
UNIVERSIDADE CATÓLICA DOM BOSCO PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
INGRID BATISTA PINTO
Caracterização dos genes matK e rbcL e da variabilidade genética entre os acessos de Araruta (Maranta arundinacea L.).
CAMPO GRANDE
2015

UNIVERSIDADE CATÓLICA DOM BOSCO PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
Caracterização dos genes matK e rbcL e da variabilidade genética entre os acessos de Araruta (Maranta arundinacea L.).
Autora: Ingrid Batista Pinto Orientadora: Carina Elisei de Oliveira
Coorientadora: Marney Pascoli Cereda
. "Dissertação apresentada, como parte das exigências para obtenção do título de MESTRE EM BIOTECNOLOGIA, no Programa de Pós-Graduação em Biotecnologia da Universidade Católica Dom Bosco - Área de concentração: Biotecnologia Aplicada à Agropecuária "
Campo Grande, Mato Grasso do Sul, Fevereiro – 2015.

ii

iii

iv
EPÍGRAFE
Os passos da Grande Obra correspondem aos da
criação do Mundo, demonstrando uma analogia entre
microcosmo e macrocosmo. Mas isto é apenas uma
analogia, porque, segundo os sábios antigos, o
'Magisterius' não é criação, e, sim, um processo gerador
realizado pelo Artista – que é conduzido pela Natureza –
e está apenas acelerando seu próprio trabalho de
purificação.
Christian Balister.

v
DEDICATÓRIA
A minha família em especial ao meu
Pai, Pedro Batista Pinto pela
dedicação, carinho e por sempre ter
me apoiado na busca e realização
dos meus sonhos.

vi
AGRADECIMENTOS
À Deus pela concretização deste trabalho, por ter me iluminado e por me dar força
interior para superar as dificuldades, me suprir em todas as minhas necessidades.
Às minhas orientadoras Professoras Carina Elisei de Oliveira e Marney Pascoli
Cereda por me apresentar o caminho da ciência e principalmente, acreditarem em
mim.
A Dra. Josimara Nolasco Rondon, pelo incentivo e dicas valiosas.
A Dra. Lucimara Chiari pelos ensinamentos.
A Dra. Renata Dias, pela contribuição significativa nesse trabalho.
À minha família, ao meu pai Pedro Batista Pinto, meu filho Pedro Fernando B.
Hormung e a minha Irmã Irina Batista Pinto, pelo apoio e carinho prestados mesmo
sem saberem exatamente, o que eu faço.
Ao meu namorado pela imensa paciência, colaboração, compreensão, amor,
carinho, por todo incentivo e pelo companheirismo incondicional.
Aos meus amigos de laboratório Guilherme A. Abrantes Souza, que compartilhou
ensinamento, ajudando-me em vários momentos, e a Poliene Costa, pelas
conversas e pela gratificante “parceria”.
A técnica do laboratório Maria Helena pela colaboração e ajuda durante a execução
deste trabalho.
A Capes/ CNPQ e a UCDB pela bolsa concedida.
Enfim a todos que contribuíram de forma direta ou indireta para resolução deste
trabalho o meu MUITO OBRIGADA.

vii
BIOGRAFIA DO AUTOR
Nascida em Campo Grande - MS em 28 de maio de 1984, Ingrid Batista Pinto, filha
de Pedro Batista Pinto e Graça Maria Colavite Pinto. Teve uma infância tranquila e
feliz.
Em 2003 entrou para o curso de Nutrição da Universidade Católica Dom Bosco,
onde cursou apenas dois anos, terminando a graduação na Universidade
Anhanguera- Uniderp na mesma área. Fez estágios em nutrição clínica no Hospital
Regional e na Unidade de Alimentação e Nutrição da Santa Casa de Campo
Grande.
Trabalhou em empresas de Alimentação e em Hospitais como nutricionista.
Em 2012 visando aperfeiçoar seus conhecimentos, decidiu se especializar iniciando
o Programa de Pós Graduação Mestrado em Biotecnologia da UCDB, onde através
da ciência encontrou seu espaço e realização.

viii
SUMÁRIO
1 INTRODUÇÃO ......................................................................................................... 1
1.1 A família Marantaceae ....................................................................................... 1
1.2 Maranta arundinacea L. ..................................................................................... 2
1.3 Importância Econômica de Maranta arundinacea . ............................................ 5
1.4 Marcadores Moleculares ................................................................................. 10
1.5 Marcador Molecular RAPD .............................................................................. 12
1.6 PCR Convencional ........................................................................................... 15
1.7 DNA barcoding ................................................................................................. 16
1.7.1 MatK ............................................................................................................ 20
1.7.2 rbcL .............................................................................................................. 21
2 OBJETIVOS ........................................................................................................... 21
REFERÊNCIAS ............................................................................................................ 22
CAPÍTULO I:. Caracterização dos genes matK e rbcL e da variabilidade
genética entre os acessos de Araruta (Maranta arundinacea L.)........................... 31
Introdução .................................................................................................................... 33
Material e Métodos ...................................................................................................... 34
Amostras .................................................................................................................... 34
Obtenção das amostras ............................................................................................. 34
Avaliação do perfil genético ....................................................................................... 35
Extração do material genético .................................................................................... 35
Verificação da quantidade e qualidade do DNA ........................................................ 36
Desenho dos Primes (iniciadores) dos genes matK e rbcL ....................................... 37
PCR convencional ..................................................................................................... 37
Análise das sequências dos genes matK e rbcL........................................................ 38
Marcadores RAPD ..................................................................................................... 38
Análise dos dados obtidos através de marcadores RAPD ........................................ 39
Resultados e Discussão ................................................................................................ 40
Variabilidade genética ............................................................................................... 44
Conclusão ..................................................................................................................... 46
Referências Bibliográficas ............................................................................................. 47
Apêndices ..................................................................................................................... 51

v
LISTA DE TABELAS
Tabela 1. Composição nutricional da araruta crua por 100 gramas ................................ 9
CAPÍTULO I
Tabela 1. Genes utilizados, primers e os números de acesso no GenBank ................. 37
Tabela 2. Lista dos primers utilizados com suas respectivas sequências de bases ..... 37
Tabela 3. Dados de quantificação via espectrofotometria das extrações de material
genético foliar de quatro acessos de araruta. A absorbância (Abs) foi medida no
comprimento de onda de 260 a 280 nanômetros .......................................................... 41
Tabela 4. Lista dos primers e suas respectivas sequências, número de fragmentos
amplificados e número de fragmentos polimórficos obtidos dos quatro acessos .......... 44
APÊNDICE
Tabela 1. Análise do acesso de Araruta Comum comparadas com sequências
depositadas no BOLD Systems de rbcL ........................................................................ 51
Tabela 2. Análise das seqências SC, Guadalupe e Seta comparadas com
sequências depositadas no BOLD Systems de rbcL .................................................... 51
Tabela 3. Análise das sequências Comum, SC, Guadalupe e Seta comparadas com
sequências depositadas no BOLD Systems de matK ................................................... 51

vi
LISTA DE FIGURAS
Figura 1. Aspecto geral da Araruta (Maranta arundinacea L.) ........................................ix
Figura 2. Aspecto geral do rizoma da Araruta ................................................................. x
Figura 3. Principais áreas de ocorrência de Maranta Arundinacea L ............................ 37
Figura 4. Esquema da reação de Polimorfismos de DNA amplifocados ao acaso
(RAPD) ........................................................................................................................ 137
Figura 5. Regiões mais utilizadas como DNA Barcoding em plantas ............................ 17
Figura 6. Esquema do genoma do cloroplasto .............................................................. 19
CAPÍTULO I
Figura 1. Imagens dos quatro acessos de M. arundinacea .......................................... 37
Figura 2. Dendograma obtido a partir da análise de Neighbor-joining (NJ), dos
acessos Comum, SC, Guadalupe e Seta, utilizando Siphonochilus decorus como
outgroup. Os valores encontrados nos ramos indicam os valores de boostrap (>50)
baseado em 1.000 interações ....................................................................................... 44
Figura 3. Dendograma de similaridade genética, obtido a partir dos produtos de
amplificação por RAPD para quatro genótipos de Araruta ............................................ 45
APÊNDICE
Figura 1. Resultado do sequenciamento de gene rbcL dos quatro acessos de M.
arundinacea .................................................................................................................. 52
Figura 2. Resultado do sequenciamento de gene matK dos quatro acessos de M.
arundinacea .................................................................................................................. 54
Figura 3. Padrão eletroforético obtido com primers OPF5 e OPAF5. PM: Peso
Molecula 1kb plus. CO: Araruta Comum. SC: Araruta SC. G: Araruta Guadalupe e
ST: Araruta Seta, C- controle negativo ......................................................................... 56
Figura 4. Padrão eletroforético obtido com primers OPA1 e OPA2. PM: Peso
Molecula 1kb plus. CO: Araruta Comum. SC: Araruta SC. G: Araruta Guadalupe e
ST: Araruta Seta, C- controle negativo ..................................................................... 3757
Figura 5. Padrão eletroforético obtido com primers OPA17. PM: Peso Molecula 1kb
plus. CO: Araruta Comum. SC: Araruta SC. G: Araruta Guadalupe e ST: Araruta
Seta, C- controle negativo ............................................................................................ 58

