
UNIVERSIDADE FEDERAL FLUMINENSE
INSTITUTO BIOMÉDICO
CURSO DE GRADUAÇÃO EM BIOMEDICINA
EFEITO DO SISTEMA ENDOCANABINOIDE NO
DESENVOLVIMENTO DE CULTURAS DE CÉLULAS DE
RETINA DE GALINHA APÓS EXPOSIÇÃO AGUDA E
CRÔNICA AO WIN 55,212-2
Bacharelado em pesquisa científica: Fisiologia e Farmacologia
AUTOR: ALEXANDER GUNTER
ORIENTADORA: REGINA CÉLIA CUSSA KUBRUSLY
Niterói, RJ
2018


ii
ALEXANDER GUNTER
EFEITO DO SISTEMA ENDOCANABINOIDE NO DESENVOLVIMENTO DE
CULTURAS DE CÉLULAS DE RETINA DE GALINHA APÓS EXPOSIÇÃO AGUDA
E CRÔNICA AO WIN 55,212-2
Orientadora:
Prof.ª Dr.ª Regina Célia Cussa Kubrusly
Niterói, RJ
2018
Trabalho de conclusão de curso submetido à
Universidade Federal Fluminense como
requisito parcial para obtenção do Grau de
Bacharel em Biomedicina. Área de
concentração: Fisiologia e Farmacologia

iii
ALEXANDER GUNTER
EFEITO DO SISTEMA ENDOCANABINOIDE NO DESENVOLVIMENTO DE
CULTURAS DE CÉLULAS DE RETINA DE GALINHA APÓS EXPOSIÇÃO AGUDA
E CRÔNICA AO WIN 55,212-2
BANCA EXAMINADORA
________________________________________________________________
Prof.ª Dr.ª Regina Célia Cussa Kubrusly
Universidade Federal Fluminense
________________________________________________________________
Prof.ª Dr.ª Ana Lúcia Marques Ventura
Universidade Federal Fluminense
________________________________________________________________
Dr.ª Luzia da Silva Sampaio
Universidade Federal do Rio de Janeiro
________________________________________________________________
Msc. Robertta Silva Martins (suplente)
Universidade Federal Fluminense
Niterói, RJ
2018
Trabalho de conclusão de curso submetido à
Universidade Federal Fluminense como
requisito parcial para obtenção do Grau de
Bacharel em Biomedicina. Área de
concentração: Fisiologia e Farmacologia

iv
Trabalho desenvolvido no laboratório de Neurofarmacologia do Departamento de
Fisiologia e Farmacologia - Instituto Biomédico, na Universidade Federal
Fluminense sob a orientação da Professora Drª Regina Célia Cussa Kubrusly.

v
A todos com interesse genuíno pela ciência
A todos que sempre estiveram ao meu lado e me apoiaram

vi
AGRADECIMENTOS
Para que esse trabalho fosse realizado, definitivamente eu tive a ajuda e apoio de
diversas pessoas sensacionais, e sem elas eu definitivamente não estaria aqui escrevendo a
minha monografia.
Gostaria de primeiramente agradecer a minha família. Aos meus pais, Claus e Sonia,
vocês sempre me apoiaram incondicionalmente, me criaram e me deram toda condição, suporte
e amor para que eu pudesse chegar até aqui. Vocês foram guerreiros a vida toda e são o exemplo
de vida que eu quero seguir. A minha irmã, Katharina, quando estamos juntos não existe tempo
ruim, a amizade que temos é algo que valorizo muito e quero que continue para sempre. Ao
meu avô Luiz e minha avó Anna, obrigado por compartilhar toda experiência de vida que vocês
têm. A Tia Eliana, Tio Márcio, Nini e Tia Li, obrigado a todos conselhos, papos cabeça e
momentos de descontração.
A todos colegas de laboratório que fizeram parte da minha rotina. Robertta, Arlete,
Augusto, Vlad, Richard, Matheus, Thaís, Marcela, Carlos e Arthur, cada um de vocês teve um
papel importante na minha trajetória no laboratório. Um agradecimento especial a quem tornou
esse projeto possível, a minha mãe científica, Regina. Você me acolheu de braços abertos desde
o primeiro dia e desde então me ensinou o que é necessário para me tornar um bom cientista.
Irei levar seus ensinamentos comigo sempre, independentemente de onde eu estiver. Agradeço
também ao Professor Ricardo Reis por sempre ter me recebido muito bem em seu laboratório.
Carol, o agradecimento agora vai para você. Nós entramos juntos na UFF e agora
estamos saindo juntos. Eu não consigo descrever em palavras o quão importante você foi para
mim nesse período de nossas vidas. Todas vezes que eu duvidei de mim mesmo, que eu estive
emocionalmente abalado, em todas elas você esteve do meu lado, me dando motivação e apoio
emocional para seguir adiante nessa rotina universitária.
A todos professores que eu tive, eu valorizo o aprendizado que tive com cada um de
vocês, isso me proporcionou poder enxergar o mundo de uma maneira completamente diferente
desde que entrei na UFF. A todos amigos que eu fiz e a equipe da coordenação. Ronald, Renata
e Nathália, vocês sempre estiveram disponíveis para tirar minhas dúvidas e sempre me ajudaram
quando eu precisei de vocês.

vii
RESUMO
Cannabis é atualmente uma das drogas mais consumidas mundialmente e dentre a
parcela de usuários se encontra um número considerável de mulheres na gravidez. O sistema
endocanabinoide possui papel fundamental no desenvolvimento do SNC e é capaz de modular
a liberação de diversos neurotransmissores, como o GABA e o glutamato. O estímulo crônico
de canabinoides também possui efeitos modulatórios na expressão de receptores CB1 e CB2,
podendo levar à down-regulation deles. O objetivo do projeto foi estudar o efeito do sistema
endocanabinoide no desenvolvimento embrionário de culturas primárias mistas de retina de
galinha Leghorn de idades E7C7 e E8C8. Para isso, foi utilizado o agonista canabinoide
sintético misto WIN 55,212-2. Primeiramente foi demonstrado por Western Blot que tanto
receptores CB1, quanto receptores CB2 estão expressos e funcionais nas culturas. Além disso,
o estímulo agudo de WIN 55,212-2 é responsável por reduzir os níveis de liberação de [³H]-
GABA a partir de receptores CB1, sem alterar os valores de captação. O pré-tratamento crônico
de 3 horas com WIN 55,212-2 leva à down-regulation dos receptores CB1, sem alterações na
expressão de receptores CB2. A retirada do tratamento crônico do WIN 55,212-2 por 24 horas
gera um aumento dos níveis de AMPc, o que indica uma down-regulation tardia de receptores
CB2. Em suma, estes resultados demonstram que os estímulos agudos e crônicos de WIN
55,212-2 em culturas mistas de retina de galinha modulam a excitabilidade celular e a expressão
de receptores canabinoides.
Palavras-chave: GABA, canabinoide, retina, desenvolvimento, AMPc

viii
ABSTRACT
Cannabis is currently one of the most consumed drugs worldwide, and within the share
of users lies a considerable number of pregnant women. The endocannabinoid system has a
major role on the development of the central nervous system and is capable of modulating the
release of many neurotransmitters, such as GABA and glutamate. The chronic stimulus of
cannabinoids also has modulatory effects on the expression of CB1 and CB2 receptors, which
may lead to their down-regulation. The objective of this project was to study the effect of the
endocannabinoid system on the embryonic development of primary mixed Leghorn chick retina
cell cultures with ages of E7C7 and E8C8. For this, the synthetic and mixed cannabinoid agonist
WIN 55,212-2 was used. First, it was shown through Western Blot analysis that CB1 and CB2
are expressed and functional on the cell cultures. Besides that, the acute stimulus of WIN
55,212-2 is responsible for reducing the levels of [³H]-GABA release through CB1 receptors,
without reducing the neurotransmitter uptake levels. The chronic pretreatment of 3 hours with
WIN 55,212-2 leads to the down-regulation of CB1 receptors, without alterations on the
expression of CB2 receptors. The withdrawal of the WIN 55,212-2 chronic treatment for 24
hours causes an increase on the cAMP levels, which indicates a late down-regulation of CB2
receptors. All in all, these results showed that the acute and chronic stimuli of WIN 55,212-2
on mixed chick retina cell cultures modulate the cellular excitability and the expression of
cannabinoid receptors.
Keywords: GABA, cannabinoid, retina, development, cAMP

ix
SUMÁRIO
INTRODUÇÃO ..................................................................................................................................... 1
1. O sistema endocanabinoide ............................................................................................. 1
a. Receptores canabinoides .............................................................................................. 1
b. Canabioides .................................................................................................................. 3
c. O uso de Cannabis durante a gravidez ........................................................................ 5
d. Uso crônico de canabinoides ....................................................................................... 6
2. Retina ............................................................................................................................... 7
a. Desenvolvimento da retina .......................................................................................... 8
b. O sistema endocanabinoide na retina ........................................................................... 9
c. A retina como modelo experimental .......................................................................... 10
3. O sistema GABAérgico ................................................................................................. 10
a. Transportadores ......................................................................................................... 12
b. Liberação de GABA .................................................................................................. 13
OBJETIVOS ........................................................................................................................................ 14
1. Objetivos gerais ............................................................................................................. 14
2. Objetivos específicos ..................................................................................................... 14
MATERIAIS E MÉTODOS .............................................................................................................. 15
1. Animais e cultura de células .......................................................................................... 15
a. Tratamentos ............................................................................................................... 15
2. Western Blot .................................................................................................................. 16
3. Ensaio de AMPc ............................................................................................................ 17
4. Liberação de [³H]-GABA .............................................................................................. 17
5. Captação de [³H]-GABA ............................................................................................... 18
6. Análise estatística .......................................................................................................... 18

x
RESULTADOS ................................................................................................................................... 19
1. Os receptores CB1 e CB2 estão expressos e funcionais em culturas mistas de retina de
galinha embrionárias ...................................................................................................... 19
2. O WIN 55,212-2 reduz a liberação de [³H]-GABA agindo em receptores CB1, sem
alterar sua captação. ....................................................................................................... 20
3. A exposição crônica das culturas ao WIN 55,212-2 reduz a expressão do receptor CB1,
mas não do CB2. ............................................................................................................ 22
DISCUSSÃO ....................................................................................................................................... 24
CONCLUSÃO ..................................................................................................................................... 32
PERSPECTIVAS ................................................................................................................................ 32
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................. 33

xi
LISTA DE ILUSTRAÇÕES
Figura 1: Efeitos da ativação dos receptores CB1 e CB2.. ......................................................... 2
Figura 2: A sinalização do sistema eCB.. ................................................................................. 4
Figura 3: A síntese dos eBCs AEA e 2-AG. ............................................................................. 5
Figura 4: A retina e sua divisão em camadas. ........................................................................... 8
Figura 5: O sistema GABAérgico. .......................................................................................... 11
Figura 6: Desenvolvimento do sistema GABAérgico e endocanabinoide em retina de galinha.
.................................................................................................................................................. 12
Figura 7: Modelo de funcionamento do transportador GAT-1. .............................................. 13
Figura 8: Linha temporal com idades e tratamento crônico com WIN 55,212-2 a 100 nM. .. 16
Figura 9: Expressão e funcionalidade de receptores canabinoides. ........................................ 19
Figura 10: Efeito do WIN 55,212-2 agudo na liberação e captação de [³H]-GABA em
culturas mistas de retina de galinha E7C7. ............................................................................... 21
Figura 11: Efeito do WIN 55,212-2 crônico na expressão e funcionalidade de receptores
canabinoides em culturas mistas de retina de galinha. ............................................................. 23

xii
LISTA DE ABREVIATURAS
Δ9-THC (−) -Δ9-tetrahidrocanabinol
[³H]- Trítio
2-AG 2-araquidonilglicerol
AEA Anandamida
AMPc Monofosfato cíclico de adenosina
CAP Capsaicina
CB1 Receptor canabinoide tipo 1
CB2 Receptor canabinoide tipo 2
CCG Camada de células ganglionares
CBD Canabidiol
CNE Camada nuclear externa
CNI Camanda nuclear interna
CPE Camada plexiforme externa
CPI Camada plexiforme interna
CPM Contagens por minuto
Cx Idade da cultura de x dias
DAG Diacilglicerol
DGL Diacilglicerol-lipase
DMEM/F-12 Meio de Eagle modificado Dulbecco: Mistura de nutrientes F-12
eCB Endocanabinoide
Ex Idade embrionária de x dias
FAAH Amida hidrolase de ácidos graxos
FGF Fator de crescimento fibroblástico
GABA Ácido gama-aminobutírico

xiii
GAD Glutamato descarboxilase
GAT Transportador de alta afinidade de GABA
GPR55 Receptor acoplado à proteína G 55
Liso-Pi Lisofosfolipídeo
Liso-PLC Lisofosfolipase C
MAPK Proteína quinase ativada por mitógeno
MGL Monoacilglicerol-lipase
NAPE N-araquinodoil-fosfatidiletanolamina
NAPE-PLD N-araquidonil-fosfatidil-etanolamina-fosfolipase-D
NAT N-aciltransferase
NMDA N-metil D-Aspartato
PKA Proteína cinase A
PLA1 Fosfolipase A1
PLC Fosfolipase C
Ro20 Rolipram
SDS-PAGE Dodecil sulfato de sódio-gel de poliacrilamida para eletroforese
SEM Erro padrão da média
SFB Soro fetal Bovino
SNC Sistema nervoso central
TCA Ácido tricloroacético
TRPV1 Receptor vanilóide de potencial transitório 1
TTBS Tampão salino de tris(hidroximetil)aminometano + Tween
VGAT Transportador vesicular de GABA
WIN 55,212-2 (R)-(+)-[2,3-Dihidro-5-metil-3(morfolinil) metil-pirrolo[1,2,3]-1,4-
benzoxazinil-(1-naftalenil) metanona mesilato

1
INTRODUÇÃO
1. O sistema endocanabinoide
As plantas do gênero Cannabis spp. produzem mais de 70 espécies moleculares de
fitocanabinoides, sendo o principal deles o (−) -Δ9-tetrahidrocanabinol (Δ9-THC) (DI MARZO
et al., 2015). Este componente é responsável pelos efeitos psicoativos e de euforia da planta e
foram descobertos no ano de 1964 a partir de seu isolamento em haxixe confiscado (GAONI;
MECHOULAM, 1964). Outro fitocanabinoide importante é o canabidiol (CBD), cuja
importância na pesquisa tem crescido muito, pois é um potente agente anti-inflamatório
(MECHOULAM; PARKER, 2013) que não possui efeitos psicoativos, além de ter potentes
efeitos anticonvulsivantes e poder agir como ansiolítico, antipsicótico e sedativo (DOS
SANTOS et al., 2015).
Historicamente, as plantas do gênero Cannabis spp. são cultivadas há mais de 5000
anos, e isso teve começo no centro e no nordeste da Ásia para posteriormente se espalhar pelo
mundo todo com o intuito de serem utilizadas para fins medicinais, recreacionais e têxteis
(ELSOHLY; SLADE, 2005). Seguindo essa descoberta, os pesquisadores da época se
perguntavam porque o nosso cérebro reage à Cannabis. A resposta veio em 1988 com a
descoberta de receptores canabinoides a partir do uso do [³H]-CP55,940, um potente agonista
canabinoide sintético (DEVANE, et al., 1988).
a. Receptores canabinoides
Os receptores CB1 e CB2 são proteínas que atravessam sete vezes a membrana
plasmática e pertencem à classe dos receptores metabotrópicos. Eles estão principalmente
acoplados a proteínas Gi e os receptores CB1, com menos frequência, a proteínas Gq/11, Gs ou Go
(COSTA et al., 2011; HOWLETT et al., 2002; BOW; RIMOLDI, 2016). Quando ativados,
esses receptores classicamente inibem a adenilil-ciclase, o que reduz a produção de AMPc pela
célula, fazendo com que ocorra um fechamento de canais de cálcio, abertura de canais de
vazamento de potássio e ativação de cinases como a MAPK (Figura 1) (MACKIE, 2007). Como
consequência, ocorre uma redução na liberação de neurotransmissores (HOWLETT et al.,
2002).

