da
capacidade natural de secreção de diferentes enzimas no meio externo (Varga et al.,
2014; O.P. Ward, Qin, Dhanjoon, Ye, & Singh, 2005). Tais enzimas degradam os
compostos e permitem que os fungos absorvam os nutrientes do ambiente (Fleiβner
& Dersch, 2010). Dentre as principais enzimas produzidas por esses fungos, podem
ser citadas celulases, xilanases e proteases (Vries & Visser, 2001).

17
Tabela 1. Características de diferentes sistemas de expressão para a produção de proteínas recombinantes.
Características E. coli Fungos filamentosos Leveduras Células de inseto Células de
mamíferos
Cultura
celular de
plantas
Crescimento celular horas a dias dias a 1 semana dias a 1 semana dias a 1 semana Semanas Meses
Custo do meio de
cultura baixo a médio baixo a médio baixo a médio alto alto médio a alto
Nível de expressão baixo a alto baixo a alto baixo a alto baixo a alto baixo a alto baixo
Capacidade de
Secreção Secreção via periplasma Secreção para o meio de
cultura
Secreção para o meio de
cultura
Secreção para o meio
de cultura
Secreção para o
meio de cultura
Secreção
para o meio
de cultura
Modificações pós-traducionais
Enovelamento da
proteína
Reenovelamento
geralmente necessário
Reenovelamento pode
ser necessário
Reenovelamento pode
ser necessário Enovelamento correto
Enovelamento
correto
Enovelamento
correto
N-glicosilação
Nenhuma
Core igual ao de
mamíferos, sem ácido
siálico, sem açúcares
humanos
High mannose, sem
ácido siálico, sem
açúcares humanos
Complexo, sem ácido
siálico, sem açúcares
humanos
Complexo, sem
açúcares humanos,
e.g. por células de
murinos
Complexo,
sem ácido
siálico, sem
açúcares
humanos
O-glicosilação Não Sim Sim Sim Sim Sim
Fosforilação Não Sim Sim Sim Sim Sim
Acetilação Não Sim Sim Sim Sim Sim
Adaptado de (Fernandez, J.M. & Hoeffler, J.P., 1999; K. M. H. Nevalainen, Te’o, & Bergquist, 2005).

18
Os fungos possuem excelentes sistemas para expressão e secreção de proteínas
homólogas e heterólogas. Muitas proteínas possuem MPTs como glicosilações,
sulfatações e fosforilações, as quais são necessárias para sua correta funcionalidade,
sendo que esta característica representa uma grande vantagem na utilização dos
fungos como modelo de expressão e secreção de proteínas (G. Liu, Qin, Li, & Qu,
2013). A facilidade de se separar da biomassa e o crescimento rápido em meios de
cultivo de baixo custo, representam ainda mais vantagens para a utilização de
Aspergillus como produtores de enzimas (Fleiβner & Dersch, 2010; K. M. H.
Nevalainen et al., 2005). Além disso, as enzimas produzidas por Aspergillus niger e
Aspergillus oryzae, por exemplo, podem ser utilizadas na indústria de maneira segura,
o que resultou com a classificação de status GRAS (Generally Regarded As Safe) (R
J Gouka, Punt, & van den Hondel, 1997).
Ainda que Aspergilli apresentem características ótimas para serem utilizados como
produtores de enzimas em escala industrial, espécies de ocorrência natural não as
produzem em altas quantidades. Com o desenvolvimento da engenharia genética e
da biologia molecular, o melhoramento de linhagens fúngicas visando a alta produção
de enzimas heterólogas e homólogas, pôde se tornar uma realidade. Muitos estudos
envolvendo fungos do gênero Aspergillus modificados para serem utilizados como
hospedeiros na produção de enzimas homólogas e heterólogas têm sido relatados
nas últimas décadas (Devchand & Gwynne, 1991; Jeenes, Mackenzie, Roberts, &
Archer, 1991; Lubertozzi & Keasling, 2009; Nayak et al., 2006; Yoon, Maruyama, &
Kitamoto, 2011).
Apesar dos fungos filamentosos possuírem um sistema de secreção de alta
capacidade, estes têm falhado na produção de grandes quantidades de proteínas
heterólogas, quando comparados com proteínas homólogas. Estes problemas
parecem estar relacionados a múltiplos fatores: (I) baixa eficiência de transformação;
(II) altos níveis de proteases ou substâncias tóxicas produzidos por algumas espécies;
(III) alterações pós-traducionais promovidas por proteases ou por baixo pH (R J Gouka
et al., 1997). Estes fatores, que influenciam negativamente a maturação das proteínas
no retículo endoplasmático (RE), parecem ser a chave para obtenção do aumento nas
taxas secreção de proteínas homólogas e heterólogas (Owen P. Ward, 2012).

19
Aspergillus nidulans se destaca dentro do gênero Aspergillus, pois é a espécie mais
bem caracterizada geneticamente (Owen P. Ward, 2012). Tal espécie é alvo de
pesquisas há mais de 60 anos, o que permitiu o avanço nos conhecimentos sobre a
fisiologia celular eucariótica, contribuindo assim para a compreensão da regulação
metabólica, controle do ciclo celular, estrutura da cromatina, controle de pH, dentre
outros (Galagan et al., 2005; Pontecorvo, Roper, Chemmons, Macdonald, & Bufton,
1953).
Secreção de proteínas recombinantes em fungos filamentosos
Nas últimas décadas, um grande esforço foi realizado para compreender os “gargalos”
da secreção de proteínas heterólogas em fungos filamentosos e como melhorar o seu
rendimento quando comparado ao de proteínas homólogas. Nevalainen e Peterson
elegantemente apresentaram a seguinte pergunta: “Produzir proteínas recombinantes
em fungos filamentosos – estamos esperando demais? (H. Nevalainen & Peterson,
2014).
Na maquinaria celular eucariótica, após tradução do mRNA maduro, as proteínas
destinadas à secreção são translocadas para o lúmen do RE, onde sofrem MPTs e
são enoveladas corretamente passando por um rígido controle de qualidade. Uma vez
corretamente enovelada, é então encaminhada ao Golgi através de vesículas de
transporte, onde sofrerá modificações como o refinamento da N-glicana (Conesa,
Punt, van Luijk, & van den Hondel, 2001; Schwarz & Aebi, 2011). Os fungos
filamentosos, assim como em Saccharomyces cerevisiae, apresentam a N-glicana do
tipo high-mannose, mas em níveis reduzidos, não apresentando hiper-glicosilação
(Deshpande, Wilkins, Packer, & Nevalainen, 2008). Por fim a vesículas de secreção
encaminharão a proteína “madura” até a membrana plasmática, onde serão
secretadas para o meio extracelular (Alberts, B. Johnson, A. Lewis, J. Raff, M. Roberts,
K. Walter, 2008). Todos estes processos apresentados são fundamentais para a
secreção de proteínas em sua funcionalidade correta, porém todos os passos podem
apresentar problemas durante a secreção de proteínas recombinantes.
Após a translocação pelo ER, as proteínas devem ser corretamente enoveladas em
sua forma nativa e biologicamente ativa. Este processo de "maturação" envolve uma
série de chaperonas e foldases, tais como BiP (bipA), dissulfeto isomerases (pdiA,
tigA, prpA), peptidil-prolyl cis-trans isomerase (cypB) e calnexina (clxA). A correlação

20
entre a superprodução de proteínas fúngicas e a super-expressão de bipA ainda não
é clara, uma vez que os níveis de BipA permanecem inalterados ou podem aumentar
dependendo da proteína que está sendo produzida (Punt et al., 2002).
O nível de produção celular de proteínas pode-se alterar em condições naturais como
fase do ciclo de vida, diferenciação celular, e mudanças de condições ambientais. No
entanto, técnicas de biotecnologia podem ser aplicadas para elevar a produção
proteica (Ron & Walter, 2007). Nessas situações, a célula pode produzir uma grande
quantidade de proteínas que excede a capacidade de enovelamento pelo RE,
podendo levar ao acúmulo de proteínas mal enoveladas (C. Rubio et al., 2011). O
acúmulo é nocivo à célula, prejudica as funções celulares, podendo desencadear
morte celular prematura (Ron & Walter, 2007).
Pesquisadores têm observado que a super-expressão de genes que codificam
enzimas de interesse, ativa um sistema de resposta ao estresse chamado de unfolded
protein response (UPR) (Saloheimo, Lund, & Penttilä, 1999). O UPR é ativado quando
a demanda por proteínas na célula excede a capacidade de enovelamento do RE,
organela na qual as proteínas direcionadas para secreção devem passar para serem
corretamente enoveladas (McCracken & Brodsky, 2000; C. Rubio et al., 2011;
Ruggiano, Foresti, & Carvalho, 2014; Walter & Ron, 2011). Assim, pela ativação desse
sistema, maior quantidade de proteínas adquirem a conformação correta e podem
deixar o RE em direção ao meio extracelular, não sendo direcionadas à degradação.
Contudo, existe alguma relação entre glicosilação e UPR? Li et al. mostraram que a
diminuição dos níveis de glicosilação, a partir da redução da expressão de genes do
complexo da oligossacaril transferase, levam às condições de estresse na célula. O
estresse desenvolvido pela baixa glicosilação de algumas proteínas promove a super-
expressão de diversos genes, entre eles genes relacionados a biogênese de parede
celular, enovelamento e degradação de proteínas mal enoveladas, genes estes
característicos da ativação do UPR (K. Li et al., 2011).
Obstáculos para a produção de proteínas recombinantes em fungos
filamentosos
Durante séculos, os fungos filamentosos foram conhecidos por sua capacidade de
secretar grandes quantidades de proteínas. O interesse biotecnológico no gênero
Aspergillus e Trichoderma promoveram o aprimoramento de técnicas de biologia

21
molecular para a produção de proteínas homólogas e heterólogas. Desde meados do
século XX, a produção de proteínas heterólogas em fungos filamentosos apresentou
vários "gargalos". Problemas no rendimento de secreção de proteínas foram
resolvidos ao longo dos anos por diferentes abordagens que serão analisadas neste
tópico.
Embora a secreção de proteínas heterólogas tenham mostrado rendimentos mais
baixos, elas ainda apresentam algumas vantagens em relação à secreção de
proteínas homólogas. Os fungos filamentosos podem secretar quantidades maiores
de proteínas com MPTs (glicosilação, fosforilação, acetilação, metilação,
palmitoilação, ubiquitinação, formação de ligações de dissulfeto, proteólise e outros)
a baixo custo (Bhadauria et al., 2007; Leach & Brown, 2012; K. M. H. Nevalainen et
al., 2005; Liping Wang, Ridgway, Gu, & Moo-Young, 2005). As MPTs são processos
covalentes muito importantes para as propriedades funcionais de proteínas, afetando
atividade, estabilidade e localização (Bhadauria et al., 2007; Jensen, 2004). Mais de
300 tipos de MPTs são atualmente conhecidas e outras novas são constantemente
descobertas (Jensen, 2004). Os obstáculos existentes no processo de secreção de
proteínas recombinantes podem ocorrer em três etapas: protocolo de transformação
de fungos, nível de transcrição e nível de tradução (Figura 3).
“Gargalos” em protocolos de transformação de fungos filamentosos
O primeiro obstáculo observado na produção de proteínas heterólogas está no
protocolo de transformação. Os primeiros protocolos de transformação desenvolvidos
foram descritos no final dos anos 70 usando Neurospora crassa (Case et al., 1979)
como hospedeiro, logo seguido por A. nidulans (Tilburn et al., 1983). O protocolo que
utiliza protoplastos é o método de transformação mais comumente empregado, no
entanto, existem vários outros métodos, incluindo a técnica de acetato de lítio,
eletroporação, biolítica/biobalística, agitação com esferas de vidro, infiltração a vácuo,

22
Figura 3. Uma visão geral da via de secreção de proteínas em fungos
filamentosos. Os diferentes estágios podem apresentar problemáticas a nível
transcricional e/ou traducional. Os números presentes nas estruturas representam
seus respectivos “gargalos” na obtenção de proteínas recombinantes. 1)
processamento de pré-mRNA incorreto e baixos níveis de transcrição, 2) baixa
estabilidade do mRNA, 3) falhas no processo de tradução, MPTs e enovelamento, 4)
transporte intracelular e secreção, 5) processamento incorreto e 6) degradação por
proteases. Adaptado de (Rubio et al., 2015)
ondas de choque e mediada pela bactéria Agrobacterium tumefaciens (Casas-Flores,
Rosales-saavedra, & Herrera-Estrella, 2004; Chakraborty, Patterson, & Kapoor, 1991;
Dhawale, Paietta, & Marzluf, 1984; M. J. A. de Groot & Bundock, 1998; Hynes, 1996;
Ruiz-Díez, 2002; Owen P. Ward, 2012). Atualmente, muitas modificações têm sido
sugeridas nas técnicas padrão de transformação, sobretudo para aumentar sua
eficiência (Chai et al., 2013; Magaña-Ortíz et al., 2013; Rivera, Magaña-Ortíz, Gómez-
Lim, Fernández, & Loske, 2014). É importante lembrar que a seleção do protocolo a
ser utilizado depende do organismo hospedeiro e da acessibilidade dessas técnicas
(Tabela 2).

23
Tabela 2. Métodos padrão para a transformação de fungos.
Método Procedimento Vantagem Desvantagem
Eletroporação
Pulsos elétricos induzem a permeabilização
da membrana, fornecendo uma força motriz
local para o transporte iônico e molecular
através dos poros.
Pode ser aplicado a qualquer fungo
in vivo ou congelado. Protocolos
eficientes: simples, rápidos e
facilmente otimizados.
Depende das características
eletrofisiológicas do fungo. Baixa
eficiência. Custo médio.
Biolítica/Biobalística Pequenas partículas cobertas de genes são
aceleradas para penetrar na parede celular.
Simples. Não é necessário um pré-
tratamento da parede celular.
Independente das propriedades
fisiológicas dos fungos.
Transformação com transgênese
múltipla possível.
DNA pode ser danificado. Produz
múltiplas cópias dos genes
introduzidos. Protocolos complexos
devido à preparação do projétil.
Baixa eficiência. Custo elevado.
Agitação com
esferas de vidro
A agitação das células fúngicas com
esferas de vidro na presença de carreador
e DNA plasmidial permite a absorção do
material genético.
Protocolo simples, rápido e seguro.
Baixo custo.
Pode causar a interrupção celular.
As células requerem suporte
osmótico. Baixa eficiência.
Infiltração a vácuo
Vácuo gera pressão atmosférica negativa
que faz com que os espaços de ar entre as
células diminuam, permitindo a infiltração
de bactérias, como Agrobacterium.
Simples e rápido. Regeneração de
fungos in vitro. Eficiência média.
Requer o uso de bactérias que
podem ter consequências
indesejadas.
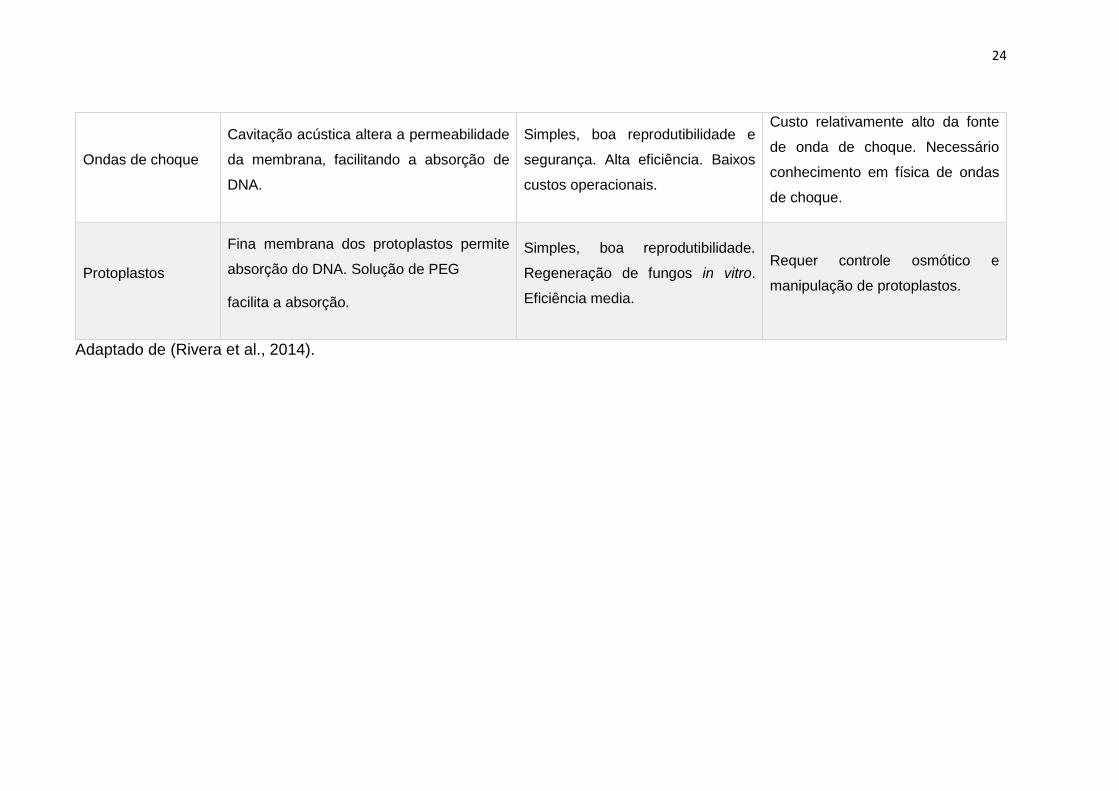
24
Ondas de choque
Cavitação acústica altera a permeabilidade
da membrana, facilitando a absorção de
DNA.
Simples, boa reprodutibilidade e
segurança. Alta eficiência. Baixos
custos operacionais.
Custo relativamente alto da fonte
de onda de choque. Necessário
conhecimento em física de ondas
de choque.
Protoplastos
Fina membrana dos protoplastos permite
absorção do DNA. Solução de PEG
facilita a absorção.
Simples, boa reprodutibilidade.
Regeneração de fungos in vitro.
Eficiência media.
Requer controle osmótico e
manipulação de protoplastos.
Adaptado de (Rivera et al., 2014).

25
“Gargalos” a nível transcricional
A produção de proteínas heterólogas pode ser limitada em nível transcricional, por
uma série de fatores tais como: a instabilidade do mRNA, o processamento incorreto
do pré-mRNA e baixos níveis de transcrição (Sharma, Katoch, Srivastava, & Qazi,
2009). A estabilidade do mRNA é afetada por alguns componentes estruturais: o cap
5' de 7-metilguanosina trifosfato, a cauda 3'-poli (A), o comprimento do mRNA, as
modificações pós-transcricionais de bases, tais como a metilação de resíduos de
adenina ou a conversão de adeninas em inosinas e sequências estabilizadoras ou
desestabilizadoras de mRNA (R J Gouka et al., 1997; Hentze, 1991).
As falhas no processo de transcrição podem gerar estruturas incorretas de mRNA. O
processamento incorreto do pré-mRNA pode produzir transcritos truncados como
mostrado na produção de α-galactosidase heteróloga por Aspergillus awamori (Robin
J Gouka, Punt, & van den Hondel, 1997). O pré-mRNA sofre alterações estruturais em
células eucarióticas, sendo que este processamento implica no reconhecimento de
regiões de poliadenilação (ricas de AU) (R J Gouka et al., 1997). Regiões de
poliadenilação que não a cauda 3'-poli (A) podem resultar em interrupção prematura
da transcrição do mRNA, produzindo mRNAs truncados como no exemplo com α-
galactosidase. Dessa maneira, os baixos níveis de RNAs podem ser resultado da
degradação devido à baixa estabilidade, bem como do processamento e conformação
incorretos.
“Gargalos” a nível traducional
Além de MPTs e o controle de qualidade do RE, algumas proteínas são submetidas à
cisão, i.e. há uma proenzima que quando clivada, resulta em uma enzima na forma
ativa (Bell & Malmberg, 1990; Hoyt, Williams-Abbott, Pitkin, & Davis, 2000). Hoyt et al.
mostrou o processamento da S-adenosil metionina descarboxilase de N. crassa
quando comparado com outros organismos. Na ausência ou falha deste
processamento, as proteínas permaneceram como proenzima afetando sua atividade
(Hoyt et al., 2000). A baixa taxa de códons no organismo hospedeiro está diretamente
relacionada ao baixo nível de tRNA para esta sequência, o que pode interromper a
transcrição prematuramente, resultando em alterações no quadro de leitura e falhas
(M. Tanaka, Tokuoka, & Gomi, 2014).

26
Apesar dos problemas de tradução descritos acima, as proteínas heterólogas ainda
precisam superar a presença de proteases nativas. Mesmo quando as proteínas
heterólogas são secretadas com sucesso, as proteases fúngicas nativas presentes
nos meios de cultura podem diminuir o seu rendimento por degradação (Archer &
Peberdy, 1997). Para compreender e regular as proteases extracelulares, algumas
foram clonadas e estudadas individualmente, através do desenvolvimento de cepas
com genes de proteases deletados ou silenciados. Estudos relataram maiores
rendimentos de proteína heteróloga usando cepas de hospedeiro com suas proteases
deletadas (Sharma et al., 2009; O.P. Ward et al., 2005). Fungos filamentosos,
geralmente, apresentam grande quantidade de genes de proteases em seu genoma;
A. nidulans, por exemplo, possui em torno de 80 genes (Sharma et al., 2009).
Estratégias para superar os principais “gargalos”
Nível transcricional
Os “gargalos” transcricionais são a baixa estabilidade do mRNA, processamento
incorreto do mRNA e o nível de transcritos heterólogos. A baixa estabilidade de mRNA
pode ser superada pela introdução de uma sequência de íntron endógena na
sequência de cDNA do gene heterólogo. Esta estratégia foi projetada para promover
o splicing correto e para aumentar a estabilidade do mRNA. Outra maneira de
melhorar a estabilidade do mRNA é a fusão do gene alvo com genes endógenos
altamente expressos. O gene heterólogo pode ser fusionado em um sinal eficiente de
expressão fúngica ou na extremidade 3' de um gene endógeno altamente expresso.
Tanaka et al. relataram aumento de 10 a 90 vezes na atividade de uma beta-
glicosidase em arroz transgênico ao usar um íntron endógeno, porém esta estratégia
ainda não foi descrita em fungos filamentosos (A. Tanaka et al., 1990).
O processamento incorreto causado por regiões ricas em AT ou códons raros pode
ser evitado otimizando-se a sequência do gene alvo. Nos genes sintéticos, as regiões
ricas em AT e os códons raros podem ser eliminados pela otimização de códons. As
otimizações de códons são realizadas com base na frequência de uso do códon do
organismo hospedeiro. Esta técnica é amplamente utilizada para expressar proteínas
heterólogas em fungos filamentosos de forma bem-sucedida (Gustafsson,
Govindarajan, & Minshull, 2004). Elevados níveis de mRNA são obtidos com a
otimização de códons, pois geralmente melhora a eficiência da transcrição. Essa

27
técnica tem sido utilizada para elevar os níveis de mRNA desde 2012, quando foi
descrita por Takada et al. em Aspergillus oryzae (M. Tanaka, Tokuoka, Shintani, &
Gomi, 2012). Chen et al. observaram que a presença de A ou U na terceira posição
de códons de baixa frequência é comum em Aspergillus spp. e outros fungos
filamentosos (Wanping Chen, Xie, Shao, & Chen, 2012). Portanto, a otimização de
códons tende a elevar o conteúdo de CG (Tokuoka et al., 2008) e o tempo de meia-
vida (M. Tanaka et al., 2012), eliminando a poliadenilação prematura. Com o objetivo
de criar um banco de dados para acadêmicos, Nakamura et al. analisaram o uso de
códons com base nas sequências de codificação de proteínas completas do GenBank
criando o “Codon Usage Database” (Nakamura, Gojobori, & Ikemura, 1998).
O número de cópias de genes pode influenciar os níveis de transcrição. Por cerca de
três décadas, ferramentas genéticas moleculares foram usadas para expressar cópias
extras de genes heterólogos de interesse em fungos filamentosos para elevar a sua
produção (Punt et al., 2002; Liping Wang et al., 2005). As cópias de genes de interesse
são geralmente expressas sob um promotor homólogo apropriado (O.P. Ward et al.,
2005). Na produção de proteínas heterólogas por Aspergillus, a álcool desidrogenase
I (alcA) de A. nidulans ou a glucoamilase (glaA) de A. niger são, geralmente, utilizados
como promotores fortes (Lubertozzi & Keasling, 2009). Veredoes et al. mostraram que
múltiplas cópias do gene heterólogo é capaz de melhorar a produção de proteínas em
A. niger, no entanto, a expressão pôde ser inativada por metilação quando grandes
quantidades (>200) do gene heterólogo foram induzidas (Archer & Peberdy, 1997;
Verdoes et al., 1993).
Nível traducional
Os problemas na iniciação da tradução são geralmente corrigidos de duas maneiras:
através da fusão do gene heterólogo à extremidade 3' de um gene endógeno
altamente expresso; ou pela fusão a um fragmento promotor endógeno. A
identificação da região endógena melhora a translocação da proteína recombinante,
o enovelamento e também evita a degradação por proteases. Além disso, a
interrupção ao longo da síntese da proteína pode ser evitada usando genes sintéticos
ou através de códons otimizados para o organismo hospedeiro. Portanto, erros no
quadro de leitura causados por códons raros não irão ocorrer e, consequentemente,

28
a eficiência da tradução será aprimorada (Fleiβner & Dersch, 2010; R J Gouka et al.,
1997).
Os fungos filamentosos são capazes de promover diversas MPTs com eficácia, sendo
que algumas delas estão descritas na Tabela 3. As modificações estão diretamente
relacionadas às propriedades funcionais e estruturais das proteínas as quais, dessa
maneira, podem ser prejudicadas caso estas sejam realizadas incorretamente. A N-
glicosilação é uma MPT muito importante e, embora esteja envolvida no correto
enovelamento de proteínas, seu reconhecimento permite a secreção de proteínas de
acordo com a via clássica. Como alternativa à via clássica de secreção de proteínas,
Sagt et al. criaram a chamada peroxisecretion. O método foi desenvolvido para
secretar proteínas intracelulares nativas, pois estas não contêm sinais de glicosilação
para o tráfego pela via secretória clássica. A técnica fusiona a proteína da membrana
peroxissomal com a proteína heteróloga. Dessa maneira, a proteína alvo enovelada
adquire a capacidade de ser transportada em uma vesícula e, posteriormente,
transferida para o citosol. A composição da membrana lipídica, semelhante às
vesículas secretoras do ER, permite a fusão com a membrana plasmática e
consequentemente sua secreção ao meio extracelular (Cees M J Sagt et al., 2009).
Os processos relacionados a via de secreção em fungos filamentosos, bem como o
controle de qualidade do enovelamento pelo ciclo da calnexina e o UPR, também
podem ser manipulados para aumentar a eficiência da secreção. Assim, a super-
expressão de genes relacionados ao UPR, como hacA, foldases e chaperonas , tem
resultado em maior produção de proteínas heterólogas em fungos filamentosos (K. M.
H. Nevalainen et al., 2005). A super-expressão de calnexina resultou no aumento da
secreção da manganês peroxidase de Phanerochaete chrysosporium (Conesa,
Jeenes, Archer, van den Hondel, & Punt, 2002). No caso de Bip, chaperona
relacionada ao UPR, a sua super-expressão resultou em um aumento de cinco vezes
da produção de eritropoietina por S. cerevisiae (R J Gouka et al., 1997).

29
Tabela 3. Tipos de modificações pós-traducionais em fungos filamentosos. Adaptado de (Rubio et al., 2015).
Modificações pós-traducionais Mecanismos Funções
Glicosilação
N-Glicosilação
O-Glicosilação
N-Glicosilação:
Anexação de uma glicana a resíduos de
asparagina em proteínas alvo através de uma
ligação amida
O-Glicosilação:
Anexação de glicanas lineares curtas através de
ligações a resíduos de serina ou treonina.
Secreção, estabilidade, localização e
reconhecimento ambiental (Deshpande et al.,
2008)
Formação de ligação dissulfeto
As ligações dissulfeto são formadas entre os
átomos de enxofre de pares de resíduos de
cisteína dentro ou entre proteínas
Estabilidade (Bulaj, 2005)
Ubiquitinação Ligação covalente da ubiquitina Estabilidade, localização e atividade proteica
(Pickart & Eddins, 2004)
Proteólise Processo de quebrar ligações peptídicas em
proteínas, realizadas por peptidases e proteases
Ativação, inativação, função proteica alterada
e regulação (Rogers & Overall, 2013)
Fosforilação Adição de um ou mais grupo fosfato a motivos
específicos, consistindo frequentemente em
Ativação e inativação da atividade enzimática
e transdução de sinal (Seo & Lee, 2004)

30
alguns resíduos-chave que envolvem o
aminoácido alvo
Palmitoilação A ligação do tioéster de palmitato a resíduos de
cisteína em proteínas
Modulação da atividade proteica, tráfico e
interações de membrana.(Smotrys & Linder,
2004)
Sumoilação O SUMO é ligado covalentemente através de uma
ligação isopeptídica ao grupo amino de resíduos
de lisina alvo em substratos de proteínas
específicos
Atividade proteica (Wong et al., 2008)
Neddilação A adição covalente do polipeptídio NEDD8 às
proteínas alvo através de uma ligação isopeptídica
entre a glicina C-terminal de NEDD8 e uma lisina
na proteína alvo
Regulação da estrutura e da função celular
(Mathewson et al., 2013)
Miristoilação Anexação de ácido mirístico, um ácido graxo
saturado de 14 carbono, a Glicina N-terminal das
proteínas
Regulação da estrutura e da função celular
(Moriya et al., 2013)
Âncora de GPI A ancoragem de GPI é um mecanismo para fixar
as proteínas à superfície da célula através de uma
ligação amida; tem sido amplamente revisado em
leveduras
Viabilidade celular (Mayor & Riezman, 2004)

31
Desde a década de 90 cepas deficientes em proteases têm sido utilizadas para
superar problemas de rendimento resultantes da ação de proteases endógenas.
Atualmente, algumas abordagens moleculares desenvolvidas visam silenciar ou
deletar esses genes. Os resultados na literatura mostraram uma melhora nos
rendimentos das proteínas heterólogas, mas essa estratégia varia de acordo com a
sensibilidade de proteínas heterólogas às proteases hospedeiras (Sharma et al.,
2009). Apesar de espécies do mesmo gênero apresentarem proporções similares de
proteases, cada fonte de carbono é capaz de induzir diferentes níveis de proteases
extracelulares (Liping Wang et al., 2005). Yoon et al. compararam rendimentos de
produção de proteínas heterólogas em uma cepa selvagem A. oryzae e cepas
knockout para cinco e dez genes de protease. A produção de proteínas heterólogas
foi 30% maior na cepa apresentando dez genes deletados quando comparado com a
cepa com cinco proteases deletadas e foi 3,8 vezes maior do que no tipo selvagem
(Yoon et al., 2011). Sharma et al. mostrou que a deleção de apenas uma protease de
A. niger é capaz de elevar a produção heteróloga de uma lacase em 42% (Sharma et
al., 2009).
Glicosilação de proteínas
Após mais de três bilhões de anos de evolução, toda célula viva livre e cada tipo de
célula dentro de organismos multicelulares permanece coberta por uma camada
densa e complexa de glicanas (Colley, Varki, & Kinoshita, 2015). A evolução tem
repetida e consistentemente selecionado glicanas como as moléculas mais diversas
e flexíveis, que estão posicionadas na interface entre a célula e o ambiente
extracelular. Os possíveis motivos para isso incluem a sua relativa hidrofilia,
flexibilidade e mobilidade em ambientes aquosos e sua extrema diversidade,
permitindo adaptações fáceis a curto e longo prazo a ambientes em mudança e
regimes patogênicos (Colley et al., 2015; Jacobs & Callewaert, 2009; Schwarz & Aebi,
2011).
O processo de glicosilação é uma das MPTs mais importantes que ocorrem na
estrutura da maioria das proteínas secretadas. O processo consiste na adição de
oligossacarídeos a proteínas ou lipídios por ligações covalentes (Helenius & Aebi,
2004; Spiro, 2002). Existem dois diferentes tipos de glicosilação em proteínas: O-

32
glicosilação, e a N-glicosilação. A N-glicosilação ocorre no RE e é extremamente
importante para diversos processos celulares tais como: resposta imune,
comunicação celular e transporte, secreção, estabilidade, enovelamento e função de
algumas proteínas.
A N-glicosilação é encontrada em todos os domínios da vida, sendo comum em
Eukaria, frequente em Archaea e raro em Bacteria (Schwarz & Aebi, 2011; Weerapana
& Imperiali, 2006). Em Bacteria, Archaea e Fungi, as glicanas apresentam papéis
estruturais críticos nas paredes celulares, oferecendo resistência a grandes diferenças
de osmolaridade entre o citoplasma e o ambiente circundante (Colley et al., 2015). Em
eucariotos, proteínas secretadas e de membrana, atravessam uma via de RE-Golgi,
sistema celular em que majoritariamente ocorrem reações de glicosilação
(Cherepanova, Shrimal, & Gilmore, 2016; Helenius & Aebi, 2004).
Para a N-ligação dos oligossacarídeos, um precursor com 14 monossacarídeos,
sintetizado na membrana do RE por diversas glicosiltransferases, é transferido ao
resíduo de asparagina alvo na cadeia polipeptídica da proteína. A estrutura desse
precursor é comum na maioria dos eucariotos e geralmente contém 3 moléculas
glicose (Glc), 9 moléculas de manose (Man) e 2 moléculas de N-acetilglicosamina
(GlcNAc) (Burda & Aebi, 1999; Helenius & Aebi, 2004; Spiro, 2002). No entanto, entre
eucariotos e procariotos pode-se encontrar diferentes composições de N-glicana
precursora devido a evolução do sistema de controle de qualidade de enovelamento
e degradação entre os diferentes organismos (Banerjee et al., 2007; Schwarz & Aebi,
2011). A biossíntese da N-glicana em eucariotos já é bem determinada, bem como os
genes envolvidos no processo (Figura 4 e Tabela 4).
A biossíntese se inicia com a transferência de grupamentos GlcNAc-fosfato a partir de
GlcNAc-UDP para o dolicol-fosfato. Em seguida, ocorre a transferência de manose a
partir de manose-GDP para GlcNAc-dolicol-difosfato. A cadeia de oligossacarídeo
formada é então translocada para o interior do RE onde ocorrerá a adição de mais
moléculas de manose e de glicose (Figura 4). Esta cadeia de oligossacarídeo é então
ligada por uma oligossacaril transferase (Ost) à asparagina da sequência consenso
Asn-Xaa-Ser/Thr (Xaa diferente de prolina), regenerando o grupo dolicol-fosfato
(Moremen, Tiemeyer, & Nairn, 2012).

33
Figura 4. Via de biossíntese da N-glicana e enzimas responsáveis em eucariotos.
As enzimas e seus respectivos produtos estão detalhados na Tabela 4 (adaptado de
Complex Carbohydrate Research Center).
Tabela 4. Genes relacionados a biossíntese da N-glicana (Asparagine-Linked
Glycosylation – ALG) em A. nidulans e seus respectivos produtos.
N Gene Locus tag EC
number Molecular function Product
Cytoplasm
1 Alg5 AN7715 2.4.1.117 Dolichol phosphate glucosyltransferase Dolichol phosphate
glucose
2 Dpm1 AN4947 2.4.1.83 Dolichol phosphate mannosyltransferase Dolichol phosphate
mannose
3 Sec59 AN11886 2.7.1.108 Dolichol kinase Dolichol phosphate
4 Alg7 AN5888 2.7.8.15 UDP-N-acetyl-D-glucosamine:dolichol
phosphate GlcNAcPP-Dol
5
6
Alg13/Alg
14
AN11802 /
AN5736 2.4.1.141
Beta-1,4-N-
acetylglucosaminyltransferase GlcNAc2PP-Dol
7 Alg1 AN5346 2.4.1.142 Beta-1,4-mannosyltransferase ManGlcNAc2PP-Dol
8 Alg2 AN6874 2.4.1.132 Alpha-1,3/alpha-1,6-
mannosyltransferase Man3GlcNAc2PP-Dol
9 Alg11 AN5725 2.4.1.- Alpha-1,6-mannosyltransferase Man5GlcNAc2PP-Dol
10 Rft1 AN7425 - Oligosaccharidyl-lipid flippase family Man5GlcNAc2PP-Dol

34
ER
11 Alg3 AN0104 2.4.1.130 Alpha-1,3-mannosyltransferase Man6GlcNAc2PP-Dol
12 Alg9 AN10118 2.4.1.- Alpha-1,2-mannosyltransferase Man7GlcNAc2PP-Dol
11 Alg12 AN3588 2.4.1.130 Alpha-1,3/alpha-1,6-
mannosyltransferase Man8GlcNAc2PP-Dol
13 Alg9 AN10118 2.4.1.- Alpha-1,2-mannosyltransferase Man9GlcNAc2PP-Dol
14 Alg6 AN4864 2.4.1.- Alpha-1,3-glucosyltransferase GlcMan9GlcNAc2PP-
Dol
15 Alg8 AN7301 2.4.1.- Alpha-1,2-glucosyltransferase Glc2Man9GlcNAc2PP-
Dol
16 Alg10 AN5902 2.4.1.- Alpha-1,2-glucosyltransferase Glc3Man9GlcNAc2PP-
Dol
17 Ost1-
alpha AN7472 2.4.1.119 Oligosaccharyltransferase Glc3Man9GlcNAc2
17 Wbp1-
beta AN4683 2.4.1.119 Oligosaccharyltransferase Glc3Man9GlcNAc2
17 Stt3 AN1455 2.4.1.119 Oligosaccharyltransferase Glc3Man9GlcNAc2
Dol: Dolicol; PP: difosfato; GlcNAc: N-acetilglicosamina; Glc: glicose; Man: manose.
O tempo de meia vida da N-glicoproteína prematura é de apenas alguns segundos,
dessa forma, duas glicoses são imediatamente removidas após a adição do
oligossacarídeo. A N-glicana contendo um único resíduo de glicose com ligação α1-3
(GlcMan9GlcNAc2) é o sinal para associar-se com a calnexina (transmembrana) e ou
calreticulina (solúvel), iniciando o controle de qualidade do enovelamento. A calnexina
possui um domínio de ligação a carboidrato e um domínio rico em prolina
responsáveis, respectivamente, pelo reconhecimento da glicana e recrutamento da
oxidoredutase associada a lectina ERp57. A ERp57 é uma dissulfeto isomerase
multifuncional responsável por catalisar a formação de pontes de dissulfeto nativas no
enovelamento da proteína (Maattanen, Kozlov, Gehring, & Thomas, 2006). A
glicoproteína, então enovelada, é liberada da calnexina por meio da remoção do
resíduo de glicose remanescente gerando a estrutura Man9GlcNAc2 (Figura 5) (Aebi,
Bernasconi, Clerc, & Molinari, 2010).

35
Figura 5. Etapas da N-glicosilação e controle de qualidade. Durante a tradução da
proteína, o complexo translocon (SEC61) transloca a proteína para o lúmen do RE,
onde ocorre a transferência de uma glicana ligada a um lipídeo (dolicol-P) para
sequencia passível de glicosilação (Asn-Xaa-Ser/Thr). Adicionada à glicana
precursora, Glc3Man9GlcNAc2, duas glicoses são removidas e a glicana remanescente
é reconhecida por lectinas, calnexina ou calreticulina, entrando no controle de
qualidade. A liberação das proteínas das lectinas se dá através da clivagem da glicose
presente na glicana. As proteínas enoveladas corretamente são encaminhadas ao
complexo de Golgi e as proteínas mal enoveladas retornam ao ciclo de controle de
qualidade por meio de uma re-glicosilação. As proteínas que não atingem o
enovelamento correto são direcionadas ao proteassoma, sinal iniciado pela ação de
α-manosidase. Antes da proteína mal enovelada ser efetivamente degradada ocorre
a remoção da N-glicana pela atividade da enzima PNGase (Moremen et al., 2012).

36
A maturação correta das glicoproteínas pode requerer mais de uma associação com
o ciclo da calnexina. Dessa maneira, o RE de eucariotos possui um sensor de
enovelamento, denominado UGGT (UDP-glucose:glycoprotein glycosyltransferase)
(Samuelson & Robbins, 2015). Esse sensor possui um domínio N-terminal capaz de
ligar em proteínas com estruturas não nativas e um domínio de carboidrato transferase
na porção C-terminal. Assim, essa enzima reconhece glicoproteínas mal enoveladas
contendo o Man9GlcNAc2 e adiciona novamente uma glicose na manose terminal,
permitindo seu retorno ao ciclo na calnexina (Aebi et al., 2010; Moremen et al., 2012).
Portanto, o ciclo da calnexina juntamente com a UGGT determinam quando as
glicoproteínas estão corretamente enoveladas.
Durante processo de adquirir a conformação correta através do ciclo da calnexina e
UGGT, as N-glicanas estão susceptíveis a ação de α-manosidases do RE que podem
remover os resíduos de manose terminais. A remoção desses resíduos dificulta o
retorno das proteínas mal enoveladas ao ciclo da calnexina e direciona esses
polipeptídios para a via de degradação associada ao RE (ER-associated degradation
- ERAD) (Banerjee et al., 2007; Ruggiano et al., 2014). Essa via tem como função
remover do RE os peptídeos e os componentes que não foram corretamente
incorporados durante a via de secreção, impedindo um estresse de RE. A N-glicana
reduzida é, então, reconhecida pela lectina Yos9, que auxilia na translocação ao
citosol via complexo SEL1L (Aebi et al., 2010). Posteriormente, as N-glicanas são
removidas pela ação da peptídeo-N-glicosidase F (PNGase F) e a proteína é
direcionada a degradação via proteassomo (Smith, Ploegh, & Weissman, 2011).
As glicoproteínas que adquiriram a conformação correta após o ciclo da calnexina são
direcionadas à via de secreção com o auxílio de algumas lectinas, tais como ERGIC-
53, VIP36, e VIPL (Helenius & Aebi, 2004). Essas lectinas são capazes de interagir
com COPI e COPII possibilitando o transporte vesicular entre o RE e o compartimento
cis-Golgi (Duden, 2003). No interior do Golgi alguns açúcares da N-glicana poderão
ser removidos e/ou adicionados, aumentando a complexidade dessa estrutura
(Stanley, Taniguchi, & Aebi, 2017). Ao contrário da biossíntese da N-glicana, as
modificações promovidas no Golgi são bastante variáveis até mesmo entre diferentes
tipos celulares (P. Wang et al., 2017). A evolução da diversidade de heterogeneidade

37
das N-glicanas pode estar relacionada com o desenvolvimento de um mecanismo de
defesa à patógenos (Gagneux, Aebi, & Varki, 2015). Em suma, as alterações
promovidas ao longo da via de secreção as glicoproteínas até a obtenção da proteína
funcional estão representadas na Figura 6.
Figura 6. Efeitos da glicosilação em uma proteína. Após a tradução (A) a proteína
é N-glicosilada (B) no interior do RE, auxiliando o seu enovelamento ao entrar no ciclo
da calnexina (C). Ao ser transportada ao Golgi a N-glicana é modificada (D) e a
proteína é O-glicosilada (E). Essas glicosilações auxiliam no direcionamento e
secreção (F). Após a secreção, as glicosilações aumentam a solubilidade (G), a
resistência a proteases, bem como a estabilidade (H). Além disso, a glicosilação
regula a ligação de: ativadores (I), cofatores (J), oligômeros (K), inibidores (L) e
substratos (M); eventualmente, as glicosilações são capazes de modular parâmetros
cinéticos em reações enzimáticas. Adaptado de (Goettig, 2016)
Diversos estudos têm provado que a N-glicosilação é determinante na secreção,
atividade, especificidade ao substrato e estabilidade de algumas enzimas. Na maioria
dos casos, a remoção de todos os sítios de N-glicosilação de uma proteína reduz
consideravelmente seu nível de secreção. No entanto, a remoção individual de sítios
de glicosilação diminui a atividade enzimática, e a deglicosilação resulta, geralmente,
em proteínas com atividade extremamente reduzida (Skropeta, 2009; Yoneda et al.,
2014). Apesar de alguns estudos relatarem alterações na secreção e nas
propriedades biofísicas das proteínas pouco se sabe sobre os mecanismos que geram

38
esse comportamento, tanto em relação aos sítios de glicosilação quanto para a
composição da glicana.
N-glicosilação e biotecnologia
Desde 1805 as glicoproteínas têm sido estudadas a fim de se compreender sua
estrutura e função (Peter-Katalinić, 2005). Dado os avanços na biotecnologia, algumas
estratégias de estudos da N-glicosilação foram desenvolvidas, tais como:
desenvolvimento de cepas knockdown ou knockout; expressão de genes da via de N-
glicosilação; e glicosilação sintética. Os avanços das “ômicas” facilitaram os estudos
envolvendo o processo de N-glicosilação a partir das abordagens de N-
glicoproteômicas e glicômicas (Weixuan Chen, Smeekens, & Wu, 2014; Dam et al.,
2013; Geyer & Geyer, 2006; Lee et al., 2016; Thaysen-Andersen & Packer, 2012; Lu
Wang et al., 2012), bem como o desenvolvimento de ferramentas de bioinformática
(Cooper, Gasteiger, & Packer, 2001).
Diferentes estratégias de biotecnologia utilizando genes relacionados ao processo de
N-glicosilação permitiram desenvolver microrganismos de grande interesse
biotecnológico e econômico. Em 2002 Wacker et al. expressaram o loci de
Campylobacter jejuni responsável pela via de N-glicosilação em E. coli com sucesso
desenvolvendo a primeira E. coli capaz de produzir glicoproteínas (Valderrama-Rincon
et al., 2012; Wacker et al., 2002; Weerapana & Imperiali, 2006). Além disso, em 2008
Kainz et al. desenvolveram cepas de Aspergillus capaz de produzir glicoproteínas
humanizadas a fim de produzir glicoproteínas de interesse biomédico (Kainz et al.,
2008). Uma vez que as N-glicanas influenciam na meia-vida na circulação, na
distribuição dos tecidos e a atividade biológica, cada glicoforma possui seu próprio
perfil farmacocinético, farmacodinâmico e de eficiência.
Estratégias empregadas para estudos de N-glicosilação
Knockdown ou knockout
A redução de atividades indesejáveis de glicosiltransferases em células foi conseguida
por estratégias de silenciamento de genes. Apesar de ter sido bem-sucedido em
plantas e Drosophila, o silenciamento não foi muito adotado, pois a baixa eficiência de
knockdown muitas vezes mantem a atividade de glicosiltransferases remanescente

39
(Nishihara et al., 2004). Knockout de genes envolvidos na biossíntese da N-glicana
tem sido aplicado, em sua maioria, em leveduras a fim de evitar a hiper-glicosilação.
(B. Liu et al., 2009; Tang et al., 2016). Nesses casos, manipular a composição da N-
glicana visa controlar a glicosilação não efetiva, pois esta pode diminuir o rendimento
de proteínas heterólogas produzidas por poder resultar em mal enovelamento da
proteína alvo (Cowcher et al., 2016; Sharma et al., 2009).
Expressão de genes da via de biossíntese de N-glicana
A adição de atividades desejáveis de glicosiltransferase pode ser obtida pela
transformação de genes da biossíntese de N-glicanas homólogos ou heterólogos. O
problema em torno dessa estratégia está no baixo controle na integração do genoma,
números de cópias e expressão gênica. A partir desse recurso, alguns trabalhos têm
detalhado a cinética enzimática das manosiltransferases Alg1, Alg2 e Alg11 (formam,
respectivamente, ManGlcNAc2, Man2GlcNAc2 e Man3GlcNAc2) e com isso
permitindo estudar o mecanismo de ação de enzimas da via de biossíntese de N-
glicanas (S. T. Li et al., 2017; Ramírez et al., 2017).
Glicosilação sintética
Sabe-se que a simples presença do motivo de N-glicosilação não garante que essa
região será glicosilada (aproximadamente 70% recebe a N-glicana (Stanley et al.,
2017)) uma vez que não existe controle genético para MPT. No entanto, a influência
que a N-glicosilação pode exercer sobre as propriedades funcionais e bioquímicas de
uma proteína são bem estudadas (Aebi, 2013; Burda & Aebi, 1999; Schwarz & Aebi,
2011; E. S. Trombetta, 2003). Dessa forma, foi desenvolvida a técnica de ligação
quimio-seletiva mediada por um grupamento tiol (Gamblin et al., 2004). Um resíduo
de cisteína é incorporado na posição desejada e o grupamento tiol de sua cadeia
lateral é subsequentemente convertido em selenil-sulfeto, seguido de uma exposição
de brometo de fenil selenil (Figura 7). Assim, a proteína é convertida em uma
glicoproteína homogênea com N-glicanas especificas em posições conhecidas (Davis,
Van Kasteren, Kramer, & Gamblin, 2007; Wright & Davis, 2017).