vii
LISTA DE ABREVIATURAS E SÍMBOLOS
LISTA DE ABREVIATURAS
atpF - Gene que codifica as subunidade CFO I da ATP sintase
atpH - Gene que codifica a subunidade CFO III da ATP sintase
BLAST -Basic Local Alignment Search Tool
CBOL - Consortium for the Barcode of Life
CIA - clorofórmio: álcool isoamilico
COI - Citocromo oxidase I
dNTP – Desoxirribonucleotideo trifosfato
ddNTP – Di Desoxirribonucleotideo trifosfato
DNA – Ácido desoxirribonucleico
EDTA - Ácido etilenodiaminotetracético
et al - E outros.
IR (a ou b) - Large Inverted Reapets
ITSs - Espacadores internos transcritos
kb - Kilo pares de bases
LSC - Large Single Copy Region
matK – Gene que codifica a proteina maturase K
mtDNA - DNA mitochondrial
ng - nanograma
pb - Pares de bases
PCR - Polymerase Chain Reaction
PlantGDB - Plant Genomic Database
psbA – Gene que codifica a proteina D1 do fotossistema II do cloroplasto
psbI – Gene que codifica o polipeptideo I de massa molecular baixa do sistema de
fotossistema II do cloroplasto
psbK - Gene que codifica o polipeptideo K de massa molecular baixa, do sistema de
fotossistema II
RAPD - Random Amplification of Polymorphic DNA
rbcL -Gene que codifica a grande subunidade da ribulose-1,5-bifosfato carboxilase/
oxigenase (RuBisCo)

viii
rDNA - DNA ribossomal
RFLPs - Restriction Fragment Lenght Polymorfisms
RNA – Acido ribonucleico
RNase – Enzima que degrada especificamente o RNA
rpoB - Gene que codifica a subunidade β da RNA polimerase
rpoC1 - Gene que codifica a subunidade β’ da RNA polimerase
sp - Espécie
spp - Espécies
SSC - Small Single Copy Region
Taq – Thermus aquaticus, bactéria termófila da qual se obtém uma das DNAs
polimerases termoestáveis utilizadas em PCR.
tRNA – Ácido ribonucleico transportador
trnH - Gene de tRNA
trnK - Gene de Trna
LISTA DE SÍMBOLOS
°C - Graus Celcius
% - Percentagem
h - Hora(s)
G - grama
μL - Microlitro(s)
μM - Micromolar
mg - Miligrama(s)
mL - Mililitro(s)
min - Minuto(s)
s - Segundo(s)

ix
RESUMO
PINTO, I. B. Caracterização dos Genes matK e rbcL e da
Variabilidade Genética entre os Acessos de Araruta (Maranta
arundinacea L.). Campo Grande, 2015. (Dissertação Mestrado em
Biotecnologia) – Universidade Católica Dom Bosco, Campo Grande,
2015.
Maranta arundinacea é uma planta nativa da América do Sul, apresenta potencial
econômico como uma matéria prima não convencional para amido, através de seus
caules subterrâneos denominados rizomas. Devido à falta de informações sobre o
número de espécies, o risco de extinção e o crescente interesse no uso e na
comercialização de seu amido fazem da M.arundinacea uma espécie ideal para
estudos moleculares. Assim, o objetivo deste trabalho foi a identificação de quatro
acessos de M. arundinacea denominados de Comum, (CeTeAgro); SC, Guadalupe
e Seta (EMBRAPA-CENERAGEM) através dos genes cloroplastidiais matK e o rbcL,
e avaliar a similaridade genética, através de marcadores moleculares RAPD. Para
análise de RAPD utilizou-se 25 primers, destes, o primer OPA 02 foi selecionado por
apresentar maior número de bandas polimórficas. Através do coeficiente de
similaridade de Jaccard, juntamente com o teste de UPGMA, foi possível
estabelecer dois grupos distintos, Grupo I (Comum) e Grupo II (SC, Guadalupe e
Seta). A identificação dos acessos por meio dos genes cloroplastidiais matK e o
rbcL, foram submetidas por busca BLAST (GenBank) e BOLD Systems, resultando
em 99,66 a 100% de identidade para o gene matK e rbcL, respectivamente, dos
quatro acessos estudados, comparados com M. arudinacea. A construção da árvore
filogenética, utilizando o gene rbcL, realizada por Neighbor-Joining, observou-se
após a separação do grupo externo, um grande grupo de M. arundinacea contendo
os acessos SC, Guadalupe, Seta e o acesso do banco de doados M. arundinacea
(3845927560), não apresentado diferenças entre eles. Outro grupo interno foi
constituído por Comum e o acesso do banco de dados M. arundinacea
(JQ592613.1), evidenciando que o Comum se diferenciou dos demais acessos SC,
Guadalupe e Seta, entretanto indicando que todos os acessos são da espécie M.
arundinacea.
Palavras-chave: DNA Barcoding, Marantaceae, RAPD.

x
ABSTRACT
PINTO, I. B. Characterization of Genes matK and rbcL and Genetics
variability between Arrowroot accesses (Maranta arundinacea L.).
Campo Grande, 2015. (Master Master in Biotechnology) - Dom Bosco
Catholic University, Campo Grande, 2015.
Maranta arundinacea is a plant native to South America, has economic potential as a
non-conventional raw material for starch, through its underground stems called
rhizomes. Due to lack of information on the number of species at risk of extinction
and the growing interest in the use and marketing of its starch M.arundinacea make
an ideal species for molecular studies. The objective of this study was the
identification of four hits M. arundinacea Joint denominated, (CeTeAgro); SC,
Guadeloupe and Arrow (EMBRAPA-CENERAGEM) through the chloroplast genes
matK and rbcL, and assess the genetic similarity using RAPD molecular markers.
For RAPD analysis was performed using 25 primers, of these, the primer OPA 02
was selected due to its higher number of polymorphic bands. Through the Jaccard
similarity coefficient, together with the UPGMA test, it was possible to establish two
groups, Group I (Common) and Group II (SC, Guadeloupe and Arrow). The
identification of the accesses through the plastid rbcL gene and matK were
submitted by BLAST search (GenBank) and BOLD Systems, resulting in 99.66 to
100% Identity to the rbcL gene and matK, respectively, of the four accesses studied,
M. arudinacea compared. The construction of the phylogenetic tree using the rbcL
gene carried by neighbor-joining, was observed after separation of the outer group, a
large group containing M. arundinacea SC accesses, Guadeloupe arrow and given
access database M . arundinacea (3845927560), did not present differences
between them. Another internal group consisted of Common and the database
access M. arundinacea (JQ592613.1), showing that the Joint different from the other
SC access, Guadeloupe and arrow, however, indicating that all accesses are the
species M. arundinacea .
Keywords: DNA Barcoding, Marantaceae, RAPD.

1
1 INTRODUÇÃO
1.1 A Família Marantaceae
Em 1890 O. G. Petersen descreveu a família Marantaceae, sendo que o
nome da família foi uma homenagem ao botânico italiano Bartolomeo Maranta, que
viveu entre 1500 e 1571.
A família Marantaceae é um grande grupo de plantas compostas por 530
espécies e 31 gêneros (KENNEDY, 1977; ANDERSSON, 1986; KENNEDY,1997,
2000). Das 530 espécies conhecidas, cerca de 80% ocorrem na América Tropical,
9% na África e 11% na Ásia (HAMMEL, 1986). Esta família pertencente à ordem
Zingiberales (NEVES et al., 2005). As espécies de Marantaceae apresentam
distribuição pantropical e são típicas de florestas úmidas tropicais, crescendo
principalmente a margens de rios e clareiras, podendo também ser encontradas em
locais com vegetação aberta em áreas encharcadas e em depressões nas margens
de estradas (COSTA et al., 2008).
De acordo com Souza e Lorenzi (2005), no Brasil a família Marantaceae,
possui 150 espécies, das quais o gênero mais representativo no país é Calathea
seguido de Maranta. Dez gêneros são encontrados na Amazônia brasileira:
Calathea, Ischnosiphon, Monotagma, Hylaeanthe, Thalia, Maranta, Koernickanthe,
Monophyllanthe, Saranthe e Myrosma (COSTA et al., 2008).
As plantas da família Marantaceae são facilmente identificadas, pela
combinação de folhas com nervuras paralelas e presença de um engrossamento do
pecíolo na junção entre este e a lâmina foliar denominado pulvino, possuem rizomas
subterrâneos, com exceção de algumas espécies que o rizoma pode estar
parcialmente fora da terra (rizomas aéreos) e as suas flores nascem em dupla,
formando uma estrutura chamada de inflorescência (COSTA et al., 2008).
As marantáceas possuem sementes com apenas um cotilédone
(monocotiledôneas), raiz fasciculada e folhas com nervuras paralelas. As
monocotiledôneas representam 22% de todas as Angiospermas (COSTA et al.,
2008).
Os principais representantes da família Marantacea apresentam grande
importância como plantas ornamentais, sendo estas Calathea, Ctenanthe, Maranta
e Stromanthe, e fonte de fibras como a espécie Ischnosiphon gracilis, conhecida

2
como “arumã”, que é utilizada para produção de cestas e criação de utensílios
domésticos de índios e caboclos (COSTA et al., 2008).
As folhas das espécies de Calathea fragilis e Calathea lutea são muito
utilizadas para embalar peixes, bolos de mandioca e milho, e outras espécies são
utilizados na alimentação humana tais como Calathea allouia e M. arundinacea L.
(ARNS, 1997; COSTA et. al., 2008). A espécie Calathea allouia, conhecida como
“ariá”, é cultivada por índios e caboclos da Amazônia, para a obtenção de seus
tubérculos, os quais são consumidos após a cocção. A espécie considerada mais
importante economicamente é M. arundinacea, que tem como principal atrativo o
rizoma que consiste da parte comestível que podem também ser consumido após o
cozimento ou usado para extração do amido (COSTA et al., 2008).
1.2 Maranta arundinacea L.
A araruta é uma planta herbácea cujo nome científico é M. arundinacea L.
(Figura 1), pertencente à família Marantaceae e à ordem Zingiberales. A araruta é
uma planta rizomatosa, que produz rizomas em forma de fuso, coberto por
escamas, os rizomas podem chegar de 10 a 25 cm de comprimento, em algumas
ocasiões dependendo do solo até 30 cm (Figura 2). Nas condições tropicais
caracteriza-se pelo pouco ou nenhum florescimento (PIO CORRÊA, 1984).
Figura 1. Aspecto geral da Araruta (Maranta arundinacea L). Fonte:<http://www.agrosoft.org.br/agropag/211821.htm#.VFvS0TTF_Qw>. Acesso em: 03 de nov. 2014.