2
Figura 1: Efeitos da ativação dos receptores CB1 e CB2. Agonistas canabinoides estimulam a inibição da adenilil-
ciclase , fechamento de canais de Ca2+ e abertura de canais de K+ (HONÓRIO; ARROIO; DA SILVA, 2006).
Outro receptor bastante discutido atualmente é o GPR55, que apesar de ter uma baixa
homologia com a sequência de aminoácidos de receptores CB1 e CB2, é ativado por
canabinoides como o Δ9-THC e a anandamida (AEA) (LAUCKNER et al., 2008). Além disso,
ele está amplamente expresso no cérebro e já foi encontrado na retina de macacos (BOUSKILA
et al., 2013). Diferente dos receptores clássicos, o GRP55 estimula a liberação de cálcio
intracelular via fosfolipase C (PLC) (MARICHAL-CANCINO et al., 2017).
Os receptores CB1 estão principalmente localizados no cérebro em terminais pré-
sinápticos (KATONA et al., 1999), embora possam ser encontrados na região pós-sináptica
(SÁNCHEZ-BLÁZQUEZ; RODRÍGUEZ-MUÑOZ; GARZÓN, 2014). Um estudo
demonstrou que em galinhas de 15 dias de idade, a retirada da retina estimulou a up-regulation
de receptores CB1 no colículo superior contralateral, o que indica uma localização pós-sináptica
desses receptores em axônios retinotectais (CHAVES et al., 2008). Os receptores canabinoides
modulam a liberação de diversos neurotransmissores, como o GABA e o glutamato, e devido a
isso possuem efeitos neuromodulatórios (HÄRING et al., 2015). No cérebro, esses receptores
foram encontrados em gânglios basais, substância nigra, globo pálido, cerebelo e hipocampo
(MECHOULAM; PARKER, 2013; ANDÓ et al., 2012). Muitas dessas regiões estão envolvidas
com funções importantes como: memória, cognição, humor e coordenação motora
(SVÍŽENSKÁ; DUBOVÝ; ŠULCOVÁ, 2008).
Os receptores CB2 estão majoritariamente localizados em células e tecidos do sistema
imune, porém também são encontrados em menores quantidades no sistema nervoso central
(SNC) associados com a resposta inflamatória em células da micróglia e em macrófagos

3
residentes (MACKIE, 2008). Evidências recentes também apontam para um papel dos
receptores CB2 na função sináptica e neuroplasticidade (KENDALL; YUDOWSKI, 2017).
b. Canabinoides
A descoberta dos receptores canabinoides despertou o interesse dos pesquisadores na
possível existência de componentes endógenos que se ligariam a eles. Sendo assim, no ano de
1992 foi encontrado o primeiro endocanabinoide (eCB), a AEA (DEVANE. et al., 1992). Este
eCB é um agonista parcial dos receptores canabinoides e possui alta afinidade por receptores
vanilóides de potencial transitório 1 (TRPV1), que são canais catiônicos não-específicos com
seletividade para Ca2+, e são majoritariamente expressos em neurônios sensoriais (TÓTH;
BLUMBERG; BOCZÁN, 2009; TOMINAGA et al., 1998). A AEA se encontra em baixas
concentrações no cérebro e em outros tecidos (SUGIURA et al., 2002), o que não é comum
para um ligante endógeno. Sendo assim, 3 anos depois foi descoberto o 2-araquidonilglicerol
(2-AG) (MECHOULAM et al., 1995). Este eCB é um agonista total de receptores CB1 e CB2 e
está presente em concentrações muito maiores no cérebro em comparação à AEA (KATONA,
ISTVÁN; FREUND, 2014). Outro componente endógeno que se liga a receptores canabinoides
é a hemopressina, que é um peptídeo derivado da hemoglobina. A hemopressina é um agonista
inverso de receptores CB1, apresentando efeitos antinociceptivos (HEIMANNN et al., 2007).
Os eCBs são mediadores lipídicos que são produzidos no terminal pós-sináptico do
neurônio a partir de precursores fosfolipídicos, sob demanda e a partir do influxo de cálcio (DE
GODOY-MATOS et al., 2006). Eles medeiam a transferência de informações para o terminal
pré-sináptico, realizando uma sinalização retrógrada (KREITZER; REGEHR, 2002). Outra
característica dos eCBs, é que eles não são armazenados em vesículas sinápticas (Figura 2).
Todos esses fatores são responsáveis por torná-los mensageiros atípicos, e não
neurotransmissores clássicos, já que para isso, os transmissores precisariam ser sintetizados no
neurônio pré-sináptico a partir de precursores específicos e armazenados em vesículas
sinápticas (SAITO; WOTJAK; MOREIRA, 2010; BURNSTOCK, 2009).

4
Figura 2: A sinalização do sistema eCB é retrógrada. Os eCBS AEA e 2-AG são produzidos sob demanda no
terminal pós-sináptico a partir do influxo de cálcio e agem nos receptores canabinoides pré-sinápticos. A enzima
FAAH degrada a AEA no terminal pós-sináptico e a MGL degrada o 2-AG no terminal pré-sináptico (SAITO;
WOTJAK; MOREIRA, 2010).
Dentre as diversas vias de síntese dos endocanabinoides, a produção da AEA ocorre
principalmente em dois passos: Primeiramente, a enzima N-aciltransferase (NAT) é
responsável por catalisar a síntese da N-araquinodoil-fosfatidiletanolamina (NAPE) a partir da
fosfatidilcolina e fosfatidiletanolamina. Essa é a etapa limitante da síntese da AEA, e é
dependente de cálcio e AMPc (PIOMELLI, 2003; CADAS; DI TOMASO; PIOMELLI, 1997).
Depois, a NAPE é hidrolisada pela N-araquidonil-fosfatidil-etanolamina-fosfolipase-D
(NAPE-PLD), que está localizada em membranas intracelulares pré- e pós-sinapticamente,
formando a AEA (Figura 3A). Sua degradação ocorre a partir da enzima amida hidrolase de
ácidos graxos (FAAH), localizada em neurônios pós-sinápticos (GULYAS et al., 2004). Sua
degradação também pode ocorrer por meio de carreadores transmembranares pré-sinápticos
(BELTRAMO et al., 1997).
O 2-AG é sintetizado a partir da enzima PLC, a qual catalisa a reação que gera 1,2-
Diacilglicerol (DAG). Esse produto é um substrato para uma enzima denominada diacilglicerol-
lipase (DGL), que está localizada na região pós-sináptica e irá converter a DAG em 2-AG
(STELLA; SCHWEITZER; PIOMELLI, 1997). Uma via alternativa da síntese do 2-AG é por
meio da fosfolipase A1 (PLA1), que catalisa a formação de um lisofosfolipídeo (liso-PI) a partir
do fosfatidilinositol. O liso-PI será hidrolizado a 2-AG pela enzima lisofosfolipase C (Liso-
PLC) (Figura 3B) (PIOMELLI, 2003). A enzima responsável pela degradação do 2-AG é a
monoacilglicerol-lipase (MGL), que possui localização pré-sináptica (SAITO; WOTJAK;
MOREIRA, 2010; GULYAS et al., 2004).

5
Figura 3: A síntese dos eBCs AEA e 2-AG. A. enzima NAT produz NAPE a partir da fosfatidiletanolamina. A
NAPE é hidrolizada pela NAPE-PLD, formando AEA. B. O fosfatidilinositol é catalisado a DAG pela PLC. A
DGL converte DAG a 2-AG. Uma via alternativa é a formação de 2-AG pela PLA1, que forma um liso-PI. Este é
catalisado a 2-AG pela Liso-PLC (adaptado de PIOMELLI, 2003).
Além dos fitocanabinoides e dos eCBs, existem os canabinoides sintéticos, que
começaram a ser produzidos em laboratórios após a descoberta da estrutura do Δ9-THC. Eles
possuem como característica principal uma maior potência em relação aos fitocanabinoides e
cada um possui afinidade e seletividade diferenciadas (SVÍŽENSKÁ; DUBOVÝ; ŠULCOVÁ,
2008). Dentre eles estão os agonistas HU-210, o CP 55,940 e o WIN 55,212-2. O último é um
agonista misto, que possui uma alta afinidade tanto por receptores CB1, quanto por receptores
CB2 e seletividade um pouco maior pelos receptores CB2 em humanos (ASHTON et al., 2008).
Além dos agonistas, também existem antagonistas sintéticos. Um exemplo é o AM-251, o qual
é um antagonista de receptores CB1 estruturalmente parecido com o rimonabanto, que também
já foi descrito como agonista inverso de receptores CB1 e agonista GPR55 (RYBERG et al.,
2007).
c. O uso de Cannabis durante a gravidez
Cannabis é atualmente uma das drogas mais consumidas mundialmente. De acordo com
dados de 2015, 3,8% da população mundial fez o consumo de Cannabis durante o ano, o que
representa aproximadamente 183 milhões de pessoas (UNITED NATIONS OFFICE OF
DRUGS AND CRIME, 2017). Entre os consumidores dessa droga psicoativa se encontram
mulheres grávidas. Um estudo demonstrou que de 4971 mulheres grávidas entre 18-44 anos,
A B

6
3,9% delas fizeram uso da marijuana no mês anterior ao estudo, 7% fizeram uso dessa droga
entre os últimos 2 e 12 meses, e dessas, 18% se encaixaram nos critérios de abuso e dependência
à droga, de acordo com os critérios do DSM-IV (KO et al., 2015). Mundialmente, se estima
que de 3% a 10% das gestantes consumam Cannabis (RIBEIRO et al., 2016).
O uso pré-natal de Cannabis pode ser prejudicial ao feto. Foi demonstrado que cerca de
1/3 do Δ9-THC presente no plasma da mulher grávida é capaz de atravessar a barreira
placentária, além de também poder ser secretado no leite materno (HUTCHINGS et al., 1989;
PEREZ-REYES; WALL, 1982). O sistema endocanabinoide possui funções importantes no
desenvolvimento do SNC, e devido a isso é controlado de forma precisa em estágios críticos do
desenvolvimento. A exposição pré-natal ao Cannabis pode interromper a sinalização
canabinoide nesse período, e como consequência, problemas cognitivos poderão ser observados
no futuro da criança (WU; JEW; LU, 2011).
d. Uso crônico de canabinoides
Sabendo-se o risco que o uso crônico da Cannabis pode gerar, é de suma importância
conhecer os mecanismos moleculares envolvidos na utilização abusiva da Cannabis e
consequente dependência a droga. De acordo com o DSM-V, o transtorno por uso de Cannabis
e a abstinência de Cannabis são considerados transtornos mentais. O critério diagnóstico para
o transtorno por uso de Cannabis é classificado como um padrão problemático do uso da droga
durante um período de 12 meses, por onde ocorre a manifestação de dois entre diversos
critérios. Entre estes critérios estão a tolerância e a abstinência ao Cannabis, além de outros,
como uso continuado de Cannabis, apesar de problemas sociais ou interpessoais persistentes
(AMERICAN PSYCHIATRY ASSOCIATION, 2013).
Tanto em roedores, quanto em humanos, foi observado que a exposição crônica a
marijuana e ao haxixe estimula a down-regulation de receptores CB1 no cérebro, que foi
observado no estriado, cerebelo e sistema límbico e cuja magnitude depende da região
encefálica (SIM-SELLEY, 2003). Essa internalização é reversível, pois após 4 semanas de
abstinência os níveis de receptor CB1 voltam ao normal (HIRVONEN et al., 2012). O termo
down-regulation implica diminuição de receptores por meio da redução da síntese,
internalização com degradação em lisossomos ou degradação não lisossômica de receptores
(VENTURA et al., 2010).