40
Figura 7. Exemplificação da metodologia de N-glicosilação sintética.
Os estudos envolvendo N-glicosilação foram realizados, inicialmente, para entender o
papel dos carboidratos em modificações biofísicas na comunicação celular, tinham
como objetivo desenvolver novas abordagens para o tratamento de doenças humanas
(Weixuan Chen et al., 2014; Eshghi, Yang, & Wang, 2014; Moremen et al., 2012). No
entanto, alguns estudos têm mostrado o efeito da glicosilação no enovelamento,
secreção e propriedades enzimáticas (Banerjee et al., 2007). O conhecimento de N-
glicosilação em CAZymes ainda são escassos e majoritariamente estão relacionados
com celobiohidrolases (Beckham et al., 2012b; Ruchi Gupta, Baldock, Fielden, &
Grieve, 2011; Jeoh, Michener, Himmel, Decker, & Adney, 2008).
A glicoengenharia de proteínas é uma ferramenta bastante promissora da
biotecnologia. Hanson et al. relataram que o core conservado da N-glicana,
ManGlcNAc2, são essenciais para a estabilidade, cinética e a termodinâmica de
enovelamento de proteínas apurando a via secretora (Hanson et al., 2009; Price et al.,
2012; Price, Powers, Powers, & Kelly, 2011; N. Wang, Li, Lu, Nakanishi, & Gao, 2017).
Segundo dados obtidos, a adição de uma única GlcNAc influencia em 65% da
estabilidade e 100% da cinética de enovelamento da proteína hCD2ad (Hanson et al.,
2009). Gusakov et al. mostrou que o processo de N-glicosilação é um componente
importante da atividade processiva de celobiohidrolases (Gusakov, Dotsenko,

41
Rozhkova, & Sinitsyn, 2017). Qi et al. descreveram a função de glicanas N-ligadas no
domínio catalítico da celobiohidrolase I de Trichoderma reesei, cujos locais de
remoção de N-glicosilação dificilmente afetariam secreção, estabilidade térmica e
atividade (Qi, Zhang, Zhang, Chen, & Liu, 2014). O mesmo efeito foi relatado em
outras enzimas, tais como celobiohidrolases (Amore et al., 2017; Goedegebuur et al.,
2017; Zoglowek, Lübeck, Ahring, & Lübeck, 2015), β-glicosidase (Wei et al., 2013), e
xilanase (Chang et al., 2017), entre outros.
Outros estudos têm demonstrado o efeito da composição de peptídeos ao redor dos
sítios de glicosilação a fim de compreender o mecanismo de N-glicosilação a nível de
reconhecimento da OST. Aproximadamente 70% dos motivos Asn-X-Ser/Thr são
glicosilados, pois existe a seleção pela OST de acordo com a estrutura do polipeptídio
nascente. Foi verificada a preferência pelos aminoácidos Phe, Gly, Ile, Ser, Tyr e Val
na posição ”X”, enquanto que os aminoácidos carregados e prolina foram encontrados
em níveis baixos quando analisado os sítios N-glicosilados validados
experimentalmente (Rao & Bernd, 2010). Somente os sítios de glicosilação acessíveis
ao lúmen do RE são conhecidos por serem N-glicosilados, no entanto a presença da
N-glicana foi relatada em muitas geometrias de superfície diferentes (Petrescu, Milac,
Petrescu, Dwek, & Wormald, 2004). A posição "X" pode reduzir a eficiência da
glicosilação, quando "X" for ácido (Asp ou Glu), ou aumentar a eficiência quando Phe
estiver em um loop reverso adjacente (Huang et al., 2017; Price et al., 2012, 2011;
Stanley et al., 2017).
Estudos voltados a composição e sequência da proteína alvo permite melhor
manipulação da sequência recombinante, bem como a escolha da melhor cepa
hospedeira. Baseado na não conservação dos sítios de glicosilação em proteínas
homólogas, Tan et al. verificaram quais as melhores mutações pontuais para substituir
uma N-glicosilação (Tan et al., 2014). Asn-Gln é a mutação sítio dirigida mais
empregada para avaliar a importância de determinados sítios de glicosilação no
enovelamento e função de glicoproteínas, porém as preferências conformacionais de
Gln são muito diferentes das de Asn, enquanto His teria conformação mais
semelhante a Asn. Apesar da semelhança conformacional entre His e Asn, essa
mutação pontual gera uma troca de natureza química e polaridade, de básico para

42
polar neutro. Foi demonstrado, também, que a substituição Asn-Asp pode suprir a
ausência da N-glicana recuperando a estabilidade da proteína (Tan et al., 2014).
Um problema importante na secreção de proteínas heterólogas é a falta ou a
inadequada N-glicosilação dentro do RE. A glicosilação é muito importante para o
enovelamento correto das proteínas e serve como informação para degradação e
controle de qualidade (Helenius & Aebi, 2004). Já foi demonstrado que fatores
externos também podem influenciar na composição e distribuição de N-glicosilação
ao longo da mesma proteína (Adav, Ravindran, & Sze, 2014; Goochee & Monica,
1990). Dessa forma, pode-se verificar o elevado nível de complexidade na produção
de proteínas recombinantes. Porém, com os avanços desses conhecimentos em
fungos filamentosos, incluindo espécies do gênero Aspergillus, será possível a
consolidação e estabelecimento de uma plataforma de alta performance para
produção de proteínas recombinantes. Pode-se assim, trazer genes heterólogos
otimizados tanto para a sequência gênica, quanto para a obtenção adequada de
MPTs.

43
Capítulo 2. Glicoproteômica e Glicômica de Aspergillus nidulans
Introdução
Neste capítulo estão detalhadamente descritos as metodologias e os resultados
obtidos e na N-glicoproteômica e glicômica de A. nidulans cultivado em glicose, xilano
e bagaço de cana-de-açúcar pré-tratado. Os experimentos realizados tiveram como
objetivo identificar a maior quantidade possível de sítios de N-glicosilação em
CAZymes. Por meio da N-glicoproteômica foi possível identificar 265 proteínas e 182
sítios de N-glicosilação. Identificou-se, também, a preferência de A. nidulans por sítios
de glicosilação compostos por N-X-T, os quais foram predominantemente encontrados
em regiões de resíduos de aminoácidos hidrofóbicos ou polares não carregados. As
mesmas proteínas secretadas foram submetidos a análise do perfil de N-glicanas
geral por meio da glicômica realizada por MALDI-TOF. As N-glicanas contendo cinco
hexoses foram prevalentes nos cultivos em glicose e bagaço de cana-de-açúcar,
enquanto, o cultivo de xilano apresentou quantidades semelhantes de 5 a 9 hexoses.
Dessa forma, esse capítulo descreve os esforços realizados para validar diferentes
sítios de N-glicosilação e composições de N-glicanas para compor o pilar inicial dos
estudos com N-glicoproteínas em nosso grupo de pesquisa. Os resultados desse
capítulo auxiliarão no estudo de produção de proteínas recombinantes em fungos
filamentosos. A determinação de uma preferência e a validação de sítios de N-
glicosilação em CAZymes, bem como a composição das N-glicanas favorecem o
engenheiramento de proteínas e otimização da produção de proteínas recombinantes.
Diversos problemas podem atrapalhar os processos de tradução, enovelamento,
transporte intracelular e secreção, no entanto a N-glicosilação correta é essencial para
esses processos.
O presente capitulo compõe o primeiro artigo desse trabalho de doutorado, o qual foi
publicado na revista Biotechnology for Biofuels em 2016 (DOI. 10.1186/s13068-016-
0580-4).

44
Mapping N-Linked Glycosylation of Carbohydrate-Active Enzymes in the
Secretome of Aspergillus nidulans Grown on Lignocellulose
Marcelo Ventura Rubio1,2, Mariane Palludeti Zubieta1,2, João Paulo Lourenço Franco
Cairo1,2, Felipe Calzado1,2, Adriana Franco Paes Leme3, Fabio Marcio Squina¹, Rolf
Alexander Prade4, André Ricardo de Lima Damásio1,2*
¹Laboratório Nacional de Ciência e Tecnologia do Bioetanol (CTBE), Centro Nacional
de Pesquisa em Energia e Materiais (CNPEM), Campinas, SP, Brasil.
²Institute of Biology, University of Campinas (UNICAMP), Campinas-SP, Brazil
3Laboratório Nacional de Biociências (LNBio), Centro Nacional de Pesquisa em
Energia e Materiais (CNPEM), Campinas, SP, Brasil.
4Department of Microbiology and Molecular Genetics, Oklahoma State University,
Stillwater-OK, USA.
Running title: Glycoproteomics of Aspergillus nidulans
*To whom correspondence should be addressed:
André R. L. Damásio; Department of Biochemistry and Tissue Biology, Institute of
Biology, University of Campinas (UNICAMP). Rua Monteiro Lobato, 255, Cidade
Universitária Zeferino Vaz, 13083-862 - Campinas-SP, Brazil; E-mail:
[email protected]; phone number: +55 19 3521-1437

45
Abstract
Background: The genus Aspergillus includes microorganisms that naturally degrade
lignocellulosic biomass, secreting large amounts of carbohydrate-active enzymes
(CAZymes) that characterize their saprophyte lifestyle. Aspergillus has the capacity to
perform post-translational modifications (PTM), which provides an additional
advantage for the use of these organisms as a host for the production of heterologous
proteins. In this study, the N-linked glycosylation of CAZymes identified in the
secretome of Aspergillus nidulans grown on lignocellulose was mapped.
Results: A. nidulans was grown in glucose, xylan and sugarcane bagasse for 96 h,
after which glycoproteomics and glycomics were carried out on the extracellular
proteins (secretome). A total of 265 proteins were identified, with 153, 210 and 182
proteins in the glucose, xylan and sugarcane bagasse (SCB) substrates, respectively.
CAZymes corresponded to more than 50% of the total secretome in xylan and SCB. A
total of 182 N-glycosylation sites were identified, of which 121 were detected in 67
CAZymes. A prevalence of the N-glyc sequon N-X-T (72.2%) was observed in N-glyc
sites compared with N-X-S (27.8%). The amino acids flanking the validated N-glyc
sites were mainly composed of hydrophobic and polar uncharged amino acids.
Selected proteins were evaluated for conservation of the N-glyc sites in Aspergilli
homologous proteins, but a pattern of conservation was not observed. A global
analysis of N-glycans released from the proteins secreted by A. nidulans was also
performed. While the proportion of N-glycans with Hex5 to Hex9 was similar in the xylan
condition, a prevalence of Hex5 was observed in the SCB and glucose conditions.
Conclusions: The most common and frequent N-glycosylated motifs, an overview of
the N-glycosylation of the CAZymes and the number of mannoses found in N-glycans
were analyzed. There are many bottlenecks in protein production by filamentous fungi,
such as folding, transport by vesicles and secretion, but N-glycosylation in the correct
context is a fundamental event for defining the high levels of secretion of target
proteins. A comprehensive analysis of the protein glycosylation processes in A.
nidulans will assist with a better understanding of glycoprotein structures, profiles,
activities and functions. This knowledge can help in the optimization of heterologous
expression and protein secretion in the fungal host.

46
Keywords: glycoproteomics, Aspergillus nidulans, carbohydrate-active enzymes,
CAZy, glycoside hydrolases, N-glycosylation, heterologous expression
Background
The dependence on energy sources derived from fossil fuels and the environmental
impact caused by their use have generated special interest from researchers and
governments regarding the use of renewable energy sources. The use of renewable
sources for fuel production has become an important alternative because they
generate fewer pollutants and allow the sustainable development of the economy and
human society. Alternatively, the use of lignocellulosic biomass, mainly composed of
cellulose, hemicellulose and lignin, is a consensus worldwide because it is the most
abundant renewable energy source on Earth (Jouzani & Taherzadeh, 2015). However,
the use of this biomass in the biorefinery concept requires its depolymerization to
mono- and oligosaccharides, which are the building blocks used to produce biofuels
and biochemicals.
Plant biomass is a complex structure rich in glycoconjugates and poly- and oligo
saccharides, and a wide variety of enzymes are necessary for the complete
degradation of this biomass (Glass et al., 2013; Segato et al., 2014). Carbohydrate-
Active Enzymes (CAZymes) participate in the breakdown, biosynthesis and
modification of the glycoconjugates and oligo- and polysaccharides that constitute the
plant cell wall. In general, CAZymes are structurally constituted by a catalytic domain,
and some CAZy families have an additional carbohydrate-binding module (CBM).
Based on structural and homology features, the CAZy database currently covers five
enzyme classes, including glycoside hydrolases (GHs), glycosyltransferases (GTs),
polysaccharide lyases (PLs), carbohydrate esterases (CEs) and auxiliary activities
(AAs) (Lombard et al., 2014).
The genus Aspergillus includes microorganisms that naturally degrade lignocellulosic
biomass and secrete large amounts of CAZymes, which characterize their saprophyte
lifestyle (Bauer, Vasu, Persson, Mort, & Somerville, 2006). This complex biomass is
partially degraded, releasing simple carbohydrates that are readily taken up by the
fungal cells to provide energy for their growth and reproduction. Due to this capacity
for secretion of a large amount and variety of enzymes, along with the abilities to

47
tolerate extreme cultivation conditions in liquid and solid state fermentation, the
Aspergillus fungus has been a successful model for enzyme production on an industrial
scale (Meyer, Wu, & Ram, 2011).
Aspergillus has the capacity to perform post-translational modifications (PTM) such as
proteolytic cleavage, disulfide bond formation and glycosylation of proteins, providing
an additional advantage for the use of these organisms as a host for the production of
heterologous proteins (K. M. H. Nevalainen et al., 2005). Asparagine-linked protein N-
glycosylation is a prevalent PTM in eukaryotic systems, and has also been described
in prokaryotic systems (Weerapana & Imperiali, 2006). The N-glycosylation consists of
the co- or post-translational attachment of an oligosaccharide to proteins by covalent
bonds in the endoplasmic reticulum (ER) lumen (Helenius & Aebi, 2004; Spiro, 2002).
N-glycosylation of proteins is essential for a range of cellular processes such as
immune responses, cellular communication, intracellular trafficking, stability, secretion,
folding and protein activity (Helenius & Aebi, 2004; Jarrell et al., 2014; Skropeta, 2009;
Solá, Rodríguez-Martínez, & Griebenow, 2007). In eukaryotes, N-linked glycosylation
occurs at the Asn-X(aa)-Ser/Thr sequon and is a co-translational process catalyzed by
oligosaccharyltransferases (OST) in the lumen of the ER (Weerapana & Imperiali,
2006).
Glycoscience, which involves N-glycosylation studies that have been performed
primarily for an understanding of the role of carbohydrates on biophysical modifications
in cell communication, is aimed at developing new approaches for the treatment of
human diseases (Weixuan Chen et al., 2014; Eshghi et al., 2014; Moremen et al.,
2012). However, some recent studies have shown the effect of glycosylation on folding,
secretion and enzymatic properties (Banerjee et al., 2007). Knowledge of N-
glycosylation of CAZymes is scarce and mainly reported for cellobiohydrolases
(Beckham et al., 2012a; Ruchi Gupta et al., 2011; Jeoh et al., 2008). The correct
glycosylation of proteins becomes an essential feature in systems for the heterologous
expression of target genes using filamentous fungi as a host because the accumulation
of unfolded or misfolded proteins is a bottleneck in the secretion pathway and also in
the protein production yield (H. Nevalainen & Peterson, 2014; Skropeta, 2009).

48
Accumulation of misfolded proteins overloads the ER processing capacity, triggering a
response called the unfolded protein response (UPR). The UPR pathway activates a
large set of genes responsible for correct protein folding, degradation of misfolded
proteins and others to recover proteostasis (C. Rubio et al., 2011; Walter & Ron, 2011).
Thus, larger amounts of proteins acquire the correct folding, can leave the ER bound
for the extracellular environment and are not targeted for degradation. The decrease
in glycosylation levels by reducing the expression of oligosaccharyltransferase genes
leads to cell stress conditions. ER stress induced by the low levels of glycosylation of
some proteins leads to the overexpression of several UPR genes, including genes
related to cell wall biogenesis, protein folding and degradation of unfolded proteins (K.
Li et al., 2011).
There are a few studies mapping the global N-glycosylation of CAZymes in filamentous
fungi (Adav et al., 2014; Lu Wang et al., 2012). In this study, the N-linked glycosylation
of CAZymes identified in the secretome of Aspergillus nidulans grown on lignocellulose
was mapped. Therefore, A. nidulans was grown in glucose, xylan and pretreated
sugarcane bagasse (SCB), followed by glycoproteomics and glycomics on the
extracellular proteins (secretome). The most common and frequent N-glycosylated
motifs, an overview of CAZymes’ N-glycosylation and the number of mannose residues
found in N-glycans were analyzed. A comprehensive analysis of protein glycosylation
processes in A. nidulans will assist with a better understanding of glycoprotein
structures, profiles, activities and functions. This knowledge can help in the
optimization of heterologous expression and protein secretion in the fungal host.
Results
Prediction of N-glycosylated CAZymes in the Aspergillus nidulans genome
In order to identify all the putative A. nidulans glycoproteins involved in lignocellulose
degradation, a comprehensive analysis of the A. nidulans ORFs (10678 entries)
downloaded from the Aspergillus Genome Database (AspGD) was performed
(Cerqueira et al., 2014). First, 428 CAZymes (4% of the A. nidulans ORFs) were
annotated by dbCAN (automated CAZymes annotation) (Yin et al., 2012). Second, 359
out of 428 CAZymes were predicted to contain at least one N-glycosylation site (N-glyc
site) by the NetNGlyc 1.0 Server. Finally, the predicted N-glycosylated CAZymes were

49
analyzed for the presence of signal peptide cleavage sites using the SignalP 4.1 Server
(Figure S1).
The majority (73%) of the 190 N-glycosylated CAZymes identified in the secretomes
were classified as glycoside hydrolases (GHs), and 7% had a C-terminal-associated
carbohydrate-binding module (CBM). The other CAZymes were predicted as auxiliary
activities (AAs; 9%), carbohydrate esterases (CEs; 7%), polysaccharide lyases (PLs;
7%) and glycosyltransferases (GTs; 4%). The number of predicted N-glyc sites in the
190 secreted CAZymes varied from 1 to 21 sites and approximately 40% of proteins
had one or two N-glyc sites (Figure S2).
Proteomics overview of A. nidulans grown on glucose, xylan and alkali
pretreated sugarcane bagasse (SCB)
A. nidulans was grown in three different substrates for 96 hours (glucose, xylan and
SCB in biological triplicates), and the secretome of each condition was evaluated by
SDS-PAGE stained with Coomassie blue for the total protein profile and with Pro-Q
Emerald for glycoprotein detection (Figure 1A).
To identify the glycoproteins occurring in each cultivation condition, the secretomes
were first enriched by ConA and then analyzed by LC-MS/MS (see the Methods
section). A total of 265 proteins was identified using one unique peptide and 0.1% of
FDR (False Discovery Rate). From the glucose, xylan and SCB conditions, 153, 210
and 182 total proteins were identified, respectively, with 99 proteins common to all
three conditions (Figure 1B). Regarding the CAZymes, 59, 111 and 107 proteins were
identified in the glucose, xylan and SCB conditions, respectively, and CAZymes
corresponded to more than 50% of the total secretome in the xylan and SCB conditions
(Table S1). Glycoside hydrolase was the most abundant class at 84%, followed by
AAs at 8% and CEs at 7% (Figure 1C). In the glucose condition, 61.5% of the proteins
were assigned as non-CAZymes. In addition, a high variation in the proportion of
CAZyme classes and families throughout the three substrates was observed, with A.
nidulans secreting the highest diversity of CAZymes in the SCB condition. A further
examination of the proteins that were exclusively identified in the SCB condition
showed a high abundance of GHs linked to CBMs. These results are directly

50
associated with the greater complexity of sugarcane bagasse and thus the requirement
for a higher range of enzymes to degrade it (Figure 1D).
In addition to the variations in the proportions of the CAZyme classes among the
substrates (Figure 2A), different compositions were also observed at the family level
(Figure 2B-E). Regarding the CEs, family CE16, known as carbohydrate
acetylesterases active on various carbohydrate acetyl esters, was the most abundant
in the SCB condition, representing 58%, followed by CE1 (feruloyl esterases) and CE2
(acetyl xylan esterases). In the xylan condition, family CE10 was the most
representative, although the members of this family are esterases that act on non-
carbohydrate substrates (Levasseur et al., 2013). Thus, the most abundant
carbohydrate esterase family was CE1 (28%), a classical family for feruloyl esterases,
followed by CE4 (acetyl xylan esterases) and CE16 (acetyl esterases) (Figure 2B).
The AA8 (flavocytochromo - cellobiose dehydrogenases) and AA9 (former copper-
dependent lytic polysaccharide monooxygenases - LPMOs) families were the most
abundant oxidative enzymes in the SCB condition, at 48.5% and 23.7% of the total
AAs, respectively. Moreover, AA8 was exclusively reported in the SCB condition. Both
enzyme families are highly correlated with the oxidative degradation of cellulose in
fungi. However, in the xylan condition, the most representative families were AA7
(oligosaccharide oxidases), AA1 (laccases) and AA9 (LPMOs) (Figure 2C). Among
the PLs, family PL1 (pectin lyase) was the most abundant in the SCB condition (68%),
followed by PL4 (rhamnogalacturonan lyase), whereas PL4 was the most
representative family in the xylan condition at 76%, followed by PL3 (pectate lyase)
(Figure 2D). The analysis of the spectrum counts showed that GH7, a classical family
of the cellobiohydrolases/exoglucanases, was the most abundant GH family in the
SCB condition, which represented 35% of the GHs, followed by GH62 (α-L-
arabinofuranosidase) and GH3 (beta-gluco/xylosidase). Families such as GH5 (endo-
glucanases) and GH6 (exo-glucanasse) were exclusively identified in the SCB
condition (Figure 2E). GH3 was the most abundant family in the xylan condition,
accounting for 16% of the total spectrum counts of GHs, followed by GH20 (beta-
hexosaminidase), GH18 (chitinase) and GH43 (xylanase and alpha-L-
arabinofuranosidase) (Figure 2E).

51
Figure 1. Overview of secretomes from A. nidulans grown on glucose, xylan and
sugarcane bagasse. (A) A. nidulans secretomes were stained with coomassie brilliant
blue and Pro-Q Emerald for detection of glycoproteins in polyacrylamide gels. L:
ladder; Glu: glucose; SCB: sugarcane bagasse. The secretomes produced on xylan
and SCB are represented in duplicate, however the experiments were performed in
triplicate. (B) Mascot searches were carried out using the Aspergillus Genome
Database (AspGD). The data were analyzed by the Scaffold software and the Venn
diagram represent the number of proteins identified in each secretome. (C) Abundance
of CAZymes classes identified in each condition. (D) CAZymes diversity shared among
the growth conditions. The intersection symbol “∩” means that proteins are common
in two or more conditions. GH: glycoside hydrolases; PL: polysaccharide lyases; CE:
carbohydrate esterases; AA: auxiliary activities; CBM: carbohydrate binding module.

52
Figure 2. CAZymes annotation in the secretomes of A. nidulans grown on xylan
and sugarcane bagasse. CAZymes in the secretomes were annotated by an HMM-
based database (dbCAN). (A) Proteins were grouped in CAZy and Non-CAZy, and the
CAZymes were grouped according to enzyme classes in carbohydrate esterases-CEs
(B), auxiliary activities-AAs (C), polysaccharide lyases-PLs (D) and glycoside
hydrolases-GHs (E). The total number of proteins in each class of enzyme was set as
100% and families representing less than 2% of the total proteins were not shown.
SCB: alkali pretreated sugarcane bagasse.

53
At the individual protein level, a GH7 cellobiohydrolase (ANID_05176) was the most
abundant protein identified in this proteomics study, accounting for 1098 peptides in
the SCB condition (Figure 3). The GH3 (ANID_02828) was the most secreted beta-
glucosidase and showed the same spectrum counts in the xylan and SCB conditions
(Figure 3).
In addition, GH47 alpha-mannosidase (ANID_00787) and GH62 alpha-L-
arabinofuranosidase were also represented, both with more spectrum counts in the
SCB condition than in the xylan or glucose conditions. One GH11 (ANID_03613) and
two GH10 xylanases (ANID_01818; ANID_07401) were also more abundant in the
SCB condition than in the xylan condition.
Figure 3. Top ten proteins secreted by A. nidulans. The total spectrum counts of a
specific protein were summed and grouped in according to their functions. (A)
Cellulases; (B) Hemicellulases; (C) Carbohydrate Esterases; (D) Auxiliary
Activities/Oxidases; (E) Polysaccharides Lyases; (F) CBMs and (G) Others functions.

54
CE16 acetyl esterase (ANID_06422) was the most abundant CE found in the SCB
condition, whereas A. nidulans secreted primarily two CE10 (ANID_01320;
ANID_01433) in the xylan condition. Considering all conditions, an AA8 cellobiose
dehydrogenase (ANID_07230) was the most abundant AA but was only detected in
the SCB condition. The AA7 gluco-oligosaccharide oxidase (ANID_07269) and AA9
LPMO (ANID_02388) were the most “regular” enzymes, with equal secretion levels in
all the conditions. However, the AA9 LPMO (ANID_06428) had peptides only reported
in the SCB condition.
All the pectinases and polysaccharide lyases were more secreted in the xylan condition
by far than in the SCB or glucose conditions. The GH105 rhamnogalactoronyl
hydrolase (ANID_09383) was the most abundant pectinase, followed by the GH28 exo-
polygalacturonase (ANID_08761), both showing high levels of secretion in the xylan
condition. The PL4 rhamnogalaturonan lyase PL4 (ANID_12097) was the most
abundant enzyme among the PLs.
Among the enzymes classified as others, a serine protease (ANID_02954) was the
most abundant enzyme in SCB, followed by a hypothetical protein (ANID_06535) and
a chitinase (ANID_04871). In the xylan condition, the most representative enzymes
were N-acetylglucosaminidase (ANID_01502) followed by catalase (ANID_09339) and
chitinase (ANID_04871).
The enzymatic activities in the A. nidulans secretomes were also analyzed (Figure
S3). The highest activity was detected on beta-glucan and xylan using the SCB
secretome. Using the xylan secretome, the highest activity was reported on xylan from
beechwood, followed by beta-glucan and mannan. Non-significant activities were
found using the glucose-condition secretome. All these enzymatic activities were in
accord with the proteome profile found for each growth condition as described above.
N-glycosylated sites detected on CAZymes
As previously mentioned, 265 proteins were detected by LC-MS/MS after enrichment
by ConA, and at least one N-glyc site was validated in 103 proteins. Considering all
the N-glyc sites predicted by the NetNGlyc server, we defined three groups of sites in
this work: 1) validated sites: N-glyc sites confirmed by our LC-MS/MS data set using
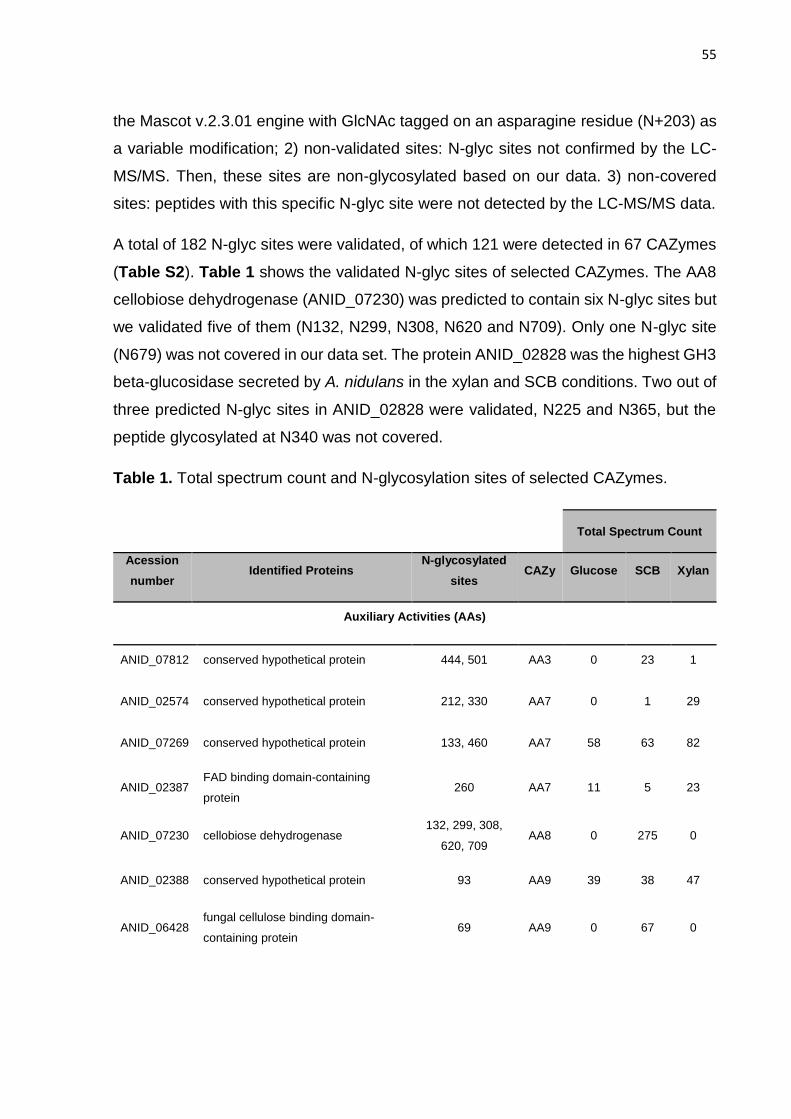
55
the Mascot v.2.3.01 engine with GlcNAc tagged on an asparagine residue (N+203) as
a variable modification; 2) non-validated sites: N-glyc sites not confirmed by the LC-
MS/MS. Then, these sites are non-glycosylated based on our data. 3) non-covered
sites: peptides with this specific N-glyc site were not detected by the LC-MS/MS data.
A total of 182 N-glyc sites were validated, of which 121 were detected in 67 CAZymes
(Table S2). Table 1 shows the validated N-glyc sites of selected CAZymes. The AA8
cellobiose dehydrogenase (ANID_07230) was predicted to contain six N-glyc sites but
we validated five of them (N132, N299, N308, N620 and N709). Only one N-glyc site
(N679) was not covered in our data set. The protein ANID_02828 was the highest GH3
beta-glucosidase secreted by A. nidulans in the xylan and SCB conditions. Two out of
three predicted N-glyc sites in ANID_02828 were validated, N225 and N365, but the
peptide glycosylated at N340 was not covered.
Table 1. Total spectrum count and N-glycosylation sites of selected CAZymes.
Total Spectrum Count
Acession
number Identified Proteins
N-glycosylated
sites CAZy Glucose SCB Xylan
Auxiliary Activities (AAs)
ANID_07812 conserved hypothetical protein 444, 501 AA3 0 23 1
ANID_02574 conserved hypothetical protein 212, 330 AA7 0 1 29
ANID_07269 conserved hypothetical protein 133, 460 AA7 58 63 82
ANID_02387 FAD binding domain-containing
protein 260 AA7 11 5 23
ANID_07230 cellobiose dehydrogenase 132, 299, 308,
620, 709 AA8 0 275 0
ANID_02388 conserved hypothetical protein 93 AA9 39 38 47
ANID_06428 fungal cellulose binding domain-
containing protein 69 AA9 0 67 0
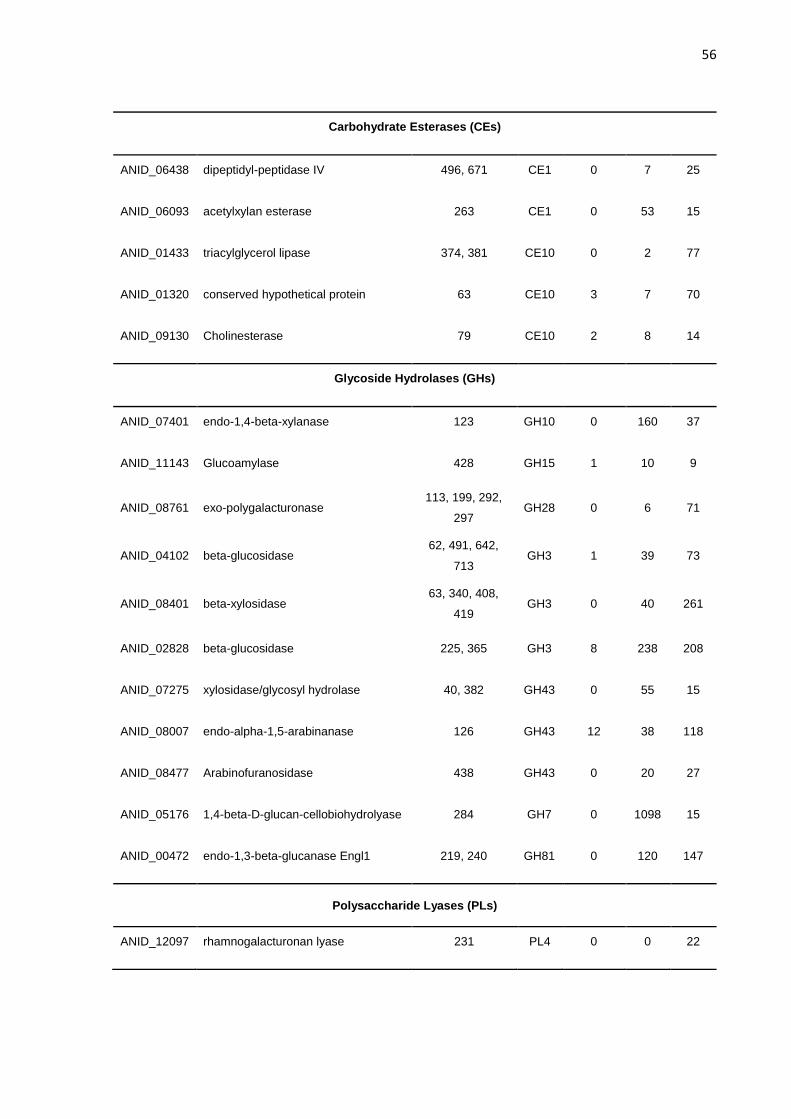
56
Carbohydrate Esterases (CEs)
ANID_06438 dipeptidyl-peptidase IV 496, 671 CE1 0 7 25
ANID_06093 acetylxylan esterase 263 CE1 0 53 15
ANID_01433 triacylglycerol lipase 374, 381 CE10 0 2 77
ANID_01320 conserved hypothetical protein 63 CE10 3 7 70
ANID_09130 Cholinesterase 79 CE10 2 8 14
Glycoside Hydrolases (GHs)
ANID_07401 endo-1,4-beta-xylanase 123 GH10 0 160 37
ANID_11143 Glucoamylase 428 GH15 1 10 9
ANID_08761 exo-polygalacturonase 113, 199, 292,
297 GH28 0 6 71
ANID_04102 beta-glucosidase 62, 491, 642,
713 GH3 1 39 73
ANID_08401 beta-xylosidase 63, 340, 408,
419 GH3 0 40 261
ANID_02828 beta-glucosidase 225, 365 GH3 8 238 208
ANID_07275 xylosidase/glycosyl hydrolase 40, 382 GH43 0 55 15
ANID_08007 endo-alpha-1,5-arabinanase 126 GH43 12 38 118
ANID_08477 Arabinofuranosidase 438 GH43 0 20 27
ANID_05176 1,4-beta-D-glucan-cellobiohydrolyase 284 GH7 0 1098 15
ANID_00472 endo-1,3-beta-glucanase Engl1 219, 240 GH81 0 120 147
Polysaccharide Lyases (PLs)
ANID_12097 rhamnogalacturonan lyase 231 PL4 0 0 22

57
We performed an additional validation of the N-glyc sites by using the Scaffold PTM
software with default statistical parameters pre-established by the program, based on
the presence and intensity of site-specific ions compared randomly (Beausoleil, Villén,
Gerber, Rush, & Gygi, 2006). From the 182 sites previously validated, 151 sites were
re-validated by the additional statistical filters, which increased the sensitivity to peptide
spectra matches (Beausoleil et al., 2006). We further analyzed the 151 re-validated N-
glyc sites to determine if there was a specific amino acid motif surrounding the N-glyc
sites. The amino acid sequence of all validated N-glycopeptides were aligned, and six
amino acid residues before and after the sequons (N-X-S/T) were analyzed. The
prevalence of the sequence N-X-T (72.2%) over N-X-S (27.8%) was observed.
Furthermore, the sequon N-X-T showed additional motif variations (Table 2 and Table
S3). In addition to the 182 N-glyc sites validated, 23 predicted N-glyc sites were not
validated by the LC-MS/MS data set. The sequon N-X-S (60.9%) was predominant for
those non-validated sites as opposed to the validated sites.
Table 2. Motifs report for the flanking sequences dataset of N-glycosylated sites.
Motif Dataset matches1 Dataset percentage2 Background percentage3
......n.S.... 42 27.8 6.60
......n.T.... 109 72.2 7.30
......nGT... 19 12.6 1.50
......nTT.... 12 7.90 0.92
......nST.... 14 9.30 0.69
......n.T.T.. 15 9.90 0.96
...P..n.T.... 15 9.90 0.80
T.....n.T.... 14 9.30 0.92
1The data set consists of the flanking sequences for N-glycosylated sites. 2Dataset percentage was
calculated based on the probability that a modification occurs in a given motif throughout all identified
peptides. 3Background percentage was calculated using all proteins loaded into the program as
background and measuring the probability that a specific amino acid appears with a motif. The dots
correspond to the amino acids flanking glycosylated asparagines (n). S: serine; T: threonine; P;
proline.

58
The amino acids flanking the validated N-glyc sites (from -6 to +6) were classified
according to the chemical properties of the side chains (Figure 4). These flanking
regions were mainly composed by hydrophobic and polar uncharged amino acids.
However, this profile was different for the non-validated N-glyc sites (Figure S4).
Figure 4. Amino acids flanking validated N-glycosylation sites. The relative
occurrence of amino acids is plotted versus sequence position -6 to +6 around an
occupied N-glyc site. Residues specified by the glycosylation sequon (0=Asn; +2=Ser
or Thr) are not plotted. Hydrophobic (Ala, Val, Leu, Ile, Met); Aromatic (Phe, Tyr, Trp);
Polar uncharged (Ser, Thr, Asn, Cys, Gln); Acidic (Asp, Glu); Basic (Lys, Arg, His);
Unique (Gly, Pro).
Conservation of N-glycosylated sites in Aspergilli
To investigate if the majority of N-glyc sites were conserved in homologous proteins,
we aligned selected proteins from A. nidulans with 19 Aspergilli genomes from the
AspGD (Table 3). Proteins were considered homologous when the E-value (Blastp)
was equal to or less than 1.00E-70. Two CAZyme sequences are shown in Figure 5.
The ANID_00472 is a GH81 endo-1,3-beta-glucanase Engl1 that was secreted by A.
nidulans in the xylan and SCB conditions. Four N-glyc sites were predicted for this
protein (N219, N240, N257, N499), two of which were validated by the
glycoproteomics. We found and aligned 20 sequences homologous to this GH81,
generating a sequence logo. The N-glyc sites N219 and N240 were highly conserved

59
throughout all the homologous sequences, 90% and 95%, respectively. Similarly,
ANID_05176 is a GH7 cellobiohydrolase with two predicted N-glyc sites (N284 and
N333), but the N333 was non-glycosylated according to our data. We found and
aligned 50 homologous sequences, and while the N284 site was conserved in 62% of
the homologous sequences, the N333 was present in only one homologous sequence.
The conservation of N-glyc sites ranges from 8 to 100% (Table 3), and therefore there
was no pattern of conservation of N-glyc sites in Aspergilli homologous sequences.
Table 3. Conservation of N-glycosylated sites of selected A. nidulans CAZymes in
homologous proteins.
Identified Proteins Accession
Number Domain
N-glyc
sites1 Conservation2
Homologous
in Aspergilli 3 Sequon4
endo-1,3-beta-glucanase
Engl1 ANID_00472 GH81 219 90,0% 20 NSS
240 95,0% 20 NAT
alpha-glucosidase AgdA ANID_02017 GH31 432 91,3% 22 NAS
beta-xylosidase ANID_02359 GH3 231 43,3% 30 NHS
673 55,9% 34 NFT
695 52,9% 34 NTT
beta-glucosidase ANID_02828 GH3 225 100% 50 NGT
365 34,0% 50 NGS
beta-glucosidase ANID_04102 GH3 62 98,0% 50 NLT
491 8,0% 50 NKT
642 92,0% 50 NQT
713 40,0% 50 NST
1,4-beta-D-glucan-
cellobiohydrolyase ANID_05176 GH7 284 62,0% 50 NTS
cellobiose
dehydrogenase ANID_07230 AA8 132 37,5% 24 NAT
299 47,4% 38 NGT
308 90,0% 40 NGT
620 7,9% 38 NVT
709 23,7% 38 NVS

60
beta-glucosidase ANID_07396 GH3 259 100% 50 NNS
438 72,0% 50 NGT
586 56,0% 50 NSS
endo-1,4-beta-xylanase ANID_07401 GH10 123 100% 11 NTT
beta-xylosidase ANID_08401 GH3 63 70,0% 30 NNT
340 30,0% 30 NET
408 46,7% 30 NGT
419 93,3% 30 NFT
arabinofuranosidase ANID_08477 GH43 438 13,6% 23 NGS
exo-polygalacturonase ANID_08761 GH28 113 62,1% 29 NDT
199 72,4% 29 NSS
292 100% 30 NIS
297 63,3% 30 NAS
exopolygalacturonase ANID_08891 GH28 65 21,9% 32 NDT
230 94,1% 17 NAS
alpha-glucuronidase ANID_09286 GH67 48 85,0% 20 NAT
315 95,0% 20 NRT
689 90,0% 20 NKS
769 15,0% 20 NST
exo-
rhamnogalacturonase B ANID_10274 GH28 34 53,3% 15 NET
340 100% 16 NCT
beta-glucosidase ANID_10482 GH3 73 100% 50 NLT
726 60,0% 50 NSS
¹N-glycosylated sites confirmed in the A. nidulans proteins by LC-MS/MS.
2Alignment gaps on the sequences shift the N-glycosylation sequons in homologous sequences. In spite of the
sequons are in different positions on primary sequence, the N-glycan is attached to similar positions at proteins
3D-level (see Figure 7).
3Proteins were considered homologous when the E-value (Blastp) was equal to or less than 1.00E-70.
4Sequon detected in A. nidulans glycoproteins by LC-MS/MS.