3
Figura 2. Aspecto geral do Rizoma da Araruta. Fonte: <http://hortas.info/como-plantar-araruta>. Acesso em: 03 de nov. 2014.
Vários autores discorrem sobre a possível origem e áreas da ocorrência
desta planta, alguns a consideram nativa das Antilhas, México e de outros países
da América Central (BENTLEY e TRIMEN 1880). Segundo León (1987) a planta é
nativa da América do Sul e das Antilhas. Peckolt e Peckolt (1893) afirmam que são
nativas do Brasil. Erdman e Erdman (1984) citam que as espécies crescem na
América do Sul, Sudeste da Ásia, Caribe, Filipinas e Índia (Figura 3).
Há indícios do cultivo da araruta pelos índios há mais de 7000 anos atrás. Os
vegetais mais cultivados nas roças dos caboclos e de índios amazônicos nas terras
baixas da América do Sul eram a mandioca (Manihot esculenta), batata-doce
(Ipomoea batatas) taioba ou taiá (Xanthosoma sp.), ariá (Maranta lutea), araruta
(Maranta arundinacea), inhame (Dioscorea alata), cupá (Cissus gongylodes) e
amendoim (Arachis sp) (MARTINS, 2005).
Os índios Aruak, que habitavam desde o Amazonas até a região do Caribe,
cultivavam a araruta e as chamavam de planta "aruaque aruá-aru", significando
"refeição das refeições", pelo fato de considerarem especiais as refeições
preparadas com o amido que destaca-se pela sua digestibilidade (NEVES et al.,
2005).

4
Os índios extraiam o amido e utilizavam para diversas aplicações como
engrossar sopas, tratar diarreia (especialmente em crianças), fortificar parturientes e
purificar o sangue. As substâncias ácidas contidas no macerado fresco dos rizomas
também eram utilizadas como compressas contra feridas provocadas por flechas ou
como antídoto nas picadas de insetos e outros animais peçonhentos (SILVA e
MONTEIRO, 1968).
As indicações da araruta na medicina tradicional são relatadas para
convalescença, dispepsia, irritação do canal digestivo, acidez estomacal,
machucados, fortificação de crianças e idosos, queimaduras de sol, picadas de
cobras e de mosquitos, para os quais são utilizados os rizomas (BERNAL e
CORREA, 1994; DELGADO e SIFUENTES, 1995; MILLIKEN e ALBERT, 1997;
PECKOLT e PECKOLT; 1893; REVILLA, 2002).
Figura 3. Principais áreas de ocorrência de Maranta arundinacea. Fonte: <http://www.boldsystems.org/index.php/Taxbrowser_Taxonpage?taxon=maranta+arundinacea&searchMenu=taxonomy&query=maranta+arundinacea>. Acesso em: 03 de nov. 2014.
A araruta é uma planta de fácil cultivo. No Brasil, adaptou-se pelas
características climáticas de chuvas bem distribuídas e temperatura média mensal
maior que 22ºC, em geral seu cultivo encontra boas condições em clima temperado
e ao nível do mar (MONTEIRO e PERESSIN, 2002).
Em relação ao solo, a araruta prefere solos arenosos e profundos os quais
são ideais por proporcionar um bom crescimento dos rizomas. O solo precisa ser
arado com até 20 cm de profundidade e com bom teor de matéria orgânica
(COELHO et al., 2005).

5
Em regiões tropicais e equatoriais, o cultivo da araruta pode ser realizado o
ano inteiro, tendo como exigência para o seu desenvolvimento presença de
umidade. Entretanto em regiões sub-tropicais ou tropicais de altitude, os meses
mais indicados são (setembro-outubro a março-abril), permanecendo a cultura em
dormência durante o período frio e/ou seco do ano (BRASIL, 2010). Apresenta
resistência a pragas e doenças, sendo que os nematóides do gênero Meloidogyne
podem causar danos aos rizomas (BRASIL, 2010).
A colheita da araruta é realizada entre 9 e 10 meses após o plantio, quando
as plantas apresentam as folhas amareladas e secas, tombadas sobre o solo. Os
tratos culturais consistem de capinas e amontoas, que podem ser manuais ou
mecanizadas. (MONTEIRO e PERESSIN, 2002, BRASIL, 2010). Após a colheita,
uma porção de rizoma é destinada ao novo plantio e a outra para extração do amido
(MARTINS, 1943).
Para a produção do amido os rizomas são processados na seguinte ordem
descascamento e lavagem, desintegração (liquidificador industrial), separação
(peneira 60 tyler), resultando em leite de amido passando pela purificação (peneira
200 tyler) novamente, pré secagem (filtro a vácuo) e secagem (flash -dryer 70oC), o
resultado é um pó fino, branco e inodoro (LEONEL, 2007).
1.3 Importância Econômica de Maranta arundinacea
Os amidos e seus derivados são importantes fontes de matéria prima, pois
são aplicados em diversas áreas de interesse econômico, tais como fabricação de
papéis, adesivos, embalagens biodegradáveis, indústria têxtil, indústria
farmacêutica, na produção de cola e principalmente na indústria alimentícia. Suas
características em alimentos permitem alterar ou controlar a textura, aparência,
umidade e consistência (ZHAO e WHISTLER, 1994 apud MEDINA, 2008).
Os amidos produzidos a partir de grãos de cereais, leguminosas e tuberosas
são importantes comercialmente, pois são muito utilizados nas indústrias,
principalmente no setor alimentício, tendo como principais fontes de matéria prima
milho, batata, trigo e mandioca (CEREDA et al., 2003). A produção mundial de
amido é de aproximadamente 48,5 milhões de toneladas de amido, sendo os EUA
responsável pela maior produção de amido de milho (24,6 milhões de toneladas) a
União Europeia a maior fabricante de amido de batata (1,8 milhão de toneladas) e

6
de trigo (2,8 milhões de toneladas), outros amidos produzidos mundialmente,
principalmente o da mandioca, correspondem a 2,5 milhões de toneladas (GUILBOT
e MERCIER, 1985; FRANCO et al., 2002).
Países da Ásia, em especial a China, produzem amidos alternativos com
diferentes propriedades, tais como arroz (Oryza sativa), araruta (Maranta
arundinacea), biri (Canna edulis), inhame (Dioscorea alata), feijão mugo (Vigna
radiata) dentre outros (CEREDA et al., 2003).
Os grânulos de amido apresentam forma e tamanhos diversos de acordo com
o grupo de planta pertencente (cereais, raízes, tubérculos e leguminosas),
apresentando também características e propriedades funcionais diferentes, inclusive
entre plantas da mesma espécie (CEREDA et al., 2002).
Segundo Madineni et al., (2012) o amido da araruta contêm grânulos que são
de pequeno, médio e grande (2,92-6,42 µm) com tamanho médio de 4,84 µm, e L /
D de 1,20 com arredondamento de 0,73. O amido tem 24,8% de teor de amilose
com uma temperatura de gelatinização de 74.8 oC.
Possui temperatura relativamente alta de gelatinização e resistência ao
cisalhamento mecânico. Portanto, apresenta textura satisfatória para alimentos
industrializados em alta temperatura ou como agente estabilizador para produtos
esterilizados tais como alimentos infantis e produtos processados em ultra-alta
temperatura (UHT) (MADINENI et al., 2012).
Estudos realizados com M. arundinacea, relatam diversas utilidades para
essa planta.
Os efeitos imunoestimulantes de extratos do rizoma de araruta, foram
analisados in vitro utilizando técnicas de cultura de células animais, e in vivo,
usando ratinhos BALB / c. Os resultados indicaram que o extrato do rizoma de
araruta estimularam a produção de IgM por células HB4C5 e imunoglobulina (IgG,
IgA e IgM) in vitro. Além disso, a análise in vivo indicou que a dieta contendo
extratos de araruta aumentou os níveis séricos de IgG, IgA e IgM em ratinhos,
revelando os efeitos imunoestimuladores in vivo bem como in vitro ( KUMALASARI,
et al., 2012).
Estudo realizado em 1984 avaliou o potencial de produção de álcool
combustível a partir de biomassa de araruta suplementado com levedura. O resíduo
grosso demonstrou qualidades adequadas para produção de papel resistente à

7
ruptura e dobramento. Além de utilização da araruta como alimentos, combustível e
recursos de fibras, ele também pode ajudar a reduzir a poluição ambiental resultante
da descarga direta do subproduto metanol não utilizado (ERDMAN e ERDMAN,
1984).
A produção de amidos não convencionais visa principalmente à obtenção de
produtos diferenciados, a serem comercializados em mercado específico
possibilitando a ascensão dos preços e a diminuição da competição entre empresas
produtoras de fécula e farinha de mandioca, pela mesma matéria prima, além de
proporcionar alimentos livres de glúten, que os tornam recomendáveis para pessoas
que apresentam intolerância alimentar a esta proteína, evitando assim o consumo
de qualquer produto contendo trigo, aveia, centeio, cevada e seus derivados.
(CEREDA et al., 2002, CEREDA et al., 2003).
Na América latina a produção de amidos alternativos, tais como biri é feita
pela Colômbia. No Brasil a produção de amido da araruta, foi substituída pela
mandioca por ser mais produtiva, entretanto inúmeras espécies tais como, açafrão,
inhame, batata doce, taioba, gengibre, araruta entre outras, são matérias primas
com potencial para a produção de amido (CEREDA et al., 2003).
Segundo Cereda (2002), a maioria dos amidos não convencionais segue a
mesma linha de produção da mandioca, sendo que no Brasil testes feitos em
fecularias, com batata doce e araruta apresentaram resultados satisfatórios tanto
em extração quanto em qualidade.
O preço do amido da araruta no mercado varia entre R$ 15,00 a R$ 20,00
reais o quilo e no mercado internacional chega a alcançar preços ainda mais
elevados (450 gramas custam cerca de US$ 22,19) (EBDA, 2013). Cerca de 50
plantas bastam para consumo próprio, rendendo de 5 a 10 quilos de fécula (amido
ou polvilho) ou de 15 a 20 quilos de farinha. Para produção comercial, ao menos
500 metros quadrados são necessários para o plantio (GLOBO RURAL, 2013). O
amido da araruta, retirado do rizoma é o principal recurso explorado nessa planta,
tem características e qualidades consideradas inigualáveis, conferindo leveza e alta
digestibilidade aos confeitos preparados (NEVES et al., 2005).
O mercado nacional e internacional de alimentos tem interesse em M.
arundinacea especialmente pela elaboração de produtos destinados a confeitaria
(LEONEL et al., 1998).