7
Além da down-regulation de receptores canabinoides causada pelo uso crônico de
canabinoides, já foi demonstrado em ratos que após 7 dias de abstinência do estímulo crônico
do WIN 55,212-2, os níveis de binding voltam aos níveis basais, o que indica uma possível up-
regulation de receptores CB1, ou seja, a ocorrência de um aumento do número de receptores na
superfície celular. Além disso, os níveis de mRNA dos receptores CB1 também voltam aos
níveis basais após 20 dias de abstinência (PERDIKARIS et al., 2018). A up-regulation de
receptores CB1 também foi observada em modelos animais de dano cerebral. Em ratos que
sofreram lesões nos neurônios dopaminérgicos nigroestriatais, ocorreu um aumento dos níveis
de mRNA de receptores CB1 em neurônios estriatais aferentes (ROMERO et al., 2000).
2. Retina
A retina é um tecido de aproximadamente 0,2 mm de espessura, que está localizada na
parte posterior do olho e é responsável pela conversão do estímulo luminoso em estímulo
elétrico. Isso ocorre porque os fotorreceptores da retina possuem proteínas denominadas
opsinas, que dependem do retinal para mudar sua conformação em resposta a fótons (KOLB,
2003). O retinal é fornecido aos fotorreceptores pelo epitélio pigmentado (SUNG; CHUANG,
2010). Posteriormente, é ativada uma via de sinalização responsável por alterações
eletroquímicas nos fotorreceptores. Esse estímulo químico é conduzido até as células
ganglionares, cujos axônios formam o nervo óptico e transmitem sinais ao cérebro, onde ocorre
o processamento e a percepção visual (KOLB, 2003).
A sinalização da retina é dividida em via vertical e via lateral (CALAZA; GARDINO,
2010). Nas células que formam a via vertical de sinalização da retina, que são os
fotorreceptores, células bipolares e ganglionares, o principal neurotransmissor liberado é o
glutamato. Na camada plexiforme externa (CPE), as células horizontais transmitem sinais
inibitórios lateralmente. O mesmo é realizado por células amácrinas na camada plexiforme
interna (CPI). Essas células fazem parte da via lateral de sinalização da retina, e nela, o GABA
é um dos principais neurotransmissores utilizados (Figura 4) (KOLB, 2009).

8
Figura 4: A retina e sua divisão em camadas. A camada nuclear externa (CNE) é constituída pelos núcleos dos
fotorreceptores. A camada plexiforme externa (CPE) abriga prolongamentos e terminais sinápticos de
fotorreceptores, células bipolares e horizontais. Na camada nuclear interna (CNI) se encontram os núcleos das
células bipolares, amácrinas, horizontais e da glia de Müller. A camada plexiforme interna (CPI) abriga processos
e terminais de células bipolares, amácrinas e ganglionares. O núcleo das células ganglionares fica na camada de
células ganglionares (CCG) (© ELSEVIER, INC. – NETTERIMAGES.COM).
a. Desenvolvimento da retina
A retina é um componente do desenvolvimento do SNC, pois ela é originária da região
posterior do prosencéfalo, o diencéfalo (HEAVNER; PEVNY, 2012). A histogênese da retina
começa quando as vesículas ópticas primárias entram em contato com o ectoderma e invaginam
para formar a vesícula óptica secundária, que é uma estrutura bilaminada. Sua lâmina interna
dará origem à retina neural, e para tal é necessário que ocorra a liberação de fatores de
crescimento FGF do ectoderma neural (MEY; THANOS, 2000). As células da retina são
originárias de células progenitoras multipotentes, as quais dão origem a 7 tipos celulares
principais em vertebrados, sendo 6 deles de origem neuronal e 1 deles glial (DOH et al., 2010).
O desenvolvimento desses tipos celulares ocorre de acordo com a seguinte ordem: Células
ganglionares, células horizontais, cones, células amácrinas, células bipolares, bastonetes e glia
de Müller (DOH et al., 2010). Na retina de galinha, a neurogênese começa na idade de E2 em
uma região posterior próxima ao disco óptico em um gradiente centro-periférico e dura

9
aproximadamente 12 dias. Também existe um gradiente dorsoventral e outro temporal para
nasal na neurogênese das células retinianas (PRADA et al., 1991).
b. O sistema endocanabinoide na retina
O sistema eCB está diretamente relacionado com o desenvolvimento retiniano e
participa de processos importantes, como proliferação celular, migração e neurogênese. Como
exemplo, um estudo comprovou que agonistas específicos para receptores CB1 e CB2 induziram
o colapso dos cones de crescimento das células ganglionares na retina e reduziram o
crescimento axonal (BOUCHARD et al., 2016). Além disso, também foi comprovado que
receptores CB1 no embrião de galinha possuem uma expressão que segue a diferenciação
neuronal, tanto espacialmente, quanto temporalmente. Isso indica que o desenvolvimento
retiniano está intimamente relacionado com esses receptores (BEGBIE; DOHERTY;
GRAHAM, 2004).
Além de ser um importante modulador do desenvolvimento retiniano, o sistema
endocanabinoide se encontra expresso no sistema retinotectal da galinha, onde receptores CB1
foram identificados na idade embrionária de E4, com marcações nas células ganglionares. Em
E16, o padrão de distribuição de receptores CB1 se encontra estabelecido, com marcações nas
CPI, CNI, CCG e em células amácrinas (LEONELLI et al., 2005). Além disso, os receptores
CB1 também estão expressos nos terminais sinápticos de fotorreceptores, células amácrinas e
ganglionares de diversos animais. Entre eles estão ratos, camundongos, galinhas, salamandras
e macacos (STRAIKER et al., 1999).
Assim como os receptores CB1, os receptores CB2, os eCBs e suas enzimas se encontram
amplamente expressos na retina de galinhas, macacos, camundongos, ratos e peixes dourados.
(SCHWITZER et al., 2016). Na retina de galinha, os receptores CB1 e CB2 foram encontrados
na idade de E5 a PE7, com uma tendência de redução na expressão durante o desenvolvimento.
A Redução dos níveis do receptor CB2 foi observada a partir de E9 (SAMPAIO et al., 2018).
Em relação à localização, os receptores CB1 foram encontrados majoritariamente na CCG e
CPI, enquanto que os receptores CB2 estão predominantemente expressos na CPI. A enzima
MGL está expressa em todas as camadas da retina, principalmente nas camadas plexiformes de
E9 a PE7, sendo encontrada principalmente em células gliais (SAMPAIO et al., 2018). Tanto
os receptores CB1, quanto os receptores CB2 estão expressos em neurônios e na glia de culturas
mistas de retina de galinha (KUBRUSLY et al., 2018).

10
c. A retina como modelo experimental
A retina de galinha tem sido utilizada como modelo para estudar o papel de neuro- e
gliotransmissores por mais de 40 anos (REIS et al., 2007). Ela se conecta ao cérebro a partir do
nervo óptico e possui diversas semelhanças com esse tecido. Uma delas é a presença de diversos
sistemas de neuromoduladores e neurotransmissores, como serotonina, GABA, dopamina e
glutamato. Além disso, as células da retina são sensíveis a drogas farmacológicas que agem na
neurotransmissão do SNC. Um exemplo são os efeitos do levodopa no eletrorretinograma de
pacientes com a doença de Alzheimer (PEPPE et al., 1998). Por último, desordens psiquiátricas,
como esquizofrenia, transtorno do espectro autista e adição a drogas proporcionam
manifestações na função da retina (LAVOIE; MAZIADE; HÉBERT, 2014). Todos esses
fatores contribuem para utilizar de maneira confiável a retina como modelo de estudo do SNC.
Ademais, a retina é mais simples de ser dissecada devido à sua localização anatômica de fácil
acesso, e suas células são divididas em camadas bem definidas, com uma organização
histológica altamente ordenada, o que torna seu estudo mais vantajoso.
3. O sistema GABAérgico
O ácido gama-aminobutírico (GABA), é o principal neurotransmissor inibitório do SNC
dos vertebrados e é de extrema importância na regulação da excitabilidade de circuitos neurais.
Na retina de galinha, neurônios que produzem GABA começam a surgir na idade de E6 e
apresentam alta taxa de geração até a idade de E9 (CALAZA; HOKOÇ; GARDINO, 2000).
Nesse período, esse neurotransmissor é principalmente sintetizado pela putrescina (DE
MELLO; BACHRACH; NIRENBERG, 1976; HOKOÇ et al., 1990). Posteriormente, a síntese
do GABA ocorre a partir da α-descarboxilação do L-glutamato, pela enzima glutamato
descarboxilase (GAD). Na literatura são descritas duas formas de GAD: GAD 65 e GAD 67,
que apresentam diferenças na cinética e distribuição nas células (KAUFMAN; HOUSER;
TOBIN, 1991). Essa enzima se encontra expressa na retina de galinha a partir da idade de E11,
e a partir desse estágio, corresponde à principal via de formação do GABA (HOKOÇ et al.,
1990). O sistema GABAérgico também é composto por receptores que se distribuem tanto pré-
quanto pós-sinapticamente. O GABAA e o GABAC são ionotrópicos e quando ativados, levam
ao influxo de Cl-. O receptor GABAB está acoplado à proteína G, pela qual ele é capaz de
interagir com outras proteínas celulares (Figura 5) (GOVINDPANI et al., 2017).

11
Figura 5: O sistema GABAérgico. O precursor do GABA é o glutamato. Quando o GABA é liberado, ele age em
receptores GABAA ou GABAB. A captação do GABA ocorre por GATs, tanto em neurônios, quanto em células
gliais (adaptado de GOVINDPANI et al., 2017).
No desenvolvimento embrionário, se acredita que o GABA possua um papel excitatório.
Na idade de E3 na retina de galinha, foi demonstrado que o GABA estimula a ativação de canais
de cálcio a partir da ativação de receptores GABAA (YAMASHITA; FUKUDA, 1993). Em E8,
o GABA passa por seu período de maior ação despolarizante, que entra em declínio e termina
na idade de E14, por onde provavelmente o GABA passa a ter seu efeito inibitório clássico
(Figura 6) (CATSICAS; MOBBS, 2001).

12
Figura 6: Desenvolvimento do sistema GABAérgico e endocanabinoide em retina de galinha. Células
GABAérgicas surgem em maior quantidade entre as idades de E6 e E9. O GABA é excitatório até E14 e sua síntese
ocorre inicialmente pela putrescina e a partir de E11 pela GAD. Os receptores CB1 começam a ser expressos em
E4 e o GAT-1 em E6. A retina apresenta seu perfil maduro em E19.
a. Transportadores
Os transportadores de alta afinidade de GABA (GATs) pertencem à família SLC6 e são
responsáveis por realizar um simporte de GABA que dependede da entrada de 2 íons de Na+
juntos a 1 íon de Cl- (Figura 7) (LOO et al., 2000; SCIMEMI, 2014). Eles são responsáveis por
regular a concentração extracelular de GABA, tanto em condições basais, quanto em eventos
sinápticos (SCIMEMI, 2014). Os GATs se encontram amplamente expressos no cérebro e são
divididos em quatro subtipos, classificados como GAT-1, GAT-2, GAT-3 e GAT-4, os quais
possuem distribuição, especificidade e sensibilidade a inibidores distintos (LIU et al., 1993).
Na retina de ratos, foi demonstrado que o GAT-1 se encontra fortemente expresso em células
amácrinas e interplexiformes. O GAT-2 foi observado no epitélio pigmentado e o GAT-3 nas
células amácrinas (JOHNSON et al., 1996).
Os GATs estão presentes tanto em neurônios, quanto em astrócitos. Nestes, o GABA
pode ser metabolizado pela GABA-transaminase a glutamina, e assim ser utilizado no neurônio
para nova produção do neurotransmissor (GOVINDPANI et al., 2017). Nas células de Müller,
devido à eficiência da GABA-transaminase, são encontrados níveis baixos de GABA

13
intracelular (MARC et al., 1998). As células da glia de Müller são responsáveis pela captação
e metabolismo do GABA, principalmente pela ação de transportadores GAT-1 e GAT-3
(BRINGMANN et al., 2013). Adicionalmente, também existe um transportador vesicular de
GABA (vGAT), que transporta o GABA para o interior das vesículas sinápticas.
Figura 7: Modelo de funcionamento do transportador GAT-1. Ocorre o simporte de 2 Na+, 1 Cl- e de 1 GABA
para dentro da célula (Adaptado de LOO et al., 2000).
b. Liberação de GABA
Classicamente, a liberação de neurotransmissores no SNC está relacionada a
mecanismos dependentes de cálcio com exocitose a partir de vesículas sinápticas. Porém, na
retina de galinha, diversos indícios apontam para a liberação do GABA por meio da reversão
de seus transportadores, por um mecanismo dependente de Na+ (CALAZA; GARDINO; DE
MELLO, 2006). Em culturas de células de retina de galinha, o L-glutamato estimulou a
liberação de [³H]-GABA, que foi bloqueada com a retirada de Na+ do meio, indicando uma
possível troca de L-glutamato e GABA por um transportador dependente de Na+. Também
foram propostas a liberação de GABA estimulada por GABA e dependente de cálcio a partir
de estímulos com o L-Aspartato (DO NASCIMENTO; DE MELLO, 1985). Além disso, foi
demonstrado que o L-Aspartato promove a liberação de [³H]-GABA agindo em receptores
NMDA, por um mecanismo que depende da captação eletrogênica de aspartato, reduzindo a
afinidade de receptores NMDA por magnésio (KUBRUSLY; DE MELLO; DE MELLO, 1998).
Por outro lado, já foi descrito na literatura a ocorrência de liberação de [³H]-GABA por um
mecanismo dependente de cálcio ocorrendo paralelamente à liberação por reversão do

14
transportador em culturas de células de retina de galinha (DUARTE et al., 1992). Sendo assim,
o projeto atual teve como proposta avaliar como o WIN 55,212-2 regula a liberação de [³H]-
GABA, além de avaliar seus efeitos crônicos em culturas mistas de retina de galinha.
OBJETIVOS
1. Objetivo geral
Avaliar o efeito dos estímulos agudos e crônicos de WIN 55,212-2 nos sistemas
GABAérgico e endocanabinoide em culturas de células mistas de retina de galinha em idade
embrionária.
2. Objetivos específicos
Avaliar a funcionalidade e expressão de receptores CB1 e CB2.
Analisar os efeitos do estímulo agudo de WIN 55,212-2 na liberação e captação de
[³H]-GABA.
Avaliar como o estímulo crônico de WIN 55,212-2 altera os níveis de AMPc,
liberação de [³H]-GABA e expressão de receptores CB1 e CB2.
Observar se a retirada do estímulo crônico de WIN 55,212-2 altera os níveis de
AMPc.