61
Figure 5. Conservation of glycosylation sites in selected proteins identified on
sugarcane bagasse-secretome. N-glyc sites of selected proteins were analyzed by
conservation in homologous proteins on AspGD. (A) AN0472 is a GH81 endo-1,3-beta-
glucanase Engl1 secreted on xylan and SCB. (B) AN5176 is a GH7 cellobiohydrolase
highly secreted in SCB.
Profile of N-glycans attached to proteins secreted by A. nidulans
A global analysis of N-glycans released from proteins secreted by A. nidulans was also
performed. The MALDI spectra of permethylated N-glycans released from each
replicate of the secretomes produced by A. nidulans in the glucose, SCB and xylan
media are shown in Figure 6. Mannose and galactofuranose are structural isomers,
have exactly the same mass and are non-distinguishable in MALDI/TOF-MS
oligosaccharide profiling. Thus, the peaks were labeled as “Hex5HexNAc2” instead of
“Man5GlcNAc2”. The relative proportion of each N-glycan component was slightly
different in the samples (Table S4) (Deshpande et al., 2008; Geysens, Whyteside, &
Archer, 2009; Maras, Van Die, Contreras, & van den Hondel, 1999). While the
proportion of the N-glycans with Hex5 to Hex9 was similar in the xylan condition, a
prevalence of Hex5 was observed in the SCB and glucose conditions. Moreover, traces
of N-glycans with Hex14 to Hex17 were only detected in the glucose condition.

62
Hex5H
exN
Ac2
Hex6H
exN
Ac2
Hex7H
exN
Ac2
Hex8H
exN
Ac2
Hex9H
exN
Ac2
Hex10H
exN
Ac2
Hex11H
exN
Ac2
Hex12H
exN
Ac2
Hex13H
exN
Ac2
Hex14H
exN
Ac2
Hex15H
exN
Ac2
Hex16H
exN
Ac2
Hex17H
exN
Ac2
0
2
4
6
8
1 0
2 0
4 0
6 0
8 0
1 0 0
G lu c o s e
S C B
X y la nR
ela
tiv
e p
ro
po
rti
on
(%
)
Figure 6. N-glycans profiling of glycoproteins secreted by A. nidulans cultured
in glucose, sugarcane bagasse and xylan. About 500 µg of total secretome was
incubated with PNGase F at 37°C overnight to release N-glycans. N-glycans were
permethylated and profiled by MALDI/TOF-MS.
Discussion
The enzymatic repertoire secreted by A. nidulans matches well to the
composition of the substrate
The N-glycoproteomic analysis of A. nidulans revealed that the abundance and the
repertoire of CAZymes for each grown condition was directly linked to substrate
composition. Our data suggested that sugar monomers and oligomers from xylan and
SCB induced the secretion of a complete repertoire of enzymes by A. nidulans. Delmas
et al. (2012) reported the transcriptional response of A. niger to complex substrates
(Delmas et al., 2012). In general, when A. niger is under starvation (no carbon source)

63
the transcription factor CreA, which act as glycoside hydrolases transcription
repressor, is derepressed allowing a basal expression of a set of GHs. After the initial
uptake of mono and oligosaccharides by the fungus, the transcription factor XlnR is
activated inducing the transcription of several CAZymes (Delmas et al., 2012). Souza
et al. (2011) also reported a complex transcriptional response of A. niger grown on
sugarcane bagasse (de Souza et al., 2011). Moreover, the transcriptional response of
ascomycetes to complex substrates, involves others activators such as AmyR, InuR,
AraR, GalR, GalX and RhaR (Daly, Munster, Raulo, & Archer, 2015).
In the xylan condition, the majority of proteins and peptides identified in the secretome
were correlated with xylan and xylooligosaccharides degradation, as previously
reported for A. fumigatus (Adav, Ravindran, & Sze, 2015). GH3 was the main family
identified in this condition, which is consistent with the secretome of Penicillium
purpurogenum grown on acetylated xylan (Navarrete, Callegari, & Eyzaguirre, 2012).
The identified GH3 enzymes were annotated as beta-xylosidases and beta-1,3/1,4-
glucosidases (Table S1), suggesting xylose production and uptake by A. nidulans. In
a secretome of A. fumigatus grown on xylan, the major families identified were GH10
and GH11, followed by GH3 (Adav et al., 2015).
Moreover, there was a higher abundance of peptides from families GH20 and GH18 in
the xylan condition, and both families are related to cell wall degradation/remodeling,
as well as to protein synthesis/degradation enzymes such as glutaminases,
tyrosinases and proteases (P. W. J. de Groot et al., 2009; Harris, 2008). In addition, a
high abundance of catalase was found in the xylan condition, an enzyme related to
fungal growth and hyphae development (Kawasaki et al., 1997). A set of proteins
related to fungi growth and development was also found in the SCB condition, which
was consistent with the secretome of A. nidulans grown on sorghum stover
(Saykhedkar et al., 2012). Adav et al. (2015) also reported a high abundance of
esterases in the secretome of A. fumigatus grown on xylan (Adav et al., 2015).
GH7, primarily composed of exo-processive cellobiohydrolases, has been described
as the major enzyme secreted by fungi to degrade cellulose and complex
lignocellulose (Glass et al., 2013; Segato et al., 2014). GH7 proteins were the most
abundant enzymes in the secretome of A. nidulans grown on sorghum stover

64
(Saykhedkar et al., 2012) and in the secretome of Trichoderma reesei grown on
sugarcane culms and bagasse, which, along with GH6, accounted for 80% of the
peptide counts (Borin et al., 2015). Ribeiro et al. (2012) reported that the GH7 family,
along with GH5 and GH6, represented the most important set of enzymes secreted by
Penicillium echinulatum grown on integral and pretreated sugarcane bagasse, as well
as on pure cellulose (Ribeiro et al., 2012).
The highest abundance of hemicellulase peptides was identified in the SCB condition
compared with the xylan condition (Table S1), such as GH3, GH62, GH10 and GH11.
This result was further validated by enzymatic activity assays with A. nidulans
secretomes, which reported higher hemicellulase activity in the SCB-derived
secretome than in the xylan secretome (Figure S3). The known composition of SCB
is 60% cellulose, 23% hemicelluloses, 8% lignin and 10% ashes, which suggest that
A. nidulans requires the secretion of different hemicellulases aimed at detaching the
xylan from the cellulose. Thus, the recalcitrant cellulose from SCB could be accessed
and degraded by a set of cellulases and oxidative enzymes.
LPMOs from families AA9 and AA10 have been reported as the major enzymes for
boosting lignocellulose breakdown in commercial cellulase cocktails (Phillips, Beeson,
Cate, & Marletta, 2011; Vaaje-kolstad et al., 2010); however, little is known regarding
their biological role in fungal and bacterial physiology (Gardner et al., 2014; Horn,
Vaaje-Kolstad, Westereng, & Eijsink, 2012). These enzymes require an electron donor
to oxidize lignocelluloses, which can be donated through a non-enzymatic donor, such
as lignin or a reducing agent, or using enzymes such as cellobiose dehydrogenases
(CDH) and oligosaccharide oxidases (GOOX) (Gardner et al., 2014; Horn et al., 2012).
Our results showed that the AA8 family (CDH) members were secreted only in the SCB
condition, whereas AA3 (CDH and GOOX) was reported in all conditions. These results
suggest that these enzymes were differentially regulated in response to substrate in A.
nidulans (Jagadeeswaran, Gainey, Prade, & Mort, 2016). AA7 (GOOX) enzymes were
the most abundant AA in the xylan condition. This family has been reported to be
glucooligosaccharide oxidases (GOO) capable of oxidizing the reducing end of
glycosyl residues of oligosaccharides (Vuong et al., 2013). Members of family AA7,
along with AA3 (GOOX), have been described as enzymes that generate hydrogen

65
peroxide as a co-product of the reactions they are involved in, which can act as a co-
factor for AA1 laccases, such as ANID_06635, another enzyme described in high
abundance in the xylan condition (van Hellemond, Leferink, Heuts, Fraaije, & van
Berkel, 2006). However, catalase B (ANID_09339) was also described in high
abundance in the xylan and SCB conditions. This type of enzyme is well known to
decompose hydrogen peroxide in biological systems. Thus, we suggest that there was
a fine control of this reactive oxygen species in the secretome, as H2O2 could be used
as a co-factor for laccases or a substrate to catalases.
Regarding the LPMOs, only one predicted AA10 enzyme (Pfam LPMO_10) was found
in the xylan and glucose conditions, suggesting AA3 and AA7 as electron donors. The
results also showed that the AA9 enzymes were found in all conditions but were most
abundant in the SCB condition, in which AA8 (CDH) could act as electron donor.
Interestingly, one AA9 (ANID_02388) was reported in all three conditions; however,
this specific enzyme was not reported in the time course secretome of A. nidulans
grown on sorghum stover for 14 days (Saykhedkar et al., 2012). The other four AA9
were only found in the SCB condition in high abundance and to a lower extent in the
glucose condition; however, they were not found in the xylan condition. These data
suggest that AA9 and AA10 enzymes were secreted according to the substrate
composition, as some AA9 enzymes were specific for certain growth conditions,
suggesting again a different regulation in the production of these enzymes, as well as
for AA3 and AA8. Although our data showed that partners LPMO/CDH-GOOX always
occurred together, we could not discard the role of lignin as electron donor for the
LPMOs (Dimarogona, Topakas, Olsson, & Christakopoulos, 2012; Rodriguez-Zuniga
et al., 2015). Aspergillus species employ significantly different approaches to degrade
plant biomass, despite their similar genomic potential. Benoit et al. (2015) showed that
the significant differences between the enzyme sets produced on wheat bran and
sugar beet pulp largely correlated with their polysaccharide composition. The data
suggest the conservation of beta-glucosidase, cellobiohydrolase, beta-galactosidase,
beta-xylosidase and alpha-arabinofuranosidase among eight species of Aspergillus,
highlighting the importance of this group of enzymes to the degradation mechanism in
this genus (Benoit et al., 2015).

66
N-glycosylation occurs preferentially at the N-X-T sequon
The sequons present in proteins are strictly targeted for glycosylation, as the majority
have an N-glycan attached (Skropeta, 2009). Thus, we asked if there were patterns or
preferences for N-glycosylation in the A. nidulans proteins. Mapping N-glyc sites is not
a trivial process and can be performed by glycoprotein- or glycopeptide-level
enrichment methods (Geyer & Geyer, 2006; Patel et al., 1993; Lu Wang et al., 2012).
In this study, the protein-level enrichment method allowed for the identification of 182
N-glyc sites. Nineteen (10.4%) out of 182 sites were previously predicted as non-
glycosylated by the NetNGlyc 1.0 Server (assuming a score <0.5). The NetNGlyc tool
was designed to discriminate what sequon will accept the N-glycan in human proteins,
validating 86% of glycosylated and 61% of non-glycosylated sites in all human proteins
tested (Ramneek Gupta & Brunak, 2002). Moreover, 23 N-glyc sites predicted by
NetNGlyc were not validated by our data set. This result suggests that there is some
inaccuracy in the prediction of N-glyc sites in fungal enzymes by NetNGlyc. Despite
this small divergence, our data show that the NetNGlyc server was a great tool for
predicting glycosylated sequons in A. nidulans proteins.
The 151 N-glyc sites with an acceptable “Ascore” (Beausoleil et al., 2006) clearly
showed the predominance of the consensus sequence N-X-T, representing 72.2% of
glycosylated sequons (Table 2), similarly to that described by Petrescu et al. (Petrescu
et al., 2004). Furthermore, there were variations of the NXT motif such as NGT
(12.6%), NST (9.3%) and NTT (7.9%). In 2010, Rao and Bernd elegantly asked if N-
glycoproteins have a preference for specific sequons. To answer this question, these
authors analyzed viral, archaeal and eukaryotic sequons with experimentally validated
N-glyc sites and detected a preference for some amino acids such as F, G, I, S, T and
V in the sequon “X” position, whereas the charged amino acids and proline were found
to be represented at a lower level (Rao & Bernd, 2010).
Hydrophobic and polar uncharged amino acids are predominant around N-
glycosylated sites
In nature, not all protein sequons (NXT/S) are glycosylated. In A. nidulans,
approximately 50% of the amino acids adjacent to N-glycosylated sites were

67
hydrophobic or polar uncharged (Figure 4). These amino acids are responsible for
producing a microenvironment able to receive the carbohydrate from the
oligosaccharyl transferase (OST). The N-glycan attached to asparagine affects the
local charge, exposing the motif region, and its interaction with the amino acid residues
is responsible for decreasing the enzyme’s dynamics and increasing the
thermostability, increasing the stability against proteolysis (Shental-Bechor & Levy,
2008; Tan et al., 2014).
The presence of hydrophilic N-glycans on the surface of hydrophobic proteins affects
primarily the thermostability, dynamics, solubility and secretion (Cheng, Edwards,
Jiang, & Gräter, 2010; Schwarz & Aebi, 2011; Shental-Bechor & Levy, 2008). Sagt et
al. (2000) showed the effect of insertion of N-glyc sites into hydrophobic proteins. The
addition of a consensus sequence in the N- or C-terminal region decreased the protein
aggregation in the ER and enhanced the secretion by 5- and 1.8-fold, respectively (C.
M J Sagt et al., 2000). Hence, the N-glycosylation in hydrophobic regions could be
related to an evolutionary process involving protein folding, stability and secretion (Lu,
Yang, & Liu, 2012).
Analyzing glycosylated sequons from proteins in the Protein Data Bank (PDB),
Petrescu and collaborators showed the presence of N-glycans attached on different
surface geometries (Petrescu et al., 2004). The surface diversity suggests that the N-
glycosylation process was carefully selected to occur depending on glycan
accessibility. Moreover, a predominance of hydrophobic followed by non-polar amino
acid residues was detected (Rao & Bernd, 2010), corroborating our data, despite the
fact that we did not clearly detect a higher frequency of aromatic amino acids before
the N-glyc site. The sequon neighborhood has also been studied to improve the
stability of target enzymes through biotechnological approaches, for example, by the
addition of an aromatic amino acid before the N-glycosylated sequon to increase
glycoprotein stability (Price et al., 2012, 2011).
The amino acids flanking the non-validated N-glyc sites (Figure S4) were interestingly
different from the validated ones.

68
The N-glycosylation sites are not completely conserved in homologous
sequences
We asked if the N-glyc sites validated in our data were conserved among Aspergilli
homologous proteins. This is a central question driving the prediction of N-
glycosylation patterns in heterologous proteins to increase heterologous secretion by
Aspergilli hosts. The analysis of selected N-glyc sites showed variable profiles of
conservation at the primary sequence level (Table 3). However, this variation can be
a consequence of the alignment, which creates some gaps in the sequences, shifting
the sequons among the homologous sequences (Thompson, Higgins, & Gibson,
1994). For example, the N-glyc sites N299 and N308 were validated by the LC-MS/MS
assay in the AA8 cellobiose dehydrogenase (AN7230), as shown in Table 3. AN7230
showed some variations at the N-glycosylated positions at the primary sequence level
in a homologous alignment, but at the tridimensional structural level, the glycosylation
position was quite similar and occurred in the same protein region (Figure 7). Several
studies have shown that the position of the N-glycan in each protein structure is
important due to the influence of the free energy in the region (Price et al., 2012, 2011).
Tan et al. showed that homologous sequences often have no conserved sequons (Tan
et al., 2014). The features that guide the N-glycan attachment to the target sequon are
extremely complex, and therefore, little is known about this phenomenon. Enzymes
that lack N-glycosylation sequons in homologous sequences most likely found different
evolutionary paths by acquiring mutations that allow for the maintenance of similar
characteristics in the microenvironment (Hanson et al., 2009). However, without a
broad study of glycoproteins, it is not possible to affirm if sequences evolved to acquire
N-glycosylation sequons or if the attachment of N-glycans was the original event and
the sequences are evolving to lack N-glycosylation sites.

69
Figure 7. Conservation of N-glycosylation sites in AN7230 and homologous
proteins. Four AN7230 homologous sequences were selected to represent the slight
differences into cellobiose dehydrogenase enzymes. The sequence alignment shows
variation of N-glycosylated sequon position at primary sequence-level in some
homologous sequences. However, the structures show that the N-glycans are attached
in the same loop at 3D-level (arrows). Selected proteins are from (A) A. nidulans
AN7230, (B) A. kawachii (Aspka1_0182079), (C) A. glaucus (Aspgl1_0179713), (D) A.
aculeatus (Aacu16872_054830) and (E) A. zonatus (Aspzo1_0090575). The 3D
protein structures were modeled using the SWISS-MODEL (Biasini et al., 2014) based
on Phanerochaete chrysosporium cellobiose dehydrogenase (PDB:1KDG).

70
A range of 5 to 9 mannose residues is predominant in A. nidulans N-glycans
We also analyzed the number of mannose residues in N-glycans released from the A.
nidulans-secreted proteins. The results show that high-mannose N-glycans were
predominant, as previously described in Aspergillus sp (Deshpande et al., 2008). It has
been reported that Aspergillus sp. can carry high-mannose type N-glycans with
galactofuranoses (Takayanagi, Kimura, Chiba, & Ajisaka, 1994). Mannose and
galactofuranose are structural isomers, have exactly the same mass and are not
distinguishable from oligosaccharide profiling by MALDI/TOF-MS. Thus, in this study,
the peaks were labeled as “Hex5HexNAc2,” but according to the literature, this is likely
to be Man5GlcNAc2 (Geysens et al., 2009; Maras et al., 1999). The genus Aspergillus
rarely displays hyperglycosylation, and the largest N-glycan was described with 18
mannose residues (Geysens et al., 2009), which is similar to our data. However, we
did not detect glucose or galactose residues in the high-mannose glycans as
previously reported for A. niger (Takayanagi et al., 1994).
A recent study showed that the difference in N-glycans composition is directly related
to the protein structure (Thaysen-Andersen & Packer, 2012). Despite the fact that N-
glycosylation occurs mainly in β-turns, the sequons could be present in a large variety
of structures with low to high accessibility (Petrescu et al., 2004; Thaysen-Andersen &
Packer, 2012). We found a total of 25, 19 and 40 exclusive proteins in the glucose,
SCB and xylan conditions, respectively. Therefore, these specific proteins found in
each secretome could explain the divergence of the N-glycans structures (Figure 6).
Recently, some studies have shown that different substrates can also influence the
composition of PTMs. Adav et al. (2014) detected changes in the N-glycosylation
profile in Phanerochaete chrysosporium when grown in glucose, cellulose and lignin.
The authors showed that the same protein had differences in the position and number
of glycosylation sites depending on the substrate (Adav et al., 2014). Moreover, Stals
et al. (2004) analyzed the N-glycosylation modification in Cel7A from T. reesei strains
(Stals et al., 2004). However, by analyzing the proteins common to all the three growth
conditions (99 proteins), we did not detect changes in the profile of N-glycosylation
when A. nidulans was cultivated in different carbon sources.

71
The knowledge of the N-glycosylation pattern of secreted proteins can assist in the
design of A. nidulans as a host for heterologous protein production. However,
understanding the N-glycosylation of wild type enzymes is important, as changes in N-
glycan composition can affect the main properties of these enzymes (Kruszewska et
al., 2008). The N-glycan profile of T. reesei has been reported because it is largely
used in industry due to its good protein secretion capabilities (Pentillä et al., 2003;
Peterson & Nevalainen, 2012). The T. reesei RUT-C30 strain was reported to have
one additional α-1,3-glucose residue in the N-glycan of the main cellobiohydrolase
(CBHI), suggesting an incorrect maturation process of the N-glycan (Peterson &
Nevalainen, 2012). The N-glycan composition could interfere at the level of protein
secretion. The enzyme secretion process is too complex to affirm that N-glycosylation
is the unique feature interfering in the level of protein secretion, although N-glycans
have been reported to contribute to the secretion process.
Conclusion
The knowledge regarding protein glycosylation in a model host such as A. nidulans is
fundamental to improving the success of heterologous protein secretion. For example,
our personal laboratory experience using A. nidulans for this purpose has shown that
only 30% of all transformed recombinant genes were effectively expressed and
secreted (unpublished data). There are many bottlenecks in protein production by
filamentous fungi, such as folding, transport by vesicles, and secretion, but N-
glycosylation at the correct sites is a fundamental event to ensure a high level of
secretion of target proteins (Lubertozzi & Keasling, 2009; H. Nevalainen & Peterson,
2014; K. M. H. Nevalainen et al., 2005; Punt et al., 2002). Our data may assist attempts
for the design of glycosylation sites of recombinant genes to be expressed in
filamentous fungal hosts. A. nidulans is a model filamentous fungus with an excellent
protein secretion system and with a GRAS (generally regarded as safe) status.
Although A. nidulans is not the main strain used for industrial biomass degradation, it
shows a specialized repertoire for biomass degradation compared with other
filamentous fungi (van Hellemond et al., 2006). Moreover, a large number of genes are
specific to A. nidulans, when compared with other Aspergillus species, and a study of
these enzymes could provide advantages (Benoit et al., 2015).

72
This is the first study to report the glycoproteomics of A. nidulans, with analysis of
proteins, N-glyc sites and N-glycans. Using glucose, xylan and SCB as substrates, we
detected 265 proteins strictly related to each substrate, as well as demonstrated
different patterns of total proteins, glycoproteins and N-glycan profiles. Glycosylation
studies rarely highlight the modifications in CAZymes, which was a focus in this study.
A. nidulans has a preference for the sequon NXT and specific variations. The results
of this study should allow for better manipulation of heterologous proteins using
Aspergillus spp. as a host.
Material and methods
Media and strain
Aspergillus nidulans strain A773 (pyrG89;wA3;pyroA4) was purchased from the Fungal
Genetics Stock Center (FGSC). A. nidulans minimal medium (MM) contained salts
solution (Pontecorvo et al., 1953) (NaNO3 6 g/L, KCl 0.52 g/L, MgSO4.7H2O 0.52 g/L,
KH2PO4 1.52 g/L), trace elements (H3BO3 0.011 g/L, MnCl2.4H2O 0.005 g/L,
FeSO4.7H2O 0.005 g/L, CoCl2.6H2O 0.0016 g/L, CuSO4.5H2O 0.0016 g/L,
Na2MoO4.4H2O 0.0011 g/L, ZnSO4.7H2O 0.022 g/L, Na2EDTA 0.050 g/L) and was
supplemented with pyridoxine (1 mg/L) and uracil/uridine (1.2 g/L each) (Segato et al.,
2012). 10 g/L of glucose, xylan from beechwood or NaOH-pretreated sugarcane
bagasse (SCB) were used as carbon source in different conditions as needed, and pH
was adjusted to 6.5 buffered with 200 mM HEPES (4-(2-hydroxyethyl)-1-
piperazineethanesulfonic acid) (Segato et al., 2012).
Growth conditions
A. nidulans A773 was cultivated in solid minimal media with glucose for 3-4 days,
spores were harvest and filtered using Miracloth (Merck Millipore). 106 spores were
inoculated into 100 ml MM glucose for 24 hours, 37 ºC and 180 rpm (Segato et al.,
2012). The mycelium was collected by filtration, washed using deionized water and
transferred to MM containing glucose, SCB (60% cellulose, 23% hemicellulose, 8%
lignin and 10% ash) or xylan for 4 days at the same conditions. Extracellular proteins
(secretome) were obtained by filtration of supernatant through one layer of Miracloth.
A triplicate was prepared to each secretome. The pretreatment of sugarcane bagasse

73
was carried out as previously described by Rocha et al. (2012) (Rocha, Gonçalves,
Oliveira, Olivares, & Rossell, 2012).
Enzymatic assays
The polysaccharides xylan from beechwood, xyloglucan from tamarind, mannan,
lichenan, beta-glucan from barley, carboxymethyl cellulose (CMC) and starch were
hydrolyzed by A. nidulans secretomes produced on sugarcane bagasse and xylan.
The enzymatic microassay were carried out using 50 μL of the substrates (0.5 %w/v),
50 mM of ammonium acetate buffer at pH 5.5 and 0.5 µg of total protein at 50°C for
120 min. The reactions were stopped using 100 μL of 3,5-dinitrosalicylic acid (DNS)
boiled at 99°C for 5 minutes and the reducing sugars were measured at 550 nm. The
FPAse activity was performed as recommended by Eveleigh et al. (2009) following the
modifications proposed by Camassola and Dillon (2012) (Camassola & J.P. Dillon,
2012; Eveleigh, Mandels, Andreotti, & Roche, 2009). All the enzymatic assays were
performed in triplicate.
Glycoprotein Enrichment and Deglycosylation
The secreted proteins (secretome) were concentrated using centrifugal filters with 10
kDa of pore size to obtain 700 ug to 1 mg of total proteins. Glycoproteins in the
secretomes were enriched by interaction in Concanavalin A (ConA - GE Healthcare)
for 2 hours. The glycoproteins were eluted using a buffered-solution of 500 mM methyl
α-D-glucopyranoside and the eluted proteins were loaded into a 10% SDS-PAGE for
30 min at 110V. The gel bands were excised and treated with 10 units of
endoglycosidase-H (Endo H; New England Biolabs) at 37 ºC during 24 hours for
deglycosylation under denaturing conditions.
Sample preparation for LC-MS/MS analysis
Proteins deglycosylated in-gel were reduced (5 mM dithiothreitol, 30 min, at room
temperature), alkylated (14 mM iodoacetamide, 30 min at room temperature in the
dark), and digested with 20 mg/ml trypsin (Promega). After peptide extraction, the
samples were dried in a vacuum concentrator. 4.5 µL of the peptide mixture was
analyzed on an ETD-enabled LTQ Velos Orbitrap mass spectrometer (Thermo Fisher

74
Scientific) coupled with LC-MS/MS by an EASY-nLC system (Proxeon Biosystems)
through a Proxeon nanoelectrospray ion source. The peptides were separated by a 2-
90% acetonitrile gradient in 0.1% formic acid using a PicoFrit Column analytical column
(20 cm x ID75 μm, 5 μm particle size, New objective) at a flow rate of 300 nL/min over
60 min. The nanoelectrospray voltage was set to 2.2 kV, and the source temperature
was 275°C. The instrument method for the LTQ Velos Orbitrap were set up in the data-
dependent acquisition mode. The full scan MS spectra (m/z 300-1,600) were acquired
in the Orbitrap analyzer after accumulation to a target value of 1e6. Resolution in the
Orbitrap was set to r = 60,000, and the 20 most intense peptide ions with charge states
≥ 2 were sequentially isolated to a target value of 5,000 and fragmented in the linear
ion trap by low-energy CID (normalized collision energy of 35%). The signal threshold
for triggering an MS/MS event was set to 1,000 counts. Dynamic exclusion was
enabled with an exclusion size list of 500, exclusion duration of 60 s, and repeat count
of 1. An activation q of 0.25 and an activation time of 10 ms were used.
The raw data files were converted to a peak list format (mgf) using the Mascot Distiller
v.2.3.2.0 software (Matrix Science Ltd.). These spectra were searched against the A.
nidulans genome from AspGD (10.560 entries) using the Mascot v.2.3.01 engine
(Matrix Science Ltd.) with oxidation of methionine and N-acetylglucosamine (GlcNAc)
tagged on asparagine residue (N+203) as variable modifications, and
carbamidomethylation as fixed modification. Additional parameters were one trypsin-
missed cleavage, a tolerance of 10 ppm for precursor ions and 1 Da for fragment ions.
Data analysis
All datasets processed using the workflow feature in the Mascot software were further
analyzed in the software ScaffoldQ+ (Proteome Software) to validate the MS/MS-
based peptide and protein identifications. Peptide identifications were accepted if they
could be established at greater than 95% probability as specified by the Peptide
Prophet algorithm (Keller, Nesvizhskii, Kolker, & Aebersold, 2002). Peptide
identifications were also required to exceed specific database search engine
thresholds. Mascot identifications required at least both the associated identity scores
and ion scores to be p<0.05. Protein identifications were accepted if they could be
established at greater than 99% probability for protein identification. Protein

75
probabilities were assigned using the Protein Prophet algorithm (Nesvizhskii, Keller,
Kolker, & Aebersold, 2003). Proteins that contained similar peptides and could not be
differentiated based on the MS/MS analysis alone were grouped to satisfy the
principles of parsimony. The scoring parameter (Peptide Probability) in the ScaffoldQ+
software obtained a false discovery rate (FDR) of 0.73%. Using the number of total
spectra output from the ScaffoldQ+ software, we identified the differentially expressed
proteins using spectral counting. Quantitative value was applied to normalize the
spectral counts. The Scaffold PTM (Proteome Software) was used to further validate
glycosylated sites assignments with confidence, based in their pre-established
parameters (Beausoleil et al., 2006; Schwartz & Gygi, 2005).
Glycomics
Around 700 ug of each secretome were treated with a mixture of chloroform and
methanol by four times, to extract lipids. The extracts were incubated at room
temperature with end-over-end agitation. After each lipid extraction procedure, the
insoluble protein-containing materials were collected by centrifugation. The final
insoluble protein pellets were further washed with cold-acetone/water (4:1, v/v) to
eliminate polysaccharides from culture media. Pellets were finally washed with cold-
acetone and dried under a stream of nitrogen. The dried samples were dissolved in
0.1 M Tris-HCl buffer, pH 8.2 containing 10mM CaCl2 and denatured by heating for 5
min at 100 °C. After cooling, the samples were digested with trypsin (37 ºC, overnight).
The samples were heated at 100º C for 5 min to inactivate trypsin and centrifuged at
3000 rpm in a refrigerated centrifuge for 15 min. The supernatants were collected and
dried. Samples were then passed through a C18 sep-pak cartridge and washed with
5% acetic acid to remove contaminants (salts, free sugar, etc.). Peptides and
glycopeptides were eluted in series with 20% iso-propanol in 5% acetic acid, 40% iso-
propanol in 5% acetic acid and 100% iso-propanol and dried in a speed vacuum
concentrator. The dried samples were combined and incubated with PNGase F at 37
ºC overnight to release N-glycans. After digestion, the samples were passed through
a C18 sep-pak cartridge and the released N-glycans was eluted with 5% acetic acid
and dried by lyophilization, and then permethylated based on the method of Anumula
and Taylor (1992) (Anumula & Taylor, 1992) and profiled by mass spectrometry.

76
MALDI/TOF-MS was performed in the reflector positive ion mode using α-
dihyroxybenzoic acid (DHBA, 20 mg/mL solution in 50% methanol:water) as a matrix.
The spectrum was obtained by using a TOF/TOF™ 5800 System (AB SCIEX).
Acknowledgment
This research was supported in part by FAPESP (grant # 2012/20549-4 to ARLD,
2014/06923-6 to FMS and 2009/54067-3 to AFPL), and by the National Institutes of
Health (NIH/NCRR) (grant # P41GM10349010) entitled ‘Integrated Technology
Resource for Biomedical Glycomics’ coordinated by Dr. Parastoo Azadi at the Complex
Carbohydrate Research Center). We are grateful to the National Council for Scientific
and Technological Development (CNPq) for the financial support (441912/2014-1 to
ARLD; 310186/2014-5 and 442333/2014-5 to FMS). MVR, MPZ and FC received
FAPESP fellowships (2013/24988-5; 2014/15403-6; 2014/23051-2). We thank the
LNBio Mass Spectrometry staff and Mayumi Ishihara from CCRC for the assistance
with LC-MS/MS and MALDI-TOF experiments, respectively.
Competing interests
The authors declare that they have no competing interests.
Authors´ contributions
Conceived and designed the experiments: ARLD and AFPL. Performed the
experiments: MVR, MPZ and FC. Analyzed the data: MVR, MPZ, FC, JPFC and ARLD.
Wrote the paper: MVR, JPFC and ARLD. Revised the manuscript: ARLD, RAP and
FMS. All authors read and approved the final manuscript.

77
Additional files
Figure S1. Aspergillus nidulans CAZymes inventory.
(A) The annotation of A. nidulans FGSC A4 CAZymes genes were determined
according to CAZy database (www.cazy.org/e185.html). The prediction of N-
glycosylation sites was carried out using NetNGlyc 1.0 Server that examine the
sequence context of Asn-Xaa-Ser/Thr sequons with threshold 0.5. CAZymes predicted
with N-glycosylation sites were analyzed for signal peptide using SignalP 4.1 Server.
(B) The 190 proteins predicted with N-glycosylation sites and signal peptide were
grouped and shown as CAZy families percentage.
A
B

78
Figure S2. Counts of A. nidulans extracellular CAZymes predicted with N-
glycosylation. The number of N-glycosylation sites of each CAZyme predicted with
signal peptide were analyzed by NetNGlyc 1.0 server.

79
Figure S3. Enzymatic repertoire in A. nidulans secretomes. Secretomes produced
on sugarcane bagasse and xylan were assayed for the hydrolysis of polysaccharides.
All the enzymatic assays were carried out at 50°C for 120 min, using 0.5 µg of total
protein. The hydrolysis were performed in triplicate.

80
Oc
cu
re
nc
e (
%)
S e q u e n c e p o s itio n
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
H y d ro p h o b ic
N S /T
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
A ro m a tic
N S /T
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
B a s ic
N S /T
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
U n iq u e
g ly c o s y la te d
n o n -g ly c o s y la te d
N S /T
N S /T
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
A cid ic
N S /T
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6
0
1 5
3 0
P o la r u n c h a rg e d
Figure S4. Amino acids distribution around N-glycosylation sites. The relative
ocurrence of amino acids is plotted versus sequence position -6 to +6 around
validated or predicted N-glyc site. Glycosylated: N-glyc sites validated by LC-MS/MS;
non-glycosylated: N-glyc sites predicted by NetNGlyc Server but not validadted by
LC-MS/MS data set. Hydrophobic (Ala, Val, Leu, Ile, Met); Aromatic (Phe, Tyr, Trp);
Polar uncharged (Ser, Thr, Asn, Cys, Gln); Acidic (Asp, Glu); Basic (Lys, Arg, His);
Unique (Gly, Pro).

81
Table S1. Identified proteins and spectrum counts.
Total Spectrum Count
Acession
number Identified Proteins Domain 1 Domain 2
Domain
3
N-X-
S/T
sites
¹
N-glyc
Position
Protei
n
covera
ge
Gluco
se A
Gluco
se B
Gluco
se C
SCB
A
SCB
B
SCB
C
Xylan
A
Xylan
B
Xylan
C
ANID_00221 | ANID_00221 | Aspergillus nidulans FGSC A4 chitinase (392 aa) GH18 6% 0 0 0 1 0 2 0 0 0
ANID_00224 | ANID_00224 | Aspergillus nidulans FGSC A4 microsomal dipeptidase (416 aa) Peptidase_M
19 38% 8 2 3 4 3 5 8 7 3
ANID_00231 | ANID_00231 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (332
aa) Tyrosinase 1 284 43% 5 9 5 41 25 31 25 18 44
ANID_00241 | ANID_00241 | Aspergillus nidulans FGSC A4 superoxide dismutase (155 aa) Sod_Cu 32% 0 0 0 1 1 1 5 0 2
ANID_00245 | ANID_00245 | Aspergillus nidulans FGSC A4 endo-1,3(4)-beta-glucanase (343 aa) GH16 12% 0 0 0 2 0 1 1 1 2
ANID_00392 | ANID_00392 | Aspergillus nidulans FGSC A4
endonuclease/exonuclease/phosphatase (435 aa)
Exo_endo_p
hos 1 272 24% 3 5 2 0 0 2 0 4 0
ANID_00393 | ANID_00393 | Aspergillus nidulans FGSC A4 glycosyl hydrolase family 76 protein
(465 aa) GH76 1 205 11% 0 0 1 1 1 0 0 2 1
ANID_00472 | ANID_00472 | Aspergillus nidulans FGSC A4 endo-1,3-beta-glucanase Engl1 (908
aa) GH81 2 219, 240 44% 0 0 0 60 27 33 46 53 48
ANID_00484 | ANID_00484 | Aspergillus nidulans FGSC A4 glycerophosphoryl diester
phosphodiesterase (410 aa) GDPD 20% 2 0 1 0 0 0 1 0 4
ANID_00494 | ANID_00494 | Aspergillus nidulans FGSC A4 1,4-beta-D-glucan-cellobiohydrolyase
(531 aa) GH7 CBM1 25% 0 0 0 50 35 24 1 0 0
ANID_00543 | ANID_00543 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (509
aa) CBM50 (5X) 8% 0 0 0 52 0 10 0 0 0
ANID_00558 | ANID_00558 | Aspergillus nidulans FGSC A4 beta glucanosyltransferase Gel2p
(474 aa) GH72 2 313, 341 22% 5 5 3 4 2 2 0 0 0
ANID_00567 | ANID_00567 | Aspergillus nidulans FGSC A4 choline dehydrogenase (625 aa) AA3 22% 0 0 0 0 0 0 7 1 4
ANID_00779 | ANID_00779 | Aspergillus nidulans FGSC A4 exo-1,3-beta-D-glucanase (965 aa) GH55 19% 3 4 1 15 12 12 13 13 2
ANID_00787 | ANID_00787 | Aspergillus nidulans FGSC A4 alpha-1,2-mannosidase (506 aa) GH47 1 88 53% 50 37 35 97 125 156 40 85 100
ANID_00933 | ANID_00933 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (405
aa) GH16 52% 34 40 37 39 2 1 16 2 3
ANID_00941 | ANID_00941 | Aspergillus nidulans FGSC A4 alpha-glucosidase (874 aa) GH31 1 713 22% 6 0 0 2 4 0 2 2 1
ANID_01058 | ANID_01058 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (315
aa) 1 166 11% 23 41 36 7 1 0 0 0 0

82
ANID_01131 | ANID_01131 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (208
aa) Sod_Cu 2 49, 116 28% 0 0 0 0 4 1 5 0 5
ANID_01142 | ANID_01142 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (606
aa) AA7 25% 2 5 6 1 0 2 1 4 4
ANID_01218 | ANID_01218 | Aspergillus nidulans FGSC A4 phosphoglycerate mutase (407 aa) His_Phos_1 1 186 29% 3 0 0 3 1 3 3 4 1
ANID_01263 | ANID_01263 | Aspergillus nidulans FGSC A4 S-adenosylhomocysteine hydrolase
(450 aa)
AdoHcyase_
NAD AdoHcyase 2% 0 1 0 0 0 0 0 0 0
ANID_01273 | ANID_01273 | Aspergillus nidulans FGSC A4 glycosyl hydrolase family 6 protein
(405 aa) GH6 43% 0 0 0 83 6 6 0 0 0
ANID_01285 | ANID_01285 | Aspergillus nidulans FGSC A4 endo-beta-1,4-glucanase (327 aa) GH5 8% 0 0 0 2 1 2 0 0 0
ANID_01318 | ANID_01318 | Aspergillus nidulans FGSC A4 tyrosinase (385 aa) Tyrosinase 1 229 22% 4 2 0 0 0 0 0 0 0
ANID_01320 | ANID_01320 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (256
aa) CE10 1 63 58% 1 0 2 2 4 1 15 28 27
ANID_01338 | ANID_01338 | Aspergillus nidulans FGSC A4 hypothetical protein (172 aa) 23% 0 1 2 3 0 3 5 1 0
ANID_01342 | ANID_01342 | Aspergillus nidulans FGSC A4 aminotransferase (387 aa) Aminotran_5 6% 0 0 0 1 0 0 3 0 0
ANID_01426 | ANID_01426 | Aspergillus nidulans FGSC A4 carboxypeptidase S1 (555 aa) Peptidase_S
10 8% 1 0 1 2 0 1 0 0 0
ANID_01433 | ANID_01433 | Aspergillus nidulans FGSC A4 triacylglycerol lipase (541 aa) CE10 2 374, 381 43% 0 0 0 0 2 0 12 16 49
ANID_01502 | ANID_01502 | Aspergillus nidulans FGSC A4 N-acetylglucosaminidase (604 aa) GH20 1 503 45% 3 0 0 3 10 12 109 69 207
ANID_01602 | ANID_01602 | Aspergillus nidulans FGSC A4 endoglucanase B (358 aa) AA9 CBM1 21% 0 0 2 6 2 0 0 0 0
ANID_01719 | ANID_01719 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (384
aa) 9% 0 6 4 0 0 0 0 0 0
ANID_01772 | ANID_01772 | Aspergillus nidulans FGSC A4 feruloyl esterase (528 aa) CE1 Tannase 3 86, 139, 356 55% 1 0 8 9 8 12 13 33 33
ANID_01792 | ANID_01792 | Aspergillus nidulans FGSC A4 GDSL Lipase/Acylhydrolase (342 aa) CE16 29% 0 0 0 0 2 2 9 11 18
ANID_01799 | ANID_01799 | Aspergillus nidulans FGSC A4 secretory lipase (451 aa) LIP 24% 1 0 7 0 0 0 0 1 0
ANID_01813 | ANID_01813 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (160
aa) 31% 5 0 1 0 0 0 3 0 1
ANID_01818 | ANID_01818 | Aspergillus nidulans FGSC A4 xylanaseF1 (310 aa) GH10 69% 1 0 0 57 40 64 43 12 10
ANID_01855 | ANID_01855 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (382
aa) 12% 2 1 2 1 1 1 0 1 0
ANID_01870 | ANID_01870 | Aspergillus nidulans FGSC A4 xylanase D (308 aa) GH43 1 87 31% 2 2 3 1 1 3 4 3 5
ANID_02017 | ANID_02017 | Aspergillus nidulans FGSC A4 alpha-glucosidase AgdA (993 aa) GH31 1 432 21% 0 1 2 4 6 5 9 3 3
ANID_02018 | ANID_02018 | Aspergillus nidulans FGSC A4 alpha-amylase AmyA (492 aa) GH13 2 39, 390 11% 0 0 0 1 0 2 0 1 2
ANID_02060 | ANID_02060 | Aspergillus nidulans FGSC A4 exo-arabinanase abnx-Penicillium
chrysogenum (379 aa) GH93 44% 0 0 1 1 2 0 5 3 11