8
As características culinárias do amido, o destacam principalmente pela
leveza dos biscoitos, mingaus e bolos, em substituição às farinhas de trigo, milho ou
mandioca, além de ser utilizada para diversos usos medicinais (COELHO et al.,
2005).
O amido da araruta pode ser qualificado através da limpeza, conteúdo de
polpa e fibras, umidade, pH, viscosidade, cinzas e o mais importante, classificação
através de sua cor que deve ser mais branca possível (PURSEGLOVE, 1985). Suas
características básicas são de um pó branco, inodoro e insípido (BERNAL e
CORREA, 1994).
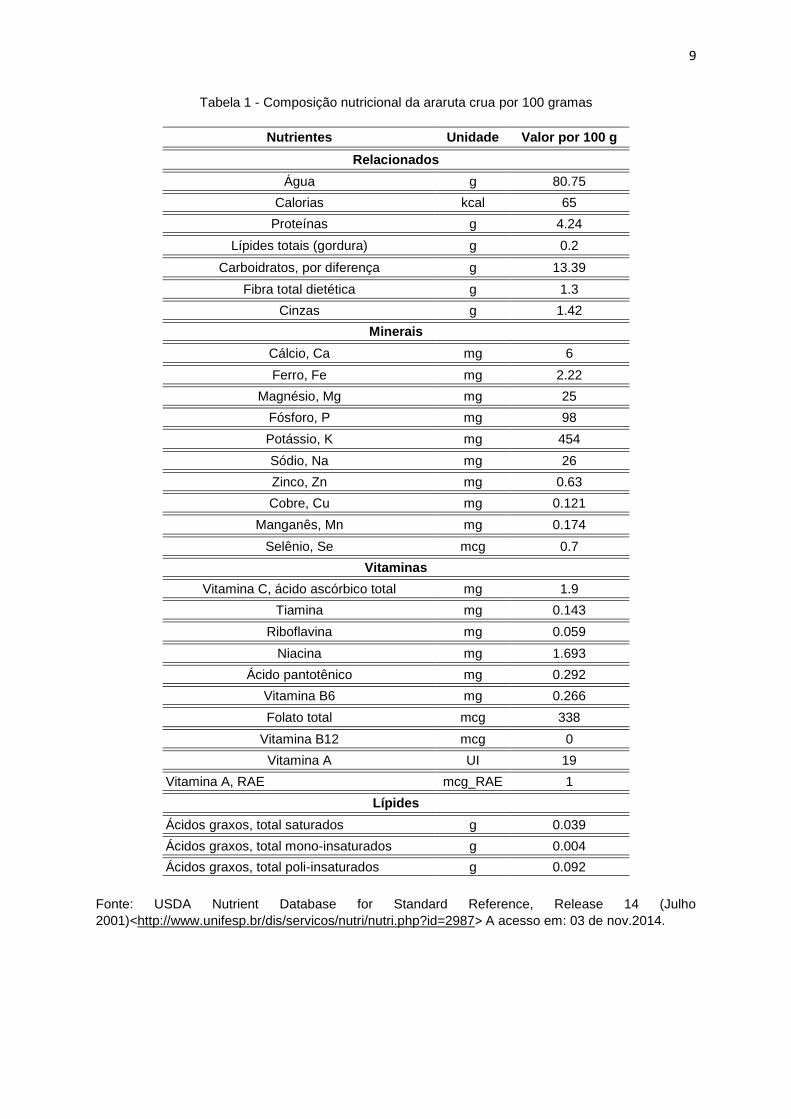
O rizoma contém, além de amido, celulose, proteína, açúcar, mucilagem e
sais minerais, betacaroteno, niacina, riboflavina, tiamina entre outros (Tabela 1),
algumas alterações desses componentes podem ocorrer devido a fatores como
clima, grau de fertilidade do solo, dentre outros fatores bióticos e abióticos
(MARTINS, 1943).
O cultivo das hortaliças não-convencionais, tais como a araruta no Brasil é
realizado principalmente por agricultores familiares. O conhecimento para o cultivo e
consumo destas plantas foi passado através das gerações. A maioria do cultivo é
através da agricultura familiar para consumo próprio, sem desígnio comercial
(BRASIL, 2010).
A araruta já foi muito cultivada no Brasil, mas perdeu espaço nos últimos 50
anos, chegando quase à extinção, devido à concorrência de outros amidos
produzidos em nível industrial como a mandioca, milho, aveia, cevada e trigo, o
cultivo da araruta foi sendo gradualmente substituído, porém, o amido de mandioca
não possui as mesmas características de fácil digestibilidade e capacidade de
gelificação (SILVA et al., 2000). Portanto, devido a sua importância é fundamental a
caracterização do seu material genético, através de marcadores moleculares, como
meio de preservação e conservação de seu germoplasma, com a possibilidade
futura da introdução de características fundamentais e que permanecem aos
diferentes genótipos, para formação de uma espécie melhorada.
9
Tabela 1 - Composição nutricional da araruta crua por 100 gramas
Nutrientes Unidade Valor por 100 g
Relacionados
Água g 80.75
Calorias kcal 65
Proteínas g 4.24
Lípides totais (gordura) g 0.2
Carboidratos, por diferença g 13.39
Fibra total dietética g 1.3
Cinzas g 1.42
Minerais
Cálcio, Ca mg 6
Ferro, Fe mg 2.22
Magnésio, Mg mg 25
Fósforo, P mg 98
Potássio, K mg 454
Sódio, Na mg 26
Zinco, Zn mg 0.63
Cobre, Cu mg 0.121
Manganês, Mn mg 0.174
Selênio, Se mcg 0.7
Vitaminas
Vitamina C, ácido ascórbico total mg 1.9
Tiamina mg 0.143
Riboflavina mg 0.059
Niacina mg 1.693
Ácido pantotênico mg 0.292
Vitamina B6 mg 0.266
Folato total mcg 338
Vitamina B12 mcg 0
Vitamina A UI 19
Vitamina A, RAE mcg_RAE 1
Lípides
Ácidos graxos, total saturados g 0.039
Ácidos graxos, total mono-insaturados g 0.004
Ácidos graxos, total poli-insaturados g 0.092
Fonte: USDA Nutrient Database for Standard Reference, Release 14 (Julho
2001)<http://www.unifesp.br/dis/servicos/nutri/nutri.php?id=2987> A acesso em: 03 de nov.2014.

10
1.4 Marcadores Moleculares
Com o advento das técnicas moleculares os marcadores de DNA tornaram-
se indicadores para o estudo de genética de vários organismos, incluído as plantas.
Estas técnicas tornaram-se rotineiramente aplicadas em pesquisa sobre a
biodiversidade, estudos de evolução e biotecnologia. Os marcadores moleculares
são características que identificam o perfil da molécula de DNA (fingerprinting) que
podem ser herdados e são capazes de diferenciar dois ou mais indivíduos (MILACH,
1998).
As técnicas aplicadas aos estudos de marcadores moleculares tornaram-se
cada vez mais rápidas, precisas e baratas, nas análises de detecção das variações
genéticas entre os organismos. No entanto, algumas técnicas são mais adequadas
do que outras.
Um marcador molecular deve apresentar algumas características
desejáveis, como: (i) ser de natureza altamente polimórfica para uso em estudos de
diversidade genética; (ii) possuir herança codominante para a determinação de
homozigose e ou heterozigose de organismos diplóides; (iii) ocorrer com frequência
no genoma, de forma uniforme e distribuída; (iv) apresentar comportamentos
seletivos neutros, pois as sequências de DNA são neutras em condições
ambientais; (v) fácil de ser detectado (disponibilidade): simples e rápido; (vi) ensaios
fáceis, rápidos e baratos; (vii) alta reprodutibilidade e (viii) fácil troca de dados entre
os laboratórios (KUMAR et al., 2009).
Entretanto, é extremamente difícil encontrar um marcador molecular, que
reúna todos os critérios acima mencionados.
Para avaliação e detecção do polimorfismo em nível de DNA, há diversas
técnicas moleculares disponíveis e a escolha da técnica ideal depende do tipo de
estudo a ser realizado. A escolha de um marcador deve ser determinada, quando
preenche pelo menos algumas das características acima citadas (WEISING et al.,
1995).
Vários tipos de marcadores moleculares são utilizados para avaliar o
polimorfismo de DNA e são geralmente baseados na hibridação e/ou na reação em
cadeia da polimerase (PCR). Na hibridização, os perfis dos tamanhos dos
fragmentos de restrição da molécula de DNA são visualizados por hibridação de
uma sonda marcada com substância radioativa e/ou fluorescente, a partir de um