15
MATERIAIS E MÉTODOS
1. Animais e cultura de células
Ovos de galinha da raça Leghorn (Gallus gallus domesticus) fertilizados foram cedidos
pelo Laboratório de Neuroquímica da Universidade Federal do Rio de Janeiro. Foram utilizados
ovos na idade E7 e E8, por onde os embriões foram retirados com o auxílio de uma pinça curva
e rapidamente decapitados. Os olhos foram eviscerados e a retina foi dissecada em meio de
cultura DMEM/F-12 + Gentamicina. Posteriormente, as retinas foram picotadas e recolhidas
em um tubo separado, formando um pool de células. Nele ocorreu a adição de 1 ml de tripsina
(TrypLE™) por 10 minutos a 37ºC. Após a incubação, a retina foi centrifugada por 30 segundos
e o sobrenadante descartado. Depois ocorreu a adição de 1ml de DMEM/F-12 + Soro fetal
bovino (SFB) a 5% no pool para posterior dissociação mecânica. Em seguida, foi adicionado
mais DMEM/F-12 + SFB a 5% até o volume necessário para o plaqueamento. Nas placas de 12
poços, foi utilizado 1 ml por poço, já nas placas de 6 poços foram utilizados 2 ml por poço. A
proporção utilizada foi de meia retina por poço. Depois da realização do plaqueamento, as
placas foram colocadas em uma estufa a 5% de CO2 e 37ºC. O meio de cultura foi trocado de
2 em 2 dias. As culturas utilizadas são mistas, ou seja, expressam tanto neurônios, quanto
células gliais. Após 7 ou 8 dias de cultivo as placas foram utilizadas para realizar os tratamentos
e experimentos.
a. Tratamentos
Para realizar o tratamento crônico, o WIN 55,212-2 a 100 nM foi filtrado junto com o
DMEM/F-12 + 5% SFB e pré-incubado por 3 horas antes de se realizar os experimentos (Grupo
1). Nos grupos em que houve a retirada do WIN 55,212-2, foi realizada a troca do meio após o
tratamento crônico e ocorreu a adição de DMEM/F-12 + 5% SFB nos poços por 24 horas
(Grupo 2). Paralelamente, para todos grupos de exposição crônica ao WIN 55,212-2 e retirada,
existiam grupos controle com tempos de exposição idênticos, porém com a utilização de
DMEM/F-12 + 5% SFB apenas (Figura 8). Todos os procedimentos realizados estão de acordo
com o protocolo de uso e cuidados com os animas em pesquisa da Universidade Federal do Rio
de Janeiro #IBCCF035.

16
Figura 8: Linha temporal com idades e tratamento crônico com WIN 55,212-2 a 100 nM.
2. Western Blot
As culturas de células mistas foram lavadas 2 vezes com Hanks 4 completo (128 mM
NaCl, 4 mM KCL, 1 mM MgCl2, 3 mM CaCl2, 20 mM Hepes e 4 mM glicose). Posteriormente
foram adicionados 100 µl de RIPA em cada poço. As células foram raspadas e recolhidas após
o pré-tratamento observado no grupo 1. As amostras forma congeladas e a concentração de
proteínas foi estimada utilizando o método de Bradford (BRADFORD, 1976).
As amostras foram diluídas em SDS e RIPA e fervidas por 10 minutos.
Aproximadamente 45 μg de proteína de cada amostra foram utilizadas na eletroforese em 10%
SDS-PAGE e transferidas para membranas de fluoreto de polivinilideno (Millipore) por um
método de transferência semi-seco. As membranas foram lavadas com TTBS e bloqueadas por
1 h com 5% leite desnatado em TTBS. Posteriormente elas foram incubadas com o anticorpo
primário anti-CB1 (1:1000 em TTBS; Proteimax) e anti-CB2 (1:1000 em TTBS; Proteimax)
overnight a 4ºC. Em seguida elas foram lavadas com TTBS 3 vezes por 10 minutos e incubadas
com os anticorpos secundários anti-coelho para os receptores CB1 e CB2 (1:2000 em TTBS;
Sigma-Aldrich) conjugados com peroxidase por 2 horas à temperatura ambiente. Após três
lavagens em TTBS (10 minutos cada), a intensidade das bandas foi obtida aplicando-se
luminata sobre as membranas e utilizando o software ImageLab no hardware ChemiDoc. Após
a obtenção das imagens, as membranas foram marcadas novamente com anticorpo anti-α-
tubulina (1:20000 em TTBS; Sigma-Aldrich), lavadas com TTBS e incubadas com anticorpo
secundário anti-camundongo (1:4000 em TTBS) conjugado com peroxidase por 2 horas à
Grupo 1
Grupo 2

17
temperatura ambiente. Após 3 lavagens com TTBS (10 minutos cada), a análise da intensidade
das bandas foi novamente realizada.
3. Ensaio de AMPc
Após os pré-tratamentos dos grupos 1 e 2, as culturas de células mistas foram lavadas 2
vezes com Hanks 4. Posteriormente, 10 µl de Rolipram (Ro20), um inibidor de fosfodiesterase,
foram adicionados a todos os poços da placa de cultura, atingindo a concentração de 500 µM
por poço e incubados por 1 hora. O WIN 55,212-2 a 100 nM foi adicionado junto ao Ro20 nos
grupos com WIN 55,212-2 agudo que não obtiveram pré-tratamento. A reação foi parada com
a adição de ácido tricloroacético (TCA) a uma concentração final de 10%, os poços foram
raspados e o conteúdo recolhido foi congelado para ser utilizado posteriormente.
O AMPc foi purificado por métodos descritos previamente (MATSUZAWA;
NIRENBERG, 1975). Em cada amostra foram adicionados 50 µl de carreador radioativo de
AMPc, e posteriormente, as amostras passaram pela etapa de separação com a utilização de
colunas de cromatografia de troca iônica para remover o TCA e outros nucleotídeos. As
triplicatas de cada amostra foram recolhidas e a contagem ocorreu por cintilação líquida. O
valor mais alto em CPM de cada triplicata foi utilizado na dosagem do AMPc de acordo com o
método de Gilman (GILMAN, 1970). Nessa etapa, cada amostra foi incubada na presença de
PKA, uma cinase ativada por AMPc, 20 µl de [³H]-AMPc em 50 mM de tampão acetato de
sódio pH 4.0, a 4°C por 100 minutos. Após esse período, a reação foi interrompida com a adição
de 200 mM de tampão fosfato pH 6.0 e cada amostra foi filtrada utilizando-se filtros Millipore.
A radiação foi determinada por cintilação liquida. Os níveis de AMPc foram normalizados por
dosagem de proteína (LOWRY et al., 1951).
4. Liberação de [³H]-GABA
As culturas de células mistas foram lavadas 3 vezes com Hanks 4 completo para
posterior incubação de GABA carreador e [³H]-GABA por 1 hora a 37°C. Após esse período
de tempo, os poços foram novamente lavados 3 vezes com Hanks 4 e foram feitas perfusões
nos poços da placa de cultura de acordo com o grupo experimental por 5 minutos cada. As
perfusões foram feitas com WIN 55,212-2, AM-251, capsaicina, WIN 55,212-2 + AM-251 e
WIN 55,212-2 + capsaicina. Após recolher todas perfusões em tubos de vidro, foi adicionado
1 ml de água milli-Q em todos os poços da placa de cultura. Ela foi congelada e as células foram
posteriormente raspadas e recolhidas. Paralelamente, foram separados tubos com 1 ml de
líquido HiSafe, que é um coquetel líquido para cintilação. 100 µl de cada amostra e o conteúdo

18
da placa foram adicionados a eles. A radioatividade foi determinada por cintilação líquida. A
quantidade de [³H]-GABA liberado foi plotada como a porcentagem do total de radioativo
captado pelas células.
5. Captação de [³H]-GABA
As culturas de células mistas de retina de galinha foram incubadas por 1 hora com o
GABA carreador e o [³H]-GABA. Antes do período de incubação, o WIN 55,212-2 foi
adicionado por 15 minutos nos poços da placa de cultura em diversas concentrações (10 nM,
100 nM ou 1µM). Posteriormente, os poços foram lavados 3 vezes com Hanks 4 para retirar a
radioatividade que não foi absorvida pelas células. 1 ml de água milli-Q foi adicionado para
causar a ruptura das membranas celulares. Após congelamento das amostras, as células foram
raspadas e recolhidas e a radioatividade celular foi determinada por cintilação líquida.
6. Análise estatística
Os dados foram processados pelo programa GraphpadPrism 6 como médias e erros
padrões da média. A significância estatística foi obtida através de uma ANOVA de uma via,
seguida de pós-teste Bonferroni para resultados com 3 ou mais grupos experimentais. O teste t
não pareado foi utilizado para resultados com 2 grupos experimentais. Os resultados foram
expressos como média ± SEM, e foram considerados estatisticamente significativos quando
p≤0,05. O n experimental representa o número de experimentos independentes.

19
RESULTADOS
1. Os receptores CB1 e CB2 estão expressos e funcionais em culturas
mistas de retina de galinha embrionárias.
Primeiramente foram realizados experimentos para comprovar a expressão e
funcionalidade de receptores canabinoides em culturas mistas de retina de galinha. A partir da
análise de Western Blot, foi comprovado que tanto receptores CB1, quanto receptores CB2 se
encontram expressos nas culturas de idade E8C8 (Figura 9A). A funcionalidade do nosso
modelo de estudo foi avaliada pela análise do efeito do WIN 55,212-2 incubado de forma aguda
nas culturas mistas de retina de galinha E7C7. O WIN 55,212-2 agudo reduziu os níveis de
AMPc em comparação aos níveis basais (Figura 9B), o que demonstra que os receptores
canabinoides ligados ao agonista possuem atividade intrínseca.
Neurochem Int. 2018 Jan; 112:27-37
55kDa 42kDa
A B
Figura 9: Expressão e funcionalidade de receptores canabinoides A. Western Blot de receptores CB1 e CB2
em culturas mistas de retina de galinha E8C8 (n=3). B. Acúmulo de AMPc em culturas mistas de retina de galinha
E7C7, Basal (69,03±4,82 pmol/ mg/ hora; n=6), WIN 55,212-2 agudo (46,85±9,303 pmol/ mg/ hora; n=4),
(p<0,05).
Ac
úm
ulo
de
AM
Pc
(pm
ol/
mg
/ho
ra
s)
Basal
WIN
ag
ud
o
0
2 0
4 0
6 0
8 0
1 0 0
*

20
2. O WIN 55,212-2 reduz a liberação de [³H]-GABA agindo em
receptores CB1, sem alterar sua captação.
Para avaliar o efeito neuroquímico do estímulo agudo de WIN 55,212-2 em culturas
mistas de retina de galinha E7C7, foram realizados experimentos de liberação e captação do
principal neurotransmissor inibitório do SNC, o GABA. Primeiramente foi observado que o
WIN 55,212-2 agudo, a uma concentração de 100 nM reduz a liberação de [³H]-GABA em
21%, quando comparado aos níveis basais (Figura 10A). Diferente do que foi observado com a
liberação, os níveis de captação de [³H]-GABA não são alterados, mesmo com o estímulo de
concentrações diferentes de WIN 55,212-2 de 10 nM, 100 nM e 1µM (Figura 10B).
Para avaliar por quais tipos de receptores a redução na liberação do [³H]-GABA pode
ocorrer, foi realizada a liberação desse neurotransmissor na presença do AM-251, antagonista
CB1, e na presença da capsaicina, que é um agonista TRPV1, ambos na concentração de 1µM.
O AM-251 não altera os níveis de liberação de [³H]-GABA, porém no grupo em que ele foi
incubado junto com o WIN 55,212-2, foi possível observar uma inibição do efeito do WIN
55,212-2, que é de reduzir a liberação do [³H]-GABA (Figura 10C). Por fim, a capsaicina não
altera os níveis basais da liberação de [³H]-GABA, porém no grupo capsaicina + WIN 55,212-
2 ocorre uma redução na liberação de [³H]-GABA a níveis idênticos aos observados no grupo
WIN 55,212-2 a 100 nM (Figura 10D), demonstrando que a capsaicina não influencia na
redução da liberação de [³H]-GABA causada pelo WIN 55,212-2.
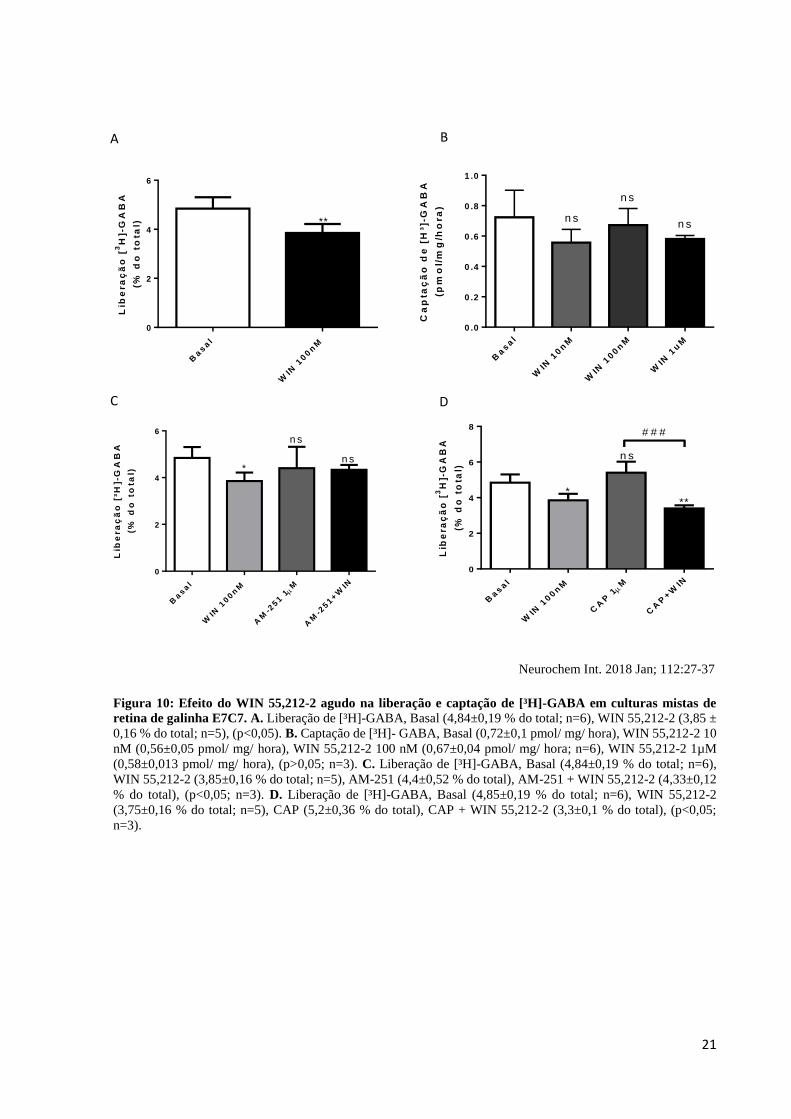
21
Neurochem Int. 2018 Jan; 112:27-37
Figura 10: Efeito do WIN 55,212-2 agudo na liberação e captação de [³H]-GABA em culturas mistas de
retina de galinha E7C7. A. Liberação de [³H]-GABA, Basal (4,84±0,19 % do total; n=6), WIN 55,212-2 (3,85 ±
0,16 % do total; n=5), (p<0,05). B. Captação de [³H]- GABA, Basal (0,72±0,1 pmol/ mg/ hora), WIN 55,212-2 10
nM (0,56±0,05 pmol/ mg/ hora), WIN 55,212-2 100 nM (0,67±0,04 pmol/ mg/ hora; n=6), WIN 55,212-2 1µM
(0,58±0,013 pmol/ mg/ hora), (p>0,05; n=3). C. Liberação de [³H]-GABA, Basal (4,84±0,19 % do total; n=6),
WIN 55,212-2 (3,85±0,16 % do total; n=5), AM-251 (4,4±0,52 % do total), AM-251 + WIN 55,212-2 (4,33±0,12
% do total), (p<0,05; n=3). D. Liberação de [³H]-GABA, Basal (4,85±0,19 % do total; n=6), WIN 55,212-2
(3,75±0,16 % do total; n=5), CAP (5,2±0,36 % do total), CAP + WIN 55,212-2 (3,3±0,1 % do total), (p<0,05;
n=3).
Lib
era
çã
o [
3H
]-G
AB
A
(% d
o t
ota
l)
Basal
WIN
100n
M
0
2
4
6
**
Lib
era
çã
o [
³H]-
GA
BA
(% d
o t
ota
l)
Basal
WIN
100n
M
AM
-251 1M
AM
-251+W
IN
0
2
4
6
*
n s
n s
Lib
era
çã
o [
3H
]-G
AB
A
(% d
o t
ota
l)
Basal
WIN
100n
M
CA
P 1M
CA
P+
WIN
0
2
4
6
8
***
# # #
n s
A
C D
B
Basal
WIN
10n
M
WIN
100n
M
WIN
1u
M
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
Ca
pta
çã
o d
e [
H³]
-GA
BA
(pm
ol/
mg
/ho
ra
)
n s
n s
n s