83
ANID_02112 | ANID_02112 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (410
aa) Lactonase 2 45, 220 64% 2 0 1 23 21 30 17 21 81
ANID_02119 | ANID_02119 | Aspergillus nidulans FGSC A4 patched sphingolipid transporter
(1272 aa) Patched 6% 1 0 1 0 0 0 4 3 2
ANID_02217 | ANID_02217 | Aspergillus nidulans FGSC A4 beta-glucosidase (780 aa) GH3 17% 0 0 0 2 10 6 2 1 5
ANID_02227 | ANID_02227 | Aspergillus nidulans FGSC A4 beta-glucosidase (839 aa) GH3 9% 0 0 0 3 1 3 0 0 0
ANID_02237 | ANID_02237 | Aspergillus nidulans FGSC A4 carboxypeptidase S1 (612 aa) Peptidase_S
10 1 494 19% 1 2 0 2 6 4 1 2 1
ANID_02325 | ANID_02325 | Aspergillus nidulans FGSC A4 glycosyl hydrolase, family 92 protein
(755 aa) GH92 4
105, 112, 118,
261 29% 0 0 0 1 3 0 7 11 17
ANID_02359 | ANID_02359 | Aspergillus nidulans FGSC A4 beta-xylosidase (801 aa) GH3 3 231, 673, 695 42% 0 0 0 18 30 33 9 15 15
ANID_02360 | ANID_02360 | Aspergillus nidulans FGSC A4 acid phosphatase (498 aa) Metallophos Metallophos_
C 2 112, 166 28% 0 0 1 0 1 0 2 1 7
ANID_02366 | ANID_02366 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (255
aa) Trypsin 58% 63 62 59 6 0 0 1 0 0
ANID_02385 | ANID_02385 | Aspergillus nidulans FGSC A4 GPI anchored endo-1,3(4)-beta-
glucanase (627 aa) GH16 24% 3 25 13 5 0 0 0 0 0
ANID_02387 | ANID_02387 | Aspergillus nidulans FGSC A4 FAD binding domain-containing
protein (503 aa) AA7 1 260 37% 5 4 2 0 2 3 1 9 13
ANID_02388 | ANID_02388 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (436
aa) AA9 1 93 32% 13 8 18 10 11 17 4 16 27
ANID_02395 | ANID_02395 | Aspergillus nidulans FGSC A4 hydrolase (614 aa) GH2 1 293 33% 0 1 0 17 26 15 14 26 27
ANID_02463 | ANID_02463 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (1024
aa) GH2 7% 0 0 0 0 0 0 2 1 6
ANID_02569 | ANID_02569 | Aspergillus nidulans FGSC A4 pectin lyase (380 aa) PL1 18% 0 0 0 9 0 3 0 0 0
ANID_02572 | ANID_02572 | Aspergillus nidulans FGSC A4 secreted dipeptidyl peptidase DppV
(723 aa) CE10 10% 0 0 0 0 0 0 2 0 2
ANID_02574 | ANID_02574 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (517
aa) AA7 2 212, 330 39% 0 0 0 1 0 0 2 11 16
ANID_02582 | ANID_02582 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (442
aa) 8% 2 5 4 0 0 0 0 0 0
ANID_02648 | ANID_02648 | Aspergillus nidulans FGSC A4 isoamyl alcohol oxidase (567 aa) AA7 1 47 23% 4 1 0 1 1 0 2 11 6
ANID_02659 | ANID_02659 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (831
aa) 5% 0 0 0 1 0 2 0 4 0
ANID_02702 | ANID_02702 | Aspergillus nidulans FGSC A4 predicted protein (228 aa) 19% 2 17 16 0 1 0 0 1 0
ANID_02704 | ANID_02704 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (579
aa) AA3 8% 0 0 3 0 0 0 0 0 0
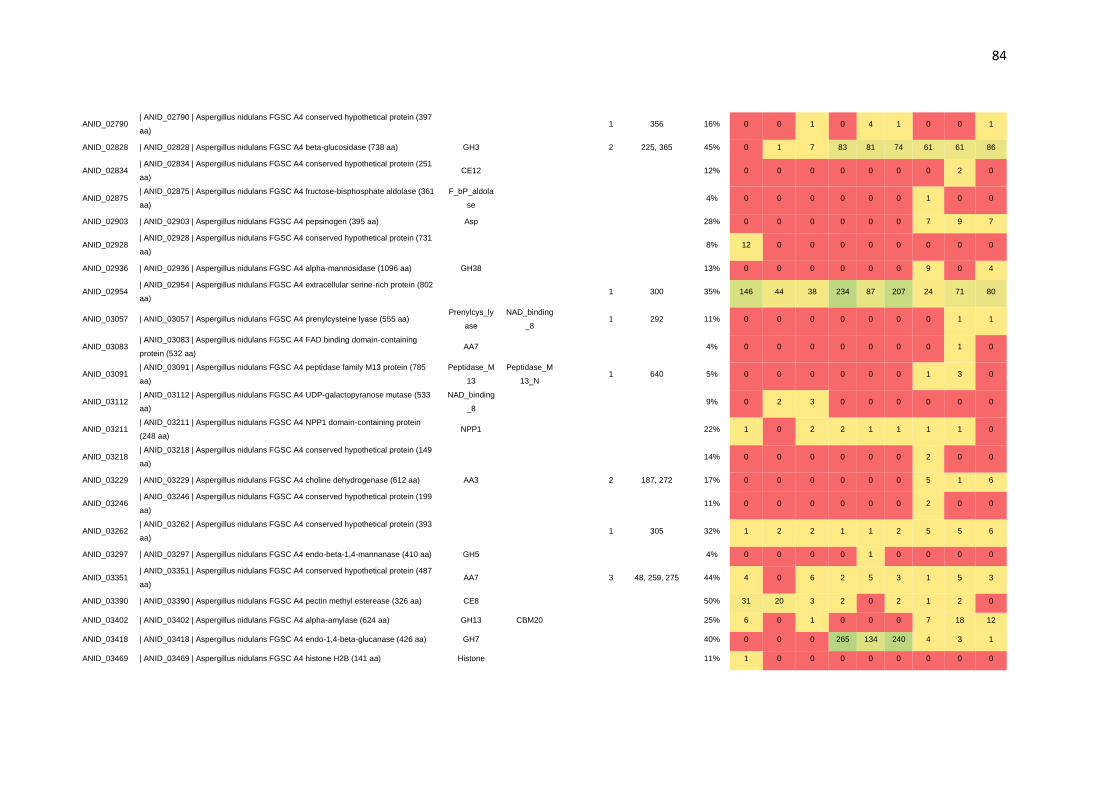
84
ANID_02790 | ANID_02790 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (397
aa) 1 356 16% 0 0 1 0 4 1 0 0 1
ANID_02828 | ANID_02828 | Aspergillus nidulans FGSC A4 beta-glucosidase (738 aa) GH3 2 225, 365 45% 0 1 7 83 81 74 61 61 86
ANID_02834 | ANID_02834 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (251
aa) CE12 12% 0 0 0 0 0 0 0 2 0
ANID_02875 | ANID_02875 | Aspergillus nidulans FGSC A4 fructose-bisphosphate aldolase (361
aa)
F_bP_aldola
se 4% 0 0 0 0 0 0 1 0 0
ANID_02903 | ANID_02903 | Aspergillus nidulans FGSC A4 pepsinogen (395 aa) Asp 28% 0 0 0 0 0 0 7 9 7
ANID_02928 | ANID_02928 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (731
aa) 8% 12 0 0 0 0 0 0 0 0
ANID_02936 | ANID_02936 | Aspergillus nidulans FGSC A4 alpha-mannosidase (1096 aa) GH38 13% 0 0 0 0 0 0 9 0 4
ANID_02954 | ANID_02954 | Aspergillus nidulans FGSC A4 extracellular serine-rich protein (802
aa) 1 300 35% 146 44 38 234 87 207 24 71 80
ANID_03057 | ANID_03057 | Aspergillus nidulans FGSC A4 prenylcysteine lyase (555 aa) Prenylcys_ly
ase
NAD_binding
_8 1 292 11% 0 0 0 0 0 0 0 1 1
ANID_03083 | ANID_03083 | Aspergillus nidulans FGSC A4 FAD binding domain-containing
protein (532 aa) AA7 4% 0 0 0 0 0 0 0 1 0
ANID_03091 | ANID_03091 | Aspergillus nidulans FGSC A4 peptidase family M13 protein (785
aa)
Peptidase_M
13
Peptidase_M
13_N 1 640 5% 0 0 0 0 0 0 1 3 0
ANID_03112 | ANID_03112 | Aspergillus nidulans FGSC A4 UDP-galactopyranose mutase (533
aa)
NAD_binding
_8 9% 0 2 3 0 0 0 0 0 0
ANID_03211 | ANID_03211 | Aspergillus nidulans FGSC A4 NPP1 domain-containing protein
(248 aa) NPP1 22% 1 0 2 2 1 1 1 1 0
ANID_03218 | ANID_03218 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (149
aa) 14% 0 0 0 0 0 0 2 0 0
ANID_03229 | ANID_03229 | Aspergillus nidulans FGSC A4 choline dehydrogenase (612 aa) AA3 2 187, 272 17% 0 0 0 0 0 0 5 1 6
ANID_03246 | ANID_03246 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (199
aa) 11% 0 0 0 0 0 0 2 0 0
ANID_03262 | ANID_03262 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (393
aa) 1 305 32% 1 2 2 1 1 2 5 5 6
ANID_03297 | ANID_03297 | Aspergillus nidulans FGSC A4 endo-beta-1,4-mannanase (410 aa) GH5 4% 0 0 0 0 1 0 0 0 0
ANID_03351 | ANID_03351 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (487
aa) AA7 3 48, 259, 275 44% 4 0 6 2 5 3 1 5 3
ANID_03390 | ANID_03390 | Aspergillus nidulans FGSC A4 pectin methyl esterease (326 aa) CE8 50% 31 20 3 2 0 2 1 2 0
ANID_03402 | ANID_03402 | Aspergillus nidulans FGSC A4 alpha-amylase (624 aa) GH13 CBM20 25% 6 0 1 0 0 0 7 18 12
ANID_03418 | ANID_03418 | Aspergillus nidulans FGSC A4 endo-1,4-beta-glucanase (426 aa) GH7 40% 0 0 0 265 134 240 4 3 1
ANID_03469 | ANID_03469 | Aspergillus nidulans FGSC A4 histone H2B (141 aa) Histone 11% 1 0 0 0 0 0 0 0 0

85
ANID_03592 | ANID_03592 | Aspergillus nidulans FGSC A4 calnexin (562 aa) Calreticulin 1 235 16% 1 0 0 0 0 0 1 5 2
ANID_03613 | ANID_03613 | Aspergillus nidulans FGSC A4 xylanase (226 aa) GH11 40% 0 0 0 110 75 125 77 13 128
ANID_03790 | ANID_03790 | Aspergillus nidulans FGSC A4 alpha-1,3-glucanase/mutanase (494
aa) GH71 8% 0 0 0 0 3 1 1 1 3
ANID_03860 | ANID_03860 | Aspergillus nidulans FGSC A4 endoglucanase (251 aa) AA9 14% 0 0 0 5 5 5 0 0 0
ANID_03918 | ANID_03918 | Aspergillus nidulans FGSC A4 aminopeptidase (503 aa) PA Peptidase_M
28 22% 2 3 1 9 6 3 7 12 25
ANID_04041 | ANID_04041 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (194
aa) 34% 0 1 0 5 2 6 4 3 13
ANID_04055 | ANID_04055 | Aspergillus nidulans FGSC A4 acid phosphatase PHOa (407 aa) Phosphoeste
rase 5
119, 150, 186,
315, 382 41% 0 1 2 14 26 21 13 23 20
ANID_04102 | ANID_04102 | Aspergillus nidulans FGSC A4 beta-glucosidase (854 aa) GH3 4 62, 491, 642, 713 45% 1 0 0 11 20 8 18 22 33
ANID_04245 | ANID_04245 | Aspergillus nidulans FGSC A4 neutral/alkaline nonlysosomal
ceramidase (759 aa)
Ceramidase_
alk 20% 0 0 0 3 7 3 3 5 2
ANID_04260 | ANID_04260 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (196
aa) 5% 1 0 1 0 0 1 1 0 0
ANID_04376 | ANID_04376 | Aspergillus nidulans FGSC A4 nadp-specific glutamate
dehydrogenase (460 aa)
ELFV_dehyd
rog
ELFV_dehydr
og_N 41% 1 4 17 2 1 1 0 0 0
ANID_04379 | ANID_04379 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (285
aa) DPBB_1 12% 0 75 44 1 0 0 0 0 0
ANID_04381 | ANID_04381 | Aspergillus nidulans FGSC A4 GPI anchored cell wall protein (175
aa) 17% 3 12 9 0 0 0 0 0 0
ANID_04390 | ANID_04390 | Aspergillus nidulans FGSC A4 GPI-anchored cell wall organization
protein Ecm33 (397 aa) Ecm33 LRR_5 (2x) 2 78, 209 16% 7 18 12 0 1 0 0 0 0
ANID_04515 | ANID_04515 | Aspergillus nidulans FGSC A4 probable glycosidase crf2 (436 aa) GH16 CBM18 26% 3 21 12 0 0 0 0 0 0
ANID_04575 | ANID_04575 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (358
aa) 19% 62 17 13 1 0 3 0 1 0
ANID_04700 | ANID_04700 | Aspergillus nidulans FGSC A4 endo-beta-1,3-glucanase (650 aa) GH17 24% 0 4 5 1 0 1 2 4 1
ANID_04793 | ANID_04793 | Aspergillus nidulans FGSC A4 aspartate-semialdehyde
dehydrogenase (364 aa)
Semialdhyde
_dh
Semialdhyde_
dhC 12% 1 1 2 0 0 0 0 0 0
ANID_04807 | ANID_04807 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (327
aa) DUF946 13% 0 1 2 0 0 0 2 1 1
ANID_04809 | ANID_04809 | Aspergillus nidulans FGSC A4 glutaminase A (689 aa) DUF1793 1 506 42% 0 1 0 34 44 50 13 54 82
ANID_04825 | ANID_04825 | Aspergillus nidulans FGSC A4 LysM domain-containing protein (901
aa) GH55 34% 0 0 0 17 11 17 18 3 12
ANID_04845 | ANID_04845 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (376
aa) 9% 0 0 0 0 0 0 2 0 0
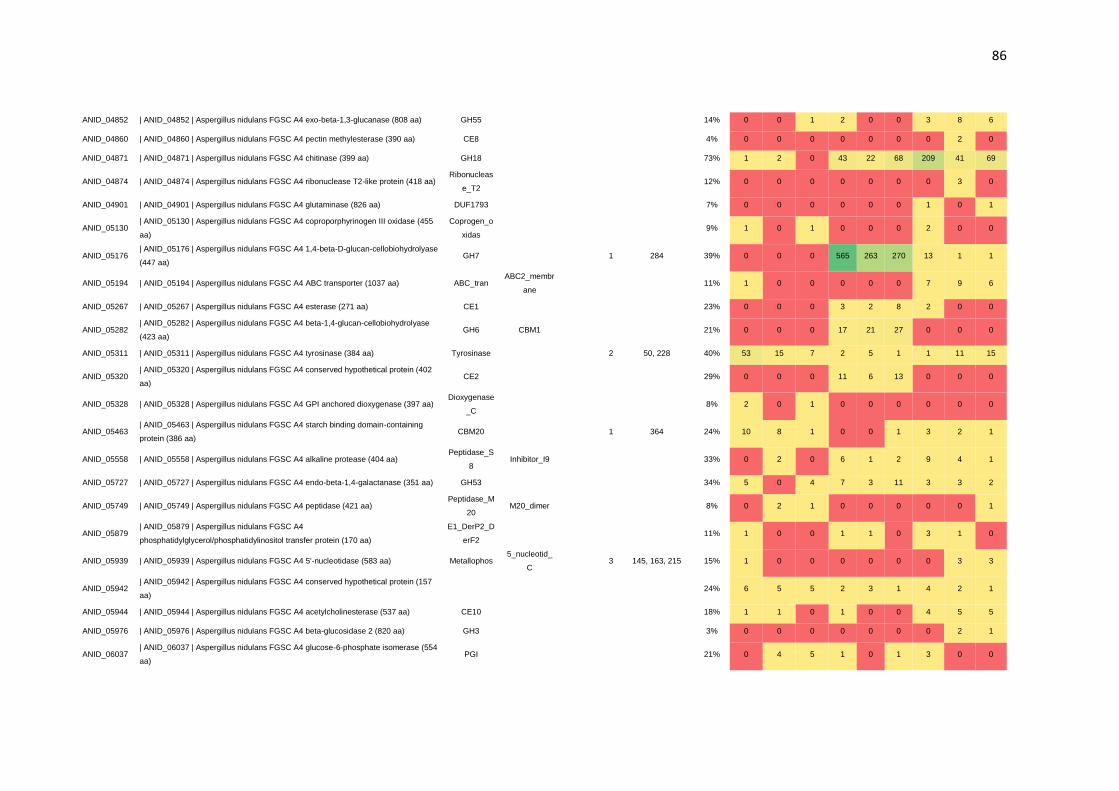
86
ANID_04852 | ANID_04852 | Aspergillus nidulans FGSC A4 exo-beta-1,3-glucanase (808 aa) GH55 14% 0 0 1 2 0 0 3 8 6
ANID_04860 | ANID_04860 | Aspergillus nidulans FGSC A4 pectin methylesterase (390 aa) CE8 4% 0 0 0 0 0 0 0 2 0
ANID_04871 | ANID_04871 | Aspergillus nidulans FGSC A4 chitinase (399 aa) GH18 73% 1 2 0 43 22 68 209 41 69
ANID_04874 | ANID_04874 | Aspergillus nidulans FGSC A4 ribonuclease T2-like protein (418 aa) Ribonucleas
e_T2 12% 0 0 0 0 0 0 0 3 0
ANID_04901 | ANID_04901 | Aspergillus nidulans FGSC A4 glutaminase (826 aa) DUF1793 7% 0 0 0 0 0 0 1 0 1
ANID_05130 | ANID_05130 | Aspergillus nidulans FGSC A4 coproporphyrinogen III oxidase (455
aa)
Coprogen_o
xidas 9% 1 0 1 0 0 0 2 0 0
ANID_05176 | ANID_05176 | Aspergillus nidulans FGSC A4 1,4-beta-D-glucan-cellobiohydrolyase
(447 aa) GH7 1 284 39% 0 0 0 565 263 270 13 1 1
ANID_05194 | ANID_05194 | Aspergillus nidulans FGSC A4 ABC transporter (1037 aa) ABC_tran ABC2_membr
ane 11% 1 0 0 0 0 0 7 9 6
ANID_05267 | ANID_05267 | Aspergillus nidulans FGSC A4 esterase (271 aa) CE1 23% 0 0 0 3 2 8 2 0 0
ANID_05282 | ANID_05282 | Aspergillus nidulans FGSC A4 beta-1,4-glucan-cellobiohydrolyase
(423 aa) GH6 CBM1 21% 0 0 0 17 21 27 0 0 0
ANID_05311 | ANID_05311 | Aspergillus nidulans FGSC A4 tyrosinase (384 aa) Tyrosinase 2 50, 228 40% 53 15 7 2 5 1 1 11 15
ANID_05320 | ANID_05320 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (402
aa) CE2 29% 0 0 0 11 6 13 0 0 0
ANID_05328 | ANID_05328 | Aspergillus nidulans FGSC A4 GPI anchored dioxygenase (397 aa) Dioxygenase
_C 8% 2 0 1 0 0 0 0 0 0
ANID_05463 | ANID_05463 | Aspergillus nidulans FGSC A4 starch binding domain-containing
protein (386 aa) CBM20 1 364 24% 10 8 1 0 0 1 3 2 1
ANID_05558 | ANID_05558 | Aspergillus nidulans FGSC A4 alkaline protease (404 aa) Peptidase_S
8 Inhibitor_I9 33% 0 2 0 6 1 2 9 4 1
ANID_05727 | ANID_05727 | Aspergillus nidulans FGSC A4 endo-beta-1,4-galactanase (351 aa) GH53 34% 5 0 4 7 3 11 3 3 2
ANID_05749 | ANID_05749 | Aspergillus nidulans FGSC A4 peptidase (421 aa) Peptidase_M
20 M20_dimer 8% 0 2 1 0 0 0 0 0 1
ANID_05879 | ANID_05879 | Aspergillus nidulans FGSC A4
phosphatidylglycerol/phosphatidylinositol transfer protein (170 aa)
E1_DerP2_D
erF2 11% 1 0 0 1 1 0 3 1 0
ANID_05939 | ANID_05939 | Aspergillus nidulans FGSC A4 5'-nucleotidase (583 aa) Metallophos 5_nucleotid_
C 3 145, 163, 215 15% 1 0 0 0 0 0 0 3 3
ANID_05942 | ANID_05942 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (157
aa) 24% 6 5 5 2 3 1 4 2 1
ANID_05944 | ANID_05944 | Aspergillus nidulans FGSC A4 acetylcholinesterase (537 aa) CE10 18% 1 1 0 1 0 0 4 5 5
ANID_05976 | ANID_05976 | Aspergillus nidulans FGSC A4 beta-glucosidase 2 (820 aa) GH3 3% 0 0 0 0 0 0 0 2 1
ANID_06037 | ANID_06037 | Aspergillus nidulans FGSC A4 glucose-6-phosphate isomerase (554
aa) PGI 21% 0 4 5 1 0 1 3 0 0

87
ANID_06093 | ANID_06093 | Aspergillus nidulans FGSC A4 acetylxylan esterase (307 aa) CE1 1 263 25% 0 0 0 25 3 25 7 2 6
ANID_06103 | ANID_06103 | Aspergillus nidulans FGSC A4 starch binding domain-containing
protein (329 aa)
Chitin_bind_
3 29% 4 0 2 0 0 0 2 4 1
ANID_06106 | ANID_06106 | Aspergillus nidulans FGSC A4 pectate lyase (234 aa) PL3 18% 0 0 0 3 0 0 1 1 2
ANID_06382 | ANID_06382 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (631
aa) 16% 0 0 0 2 2 2 10 1 2
ANID_06405 | ANID_06405 | Aspergillus nidulans FGSC A4 secreted glycosyl hydrolase (247 aa) GH25 1 183 32% 0 10 1 0 0 0 0 1 0
ANID_06422 | ANID_06422 | Aspergillus nidulans FGSC A4 fungal cellulose binding domain-
containing protein (306 aa) CE16 19% 0 0 0 56 89 98 0 1 0
ANID_06428 | ANID_06428 | Aspergillus nidulans FGSC A4 fungal cellulose binding domain-
containing protein (232 aa) AA9 1 69 33% 0 0 0 13 38 16 0 0 0
ANID_06438 | ANID_06438 | Aspergillus nidulans FGSC A4 dipeptidyl-peptidase IV (774 aa) CE1 2 496, 671 30% 0 0 0 4 3 0 5 11 9
ANID_06470 | ANID_06470 | Aspergillus nidulans FGSC A4 N,O-diacetylmuramidase (217 aa) GH25 24% 0 2 0 1 0 2 2 1 2
ANID_06473 | ANID_06473 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (693
aa) 1 459 10% 0 0 0 0 1 0 1 2 1
ANID_06476 | ANID_06476 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (135
aa) 29% 0 0 0 6 2 6 0 0 0
ANID_06525 | ANID_06525 | Aspergillus nidulans FGSC A4 NAD-dependent formate
dehydrogenase AciA/Fdh (366 aa) 2-Hacid_dh
2-
Hacid_dh_C 20% 1 1 2 0 0 1 0 0 0
ANID_06535 | ANID_06535 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (217
aa) 1 51 73% 61 65 69 92 10 55 65 34 97
ANID_06620 | ANID_06620 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (389
aa) GH16 1 166 31% 20 10 13 2 4 4 7 2 6
ANID_06621 | ANID_06621 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (384
aa)
Abhydrolase
_6 40% 9 11 2 2 4 1 1 3 6
ANID_06635 | ANID_06635 | Aspergillus nidulans FGSC A4 laccase (610 aa) AA1 Cu-oxidase
(3x) 1 403 38% 0 0 0 0 0 0 11 7 22
ANID_06673 | ANID_06673 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (808
aa) GH95 26% 0 0 0 1 4 1 4 11 9
ANID_06697 | ANID_06697 | Aspergillus nidulans FGSC A4 SUN domain-containing protein (411
aa) GH132 27% 15 7 14 0 0 0 0 0 0
ANID_06795 | ANID_06795 | Aspergillus nidulans FGSC A4 antigenic cell wall
galactomannoprotein (189 aa) HsbA 13% 0 0 0 0 0 2 0 0 0
ANID_06819 | ANID_06819 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (308
aa) GH16 37% 11 12 13 27 15 45 15 20 14
ANID_06927 | ANID_06927 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (206
aa) WSC 12% 0 3 2 0 0 0 0 0 0
ANID_07041 | ANID_07041 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (822
aa) 3% 1 1 2 0 0 0 0 0 0

88
ANID_07046 | ANID_07046 | Aspergillus nidulans FGSC A4 carboxylesterase (580 aa) CE10 9% 0 0 0 0 0 0 2 5 2
ANID_07087 | ANID_07087 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (274
aa) 62% 1 0 0 11 7 16 11 2 2
ANID_07102 | ANID_07102 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (173
aa) 34% 2 0 0 0 1 2 0 2 1
ANID_07135 | ANID_07135 | Aspergillus nidulans FGSC A4 rhamnogalacturonase B (531 aa) PL4 12% 0 0 0 3 2 2 0 0 0
ANID_07151 | ANID_07151 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (879
aa) GH78 CBM67 3 36, 58, 586 24% 0 0 0 0 0 0 3 12 11
ANID_07152 | ANID_07152 | Aspergillus nidulans FGSC A4 alpha-galactosidase (641 aa) GH27 1 532 35% 0 0 0 16 21 18 5 8 8
ANID_07159 | ANID_07159 | Aspergillus nidulans FGSC A4 tripeptidyl-peptidase (605 aa) Peptidase_S
8
Pro-
kuma_activ 17% 1 2 1 0 0 1 0 1 0
ANID_07181 | ANID_07181 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (453
aa) 27% 0 1 1 3 0 3 1 1 5
ANID_07214 | ANID_07214 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (464
aa)
NAD_binding
_8 10% 1 3 1 0 0 0 0 0 0
ANID_07230 | ANID_07230 | Aspergillus nidulans FGSC A4 cellobiose dehydrogenase (797 aa) AA8 5 132, 299, 308,
620, 709 47% 0 0 0 45 190 40 0 0 0
ANID_07231 | ANID_07231 | Aspergillus nidulans FGSC A4 extracelular serine carboxypeptidase
(520 aa)
Peptidase_S
28 4
282, 294, 309,
312 42% 2 0 4 3 6 5 6 13 6
ANID_07269 | ANID_07269 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (497
aa) AA7 2 133, 460 61% 27 10 21 19 25 19 21 29 32
ANID_07275 | ANID_07275 | Aspergillus nidulans FGSC A4 xylosidase/glycosyl hydrolase (517
aa) GH43 2 40, 382 32% 0 0 0 24 7 24 4 7 4
ANID_07307 | ANID_07307 | Aspergillus nidulans FGSC A4 DUF1237 domain-containing protein
(538 aa) GH125 1 68 26% 0 0 4 2 8 8 5 4 8
ANID_07331 | ANID_07331 | Aspergillus nidulans FGSC A4 cyanate hydratase (161 aa) Cyanate_lya
se 34% 0 0 0 1 0 0 4 0 1
ANID_07345 | ANID_07345 | Aspergillus nidulans FGSC A4 alpha/beta-glucosidase (895 aa) GH31 1 536 3% 0 0 0 0 1 0 0 0 1
ANID_07349 | ANID_07349 | Aspergillus nidulans FGSC A4 mutanase (432 aa) GH71 13% 0 0 0 0 0 0 4 2 0
ANID_07389 | ANID_07389 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (581
aa) AA1
Cu-oxidase
(3x) 2 55, 472 18% 0 0 0 4 14 7 0 2 3
ANID_07396 | ANID_07396 | Aspergillus nidulans FGSC A4 beta-glucosidase (773 aa) GH3 3 259, 438, 586 24% 0 0 0 2 7 6 3 4 5
ANID_07401 | ANID_07401 | Aspergillus nidulans FGSC A4 endo-1,4-beta-xylanase (382 aa) GH10 CBM1 1 123 43% 0 0 0 39 65 56 26 5 6
ANID_07402 | ANID_07402 | Aspergillus nidulans FGSC A4 glucan 1,4-alpha-glucosidase (635
aa) GH15 CBM20 24% 15 3 0 2 0 0 0 3 0
ANID_07511 | ANID_07511 | Aspergillus nidulans FGSC A4 1,3-beta-glucanosyltransferase (540
aa) GH72 CBM43 2 36, 415 26% 9 7 4 12 7 10 3 16 9
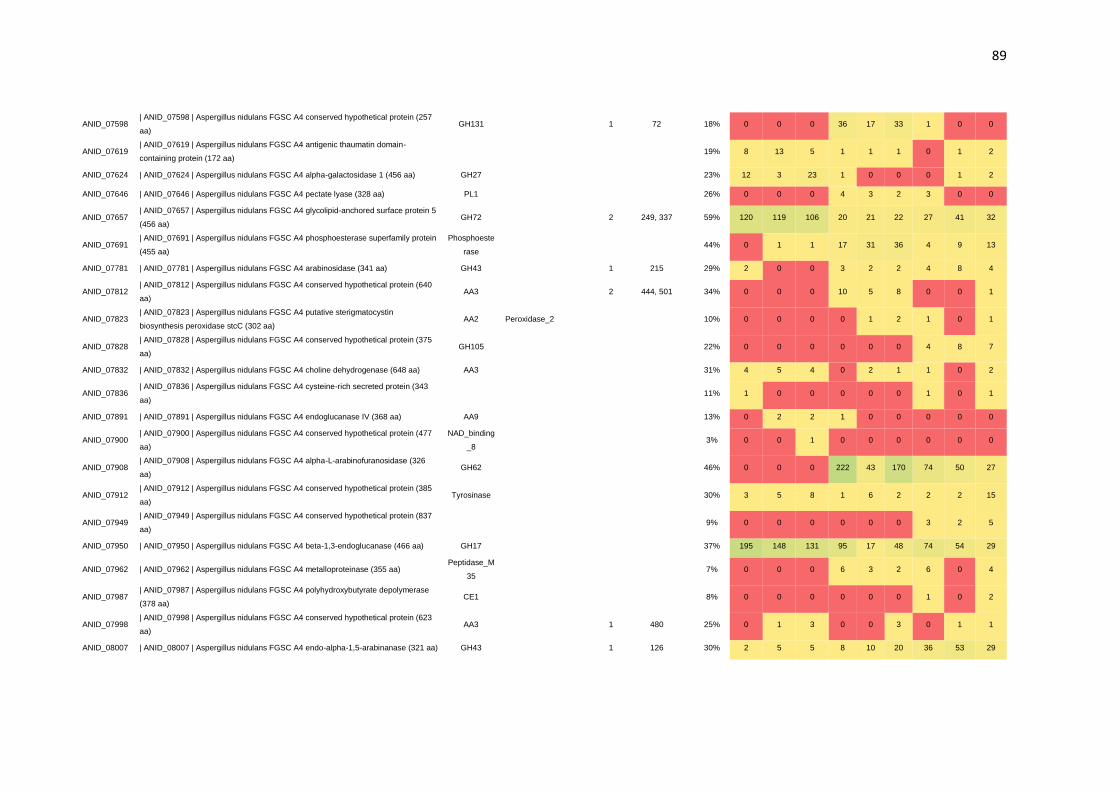
89
ANID_07598 | ANID_07598 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (257
aa) GH131 1 72 18% 0 0 0 36 17 33 1 0 0
ANID_07619 | ANID_07619 | Aspergillus nidulans FGSC A4 antigenic thaumatin domain-
containing protein (172 aa) 19% 8 13 5 1 1 1 0 1 2
ANID_07624 | ANID_07624 | Aspergillus nidulans FGSC A4 alpha-galactosidase 1 (456 aa) GH27 23% 12 3 23 1 0 0 0 1 2
ANID_07646 | ANID_07646 | Aspergillus nidulans FGSC A4 pectate lyase (328 aa) PL1 26% 0 0 0 4 3 2 3 0 0
ANID_07657 | ANID_07657 | Aspergillus nidulans FGSC A4 glycolipid-anchored surface protein 5
(456 aa) GH72 2 249, 337 59% 120 119 106 20 21 22 27 41 32
ANID_07691 | ANID_07691 | Aspergillus nidulans FGSC A4 phosphoesterase superfamily protein
(455 aa)
Phosphoeste
rase 44% 0 1 1 17 31 36 4 9 13
ANID_07781 | ANID_07781 | Aspergillus nidulans FGSC A4 arabinosidase (341 aa) GH43 1 215 29% 2 0 0 3 2 2 4 8 4
ANID_07812 | ANID_07812 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (640
aa) AA3 2 444, 501 34% 0 0 0 10 5 8 0 0 1
ANID_07823 | ANID_07823 | Aspergillus nidulans FGSC A4 putative sterigmatocystin
biosynthesis peroxidase stcC (302 aa) AA2 Peroxidase_2 10% 0 0 0 0 1 2 1 0 1
ANID_07828 | ANID_07828 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (375
aa) GH105 22% 0 0 0 0 0 0 4 8 7
ANID_07832 | ANID_07832 | Aspergillus nidulans FGSC A4 choline dehydrogenase (648 aa) AA3 31% 4 5 4 0 2 1 1 0 2
ANID_07836 | ANID_07836 | Aspergillus nidulans FGSC A4 cysteine-rich secreted protein (343
aa) 11% 1 0 0 0 0 0 1 0 1
ANID_07891 | ANID_07891 | Aspergillus nidulans FGSC A4 endoglucanase IV (368 aa) AA9 13% 0 2 2 1 0 0 0 0 0
ANID_07900 | ANID_07900 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (477
aa)
NAD_binding
_8 3% 0 0 1 0 0 0 0 0 0
ANID_07908 | ANID_07908 | Aspergillus nidulans FGSC A4 alpha-L-arabinofuranosidase (326
aa) GH62 46% 0 0 0 222 43 170 74 50 27
ANID_07912 | ANID_07912 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (385
aa) Tyrosinase 30% 3 5 8 1 6 2 2 2 15
ANID_07949 | ANID_07949 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (837
aa) 9% 0 0 0 0 0 0 3 2 5
ANID_07950 | ANID_07950 | Aspergillus nidulans FGSC A4 beta-1,3-endoglucanase (466 aa) GH17 37% 195 148 131 95 17 48 74 54 29
ANID_07962 | ANID_07962 | Aspergillus nidulans FGSC A4 metalloproteinase (355 aa) Peptidase_M
35 7% 0 0 0 6 3 2 6 0 4
ANID_07987 | ANID_07987 | Aspergillus nidulans FGSC A4 polyhydroxybutyrate depolymerase
(378 aa) CE1 8% 0 0 0 0 0 0 1 0 2
ANID_07998 | ANID_07998 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (623
aa) AA3 1 480 25% 0 1 3 0 0 3 0 1 1
ANID_08007 | ANID_08007 | Aspergillus nidulans FGSC A4 endo-alpha-1,5-arabinanase (321 aa) GH43 1 126 30% 2 5 5 8 10 20 36 53 29

90
ANID_08068 | ANID_08068 | Aspergillus nidulans FGSC A4 extracellular endoglucanase (573 aa) GH5 CBM46 1 275 23% 0 0 0 5 16 13 0 0 0
ANID_08086 | ANID_08086 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (154
aa)
GPI-
anchored 2 43, 73 78% 22 11 10 3 10 9 14 24 36
ANID_08145 | ANID_08145 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (171
aa) Cupin_5 2 40, 46 25% 1 2 1 1 0 0 2 1 3
ANID_08218 | ANID_08218 | Aspergillus nidulans FGSC A4 sulphydryl oxidase Sox patent-A1-
Aspergillus niger (386 aa) Pyr_redox_2 50% 22 8 9 11 13 17 17 12 22
ANID_08265 | ANID_08265 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (234
aa) 1 62 20% 2 5 2 0 0 0 0 0 1
ANID_08333 | ANID_08333 | Aspergillus nidulans FGSC A4 PhiA protein (183 aa) 2 57, 107 56% 29 26 24 6 2 1 23 11 61
ANID_08389 | ANID_08389 | Aspergillus nidulans FGSC A4 histidine acid phosphatase (475 aa) His_Phos_2 1 356 14% 0 0 0 0 0 0 1 0 3
ANID_08401 | ANID_08401 | Aspergillus nidulans FGSC A4 beta-xylosidase (764 aa) GH3 4 63, 340, 408, 419 60% 0 0 0 11 18 11 41 101 119
ANID_08432 | ANID_08432 | Aspergillus nidulans FGSC A4 six-hairpin glycosidase (685 aa) 4 267, 283, 309,
552 23% 0 0 2 8 8 4 1 5 1
ANID_08445 | ANID_08445 | Aspergillus nidulans FGSC A4 aminopeptidase Y (504 aa) PA Peptidase_M
28 35% 1 2 7 13 22 10 27 73 86
ANID_08477 | ANID_08477 | Aspergillus nidulans FGSC A4 arabinofuranosidase (547 aa) GH43 1 438 34% 0 0 0 3 13 4 7 12 8
ANID_08484 | ANID_08484 | Aspergillus nidulans FGSC A4 predicted protein (483 aa) HsbA 4% 0 0 0 0 0 2 0 0 0
ANID_08544 | ANID_08544 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (265
aa) 17% 0 0 2 0 0 0 0 0 0
ANID_08546 | ANID_08546 | Aspergillus nidulans FGSC A4 phosphatidylglycerol specific
phospholipase (509 aa)
Phosphoeste
rase 4% 0 0 0 0 0 0 0 1 1
ANID_08602 | ANID_08602 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (438
aa) 7% 1 5 1 2 2 5 1 7 3
ANID_08730 | ANID_08730 | Aspergillus nidulans FGSC A4 N-acetylMuramoyl-L-alanine amidase
(371 aa) 1 29 12% 0 1 2 0 0 0 0 0 0
ANID_08761 | ANID_08761 | Aspergillus nidulans FGSC A4 exo-polygalacturonase (435 aa) GH28 4 113, 199, 292,
297 27% 0 0 0 1 4 1 9 27 35
ANID_08891 | ANID_08891 | Aspergillus nidulans FGSC A4 exopolygalacturonase (441 aa) GH28 2 65, 230 27% 0 0 0 0 0 0 3 7 4
ANID_08908 | ANID_08908 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (279
aa) CE14 7% 0 0 0 0 0 0 2 0 0
ANID_08953 | ANID_08953 | Aspergillus nidulans FGSC A4 alpha-glucosidase B (956 aa) GH31 3 188, 191, 843 39% 22 1 0 0 0 0 21 49 46
ANID_08977 | ANID_08977 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (354
aa) SGL 28% 0 0 0 0 4 0 4 4 4
ANID_09042 | ANID_09042 | Aspergillus nidulans FGSC A4 mutanase (643 aa) GH71 CBM24 16% 0 0 0 17 4 11 5 9 7
ANID_09130 | ANID_09130 | Aspergillus nidulans FGSC A4 cholinesterase (729 aa) CE10 1 79 22% 1 0 1 2 6 0 3 7 4
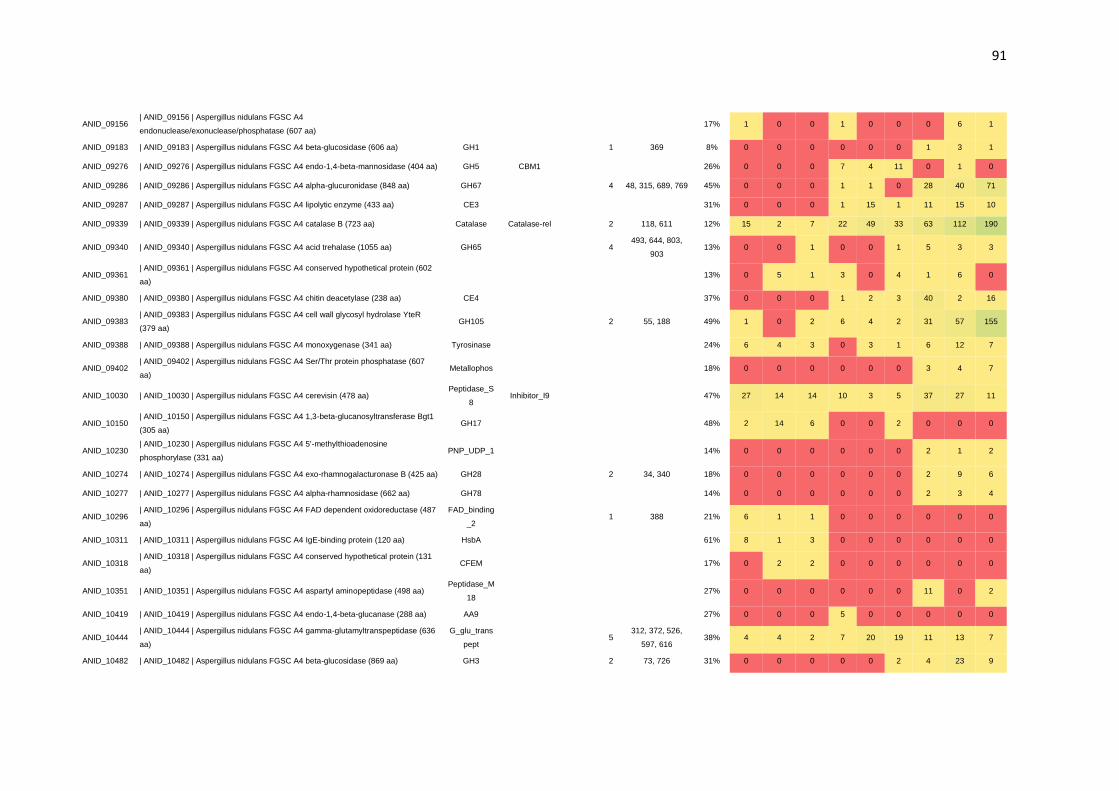
91
ANID_09156 | ANID_09156 | Aspergillus nidulans FGSC A4
endonuclease/exonuclease/phosphatase (607 aa) 17% 1 0 0 1 0 0 0 6 1
ANID_09183 | ANID_09183 | Aspergillus nidulans FGSC A4 beta-glucosidase (606 aa) GH1 1 369 8% 0 0 0 0 0 0 1 3 1
ANID_09276 | ANID_09276 | Aspergillus nidulans FGSC A4 endo-1,4-beta-mannosidase (404 aa) GH5 CBM1 26% 0 0 0 7 4 11 0 1 0
ANID_09286 | ANID_09286 | Aspergillus nidulans FGSC A4 alpha-glucuronidase (848 aa) GH67 4 48, 315, 689, 769 45% 0 0 0 1 1 0 28 40 71
ANID_09287 | ANID_09287 | Aspergillus nidulans FGSC A4 lipolytic enzyme (433 aa) CE3 31% 0 0 0 1 15 1 11 15 10
ANID_09339 | ANID_09339 | Aspergillus nidulans FGSC A4 catalase B (723 aa) Catalase Catalase-rel 2 118, 611 12% 15 2 7 22 49 33 63 112 190
ANID_09340 | ANID_09340 | Aspergillus nidulans FGSC A4 acid trehalase (1055 aa) GH65 4 493, 644, 803,
903 13% 0 0 1 0 0 1 5 3 3
ANID_09361 | ANID_09361 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (602
aa) 13% 0 5 1 3 0 4 1 6 0
ANID_09380 | ANID_09380 | Aspergillus nidulans FGSC A4 chitin deacetylase (238 aa) CE4 37% 0 0 0 1 2 3 40 2 16
ANID_09383 | ANID_09383 | Aspergillus nidulans FGSC A4 cell wall glycosyl hydrolase YteR
(379 aa) GH105 2 55, 188 49% 1 0 2 6 4 2 31 57 155
ANID_09388 | ANID_09388 | Aspergillus nidulans FGSC A4 monoxygenase (341 aa) Tyrosinase 24% 6 4 3 0 3 1 6 12 7
ANID_09402 | ANID_09402 | Aspergillus nidulans FGSC A4 Ser/Thr protein phosphatase (607
aa) Metallophos 18% 0 0 0 0 0 0 3 4 7
ANID_10030 | ANID_10030 | Aspergillus nidulans FGSC A4 cerevisin (478 aa) Peptidase_S
8 Inhibitor_I9 47% 27 14 14 10 3 5 37 27 11
ANID_10150 | ANID_10150 | Aspergillus nidulans FGSC A4 1,3-beta-glucanosyltransferase Bgt1
(305 aa) GH17 48% 2 14 6 0 0 2 0 0 0
ANID_10230 | ANID_10230 | Aspergillus nidulans FGSC A4 5'-methylthioadenosine
phosphorylase (331 aa) PNP_UDP_1 14% 0 0 0 0 0 0 2 1 2
ANID_10274 | ANID_10274 | Aspergillus nidulans FGSC A4 exo-rhamnogalacturonase B (425 aa) GH28 2 34, 340 18% 0 0 0 0 0 0 2 9 6
ANID_10277 | ANID_10277 | Aspergillus nidulans FGSC A4 alpha-rhamnosidase (662 aa) GH78 14% 0 0 0 0 0 0 2 3 4
ANID_10296 | ANID_10296 | Aspergillus nidulans FGSC A4 FAD dependent oxidoreductase (487
aa)
FAD_binding
_2 1 388 21% 6 1 1 0 0 0 0 0 0
ANID_10311 | ANID_10311 | Aspergillus nidulans FGSC A4 IgE-binding protein (120 aa) HsbA 61% 8 1 3 0 0 0 0 0 0
ANID_10318 | ANID_10318 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (131
aa) CFEM 17% 0 2 2 0 0 0 0 0 0
ANID_10351 | ANID_10351 | Aspergillus nidulans FGSC A4 aspartyl aminopeptidase (498 aa) Peptidase_M
18 27% 0 0 0 0 0 0 11 0 2
ANID_10419 | ANID_10419 | Aspergillus nidulans FGSC A4 endo-1,4-beta-glucanase (288 aa) AA9 27% 0 0 0 5 0 0 0 0 0
ANID_10444 | ANID_10444 | Aspergillus nidulans FGSC A4 gamma-glutamyltranspeptidase (636
aa)
G_glu_trans
pept 5
312, 372, 526,
597, 616 38% 4 4 2 7 20 19 11 13 7
ANID_10482 | ANID_10482 | Aspergillus nidulans FGSC A4 beta-glucosidase (869 aa) GH3 2 73, 726 31% 0 0 0 0 0 2 4 23 9

92
ANID_10563 | ANID_10563 | Aspergillus nidulans FGSC A4 alkaline phosphatase (607 aa) Alk_phospha
tase 1 248 46% 9 2 7 8 9 9 18 21 20
ANID_10577 | ANID_10577 | Aspergillus nidulans FGSC A4 dihydrolipoyl dehydrogenase (149
aa)
Pyr_redox_di
m 30% 0 0 0 0 0 0 1 0 0
ANID_10919 | ANID_10919 | Aspergillus nidulans FGSC A4 endo-1,4-beta-xylanase D (414 aa) GH43 CBM6 30% 0 0 0 26 13 36 17 28 30
ANID_10930 | ANID_10930 | Aspergillus nidulans FGSC A4 predicted protein (486 aa) AA7 21% 0 0 0 0 0 0 7 7 9
ANID_11070 | ANID_11070 | Aspergillus nidulans FGSC A4 aspartyl aminopeptidase (520 aa) Peptidase_M
18 29% 0 0 1 1 2 0 4 0 5
ANID_11121 | ANID_11121 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (729
aa)
Abhydrolase
_1
Abhydrolase_
4 1 161 5% 0 0 1 0 0 0 0 0 0
ANID_11143 | ANID_11143 | Aspergillus nidulans FGSC A4 glucoamylase (696 aa) GH15 CBM20 1 428 13% 0 1 0 3 5 2 0 5 4
ANID_11152 | ANID_11152 | Aspergillus nidulans FGSC A4 1,3-beta-glucanosyltransferase (525
aa) GH72 CBM43 2 54, 253 20% 3 2 6 0 0 0 0 0 0
ANID_11698 | ANID_11698 | Aspergillus nidulans FGSC A4 tripeptidyl peptidase SED3 (592 aa) Peptidase_S
8
Pro-
kuma_activ 4% 0 0 0 0 0 0 0 0 2
ANID_11714 | ANID_11714 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (162
aa) DUF3237 1 69 48% 0 0 0 1 3 4 0 0 9
ANID_11778 | ANID_11778 | Aspergillus nidulans FGSC A4 fructan beta-fructosidase (548 aa) GH32 2 403, 465 40% 27 22 20 6 12 8 10 63 66
ANID_11897 | ANID_11897 | Aspergillus nidulans FGSC A4 ribonuclease T2 (261 aa) Ribonucleas
e_T2 31% 2 0 0 2 1 3 1 8 4
ANID_11920 | ANID_11920 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (354
aa) Tyrosinase 27% 5 3 4 0 0 0 0 1 0
ANID_11979 | ANID_11979 | Aspergillus nidulans FGSC A4 extracellular cell wall glucanase
Crf1/allergen Asp F9 (329 aa) GH16 13% 2 1 0 0 0 0 0 0 0
ANID_11981 | ANID_11981 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (498
aa) AA7 1 33 40% 1 1 9 2 10 5 5 4 2
ANID_12097 | ANID_12097 | Aspergillus nidulans FGSC A4 rhamnogalacturonan lyase (660 aa) PL4 1 231 30% 0 0 0 0 0 0 5 9 8
ANID_12193 | ANID_12193 | Aspergillus nidulans FGSC A4 conserved hypothetical protein (327
aa) CE10 1 169 18% 0 0 0 3 0 2 0 0 0
ANID_12420 | ANID_12420 | Aspergillus nidulans FGSC A4 class V chitinase (443 aa) GH18 17% 0 0 0 0 0 0 4 2 1
ANID_12465 | ANID_12465 | Aspergillus nidulans FGSC A4 dihydrolipoamide dehydrogenase
(419 aa) Pyr_redox
Pyr_redox_di
m
Pyr_redo
x_2 24% 0 0 1 0 0 0 7 2 2
¹ Number of N-glycosylated sites confirmed by LC-MS/MS.