11
fragmento de DNA de origem conhecida. A PCR é baseada na amplificação in vitro
de sequências de DNA específicas ou loci, orientada por oligonucleotídeos
específicos ou aleatórios (MATIOLI, 2001).
Em relação ao tipo de marcador é importante diferenciar aqueles que
fornecem dados de um único loci, daqueles que possuem múltiplos loci. Esta
informação é extremamente importante devido à quantidade de informações e
interpretação dos dados (MATIOLI, 2001).
Os marcadores multilocos utilizados em DNA fingerprinting, são as técnicas
denominadas de DNA polimórfico amplificado ao acaso (RAPD), polimorfismo de
tamanho de fragmentos amplificados (AFLP), Microssatélites e Minissatélites, os
quais são mais adequados para estudos de identificação genética, teste de
paternidade e estudo de variabilidade genética intra-espécie. Enquanto os
marcadores de Amplified Fragment Length Polymorphism (RFLPs) obtidos de DNA
mitocondrial (mtDNA), DNA de cloroplasto (cpDNA), DNA ribosomal (rDNA) são
mais apropriados para análises de espécies relacionadas (interespécies) (MATIOLI,
2001).
O uso da técnica da reação em cadeia da polimerase (PCR), utilizada para
vários marcadores, como Random Amplified Polymorphic DNA (RAPD) ou DNA
amplificado ao acaso (WILLIAMS et al., 1990), AFLP ou polimorfismo de
comprimento de fragmento amplificado (VOS et al., 1995) e marcadores
microssatélites ou sequências simples repetidas (SSR) (MORGANTE e OLIVIERI,
1993).
As informações obtidas por meio dos marcadores moleculares são muito
úteis para auxiliar na identificação dos genótipos contrastantes em programas de
melhoramento de plantas o qual incluem diversos padrões para obtenção de
variabilidade genética, seleção de indivíduos superiores, avaliação de materiais
genéticos promissores para o comércio. Nesses programas os usos de marcadores
de DNA são aplicados no monitoramento e organização da variabilidade genética, a
seleção assistida por marcadores moleculares no qual detecta diferenças genético-
moleculares, representando o futuro promissor na agricultura e a proteção de
cultivares (FARALDO et al., 2003, FUKUDA et al., 1994; LEE, 1995).
Os marcadores moleculares constituem ferramentas de grande importância
para a identificação e caracterização dos acessos mantidos em Bancos de
Germoplasma de várias plantas tais como: mandioca, milho, feijão, soja, entre

12
outras, cujo principal objetivo é a manutenção da variabilidade genética (FALEIRO,
2007).
Sabe-se que muitos pesquisadores melhoristas, evitam trabalhar com
materiais disponíveis nas coleções de genótipos silvestres e etnovariedades por
falta de identificação e caracterização de genes com potencial para melhoramento
genético (FARALDO et al., 2003; NASS, 2001).
A escolha de genitores e o planejamento de cruzamento em programas de
melhoramento genético são etapas fundamentais para o sucesso do programa. Para
balizar este processo são essenciais a realização de caracterização genética
apuradas e precisas da coleção de acesso do banco de germoplasma, a qual é
obtida com base na avaliação acurada de características morfológicas, fisiológicas e
agronômicas, além de dados de pedigree (FALEIRO, 2007).
Dados adicionais sobre a diversidade genética de potenciais genitores
obtidos com base em marcadores moleculares podem auxiliar na escolha dos
genitores e no planejamento dos cruzamentos visando o melhoramento do efeito da
heterose e das recombinações gênicas desejadas (FALEIRO, 2007).
1.5 Marcador Molecular RAPD
A técnica de PCR permitiu o desenvolvimento de várias classes de
marcadores moleculares os quais são cada vez mais utilizados em várias espécies
de plantas. Dentre esses marcadores, destaca-se o Random Amplified Polymorphic
DNA (RAPD) ou DNA amplificado ao acaso (WILLIAMS et al.,1990).
A técnica de RAPD (Figura 6) é baseada na repetição cíclica da extensão
enzimática de iniciadores, como pequenos primers complementares, que se
hibridizam ao DNA e servem como molde na amplificação através da PCR. Os locos
que apresentam duas sequências alvo em fitas opostas com orientação e
espaçamento adequado são amplificados pelo processo de PCR (TINGEY et al.,
1992). Nesse método um único primer é utilizado o qual é normalmente formado por
diferentes combinações das quatro bases nitrogenadas que apresentam uma
sequência predominante de bases citosina e guanina (C/G 60 a 80%) e com alvos
desconhecidos. O RAPD tem sido adotado por alguns pesquisadores envolvidos no
desenvolvimento de métodos de identificação de cultivares, principalmente por ser
uma técnica simples que não requer nenhuma informação prévia sobre as

13
sequências de nucleotídeos do genoma da espécie, além de ser bastante acessível
e de custo relativamente baixo e pode ser utilizado sobre as informações
moleculares escassas (FRITSCH e RIESEBERG,1996; WEEDEN, 1992; MENEZES
et al., 2002).
Figura 4. Representação esquemática da técnica (RAPD).Fonte adaptada pela autora:
Disponível em: <http://www.usask.ca/.../projects_2002/pawlin/resources/rapd1.jpg>. Acesso em: 03
de nov. 2014.
Por apresentarem primers de sequência curta, há grande probabilidade de
que esses primers encontrem regiões do genoma onde se anelarem, fazendo com
que diversos fragmentos de tamanhos diferentes resultem de uma reação
(WILLIAMS et al., 1990).
Os polimorfismos são identificados pela presença de um fragmento específico
de DNA amplificado em um determinado genoma, comparado à ausência do mesmo
fragmento de outro genoma por apresentar de diferentes comprimentos, podem ser
separados e distinguidos por eletroforese (TINGEY et al., 1992). Apresentam
algumas limitações por serem marcadores dominantes, ou seja, indivíduos
homozigotos dominantes e heterozigotos não podem ser diferenciados, pois ambos
serão visualizados pela presença de uma banda no gel (WILLIAMS et al., 1990).
Contudo existem muitos trabalhos demonstrando resultados de identificação de

14
acessos com marcadores RAPD, utilizados na alimentação humana, tais como
mandioca, milho, arroz, cúrcuma longa e pimenta são alvos dessa técnica.
O uso da técnica RAPD produz um grande número de marcadores
moleculares, permitindo encontrar marcadores específicos de gêneros, espécies,
subespécies, ou raças, sendo de grande interesse para o estabelecimento de
relações taxonômicas (HADRYS et al., 1992; AAGAARD et al., 1995), para o
estabelecimento de relações filogenéticas e diferenciação de espécies próximas
(ARNOLD et. al., 1991; LINFANTE e AGUINAGALDE, 1996; WOLFF e MORGAN-
RICHARDS, 1998), e para detecção de híbridos (DAEHLER e STRONG, 1997; DE
GREEF e TRIEST, 1999; KUEHN et al., 1999).
Nos estudos de Angel et al. (2008), utilizando 11 espécies de cúrcuma,
testados com 20 primers, no total de 274 bandas foram geradas, das quais 264
foram polimórficas revelando um elevado grau de polimorfismo, possibilitando a
identificação da espécie.
A técnica de RAPD e ISSR foram utilizadas para avaliar a variação genética
entre 13 cultivares de Calathea em 11 espécies. No total, 262 bandas foram
encontradas, das quais 252 eram polimórficas, resultando em alto grau de
polimorfismo 96,1%. Assim, estes marcadores têm o potencial para a identificação
de espécies / variedades e útil em programas de melhoramento genético (ROUT, et
al.,2007).
Outro estudo analisou a variabilidade genética entre Açafrão (Curcuma longa)
entre 20 acessos obtidos de diferentes regiões do Brasil, onde foram utilizados 45
loci polimórficos. Através de teste estatístico AMOVA houve a formação de dois
grupos. A proporção da variabilidade genética existente entre os três acessos pré-
estabelecidos permitiu verificar 44,49% de variabilidade genética entre os grupos
(PINHEIRO et al., 2003).
Estudos já publicados envolvendo plantas da mesma família, como o açafrão
e o gengibre, foram utilizados como referência para o desenvolvimento deste
trabalho, utilizando a M. arundinacea. Existem muitos trabalhos na literatura
utilizando marcadores de DNA para a caracterização da variabilidade genética em
muitas espécies de populações naturais (JAGGI et al., 2000; BOUZAT, 2001).