22
3. A exposição crônica das culturas ao WIN 55,212-2 reduz a expressão
do receptor CB1, mas não do CB2.
Após avaliar o efeito agudo do WIN 55,212-2, foi estudado o efeito crônico desse
agonista sintético misto nas culturas mistas de retina de galinha. O pré-tratamento crônico de 3
horas com o WIN 55,212-2 é responsável por aumentar em 30% a liberação de [³H]-GABA em
comparação ao grupo que obteve apenas o estímulo agudo do WIN 55,212-2 a 100 nM, o qual
reduz a liberação de [³H]-GABA (Figura 11A). Além disso, por análise de Western Blot é
possível observar que o pré-tratamento crônico de 3 horas com o WIN 55,212-2 também é
responsável por reduzir a quantidade total de receptores CB1, sem alterar os níveis de receptores
CB2 nas culturas, quando comparado aos respectivos valores controle (Figuras 11B e C).
Por último, foi realizado um AMPc em que os valores de cada grupo experimental foram
divididos pelos seus níveis basais, gerando uma relação estimulado/basal. O WIN 55,212-2
agudo a 100 nM reduz os níveis de AMPc das culturas, enquanto que com o pré-tratamento de
3 horas do agonista canabinoide misto já é possível observar um aumento acima dos níveis
basais, porém não significativo em comparação ao grupo agudo. Já com a retirada de 24 horas
do WIN 55,212-2 após o estímulo crônico, os níveis de AMPc aumentam de forma significativa
quando comparado ao grupo WIN 55,212-2 agudo (Figura 11D).
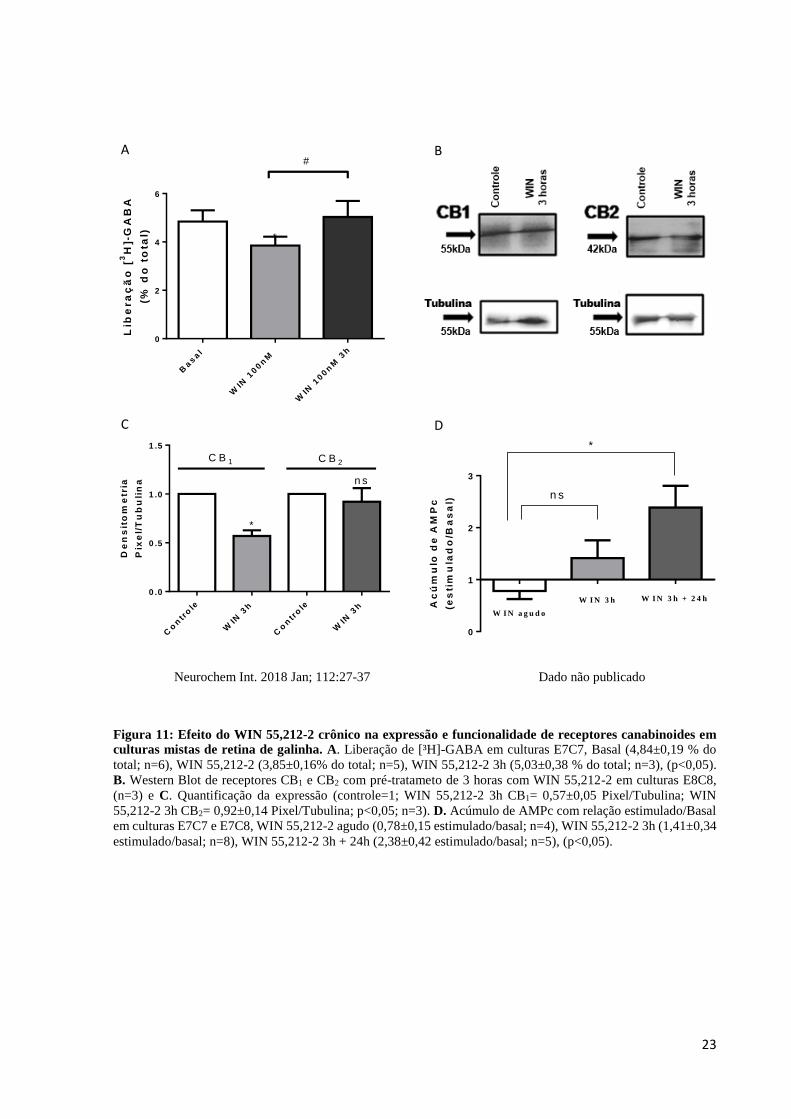
23
Neurochem Int. 2018 Jan; 112:27-37 Dado não publicado
Figura 11: Efeito do WIN 55,212-2 crônico na expressão e funcionalidade de receptores canabinoides em
culturas mistas de retina de galinha. A. Liberação de [³H]-GABA em culturas E7C7, Basal (4,84±0,19 % do
total; n=6), WIN 55,212-2 (3,85±0,16% do total; n=5), WIN 55,212-2 3h (5,03±0,38 % do total; n=3), (p<0,05).
B. Western Blot de receptores CB1 e CB2 com pré-tratameto de 3 horas com WIN 55,212-2 em culturas E8C8,
(n=3) e C. Quantificação da expressão (controle=1; WIN 55,212-2 3h CB1= 0,57±0,05 Pixel/Tubulina; WIN
55,212-2 3h CB2= 0,92±0,14 Pixel/Tubulina; p<0,05; n=3). D. Acúmulo de AMPc com relação estimulado/Basal
em culturas E7C7 e E7C8, WIN 55,212-2 agudo (0,78±0,15 estimulado/basal; n=4), WIN 55,212-2 3h (1,41±0,34
estimulado/basal; n=8), WIN 55,212-2 3h + 24h (2,38±0,42 estimulado/basal; n=5), (p<0,05).
Lib
era
çã
o [
3H
]-G
AB
A
(% d
o t
ota
l)
Basal
WIN
100n
M
WIN
100n
M 3
h
0
2
4
6
*
#A
C A
cú
mu
lo d
e A
MP
c
(es
tim
ula
do
/Ba
sa
l)
0
1
2
3
W I N a g u d o
W I N 3 h W I N 3 h + 2 4 h
n s
*
D
B
De
ns
ito
me
tria
Pix
el/
Tu
bu
lin
a
Co
ntr
ole
WIN
3h
Co
ntr
ole
WIN
3h
0 .0
0 .5
1 .0
1 .5
n s
*
C B 1 C B 2

24
DISCUSSÃO
Neste projeto foi avaliado como o sistema endocanabinoide modula o sistema
GABAérgico e a disponibilidade de receptores canabinoides no período de desenvolvimento
embrionário em culturas de células mistas de retina de galinha após exposições agudas e
crônicas ao agonista canabinoide WIN 55,212-2. Primeiramente, foi demonstrado que os
receptores CB1 e CB2 se encontram expressos e funcionais. Também demonstramos que a
ativação de receptores CB1 leva à redução da liberação de GABA, sem alterar seus níveis de
captação, e que a exposição crônica a canabinoides leva a uma redução da expressão desses
receptores. Por último, a retirada do estímulo crônico por 24 horas é capaz de elevar os níveis
de AMPc celulares, por um mecanismo que pode envolver receptores CB2.
Para avaliar a funcionalidade do nosso modelo de estudo, foi comprovado que tanto
receptores CB1, quanto receptores CB2 se encontram expressos na retina de galinha nas idades
de E8C8, cuja idade equivalente é de E16. A maioria dos nossos experimentos foram realizados
na idade de E7C7, com idade equivalente de E14. Entre essas idades, foi demonstrado que
ocorre uma intensa formação de sinapses convencionais, com anatomia característica de
sinapses realizadas por células amácrinas e bipolares, logo nossos experimentos foram
realizados em um período de sinaptogênese ativa (GLEASON; WILSON, 1989). Como as
nossas culturas são mistas, nós levamos em consideração que tanto neurônios, quanto células
da glia de Müller expressam os receptores canabinoides. Em culturas mistas de retina de galinha
E8C6, foi demonstrado por imunocitoquímica que tanto receptores CB1, quanto CB2, se
encontram expressos em neurônios e nas células da glia de Müller (KUBRUSLY et al., 2018).
Recentemente, foi demonstrado que na retina de galinha os receptores CB1 e CB2 estão
expressos nas idades embrionárias de E5, E7, E9, E14 (SAMPAIO et al., 2018). Em E14 e E16,
os receptores CB1 foram encontrados expressos majoritariamente na CPI e CCG, e os receptores
CB2 na CPI (SAMPAIO et al., 2018; LEONELLI et al., 2005). Em relação à expressão do
sistema GABAérgico na retina de galinha embrionária, foi encontrada imunoreatividade para o
GABA em células horizontais, amácrinas e em células da CCG (POHL-GUIMARÃES et al.,
2010). Logo, levando em consideração a idade equivalente das nossas culturas de E14 e E16, e
analisando o perfil da formação de sinapses, expressão de receptores canabinoides e de GABA
no desenvolvimento embrionário da retina de galinha, a localização provável dos receptores
canabinoides em nosso modelo de estudo está em células amácrinas da CPI e/ou em células
ganglionares da CCG.

25
Apesar de os receptores CB1 poderem estar acoplados a proteínas Gq/11, Gs ou Go
(HOWLETT et al., 2002; BOW; RIMOLDI, 2016), ou ativar outras vias de sinalização, como
a via da MAPK (DI MARZO et al., 2015), nossos resultados demonstraram que em culturas
mistas de retina galinha os receptores CB1 e CB2 ativam proteínas Gi, o que inibe a adenilil-
ciclase e reduz os níveis de AMPc. Essa conclusão foi tirada, pois o estímulo do WIN 55,212-
2 de forma aguda nas culturas levou à redução dos níveis de AMPc, o que demonstrou que elas
se encontram funcionais. O mesmo efeito foi observado em neurônios hipocampais, onde foi
demonstrado que o WIN 55,212-2 aumentou correntes de K+ dependentes de voltagem a partir
da inibição da adenilil-ciclase e redução dos níveis de AMPc (DEADWYLER et al., 1995).
A alteração dos níveis de AMPc é capaz de afetar diretamente o desenvolvimento
correto das células. Em neurônios da retina de galinha já foi demonstrado que a elevação dos
níveis de AMPc inibe a motilidade de cones de crescimento (LANKFORD; DE MELLO;
KLEIN, 1988). No SNC em desenvolvimento, já foi demonstrado que o AMPc estimula a
diferenciação neuronal, além de influenciar na sobrevivência celular, plasticidade sináptica e
formação de memórias (INDA et al., 2017). Sendo assim, é possível que o sistema
endocanabinoide no nosso modelo de estudo esteja afetando o desenvolvimento de neurônios
no período embrionário pela regulação dos níveis de AMPc.
Nós também demonstramos que há uma alteração neuroquímica do sistema
GABAérgico quando exposto a canabinoides, em que o WIN 55,212-2 agudo foi capaz de
reduzir a liberação do [³H]-GABA em relação aos níveis basais. Nós utilizamos a concentração
de 100 nM do agonista canabinoide misto, que também foi capaz de reduzir a liberação de
GABA em interneurônios hipocampais de ratos de maneira dose-dependente (0,01-3µM)
(KATONA et al., 1999). O WIN 55,212-2 também reduziu a liberação do GABA em neurônios
nigroestriatais (WALLMICHRATH; SZABO, 2002). Apesar de o WIN 55,212-2 ser mais
reconhecido por seus efeitos inibitórios na liberação de [³H]-GABA, já foi descrito que no globo
pálido de ratos o WIN 55,212-2 possui um efeito bimodal na liberação de [³H]-GABA, ou seja,
de acordo com a concentração ele pode aumentar ou reduzi-la (GONZALEZ et al., 2009).
Apesar de o efeito do sistema endocanabinoide na regulação da liberação de diversos
neurotransmissores ser bem caracterizado no sistema nervoso maduro, o nosso projeto aborda
esse efeito no desenvolvimento embrionário da retina de galinha, que é um tema com poucos
estudos sobre.