93
Table S2. Identified CAZymes and spectrum counts.
Total spectrum count
CAZymes (67)
Accession
Number
N-glyc
sites¹ Position (Asn)²
Domai
n 1
Domain
2
Glucos
e A
Glucos
e B
Glucos
e C
SCB
A
SCB
B
SCB
C
Xyla
n A
Xyla
n B
Xyla
n C
| ANID_00393 | Aspergillus nidulans FGSC A4 glycosyl hydrolase family 76
protein (465 aa) ANID_00393 1 205 GH76 0 0 1 1 1 0 0 2 1
| ANID_00472 | Aspergillus nidulans FGSC A4 endo-1,3-beta-glucanase
Engl1 (908 aa) ANID_00472 2 219, 240 GH81 0 0 0 60 27 33 46 53 48
| ANID_00558 | Aspergillus nidulans FGSC A4 beta glucanosyltransferase
Gel2p (474 aa) ANID_00558 2 313, 341 GH72 5 5 3 4 2 2 0 0 0
| ANID_00787 | Aspergillus nidulans FGSC A4 alpha-1,2-mannosidase
(506 aa) ANID_00787 1 88 GH47 50 37 35 97 125 156 40 85 100
| ANID_00941 | Aspergillus nidulans FGSC A4 alpha-glucosidase (874 aa) ANID_00941 1 713 GH31 6 0 0 2 4 0 2 2 1
| ANID_01320 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (256 aa) ANID_01320 1 63 CE10 1 0 2 2 4 1 15 28 27
| ANID_01433 | Aspergillus nidulans FGSC A4 triacylglycerol lipase (541
aa) ANID_01433 2 374, 381 CE10 0 0 0 0 2 0 12 16 49
| ANID_01502 | Aspergillus nidulans FGSC A4 N-acetylglucosaminidase
(604 aa) ANID_01502 1 503 GH20 3 0 0 3 10 12 109 69 207
| ANID_01772 | Aspergillus nidulans FGSC A4 feruloyl esterase (528 aa) ANID_01772 3 86, 139, 356 CE1 Tannase 1 0 8 9 8 12 13 33 33
| ANID_01870 | Aspergillus nidulans FGSC A4 xylanase D (308 aa) ANID_01870 1 87 GH43 2 2 3 1 1 3 4 3 5
| ANID_02017 | Aspergillus nidulans FGSC A4 alpha-glucosidase AgdA
(993 aa) ANID_02017 1 432 GH31 0 1 2 4 6 5 9 3 3
| ANID_02018 | Aspergillus nidulans FGSC A4 alpha-amylase AmyA (492
aa) ANID_02018 2 39, 390 GH13 0 0 0 1 0 2 0 1 2
| ANID_02325 | Aspergillus nidulans FGSC A4 glycosyl hydrolase, family
92 protein (755 aa) ANID_02325 4
105, 112, 118,
261 GH92 0 0 0 1 3 0 7 11 17
| ANID_02359 | Aspergillus nidulans FGSC A4 beta-xylosidase (801 aa) ANID_02359 3 231, 673, 695 GH3 0 0 0 18 30 33 9 15 15
| ANID_02387 | Aspergillus nidulans FGSC A4 FAD binding domain-
containing protein (503 aa) ANID_02387 1 260 AA7 5 4 2 0 2 3 1 9 13
| ANID_02388 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (436 aa) ANID_02388 1 93 AA9 13 8 18 10 11 17 4 16 27

94
| ANID_02395 | Aspergillus nidulans FGSC A4 hydrolase (614 aa) ANID_02395 1 293 GH2 0 1 0 17 26 15 14 26 27
| ANID_02574 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (517 aa) ANID_02574 2 212, 330 AA7 0 0 0 1 0 0 2 11 16
| ANID_02648 | Aspergillus nidulans FGSC A4 isoamyl alcohol oxidase
(567 aa) ANID_02648 1 47 AA7 4 1 0 1 1 0 2 11 6
| ANID_02828 | Aspergillus nidulans FGSC A4 beta-glucosidase (738 aa) ANID_02828 2 225, 365 GH3 0 1 7 83 81 74 61 61 86
| ANID_03229 | Aspergillus nidulans FGSC A4 choline dehydrogenase (612
aa) ANID_03229 2 187, 272 AA3 0 0 0 0 0 0 5 1 6
| ANID_03351 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (487 aa) ANID_03351 3 48, 259, 275 AA7 4 0 6 2 5 3 1 5 3
| ANID_04102 | Aspergillus nidulans FGSC A4 beta-glucosidase (854 aa) ANID_04102 4 62, 491, 642,
713 GH3 1 0 0 11 20 8 18 22 33
| ANID_05176 | Aspergillus nidulans FGSC A4 1,4-beta-D-glucan-
cellobiohydrolyase (447 aa) ANID_05176 1 284 GH7 0 0 0 565 263 270 13 1 1
| ANID_05463 | Aspergillus nidulans FGSC A4 starch binding domain-
containing protein (386 aa) ANID_05463 1 364
CBM2
0 10 8 1 0 0 1 3 2 1
| ANID_06093 | Aspergillus nidulans FGSC A4 acetylxylan esterase (307
aa) ANID_06093 1 263 CE1 0 0 0 25 3 25 7 2 6
| ANID_06405 | Aspergillus nidulans FGSC A4 secreted glycosyl hydrolase
(247 aa) ANID_06405 1 183 GH25 0 10 1 0 0 0 0 1 0
| ANID_06428 | Aspergillus nidulans FGSC A4 fungal cellulose binding
domain-containing protein (232 aa) ANID_06428 1 69, 143 AA9 0 0 0 13 38 16 0 0 0
| ANID_06438 | Aspergillus nidulans FGSC A4 dipeptidyl-peptidase IV (774
aa) ANID_06438 2 496, 671 CE1 0 0 0 4 3 0 5 11 9
| ANID_06620 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (389 aa) ANID_06620 1 166 GH16 20 10 13 2 4 4 7 2 6
| ANID_06635 | Aspergillus nidulans FGSC A4 laccase (610 aa) ANID_06635 1 403 AA1
Cu-
oxidase
(3x)
0 0 0 0 0 0 11 7 22
| ANID_07151 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (879 aa) ANID_07151 3 36, 58, 586 GH78 CBM67 0 0 0 0 0 0 3 12 11
| ANID_07152 | Aspergillus nidulans FGSC A4 alpha-galactosidase (641
aa) ANID_07152 1 532 GH27 0 0 0 16 21 18 5 8 8

95
| ANID_07230 | Aspergillus nidulans FGSC A4 cellobiose dehydrogenase
(797 aa) ANID_07230 5
132, 299, 308,
620, 709 AA8 0 0 0 45 190 40 0 0 0
| ANID_07269 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (497 aa) ANID_07269 2 133, 460 AA7 27 10 21 19 25 19 21 29 32
| ANID_07275 | Aspergillus nidulans FGSC A4 xylosidase/glycosyl
hydrolase (517 aa) ANID_07275 2 40, 382 GH43 0 0 0 24 7 24 4 7 4
| ANID_07307 | Aspergillus nidulans FGSC A4 DUF1237 domain-
containing protein (538 aa) ANID_07307 1 68 GH125 0 0 4 2 8 8 5 4 8
| ANID_07345 | Aspergillus nidulans FGSC A4 alpha/beta-glucosidase (895
aa) ANID_07345 1 536 GH31 0 0 0 0 1 0 0 0 1
| ANID_07389 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (581 aa) ANID_07389 2 55, 472 AA1
Cu-
oxidase
(3x)
0 0 0 4 14 7 0 2 3
| ANID_07396 | Aspergillus nidulans FGSC A4 beta-glucosidase (773 aa) ANID_07396 3 259, 438, 586 GH3 0 0 0 2 7 6 3 4 5
| ANID_07401 | Aspergillus nidulans FGSC A4 endo-1,4-beta-xylanase
(382 aa) ANID_07401 1 123 GH10 CBM1 0 0 0 39 65 56 26 5 6
| ANID_07511 | Aspergillus nidulans FGSC A4 1,3-beta-
glucanosyltransferase (540 aa) ANID_07511 2 36, 415 GH72 CBM43 9 7 4 12 7 10 3 16 9
| ANID_07598 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (257 aa) ANID_07598 1 72 GH131 0 0 0 36 17 33 1 0 0
| ANID_07657 | Aspergillus nidulans FGSC A4 glycolipid-anchored surface
protein 5 (456 aa) ANID_07657 2 249, 337 GH72 120 119 106 20 21 22 27 41 32
| ANID_07781 | Aspergillus nidulans FGSC A4 arabinosidase (341 aa) ANID_07781 1 215 GH43 2 0 0 3 2 2 4 8 4
| ANID_07812 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (640 aa) ANID_07812 2 444, 501 AA3 0 0 0 10 5 8 0 0 1
| ANID_07998 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (623 aa) ANID_07998 1 480 AA3 0 1 3 0 0 3 0 1 1
| ANID_08007 | Aspergillus nidulans FGSC A4 endo-alpha-1,5-arabinanase
(321 aa) ANID_08007 1 126 GH43 2 5 5 8 10 20 36 53 29
| ANID_08068 | Aspergillus nidulans FGSC A4 extracellular endoglucanase
(573 aa) ANID_08068 1 275 GH5 CBM46 0 0 0 5 16 13 0 0 0
| ANID_08401 | Aspergillus nidulans FGSC A4 beta-xylosidase (764 aa) ANID_08401 4 63, 340, 408,
419 GH3 0 0 0 11 18 11 41 101 119

96
| ANID_08477 | Aspergillus nidulans FGSC A4 arabinofuranosidase (547
aa) ANID_08477 1 438 GH43 0 0 0 3 13 4 7 12 8
| ANID_08761 | Aspergillus nidulans FGSC A4 exo-polygalacturonase (435
aa) ANID_08761 4
113, 199, 292,
297 GH28 0 0 0 1 4 1 9 27 35
| ANID_08891 | Aspergillus nidulans FGSC A4 exopolygalacturonase (441
aa) ANID_08891 2 65, 230 GH28 0 0 0 0 0 0 3 7 4
| ANID_08953 | Aspergillus nidulans FGSC A4 alpha-glucosidase B (956
aa) ANID_08953 3 188, 191, 843 GH31 22 1 0 0 0 0 21 49 46
| ANID_09130 | Aspergillus nidulans FGSC A4 cholinesterase (729 aa) ANID_09130 1 79 CE10 1 0 1 2 6 0 3 7 4
| ANID_09183 | Aspergillus nidulans FGSC A4 beta-glucosidase (606 aa) ANID_09183 1 369 GH1 0 0 0 0 0 0 1 3 1
| ANID_09286 | Aspergillus nidulans FGSC A4 alpha-glucuronidase (848
aa) ANID_09286 4
48, 315, 689,
769 GH67 0 0 0 1 1 0 28 40 71
| ANID_09340 | Aspergillus nidulans FGSC A4 acid trehalase (1055 aa) ANID_09340 4 493, 644, 803,
903 GH65 0 0 1 0 0 1 5 3 3
| ANID_09383 | Aspergillus nidulans FGSC A4 cell wall glycosyl hydrolase
YteR (379 aa) ANID_09383 2 55, 188 GH105 1 0 2 6 4 2 31 57 155
| ANID_10274 | Aspergillus nidulans FGSC A4 exo-rhamnogalacturonase B
(425 aa) ANID_10274 2 34, 340 GH28 0 0 0 0 0 0 2 9 6
| ANID_10482 | Aspergillus nidulans FGSC A4 beta-glucosidase (869 aa) ANID_10482 2 73, 726 GH3 0 0 0 0 0 2 4 23 9
| ANID_11143 | Aspergillus nidulans FGSC A4 glucoamylase (696 aa) ANID_11143 1 428 GH15 CBM20 0 1 0 3 5 2 0 5 4
| ANID_11152 | Aspergillus nidulans FGSC A4 1,3-beta-
glucanosyltransferase (525 aa) ANID_11152 2 54, 253 GH72 CBM43 3 2 6 0 0 0 0 0 0
| ANID_11778 | Aspergillus nidulans FGSC A4 fructan beta-fructosidase
(548 aa) ANID_11778 2 403, 465 GH32 27 22 20 6 12 8 10 63 66
| ANID_11981 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (498 aa) ANID_11981 1 33 AA7 1 1 9 2 10 5 5 4 2
| ANID_12097 | Aspergillus nidulans FGSC A4 rhamnogalacturonan lyase
(660 aa) ANID_12097 1 231 PL4 0 0 0 0 0 0 5 9 8
| ANID_12193 | Aspergillus nidulans FGSC A4 conserved hypothetical
protein (327 aa) ANID_12193 1 169 CE10 0 0 0 3 0 2 0 0 0
¹ Total number of N-glycosylated sites confirmed by LC-MS/MS.
² Position of N-glycosylated asparagine confirmed by LC-MS/MS.

97
Table S3. Identified motifs on N-glycosylated sites.
Modification Motif1 Motif Score Surrounding Sequence Accession N-glyc Site Best Ambiguity score2 Localization
Probability2
HexNAc ......n.S.... 25.13 WTFVWRNASDIST ANID_08730 N29 1000 100%
HexNAc ......n.S.... 25.13 WYQLVANSSDVLF ANID_08761 N199 1000 100%
HexNAc ......n.S.... 25.13 PYTVAKNSSDYGS ANID_07396 N586 1000 100%
HexNAc ......n.S.... 25.13 EQSFLYNASAPSA ANID_03351 N275 40.35 100%
HexNAc ......n.S.... 25.13 VRKNGVNSSNPIE ANID_00472 N219 22.45 99%
HexNAc ......n.S.... 25.13 WQPSSSNASAVTV ANID_09340 N903 64.95 100%
HexNAc ......n.S.... 25.13 ANTFRVNSSDSVL ANID_07151 N58 1000 100%
HexNAc ......n.S.... 25.13 QTYASHNGSLAEQ ANID_11143 N428 1000 100%
HexNAc ......n.S.... 25.13 EKHQPGNLSEATA ANID_07812 N444 1000 100%
HexNAc ......n.S.... 25.13 NDYPTGNYSLWLH ANID_08086 N73 28.13 100%
HexNAc ......n.S.... 25.13 CSYNRVNNSYACQ ANID_07396 N259 33.98 100%
HexNAc ......n.S.... 25.13 INAYSTNASAEPK ANID_08891 N230 61.26 100%
HexNAc ......n.S.... 25.13 VWESGPNRSYTVP ANID_05463 N364 1000 100%
HexNAc ......n.S.... 25.13 MNYGGGNTSADVR ANID_06635 N403 22.07 100%
HexNAc ......n.S.... 25.13 PNSATYNSSGSSY ANID_03351 N48 48.91 100%
HexNAc ......n.S.... 25.13 TEYPVGNVSVPGV ANID_08432 N552 1000 100%
HexNAc ......n.S.... 25.13 DIESWNNHSRLGN ANID_02359 N231 27.96 100%
HexNAc ......n.S.... 25.13 VHGHFSNGSTSGD ANID_12097 N231 21.91 100%

98
HexNAc ......n.S.... 25.13 KNVLVYNISMYNA ANID_08761 N292 14.19 98%
HexNAc ......n.S.... 25.13 ADAAVANSSKTYP ANID_10482 N726 121.98 100%
HexNAc ......n.S.... 25.13 WAYPKANVSMAEH ANID_07230 N709 118.88 100%
HexNAc ......n.S.... 25.13 TNLTIRNVSSVSL ANID_04390 N209 1000 100%
HexNAc ......n.S.... 25.13 YAPLHTNSSYVGG ANID_02237 N494 1000 100%
HexNAc ......n.S.... 25.13 RYSVLVNGSSPGP ANID_07389 N55 1000 100%
HexNAc ......n.S.... 25.13 DRISASNASQTSV ANID_00558 N341 1000 100%
HexNAc ......n.S.... 25.13 IECRTFNSSAVEQ ANID_07307 N68 1000 100%
HexNAc ......n.S.... 25.13 FTPENVNLSTHRL ANID_10444 N372 26.31 100%
HexNAc ......n.S.... 25.13 DRRASGNTSFDET ANID_11778 N465 1000 100%
HexNAc ......n.S.... 25.13 AATAHINSSTSGS ANID_07152 N532 1000 100%
HexNAc ......n.S.... 25.13 TATIHLNISSMAS ANID_07275 N382 1000 100%
HexNAc ......n.S.... 25.13 NSYRMGNTSFYGP ANID_05176 N284 1000 100%
HexNAc ......n.S.... 25.13 EEYALPNISYFEL ANID_06438 N496 1000 100%
HexNAc ......n.S.... 25.13 GYTTGDNASGPRN ANID_08333 N107 1000 100%
HexNAc ......n.S.... 25.13 MAVTTSNASEITL ANID_01772 N86 1000 100%
HexNAc ......n.S.... 25.13 RTEEFANYSVPVF ANID_11152 N253 34.87 100%
HexNAc ......n.S.... 25.13 NEAGLWNRSTFQY ANID_07151 N586 31.78 100%
HexNAc ......n.S.... 25.13 FLRIPANVSTVVD ANID_04055 N119 1000 100%
HexNAc ......n.S.... 25.13 VATFAANISDPKT ANID_02387 N260 1000 100%
HexNAc ......n.S.... 25.13 VKAAGYNASNTTL ANID_07231 N309 1000 100%

99
HexNAc ......n.S.... 25.13 HNVHRANHSSPDV ANID_01058 N166 1000 100%
HexNAc ......n.S.... 25.13 KHPAILNFSGIPW ANID_01870 N87 64.98 100%
HexNAc ......n.S.... 25.13 YNISMYNASDMAR ANID_08761 N297 55.37 100%
HexNAc ......n.T.... 36.45 FMLNTTNYTRTIY ANID_08953 N191 30.77 100%
HexNAc ......n.T.... 36.45 RFATLYNLTSELQ ANID_02574 N330 1000 100%
HexNAc ......n.T.... 36.45 KALQAANGTMTLE ANID_10444 N312 1000 100%
HexNAc ......n.T.... 36.45 ILEIVANKTGYVN ANID_04055 N315 120.62 100%
HexNAc ......n.T.... 36.45 GGDGLYNATANEI ANID_07389 N472 47.31 100%
HexNAc ......n.T.... 36.45 PGVEKQNGTGLDY ANID_01318 N229 74.39 100%
HexNAc ......n.T.... 36.45 YPTVDMNSTDFTE ANID_12193 N169 1000 100%
HexNAc ......n.T.... 36.45 PGPFTYNRTLADG ANID_09286 N315 139.62 100%
HexNAc ......n.T.... 36.45 RALKTANITFGAL ANID_01218 N186 1000 100%
HexNAc ......n.T.... 36.45 RIVNANNNTNVEL ANID_02574 N212 14.02 92%
HexNAc ......n.T.... 36.45 YSDPTWNATKWTI ANID_01772 N356 1000 100%
HexNAc ......n.T.... 36.45 LDREGTNSTTYLN ANID_01320 N63 54.91 100%
HexNAc ......n.T.... 36.45 FYVNNENVTYWDP ANID_08432 N283 40 100%
HexNAc ......n.T.... 36.45 SPVTEKNSTSVNA ANID_00941 N713 81.69 100%
HexNAc ......n.T.... 36.45 PSIGLWNGTMKPD ANID_07230 N299 1000 100%
HexNAc ......n.T.... 36.45 FDGTVSNWTQSAY ANID_07151 N36 87.68 100%
HexNAc ......n.T.... 36.45 GIPYYTNGTKDYY ANID_11121 N161 57.3 100%
HexNAc ......n.T.... 36.45 TSTGGANITSLSA ANID_04390 N78 1000 100%

100
HexNAc ......n.T.... 36.45 YSGPTENSTLTTG ANID_08086 N43 59.35 100%
HexNAc ......n.T.... 36.45 STRPFPNATRYWR ANID_01433 N381 1000 100%
HexNAc ......n.T.... 36.45 AVWQIRNETEVNR ANID_05939 N215 78.33 100%
HexNAc ......n.T.... 36.45 SEYKNPNSTLDEA ANID_07598 N72 18.58 99%
HexNAc ......n.T.... 36.45 DRMRALNGTTMYT ANID_00231 N284 23.83 100%
HexNAc ......n.T.... 36.45 AGLPKQNATCEGT ANID_08333 N57 1000 100%
HexNAc ......n.T.... 36.45 AAGSLSNKTMNTD ANID_07345 N536 107.16 100%
HexNAc ......n.T.... 36.45 TINPALNHTTIGR ANID_02360 N166 33.81 100%
HexNAc ......n.T.... 36.45 GTQPTYNYTSPFP ANID_04055 N382 1000 100%
HexNAc ......n.T.... 36.45 MTGRFPNTTVTYD ANID_03229 N187 101.26 100%
HexNAc ......n.T.... 36.45 ILSLPANSTRNLG ANID_11778 N403 1000 100%
HexNAc ......n.T.... 36.45 RGTVTINTTNTAD ANID_07812 N501 28.73 100%
HexNAc ......n.T.... 36.45 QQDYTTNSTSSSD ANID_11152 N54 1000 100%
HexNAc ......n.T.... 36.45 AIVTHPNGTTFFI ANID_10444 N526 1000 100%
HexNAc ......n.T.... 36.45 GRIQFTNDTDYWQ ANID_08761 N113 127.03 100%
HexNAc ......n.T.... 36.45 TIRSSHNDTDDVS ANID_08891 N65 1000 100%
HexNAc ......n.T.... 36.45 RRRQIKNFTHFDD ANID_04055 N186 1000 100%
HexNAc ......n.T.... 36.45 PRFSGNNGTGEEG ANID_06620 N166 11.06 93%
HexNAc ......n.T.... 36.45 MKPDWLNGTDLTR ANID_07230 N308 1000 100%
HexNAc ......n.T.... 36.45 QSLKTTNLTVYLH ANID_03229 N272 1000 100%
HexNAc ......n.T.... 36.45 FDQAIGNSTWRLF ANID_09286 N769 98.53 100%

101
HexNAc ......n.T.... 36.45 SPQEVQNDTPVAG ANID_06093 N263 55.21 100%
HexNAc ......n.T.... 36.45 TYTGSKNATYIDP ANID_09340 N493 1000 100%
HexNAc ......n.T.... 36.45 WALDKVNKTASES ANID_04102 N491 78.02 100%
HexNAc ......n.T.... 36.45 EGDGGYNKTGGAN ANID_07657 N337 189.46 100%
HexNAc ......n.T.... 36.45 NYDFTLNLTNAQA ANID_01502 N503 56.34 100%
HexNAc ......n.T.... 36.45 RANVQLNHTVTSI ANID_03057 N292 70.03 100%
HexNAc ......n.T.... 36.45 VQQYLWNTTFNHF ANID_08432 N267 47.31 100%
HexNAc ......n.T.... 36.45 TSGSWTNTTLTAA ANID_07401 N123 34.87 100%
HexNAc ......n.T.... 36.45 ASVTDTNTTKAIA ANID_02954 N300 1000 100%
HexNAc ......n.T.... 36.45 RSDLYKNGTLYVG ANID_07396 N438 14.66 92%
HexNAc ......n.T.... 36.45 RCHFLPNTTDGSC ANID_10563 N248 1000 100%
HexNAc ......n.T.... 36.45 SEVESLNETIKNY ANID_05939 N163 1000 100%
HexNAc ......n.T.... 36.45 FRHYCDNITSTEP ANID_07231 N282 52.1 100%
HexNAc ......n.T.... 36.45 TALVYVNTTAGPA ANID_02359 N695 1000 100%
HexNAc ......n.T.... 36.45 QKADAVNGTYGAY ANID_07511 N415 1000 100%
HexNAc ......n.T.... 36.45 AKWNALNSTVGGR ANID_02648 N47 54.7 100%
HexNAc ......n.T.... 36.45 CSYNKINGTWACE ANID_02828 N225 181.16 100%
HexNAc ......n.T.... 36.45 YYLQDVNSTRPFP ANID_01433 N374 1000 100%
HexNAc ......n.T.... 36.45 YKIVSTNSTVDHF ANID_02360 N112 1000 100%
HexNAc ......n.T.... 36.45 SPNLQPNWTDHGA ANID_08007 N126 35.13 100%
HexNAc ......n.T.... 36.45 WDQKVKNFTGYGL ANID_07657 N249 123.87 100%
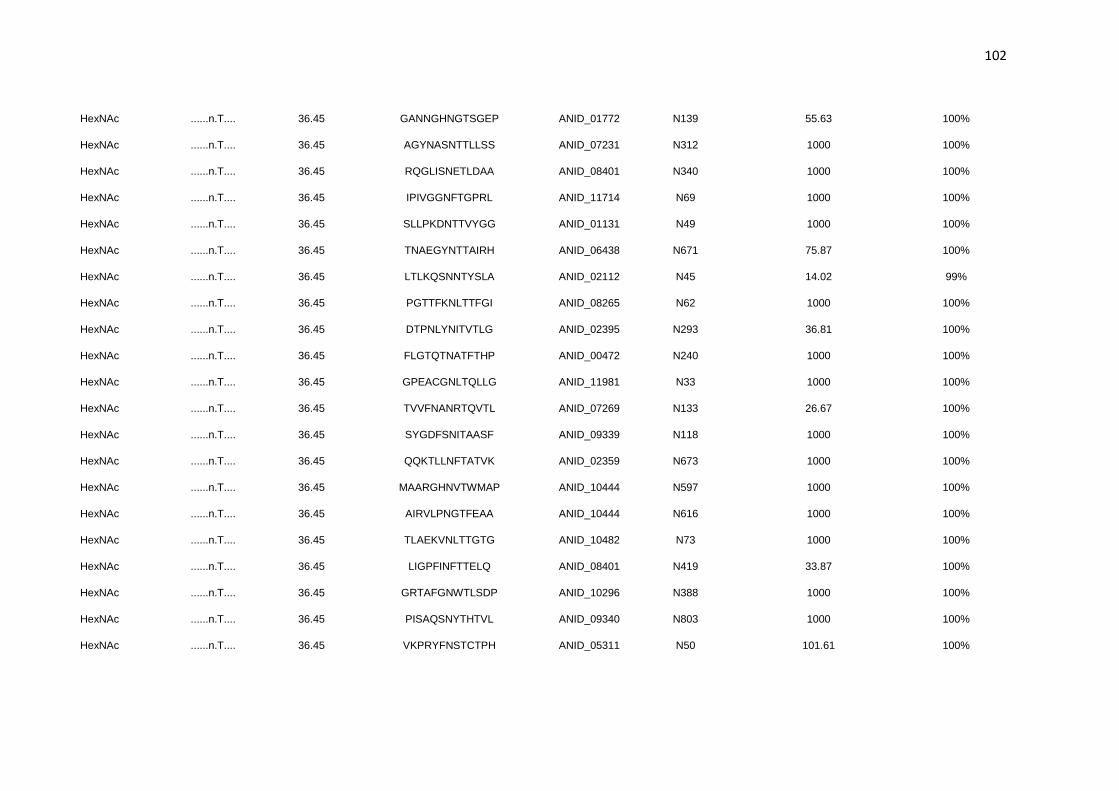
102
HexNAc ......n.T.... 36.45 GANNGHNGTSGEP ANID_01772 N139 55.63 100%
HexNAc ......n.T.... 36.45 AGYNASNTTLLSS ANID_07231 N312 1000 100%
HexNAc ......n.T.... 36.45 RQGLISNETLDAA ANID_08401 N340 1000 100%
HexNAc ......n.T.... 36.45 IPIVGGNFTGPRL ANID_11714 N69 1000 100%
HexNAc ......n.T.... 36.45 SLLPKDNTTVYGG ANID_01131 N49 1000 100%
HexNAc ......n.T.... 36.45 TNAEGYNTTAIRH ANID_06438 N671 75.87 100%
HexNAc ......n.T.... 36.45 LTLKQSNNTYSLA ANID_02112 N45 14.02 99%
HexNAc ......n.T.... 36.45 PGTTFKNLTTFGI ANID_08265 N62 1000 100%
HexNAc ......n.T.... 36.45 DTPNLYNITVTLG ANID_02395 N293 36.81 100%
HexNAc ......n.T.... 36.45 FLGTQTNATFTHP ANID_00472 N240 1000 100%
HexNAc ......n.T.... 36.45 GPEACGNLTQLLG ANID_11981 N33 1000 100%
HexNAc ......n.T.... 36.45 TVVFNANRTQVTL ANID_07269 N133 26.67 100%
HexNAc ......n.T.... 36.45 SYGDFSNITAASF ANID_09339 N118 1000 100%
HexNAc ......n.T.... 36.45 QQKTLLNFTATVK ANID_02359 N673 1000 100%
HexNAc ......n.T.... 36.45 MAARGHNVTWMAP ANID_10444 N597 1000 100%
HexNAc ......n.T.... 36.45 AIRVLPNGTFEAA ANID_10444 N616 1000 100%
HexNAc ......n.T.... 36.45 TLAEKVNLTTGTG ANID_10482 N73 1000 100%
HexNAc ......n.T.... 36.45 LIGPFINFTTELQ ANID_08401 N419 33.87 100%
HexNAc ......n.T.... 36.45 GRTAFGNWTLSDP ANID_10296 N388 1000 100%
HexNAc ......n.T.... 36.45 PISAQSNYTHTVL ANID_09340 N803 1000 100%
HexNAc ......n.T.... 36.45 VKPRYFNSTCTPH ANID_05311 N50 101.61 100%

103
HexNAc ......n.T.... 36.45 PLYPANNATALLP ANID_07231 N294 11.1 96%
HexNAc ......n.T.... 36.45 TLPLKANGTLALI ANID_08401 N408 31.82 100%
HexNAc ......n.T.... 36.45 TLDEKVNLTTGTG ANID_04102 N62 1000 100%
HexNAc ......n.T.... 36.45 AFWKFKNSTVDPY ANID_03351 N259 83.51 100%
HexNAc ......n.T.... 36.45 ITNPLSNITVARN ANID_02325 N112 11.72 95%
HexNAc ......n.T.... 36.45 DGVTAQNGTGLDY ANID_05311 N228 24.78 100%
HexNAc ......n.T.... 36.45 TAIMMRNATIVNQ ANID_00787 N88 32.32 100%
HexNAc ......n.T.... 36.45 IDIIRVNDTFYYS ANID_07275 N40 43 100%
HexNAc ......n.T.... 36.45 ASAPAANRTELAH ANID_09130 N79 1000 100%
HexNAc ......n.T.... 36.45 LEYTTMNGTAAVK ANID_09340 N644 64.17 100%
HexNAc ......n.T.... 36.45 TLEEKINNTGHEA ANID_08401 N63 17.08 100%
HexNAc ......n.T.... 36.45 PSVYSGNATLTQI ANID_07230 N132 1000 100%
HexNAc ......n.T.... 36.45 KLFYSNNGTEFFI ANID_07511 N36 13.8 96%
HexNAc ......n.T.... 36.45 DASTNGNATETLP ANID_06428 N69 42.28 100%
HexNAc ......n.T.... 36.45 RDEVINNRTFVEG ANID_03262 N305 20 99%
HexNAc ......n.T.... 36.45 DGAAVKNGTLLED ANID_03592 N235 63.65 100%
HexNAc ......n.T.... 36.45 NACPSLNYTLGGG ANID_02112 N220 49.8 100%
HexNAc ......n.T.... 36.45 QVTGSDNVTRQLQ ANID_07230 N620 147.98 100%
HexNAc ......n.T.... 36.45 TYINLGNETAIDL ANID_07781 N215 14.08 96%
HexNAc ......n.T.... 36.45 SDSFMLNTTNYTR ANID_08953 N188 20.37 99%
HexNAc ......n.T.... 36.45 KSQSCANATDFVS ANID_06535 N51 1000 100%

104
HexNAc ......n.T.... 36.45 YMKPVANATSDNN ANID_02388 N93 71.64 100%
HexNAc ......n.T.... 36.45 FIYPWINSTDLKE ANID_04102 N713 1000 100%
HexNAc ......n.T.... 36.45 RHFDRTNQTPIYE ANID_04102 N642 1000 100%
HexNAc ......n.T.... 36.45 NITVARNGTDEAE ANID_02325 N118 1000 100%
HexNAc ......n.T.... 36.45 QRIESANTTTALR ANID_08953 N843 1000 100%
HexNAc ......n.T.... 36.45 FAGPLANQTNLAL ANID_04809 N506 15.26 97%
HexNAc ......n.T.... 36.45 GGVFSFNDTEVIS ANID_02325 N261 1000 100%
HexNAc ......n.T.T.. 44.96 FMLNTTNYTRTIY ANID_08953 N191 30.77 100%
HexNAc ......n.T.T.. 44.96 KALQAANGTMTLE ANID_10444 N312 1000 100%
HexNAc ......n.T.T.. 44.96 YSGPTENSTLTTG ANID_08086 N43 59.35 100%
HexNAc ......n.T.T.. 44.96 MTGRFPNTTVTYD ANID_03229 N187 101.26 100%
HexNAc ......n.T.T.. 44.96 RGTVTINTTNTAD ANID_07812 N501 28.73 100%
HexNAc ......n.T.T.. 44.96 RANVQLNHTVTSI ANID_03057 N292 70.03 100%
HexNAc ......n.T.T.. 44.96 TSGSWTNTTLTAA ANID_07401 N123 34.87 100%
HexNAc ......n.T.T.. 44.96 FRHYCDNITSTEP ANID_07231 N282 52.1 100%
HexNAc ......n.T.T.. 44.96 DTPNLYNITVTLG ANID_02395 N293 36.81 100%
HexNAc ......n.T.T.. 44.96 FLGTQTNATFTHP ANID_00472 N240 1000 100%
HexNAc ......n.T.T.. 44.96 QQKTLLNFTATVK ANID_02359 N673 1000 100%
HexNAc ......n.T.T.. 44.96 PISAQSNYTHTVL ANID_09340 N803 1000 100%
HexNAc ......n.T.T.. 44.96 VKPRYFNSTCTPH ANID_05311 N50 101.61 100%
HexNAc ......n.T.T.. 44.96 PSVYSGNATLTQI ANID_07230 N132 1000 100%

105
HexNAc ......n.T.T.. 44.96 DASTNGNATETLP ANID_06428 N69 42.28 100%
HexNAc ......nGT.... 62.11 KALQAANGTMTLE ANID_10444 N312 1000 100%
HexNAc ......nGT.... 62.11 PGVEKQNGTGLDY ANID_01318 N229 74.39 100%
HexNAc ......nGT.... 62.11 PSIGLWNGTMKPD ANID_07230 N299 1000 100%
HexNAc ......nGT.... 62.11 GIPYYTNGTKDYY ANID_11121 N161 57.3 100%
HexNAc ......nGT.... 62.11 DRMRALNGTTMYT ANID_00231 N284 23.83 100%
HexNAc ......nGT.... 62.11 AIVTHPNGTTFFI ANID_10444 N526 1000 100%
HexNAc ......nGT.... 62.11 PRFSGNNGTGEEG ANID_06620 N166 11.06 93%
HexNAc ......nGT.... 62.11 MKPDWLNGTDLTR ANID_07230 N308 1000 100%
HexNAc ......nGT.... 62.11 RSDLYKNGTLYVG ANID_07396 N438 14.66 92%
HexNAc ......nGT.... 62.11 QKADAVNGTYGAY ANID_07511 N415 1000 100%
HexNAc ......nGT.... 62.11 CSYNKINGTWACE ANID_02828 N225 181.16 100%
HexNAc ......nGT.... 62.11 GANNGHNGTSGEP ANID_01772 N139 55.63 100%
HexNAc ......nGT.... 62.11 AIRVLPNGTFEAA ANID_10444 N616 1000 100%
HexNAc ......nGT.... 62.11 TLPLKANGTLALI ANID_08401 N408 31.82 100%
HexNAc ......nGT.... 62.11 DGVTAQNGTGLDY ANID_05311 N228 24.78 100%
HexNAc ......nGT.... 62.11 LEYTTMNGTAAVK ANID_09340 N644 64.17 100%
HexNAc ......nGT.... 62.11 KLFYSNNGTEFFI ANID_07511 N36 13.8 96%
HexNAc ......nGT.... 62.11 DGAAVKNGTLLED ANID_03592 N235 63.65 100%
HexNAc ......nGT.... 62.11 NITVARNGTDEAE ANID_02325 N118 1000 100%
HexNAc ......nST.... 30.92 YPTVDMNSTDFTE ANID_12193 N169 1000 100%

106
HexNAc ......nST.... 30.92 LDREGTNSTTYLN ANID_01320 N63 54.91 100%
HexNAc ......nST.... 30.92 SPVTEKNSTSVNA ANID_00941 N713 81.69 100%
HexNAc ......nST.... 30.92 YSGPTENSTLTTG ANID_08086 N43 59.35 100%
HexNAc ......nST.... 30.92 SEYKNPNSTLDEA ANID_07598 N72 18.58 99%
HexNAc ......nST.... 30.92 ILSLPANSTRNLG ANID_11778 N403 1000 100%
HexNAc ......nST.... 30.92 QQDYTTNSTSSSD ANID_11152 N54 1000 100%
HexNAc ......nST.... 30.92 FDQAIGNSTWRLF ANID_09286 N769 98.53 100%
HexNAc ......nST.... 30.92 AKWNALNSTVGGR ANID_02648 N47 54.7 100%
HexNAc ......nST.... 30.92 YYLQDVNSTRPFP ANID_01433 N374 1000 100%
HexNAc ......nST.... 30.92 YKIVSTNSTVDHF ANID_02360 N112 1000 100%
HexNAc ......nST.... 30.92 VKPRYFNSTCTPH ANID_05311 N50 101.61 100%
HexNAc ......nST.... 30.92 AFWKFKNSTVDPY ANID_03351 N259 83.51 100%
HexNAc ......nST.... 30.92 FIYPWINSTDLKE ANID_04102 N713 1000 100%
HexNAc ......nTT.... 45.41 MTGRFPNTTVTYD ANID_03229 N187 101.26 100%
HexNAc ......nTT.... 45.41 RGTVTINTTNTAD ANID_07812 N501 28.73 100%
HexNAc ......nTT.... 45.41 VQQYLWNTTFNHF ANID_08432 N267 47.31 100%
HexNAc ......nTT.... 45.41 TSGSWTNTTLTAA ANID_07401 N123 34.87 100%
HexNAc ......nTT.... 45.41 ASVTDTNTTKAIA ANID_02954 N300 1000 100%
HexNAc ......nTT.... 45.41 RCHFLPNTTDGSC ANID_10563 N248 1000 100%
HexNAc ......nTT.... 45.41 TALVYVNTTAGPA ANID_02359 N695 1000 100%
HexNAc ......nTT.... 45.41 AGYNASNTTLLSS ANID_07231 N312 1000 100%

107
HexNAc ......nTT.... 45.41 SLLPKDNTTVYGG ANID_01131 N49 1000 100%
HexNAc ......nTT.... 45.41 TNAEGYNTTAIRH ANID_06438 N671 75.87 100%
HexNAc ......nTT.... 45.41 SDSFMLNTTNYTR ANID_08953 N188 20.37 99%
HexNAc ......nTT.... 45.41 QRIESANTTTALR ANID_08953 N843 1000 100%
HexNAc ...P..n.T.... 56.65 YSDPTWNATKWTI ANID_01772 N356 1000 100%
HexNAc ...P..n.T.... 56.65 YSGPTENSTLTTG ANID_08086 N43 59.35 100%
HexNAc ...P..n.T.... 56.65 STRPFPNATRYWR ANID_01433 N381 1000 100%
HexNAc ...P..n.T.... 56.65 AGLPKQNATCEGT ANID_08333 N57 1000 100%
HexNAc ...P..n.T.... 56.65 TINPALNHTTIGR ANID_02360 N166 33.81 100%
HexNAc ...P..n.T.... 56.65 GTQPTYNYTSPFP ANID_04055 N382 1000 100%
HexNAc ...P..n.T.... 56.65 SLLPKDNTTVYGG ANID_01131 N49 1000 100%
HexNAc ...P..n.T.... 56.65 LIGPFINFTTELQ ANID_08401 N419 33.87 100%
HexNAc ...P..n.T.... 56.65 PLYPANNATALLP ANID_07231 N294 11.1 96%
HexNAc ...P..n.T.... 56.65 ITNPLSNITVARN ANID_02325 N112 11.72 95%
HexNAc ...P..n.T.... 56.65 ASAPAANRTELAH ANID_09130 N79 1000 100%
HexNAc ...P..n.T.... 56.65 NACPSLNYTLGGG ANID_02112 N220 49.8 100%
HexNAc ...P..n.T.... 56.65 YMKPVANATSDNN ANID_02388 N93 71.64 100%
HexNAc ...P..n.T.... 56.65 FIYPWINSTDLKE ANID_04102 N713 1000 100%
HexNAc ...P..n.T.... 56.65 FAGPLANQTNLAL ANID_04809 N506 15.26 97%
HexNAc T.....n.T.... 37.4 TSTGGANITSLSA ANID_04390 N78 1000 100%
HexNAc T.....n.T.... 37.4 TINPALNHTTIGR ANID_02360 N166 33.81 100%