15
1.6 PCR Convencional
As técnicas que envolvem o uso de DNA apresentam alto poder de
resolução, com as diferenças entre os indivíduos detectadas nas sequências de
nucleotídeos distribuídas pelo genoma (FERREIRA e GRATTAPAGLIA, 1998), que
permitem a distinção entre genótipos com elevada facilidade, utilizando primers
pequenos e de sequência arbitrária, revelando ser uma técnica muito simples e de
custo moderado, permitindo a análise genética de varias espécies (HADRYS et al.,
1992).
Diversas técnicas estão disponíveis, as baseadas na reação da polimerase
em cadeia (PCR) oferecem vantagens em relação a outros métodos, pois as
quantidades de DNA utilizadas são mínimas na proporção de picogramas ou
nanogramas, permitindo a amplificação do produto, resultando em moléculas de
DNA com sequência homóloga ao DNA, que serviu de origem e os perfis
eletroforéticos são obtidos com maior rapidez (FERREIRA e GRATTAPAGLIA,
1998; GUIMARÃES, 1999; TAYLOR, 1993).
Kary Mullis nos anos 80 desenvolveu a técnica de PCR Polymerase Chain
Reaction ou Reação em Cadeia de Polimerase, realizada em um termociclador,
aparelho que permite a realização de três etapas fundamentais para amplificação do
DNA, que são desnaturação, anelamento e extensão.
A desnaturação do DNA consiste na elevação da temperatura a 92/95°C e
no rompimento das pontes de hidrogênio, separando a fita dupla de DNA. As fitas
simples de DNA são expostas, criando um molde, nas quais serão anelados o
primer ou iniciador, complementando sua sequência (EISENSTEIN, 1990,
SCHEINERT et al., 2005). Na etapa de anelamento ocorre uma variação na
temperatura de 35/60°C dependendo essencialmente do tamanho e sequencia dos
iniciadores utilizados, permitindo a hibridização DNA-DNA de cada iniciador com as
sequências complementares da fita simples (molde) que flanqueiam a região alvo.
Na última etapa ocorre a extensão no sentido 5’- 3’, através da ativação da enzima
DNA polimerase, em uma temperatura elevada 72°C (TAYLOR, 1993) .
Um fator importante deve-se aos seus sítios de anelamentos que devem
estar em uma distância suficiente para que não ocorra o anelamento entre os
primers, assim permitindo a síntese subsequente de um novo produto. Para uma
amplificação perfeita se faz necessária à padronização da reação e esta consiste na

16
otimização dos reagentes utilizados, como o tampão da Taq DNA polimerase,
MgCl2, iniciadores (com pequenas sequências de nucleotídeos, denominados de
primers), desoxirribonucleotídeos trifosfatados (dATP, dCTP, dGTP,dTTP) e a
enzima termoestável Taq DNA polimerase, isolada da bactéria termofílica Thermus
aquaticus (EISENSTEIN, 1990).
A quantidade de ciclos geralmente é determinada pelo pesquisador e/ou de
acordo com o tamanho do produto a ser amplificado e a especificidade do primer, o
ciclo é repetido por algumas dezenas de vezes, as fitas de DNA se multiplicam à
medida que os ciclos vão se repetindo e os primers consumidos, esses primers irão
definir a sequência a ser replicada, resultando na amplificação de uma determinada
sequência de DNA com bilhões de cópias (EISENSTEIN, 1990; MULLIS, 1987).
1.7 DNA barcoding
Nas plantas, as sequências de DNA podem ser obtidas a partir de três
genomas: cloroplasto (cpDNA), mitocôndria (mtDNA) e núcleo (nDNA). As variações
que ocorrem nesses genomas, tais como mutações pontuais e rearranjos, incluindo
inversões, deleções e inserções, são os caracteres utilizados em estudos de
filogenia (Figura 4).
Estas sequências de DNA de regiões padrão têm sido utilizadas como
ferramentas para a identificação de espécies de organismos e são denominadas de
barcoding (código de barra).
Segundo o - Consortium for the Barcode of Life - CBOL (2009), DNA
barcoding envolve o sequenciamento de uma região padrão de DNA como uma
ferramenta para a identificação de espécies. Lahaye et al., (2008), afirmam que
DNA barcoding é uma técnica em que a identificação de espécies é realizada
usando sequências de DNA a partir de um pequeno fragmento do genoma, com o
objetivo de contribuir nos estudos ecológicos e de conservação em que a
identificação taxonômica tradicional seria inviável.

17
Figura 5. Regiões mais utilizadas como DNA Barcoding em plantas Fonte modificada pela autora de: <http://www.sobiologia.com.br/conteudos/Morfofisiologia_vegetal/morfovegetal17.php>
<http://losrelatosdesamid.blogspot.com.br/2013/05/mitocondriashtml>
<http://cmapspublic2.ihmc.us/rid=1N2FN04F7JMPKS2109/Recursos%20para%20organelas%20citopl
asmaticos>. Acesso em: 03 de nov. 2014.
Para uma região do gene se tornar um DNA barcoding foram definidos três
critérios: conter variabilidade genética entre espécies em nível significativo de
divergência, possuir locais de flancos conservados para o desenvolvimento de
primers universais para PCR a fim de serem aplicados em estudos de taxonomia,
além de terem um comprimento de sequência curta, para facilitar a extração e
amplificação do DNA (KRESS et al., 2008).
O marcador barcoding, mais utilizado é uma região do gene citocromo c
oxidase mitocondrial (COI ou cox1), muito empregado nos grupos de animais. O
mesmo barcoding foi testado para as plantas terrestres, entretanto não
demonstraram sucesso, a dificuldade está nas 300,000 espécies de plantas
terrestres (HEBERT et al., 2003).
Portanto, é inviável sua utilização em plantas principalmente as
Angiospermas, devido a divergências das regiões codificantes do gene COI e por
rapidamente terem a estrutura do seu genoma mitocondrial modificada, além de

18
apresentarem baixas taxas de substituição de bases no DNA mitocondrial,
características que levaram a busca de regiões barcoding alternativos, como as
regiões do DNA plastidial (KRESS et al., 2005; FAZEKAS et al., 2008; CBOL,
2009).
O Consortium for the Barcode of Life criado em 2004 com intuito de facilitar e
formalizar a seleção de um código de barras de plantas. Para isso realizou-se a
formação de um grupo de trabalho com representantes de consórcios de diferentes
grupos de pesquisa por parte da comunidade sistemática que propôs, os sete
principais marcadores barcoding candidatos, presentes no DNA de cloroplasto
(HOLLINGSWORTH et al., 2011).
O cloroplasto é uma organela citoplasmática, abundante nas células vegetais
constituído por apenas um cromossomo circular facilmente isolado (PALMER,
1987). O genoma plastidial possui 120-160 kilo pares de bases (kpb) a estrutura
geral é representada por duas regiões a Large Single Copy Region (LSC) e a Small
Single Copy Region (SSC) interrompidas por duas copias de Large Inverted
Reapets (IRa e IRb) (Figura 5). O genoma do cloroplasto contem todos os genes
ribosomais (rRNA) e transportadores para a síntese das proteínas fundamentais
para seu funcionamento (SHINOZAKI et al., 1986).
Geralmente, o cpDNA apresenta herança unipariental, sendo transmitido
apenas pelo gameta feminino na maioria das angiospermas (DEMESURE et al.,
1995). Além disso, é altamente conservado em termos de tamanho, conteúdo,
estrutura e ordem dos genes (JUDD et al., 2002). Isso permite que iniciadores
específicos desse genoma sejam utilizados em quase todo o reino vegetal,
facilitando a comparação entre os diferentes táxons.
A recomendação de um código de barras padrão de plantas foi realizada pelo
CBOL (2009), que comparou o desempenho das sete principais regiões presentes
no cpDNA, denominadas de matK, rbcL, rpoB, e rpoC1,ATPF-atpH, trnH-psbA, e
psbK-psbI (KRESS et al., 2005; SHAW et al., 2007; ŠTORCHOVÁ e OLSON, 2007).

19
Figura 6. Esquema do Genoma do Cloroplasto. Fonte:<http://pt.slideshare.net/lonachaliha/chloroplast-21493415>. Acesso em 03 de nov.
2014.
.
O gene matK codifica a proteína maturase K, o gene rbcL codifica a
subunidade grande da ribulose-1,5- bifosfato carboxilase/oxigenase, o gene rpoB
codifica a subunidade β da enzima RNA polimerase do cloroplasto, e o gene rpoC1
codifica a subunidade β’ da enzima RNA polimerase do cloroplasto. As regimer
atpF-atpH, trnH-psbA e psbI-psbK são espaçadores Intergênicos. Os genes atpF e
atpH codificam as subunidades CFO I e CFO III da ATP síntase, respectivamente.
Os genes psbK e psbI codificam dois polipeptídios de baixa massa molecular do
fotossistema II; estas duas regiões são altamente conservadas desde as algas até
as plantas terrestres e parasitarias (LAHAYE et al., 2008, LOPES 2011). O gene
trnH e um gene de tRNA e psbA codifica a proteina D1 do fotossistema II. Todos

20
estes dez genes são cloroplastidiais e localizam-se na região Large Single Copy
(LSC) do genoma do cloroplasto (LOPES, 2011).
Após a avaliação das propostas e inúmeros trabalhos na busca barcoding em
plantas o comitê executivo da CBOL concluiu que só rbcL (marcador de taxa
evolutiva lenta, porém conservada) e matK (mais variável porém de maior
resolução) estão entre as regiões de código de barras aprovados para as plantas
terrestres (CBOL, 2009).
Portanto, o CBOL ao recomendar os genes rbcL e matK como o código de
barras para as plantas terrestres, considerou ser necessário a introdução de novos
dados para a caracterização desses genes, assim como os progressos obtidos
através dos primers do matK e rbcL.
1.7.1 Maturase K
O gene matK é utilizado como um marcador na construção filogenética e tem
mostrado alto poder de distinção taxonômico tanto em níveis de ordem e família
quanto em níveis de gênero e espécies de plantas, devido a sua rápida evolução,
presença em todas as plantas e elevado poder de discriminação entre espécies
(HILU e LIANG, 1997).
O gene matK tem aproximadamente 1500 pb está localizado dentro de um
íntron classe II, entre os exons 5’ e 3’ do gene que codifica o trnK. (SUGITA et al.,
1985; STEANE, 2005; DANIELL et al., 2006; TURMEL et al., 2006). Este gene é de
cópia única contém altas taxas de substituição dentro da espécie e está emergindo
como potencial candidato para estudos de sistemática vegetal e evolução. Desta
forma, filogeneticamente, a taxa de evolução do matK é adequada para resolver
relações intergênero, bem como as relações interespécies em muitas plantas
angiospermas (HILU e LIANG, 1997) .
O complexo gene matK - trnK é comumente utilizado para estudos de
evolução de plantas e aborda a solução para vários níveis taxonômicos. O gene
matK tem o tamanho ideal, elevada taxa de substituição, grande proporção da
variação em nível do ácido nucleico, na primeira e segunda posição do códon,
baixo razão de transição/ transversão, e a presença de setores mutacionalmente
conservadas (HILU e LIANG, 1997) .
Para os estudos de barcoding o segmento utilizado de matK compreende em
aproximadamente 790 pb (STOECKLE et al., 2011). Devido as características de