26
Para descobrir em qual receptor o WIN 55,212-2 estava agindo para reduzir a liberação
de [³H]-GABA, nós avaliamos como o antagonista CB1, AM-251, e o agonista de receptores
TRPV1, capsaicina, modulariam o efeito do WIN 55,212-2. Nós utilizamos a capsaicina, pois
já foi demonstrado que a AEA também é capaz de ativar os receptores TRPV1 (TÓTH;
BLUMBERG; BOCZÁN, 2009), logo há a possibilidade de uma interação entre o sistema
endocanabinoide e o receptor TRPV1 regulando a liberação de [³H]-GABA.
Quando o WIN 55,212-2 foi incubado junto com o AM-251, foi observada a inibição
da redução na liberação de [³H]-GABA. Isso demonstrou que a redução na liberação desse
neurotransmissor está ocorrendo por receptores CB1, já que seu antagonista impediu a ação do
agonista canabinoide misto. Além disso, pelo fato de o AM-251 poder ativar receptores GPR55
(RYBERG et al., 2007), que aumentam a quantidade de cálcio intracelular, é pouco provável
que a liberação de [³H]-GABA esteja ocorrendo por via exocítica em nosso modelo de estudo,
já que o AM-251 incubado sozinho não aumentou a liberação de [³H]-GABA.
A ação de antagonista ou agonista inverso do AM-251 depende da concentração em que
ele está sendo utilizado. Em concentrações nanomolar, o AM-251 age como antagonista
seletivo, e em concentrações micromolar já foi observada atividade de agonista inverso
(SAVINAINEN et al., 2003). Porém em concentrações micromolar baixas, como a de 1 µM, já
foi demonstrado em cerebelos de ratos que o AM-251 não possui atividade de agonista inverso
(SAVINAINEN et al., 2001). Em nossos resultados, o AM-251 não alterou os níveis de
liberação de [³H]-GABA em comparação aos níveis basais. Isso demonstra que provavelmente
ele está agindo como antagonista e não como agonista inverso, já que se esperaria de um
agonista inverso um aumento da liberação de [³H]-GABA. Em hipocampos de ratos já foi
demonstrado que o AM-251 a 2 µM aumenta a liberação de GABA (PERTWEE, 2005), o que
indica um antagonismo inverso que não foi observado em nossos resultados.
Nós descartamos a ação de receptores TRPV1 modulando a liberação de [³H]-GABA,
pois no grupo CAP + WIN não foram observadas alterações na liberação de [³H]-GABA,
quando comparado ao efeito do WIN 55,212-2, apenas. Isso indica que o WIN 55,212-2 está
reduzindo a liberação de [³H]-GABA sem que ocorra ação modulatória da capsaicina. Nós
também desconsideramos o efeito do WIN 55,212-2 sobre receptores GPR55, pois já foi
descrito que ele não ativa e nem se liga a essa classe de receptores (LAUCKNER et al., 2008).
Em relação à localização celular dos receptores canabinoides, foi demonstrado na retina
de galinha que na idade de E14 os receptores CB1 possuem localização pré-sináptica, devido a

27
sua co-localização com a sinaptofisina, que é um marcador pré-sináptico, na CPI e na CCG
(KUBRUSLY et al., 2018). A liberação de GABA na retina de galinha ocorre majoritariamente
por reversão de seu transportador, que é dependente de Na+ (CALAZA; GARDINO; DE
MELLO, 2006). Vários mecanismos foram propostos para essa reversão, que pode ser induzida
por despolarização da célula, a qual pode ser estimulada pelo aumento extracelular de K+ ou
pelo glutamato. Também é possível que ocorra por homotroca com o GABA (SCHWARTZ,
1982). Na retina de peixes dourados, foi demonstrado que a liberação de [³H]-GABA ocorre
principalmente pelo transportador de alta afinidade do GABA a partir do influxo de Na+ em
células horizontais (YAZULLA; KLEINSCHMIDT, 1983). Em culturas de célula de retina de
galinha de idade E8C7, o uso de antagonistas para o GAT-1 demonstrou que a liberação de
GABA provavelmente ocorre pela reversão de um transportador que apresenta apenas algumas
características farmacológicas do GAT-1 (DO NASCIMENTO; VENTURA; PAES DE
CARVALHO, 1998).
Um mecanismo proposto para a redução da liberação de [³H]-GABA pelo WIN 55,212-
2 agudo pode ser a hiperpolarização causada pela ativação de receptores canabinoides pré-
sinápticos, os quais aumentam a condutância de K+ pela redução da ativação da PKA
(SVÍŽENSKÁ; DUBOVÝ; ŠULCOVÁ, 2008), fazendo com que o K+ saia da célula e possa
reduzir a atividade dos transportadores de alta afinidade do GABA. Os GATs na retina de
galinha também podem ser regulados pela ação do Ca2+ ou pela fosforilação de sítios
específicos (DO NASCIMENTO et al., 2013). Em retina de galinha, foi demonstrado que a
cafeína estimula a liberação de [³H]-GABA via GAT-1, por um mecanismo que ativa a via da
PKA (FERREIRA et al., 2014). Em terminais nervosos, foi observado que o aumento de AMPc
e PKA estimula o transporte de GABA por redução da atividade inibitória da PKC sobre o GAT-
1 (CRISTÓVÃO-FERREIRA et al., 2009). Nossos resultados demonstraram que o WIN
55,212-2 reduz os níveis de AMPc, o que leva a uma redução da ativação da PKA. Logo, outro
mecanismo possível é a modulação da fosforilação do GAT-1 pela PKA, reduzindo a liberação
de [³H]-GABA.
No entanto, uma redução na liberação de um neurotransmissor não significa
necessariamente que sua quantidade nas células da retina está sendo alterada. Ela pode estar
sendo acompanhada por uma redução na captação a níveis correspondentes, fazendo com que
no fim das contas não ocorram alterações na quantidade de GABA celular. Nossos resultados
demonstraram que concentrações diferentes de WIN 55,212-2 (10 nM, 100 nM e 1 µM) não
alteram os níveis de captação do [³H]-GABA em relação aos níveis basais nas culturas mistas

28
de retina de galinha. Como consequência, a ativação de receptores canabinoides está sendo
responsável por causar uma alteração na homeostase do GABA, com redução na liberação,
porém sem alteração na captação. Esse efeito também foi observado no globo pálido de ratos,
onde foi demonstrado que o WIN 55,212-2 não altera a captação de [³H]-GABA em diferentes
concentrações, de 1 a 100 µM (VENDEROVA; BROWN; BROTCHIE, 2005).
A modulação do sistema GABAérgico pelo canabinoide no desenvolvimento
embrionário é relevante, pois o sistema GABAérgico é de extrema importância para o
desenvolvimento correto do SNC. Na retina, o GABA atua como fator trófico, influenciando na
disponibilidade de receptores GABAérgicos (MADTES; REDBURN, 1983). O GABA também
influencia o ciclo celular na fase de desenvolvimento neuronal, tendo efeitos na proliferação,
diferenciação celular e sinaptogênese (LOTURCO et al., 1995). Em relação ao sistema
endocanabinoide no desenvolvimento embrionário, se sabe que canabinoides são capazes de
modular a proliferação de progenitores e influenciar o destino de células-tronco neurais, além
de refinar funções sinápticas em circuitarias neuronais complexas durante o desenvolvimento
(HARKANY; MACKIE; DOHERTY, 2008). Logo, além de alterar a excitabilidade de
circuitarias de células neuronais e gliais na retina de galinha, o sistema endocanabinoide pode
estar modulando a formação de células retinianas.
Além de analisar o efeito agudo, também foi analisado o efeito crônico do WIN 55,212-
2 sobre as culturas mistas de retina de galinha. Primeiramente nós observamos que o pré-
tratamento crônico com o WIN 55,212-2 reverteu a redução na liberação de [³H]-GABA, que
foi observada no grupo WIN 100 nM agudo, o que aponta para uma down-regulation de
receptores canabinoides, pois estando eles menos expressos nas membranas celulares, é
esperado que eles não reduzam mais a liberação do [³H]-GABA. Corroborando com a alteração
neuroquímica observada, também foi observado que o pré-tratamento crônico com WIN
55,212-2 nas culturas mistas de retina de galinha reduziu a expressão proteica de receptores
CB1, mas não de receptores CB2. Logo chegamos à conclusão que em nosso modelo de estudo,
está ocorrendo a down-regulation de receptores CB1 pelo estímulo crônico de 3 horas do
canabinoide misto WIN 55,212-2.
Em ratos Sprague-Dawley foi demonstrado que um tratamento crônico de 4 dias com
Δ9-THC reduziu o número de locais de alta afinidade para o [³H]-WIN 55,212-2 em regiões
como o córtex frontal, cerebelo, hipocampo e gânglios basais. Esses resultados indicam a
ocorrência da down-regulation de receptores canabinoides (BREIVOGEL et al., 2003). No

29
mesmo modelo animal, um tratamento crônico com injeção diária de WIN 55,212-2 por 20 dias
foi responsável por reduzir os níveis de binding específico ao agonista canabinoide [³H]-
CP55,940, além de reduzir a expressão do mRNA de receptores CB1 no estriado dos animais
(PERDIKARIS et al., 2018). Um estudo que avaliou a down-regulation de receptores
canabinoides após exposições crônicas de THC por tempos diferentes diferentes (1, 3, 7 ou 14
dias) em diversas regiões cerebrais, mostrou que no hipocampo ocorre uma redução do binding
após a primeira injeção e no globo pálido após mais de 3 dias, ou seja, ela é variável de acordo
com a região analisada (ROMERO et al., 1998).
A maioria dos estudos publicados envolvendo o efeito crônico de canabinoides é
realizado no cérebro e com animais em idades pós-natais. Nós analisamos o efeito do WIN
55,212-2 crônico no período embrionário em culturas de células mistas de retina de galinha, e
o tempo de exposição de 3 horas foi considerado crônico devido ao nosso modelo de estudo,
que avalia o SNC em proporções menores quando comparado aos outros estudos que utilizaram
modelos in vivo com sistemas biológicos mais complexos, e devido a isso fizeram exposições
mais longas de canabinoides.
O pré-tratamento crônico de 3 horas com o WIN 55,212-2 também foi responsável por
elevar os níveis de AMPc, porém foi após a retirada por 24 horas do estímulo crônico do WIN
55,212-2 que foi possível observar um aumento significativo dos níveis de AMPc. Pelo fato de
os receptores canabinoides classicamente reduzirem os níveis de AMPc, se espera que com a
down-regulation desses receptores ocorra um aumento de AMPc. No grupo em que ocorreu o
tratamento crônico com o WIN 55,212-2 por 3 horas, está ocorrendo apenas a down-regulation
dos receptores CB1, mas não dos receptores CB2. Como consequência, os receptores CB2 ainda
estão expressos e funcionais nas membranas celulares, e contribuindo para que os níveis de
AMPc não subam para níveis significativos em comparação ao grupo WIN 55,212-2 agudo.
Porém após as 24 horas de retirada, é possível que os receptores CB2 tenham sofrido
down-regulation. Pelo fato de o WIN 55,212-2 possuir uma afinidade muito mais elevada pelos
receptores canabinoides em comparação aos endocanabinoides, é provável que mesmo após a
lavagem dos poços das placas de cultura e retirada dos canabinoides do meio, o WIN 55,212-2
tenha continuado ligado aos receptores CB2. Em receptores CB2 humanos clonados, foi
demonstrado que a constante de inibição para o WIN 55,212-2 é aproximadamente 600 vezes
menor que a constante da AEA, o que mostra uma afinidade maior do WIN 55,212-2 pelo
receptor CB2 em comparação a AEA (FELDER et al., 1995).

30
Além disso, foi demonstrado que em humanos o WIN 55,212,2 apresenta uma afinidade
maior para os receptores CB2 que para os receptores CB1, com constantes de dissociação de 3,7
nM e 16,7 nM, respectivamente (MCPARTLAND; GLASS; PERTWEE, 2007). Porém em ratos
foi observado o oposto, onde as constantes de dissociação para os receptores CB2 e CB1 foram
de 35,6 nM e 2,4 nM, respectivamente (MCPARTLAND; GLASS; PERTWEE, 2007),
demonstrando uma afinidade maior do WIN 55,212-2 por receptores CB1 em ratos. Sendo
assim, é correto afirmar que o WIN 55,212-2 possui afinidades distintas para ambos receptores
canabinoides, as quais variam de acordo com a espécie analisada. Isso pode ser o responsável
direto pela down-regulation dos receptores CB1 e CB2 em tempos diferentes de tratamento em
culturas mistas de retina de galinha. Logo, é possível que a down-regulation tardia dos
receptores CB2 esteja aumentando os níveis de AMPc, já que nesse estágio do tratamento,
ambos receptores canabinoides estariam internalizados nas células e sem funcionalidade.
Outra possibilidade para explicar o aumento de AMPc após as 24 horas de retirada do
estímulo crônico de WIN 55,212-2 é o possível aumento da produção de endocanabinoides
como um mecanismo compensatório para a down-regulation de receptores CB1 após o estímulo
crônico do agonista canabinoide. Como consequência, os receptores CB2 podem estar sofrendo
uma down-regulation tardia por efeito dos próprios endocanabinoides. Nós descartamos a
ocorrência de uma up-regulation de receptores CB1 após a retirada de 24 horas do estímulo
crônico, pois nesse caso se esperaria que não ocorresse uma elevação dos níveis de AMPc.
O abuso de Cannabis durante a gravidez é considerado um fator de risco na toxicologia
reprodutiva e está relacionado com danos ao feto (SILVA; YONAMINE, 2018). Já foi
demonstrado que o uso pré-natal de Cannabis pode gerar alterações comportamentais na
criança, como hiperatividade, impulsividade e desatenção, além de poder causar alterações
cognitivas executivas, como capacidade de resolver problemas, atenção prolongada, raciocínio
e memória de trabalho (HUIZINK; MULDER, 2006). Uma teoria proposta para o fato de o uso
de Cannabis durante a gravidez não gerar inicialmente problemas graves no feto, é a teoria do
double-hit. Ela se baseia na ideia de que o uso de canabinoides no período pré-natal pode agir
apenas como um primeiro fator para o futuro desenvolvimento de problemas neurológicos mais
acentuados. A Cannabis pré-natal estaria desregulando o sistema endocanabinoide, e isso pode
ser responsável pela vulnerabilidade do feto a fatores externos no futuro, o que seria o segundo
hit responsável por gerar desordens neurológicas (RICHARDSON; HESTER; MCLEMORE,
2016).