108
HexNAc T.....n.T.... 37.4 TIRSSHNDTDDVS ANID_08891 N65 1000 100%
HexNAc T.....n.T.... 37.4 TYTGSKNATYIDP ANID_09340 N493 1000 100%
HexNAc T.....n.T.... 37.4 TSGSWTNTTLTAA ANID_07401 N123 34.87 100%
HexNAc T.....n.T.... 37.4 TALVYVNTTAGPA ANID_02359 N695 1000 100%
HexNAc T.....n.T.... 37.4 TNAEGYNTTAIRH ANID_06438 N671 75.87 100%
HexNAc T.....n.T.... 37.4 TVVFNANRTQVTL ANID_07269 N133 26.67 100%
HexNAc T.....n.T.... 37.4 TLAEKVNLTTGTG ANID_10482 N73 1000 100%
HexNAc T.....n.T.... 37.4 TLPLKANGTLALI ANID_08401 N408 31.82 100%
HexNAc T.....n.T.... 37.4 TLDEKVNLTTGTG ANID_04102 N62 1000 100%
HexNAc T.....n.T.... 37.4 TAIMMRNATIVNQ ANID_00787 N88 32.32 100%
HexNAc T.....n.T.... 37.4 TLEEKINNTGHEA ANID_08401 N63 17.08 100%
HexNAc T.....n.T.... 37.4 TYINLGNETAIDL ANID_07781 N215 14.08 96%

109
Table S4. N-linked glycans from glycoproteins secreted by A. nidulans detected by MALDI TOF/TOF MS.
Assignment m/z
Glucose SCB1 Xylan
Rep1 Rep2 Rep3 Rep1 Rep2 Rep3 Rep1 Rep2 Rep3
Hex5HexNAc2 1579.8 32.8% 40.7% 28.1% 21.8% 43.1% 56.4% 14.8% 23.7% 25.7%
Hex6HexNAc2 1783.9 28.2% 20.4% 26.6% 18.3% 21.2% 19.1% 17.3% 23.0% 20.0%
Hex7HexNAc2 1988.0 12.2% 13.2% 13.8% 14.0% 13.8% 10.5% 14.7% 15.2% 14.1%
Hex8HexNAc2 2192.0 14.5% 16.1% 16.4% 19.2% 11.9% 7.9% 22.0% 18.3% 18.2%
Hex9HexNAc2 2396.2 9.5% 7.8% 9.9% 20.0% 7.2% 4.6% 25.7% 15.2% 16.7%
Hex10HexNAc2 2600.3 2.0% 1.3% 2.8% 5.3% 2.2% 1.3% 5.0% 4.0% 4.6%
Hex11HexNAc2 2804.4 0.4% 0.4% 0.9% 0.9% 0.5% 0.2% 0.6% 0.5% 0.6%
Hex12HexNAc2 3008.4 0.2% 0.2% 0.6% 0.4% n.d 0.1% n.d 0.2% 0.2%
Hex13HexNAc2 3212.5 0.1% n.d 0.4% 0.2% n.d n.d n.d n.d 0.1%
Hex14HexNAc2 3416.5 n.d n.d 0.2% n.d n.d n.d n.d n.d n.d
Hex15HexNAc2 3620.6 n.d n.d 0.1% n.d n.d n.d n.d n.d n.d
Hex16HexNAc2 3824.6 n.d n.d 0.1% n.d n.d n.d n.d n.d n.d
Hex17HexNAc2 4028.7 n.d n.d 0.1% n.d n.d n.d n.d n.d n.d
100.0% 100.0% 100.0% 100.0% 100.0% 100.0% 100.0% 100.0% 100.0%
1 Sugarcane bagasse

110
Capítulo 3 – Influência da N-glicosilação na produção e função de uma β-
xilosidase de A. nidulans
Introdução
A N-glicosilação é um processo bastante complexo, influenciando desde a síntese do
polipeptídeo até suas propriedades estruturais e funcionais. No entanto, os efeitos
global e específico de cada N-glicosilação ainda não são totalmente compreendidos.
Com base nessas informações, esse capítulo visa contribuir na compreensão da
influência da N-glicosilação no enovelamento, na secreção, e na cinética enzimática
de uma CAZyme de A. nidulans.
Com base na proteômica apresentada no capítulo 2, a β-xilosidase BxlB da família
GH3 foi selecionada como modelo para compreender tais influências. β-xilosidases
catalizam a hidrólise de ligações β 1,4 da extremidade não redutora de
xilooligossacarídeos liberando resíduos de xilose. Assim, nesse capítulo estão
descritos os experimentos realizados no engenheiramento da N-glicosilação,
caracterização estrutural e funcional de BxlB. Os glicomutantes de BxlB foram
desenhados, incialmente, para contemplar as variantes: não glicosilada (BxlBDeglyc),
parcialmente glicosilada (BxlBN1;5;7), totalmente glicosilada (BxlBwt) e com um novo
padrão de sítios de glicosilação (BxlBCC). Posteriormente, para refinar a influência de
cada sítio, quatro novas variantes foram criadas: BxlBN1 BxlBN5, BxlBN7 e BxlBN5;7.
Com base nos experimentos conduzidos, observou-se que a alteração no perfil de N-
glicosilação pode influenciar negativamente na secreção enzimática mesmo havendo
elevados níveis de mRNA e os sítios N5 e N7 são muito importantes para secreção e
atividade enzimática de BxlB.
O presente capítulo irá compor o segundo artigo desse trabalho de doutorado, para
submissão ao periódico Journal of Biological Chemistry. Posteriormente, serão
adicionados experimentos de: dinâmica molecular dos glicomutantes BxlBN1;5;7,
BxlBN5;7 e BxlBCC; simulação computacional do glicomutante BxlBCC; novos
experimentos de Western blot dos glicomutantes de BxlB; e possivelmente a estrutura
cristalizada da BxlB. Estes experimentos têm como objetivo refinar a discussão sobre
a influência da N-glicosilação no enovelamento e secreção, e compreender como a
alteração nos padrões de glicosilação em BxlBCC afetaram sua secreção e atividade
enzimática.

111
Redesigning N-glycosylation sites in a GH3 β-xylosidase improves enzyme
efficiency in Aspergillus nidulans
Marcelo V. Rubio1, César R. F. Terrasan1, Fabiano J. Contesini1, Mariane P. Zubieta1,
Jaqueline A. Gerhardt1, André Damasio1*
1Department of Biochemistry and Tissue Biology, Institute of Biology, University of
Campinas (UNICAMP), Campinas, SP, Brazil. 13083-862
Running title: Mutation of N-glycosylation sites improves β-xylosidase kinetics
efficiency
*To whom correspondence should be addressed:
André Damasio; Department of Biochemistry and Tissue Biology, Institute of Biology,
University of Campinas (UNICAMP), Campinas-SP, Brazil Email:
Keywords: GH3, β-xylosidase, Aspergillus nidulans, N-glycosylation, enzyme
secretion, glycomutants

112
Abstract
β-xylosidases are glycoside hydrolases (GHs) that cleave xylooligosaccharides into
shorter oligosaccharides and xylose. Aspergillus nidulans is an established genetic
model and a good source of carbohydrate-active enzymes (CAZymes). N-glycosylation
of fungal proteins deserves attention in order to overcome bottlenecks in enzyme
secretion and application. In this study, a highly secreted A. nidulans GH3 with β-
xylosidase activity (BxlB) harboring several N-glycosylation sites was selected for N-
glycosylation engineering. Glycomutants were designed in order to investigate the
influence of glycosylation on β-xylosidase secretion and function. The deglycosylated
mutant (BxlBDeglyc) showed similar results regarding enzyme secretion and activity
compared to the wild-type (BxlBwt). Interestingly, a partially glycosylated mutant
(BxlBN1;5;7) showed increased activity and secretion levels. On the other hand, the
mutant BxlBCC, in which the glycosylation context was changed by the design of four
new N-glycosylation sites, was expressed but not secreted in A. nidulans. BxlBwt,
BxlBDeglyc and BxlBN1;5;7 showed similar secondary structure, although the mutants had
lower melting temperature compared to the wild-type. Moreover, an additional BxlB
glycomutant maintaining only two N-glycosylated sites (BxlBN5;7) showed improved
catalytic efficiency. This study showed the influence of N-glycosylation on β-xylosidase
function and production in A. nidulans, reinforcing that proteins glycoengineering is a
promising tool to enhance thermal stability, secretion and enzymatic activity. Our report
may also support N-glycosylation modification in CAZymes to biotechnological
applications.
Introduction
Filamentous fungi are important sources of carbohydrate-active enzymes (CAZymes)
since they hold several genes related to plant biomass degradation in the genome and
present high capability of proteins secretion (Benoit et al., 2015). However, an
important bottleneck in the production of plant cell wall degrading enzymes is the level
of protein secretion that still requires improvement to make enzymes application
economically feasible. Enzymes of the glycoside hydrolases family 3 (GH3) present a
variety of activities such as β-xylosidase, β-glucosidase, α-L-arabinofuranosidase and
exo-1,3-1,4-glucanase and are important enzymes produced by Aspergillus spp. GH3
enzymes with β-xylosidase activity (xylan 1,4-β-D-xylosidase, EC 3.2.1.37) are

113
responsible for completing hemicellulose degradation, by hydrolyzing non-reducing
ends of xylooligosaccharides releasing xylose. These enzymes play an important role
in plant biomass degradation, hence presenting different industrial applications in
biofuel, paper, food and animal feed industries (Jordan & Wagschal, 2010).
Several post-translational modifications (PTMs) occur in proteins produced by all types
of organisms, including bacteria and fungi. N-glycosylation is one of the most important
types because it can influence protein secretion, stability, activity, signalization and
provide protection from proteases (Larkin & Imperiali, 2011). In addition, this PTM
occurs in many proteins from filamentous fungi, including those secreted by Aspergillus
nidulans (M. V. Rubio et al., 2016). This modification is catalyzed by
oligosaccharyltransferases in the lumen of the endoplasmic reticulum (ER) and
involves the attachment of N-glycans to asparagine (Asn, N) residue, and it is found in
70% of the predicted N-glycosylation consensus sequence or sequons (Asn-X(aa)-
Ser/Thr) (Weerapana & Imperiali, 2006).
There are interesting examples regarding the influence of N-glycans on CAZymes
properties. A thermophilic GH10 xylanase from Aspergillus fumigatus was expressed
in Pichia pastoris, and the N-glycosylated enzyme showed improved activity and
thermal stability in relation to the non-glycosylated form (Chang et al., 2017). In
addition, the position of N-linked glycan can positively or negatively influence the
processivity of a GH6 cellobiohydrolase from Penicillium verruculosum (Gusakov et
al., 2017). Glycosylation sites engineering showed that N-glycans can improve the
thermal stability of the cutinase C from Aspergillus oryzae by inhibiting its thermal
aggregation behavior (Shirke et al., 2017).
A. nidulans is a model organism for studies regarding secretion of recombinant
enzymes (Lubertozzi & Keasling, 2009; Segato et al., 2012; Zubieta et al., 2018).
However, studies investigating the role of N-glycosylation in enzyme secretion in
filamentous fungi are scarce (Contesini et al., 2017b; Qi et al., 2014; Wei et al., 2013;
Yang et al., 2015). A recent study used N-glycoproteomics and glycomics approaches
in order to perform an overview of N-glycosylation in secreted proteins from A. nidulans
grown in glucose, xylan and NaOH-pretreated sugarcane bagasse. Within 265
identified N-glycoproteins, more than 50% were classified as CAZymes (M. V. Rubio

114
et al., 2016). Among them, some industrially relevant enzymes were highly secreted
representing relevant targets for an investigation regarding the influence of N-
glycosylation on enzymatic properties and secretion. Here a highly secreted GH3
(BxlBwt) from A. nidulans A773 with high activity towards p-nitrophenyl β-D-
xylopyranoside (ρNP-X) was selected as a model for N-glycosylation engineering.
BxlBwt harbored seven putative N-glycosylation sites, and five of them were validated
by LC-MS/MS. Site-directed mutagenesis was applied to design BxlB glycomutants
with different N-glycosylation profiles in order to investigate whether the absence or
the addition of specific N-glycosylation sites could influence enzyme
production/secretion, activity, kinetics and stability. We observed that two N-
glycosylation sites are effectively important for enzyme secretion catalytic efficiency.
In addition, the synthesis of the BxlB was completely impaired by changing the N-
glycosylation context.
Results
BxlBwt has seven predicted N-glycosylation sites
The BxlBwt was identified in the top five enzymes secreted by A. nidulans cultivated in
different polymeric substrates, and it was the most secreted hemicellulase when
cultivation was performed on beechwood xylan (M. V. Rubio et al., 2016). BxlBwt is a
highly N-glycosylated enzyme belonging to the GH3 family harboring seven predicted
N-glycosylation sites (NetNGlyc 1.0 Server). Five sites were validated by mass
spectrometry analysis. Furthermore, in silico analysis indicated that N-glycosylation
enhances the BxlBwt folding by the absence of one folding-stage in comparison to the
non-glycosylated enzyme (Figure S1 and S2). Based on these data, BxlBwt was
chosen as a model glycoprotein for studies regarding the influence of N-glycosylation
on enzyme production/secretion and function.
In order to design BxlB glycomutants, N-glycosylation sequons were mutated by
replacing asparagine to glutamine. Three BxlB N-glycomutants were designed: 1) all
N-glycosylation sites were replaced giving rise to the deglycosylated variant
(BxlBDeglyc); 2) a partially deglycosylated variant was designed by replacing the N340,
N408, N419 and N621 (BxlBN1;5;7); and 3) the N-glycosylation context was completely
changed by the addition of four new N-glycosylation sites using the BxlBDeglyc as

115
template (BxlBCC). The new sites were designed based on the homology with 33
Aspergilli sequences (Figure S3) and by calculating the accessible surface area (ASA)
for each new N-glycosylation site candidate (Figure 1 and Table S1). The design of
BxlBCC enabled to specifically verify the importance of the sequons position for the β-
xylosidase production/secretion and function.
Figure 1. Overview of BxlB glycomutants. Seven N-glycosylation sites were
predicted in the BxlBwt by the NetNGlyc server and five sites were confirmed by LC-
MS/MS (orange circles). Three glycomutants were designed by site-directed
mutagenesis: BxlBN1;5;7, N/Q mutation of four validated N-glycosylated sites; BxlBDeglyc,
N/Q mutation of the seven predicted N-glycosylation sites; BxlBCC, the N-glycosylation
context was changed (purple circles) by the addition of four new sites using BxlBDeglyc
as template, with N121 (A123T), Q163N, Q391N, and N448 (L450T). SP: signal
peptide, N: amino-terminus, C: carboxyl-terminus, grey circles: site non-covered by
LC-MS/MS.
Enzyme secretion analysis suggests that BxlBN1;5;7 was more secreted than the
BxlBwt
All the bxlB genes were cloned into the pEXPYR vector controlled by the glaA promoter
and the glucoamylase signal peptide (SP) from A. niger and then transformed into the
A. nidulans A773 parental strain (Segato et al., 2012). After cultivation on maltose, the
crude supernatants analysis revealed that the complete deglycosylation (BxlBDeglyc)
resulted in similar enzyme secretion and activity (Figure 2A). However, the BxlBN1;5;7
mutant showed higher enzymatic activity (1.5x) and secretion (2.5x) compared to the
wild-type enzyme, according to the protein band intensity quantification using the
Image Lab 5.2.1 Software (Bio-Rad) (data not shown). On the other hand, the BxlBCC
activity and secretion were not detected (Figure 2B), despite some peptides were
detected by mass spectrometry (Table S2).

116
Considering the absence of BxlBCC secretion, intracellular proteins were extracted to
evaluate the presence of BxlBCC. A very low BxlBCC intracellular activity was detected
(Figure S4), and an intermediate level of β-xylosidase activity was found for BxlBwt and
BxlBDeglyc. Furthermore, the intracellular activity of BxlBN1;5;7 was 4-fold higher than
BxlBwt. Moreover, real-time PCR revealed that all BxlB glycomutants were properly
expressed and there was no direct relationship between gene expression and enzyme
production (Figure S5).
Figure 2. Analysis of BxlB glycomutants secretion by homologous expression
in A. nidulans. (A) The secretion of BxlB glycomutants was quantified with the ρNP-
X assay. The reaction was carried out at 50 °C for 15 min and pH 5.0. (B) The
secretomes profile (20 µg of protein) were evaluated by SDS-PAGE stained with
Coomassie brilliant blue and the relative amount of BxlB was estimated using the
Image Lab 5.2.1 software (Bio-Rad). Arrows indicate the quantified band. L: ladder;
A773: A. nidulans parental strain. *ANOVA with Bonferroni post-test, p<0.05.
The removal of N-glycosylation sites decreased the BxlB thermal stability
BxlBwt, BxlBDeglyc and BxlBN1;5;7 were purified by sequential ion-exchange DEAE-
Sepharose and size exclusion Superdex 200 16/600 chromatographies and assayed
to determine the kinetic parameters (Figure 3). Circular dichroism (CD) showed that
the BxlBwt and the glycomutants had similar secondary structure composition,
corresponding to 39.4% α-helix and 11.5% β-strand, nevertheless, the temperature of
melting was reduced from 63.8 °C in the BxlBwt to 61.6 and 59.5 °C in BxlBN1;5;7-and

117
BxlBDeglyc, respectively. Enzymatic activity of purified enzymes showed no difference
between BxlBwt and BxlBDeglyc, however the BxlBN1;5;7 activity was 1.75x higher than
the wild-type.
Figure 3. Functional and structural analysis of BxlB glycomutants. (A) SDS-
PAGE (0.5 µg per lane) and enzymatic activity of BxlB glycomutants quantified by the
ρNP-X assay. The reaction was carried out with 0.1 µg of purified enzymes for 30 min
at 60 °C and pH 5.0. (B) Structural analysis was conducted by CD using a JASCO J-
815 Spectropolarimeter. *ANOVA p<0.05.
BxlBN1;5;7 presented a higher catalytic efficiency
The optimal temperature and pH were 60 °C and 5.0, respectively, for all the BxlB
glycomutants and the wild-type. The BxlBwt, BxlBN1;5;7 and BxlBDeglyc Km values were
2.06, 2.49 and 2.36 mM, while Vmax values were 9.56, 17.28 and 10.12 U/mg,
respectively. Therefore, the BxlBN1;5;7 glycomutant showed 50% higher catalytic
efficiency compared to the wild-type, while the catalytic efficiency of the BxlBDeglyc was
decreased by 7% (Table 1).

118
Designing additional mutants by combining mutations in the sites N1, N5 and N7
For a deeper understanding of the BxlBN1;5;7 improved secretion and catalytic
efficiency, four additional glycomutants were designed by maintaining the following N-
glycosylation sites: N1 (BxlBN1); N5 (BxlBN5); N7 (BxlBN7); and both N5 and N7
(BxlBN5;7) (Figure 4). These newly designed genes were also cloned into the pEXPYR
vector and then transformed into A. nidulans A773.
Figure 4. Overview of additional BxlB glycomutants. Four additional mutants were
designed by site-directed mutagenesis using the BxlBDeglyc as template. SP: signal
peptide, N: amino-terminus, C: carboxyl-terminus, orange circles: detected as
glycosylated by LC-MS/MS, grey circles: non-covered site by LC-MS/MS.
Initial analysis of BxlBN1, BxlBN5 and BxlBN7 showed lower secretion levels compared
to the wild-type, while BxlBN5;7 secretion was improved by achieving similar levels to
BxlBN1;5;7 (Figure S6 and Table 1). Moreover, kinetic parameters showed that the
catalytic efficiency of BxlBN5 and BxlBN7 was drastically reduced, while this parameter
was partially affected in the BxlBN1. Furthermore, BxlBN5;7 presented the highest
catalytic efficiency corresponding to a 57% increase when compared to the wild-type
(Table 1).
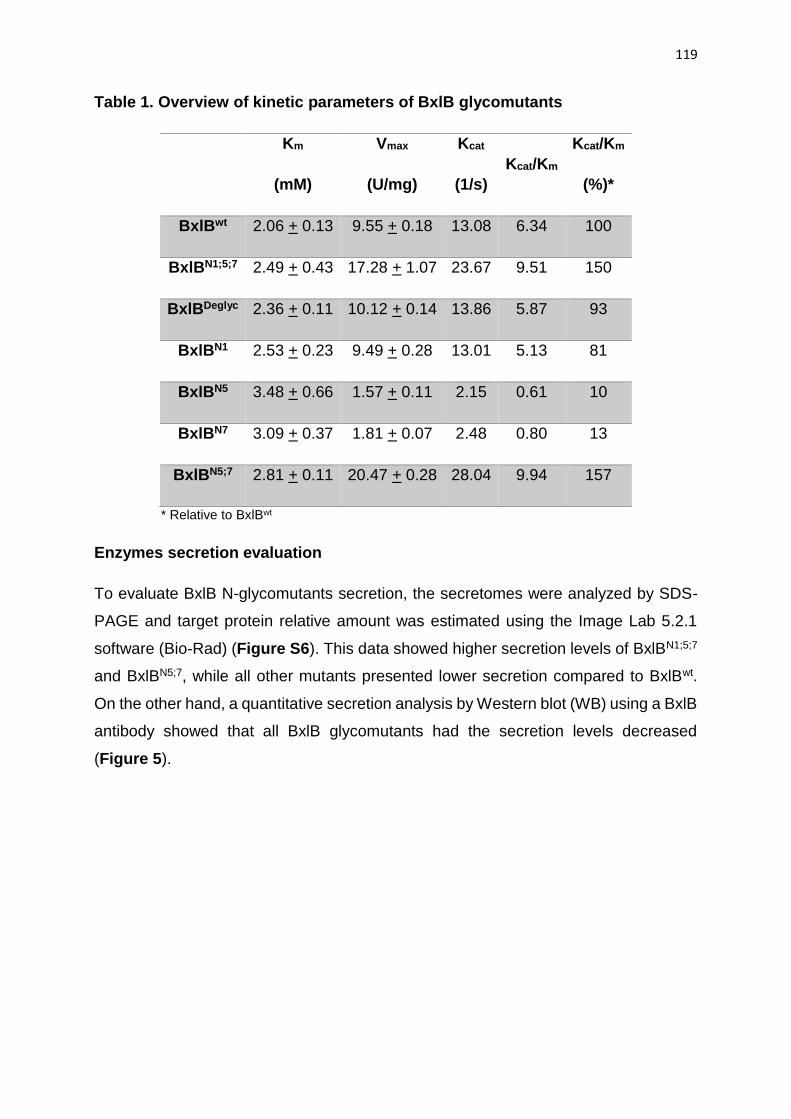
119
Table 1. Overview of kinetic parameters of BxlB glycomutants
Km
(mM)
Vmax
(U/mg)
Kcat
(1/s)
Kcat/Km
Kcat/Km
(%)*
BxlBwt 2.06 + 0.13 9.55 + 0.18 13.08 6.34 100
BxlBN1;5;7 2.49 + 0.43 17.28 + 1.07 23.67 9.51 150
BxlBDeglyc 2.36 + 0.11 10.12 + 0.14 13.86 5.87 93
BxlBN1 2.53 + 0.23 9.49 + 0.28 13.01 5.13 81
BxlBN5 3.48 + 0.66 1.57 + 0.11 2.15 0.61 10
BxlBN7 3.09 + 0.37 1.81 + 0.07 2.48 0.80 13
BxlBN5;7 2.81 + 0.11 20.47 + 0.28 28.04 9.94 157
* Relative to BxlBwt
Enzymes secretion evaluation
To evaluate BxlB N-glycomutants secretion, the secretomes were analyzed by SDS-
PAGE and target protein relative amount was estimated using the Image Lab 5.2.1
software (Bio-Rad) (Figure S6). This data showed higher secretion levels of BxlBN1;5;7
and BxlBN5;7, while all other mutants presented lower secretion compared to BxlBwt.
On the other hand, a quantitative secretion analysis by Western blot (WB) using a BxlB
antibody showed that all BxlB glycomutants had the secretion levels decreased
(Figure 5).

120
Figure 5. Secretion yield evaluated by Western blot. The WB was performed in
triplicate using BxlB glycomutants crude extracts (60 ug) and polyclonal BxlB antibody.
The relative protein amount corresponding to the BxlB band was estimated using the
BxlBwt as a reference. Asterisks in WB indicate the quantified band. A773: A. nidulans
parental strain. *ANOVA with Tukey posttest p<0.05.
Monitoring BxlBCC secretion using C-linked GFP
As previously showed, the absence of BxlBCC secretion cannot be attributed to
transcriptional impairment. In addition, real-time PCR showed non-significant
difference in the transcription of bipA (ER chaperone) and pdiA (protein disulfide
isomerase), genes normally associated to ER stress (data not shown) (Heimel, 2014;
Schröder, 2008; Sims et al., 2005). For further investigation on the low secretion of
BxlBCC, GFP was fused to the enzyme C-terminus, allowing protein monitoring by
fluorescence microscopy. The glucoamylase SP was maintained in the cloning of
bxlBwt and bxlBCC genes into the pmcB17 vector, which presents an alcA promoter.
Real-time PCR showed that both fused genes were expressed at similar levels in A.
nidulans (Figure 6).

121
G F P -B x lBw t
G F P -B x lBCC
0
1 0
2 0
3 0
4 0
5 0
Fo
ld C
ha
ng
e (
log
2)
***
Figure 6. Real-time PCR of BxlBwt using the pmcB17 expression vector. Data
were analyzed by the ΔΔCt method using tubC as endogenous control and A. nidulans
A773 was established as control strain. *ANOVA p<0.05.
After transcriptional analysis, intra and extracellular β-xylosidase activity were assayed
to monitor enzyme production. The activities of A. nidulans GFP-BxlBCC and A.
nidulans A773 crude extracts were very similar, showing the absence of GFP-BxlBCC
secretion. However, both intra and extracellular activities were detected for A. nidulans
GFP-BxlBwt. Despite the absence of functional activity, WB showed the intracellular
production of both enzymes at 48 h, and a weak extracellular signal detected for GFP-
BxlBCC at 24 and 48 h (Figure 7). In order to confirm the intracellular GFP signals, the
same strains were examined by confocal microscopy. Autofluorescence was avoided
by calibrating the microscope using A. nidulans A773 as control, and all images were
taken using the same measure settings. In spite of the absence of intracellular GFP
detection by WB at 24 h, confocal microscopy showed signal in the recombinant strains
(Figure S7). These results were further validated in the recombinant strains by using
a polyclonal BxlBwt antibody. Excepting for BxlBCC, all the BxlB glycomutants were
detected in the extracellular fraction (Figure S8A). However, a strong intracellular
signal was detected BxlBwt, while the BxlBCC presented weak signals (Figure S8B).

122
Figure 7. Enzymatic activity and WB analysis of BxlB glycovariants cloned into
the pmcB17 vector. The secretion of GFP-tagged BxlB glycomutants was quantified
by ρNP-X assay and monitored by WB using anti-GFP antibody (below X-axis at the
respective enzymatic activity). *ANOVA p<0.05.
Discussion
N-glycosylation enhances BxlB folding
The bxlBwt gene from A. nidulans encoding for a β-xylosidase was selected among 265
N-glycoproteins. The high secretion levels when cultivation was performed in
beechwood xylan revealed that this enzyme is very important for biomass
deconstruction (M. V. Rubio et al., 2016). β-xylosidases have been detected in various
prokaryotes and eukaryotes, however only eleven eukaryotic GH3 β-xylosidases have
been characterized to date, according to the MycoClap database (Murphy, Powlowski,
Wu, Butler, & Tsang, 2011; Strasser et al., 2015). Among the characterized β-
xylosidases, four correspond to recombinant enzymes produced in hosts such as
Aspergillus spp, P. pastoris and Saccharomyces cerevisiae (Table S3). Usually, well-
known vectors and homologous expression systems are selected to prevent concerns
related to correct gene expression and protein production. In addition to the high

123
secretion in lignocellulosic substrates, BxlBwt is a good model for studies regarding the
influence of N-glycosylation on enzyme secretion and functional parameters due to its
high levels of N-glycosylation and enzymatic activity.
As observed in the free energy landscape (Figure S1 and Figure S2), the glycosylated
protein loses an intermediate state (arrow at point 2), facilitating the folding process.
The arrows 1 and 2 at the Figure S2 show that the non-glycosylated protein may have
difficulties in reaching the folded state. N-glycans contact polypeptide chains, changing
both the free energy and specific conformational ensembles. In this sense, secretion
of glycoproteins is generally facilitated by their higher stability and the presence of the
calnexin/calreticulin quality control (Helenius & Aebi, 2004). Furthermore, low
thermodynamic stability decreases proteins export efficiency, i.e., very unstable
proteins are poorly exported (Wiseman, Powers, Buxbaum, Kelly, & Balch, 2007).
Influence of N-glycan composition in protein folding was also studied in the human
immune cell receptor (hCD2ad), and it was reported that the presence of complete N-
glycans accelerates folding by 4-fold and also stabilizes protein structure (Hanson et
al., 2009).
N-glycans at the positions 1, 5 and 7 are important for enzyme secretion
The secretion of BxlB, BxlBN1;5;7, BxlBCC and BxlBDeglyc were evaluated by homologous
expression in A. nidulans avoiding concerns on codon usage and gene expression.
SDS-PAGE and activity assays using the extracellular enzymes revealed that
complete deglycosylation did not affect BxlBDeglyc secretion and activity. On the other
hand, the improvement of secretion and enzymatic activity of the BxlBN1;5;7 indicates
that N63, N458 and N760 sites are important for protein folding and export. Moreover,
the BxlBCC mutant activity and secretion were not detected in the extracellular fraction
indicating some impairment in gene expression, protein production or in the secretion
pathway. Real-time PCR showed high transcription levels of wild-type, BxlBN1;5;7,
BxlBCC and BxlBDeglyc, suggesting that the transcription is not a bottleneck for BxlBCC
production. There are few studies dealing with N-glycan influence in protein secretion,
and some of them reported different results than we found in this study. By one side,
changes in two N-glycosylation sites (N14Q and N48Q) of a heterologous lipase
expressed in P. pastoris did not affect its secretion, while the N60Q mutation

124
completely abolished the enzyme secretion (Yang et al., 2015). Hence, N-glycans
attached to specific positions in the protein might be essential to folding kinetics and
quality control process in the secretion pathway.
Deglycosylation of BxlBwt reduces the thermal stability
Secondary structure analysis of BxlBwt and glycomutants revealed a classical profile of
α-helix as the predominant secondary structure in all recombinant enzymes (Figure 3)
(Kelly, Jess, & Price, 2005). The similarity among CD data was confirmed by
deconvolution analysis that verified the same rate of α-helix and β-strand. The
deconvolution of CD data showed high similarity to the A. niger β-xylosidase with 41%
α-helix and 16% β-sheet, however a T. reesei β-xylosidase presented lower similarity
with 23% α-helix, 27% β-sheet (Díaz-Malváez, García-Almendárez, Hernández-Arana,
Amaro-Reyes, & Regalado-González, 2013; Rojas et al., 2005).
Generally, glycoproteins present higher stability when compared to their partially or
non-glycosylated counterparts, in spite of the absence of structural links associated
with N-glycosylation (Imperiali & Connor, 1999). The same result was observed in this
study, i.e., the thermal stability decreased according to deglycosylation level, with the
deglycosylated form the most unstable variant (Figure 3). Recently, thermal stability
was evaluated in a T. reesei cellobiohydrolase (TrCel7A) by differential scanning
calorimetry and lower stability was reported for all the fifteen N-glycosylation mutants
(Amore et al., 2017). Commonly, interactions between N-glycan sugars and amino acid
residues stabilize the protein structure, consequently, the glycoproteins are more
stable than the non-glycosylated form (Imperiali & Connor, 1999; Shental-Bechor &
Levy, 2008).
Glycosylations at N63, N458 and N760 sites increase BxlB kinetic parameters
The optimal activity conditions to all BxlB glycomutant were pH 5.0 and 60 °C using
the substrate ρNP-X, while this same enzyme expressed in P pastoris, showed optimal
activity at pH 4.4 and 48 °C using rye arabinoxylan and xylohexaose as substrates
(Bauer et al., 2006). This difference in reaction conditions due to N-glycosylation
composition was also reported for a β-glucosidase from A. terreus expressed in P.
pastoris and T. reesei (Wei et al., 2013). Kinetic parameters showed that removal of

125
any N-glycosylation site decreases the BxlBwt affinity for ρNP-X, indicating that N-
glycans should help substrate recognition or affect catalytic site flexibility. Structural
dynamics can influence catalytic site residues position and flexibility, impacting in the
kinetic parameters (Solá et al., 2007). Wei et al. showed that the specific activity of A.
terreus β-glucosidase produced in filamentous fungi or yeast decreases by removing
some N-glycosylation site (Wei et al., 2013). Removal of four N-glycosylation sites in
the BxlBN1;5;7 increased Vmax and Km suggesting important changes in structural
dynamics.
N5 and N7 are essential glycosylation sites to BxlB catalytic efficiency
Four additional mutants were designed to further understanding the influence of each
N-glycosylation site in folding, secretion and functional parameters of BxlBwt. The
secretion and catalytic efficiency of BxlBN1 decreased in comparison to the BxlBDeglyc.
A similar behavior was showed for BxlBN1;5;7 in relation to BxlBN5;7. These data suggest
that the N1 site had a slight negative influence on BxlBwt kinetic parameters and
secretion levels. On the other hand, for the glycomutants containing a unique N-
glycosylation site, maintaining the N1 site (BxlBN1) do not drastically affect secretion
and enzyme function. The sites N5 and N7 individually had a more detrimental
influence on enzyme function; however, together these sites had the highest positive
influence on the catalytic efficiency. These alterations in secretion and enzyme activity
were probably impacted by the N-glycan position in the 3D structure, protein dynamics
and stability (Nagae & Yamaguchi, 2012; Solá et al., 2007). Despite the BxlBwt has
apparently two essential N-glycosylation site (N5 and N7), previous studies
demonstrated the critical importance of a single N-glycosylation site to the folding and
enzymatic activity, for example, the P. pastoris BglS N224 is important to enzyme
activity and production and the N428 is essential to N-Acetylglucosamine-6-
sulfotransferase-1 enzyme activity (Desko, Gross, & Kohler, 2009; Wei et al., 2013).
Changing the BxlBwt N-glycosylation context impaired the protein folding and
secretion
Our results show that the misposition of N-glycosylation sites can impair the production
of recombinant enzymes in A. nidulans. In order to understand the impairment of
BxlBCC secretion, experiments were designed to monitoring enzyme secretion.

126
Designing hybrid enzymes tagging another enzyme at C terminus are generally used
to improve secretion and monitor target enzymes (Fleiβner & Dersch, 2010; H.
Nevalainen & Peterson, 2014; Owen P. Ward, 2012). The absence of BxlBCC secretion
did not take place due to transcriptional issues, therefore the BxlBwt was GFP-fused in
order to follow the enzyme along the secretion pathway. GFP was tagged by gene
cloning into a pmcB17 vector and in spite of using the strong alcA promoter, lower
mRNA levels were detected compared to the expression under glaA promoter (Figure
5 and S5). Despite this lower expression, the GFP-BxlBwt (Figure 6) was functionally
secreted but with lower activity when compared to the BxlBwt (Figure 2). GFP-BxlBCC
had no activity on ρNP-X, however weak extracellular and stronger intracellular signals
were detected at 48 h. The microscopy data corroborates the intracellular signal
detection, indicating that the mutations designed in BxlBCC affected the folding and
secretion yield. Probably, the misfolding of BxlBCC triggered protein degradation and a
small amount was secreted in a non-functional form.
Proteins with difficulty in achieving the correct folding remain longer in the ER, which
has a strict quality control in order to facilitates the folding or to direct misfolded proteins
to degradation (Helenius & Aebi, 2004; E. Sergio Trombetta & Parodi, 2003). The
quality control in A. nidulans is represented by the calnexin cycle. Calnexin recognizes
the N-glycan attached to proteins, assists the disulfide bonds and, consequently
facilitates the correct folding and secretion (Moremen et al., 2012; Schwarz & Aebi,
2011). Here, the confocal microscopy showed that the GFP-BxlBCC was retained in the
intracellular fraction suggesting that the misposition of N-glycosylation sites triggered
the protein for degradation.
Experimental procedure
Strains, plasmids and media
The reference strain A. nidulans A773 (pyrG89;wA3;pyroA4) was obtained from the
Fungal Genetics Stock Center (FGSC). The A. nidulans A773 and recombinant strains
were regularly maintained in minimal medium (MM) 1% glucose (m/v), pH 6.5
(Pontecorvo et al., 1953; Segato et al., 2012). Plasmids were propagated in E. coli
DH5α maintained in Luria-Bertani (LB) medium.

127
Site-directed mutagenesis in GH3 β-xylosidase
The gene bxlBwt that encodes BxlBwt were amplified from A. nidulans A773 genomic
DNA by PCR using specific primers (BxlBwt Fwd and BxlBwt Rev) (Table S4). The gene
was cloned into NotI and XbaI sites of the pEXPYR shuttle vector previously described
(Segato et al., 2012). The GFP-tagged glycomutants were obtained by the gene
cloning in pmcB17 vector (Fernández-Abalos, Fox, Pitt, Wells, & Doonan, 1998).
The residues substitutions in BxlBwt sequence generated BxlBN1;5;7 (N340Q, N408Q,
N419Q and N621Q), BxlBDeglyc (N63Q, A123T, Q163N, N340Q, Q391N, N408Q,
N419Q, N458Q, N621Q and N760Q) and BxlBCC (N63Q, A123T, N340Q, N408Q,
N419Q, L450T, N458Q, N621Q and N760Q) mutants synthesized by GenOne (Rio de
Janeiro, Brazil). The other four mutants: BxlBN1 (N340Q, N408Q, N419Q, N458Q,
N621Q and N760Q), BxlBN5 (N63Q, N340Q, N408Q, N419Q, N458Q and N621Q),
BxlBN7 (N63Q, N340Q, N408Q, N419Q, N458Q and N621Q) and BxlBN5;7 (N63Q,
N340Q, N408Q, N419Q, N621Q) were designed using Q5® Site-Directed
Mutagenesis Kit (New Englands Biolabs). All primer sequences used in this study are
reported in the supplementary Table S4.
A. nidulans transformation
The genes cloned into the pEXPYR plasmid were transformed into calcium competent
E. coli cells by heat shock and confirmation was carried out by colony PCR. Positive
colonies were cultivated overnight, and plasmids extracted and used in fungal
transformation.
The fungal transformation was carried out as previously described with modifications
(Szewczyk et al., 2006). Spores of A. nidulans A773 were inoculated on YG medium
(20g.L-1 glucose, 5 g.L-1 yeast extract, 1× trace elements, 1 mg l−1pyridoxine and
2.5 mg l−1 uracil/uridine) and incubated at 30°C/130 rpm for 13 hours. The protoplast
was prepared by hydrolysis of the fungal cell wall with 125 mg of lysozyme from
chicken egg white (Sigma L7651) and 1.02 g of Vinotaste Pro (Novozymes) for 2 hours.
Approximately 10 µg of recombinant DNA was mixed with protoplasts solution and
PEG 25% (w/v). Protoplast-recovery were performed in MM plates supplemented with
1.2 M sorbitol and pyridoxine incubated at 37°C for three days.

128
In situ analysis
The stability and free landscape energy were performed using simplified contact of
structural units (CSU software) interaction type. This method analyzes interatomic
structures contacts, such as helices, sheets, strands and residues (Sobolev, Sorokine,
Prilusky, Abola, & Edelman, 1999). The structures were simplified to alfa-carbon
interactions. The CSU software has been designed to assist the molecular biologist in
understanding the structural consequences of modifying a ligand and/or protein. The
analysis was conducted in function of temperature and free energy in order to evaluate
stability and folding, respectively.
Biochemical characterization of β-xylosidase and glycomutants
Enzymatic activity and protein determination
107–108 fresh spores of A. nidulans A773 and recombinant strains were inoculated in
MM supplemented with 2% (w/v) maltose, pH 6.5 for 36 h at 37°C. Proteins in the crude
extracts were quantified by the Bradford method (Bradford, 1976). About 20 ug of
supernatant proteins were loaded in SDS-PAGE (Laemmli, 1970) and the target
protein relative secretion was measured by Image Lab 5.2.1 Software (Bio-Rad) using
the BxlBwt band as reference. Enzymatic activity was determined using 5 mM ρNP-X
as substrate and ρ-nitrophenol (ρNP) release was determined at 400 nm. One unit (U)
of β-xylosidase activity was defined as the amount of enzyme releasing 1 µmol of ρNP
per minute under the assay conditions.
Protein purification
All enzymes were purified by two-steps using HiPrepTM DEAE FF 16/10 (GE
Healthcare) followed by HiLoadTM 16/600 SuperdexTM 200 pg (GE healthcare)
columns, as previously described (Contesini et al., 2017a). Purification was evaluated
by SDS-PAGE and protein concentration was determined by reading the absorbance
at 280 nm and using the molar extinction coefficient (Kelly et al., 2005) calculated from
amino acid composition (http://web.expasy.org/protparam/).

129
Structural characterization by Circular Dichroism (CD) spectroscopy
CD analysis was carried out using a JASCO 815 spectropolarimeter (JASCO Inc.,
Tokyo, Japan) equipped with a Peltier temperature control unit and a 0.1 cm path
length cuvette as previously described (Cota et al., 2011). Data from 260 to 190 nm
were collected using 100 nm/min scanning speed, 1 nm spectral bandwidth and 0.5 s
response time. Melting temperature was evaluated by spectra measurement at 20 °C
to 100 °C.
Effect of temperature and pH on enzyme activity
To determine the optimal temperature of all recombinant proteins, enzyme activity was
assayed from 35 to 70 °C at pH 5.0, and for optimal pH, activity was determined using
different buffers in a pH range from 3.0 to 12.5 at 50 °C and ρNP-X as substrate.
Kinetic parameters
Maximum velocity (Vmax), Michaelis-Menten constant (Km), catalytic constant (Kcat) and
catalytic efficiency (Kcat/Km) were determined for all mutants using different
concentrations of ρNP-X (1-20 mM).
RNA extraction, transcript analysis by qPCR (quantitative real-time PCR) and
primer design
Total RNA was extracted by grinding frozen mycelia with a mortar and pestle under
liquid nitrogen, followed by extraction using Direct-Zol RNA Miniprep from Zymo
Research according to the manufacturer’s instructions. Total RNA (DNA free) was
assayed for reverse transcription using the Maxima First Strand cDNA Synthesis Kit
for RT-qPCR, with dsDNase (ThermoFisher Scientific). cDNA samples were diluted
and each qPCR reaction containing cDNA (100 ng), SYBR Green (Life Technologies),
forward and reverse primers (Table S4) and nuclease-free water was carried out using
ViiA™ 7 real-time PCR system (Life Technologies). All PCR reactions were carried out
in biological triplicate. Gene expression levels in the different samples were determined
using the ΔΔCt method and β-tubulin (tubC) as reference gene.