21
alto poder de discriminação e universalidade das espécies o CBOL recomendou
como código de barras de plantas o matK em combinação com rbcL (VIJAYAN e
TSOU, 2010).
1.7.2 rbcL
Pela sua característica a maior subunidade da ribulose-1,5-bifosfato
carboxilase oxidase (RuBisCO ) desperta o interesse de engenheiros genéticos e
filogenistas de plantas, por ser um gene codificante de uma proteína altamente
conservada o rbcL. O sítio ativo, que é responsável por quase toda a fixação de
carbono na terra (KELLOGG e NICKOLAS, 1997) é o gene mais comum utilizado
para fornecer dados filogenéticos.
Este gene está localizado na região Large Single Copy do cloroplasto, com
aproximadamente 1.430 pares de bases, e apresenta uma taxa de evolução lenta e
bastante conservada, apresentando boa resolução em níveis taxonômicos mais
elevados, como família. Como a RuBisCo é uma enzima fotossintética muito
importante, o rbcL foi o primeiro gene das plantas a ser sequenciado (ZURAWSKI et
al., 1981). Este gene tem sido utilizado amplamente em estudos filogenéticos de
plantas com mais de 10000 sequências do rbcL, já disponíveis no GenBank
(CHASE et al., 2007).
2 OBJETIVOS
Identificar os acessos de M. arundinacea por meio dos genes cloroplastidiais
matK (maturase K) e o rbcL (ribulose-1,5-bifosfato carboxilase/oxigenase).
Determinar a similaridade genética entre os acessos de M. arundinacea
(intraespecífica), por meio de marcadores RAPD.

22
REFERÊNCIAS BIBLIOGRÁFICAS
AAGAARD, J. E.; VOLMER, S. S.; SORENSEN, F. C.; STRAUSS, S. H. Mitochondrial DNA products among RAPD profiles are frequent and strongly differentiated between races of Douglas-fir. Molecular Ecology, v.4, n.4, p.441-447, 1995. ANDERSSON L.. Revision of Maranta subgen. Maranta (Marantaceae). Nordic Journal of Botany,v. 6, n.6, p.729–756, 1986. ANGEL, G.R., MAKESHKUMAR, T., MOHAN, C., VIMALA, B. and NAMBISAN, B. Genetic diversity analysis of starchy Curcuma species using RAPD markers. Journal of Plant Biochemistry and Biotechnology, v.17, n.2, p.173-176, 2008.
ARNOLD, M. L.; BUCKNER, C. M. & ROBINSON, J. J. Pollenmediated introgression and hybrid speciation in Louisiana irises. Proceeding of the National Academy of Science of USA, v.88, n.4, p. 1398-1402, 1991. ARNS, Y.K.N. Estudo Taxonômico dos grupos Monotagma, Maranta e Myrosma (Marantaceae) em Pernambuco. 1997.105p. Dissertação de Mestrado – Biologia Vegetal. Universidade Federal de Pernambuco, Pernambuco,1997. BENTLEY, R.; TRIMEN, H. Medicinal plants. London: J. and A. Churchill, 1880. v.4. p.228-306. BERNAL, H.Y.; CORREA, J.E. (eds.). Espécies vegetales promisorias de los paises Del convenio Andrés Bello. Bogotá: Guadalupe, 1994. 547p. Tomo 10. (PREVECAB. Série Ciência e Tecnologia, 53). BOUZAT, J. L. The population genetic structure of the Greater Rhea (Rhea Americana) in an agricultural landscape. Biological Conservation, v.99, n.3, p. 277-284, 2001. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Manual de hortaliças não-convencionais / Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Desenvolvimento Agropecuário e Cooperativismo. – Brasília : Mapa/ACS, 2010. 92 p CBOL Plant Working Group. 2009. A DNA barcode for land plants. Proceedings of the National Academy of Sciences, v.106, n.31, p.12794—12797, 2009. CEREDA, M.P. Propriedades gerais do amido. São Paulo, Fundação Cargill, 89-100 p. (Série: Culturas de Tuberosas Amiláceas Latino-americanas, v. 1) 2002.
CEREDA, M. P. Produção de fécula a partir do biri (Canna edulis).VILPOUX, O. (Org.) Tecnologia, uso e potencialidades de tuberosas amiláceas

23
latinoamericanas. São Paulo: Fundação Cargill, 2003. 191-199 p. (Série: Culturas de Tuberosas Amiláceas Latinoamericanas, v.3). CEREDA, M. P. Massas alimentícias alternativas à base de amido. VILPOUX, O. (Org.) Tecnologia, uso e potencialidades de tuberosas amiláceas latinoamericanas. São Paulo: Fundação Cargill, 2003. 201-219 p. (Série: Culturas de Tuberosas Amiláceas Latinoamericanas, v.3).
CHASE, M.W.; COWAN, R.S.; HOLLINGSWORTH, P.M.; BERG, C.; MADRINAN, S.; PETERSEN, G.;SEBERG, O.; JORGSENSEN, T.; CAMERO, K.M.; CARINE, M.; PEDERSEN, N.; HEDDERSON, T.A.J.;CONRAD, F.;SALAZAR, G.A.; RICHARDSON, J.E.;HOLLINGSWORTH, M.L.; BARRACLOUGH, T.G.; KELLY,L. ; WILKINSON, M. A proposal for a standardized protocol to barcode all land plants. Taxon 56, v.2, n.2, p. 295–299, 2007.
COELHO, I. da S; SANTOS, M. da C. F; ALMEIDA, D. L. de; SILVA, E. M. R. da; NEVES, M. C. P. Como plantar e usar a araruta. Brasília: Embrapa Informação Tecnológica; Seropédica: Embrapa Agrobiologia, 2005. 55 p.
COSTA, FLÁVIA R. C.; ESPINELLI, F. P.; FIGUEIREDO, F. O. G. Guia de marantáceas da Reserva Ducke e da Reserva Biológicado Uatumã = Guide to the Marantaceae of the Reserva Ducke and Reserva Biológica do Uatumã, Manaus : INPA, 2008, 154p. DAEHLER, C. C.; STRONG, D. R. Hybridization between introduced smooth cordgrass (Spartina alterniflora; Poaceae) and native California cordgrass (S. foliosa) in San Francisco Bay. American Journal of Botany, v.84,n.5, p. 607-611, 1997. DANIELL, H.; LEE, S.B.; GREVICH, J.; SASKI, C.; QUESADAVARGAS, T.; GUDA, C.; TOMKINS, J.; JANSEN, R.K. Complete chloroplast genome sequences of Solanum bulbocastanum, Solanum lycopersicum and comparative analyses with other Solanaceae genomes. Theoretical and Applied Genetics, v.112, n.8, p. 1503–1518, 2006. DE GREEF, B.; TRIEST, L. The use of random amplified polymorphic DNA (RAPD) for hybrid detection in Scirpus from the river Schelde (Belgium). Molecular Ecology, v.8, n.3, p.379-386, 1999. DELGADO, H.S.; SIFUENTES, T.C. Plantas medicinales del Jardin Botânico IMETIPSS. Iquitos: Instituto Peruano de Seguridad Social/Instituto de Medicina Tradicional, 1995. 85p. DEMESURE, B.; SODZI, N.; PETITI, R.J. A set of universal primers for amplification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Molecular Ecology, v.4, n.1, p.129-131, 1995.

24
EBDA – Empresa baiana de Desenvolvimento Agrícola S.A. Disponível em: <http://www.ebda.ba.gov.br/ebda-resgata-a-cultura-da-araruta-em-cruz-das-almas/> Acesso em : 03/11/2014. EISENSTEIN, B. I. The polymerase chain reaction: A new method of using molecular genetics for medical diagnosis. The New England Journal of Medicine, v.322, n. 3, p. 178-183, 1990. ERDMAN, M.D.; ERDMAN, B.A. Arrowroot (Maranta arundinacea): food, feed, fuel, and fiber resource. Economic Botany, v. 38, n.3, p. 322-341, 1984. FALEIRO, F. Marcadores moleculares aplicados a programas de conservação e uso de recursos genéticos. Planaltina-DF: Embrapa Cerrados, 2007. 102 p.
FARALDO, M.I.F. Marcadores moleculares em mandioca. In: CEREDA, M.P. Agricultura: Tuberosas Amiláceas Latino Americano, São Paulo: Fundação Cargill, 2003. v.2, p.101-117. FAZEKAS, A.J.; BURGESS, K.S.; KESANAKURTI, P.R.; GRAHAM, S.W.; NEWMASTER, S.G.; HUSBAND, B.C.; PERCY, D.M.; HAJIBABAEI, M.; BARRETT, S. C. H. Multiple Multilocus DNA Barcodes from the Plastid Genome Discriminate Plant Species Equally Well. PLoS ONE. v. 3, n.7, p. e2802, 2008. FERREIRA, M. E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores moleculares em análise genética. 3.ed. Brasília:EMBRAPA-CENARGEN, 1998. 220p. FRANCO, C. M. L. et al. Structural and functional characteristics of selected soft wheat starches. Cereal Chemistry, v. 79,n.2, p. 243-248, 2002. FRITSCH, P.; RIESEBERG, L. H. The use of Random Amplified Polymorphic DNA (RAPD) in conservation genetics. In: SMITH, T. B.; WAYNE, R. K. (Ed.). Molecular genetic approaches in conservation, New York: Oxford University Press, 1996, p. 54-73.
FUKUDA, W. M. G.; CAVALCANTI J.; COSTA I.R.S. . Variabilidade genética e melhoramento da mandioca (Manihot esculenta Crantz). Recursos Genéticos e Melhoramento de Plantas para o Nordeste Brasileiro. 1994. Disponível em: <http://www.cpatsa.embrapa.br/catalogo/livrorg/mandioca.pdf>. Acesso: 03 de novembro de 2014. GLOBO RURAL. Araruta. Revista Globo Rural, v.11, n.131, p.56-59, 1996. GLOBO RURAL. Como plantar araruta. Revista Globo Rural, 2013. Disponível em: http://revistagloborural.globo.com/vida-na -