31
Apesar de existirem evidências para os efeitos negativos do uso pré-natal de Cannabis
no desenvolvimento embrionário, esse é um tema ainda muito discutido, devido a limitações
metodológicas em estudos com humanos. O uso de animais permite o controle das condições
ambientais dos experimentos, e logo pode ser uma ferramenta útil para a exploração sistemática
de mecanismos neuroquímicos.
Portanto, levando em consideração o papel fundamental do sistema endocanabinoide no
desenvolvimento embrionário do SNC, foi demonstrado nesse projeto que a adição exógena de
canabinoides pode ser responsável por desregular seu desenvolvimento, com a ocorrência da
modulação da excitabilidade celular em culturas mistas de retina de galinha embrionárias, além
da regulação da expressão de receptores canabinoides após respectivos estímulos agudos e
crônicos do WIN 55,212-2.

32
CONCLUSÃO
Os receptores CB1 e CB2 estão expressos e funcionais em culturas mistas de retina
de galinha embrionárias.
O estímulo agudo de WIN 55,212-2 nas culturas reduz a liberação de [³H]-GABA,
porém concentrações diferentes do mesmo agonista não alteram os níveis de
captação. Logo está ocorrendo uma desregulação na homeostase do GABA.
O WIN 55,212-2 reduz a liberação de [³H]-GABA agindo em receptores CB1.
O estímulo crônico de 3 horas com WIN 55,212-2 causa a down-regulation de
receptores CB1, mas não de receptores CB2.
A retirada do estímulo crônico do WIN 55,212-2 por 24 horas leva a down-
regulation dos receptores CB2.
PERSPECTIVAS
Analisar a expressão de receptores CB1 e CB2 após a retirada de 24 horas do estímulo
crônico de WIN 55,212-2 por Western Blot.
Analisar a expressão de tirosina hidroxilase após estímulos crônicos e retirada do
WIN 55,212-2 por Western Blot e imunocitoquímica.
Analisar a expressão de receptores CB1 e CB2 após a retirada de 24 horas do estímulo
crônico de WIN 55,212-2 por imunocitoquímica.
Avaliar se ocorre alteração na liberação de [³H]-GABA após 24 horas de retirada do
estímulo crônico de WIN 55,212-2.
Identificar por qual via de sinalização o WIN 55,212-2 reduz a liberação de [³H]-
GABA, utilizando a forscolina simultaneamente ao estímulo com WIN 55,212-2 e
analisando se ocorre prevenção da redução na liberação de [³H]-GABA.
Analisar os níveis de anandamida, 2-AG e enzimas de degradação após estímulos
crônicos e de retirada do WIN 55,212-2.

33
REFERÊNCIAS BIBLIOGRÁFICAS
AMERICAN PSYCHIATRY ASSOCIATION. DSM-V-TR - Manual Diagnóstico e
Estatístico de Transtornos Mentais. Porto Alegre: ARTMED, 2013.
ANDÓ, R. D. et al. The inhibitory action of exo- and endocannabinoids on [3H]GABA
release are mediated by both CB1and CB2receptors in the mouse hippocampus.
Neurochemistry International, v. 60, n. 2, p. 145–152, 2012.
ASHTON, J. C. et al. Cannabinoid CB1 and CB2 receptor ligand specificity and the
development of CB2-selective agonists. Current Medicinal Chemistry, v. 15, n. 14, p. 1428–
1443, 2008.
BEGBIE, J.; DOHERTY, P.; GRAHAM, A. Cannabinoid receptor, CB1, expression follows
neuronal differentiation in the early chick embryo. Journal of Anatomy, v. 205, n. 3, p. 213–
218, 2004.
BELTRAMO, M. et al. Functional role of high-affinity anandamide transport, as revealed by
selective inhibition. Science (New York, N.Y.), v. 277, n. 5329, p. 1094–7, 22 ago. 1997.
BOUCHARD, J. F. et al. Expression and function of the endocannabinoid system in the retina
and the visual brain. Neural Plasticity, v. 2016, p. 1-14, 2016.
BOUSKILA, J. et al. Rod photoreceptors express GPR55 in the adult vervet monkey retina.
PLoS ONE, v. 8, n. 11, 2013.
BOW, E. W.; RIMOLDI, J. M. The Structure – Function Relationships of Classical
Cannabinoids : CB1 / CB2 Modulation. Perspectives in Medicinal Chemistry, v. 1940, p. 17–
39, 2016.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, v.
72, n. 1–2, p. 248–254, 1976.
BREIVOGEL, C. S. et al. The effects of Δ9-tetrahydrocannabinol physical dependence on
brain cannabinoid receptors. European Journal of Pharmacology, v. 459, n. 2–3, p. 139–150,
2003.
BRINGMANN, A. et al. GABA and glutamate uptake and metabolism in retinal glial
(Müller) cells. Frontiers in Endocrinology, v. 4, n. APR, p. 1–14, 2013.
BURNSTOCK, G. Autonomic Neurotransmission: 60 Years Since Sir Henry Dale. Annual
Review of Pharmacology and Toxicology, v. 49, n. 1, p. 1–30, 2009.
CADAS, H.; DI TOMASO, E.; PIOMELLI, D. Occurrence and biosynthesis of endogenous
cannabinoid precursor, N-arachidonoyl phosphatidylethanolamine, in rat brain. The Journal of
Neuroscience, v. 17, n. 4, p. 1226–42, 1997.
CALAZA, K. C.; GARDINO, P. F.; DE MELLO, F. G. Transporter mediated GABA release
in the retina: Role of excitatory amino acids and dopamine. Neurochemistry International, v.
49, n. 8, p. 769–777, 2006.
CALAZA, K. C.; GARDINO, P. F. Neurochemical phenotype and birthdating of specific cell
populations in the chick retina. Anais da Academia Brasileira de Ciencias, v. 82, n. 3, p. 595–
608, 2010.

34
CALAZA, K. C.; HOKOÇ, J. N.; GARDINO, P. F. Neurogenesis of GABAergic cells in the
chick retina. International journal of developmental neuroscience : the official journal of the
International Society for Developmental Neuroscience, v. 18, n. 8, p. 721–6, dez. 2000.
CATSICAS, M; MOBBS, P. GABAb receptors regulate chick retinal calcium waves. The
Journal of neuroscience : the official journal of the Society for Neuroscience, v. 21, n. 3, p.
897–910, 2001.
CHAVES, G. P. et al. Retinal removal up-regulates cannabinoid CB1 receptors in the chick
optic tectum. Journal of Neuroscience Research, v. 86, n. 7, p. 1626–1634, 2008.
COSTA, J. L. P. G. et al. Neurobiology of Cannabis: from the endocannabinoid system to
cannabis-related disorder. Jornal Brasileiro de Psiquiatria, v. 60, n. 11, p. 111–122, 2011.
CRISTÓVÃO-FERREIRA, S. et al. Adenosine A2A receptors enhance GABA transport into
nerve terminals by restraining PKC inhibition of GAT-1. Journal of Neurochemistry, v. 109,
n. 2, p. 336–347, 2009.
DE GODOY-MATOS, A. F. et al. The endocannabinoid system: a new paradigm in the
metabolic syndrome treatment. Arquivos brasileiros de endocrinologia e metabologia, v. 50,
n. 2, p. 390–399, 2006.
DE MELLO, F G; BACHRACH, U; NIRENBERG, M. Ornithine and glutamic acid
decarboxylase activities in the developing chick retina. J Neurochem, v. 27, n. 4, p. 847–851,
1976.
DEADWYLER, S. A. et al. Cannabinoids modulate voltage sensitive potassium A-current in
hippocampal neurons via a cAMP-dependent process. The Journal of pharmacology and
experimental therapeutics, v. 273, n. 2, p. 734–43, maio 1995.
DEVANE, W. A. et al. Isolation and structure of a brain constituent that binds to the
cannabinoid receptor. Science, v. 258, n. 5090, p. 1946–1949, 1992.
DEVANE, W. A. et al. Determination and characterization of a cannabinoid receptor in rat
brain. Molecular Pharmacology, v. 34, n. 5, p. 605–613, 1988.
DI MARZO, V. et al. Endocannabinoid signalling and the deteriorating brain. Nature Reviews
Neuroscience, v. 16, n. 1, p. 30–42, 2015.
DO NASCIMENTO, J. L. et al. GABA and glutamate transporters: New events and function
in the vertebrate retina. Psychology and Neuroscience, v. 6, n. 2, p. 145–150, 2013.
DO NASCIMENTO, J. L.; DE MELLO, F. G. Induced release of gamma-aminobutyric acid
by a carrier-mediated, high-affinity uptake of L-glutamate in cultured chick retina cells.
Journal of neurochemistry, v. 45, n. 6, p. 1820–7, dez. 1985.
DO NASCIMENTO, J. L.; VENTURA, A. L.; PAES DE CARVALHO, R. Veratridine- and
glutamate-induced release of [3H]-GABA from cultured chick retina cells: Possible
involvement of a GAT-1-like subtype of GABA transporter. Brain Research, v. 798, n. 1–2,
p. 217–222, 1998.
DOH, S. T. et al. Analysis of retinal cell development in chick embryo by
immunohistochemistry and in ovo electroporation techniques. BMC Development Biology, v.
10, n. 8, p. 1–16, 2010.

35
DOS SANTOS, R. G. et al. Phytocannabinoids and epilepsy. Journal of Clinical Pharmacy
and Therapeutics, v. 40, n. 2, p. 135–143, 2015.
DUARTE, C. B. et al. Ca2+-dependent release of [3H]GABA in cultured chick retina cells.
Brain Research, v. 591, n. 1, p. 27–32, 1992.
ELSOHLY, M. A.; SLADE, D. Chemical constituents of marijuana: The complex mixture of
natural cannabinoids. Life Sciences, v. 78, n. 5, p. 539–548, 2005.
FELDER, C. C. et al. Comparison of the pharmacology and signal transduction of the human
cannabinoid CB1 and CB2 receptors. Mol Pharmacol, v. 48, n. 3, p. 443–450, 1995.
FERREIRA, D. D. P. et al. Caffeine potentiates the release of GABA mediated by NMDA
receptor activation: Involvement of A1adenosine receptors. Neuroscience, v. 281, n. October,
p. 208–215, 2014.
GAONI, Y.; MECHOULAM, R. Isolation, Structure, and Partial Synthesis of an Active
Constituent of Hashish. Journal of the American Chemical Society, v. 86, n. 8, p. 1646–1647,
1964.
GILMAN, A. G. A protein binding assay for adenosine 3’:5’-cyclic monophosphate.
Proceedings of the National Academy of Sciences of the United States of America, v. 67, n. 1,
p. 305–12, 1970.
GLEASON, E.; WILSON, M. Development of synapses between chick retinal neurons in
dispersed culture. The Journal of Comparative Neurology, v. 287, n. 2, p. 213–224, 8 set.
1989.
GONZALEZ, B. et al. Cannabinoid agonists stimulate [3H]GABA release in the globus
pallidus of the rat when G(i) protein-receptor coupling is restricted: role of dopamine D2
receptors. The Journal of pharmacology and experimental therapeutics, v. 328, n. 3, p. 822–
828, 2009.
GOVINDPANI, K. et al. Towards a better understanding of GABAergic remodeling in
alzheimer’s disease. International Journal of Molecular Sciences, v. 18, n. 8, 2017.
GULYAS, A. I. et al. Segregation of two endocannabinoid-hydrolyzing enzymes into pre-
and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. European
Journal of Neuroscience, v. 20, n. 2, p. 441–458, 2004.
HÄRING, M. et al. Cannabinoid type-1 receptor signaling in central serotonergic neurons
regulates anxiety-like behavior and sociability. Frontiers in Behavioral Neuroscience, v. 9, n.
September, p. 1–13, 2015.
HARKANY, T; MACKIE, K.; DOHERTY, P. Wiring and firing neuronal networks:
endocannabinoids take center stage. Current Opinion in Neurobiology, v. 18, n. 3, p. 338–
345, jun. 2008.
HEAVNER, W.; PEVNY, L. Eye development and retinogenesis. Cold Spring Harbor
Perspectives in Biology, v. 4, n. 12, 2012.
HEIMANN, A. S. et al. Hemopressin is an inverse agonist of CB1 cannabinoid receptors.
Proceedings of the National Academy of Sciences of the United States of America, v. 104, n.
51, p. 20588–93, 2007.

36
HIRVONEN, J. et al. Reversible and regionally selective downregulation of brain
cannabinoid CB1 receptors in chronic daily cannabis smokers. Molecular psychiatry, v. 17, n.
6, p. 642–9, jun. 2012.
HOKOÇ, J. N. et al. Developmental immunoreactivity for GABA and GAD in the avian
retina: possible alternative pathway for GABA synthesis. Brain Research, v. 532, n. 1–2, p.
197–202, 1990.
HONÓRIO, K. M.; ARROIO, A.; DA SILVA, A. B. F. Aspectos terapêuticos de compostos
da planta Cannabis sativa. Quimica Nova, v. 29, n. 2, p. 318–325, 2006.
HOWLETT, A. C. et al. International Union of Pharmacology. XXVII. Classification of
cannabinoid receptors. Pharmacological reviews, v. 54, n. 2, p. 161–202, jun. 2002.
HUIZINK, A. C.; MULDER, E. J. H. Maternal smoking, drinking or cannabis use during
pregnancy and neurobehavioral and cognitive functioning in human offspring. Neuroscience
and Biobehavioral Reviews, v. 30, n. 1, p. 24–41, 2006.
HUTCHINGS, D. E. et al. Plasma concentrations of delta-9-tetrahydrocannabinol in dams
and fetuses following acute or multiple prenatal dosing in rats. Life Sciences, v. 44, n. 11, p.
697–701, 1989.
INDA, C. et al. CAMP-dependent cell differentiation triggered by activated CRHR1 in
hippocampal neuronal cells. Scientific Reports, v. 7, n. 1, p. 1–17, 2017.
JOHNSON, J. et al. Multiple gamma-Aminobutyric acid plasma membrane transporters
(GAT-1, GAT-2, GAT-3) in the rat retina. The Journal of comparative neurology, v. 375, n.
2, p. 212–24, 11 nov. 1996.
KATONA, I. et al. Presynaptically located CB1 cannabinoid receptors regulate GABA
release from axon terminals of specific hippocampal interneurons. The Journal of
neuroscience : the official journal of the Society for Neuroscience, v. 19, n. 11, p. 4544–4558,
1999.
KATONA, I.; FREUND, T. F. Multiple Functions of Endocannabinoid Signaling in the Brain.
Annual Review of Neuroscience, v. 35, p. 529–558, 2014.
KAUFMAN, D. L.; HOUSER, C. R.; TOBIN, A. J. Two Forms of the γ‐Aminobutyric Acid
Synthetic Enzyme Glutamate Decarboxylase Have Distinct Intraneuronal Distributions and
Cofactor Interactions. Journal of Neurochemistry, v. 56, n. 2, p. 720–723, 1991.
KENDALL, D. A.; YUDOWSKI, G. A. Cannabinoid Receptors in the Central Nervous
System: Their Signaling and Roles in Disease. Frontiers in Cellular Neuroscience, v. 10, n.
January, p. 1–10, 2017.
KO, J. Y. et al. Prevalence and patterns of marijuana use among pregnant and nonpregnant
women of reproductive age. American Journal of Obstetrics and Gynecology, v. 213, n. 2, p.
201.e1-201.e10, 2015.
KOLB, H. How the retina works. American Scientist 91:28–35, 2003.
KOLB, H. The Organization of the Retina and Visual System [Internet]: Neurotransmitters in
the Retina. 2005 Mai 1 [Updated 2009 Jun 1]. Disponível em:
<https://www.ncbi.nlm.nih.gov/books/NBK11546/> Acesso em: 22 fev. 2018.