130
Confocal microscopy
Microscopic analyses were carried out by confocal microscopy based on Fischer-
Parton et al. (2000) with modifications. Mycelium was obtained after 24 and 48 h
cultivation of A. nidulans strains, at 37 °C and 200 rpm in MM. The images were
obtained by an LSM 510 Axiovert 200 M (Carls Zeiss) confocal inverted microscope,
fitted with an argon laser. The laser intensity was set using the A. nidulans A773 strain
to avoid autofluorescence at 488 nm excitation and fluorescence emission at 530 nm.
All samples were spread over a microscope slide, in natura, then analyzed right after
the cultivation.
Western Blot
WB analysis was carried out as described by Nutzmann et al. (2011). Sixty μg of
protein were separated by SDS-PAGE followed by transfer to PVDF membrane by
using a wet blotting system (Bio-Rad). The membrane was blocked with BSA 5%, then
incubated overnight with primary antibodies (GFP or BxlB), gently shaken at room
temperature, and posteriorly the secondary antibody anti-rabbit immunoglobulin G
labeled with peroxidase. Protein detection was carried out using Clarity Western ECL
Substrate chemiluminescence detection kit (Bio-Rad), as described by manufacturer's
protocol.
Conclusion
The study of N-glycosylation patterns on fungal CAZymes is important to shed light on
the effect of N-glycans position on proteins properties such as structure, dynamics,
stability and function. Here we demonstrate that N-glycosylation facilitates the correct
folding of a GH3 β-xylosidase by losing an intermediate state. The change of N-
glycosylation context in the BxlB resulted in secretion impairment even with high levels
of mRNA. Secondary structures were preserved in the BxlB glycomutants but their
thermal stability were reduced. Despite the secretion and kinetics parameters of BxlBwt
were affected by N-glycosylation sites mutations, the completely deglycosylated
enzymes (BxlBDeglyc) was secreted in a functional state. There is strong evidence that
misposition of BxlB N-glycosylation sites (BxlBCC) resulted in unfolded/misfolded and
non-functional enzyme. At the individual level, the N-glycosylation sites N5 and N7 are

131
essential to BxlB, improving enzyme catalytic efficiency. Moreover, BxlB glycomutants
showed how complex the N-glycosylation effect is by positively and/or negatively
affecting the folding process, secretion and kinetics parameters. N-glycosylation
engineering can be a promising tool to enhance target enzymes secretion, activity and
thermal stability.
Acknowledgments
This research was supported by FAPESP (grant 2012/20549-4 to ARLD). We are
grateful to the National Council for Scientific and Technological Development (CNPq)
for the financial support (441912/2014-1 and 304816/2017-5 to ARLD). MVR, CRFT,
FJC and MPZ received FAPESP fellowships (13/24988-5, 16/16306-0, 17/10083-1
and 14/15403-6). We thank the LNBio Mass Spectrometry staff for the assistance with
LC-MS/MS.
Competing interests
The authors declare that they have no competing interests.
Author’s contributions
ARLD conceived and designed the experiments. MVR, CRFT and JAG participated in
the design of the study and performed the experiments. MVR, CRFT and JAG and
ARLD analyzed the data. MVR, CRFT, FJC and MPZ drafted the manuscript. ARLD
revised the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
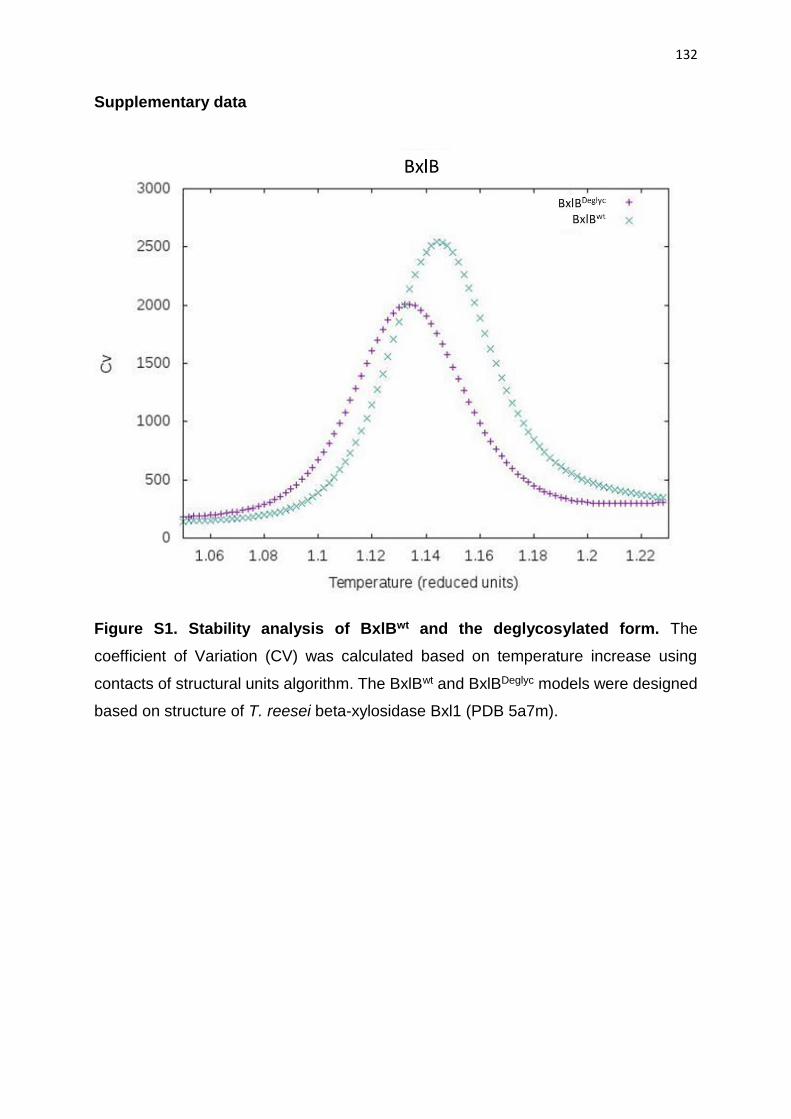
132
Supplementary data
Figure S1. Stability analysis of BxlBwt and the deglycosylated form. The
coefficient of Variation (CV) was calculated based on temperature increase using
contacts of structural units algorithm. The BxlBwt and BxlBDeglyc models were designed
based on structure of T. reesei beta-xylosidase Bxl1 (PDB 5a7m).

133
Figure S2. Free landscape energy analysis of BxlBwt and deglycosylated form.
The number of contacts indicates the transition of unfolded to folded form. The two
states with higher free energy (1 and 2 arrows) indicates that N-glycans facilitates the
folding. The free-energy profile in which an interaction number close to the value zero
indicates the unfolded state and when close to the value one, the folded state (0.9).

134
Figure S3. Analysis of N-glycosylation sites conservation by the alignment of
thirty-three BxlBwt homologous sequences. The alignment was performed by the
ClustalW algorithm. The homologous (E-value < 10-40) sequences were obtained in
Aspergillus Genome Database (AspGD).
BxlB
wt
BxlB
N1;5
;7
BxlB
CC
BxlB
Degly
c
A773
0
5
1 0
1 5
2 0
2 5
6 0
7 0
8 0
9 0
Re
lati
ve
ac
tiv
ity
(A
77
3) * * *
*
*
Figure S4. Analysis of BxlB glycomutants intracellular activity by homologous
expression in A. nidulans. The β-xylosidase activity was measured using the ρNP-X
assay. The reaction was carried out at 50 °C for 15 min and pH 5.0. *ANOVA p<0.05.

135
BxlB
wt
BxlB
N1;5
;7
BxlB
CC
BxlB
Degly
c
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
Fo
ld C
ha
ng
e (
log
2)
Figure S5. Real-time PCR analysis of BxlB glycomutants. The data was analyzed
by ΔΔCt method using the tubC as endogenous gene and A. nidulans A773 as control
strain. The gene induction was performed on maltose 2% and 36h. ANOVA statistics
were performed and no significant difference was observed.
Figure S6. Quantification of BxlB glycomutants secretion in A. nidulans.
Extracellular extracts (20 ug) were analyzed in biological replicates by Image Lab 5.2.1
software (Bio-Rad). Arrows indicate the quantified band. L: ladder.

136
Figure S7. In vivo monitoring of GFP-tagged BxlB mutants by confocal
microscopy. The intracellular GFP-tagged BxlB glycomutants were monitored after
24 h of induction with ethanol 2%. All images were captured using the same
parameters and the A. nidulans A773 strain was set as control to avoid
autofluorescence.

137
Figure S8. WB analysis of BxlB glycomutants. A) A. nidulans secretomes producing
BxlB glycomutants and the control strain A773 (30 ug) were analyzed using anti-BxlB
polyclonal antibody. B) The same analysis was performed in triplicate using 100 ug of
intracellular fraction and 0.5 ug of purified BxlBwt as positive control (C).
Table S1. Overview of BxlBwt putative N-glycosylated sites.
BxlBwt (PDB 3u48)
Asn number Sequence Asn position ASA Asn condition
(LC-MS/MS)
1 KIN*NT 63 86.8 + Glycosylated
2 ISN*ET 340 54.8 + Glycosylated
3 KAN*GT 408 69.9 + Glycosylated
4 FIN*FT 419 60.4 + Glycosylated
5 EVN*ST 458 79.3 + Not covered
6 TFN*VS 621 71.0 + Glycosylated
7 SGN*DS 760 NC Non glycosylated
ASA: Accessible Surface Area
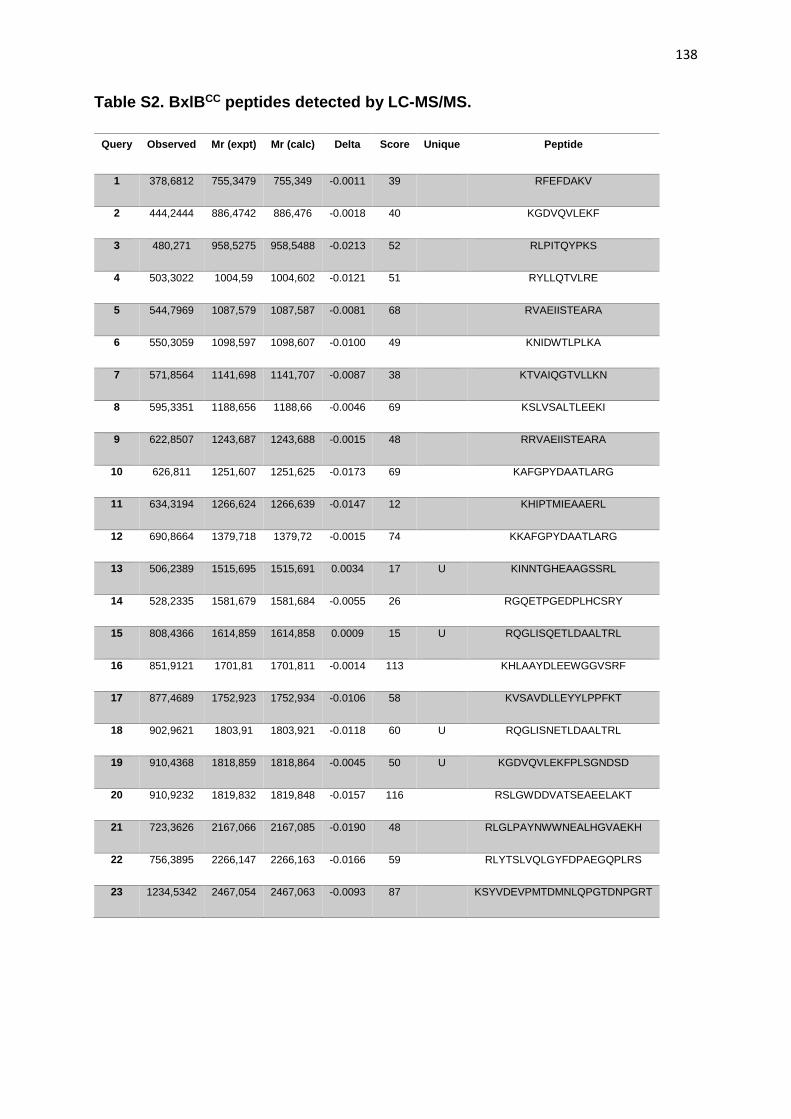
138
Table S2. BxlBCC peptides detected by LC-MS/MS.
Query Observed Mr (expt) Mr (calc) Delta Score Unique Peptide
1 378,6812 755,3479 755,349 -0.0011 39 RFEFDAKV
2 444,2444 886,4742 886,476 -0.0018 40 KGDVQVLEKF
3 480,271 958,5275 958,5488 -0.0213 52 RLPITQYPKS
4 503,3022 1004,59 1004,602 -0.0121 51 RYLLQTVLRE
5 544,7969 1087,579 1087,587 -0.0081 68 RVAEIISTEARA
6 550,3059 1098,597 1098,607 -0.0100 49 KNIDWTLPLKA
7 571,8564 1141,698 1141,707 -0.0087 38 KTVAIQGTVLLKN
8 595,3351 1188,656 1188,66 -0.0046 69 KSLVSALTLEEKI
9 622,8507 1243,687 1243,688 -0.0015 48 RRVAEIISTEARA
10 626,811 1251,607 1251,625 -0.0173 69 KAFGPYDAATLARG
11 634,3194 1266,624 1266,639 -0.0147 12 KHIPTMIEAAERL
12 690,8664 1379,718 1379,72 -0.0015 74 KKAFGPYDAATLARG
13 506,2389 1515,695 1515,691 0.0034 17 U KINNTGHEAAGSSRL
14 528,2335 1581,679 1581,684 -0.0055 26 RGQETPGEDPLHCSRY
15 808,4366 1614,859 1614,858 0.0009 15 U RQGLISQETLDAALTRL
16 851,9121 1701,81 1701,811 -0.0014 113 KHLAAYDLEEWGGVSRF
17 877,4689 1752,923 1752,934 -0.0106 58 KVSAVDLLEYYLPPFKT
18 902,9621 1803,91 1803,921 -0.0118 60 U RQGLISNETLDAALTRL
19 910,4368 1818,859 1818,864 -0.0045 50 U KGDVQVLEKFPLSGNDSD
20 910,9232 1819,832 1819,848 -0.0157 116 RSLGWDDVATSEAEELAKT
21 723,3626 2167,066 2167,085 -0.0190 48 RLGLPAYNWWNEALHGVAEKH
22 756,3895 2266,147 2266,163 -0.0166 59 RLYTSLVQLGYFDPAEGQPLRS
23 1234,5342 2467,054 2467,063 -0.0093 87 KSYVDEVPMTDMNLQPGTDNPGRT

139
Table S3. Characterized β-xylosidases using pNP-X according to the MycoClap*.
Entry Name Species Host (for
recombinant
expression)
Specific
Activity
pH Temp
(°C)
Genbank Protein ID Uniprot ID Literature PMID
XYL3A_TRIRE Trichoderma
reesei
Saccharomyces
cerevisiae
active CAA93248 Q92458 8837440
XYL3D_ASPNG Aspergillus
niger
native 60.2 U/mg CAB06417, CAB59162,
CAK37179, CAW52627
O00089 9128738
XYL3A_EMENI Emericella
nidulans
Aspergillus
nidulans G191
active 5.0 50 CAA73902 9546179
XYL3A_ASPOR Aspergillus
oryzae
native 76 U/mg 4.0 60 BAA28267 9872754
XYL3A_ASPAW Aspergillus
awamori
native active BAE19756 Q4AEG8 16202538
XYL3A_ASPJA Aspergillus
japonicus
native 112 U/mg 4.0 70 BAG82824 B6EY09 19000618

140
XYL3A_AURPU Aureobasidiu
m pullulans
native 288 U/mg 3.5 70 BAI82526 D4AHT5 20547381
XYL3A_TRIRE Trichoderma
reesei
native 28 U/mg 4.0 60 DOI: 10.1007/BF00268208
XYL3A_ASPAW Aspergillus
awamori
native 20 U/mg 4.0 70 DOI: 10.1271/bbb.61.2010
XYL3A_EMENI Emericella
nidulans
native 107 U/mg http://onlinelibrary.wiley.com
/doi/10.1111/j.1574-
6968.1996.tb08003.x/pdf
XYL3A_ASPJA Aspergillus
japonicus
Pichia pastoris
GS115
19 U/mg
* MycoClap site: https://mycoclap.fungalgenomics.ca/mycoCLAP/

141
Table S4. Oligonucleotides used in this study.
Primer name Sequence
BxlBwt Fwd 5’- TATAGCGGCCGCTACCCGGACTGCACAA -3’
BxlBwt Rev 5’- TATATCTAGATAATCACTGTCGTTACCTGACA -3’
RT BxlBwt Fwd 5’- ACCATGATCGAGGCAGCAG -3’
RT BxlBwt Rev 5’- GGGCATCATCGAATCCGTCT -3’
N1 Fwd 5’- AGAGAAAATCAACAACACGGGCCACG -3’
N1 Rev 5’- TCGAGTGTCAGGGCAGAG -3’
N5 Fwd 5’- GACAGAGGTGAATTCGACAAGCACAGACG -3’
N5 Rev 5’- CCTGGCGCGGTGAGAACG -3’
N7 Fwd 5’- GTTGTCAGGTAACGACAGTGATTATC -3’
N7 Rev 5’- GGAAACTTTTCAAGCACC -3’
The NotI and XbaI restriction sites are shown in bold type

142
Capítulo 4. Considerações finais
Historicamente, estudos relacionados à glicosilação de proteínas foram desenvolvidos
a fim de compreender o papel do carboidrato em modificações biofísicas e
comunicação celular, com foco no desenvolvimento de novos tratamentos para
doenças humanas. Apesar de escassos, alguns estudos têm apresentado resultados
bastante promissores ao analisar o efeito das glicosilações no enovelamento,
secreção e propriedades funcionais de diferentes enzimas.
A N-glicosilação efetiva e correta é essencial para a produção de proteínas
heterólogas em fungos filamentosos. O acúmulo de proteínas não enoveladas ou mal
enoveladas representa um grande problema na via de secreção e,
consequentemente, no rendimento final da proteína alvo. Assim, estudar o processo
de N-glicosilação em A. nidulans, um organismo modelo, é fundamental para propor
novas estratégias para melhorar a secreção de proteínas recombinantes nesta
linhagem e em fungos filamentosos de maneira geral.
A glicoproteômica foi o primeiro trabalho de A. nidulans analisando proteínas, sítios
de N-glicosilação e N-glicanas. Apesar de não ser uma linhagem amplamente utilizada
na indústria, nossos resultados mostraram que A. nidulans possui um repertório de
CAZymes capaz de degradar as principais ligações glicosídicas presentes na
biomassa lignocelulósica. Dentre as proteínas identificadas, observou-se que o
repertório enzimático está diretamente relacionado com a composição do substrato
indutor.
As N-glicanas contendo cinco hexoses foram prevalentes nos cultivos em glicose e
bagaço de cana-de-açúcar, enquanto, o cultivo com xilano apresentou quantidades
semelhantes de 5 a 9 hexoses. Identificou-se, também, a preferência de A. nidulans
por sítios de glicosilação compostos por N-X-T, os quais foram predominantemente
encontrados em regiões da proteína com predominância de resíduos de aminoácidos
hidrofóbicos ou polares não carregados.
Os resultados da glicoproteômica poderão facilitar a manipulação e desenho correto
de sítios de N-glicosilação em genes recombinantes a serem expressos em A.
nidulans. Dessa forma, a β-xilosidase BxlB da família GH3 foi selecionada como

143
modelo para compreender a influência e efeitos da N-glicosilação no enovelamento,
na secreção, e na cinética enzimática de uma CAZyme de A. nidulans. BxlB e seus
respectivos glicomutantes foram caracterizados estrutural e funcionalmente. Com
base nos experimentos conduzidos, concluiu-se que a alteração no perfil de N-
glicosilação pode influenciar negativamente a secreção enzimática mesmo havendo
elevados níveis de mRNA. Os glicomutantes BxlBN1;5;7 e BxlBN5;7 apresentaram maior
eficiência catalítica. Dessa forma, concluiu-se que os sítios N5 e N7 são essenciais
para cinética enzimática da BxlB.
Em suma, a manipulação do processo de N-glicosilação é uma ferramenta bastante
promissora na otimização da secreção, atividade enzimática e estabilidade térmica.
Dessa forma, o estudo de sítios de N-glicosilação se torna fundamental para o correto
engenheiramento de proteínas recombinantes. Os resultados obtidos com os
glicomutantes de BxlB confirmam o grande potencial da manipulação dos sítios de N-
glicosilação em uma CAZyme alvo. Além disso, esse trabalho irá contribuir tanto com
a determinação de CAZymes alvo para estudos posteriores, quanto como modelo de
manipulação de N-glicosilação de β-xilosidases. Assim, será possível avançar no
desenvolvimento de excelentes catalizadores biológicos de acordo com a demanda
de processos biotecnológicos.

144
Referências
Adav, S. S., Ravindran, A., & Sze, S. K. (2014). Study of Phanerochaete
chrysosporium Secretome Revealed Protein Glycosylation as a Substrate-
Dependent Post-Translational Modification. Journal of Proteome Research,
13(10), 4272–4280. https://doi.org/10.1021/pr500385y
Adav, S. S., Ravindran, A., & Sze, S. K. (2015). Data for iTRAQ secretomic analysis
of Aspergillus fumigatus in response to different carbon sources. Data in Brief, 3,
175–179. https://doi.org/10.1016/j.dib.2015.03.001
Aebi, M. (2013). N-linked protein glycosylation in the ER. Biochimica et Biophysica
Acta - Molecular Cell Research, 1833(11), 2430–2437.
https://doi.org/10.1016/j.bbamcr.2013.04.001
Aebi, M., Bernasconi, R., Clerc, S., & Molinari, M. (2010). N-glycan structures:
recognition and processing in the ER. Trends in Biochemical Sciences, 35(2), 74–
82. https://doi.org/10.1016/j.tibs.2009.10.001
Alberts, B. Johnson, A. Lewis, J. Raff, M. Roberts, K. Walter, P. (2008). Molecular
Biology of the Cell, 5th Edition. Perspective.
Alvarez-Leefmans, F., & Delpire, E. (2010). Physiology and Pathology of Chloride
Transporters and Channels in the Nervous System. Physiology and Pathology of
chloride transporters and channels in the nervous system. Elsevier.
https://doi.org/10.1016/B978-0-12-374373-2.X0001-5
Amore, A., Knott, B. C., Supekar, N. T., Shajahan, A., Azadi, P., Zhao, P., … Taylor,
L. E. (2017). Distinct roles of N- and O-glycans in cellulase activity and stability.
Proceedings of the National Academy of Sciences, 114(52), 13667–13672.
https://doi.org/10.1073/pnas.1714249114
Anumula, K. R., & Taylor, P. B. (1992). A comprehensive procedure for preparation of
partially methylated alditol acetates from glycoprotein carbohydrates. Analytical
Biochemistry, 203(1), 101–108. https://doi.org/10.1016/0003-2697(92)90048-C
Archer, D. B., & Peberdy, J. F. (1997). The molecular biology of secreted enzyme

145
production by fungi. Critical Reviews in Biotechnology, 17(4), 273–306.
https://doi.org/10.3109/07388559709146616
Banerjee, S., Vishwanath, P., Cui, J., Kelleher, D. J., Gilmore, R., Robbins, P. W., &
Samuelson, J. (2007). The evolution of N-glycan-dependent endoplasmic
reticulum quality control factors for glycoprotein folding and degradation.
Proceedings of the National Academy of Sciences of the United States of America,
104(28), 11676–11681. https://doi.org/10.1073/pnas.0704862104
Bauer, S., Vasu, P., Persson, S., Mort, A. J., & Somerville, C. R. (2006). Development
and application of a suite of polysaccharide-degrading enzymes for analyzing
plant cell walls. Proceedings of the National Academy of Sciences of the United
States of America, 103(30), 11417–11422.
https://doi.org/10.1073/pnas.0604632103
Beausoleil, S. A., Villén, J., Gerber, S. A., Rush, J., & Gygi, S. P. (2006). A probability-
based approach for high-throughput protein phosphorylation analysis and site
localization. Nature Biotechnology, 24(10), 1285–92.
https://doi.org/10.1038/nbt1240
Beckham, G. T., Dai, Z., Matthews, J. F., Momany, M., Payne, C. M., Adney, W. S., …
Himmel, M. E. (2012a). Harnessing glycosylation to improve cellulase activity.
Current Opinion in Biotechnology, 23(3), 338–345.
https://doi.org/10.1016/j.copbio.2011.11.030
Beckham, G. T., Dai, Z., Matthews, J. F., Momany, M., Payne, C. M., Adney, W. S., …
Himmel, M. E. (2012b). Harnessing glycosylation to improve cellulase activity.
Current Opinion in Biotechnology, 23(3), 338–345.
https://doi.org/10.1016/j.copbio.2011.11.030
Bell, E., & Malmberg, R. L. (1990). Analysis of a cDNA encoding arginine
decarboxylase from oat reveals similarity to the Escherichia coli arginine
decarboxylase and evidence of protein processing. Molecular and General
Genetics MGG, 1(224), 431–436. Retrieved from
http://link.springer.com/article/10.1007/BF00262438

146
Benoit, I., Culleton, H., Zhou, M., DiFalco, M., Aguilar-Osorio, G., Battaglia, E., … de
Vries, R. P. (2015). Closely related fungi employ diverse enzymatic strategies to
degrade plant biomass. Biotechnology for Biofuels, 8(1), 107.
https://doi.org/10.1186/s13068-015-0285-0
Bhadauria, V., Zhao, W.-S. S., Wang, L.-X. X., Zhang, Y., Liu, J.-H. H., Yang, J., …
Peng, Y.-L. L. (2007). Advances in fungal proteomics. Microbiological Research,
162(3), 193–200. https://doi.org/10.1016/j.micres.2007.03.001
Biasini, M., Bienert, S., Waterhouse, A., Arnold, K., Studer, G., Schmidt, T., …
Schwede, T. (2014). SWISS-MODEL: Modelling protein tertiary and quaternary
structure using evolutionary information. Nucleic Acids Research, 42(W1).
https://doi.org/10.1093/nar/gku340
Borin, G. P., Sanchez, C. C., de Souza, A. P., de Santana, E. S., de Souza, A. T.,
Leme, A. F. P., … Oliveira, J. V. D. C. (2015). Comparative Secretome Analysis
of Trichoderma reesei and Aspergillus niger during Growth on Sugarcane
Biomass. Plos One, 10(6), e0129275.
https://doi.org/10.1371/journal.pone.0129275
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Analytical
Biochemistry, 72(1–2), 248–254. https://doi.org/10.1016/0003-2697(76)90527-3
Bulaj, G. (2005). Formation of disulfide bonds in proteins and peptides. Biotechnology
Advances, 23(1), 87–92. https://doi.org/10.1016/j.biotechadv.2004.09.002
Burda, P., & Aebi, M. (1999). The dolichol pathway of N-linked glycosylation.
Biochimica et Biophysica Acta - General Subjects, 1426(2), 239–257.
Camassola, M., & J.P. Dillon, A. (2012). Cellulase Determination: Modifications to
Make the Filter Paper Assay Easy, Fast, Practical and Efficient. Journal of
Analytical & Bioanalytical Techniques, 01(S1), 10–13.
https://doi.org/10.4172/scientificreports.125
Canilha, L., Rodrigues, R. D. C. L. B., Antunes, F. A. F., Milessi, T. S. D. S., Felipe, M.

147
D. G. A., & Silva, S. S. Da. (2013). Bioconversion of Hemicellulose from
Sugarcane Biomass Into Sustainable Products. In Sustainable Degradation of
Lignocellulosic Biomass - Techniques, Applications and Commercialization (pp.
15–45). https://doi.org/dx.doi.org/10.5772/53832
Cantarel, B. I., Coutinho, P. M., Rancurel, C., Bernard, T., Lombard, V., & Henrissat,
B. (2009). The Carbohydrate-Active EnZymes database (CAZy): An expert
resource for glycogenomics. Nucleic Acids Research, 37(SUPPL. 1), D233-8.
https://doi.org/10.1093/nar/gkn663
Casas-Flores, S., Rosales-saavedra, T., & Herrera-Estrella, A. (2004). Three decades
of fungal transformation: novel technologies. Methods in Molecular Biology
(Clifton, N.J.), 267, 315–325. https://doi.org/10.1385/1-59259-774-2:315
Case, M. E., Schweizer, M., Kushner, S. R., Giles, N. H., Kushnert, S. R., & Giles, N.
H. (1979). Efficient transformation of Neurospora crassa by utilizing hybrid plasmid
DNA. Proceedings of the National Academy of Sciences of the United States of
America, 76(10), 5259–5263. https://doi.org/10.1073/pnas.76.10.5259
Cerqueira, G. C., Arnaud, M. B., Inglis, D. O., Skrzypek, M. S., Binkley, G., Simison,
M., … Wortman, J. R. (2014). The Aspergillus Genome Database: Multispecies
curation and incorporation of RNA-Seq data to improve structural gene
annotations. Nucleic Acids Research. https://doi.org/10.1093/nar/gkt1029
Chai, R., Zhang, G., Sun, Q., Zhang, M., Zhao, S., & Qiu, L. (2013). Liposome-
mediated mycelial transformation of filamentous fungi. Fungal Biology, 117(9),
577–83. https://doi.org/10.1016/j.funbio.2013.06.008
Chakraborty, B. N., Patterson, N. A., & Kapoor, M. (1991). An electroporation-based
system for high-efficiency transformation of germinated conidia of filamentous
fungi. Can. J. Microbiol, 37(11), 858–863. https://doi.org/10.1139/m91-147
Chang, X., Xu, B., Bai, Y., Luo, H., Ma, R., Shi, P., & Yao, B. (2017). Role of N-linked
glycosylation in the enzymatic properties of a thermophilic GH 10 xylanase from
Aspergillus fumigatus expressed in Pichia pastoris. PLoS ONE, 12(2), 1–13.
https://doi.org/10.1371/journal.pone.0171111

148
Chen, W., Smeekens, J. M., & Wu, R. (2014). Comprehensive analysis of protein N-
glycosylation sites by combining chemical deglycosylation with LC-MS. Journal of
Proteome Research, 13(3), 1466–1473. https://doi.org/10.1021/pr401000c
Chen, W., Xie, T., Shao, Y., & Chen, F. (2012). Genomic characteristics comparisons
of 12 food-related filamentous fungi in tRNA gene set, codon usage and amino
acid composition. Gene, 497(1), 116–124.
https://doi.org/10.1016/j.gene.2012.01.016
Cheng, S., Edwards, S. a., Jiang, Y., & Gräter, F. (2010). Glycosylation Enhances
Peptide Hydrophobic Collapse by Impairing Solvation. ChemPhysChem, 11(11),
2367–2374. https://doi.org/10.1002/cphc.201000205
Cherepanova, N., Shrimal, S., & Gilmore, R. (2016). N-linked glycosylation and
homeostasis of the endoplasmic reticulum. Current Opinion in Cell Biology, 41,
57–65. https://doi.org/10.1016/j.ceb.2016.03.021
Chung, D., Cha, M., Guss, A. M., & Westpheling, J. (2014). Direct conversion of plant
biomass to ethanol by engineered Caldicellulosiruptor bescii. Proceedings of the
National Academy of Sciences of the United States of America, 111(24), 8931–6.
https://doi.org/10.1073/pnas.1402210111
Colley, K. J., Varki, A., & Kinoshita, T. (2015). Cellular Organization of Glycosylation.
Essentials of Glycobiology. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/28876808
Conesa, A., Jeenes, D., Archer, D. B., van den Hondel, C. A. M. J. J., & Punt, P. J.
(2002). Calnexin Overexpression Increases Manganese Peroxidase Production in
Aspergillus niger. Applied and Environmental Microbiology, 68(2), 846–851.
https://doi.org/10.1128/AEM.68.2.846-851.2002
Conesa, A., Punt, P. J., van Luijk, N., & van den Hondel, C. A. M. J. J. (2001). The
secretion pathway in filamentous fungi: a biotechnological view. Fungal Genetics
and Biology : FG & B, 33(3), 155–171. https://doi.org/10.1006/fgbi.2001.1276
Contesini, F. J., Liberato, M. V., Rubio, M. V., Calzado, F., Zubieta, M. P., Riaño-

149
Pachón, D. M., … Damasio, A. R. (2017a). Structural and functional
characterization of a highly secreted α- l -arabinofuranosidase (GH62) from
Aspergillus nidulans grown on sugarcane bagasse. Biochimica et Biophysica Acta
(BBA) - Proteins and Proteomics, 1865(12), 1758–1769.
https://doi.org/10.1016/j.bbapap.2017.09.001
Contesini, F. J., Liberato, M. V., Rubio, M. V., Calzado, F., Zubieta, M. P., Riaño-
Pachón, D. M., … Damasio, A. R. (2017b). Structural and functional
characterization of a highly secreted α-L-arabinofuranosidase (GH62) from
Aspergillus nidulans grown on sugarcane bagasse. Biochimica et Biophysica Acta
- Proteins and Proteomics, 1865(12), 1758–1769.
https://doi.org/10.1016/j.bbapap.2017.09.001
Cooper, C. A., Gasteiger, E., & Packer, N. H. (2001). GlycoMod - A software tool for
determining glycosylation compositions from mass spectrometric data.
Proteomics, 1(2), 340–349. https://doi.org/10.1002/1615-
9861(200102)1:2<340::AID-PROT340>3.0.CO;2-B
Cota, J., Alvarez, T. M., Citadini, A. P., Santos, C. R., de Oliveira Neto, M., Oliveira, R.
R., … Squina, F. M. (2011). Mode of operation and low-resolution structure of a
multi-domain and hyperthermophilic endo-β-1,3-glucanase from Thermotoga
petrophila. Biochemical and Biophysical Research Communications, 406(4), 590–
594. https://doi.org/10.1016/j.bbrc.2011.02.098
Cowcher, D. P., Deckert-Gaudig, T., Brewster, V. L., Ashton, L., Deckert, V., &
Goodacre, R. (2016). Detection of Protein Glycosylation Using Tip-Enhanced
Raman Scattering. Analytical Chemistry, 88(4), 2105–2112.
https://doi.org/10.1021/acs.analchem.5b03535
Daly, P., Munster, J. M. Van, Raulo, R., & Archer, D. B. (2015). Transcriptional
Regulation and Responses in Filamentous Fungi Exposed to Lignocellulose. In
Mycology: Current and Future Developments (Vol. 1, pp. 82–127).
Dam, S., Thaysen-Andersen, M., Stenkjær, E., Lorentzen, A., Roepstorff, P., Packer,
N. H., & Stougaard, J. (2013). Combined N-glycome and N-glycoproteome

150
analysis of the Lotus japonicus seed globulin fraction shows conservation of
protein structure and glycosylation in legumes. Journal of Proteome Research,
12(7), 3383–92. https://doi.org/10.1021/pr400224s
Davis, B. G., Van Kasteren, S. I., Kramer, H. B., & Gamblin, D. P. (2007). Site-selective
glycosylation of proteins: Creating synthetic glycoproteins. Nature Protocols,
2(12), 3185–3194. https://doi.org/10.1038/nprot.2007.430
de Groot, P. W. J., Brandt, B. W., Horiuchi, H., Ram, A. F. J., de Koster, C. G., & Klis,
F. M. (2009). Comprehensive genomic analysis of cell wall genes in Aspergillus
nidulans. Fungal Genetics and Biology : FG & B, 46(1), S72–S81.
https://doi.org/10.1016/j.fgb.2008.07.022
de Souza, W. R., de Gouvea, P. F., Savoldi, M., Malavazi, I., de Souza Bernardes, L.
a, Goldman, M. H. S., … Goldman, G. H. (2011). Transcriptome analysis of
Aspergillus niger grown on sugarcane bagasse. Biotechnology for Biofuels, 4(1),
40. https://doi.org/10.1186/1754-6834-4-40
Decker, S. R., Siika-Aho, M., & Viikari, L. (2009). Enzymatic Depolymerization of Plant
Cell Wall Hemicelluloses. In Biomass Recalcitrance: Deconstructing the Plant Cell
Wall for Bioenergy (pp. 352–373). Blackwell Publishing Ltd.
https://doi.org/10.1002/9781444305418.ch10
Delmas, S., Pullan, S. T., Gaddipati, S., Kokolski, M., Malla, S., Blythe, M. J., … Archer,
D. B. (2012). Uncovering the Genome-Wide Transcriptional Responses of the
Filamentous Fungus Aspergillus niger to Lignocellulose Using RNA Sequencing.
PLoS Genetics, 8(8), e1002875. https://doi.org/10.1371/journal.pgen.1002875
Deshpande, N., Wilkins, M. R., Packer, N., & Nevalainen, H. (2008). Protein
glycosylation pathways in filamentous fungi. Glycobiology, 18(8), 626–637.
https://doi.org/10.1093/glycob/cwn044
Desko, M. M., Gross, D. A., & Kohler, J. J. (2009). Effects of N-glycosylation on the
activity and localization of GlcNAc-6-sulfotransferase 1. Glycobiology, 19(10),
1068–1077. https://doi.org/10.1093/glycob/cwp092

151
Devchand, M., & Gwynne, D. I. (1991). Expression of heterologous proteins in
Aspergillus. Journal of Biotechnology, 17(1), 3–9. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/16814894
Dhawale, S. S., Paietta, J. V, & Marzluf, G. A. (1984). A new, rapid and efficient
transformation procedure for Neurospora. Current Genetics, 8(1), 77–79.
https://doi.org/10.1007/BF00405435
Díaz-Malváez, F. I., García-Almendárez, B. E., Hernández-Arana, A., Amaro-Reyes,
A., & Regalado-González, C. (2013). Isolation and properties of β-xylosidase from
Aspergillus niger GS1 using corn pericarp upon solid state fermentation. Process
Biochemistry, 48(7), 1018–1024. https://doi.org/10.1016/j.procbio.2013.05.003
Dimarogona, M., Topakas, E., Olsson, L., & Christakopoulos, P. (2012). Lignin boosts
the cellulase performance of a GH-61 enzyme from Sporotrichum thermophile.
Bioresource Technology, 110, 480–487.
https://doi.org/10.1016/j.biortech.2012.01.116
Duden, R. (2003). ER-to-Golgi transport: COP I and COP II function. Molecular
Membrane Biology. https://doi.org/10.1080/0968768031000122548
Eshghi, S. T., Yang, S., & Wang, X. (2014). Imaging of N-Linked Glycans from
Formalin-Fixed Paraffin-Embedded Tissue Sections Using MALDI Mass
Spectrometry. ACS Chemical Biology. Retrieved from
http://pubs.acs.org/doi/abs/10.1021/cb500405h
Eveleigh, D. E., Mandels, M., Andreotti, R., & Roche, C. (2009). Measurement of
saccharifying cellulase. Biotechnology for Biofuels, 2(1), 21.
https://doi.org/10.1186/1754-6834-2-21
Fernandes, P. (2010). Enzymes in Food Processing: A Condensed Overview on
Strategies for Better Biocatalysts. Enzyme Research, 2010, 1–19.
https://doi.org/10.4061/2010/862537
Fernández-Abalos, J. M., Fox, H., Pitt, C., Wells, B., & Doonan, J. H. (1998). Plant-
adapted green fluorescent protein is a versatile vital reporter for gene expression,

152
protein localization and mitosis in the filamentous fungus, Aspergillus nidulans.
Molecular Microbiology, 27(1), 121–130. https://doi.org/10.1111/j.1365-
2958.2009.06936.x
Fernandez, J.M. & Hoeffler, J.P., E. (1999). Gene Expression Systems. Using nature
for the art of expression. Retrieved from
http://www.embl.de/pepcore/pepcore_services/cloning/choice_expression_syste
ms/comparison_expression_systems/index.html
Fischer-Parton, S., Parton, R. M., Hickey, P. C., Dijksterhuis, J., Atkinson, H. A., &
Read, N. D. (2000). Confocal microscopy of FM4-64 as a tool for analysing
endocytosis and vesicle trafficking in living fungal hyphae. In Journal of
Microscopy (Vol. 198, pp. 246–259). https://doi.org/10.1046/j.1365-
2818.2000.00708.x
Flamm, E. L. (1991). How FDA approved chymosin: A case history. Bio/Technology,
9(4), 349–351. https://doi.org/10.1038/nbt0491-349
Fleiβner, A., & Dersch, P. (2010). Expression and export: Recombinant protein
production systems for Aspergillus. Applied Microbiology and Biotechnology,
87(4), 1255–70. https://doi.org/10.1007/s00253-010-2672-6
Gagneux, P., Aebi, M., & Varki, A. (2015). Evolution of Glycan Diversity. Essentials of
Glycobiology. https://doi.org/28876829
Galagan, J. E., Calvo, S. E., Cuomo, C., Ma, L.-J., Wortman, J. R., Batzoglou, S., …
Birren, B. W. (2005). Sequencing of Aspergillus nidulans and comparative
analysis with A. fumigatus and A. oryzae. Nature, 438(7071), 1105–1115.
https://doi.org/10.1038/nature04341
Gamblin, D. P., Garnier, P., van Kasteren, S., Oldham, N. J., Fairbanks, A. J., & Davis,
B. G. (2004). Glyco-SeS: selenenylsulfide-mediated protein glycoconjugation--a
new strategy in post-translational modification. Angewandte Chemie (International
Ed. in English), 43(7), 828–33. https://doi.org/10.1002/anie.200352975
Gardner, J. G., Crouch, L., Labourel, A., Forsberg, Z., Bukhman, Y. V., Vaaje-Kolstad,

153
G., … Keating, D. H. (2014). Systems biology defines the biological significance
of redox-active proteins during cellulose degradation in an aerobic bacterium.
Molecular Microbiology, 94(5), 1121–1133. https://doi.org/10.1111/mmi.12821
Geyer, H., & Geyer, R. (2006). Strategies for analysis of glycoprotein glycosylation.
Biochimica et Biophysica Acta, 1764(12), 1853–69.
https://doi.org/10.1016/j.bbapap.2006.10.007
Geysens, S., Whyteside, G., & Archer, D. B. (2009). Genomics of protein folding in the
endoplasmic reticulum, secretion stress and glycosylation in the aspergilli. Fungal
Genetics and Biology, 46(1), S121–S140.
https://doi.org/10.1016/j.fgb.2008.07.016
Ghaffar, S. H., & Fan, M. (2013). Structural analysis for lignin characteristics in biomass
straw. Biomass and Bioenergy, 57, 264–279.
https://doi.org/10.1016/j.biombioe.2013.07.015
Glass, N. L., Schmoll, M., Cate, J. H. D., & Coradetti, S. (2013). Plant cell wall
deconstruction by ascomycete fungi. Annual Review of Microbiology, 67(June),
477–98. https://doi.org/10.1146/annurev-micro-092611-150044
Goedegebuur, F., Dankmeyer, L., Gualfetti, P., Karkehabadi, S., Hansson, H., Jana,
S., … Sandgren, M. (2017). Improving the thermal stability of cellobiohydrolase
Cel7A from Hypocrea jecorina by directed evolution. Journal of Biological
Chemistry, jbc.M117.803270. https://doi.org/10.1074/jbc.M117.803270
Goettig, P. (2016). Effects of glycosylation on the enzymatic activity and mechanisms
of proteases. International Journal of Molecular Sciences, 17(12), 1–24.
https://doi.org/10.3390/ijms17121969
Goochee, C. F., & Monica, T. (1990). Environmental effects on protein glycosylation.
Nature Biotechnology, 8(5), 421–7. https://doi.org/10.1038/nbt0898-773
Gouka, R. J., Punt, P. J., & van den Hondel, C. A. M. J. J. (1997). Efficient production
of secreted proteins by Aspergillus : progress, limitations and prospects. Applied
Microbiology and Biotechnology, 47(1), 1–11.