37
KREITZER, A. C.; REGEHR, W. G. Retrograde signaling by endocannabinoids. Current
Opinion in Neurobiology, v. 12, n. 3, p. 324–330, 2002.
KUBRUSLY, R. C. C. et al. Neuro-glial cannabinoid receptors modulate signaling in the
embryonic avian retina. Neurochemistry International, v. 112, p. 27–37, 2018.
KUBRUSLY, R. C. C.; DE MELLO, M. C. F.; DE MELLO, F. G. Aspartate as a selective
NMDA receptor agonist in cultured cells from the avian retina. Neurochemistry International,
v. 32, n. 1, p. 47–52, 1998.
LANKFORD, K. L.; DE MELLO, F. G.; KLEIN, W. L. D1-type dopamine receptors inhibit
growth cone motility in cultured retina neurons: evidence that neurotransmitters act as
morphogenic growth regulators in the developing central nervous system. Proceedings of the
National Academy of Sciences, v. 85, n. 8, p. 2839–2843, 1988.
LAUCKNER, J. E. et al. GPR55 is a cannabinoid receptor that increases intracellular calcium
and inhibits M current. Proceedings of the National Academy of Sciences, v. 105, n. 7, p.
2699–2704, 2008.
LAVOIE, J.; MAZIADE, M.; HÉBERT, M. The brain through the retina: The flash
electroretinogram as a tool to investigate psychiatric disorders. Progress in Neuro-
Psychopharmacology and Biological Psychiatry, v. 48, p. 129–134, 3 jan. 2014.
LEONELLI, M. et al. Developmental expression of cannabinoid receptors in the chick
retinotectal system. Developmental Brain Research, v. 156, n. 2, p. 176–182, 2005.
LIU, Q. R. et al. Molecular characterization of four pharmacologically distinct gamma-
aminobutyric acid transporters in mouse brain [corrected]. The Journal of biological
chemistry, v. 268, n. 28, p. 2106–2112, 1993.
LOO, D. D. F. et al. Role of Cl-in electrogenic NA+-coupled cotransporters GAT1 and
SGLT1. Journal of Biological Chemistry, v. 275, n. 48, p. 37414–37422, 2000.
LOTURCO, J. J. et al. GABA and glutamate depolarize cortical progenitor cells and inhibit
DNA synthesis. Neuron, v. 15, n. 6, p. 1287–1298, 1995.
LOWRY, O. H. et al. Protein measurement with the Folin phenol reagent. The Journal of
biological chemistry, v. 193, n. 1, p. 265–275, 1951.
MACKIE, K. Cannabinoid receptors: Where they are and what they do. Journal of
Neuroendocrinology, v. 20, n. SUPPL. 1, p. 10–14, 2008.
MACKIE, K. From active ingredients to the discovery of the targets: The cannabinoid
receptors. Chemistry and Biodiversity, v. 4, n. 8, p. 1693–1706, 2007.
MADTES, P.; REDBURN, D. A. GABA as a trophic factor during development. Life
Sciences, v. 33, n. 10, p. 979–984, 1983.
MARC, R. E. et al. Amino acid signatures in the normal cat retina. Investigative
ophthalmology & visual science, v. 39, n. 9, p. 1685–93, ago. 1998.
MARICHAL-CANCINO, B. A. et al. Advances in the Physiology of GPR55 in the Central
Nervous System. Current Neuropharmacology, v. 15, n. 5, p. 771–778, 2017.
MATSUZAWA, H.; NIRENBERG, M. Receptor-mediated shifts in cGMP and cAMP levels
in neuroblastoma cells. Proceedings of the National Academy of Sciences of the United States
of America, v. 72, n. 9, p. 3472–3476, 1975.

38
MCPARTLAND, J. M.; GLASS, M.; PERTWEE, R. G. Meta-analysis of cannabinoid ligand
binding affinity and receptor distribution: Interspecies differences. British Journal of
Pharmacology, v. 152, n. 5, p. 583–593, 2007.
MECHOULAM, R. et al. Identification of an endogenous 2-monoglyceride, present in canine
gut, that binds to cannabinoid receptors. Biochemical Pharmacology, v. 50, n. 1, p. 83–90,
1995.
MECHOULAM, R.; PARKER, L. A. The Endocannabinoid System and the Brain. Annual
Review of Psychology, v. 64, n. 1, p. 21–47, 2013.
MEY, J; THANOS, S. Development of the visual system of the chick. I. Cell differentiation
and histogenesis. Brain research. Brain research reviews, v. 32, n. 2–3, p. 343–379, 2000.
PEPPE, A. et al. Does pattern electroretinogram spatial tuning alteration in Parkinson’s
disease depend on motor disturbances or retinal dopaminergic loss? Electroencephalography
and clinical neurophysiology, v. 106, n. 4, p. 374–82, abr. 1998.
PERDIKARIS, P. et al. Long lasting effects of chronic WIN55,212-2 treatment on
mesostriatal dopaminergic and cannabinoid systems in the rat brain. Neuropharmacology, v.
129, p. 1–15, 2018.
PEREZ-REYES, M; WALL, M E. Presence of Δ9-Tetrahydrocannabinol in Human Milk.
New England Journal of Medicine, v. 307, n. 13, p. 819–820, 23 set. 1982.
PERTWEE, R. G. Inverse agonism and neutral antagonism at cannabinoid CB1receptors. Life
Sciences, v. 76, n. 12, p. 1307–1324, 2005.
PIOMELLI, D. The molecular logic of endocannabinoid signalling. Nature Reviews
Neuroscience, v. 4, n. 11, p. 873–884, 2003.
POHL-GUIMARÃES, F. et al. Ethanol increases GABA release in the embryonic avian
retina. International Journal of Developmental Neuroscience, v. 28, n. 2, p. 189–194, 2010.
PRADA, C. et al. Spatial and Temporal Patterns of Neurogenesis in the Chick Retina. The
European journal of neuroscience, v. 3, n. 11, p. 1187, 1991.
REIS, R. A. M. et al. Dopaminergic signaling in the developing retina. Brain Research
Reviews, v. 54, n. 1, p. 181–188, 2007.
RIBEIRO, H. L. et al. Efeitos do consumo de cannabis na gravidez e no período pós parto.
Revista Debates em psiquiatria, v. Mar/Abr, p. 16–24, 2016.
RICHARDSON, K. A.; HESTER, A. K.; MCLEMORE, G. L. Prenatal cannabis exposure -
The “first hit” to the endocannabinoid system. Neurotoxicology and Teratology, v. 58, p. 5–
14, 2016.
ROMERO, J. et al. Time-course of the cannabinoid receptor down-regulation in the adult rat
brain caused by repeated exposure to delta9-tetrahydrocannabinol. Synapse (New York, N.Y.),
v. 30, n. 3, p. 298–308, 1998.
ROMERO, J. et al. Unilateral 6-hydroxydopamine lesions of nigrostriatal dopaminergic
neurons increased CB1 receptor mRNA levels in the caudate-putamen. Life sciences, v. 66, n.
6, p. 485–94, 2000.
RYBERG, E. et al. The orphan receptor GPR55 is a novel cannabinoid receptor. British
Journal of Pharmacology, v. 152, n. 7, p. 1092–1101, 2007.

39
SAITO, V. M.; WOTJAK, C. T.; MOREIRA, F. A. Exploração farmacológica do sistema
endocanabinoide: novas perspectivas para o tratamento de transtornos de ansiedade e
depressão? Revista Brasileira de Psiquiatria, v. 32, n. suppl 1, p. 57–514, maio 2010.
SAMPAIO, L. et al. Cannabinoid Receptor Type 1 Expression in the Developing Avian
Retina: Morphological and Functional Correlation With the Dopaminergic System. Frontiers
in cellular neuroscience, v. 12, n. march, p. 58, 2018.
SÁNCHEZ-BLÁZQUEZ, P.; RODRÍGUEZ-MUÑOZ, M.; GARZÓN, J. The cannabinoid
receptor 1 associates with NMDA receptors to produce glutamatergic hypofunction:
Implications in psychosis and schizophrenia. Frontiers in Pharmacology, v. 4 JAN, n.
January, p. 1–10, 2014.
SAVINAINEN, J. R. et al. Despite substantial degradation, 2-arachidonoylglycerol is a potent
full efficacy agonist mediating CB(1) receptor-dependent G-protein activation in rat
cerebellar membranes. British journal of pharmacology, v. 134, n. 3, p. 664–72, 2001.
SAVINAINEN, J. R. et al. An optimized approach to study endocannabinoid signaling:
evidence against constitutive activity of rat brain adenosine A1 and cannabinoid CB1
receptors. British journal of pharmacology, v. 140, n. 8, p. 1451–1459, 2003.
SCHWARTZ, E. A. Calcium-independent release of GABA from isolated horizontal cells of
the toad retina. The Journal of physiology, v. 323, p. 211–27, fev. 1982.
SCHWITZER, T. et al. The Endocannabinoid System in the Retina: From Physiology to
Practical and Therapeutic Applications. Neural Plasticity, v. 2016, p. 1-10, 2016.
SCIMEMI, A. Structure, function, and plasticity of GABA transporters. Frontiers in Cellular
Neuroscience, v. 8, n. June, p. 1–14, 2014.
SILVA, J. P.; YONAMINE, M. Bioanalytical and methodological challenges in the
evaluation of fetal Cannabis exposure. Bioanalysis, v. 10, n. 10, p. 713-716, 2018.
SIM-SELLEY, L. J. Regulation of cannabinoid CB1 receptors in the central nervous system
by chronic cannabinoids. Critical reviews in neurobiology, v. 15, n. 2, p. 91–119, 2003.
STELLA, N.; SCHWEITZER, P.; PLOMELLI, D. A. second endogenous’ cannabinoid that
modulates long-term potentiation. Nature, v. 388, n. 6644, p. 773–778, 1997.
STRAIKER, A. et al. Cannabinoid CB1 receptors and ligands in vertebrate retina:
localization and function of an endogenous signaling system. Proceedings of the National
Academy of Sciences of the United States of America, v. 96, n. 25, p. 14565–70, 1999.
SUGIURA, T. et al. Biosynthesis and degradation of anandamide and 2-arachidonoylglycerol
and their possible physiological significance. Prostaglandins Leukotrienes and Essential
Fatty Acids, v. 66, n. 2–3, p. 173–192, 2002.
SUNG, C. H.; CHUANG, J. Z. The cell biology of vision. Journal of Cell Biology, v. 190, n.
6, p. 953–963, 2010.
SVÍŽENSKÁ, I.; DUBOVÝ, P.; ŠULCOVÁ, A. Cannabinoid receptors 1 and 2 (CB1 and
CB2), their distribution, ligands and functional involvement in nervous system structures - A
short review. Pharmacology Biochemistry and Behavior, v. 90, n. 4, p. 501–511, 2008.
THE RETINA: RETINAL LAYERS. Disponível em :<
http://www.netterimages.com/image/29421.htm> Acesso em: 11 jun. 2018.

40
TOMINAGA, M. et al. The cloned capsaicin receptor integrates multiple pain-producing
stimuli. Neuron, v. 21, n. 3, p. 531–543, 1998.
TÓTH, A.; BLUMBERG, P. M.; BOCZÁN, J. Chapter 15 Anandamide and the Vanilloid
Receptor (TRPV1). Vitamins and Hormones. v. 81, n. C, p. 389-419 , 2009.
UNITED NATIONS OFFICE OF DRUGS AND CRIME. Market Analysis of Plant-Based
Drugs. [S.l: s.n.], 2017.
VENDEROVA, K.; BROWN, T. M.; BROTCHIE, J. M. Differential effects of
endocannabinoids on [3H]-GABA uptake in the rat globus pallidus. Experimental Neurology,
v. 194, n. 1, p. 284–287, 2005.
VENTURA, A. L. et al. Sistema colinérgico: Revisitando receptores, regulação e a relação
com a doença de Alzheimer, esquizofrenia, epilepsia e tabagismo. Revista de Psiquiatria
Clinica, v. 37, n. 2, p. 74–80, 2010.
WALLMICHRATH, I.; SZABO, B. Cannabinoids inhibit striatonigral GABAergic
neurotransmission in the mouse. Neuroscience, v. 113, n. 3, p. 671–682, 2002.
WU, C. S.; JEW, C. P.; LU, H.C. Lasting impacts of prenatal cannabis exposure and the role
of endogenous cannabinoids in the developing brain. Future Neurology, v. 6, n. 4, p. 459–
480, 2011.
YAMASHITA, M.; FUKUDA, Y. Calcium channels and GABA receptors in the early
embryonic chick retina. Journal of Neurobiology, v. 24, n. 12, p. 1600–1614, 1993.
YAZULLA, S.; KLEINSCHMIDT, J. Carrier-mediated release of GABA from retinal
horizontal cells. Brain Research, v. 263, n. 1, p. 63–75, 1983.