154
https://doi.org/10.1007/s002530050880
Gouka, R. J., Punt, P. J., & van den Hondel, C. A. M. J. J. (1997). Glucoamylase Gene
Fusions Alleviate Limitations for Protein Production in Aspergillus awamori at the
Transcriptional and ( Post ) Translational Levels. Applied and Environmental
Microbiology, 63(2), 488–497.
Groot, M. J. A. de, & Bundock, P. (1998). Agrobacterium tumefaciens-mediated
transformation of flamentous fungi. Nature Biotechnology, 16, 839–842.
Gupta, R., Baldock, S. J., Fielden, P. R., & Grieve, B. D. (2011). Capillary zone
electrophoresis for the analysis of glycoforms of cellobiohydrolase. Journal of
Chromatography. A, 1218(31), 5362–8.
https://doi.org/10.1016/j.chroma.2011.06.036
Gupta, R., & Brunak, S. (2002). Prediction of glycosylation across the human proteome
and the correlation to protein function. Pacific Symposium on Biocomputing., 322,
310–322. https://doi.org/10.1142/9789812799623_0029
Gusakov, A. V. (2011). Alternatives to Trichoderma reesei in biofuel production. Trends
in Biotechnology, 29, 419–425. https://doi.org/10.1016/j.tibtech.2011.04.004
Gusakov, A. V., Dotsenko, A. S., Rozhkova, A. M., & Sinitsyn, A. P. (2017). N-Linked
glycans are an important component of the processive machinery of
cellobiohydrolases. Biochimie, 132, 102–108.
https://doi.org/10.1016/j.biochi.2016.11.004
Gustafsson, C., Govindarajan, S., & Minshull, J. (2004). Codon bias and heterologous
protein expression. Trends in Biotechnology, 22(7), 346–353.
https://doi.org/10.1016/j.tibtech.2004.04.006
Hanson, S. R., Culyba, E. K., Hsu, T., Wong, C.-H., Kelly, J. W., & Powers, E. T. (2009).
The core trisaccharide of an N-linked glycoprotein intrinsically accelerates folding
and enhances stability. Proceedings of the National Academy of Sciences of the
United States of America, 106(9), 3131–3136.
https://doi.org/10.1073/pnas.0810318105

155
Harris, S. D. (2008). Branching of fungal hyphae: regulation, mechanisms and
comparison with other branching systems. Mycologia, 100(6), 823–832.
https://doi.org/10.3852/08-177
Heimel, K. (2014). Unfolded protein response in filamentous fungi-implications in
biotechnology. Applied Microbiology and Biotechnology, 99(1), 121–132.
https://doi.org/10.1007/s00253-014-6192-7
Helenius, A., & Aebi, M. (2004). Roles of N-linked glycans in the endoplasmic
reticulum. Annual Review of Biochemistry, 73, 1019–49.
https://doi.org/10.1146/annurev.biochem.73.011303.073752
Hentze, M. W. (1991). Determinants and regulation of cytoplasmic mRNA stability in
eukaryotic cells. Biochimica et Biophysica Acta - Gene Structure and Expression,
1090(3), 281–292. https://doi.org/10.1016/0167-4781(91)90191-N
Horn, S. J., Vaaje-Kolstad, G., Westereng, B., & Eijsink, V. G. (2012). Novel enzymes
for the degradation of cellulose. Biotechnology for Biofuels, 5(1), 45.
https://doi.org/10.1186/1754-6834-5-45
Hoyt, M. A., Williams-Abbott, L. J., Pitkin, J. W., & Davis, R. H. (2000). Cloning and
expression of the S-adenosylmethionine decarboxylase gene of Neurospora
crassa and processing of its product. Molecular & General Genetics : MGG,
263(4), 664–673. https://doi.org/10.1007/s004380051215
Huang, Y.-W., Yang, H.-I., Wu, Y.-T., Hsu, T.-L., Lin, T.-W., Kelly, J. W., & Wong, C.-
H. (2017). Residues Comprising the Enhanced Aromatic Sequon Influence Protein
N-glycosylation Efficiency. Journal of the American Chemical Society,
jacs.7b03868. https://doi.org/10.1021/jacs.7b03868
Hynes, M. J. (1996). Genetic transformation of filamentous fungi. Journal of Genetics,
75(3), 297–311. https://doi.org/10.1007/BF02966310
Imperiali, B., & Connor, S. E. O. (1999). Effect of N -linked glycosylation on
glycopeptide and glycoprotein structure, 9, 643–649.
Jacobs, P. P., & Callewaert, N. (2009). N-glycosylation engineering of

156
biopharmaceutical expression systems. Current Molecular Medicine, 9(7), 774–
800. https://doi.org/10.2174/156652409789105552
Jagadeeswaran, G., Gainey, L., Prade, R., & Mort, A. J. (2016). A family of AA9 lytic
polysaccharide monooxygenases in Aspergillus nidulans is differentially regulated
by multiple substrates and at least one is active on cellulose and xyloglucan.
Applied Microbiology and Biotechnology, 100(10), 4535–4547.
https://doi.org/10.1007/s00253-016-7505-9
Jarrell, K. F., Ding, Y., Meyer, B. H., Albers, S.-V., Kaminski, L., & Eichler, J. (2014).
N-Linked Glycosylation in Archaea: a Structural, Functional, and Genetic Analysis.
Microbiology and Molecular Biology Reviews, 78(2), 304–341.
https://doi.org/10.1128/MMBR.00052-13
Jeenes, D. J., Mackenzie, D. A., Roberts, I. N., & Archer, D. B. (1991). Heterologous
protein production by filamentous fungi. Biotechnology & Genetic Engineering
Reviews, 9(1), 327–67. https://doi.org/10.1080/02648725.1991.10647884
Jensen, O. N. (2004). Modification-specific proteomics: Characterization of post-
translational modifications by mass spectrometry. Current Opinion in Chemical
Biology, 8(1), 33–41. https://doi.org/10.1016/j.cbpa.2003.12.009
Jeoh, T., Michener, W., Himmel, M. E., Decker, S. R., & Adney, W. S. (2008).
Implications of cellobiohydrolase glycosylation for use in biomass conversion.
Biotechnology for Biofuels, 1(1), 10. https://doi.org/10.1186/1754-6834-1-10
Jordan, D. B., & Wagschal, K. (2010). Properties and applications of microbial β-D-
xylosidases featuring the catalytically efficient enzyme from Selenomonas
ruminantium. Applied Microbiology and Biotechnology.
https://doi.org/10.1007/s00253-010-2538-y
Jouzani, G. S., & Taherzadeh, M. J. (2015). Advances in consolidated bioprocessing
systems for bioethanol and butanol production from biomass: a comprehensive
review. Biofuel Research Journal, 5, 152–195.
Kainz, E., Gallmetzer, A., Hatzl, C., Nett, J. H., Li, H., Schinko, T., … Strauss, J. (2008).

157
N-glycan modification in Aspergillus species. Applied and Environmental
Microbiology, 74(4), 1076–1086. https://doi.org/10.1128/AEM.01058-07
Kamm, B., Kamm, M., Schmidt, M., Hirth, T., & Schulze, M. (2006). Lignocellulose-
based Chemical Products and Product Family Trees. Biorefineries-Industrial
Processes and Products (Vol. 2). https://doi.org/10.1002/9783527619849.ch20
Kawasaki, L., Kawasaki, L., Wysong, D., Wysong, D., Diamond, R., Diamond, R., …
Notably, R. (1997). Two Divergent Catalase Genes Are Differentially Regulated
during Aspergillus nidulans Development and Oxidative Stress. Microbiology,
179(10), 3284–3292.
Keller, A., Nesvizhskii, A. I., Kolker, E., & Aebersold, R. (2002). Empirical Statistical
Model To Estimate the Accuracy of Peptide Identifications Made by MS/MS and
Database Search. Analytical Chemistry, 74(20), 5383–5392.
https://doi.org/10.1021/ac025747h
Kelly, S. M., Jess, T. J., & Price, N. C. (2005). How to study proteins by circular
dichroism. Biochimica et Biophysica Acta, 1751(2), 119–39.
https://doi.org/10.1016/j.bbapap.2005.06.005
Kirk, O., Borchert, T. V., & Fuglsang, C. C. (2002). Industrial enzyme applications.
Current Opinion in Biotechnology, 13(4), 345–351. https://doi.org/10.1016/S0958-
1669(02)00328-2
Kruszewska, J. S., Perlińska-Lenart, U., Górka-Nieć, W., Orłowski, J., Zembek, P., &
Palamarczyk, G. (2008). Alterations in protein secretion caused by metabolic
engineering of glycosylation pathways in fungi. Acta Biochimica Polonica, 55(3),
447–56. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/18797519
Kumar, P., Barrett, D. M., Delwiche, M. J., & Stroeve, P. (2009). Methods for
pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel
production. Industrial and Engineering Chemistry Research, 48(8), 3713–3729.
https://doi.org/10.1021/ie801542g
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head

158
of bacteriophage T4. Nature, 227(5259), 680–685.
https://doi.org/10.1038/227680a0
Larkin, A., & Imperiali, B. (2011). The expanding horizons of asparagine-linked
glycosylation. Biochemistry, 50(21), 4411–26. https://doi.org/10.1021/bi200346n
Leach, M. D., & Brown, A. J. P. (2012). Posttranslational modifications of proteins in
the pathobiology of medically relevant fungi. Eukaryotic Cell, 11(2), 98–108.
https://doi.org/10.1128/EC.05238-11
Lee, L. Y., Moh, E. S. X., Parker, B. L., Bern, M., Packer, N. H., & Thaysen-Andersen,
M. (2016). Toward Automated N-Glycopeptide Identification in Glycoproteomics.
Journal of Proteome Research, 15(10), 3904–3915.
https://doi.org/10.1021/acs.jproteome.6b00438
Levasseur, A., Drula, E., Lombard, V., Coutinho, P. M., & Henrissat, B. (2013).
Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary
redox enzymes. Biotechnology for Biofuels, 6(1), 41. https://doi.org/10.1186/1754-
6834-6-41
Li, K., Ouyang, H., Lü, Y., Liang, J., Wilson, I. B. H., & Jin, C. (2011). Repression of N-
glycosylation triggers the unfolded protein response (UPR) and overexpression of
cell wall protein and chitin in aspergillus fumigatus. Microbiology, 157(7), 1968–
1979. https://doi.org/10.1099/mic.0.047712-0
Li, S. T., Wang, N., Xu, S., Yin, J., Nakanishi, H., Dean, N., & Gao, X. D. (2017).
Quantitative study of yeast Alg1 beta-1, 4 mannosyltransferase activity, a key
enzyme involved in protein N-glycosylation. Biochimica et Biophysica Acta -
General Subjects, 1861(1), 2934–2941.
https://doi.org/10.1016/j.bbagen.2016.09.023
Liu, B., Gong, X., Chang, S., Yang, Y., Song, M., Duan, D., … Wu, J. (2009). Disruption
of the OCH1 and MNN1 genes decrease N-glycosylation on glycoprotein
expressed in Kluyveromyces lactis. Journal of Biotechnology, 143(2), 95–102.
https://doi.org/10.1016/j.jbiotec.2009.06.016

159
Liu, G., Qin, Y., Li, Z., & Qu, Y. (2013). Development of highly efficient, low-cost
lignocellulolytic enzyme systems in the post-genomic era. Biotechnology
Advances, 31(6), 962–975. https://doi.org/10.1016/j.biotechadv.2013.03.001
Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P. M., & Henrissat, B. (2014).
The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids
Research, 42(D1), D490-5. https://doi.org/10.1093/nar/gkt1178
Lu, D., Yang, C., & Liu, Z. (2012). How hydrophobicity and the glycosylation site of
glycans affect protein folding and stability: A molecular dynamics simulation.
Journal of Physical Chemistry B, 116(1), 390–400.
https://doi.org/10.1021/jp203926r
Lubertozzi, D., & Keasling, J. D. (2009). Developing Aspergillus as a host for
heterologous expression. Biotechnology Advances, 27(1), 53–75.
https://doi.org/10.1016/j.biotechadv.2008.09.001
Maattanen, P., Kozlov, G., Gehring, K., & Thomas, D. Y. (2006). ERp57 and PDI:
multifunctional protein disulfide isomerases with similar domain architectures but
differing substrate–partner associationsThis paper is one of a selection of papers
published in this Special Issue, entitled CSBMCB — Membrane Proteins in He.
Biochemistry and Cell Biology, 84(6), 881–889. https://doi.org/10.1139/o06-186
Magaña-Ortíz, D., Coconi-Linares, N., Ortiz-Vazquez, E., Fernández, F., Loske, A. M.,
& Gómez-Lim, M. A. (2013). A novel and highly efficient method for genetic
transformation of fungi employing shock waves. Fungal Genetics and Biology : FG
& B, 56(April), 9–16. https://doi.org/10.1016/j.fgb.2013.03.008
Maloy, S., & Schaechter, M. (2006). The era of microbiology: A Golden Phoenix.
International Microbiology. https://doi.org/im2306001 [pii]
Maras, M., Van Die, I., Contreras, R., & van den Hondel, C. A. M. J. J. (1999).
Filamentous fungi as production organisms for glycoproteins of bio-medical
interest. Glycoconjugate Journal, 16(2), 99–107.
https://doi.org/10.1023/A:1026436424881

160
Mathewson, N., Toubai, T., Kapeles, S., Sun, Y. Y., Oravecz-Wilson, K., Tamaki, H.,
… Reddy, P. (2013). Neddylation plays an important role in the regulation of
murine and human dendritic cell function. Blood, 122(12), 2062–2073.
https://doi.org/10.1182/blood-2013-02-486373
Mayor, S., & Riezman, H. (2004). Sorting GPI-anchored proteins. Nature Reviews.
Molecular Cell Biology, 5(2), 110–120. https://doi.org/10.1038/nrm1309
McCracken, A. A., & Brodsky, J. L. (2000). A molecular portrait of the response to
unfolded proteins. Genome Biology, 1(2), REVIEWS1013.
https://doi.org/10.1186/gb-2000-1-2-reviews1013
Meyer, V., Wu, B., & Ram, A. F. J. (2011). Aspergillus as a multi-purpose cell factory:
Current status and perspectives. Biotechnology Letters, 33(3), 469–476.
https://doi.org/10.1007/s10529-010-0473-8
Moremen, K. W. K., Tiemeyer, M., & Nairn, A. V. A. (2012). Vertebrate protein
glycosylation: diversity, synthesis and function. Nature Reviews. Molecular Cell
Biology, 13(7), 448–462. https://doi.org/10.1038/nrm3383
Moriya, K., Nagatoshi, K., Noriyasu, Y., Okamura, T., Takamitsu, E., Suzuki, T., &
Utsumi, T. (2013). Protein N-myristoylation plays a critical role in the endoplasmic
reticulum morphological change induced by overexpression of protein lunapark,
an integral membrane protein of the endoplasmic reticulum. PLoS ONE, 8(11), 1–
13. https://doi.org/10.1371/journal.pone.0078235
Murphy, C., Powlowski, J., Wu, M., Butler, G., & Tsang, A. (2011). Curation of
characterized glycoside hydrolases of Fungal origin. Database, 2011, 1–14.
https://doi.org/10.1093/database/bar020
Nagae, M., & Yamaguchi, Y. (2012). Function and 3D Structure of the N-Glycans on
Glycoproteins. International Journal of Molecular Sciences, 13(12), 8398–8429.
https://doi.org/10.3390/ijms13078398
Nakamura, Y., Gojobori, T., & Ikemura, T. (1998). Codon usage tabulated from
international DNA sequence databases: status for the year 2000. Nucleic Acids

161
Research, 26(1), 334. https://doi.org/10.1093/nar/26.1.334
Navarrete, M., Callegari, E., & Eyzaguirre, J. (2012). The effect of acetylated xylan and
sugar beet pulp on the expression and secretion of enzymes by Penicillium
purpurogenum. Applied Microbiology and Biotechnology, 93(2), 723–741.
https://doi.org/10.1007/s00253-011-3744-y
Nayak, T., Szewczyk, E., Oakley, C. E., Osmani, A., Ukil, L., Murray, S. L., … Oakley,
B. R. (2006). A versatile and efficient gene-targeting system for Aspergillus
nidulans. Genetics, 172(3), 1557–66.
https://doi.org/10.1534/genetics.105.052563
Nesvizhskii, A. I., Keller, A., Kolker, E., & Aebersold, R. (2003). A Statistical Model for
Identifying Proteins by Tandem Mass Spectrometry abilities that proteins are
present in a sample on the basis. Analytical Chemistry, 75(17), 4646–4658.
https://doi.org/10.1021/ac0341261
Nevalainen, H., & Peterson, R. (2014). Making recombinant proteins in filamentous
fungi- Are we expecting too much? Frontiers in Microbiology, 5(FEB), 75.
https://doi.org/10.3389/fmicb.2014.00075
Nevalainen, K. M. H., Te’o, V. S. J., & Bergquist, P. L. (2005). Heterologous protein
expression in filamentous fungi. Trends in Biotechnology, 23(9), 468–74.
https://doi.org/10.1016/j.tibtech.2005.06.002
Nishihara, S., Ueda, R., Goto, S., Toyoda, H., Ishida, H., & Nakamura, M. (2004).
Approach for functional analysis of glycan using RNA interference. Glycoconj J,
21(1–2), 63–68. https://doi.org/5277453
[pii]\r10.1023/B:GLYC.0000043750.80389.14
Nutzmann, H.-W., Reyes-Dominguez, Y., Scherlach, K., Schroeckh, V., Horn, F.,
Gacek, A., … Brakhage, A. A. (2011). Bacteria-induced natural product formation
in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone
acetylation. Proceedings of the National Academy of Sciences, 108(34), 14282–
14287. https://doi.org/10.1073/pnas.1103523108

162
Pasternak, J. J. (2005). Recombinant DNA Technology. In An Introduction to Human
Molecular Genetics (Second, pp. 107–152). Hoboken, NJ, USA: John Wiley &
Sons, Inc. https://doi.org/10.1002/0471719188.ch5
Patel, T., Bruce, J., Merry, a, Bigge, C., Wormald, M., Jaques, a, & Parekh, R. (1993).
Use of hydrazine to release in intact and unreduced form both N- and O-linked
oligosaccharides from glycoproteins. Biochemistry, 32(2), 679–693.
Pentillä, M., Limón, C., Nevalainen, H., Penttilä, M., Limón, C., Nevalainen, H., …
Nevalainen, H. (2003). Molecular biology of Trichoderma and biotechnological
applications. In D. K. Arora (Ed.), Handbook of Fungal Biotechnology (2nd ed.,
Vol. 20031860). CRC Press. https://doi.org/10.1201/9780203027356.ch31
Peter-Katalinić, J. (2005). Methods in enzymology: O-glycosylation of proteins.
Methods in Enzymology, 405(05), 139–71. https://doi.org/10.1016/S0076-
6879(05)05007-X
Peterson, R., & Nevalainen, H. (2012). Trichoderma reesei RUT-C30 - thirty years of
strain improvement. Microbiology, 158(1), 58–68.
https://doi.org/10.1099/mic.0.054031-0
Petrescu, A.-J. A.-J., Milac, A.-L., Petrescu, S. M., Dwek, R. A., & Wormald, M. R.
(2004). Statistical analysis of the protein environment of N-glycosylation sites:
implications for occupancy, structure, and folding. Glycobiology, 14(2), 103–14.
https://doi.org/10.1093/glycob/cwh008
Phillips, C. M., Beeson, W. T., Cate, J. H., & Marletta, M. A. (2011). Cellobiose
dehydrogenase and a copper-dependent polysaccharide monooxygenase
potentiate cellulose degradation by Neurospora crassa. ACS Chemical Biology,
6(12), 1399–1406. https://doi.org/10.1021/cb200351
Pickart, C. M., & Eddins, M. J. (2004). Ubiquitin: Structures, functions, mechanisms.
Biochimica et Biophysica Acta - Molecular Cell Research, 1695(1–3), 55–72.
https://doi.org/10.1016/j.bbamcr.2004.09.019
Pontecorvo, G., Roper, J. A., Chemmons, L. M., Macdonald, K. D., & Bufton, A. W. J.

163
(1953). The Genetics of Aspergillus nidulans. Advances in Genetics, 5(C), 141–
238. https://doi.org/10.1016/S0065-2660(08)60408-3
Price, J. L., Culyba, E. K., Chen, W., Murray, A. N., Hanson, S. R., Wong, C. H., …
Kelly, J. W. (2012). N-glycosylation of enhanced aromatic sequons to increase
glycoprotein stability. Biopolymers, 98(3), 195–211.
https://doi.org/10.1002/bip.22030
Price, J. L., Powers, D. L., Powers, E. T., & Kelly, J. W. (2011). Glycosylation of the
enhanced aromatic sequon is similarly stabilizing in three distinct reverse turn
contexts. Proceedings of the National Academy of Sciences of the United States
of America, 108(34), 14127–14132. https://doi.org/10.1073/pnas.1105880108
Punt, P. J., van Biezen, N., Conesa, A., Albers, A., Mangnus, J., & van den Hondel, C.
A. M. J. J. (2002). Filamentous fungi as cell factories for heterologous protein
production. Trends in Biotechnology, 20(5), 200–6. https://doi.org/10.1016/S0167-
7799(02)01933-9
Qi, F., Zhang, W., Zhang, F., Chen, G., & Liu, W. (2014). Deciphering the effect of the
different N-glycosylation sites on the secretion, activity, and stability of
cellobiohydrolase I from Trichoderma reesei. Applied and Environmental
Microbiology, 80(13), 3962–3971. https://doi.org/10.1128/AEM.00261-14
Ramírez, A. S., Boilevin, J., Lin, C. W., Ha Gan, B., Janser, D., Aebi, M., … Locher, K.
P. (2017). Chemo-enzymatic synthesis of lipid-linked
GlcNAc2Man5oligosaccharides using recombinant Alg1, Alg2 and Alg11 proteins.
Glycobiology, 27(8), 726–733. https://doi.org/10.1093/glycob/cwx045
Rao, R. S. P., & Bernd, W. (2010). Do N-glycoproteins have preference for specific
sequons? Bioinformation, 5(5), 208–212.
https://doi.org/10.6026/97320630005208
Ribeiro, D. a, Cota, J., Alvarez, T. M., Brüchli, F., Bragato, J., Pereira, B. M. P., …
Squina, F. M. (2012). The Penicillium echinulatum secretome on sugar cane
bagasse. PloS One, 7(12), e50571. https://doi.org/10.1371/journal.pone.0050571

164
Rivera, A. L., Magaña-Ortíz, D., Gómez-Lim, M., Fernández, F., & Loske, A. M. (2014).
Physical methods for genetic transformation of fungi and yeast. Physics of Life
Reviews, 11(2), 184–203. https://doi.org/10.1016/j.plrev.2014.01.007
Rocha, G. J. M., Gonçalves, A. R., Oliveira, B. R., Olivares, E. G., & Rossell, C. E. V.
(2012). Steam explosion pretreatment reproduction and alkaline delignification
reactions performed on a pilot scale with sugarcane bagasse for bioethanol
production. Industrial Crops and Products, 35(1), 274–279.
https://doi.org/10.1016/j.indcrop.2011.07.010
Rodriguez-Zuniga, U. F., Cannella, D., Giordano, R. D. C., Giordano, R. D. L. C.,
Jørgensen, H., & Felby, C. (2015). Lignocellulose pretreatment technologies affect
the level of enzymatic cellulose oxidation by LPMO. Green Chem., 2896–2903.
https://doi.org/10.1039/C4GC02179G
Rogers, L. D., & Overall, C. M. (2013). Proteolytic post-translational modification of
proteins: proteomic tools and methodology. Molecular & Cellular Proteomics :
MCP, 12(12), 3532–42. https://doi.org/10.1074/mcp.M113.031310
Rojas, A. L., Fischer, H., Eneiskaya, E. V., Kulminskaya, A. A., Shabalin, K. A.,
Neustroev, K. N., … Polikarpov, I. (2005). Structural insights into the β-xylosidase
from Trichoderma reesei obtained by synchrotron small-angle x-ray scattering and
circular dichroism spectroscopy. Biochemistry, 44(47), 15578–15584.
https://doi.org/10.1021/bi050826j
Ron, D., & Walter, P. (2007). Signal integration in the endoplasmic reticulum unfolded
protein response. Nature Reviews. Molecular Cell Biology, 8(7), 519–529.
https://doi.org/10.1038/nrm2199
Rosa, M. F., Souza Filho, M. S. M., Figueiredo, M. C. B., Morais, J. P. S., Santaella,
S. T., & Leitão, R. C. (2011). Valorização De Resíduos Da Agroindústria. II
Simpósio Internacional Sobre Gerenciamento de Resíduos Agropecuários e
Agroindustriais, I, 98–105. https://doi.org/10.1021/jf050995a
Rubio, C., Pincus, D., Korennykh, A., Schuck, S., El-Samad, H., & Walter, P. (2011).
Homeostatic adaptation to endoplasmic reticulum stress depends on Ire1 kinase

165
activity. Journal of Cell Biology, 193, 171–184.
https://doi.org/10.1083/jcb.201007077
Rubio, M. V., Tramontina, R., Gonçalves, T. A., Uchima, C. A., Segato, F., Squina, F.
M., & Damasio, A. R. de L. (2015). Mycology: Current and Future Developments.
(R. Silva, Ed.), Mycology: Current and Future Developments (Vol. 1). BENTHAM
SCIENCE PUBLISHERS. https://doi.org/10.2174/97816810807411150101
Rubio, M. V., Zubieta, M. P., Franco Cairo, J. P. L., Calzado, F., Paes Leme, A. F.,
Squina, F. M., … de Lima Damásio, A. R. (2016). Mapping N-linked glycosylation
of carbohydrate-active enzymes in the secretome of Aspergillus nidulans grown
on lignocellulose. Biotechnology for Biofuels, 9(1), 168.
https://doi.org/10.1186/s13068-016-0580-4
Ruggiano, A., Foresti, O., & Carvalho, P. (2014). ER-associated degradation: Protein
quality control and beyond. Journal of Cell Biology, 204(6), 869–879.
https://doi.org/10.1083/jcb.201312042
Ruiz-Díez, B. (2002). A Review: Strategies for the transformation of filamentous fungi.
Journal of Applied Microbiology, 92(2), 189–195. https://doi.org/10.1046/j.1365-
2672.2002.01516.x
Sagt, C. M. J., Kleizen, B., Verwaal, R., De Jong, M. D. M., Muller, W. H., Smits, a.,
… Verrips, C. T. (2000). Introduction of an N-glycosylation site increases secretion
of heterologous proteins in yeasts. Applied and Environmental Microbiology,
66(11), 4940–4944. https://doi.org/10.1128/AEM.66.11.4940-4944.2000
Sagt, C. M. J., ten Haaft, P. J., Minneboo, I. M., Hartog, M. P., Damveld, R. a, van der
Laan, J. M., … de Winde, J. H. (2009). Peroxicretion: a novel secretion pathway
in the eukaryotic cell. BMC Biotechnology, 9, 1–11. https://doi.org/10.1186/1472-
6750-9-48
Saloheimo, M., Lund, M., & Penttilä, M. E. (1999). The protein disulphide isomerase
gene of the fungus Trichoderma reesei is induced by endoplasmic reticulum stress
and regulated by the carbon source. Molecular and General Genetics, 262(1), 35–
45. https://doi.org/10.1007/s004380051057

166
Samuelson, J., & Robbins, P. W. (2015). Effects of N-glycan precursor length diversity
on quality control of protein folding and on protein glycosylation. Seminars in Cell
and Developmental Biology, 41, 121–128.
https://doi.org/10.1016/j.semcdb.2014.11.008
Sani, R. K., & Krishnaraj, R. N. (2017). Extremophilic enzymatic processing of
lignocellulosic feedstocks to bioenergy. Extremophilic Enzymatic Processing of
Lignocellulosic Feedstocks to Bioenergy. https://doi.org/10.1007/978-3-319-
54684-1
Saykhedkar, S., Ray, A., Ayoubi-Canaan, P., Hartson, S. D., Prade, R., & Mort, A. J.
(2012). A time course analysis of the extracellular proteome of Aspergillus
nidulans growing on sorghum stover. Biotechnology for Biofuels, 5(1), 52.
https://doi.org/10.1186/1754-6834-5-52
Scheller, H. V., & Ulvskov, P. (2010). Hemicelluloses. Annual Review of Plant Biology,
61, 263–89. https://doi.org/10.1146/annurev-arplant-042809-112315
Schröder, M. (2008). Endoplasmic reticulum stress responses. Cellular and Molecular
Life Sciences (Vol. 65). https://doi.org/10.1007/s00018-007-7383-5
Schwartz, D., & Gygi, S. P. (2005). An iterative statistical approach to the identification
of protein phosphorylation motifs from large-scale data sets. Nature
Biotechnology, 23(11), 1391–1398. https://doi.org/10.1038/nbt1146
Schwarz, F., & Aebi, M. (2011). Mechanisms and principles of N-linked protein
glycosylation. Current Opinion in Structural Biology, 21(5), 576–582.
https://doi.org/10.1016/j.sbi.2011.08.005
Segato, F., Damásio, A. R. L., de Lucas, R. C., Squina, F. M., & Prade, R. A. (2014).
Genomics Review of Holocellulose Deconstruction by Aspergilli. Microbiology and
Molecular Biology Reviews, 78(4), 588–613.
https://doi.org/10.1128/MMBR.00019-14
Segato, F., Damásio, A. R. L. L., Gonçalves, T. A., de Lucas, R. C., Squina, F. M.,
Decker, S. R., & Prade, R. A. (2012). High-yield secretion of multiple client proteins

167
in Aspergillus. Enzyme and Microbial Technology, 51(2), 100–106.
https://doi.org/10.1016/j.enzmictec.2012.04.008
Seo, J.-W., & Lee, K.-J. (2004). Post-translational modifications and their biological
functions: proteomic analysis and systematic approaches. Journal of Biochemistry
and Molecular Biology, 37(1), 35–44.
https://doi.org/10.5483/BMBRep.2004.37.1.035
Sharma, R., Katoch, M., Srivastava, P. S., & Qazi, G. N. (2009). Approaches for
refining heterologous protein production in filamentous fungi. World Journal of
Microbiology and Biotechnology, 25(12), 2083–2094.
https://doi.org/10.1007/s11274-009-0128-x
Shental-Bechor, D., & Levy, Y. (2008). Effect of glycosylation on protein folding: a close
look at thermodynamic stabilization. Proceedings of the National Academy of
Sciences of the United States of America, 105(24), 8256–61.
https://doi.org/10.1073/pnas.0801340105
Shirke, A. N., Su, A., Jones, J. A., Butterfoss, G. L., Koffas, M. A. G., Kim, J. R., &
Gross, R. A. (2017). Comparative thermal inactivation analysis of Aspergillus
oryzae and Thiellavia terrestris cutinase: Role of glycosylation. Biotechnology and
Bioengineering, 114(1), 63–73. https://doi.org/10.1002/bit.26052
Sims, A. H., Gent, M. E., Lanthaler, K., Dunn-Coleman, N. S., Oliver, S. G., & Robson,
G. D. (2005). Transcriptome analysis of recombinant protein secretion by
Aspergillus nidulans and the unfolded-protein response in vivo. Applied and
Environmental Microbiology, 71(5), 2737–2747.
https://doi.org/10.1128/AEM.71.5.2737-2747.2005
Singh Nee Nigam, P., & Pandey, A. (2009). Biotechnology for agro-industrial residues
utilisation: Utilisation of agro-residues. Biotechnology for Agro-Industrial Residues
Utilisation: Utilisation of Agro-Residues. https://doi.org/10.1007/978-1-4020-9942-
7
Skropeta, D. (2009). The effect of individual N-glycans on enzyme activity. Bioorganic
& Medicinal Chemistry, 17(7), 2645–53.

168
https://doi.org/10.1016/j.bmc.2009.02.037
Smith, M. H., Ploegh, H. L., & Weissman, J. S. (2011). Road to ruin: Targeting proteins
for degradation in the endoplasmic reticulum. Science.
https://doi.org/10.1126/science.1209235
Smotrys, J. E., & Linder, M. E. (2004). Palmitoylation of intracellular signaling proteins:
regulation and function. Annual Review of Biochemistry, 73, 559–587.
https://doi.org/10.1146/annurev.biochem.73.011303.073954
Sobolev, V., Sorokine, A., Prilusky, J., Abola, E. E., & Edelman, M. (1999). Automated
analysis of interatomic contacts in proteins. Bioinformatics, 15(4), 327–332.
https://doi.org/10.1093/bioinformatics/15.4.327
Solá, R. J., Rodríguez-Martínez, J. a., & Griebenow, K. (2007). Modulation of protein
biophysical properties by chemical glycosylation: Biochemical insights and
biomedical implications. Cellular and Molecular Life Sciences, 64(16), 2133–2152.
https://doi.org/10.1007/s00018-007-6551-y
Spiro, R. G. (2002). Protein glycosylation: nature, distribution, enzymatic formation,
and disease implications of glycopeptide bonds. Glycobiology, 12(4), 43R–56R.
https://doi.org/10.1093/glycob/12.4.43R
Stals, I., Sandra, K., Geysens, S., Contreras, R., Van Beeumen, J., & Claeyssens, M.
(2004). Factors influencing glycosylation of Trichoderma reesei cellulases. I:
Postsecretorial changes of the O- and N-glycosylation pattern of Ce17A.
Glycobiology, 14(8), 713–724. https://doi.org/10.1093/glycob/cwh080
Stanley, P., Taniguchi, N., & Aebi, M. (2017). N-Glycans. Essentials of Glycobiology.
Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/28876855
Strasser, K., McDonnell, E., Nyaga, C., Wu, M., Wu, S., Almeida, H., … Tsang, A.
(2015). MycoCLAP, the database for characterized lignocellulose-active proteins
of fungal origin: Resource and text mining curation support. Database, 2015.
https://doi.org/10.1093/database/bav008
Szewczyk, E., Nayak, T., Oakley, C. E., Edgerton, H., Xiong, Y., Taheri-Talesh, N., …

169
Oakley, B. R. (2006). Fusion PCR and gene targeting in Aspergillus nidulans.
Nature Protocols, 1(6), 3111–3120. https://doi.org/10.1038/nprot.2006.405
Takayanagi, T., Kimura, A., Chiba, S., & Ajisaka, K. (1994). Novel structures of N-
linked high-mannose type oligosaccharides containing α-D-galactofuranosyl
linkages in Aspergillus niger α-D-glucosidase. Carbohydrate Research, 256(1),
149–158. https://doi.org/10.1016/0008-6215(94)84234-5
Tan, N. Y., Bailey, U.-M., Jamaluddin, M. F., Mahmud, S. H. B., Raman, S. C., &
Schulz, B. L. (2014). Sequence-based protein stabilization in the absence of
glycosylation. Nature Communications, 5, 1–7.
https://doi.org/10.1038/ncomms4099
Tanaka, A., Mita, S., Ohta, S., Kyozuka, J., Shimamoto, K., & Nakamura, K. (1990).
Enhancement of foreign gene expression by a dicot intron in rice but not in tobacco
is correlated with an increased level of mRNA and an efficient splicing of the intron.
Nucleic Acids Research, 18(23), 6767–6770.
https://doi.org/10.1093/nar/18.23.6767
Tanaka, M., Tokuoka, M., & Gomi, K. (2014). Effects of codon optimization on the
mRNA levels of heterologous genes in filamentous fungi. Applied Microbiology
and Biotechnology, 98(9), 3859–3867. https://doi.org/10.1007/s00253-014-5609-
7
Tanaka, M., Tokuoka, M., Shintani, T., & Gomi, K. (2012). Transcripts of a
heterologous gene encoding mite allergen der f 7 are stabilized by codon
optimization in Aspergillus oryzae. Applied Microbiology and Biotechnology, 96(5),
1275–1282. https://doi.org/10.1007/s00253-012-4169-y
Tang, H., Wang, S., Wang, J., Song, M., Xu, M., Zhang, M., … Bao, X. (2016). N-
hypermannose glycosylation disruption enhances recombinant protein production
by regulating secretory pathway and cell wall integrity in Saccharomyces
cerevisiae. Scientific Reports, 6(January), 1–13.
https://doi.org/10.1038/srep25654
Thaysen-Andersen, M., & Packer, N. H. (2012). Site-specific glycoproteomics confirms

170
that protein structure dictates formation of N-glycan type, core fucosylation and
branching. Glycobiology, 22(11), 1440–1452.
https://doi.org/10.1093/glycob/cws110
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). CLUSTAL W: improving the
sensitivity of progressive multiple sequence alignment through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids
Research, 22(22), 4673–4680. https://doi.org/10.1093/nar/22.22.4673
Tilburn, J., Scazzocchio, C., Taylor, G. G., Zabicky-Zissman, J. H., Lockington, R. A.,
& Davies, R. W. (1983). Transformation by integration in Aspergillus nidulans.
Gene, 26(2–3), 205–221. https://doi.org/10.1016/0378-1119(83)90191-9
Tokuoka, M., Tanaka, M., Ono, K., Takagi, S., Shintani, T., & Gomi, K. (2008). Codon
optimization increases steady-state mRNA levels in Aspergillus oryzae
heterologous gene expression. Applied and Environmental Microbiology, 74(21),
6538–46. https://doi.org/10.1128/AEM.01354-08
Trombetta, E. S. (2003). The contribution of N-glycans and their processing in the
endoplasmic reticulum to glycoprotein biosynthesis. Glycobiology, 13(9), 77R–
91R. https://doi.org/10.1093/glycob/cwg075
Trombetta, E. S., & Parodi, A. J. (2003). Quality Control and Protein Folding in the
Secretory Pathway. Annual Review of Cell and Developmental Biology, 19(1),
649–676. https://doi.org/10.1146/annurev.cellbio.19.110701.153949
Vaaje-kolstad, G., Westereng, B., Horn, S. J., Liu, Z., Zhai, H., Sorlie, M., … Eijsink,
V. G. H. (2010). An Oxidative Enzyme Boosting the Enzymatic Conversion of
Recalcitrant Polysaccharides. Science, 330(6001), 219–222.
https://doi.org/10.1126/science.1192231
Valderrama-Rincon, J. D., Fisher, A. C., Merritt, J. H., Fan, Y.-Y., Reading, C. A.,
Chhiba, K., … DeLisa, M. P. (2012). An engineered eukaryotic protein
glycosylation pathway in Escherichia coli. Nature Chemical Biology, 8(5), 434–
436. https://doi.org/10.1038/nchembio.921

171
van Hellemond, E. W., Leferink, N. G. H., Heuts, D. P. H. M., Fraaije, M. W., & van
Berkel, W. J. H. (2006). Occurrence and Biocatalytic Potential of Carbohydrate
Oxidases. Advances in Applied Microbiology, 60(Table I), 17–54.
https://doi.org/10.1016/S0065-2164(06)60002-6
Varga, J., Szigeti, G., Baranyi, N., Kocsubé, S., O’Gorman, C. M., & Dyer, P. S. (2014).
Aspergillus: Sex and Recombination. Mycopathologia.
https://doi.org/10.1007/s11046-014-9795-8
Verdoes, J. C., Punt, P. J., Schrickx, J. M., van Verseveld, H. W., Stouthamer, A. H.,
& van den Hondel, C. A. M. J. J. (1993). Glucoamylase overexpression in
Aspergillus niger: Molecular genetic analysis of strains containing multiple copies
of the glaA gene. Transgenic Research, 2(2), 84–92.
https://doi.org/10.1007/BF01969381
Vries, R. P. De, & Visser, J. (2001). Aspergillus Enzymes Involved in Degradation of
Plant Cell Wall Polysaccharides Aspergillus Enzymes Involved in Degradation of
Plant Cell Wall Polysaccharides. Microbiology and Molecular Biology Reviews,
65(4), 497–522. https://doi.org/10.1128/MMBR.65.4.497
Vuong, T. V, Vesterinen, A.-H., Foumani, M., Juvonen, M., Seppälä, J., Tenkanen, M.,
& Master, E. R. (2013). Xylo- and cello-oligosaccharide oxidation by gluco-
oligosaccharide oxidase from Sarocladium strictum and variants with reduced
substrate inhibition. Biotechnology for Biofuels, 6(1), 148.
https://doi.org/10.1186/1754-6834-6-148
Wacker, M., Linton, D., Hitchen, P. G., Nita-Lazar, M., Haslam, S. M., North, S. J., …
Aebi, M. (2002). N-linked glycosylation in Campylobacter jejuni and its functional
transfer into E. coli. Science, 298(5599), 1790–1793.
https://doi.org/10.1126/science.298.5599.1790
Walter, P., & Ron, D. (2011). The Unfolded Protein Response: From Stress Pathway
to Homeostatic Regulation. Science, 334(6059), 1081–1086.
https://doi.org/10.1126/science.1209038
Wang, L., Aryal, U. K., Dai, Z., Mason, A. C., Monroe, M. E., Tian, Z.-X. X., … Qian,

172
W.-J. J. (2012). Mapping N-linked glycosylation sites in the secretome and whole
cells of aspergillus niger using hydrazide chemistry and mass spectrometry.
Journal of Proteome Research, 11(1), 143–156.
https://doi.org/10.1021/pr200916k
Wang, L., Ridgway, D., Gu, T., & Moo-Young, M. (2005). Bioprocessing strategies to
improve heterologous protein production in filamentous fungal fermentations.
Biotechnology Advances, 23(2), 115–29.
https://doi.org/10.1016/j.biotechadv.2004.11.001
Wang, N., Li, S., Lu, T., Nakanishi, H., & Gao, X. (2017). Approaches towards the core
pentasaccharide in N- linked glycans. Chinese Chemical Letters, 1–5.
https://doi.org/10.1016/j.cclet.2017.09.044
Wang, P., Wang, H., Gai, J., Tian, X., Zhang, X., Lv, Y., & Jian, Y. (2017). Evolution of
protein N-glycosylation process in Golgi apparatus which shapes diversity of
protein N-glycan structures in plants, animals and fungi. Scientific Reports,
7(October 2016), 1–13. https://doi.org/10.1038/srep40301
Ward, O. P. (2012). Production of recombinant proteins by filamentous fungi.
Biotechnology Advances, 30(5), 1119–1139.
https://doi.org/10.1016/j.biotechadv.2011.09.012
Ward, O. P., Qin, W. M., Dhanjoon, J., Ye, J., & Singh, A. (2005). Physiology and
Biotechnology of Aspergillus. In Advances in Applied Microbiology (Vol. 58, pp. 1–
75). https://doi.org/10.1016/S0065-2164(05)58001-8
Weerapana, E., & Imperiali, B. (2006). Asparagine-linked protein glycosylation: from
eukaryotic to prokaryotic systems. Glycobiology, 16(6), 91R–101R.
https://doi.org/10.1093/glycob/cwj099
Wei, W., Chen, L., Zou, G., Wang, Q., Yan, X., Zhang, J., … Zhou, Z. (2013). N -
glycosylation affects the proper folding, enzymatic characteristics and production
of a fungal β-glucosidase. Biotechnology and Bioengineering, 110(12), 3075–
3084. https://doi.org/10.1002/bit.24990

173
White, R. J. (2015). The Search for Functional Porous Carbons from Sustainable
Precursors. Porous Carbon Materials from Sustainable Precursors, (32), 3–49.
https://doi.org/10.1039/9781782622277-00003
Wiseman, R. L., Powers, E. T., Buxbaum, J. N., Kelly, J. W., & Balch, W. E. (2007). An
Adaptable Standard for Protein Export from the Endoplasmic Reticulum. Cell,
131(4), 809–821. https://doi.org/10.1016/j.cell.2007.10.025
Wong, K. H., Todd, R. B., Oakley, B. R., Oakley, C. E., Hynes, M. J., & Davis, M. a.
(2008). Sumoylation in Aspergillus nidulans: sumO inactivation, overexpression
and live-cell imaging. Fungal Genetics and Biology, 45(5), 728–737.
https://doi.org/10.1016/j.fgb.2007.12.009
Wright, T. H., & Davis, B. G. (2017). Post-translational mutagenesis for installation of
natural and unnatural amino acid side chains into recombinant proteins. Nature
Protocols, 12(10), 2243–2250. https://doi.org/10.1038/nprot.2017.087
Yang, M., Yu, X.-W., Zheng, H., Sha, C., Zhao, C., Qian, M., & Xu, Y. (2015). Role of
N-linked glycosylation in the secretion and enzymatic properties of Rhizopus
chinensis lipase expressed in Pichia pastoris. Microbial Cell Factories, 14(1), 40.
https://doi.org/10.1186/s12934-015-0225-5
Yin, Y., Mao, X., Yang, J., Chen, X., Mao, F., & Xu, Y. (2012). dbCAN: A web resource
for automated carbohydrate-active enzyme annotation. Nucleic Acids Research,
40, W445–W451. https://doi.org/10.1093/nar/gks479
Yoneda, A., Kuo, H.-W. D., Ishihara, M., Azadi, P., Yu, S.-M., & Ho, T. D. (2014).
Glycosylation Variants of a β-Glucosidase Secreted by a Taiwanese Fungus,
Chaetomella raphigera, Exhibit Variant-Specific Catalytic and Biochemical
Properties. PLoS ONE, 9(9), e106306.
https://doi.org/10.1371/journal.pone.0106306
Yoon, J., Maruyama, J. I., & Kitamoto, K. (2011). Disruption of ten protease genes in
the filamentous fungus Aspergillus oryzae highly improves production of
heterologous proteins. Applied Microbiology and Biotechnology, 89(3), 747–59.
https://doi.org/10.1007/s00253-010-2937-0

174
Zoglowek, M., Lübeck, P. S., Ahring, B. K., & Lübeck, M. (2015). Heterologous
expression of cellobiohydrolases in filamentous fungi – An update on the current
challenges, achievements and perspectives. Process Biochemistry, 50(2), 211–
220. https://doi.org/10.1016/j.procbio.2014.12.018
Zubieta, M. P., Contesini, F. J., Rubio, M. V., Gonçalves, A. E. de S. S., Gerhardt, J.
A., Prade, R. A., & Damasio, A. R. de L. (2018). Protein profile in Aspergillus
nidulans recombinant strains overproducing heterologous enzymes. Microbial
Biotechnology, 11(2). https://doi.org/10.1111/1751-7915.13027

175
Anexos
Termo de aprovação da pesquisa pela Comissão de Biossegurança

176
Declaração referente aos direitos autorais