25
fazenda/como_plantar/noticia/2013/12/como-plantar-araruta.html Acesso : 03 de novembro de 2014. GUILBOT, A., and MERCIER, C. Starch Pages. In: The Polysaccharides, Academic Press: Orlando, 1985. v. 3, p.209-292. GUIMARÃES, C. T.; MOREIRA, M. A. Genética molecular aplicada ao melhoramento de plantas. In: BORÉM, A. Melhoramento de espécies cultivadas. Viçosa: Editora UFV, 1999. p.715-740. HADRYS, H.; BALICK, M.; SCHIERWATER, B. Applications of Random amplified polymorphic DNA (RAPD) in molecular ecology. Molecular Ecology, v.1, n.1, p.55-64, 1992. HAMMEL, B.E. The vascular flora of La Selva Biological Station, Costa Rica. Marantaceae. Selbyana, Sorasota, v. 9, n.1, p.234–242, oct.1986. HEBERT, P.D.N.; CYWINSKA, A.; BALL S.L.; DEWAARD, J.R. Biological identifications through DNA barcodes. The Royal Society, London, v. 270, n.1512, p.313-321, jan.2003. HERBERT, P.D.N; RATNASINGHAM, S.; DeWAARD J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. The Royal Society, v.270, n.1, p. 96–99, 2003. HILU, K.W.; e LIANG H. The matK gene: sequence variation and application in plant systematics. American Journal of Botany, v.84, n.6, p. 830– 839, 1997. HOLLINGSWORTH, P.M; GRAHAM, S.W; LITTLE, D.P. Choosing and Using aPlant DNA Barcode. PLoS ONE, v. 6, n.5, p. e19254, 2011. JAGGI,C.; WIRTH, T.; BAUR, B. Genetc variability in subpopulations of the asp viper (Vipera aspis) in the Swiss Jura Mountains: implications for a conservation strategy. Biological Conservation, v.94, n.1, p.69-77, 2000. JUDD, W. S.; CAMPBELL, C. S.; KELLOGG, E. A.; STEVENS, P. F. and DONOGHUE, M. J. Plant Systematics: a phylogenetic. Systematic Biology, v.53, n.3, p.517-518, 2002. KELLOGG, E. A. and NICKOLAS, D. J. The Structure and Function of Rubisco and their Implications for Systematic Studies. American Journal of Botany, v.84, n.3, p. 413–428, 1997. KENNEDY, H. Systematics and pollination of the ‘closedflowered’ species of Calathea (Marantaceae). University of California Publications in Botany, 1977. v.71, p.1–90. KENNEDY, H. New species of Calathea (Marantaceae) endemic to Costa Rica. Canadian Journal of Botany, v.75, n.8, p.1356-1362, 1997.

26
KENNEDY, H. Diversification in pollination mechanisms in the Marantaceae. In: Wilson, KL, Morrisson DA, (ed). Monocots – Systematics and evolution. Melbourne: CSIRO, 2000, p.335–343. KRESS, W.J.; ERICKSON, D.L. DNA barcodes: Genes, genomics, and bioinformatics. Proceedings of the National Academy of Sciences, v.105, n.8, p.2761–2762, 2008. KRESS, W. J.; WURDACK, K. J.; ZIMMER, E. A.; WEIGT, L. A.; JANZEN, D. H. Use of DNA barcodes to identify flowering plants. Proceedings of the National Academy of Sciences, v. 102, n.23, p.8369–74, 2005. KUEHN, M. M.; MINOR, J. E.; WHITE, B. N. An examination of hybridization between the cattail species Typha latifolia and Typha angustifolia using random amplified polymorphic DNA and chloroplast DNA markers. Molecular Ecology, v.8, n.12, p. 1981-1990, 1999. KUMALASARI, I. D.; HARMAYANI, E.; LESTARI, L. A.; RAHARJO, S.; ASMARA, W.; NISHI, K.; AND SUGAHARA, T. Evaluation of immunostimulatory effect of the arrowroot (Maranta arundinacea. L) in vitro and in vivo. Cytotechnology, v.64, n.2, p. 131–137, 2012.
KUMAR, P.; GUPTA, V.K.; MISRA, A.K.; MODI, D. R. and PANDEY, B. K. Potential of Molecular Markers in Plant Biotechnology. Plant Omics Journal v.2, n.4, p.141-162, 2009. LAHAYE, R.; van der BANK, M.; BOGARIN, D.; WARNER, J.; PUPULIN, F.; GIGOT, G.; MAURIN, O.; DUTHOIT, S.; BARRACLOUGH, T. G.; SAVOLAINEN, V. DNA Barcoding the floras of biodiversity hotspots. Proceedings of the National Academy of Sciences, v. 105, n. 8, p. 2923-2928, 2008. LEE, M. DNA markers and plant breeding programs. Advances in Agronomy, San Diego, v.55, p.265-343, 1995. LEÓN, J. Botânica de los cultivos tropicales. San Jose: IICA, 1987. 445p. LEONEL, M. Análise da forma e tamanho de grânulos de amidos de diferentes fontes botânicas. Ciência e Tecnologia de Alimentos, Campinas, v. 27. n. 3. p. 579-588, 2007.
LEONEL, M.; JACKEY, S.; CEREDA, M.P.. Processamento industrial de fécula de mandioca e batata doce - um estudo de caso. Ciência e Tecnologia de Alimentos, v. 18, n. 3, p.343-345,1998. LEONEL, M.; SARMENTO, S.B.S. Características dos rizomas e fécula de araruta (Maranta arundinacea) em diferentes estágios de desenvolvimento da planta. Brazilian Journal of Food Technology, v. 8, n.2, p. 93 - 98, 2004.

27
LINFANTE, Z. D.; AGUINAGALDE, I. The use of Random Amplified Polymorphic DNA (RAPD) markers for the study of taxonomical relationships among species of Asphodelus sect. Verinea (Asphodelaceae). American Journal of Botany, v. 83, n.7, p. 949-953, 1996. LOPES, S. M. M. de B. Botânica Molecular Forense: O DNA na identificação de espécies vegetais. 2011.43 f. Dissertação (Mestrado em Ciências Forenses) – Unversidade do Porto, Portugal, 2011. MADINENI, M.,N.; FAIZA,S.; SUREKHA,R.,S.; RAVI,R.; AND GUHA, M. Morphological, Structural, and Functional Properties of Maranta (Maranta arundinacea L) Starch. Food Science Biotechnology, v. 21, n.3, p. 747-752, 2012. MARTINS, A. Q. Araruta. Revista da Flora Medicinal, v. 10, n.4, p.147-173, 1943. MARTINS, P. S. Dinâmica evolutiva em roças de caboclos amazônicos. Estudos Avançados, v.19, n.53, p. 209-220, 2005.
MATIOLI, S. R. (Org.). Biologia molecular e evolução. 1. ed. Ribeirão Preto, SP: Holos, Editora, 2001. v. 1. 202 p. MEDINA, M.H; UCO, J.G.T.; GUERRERO,L.C.; ANCONA,D.B. Caracterización fisicoquímica de almidones de tubérculos cultivados en Yucatán, México. Ciência e Tecnologia de Alimentos, v.28, n.3, p.718-726, 2008. MENEZES, C.C.E. et al. Análise da pureza genética e discriminação de cultvares de vinca (Catharanthus roseus(L.) G. Don) usando “Randon Amplified Polymorphic DNA” em DNA extraído de sementes e folhas. Revista Brasileira de Sementes, v. 24, n. 1, p. 279-285, 2002. MILACH S.C.K. Principais Tipos de Marcadores e suas Características. In: Milach, S. Marcadores Moleculares em Plantas. Porto Alegre: S. C. K. Milach, 1998, p. 17-28. MILLIKEN, W.; ALBERT, B. The use of medicinal plants by the Yanomami indians of Brazil. Part II. Economic Botany, v.51, n.3, p.264-278, 1997.
MONTEIRO, D. A.; PERESSIN, V. A. Cultura da araruta. In:CEREDA, M. P. (Coord.). Agricultura: tuberosas amiláceas Latino Americanas. São Paulo: Fundação Cargill, 2002, v. 2, p. 440-447. MORGANTE, M.; OLIVIERI, A.M. PCR-amplified microsatellites as markers in plant genetic. The Plant Journal, v.3, n.1, p.175-182, 1993. MULLIS, K. B.; FALOONA, F. Specific synthesis of DNA in vitro via a polymerasecatalyzed chain reaction. Methods in Enzymology, v. 155, n.C, p. 335-350, 1987.

28
NASS, L.L. Utilização de recursos genéticos vegetais no melhoramento. In: NASS,L.L.; VALOIS, A.C.C.; VALADARES- INGLIS, M.C.(Ed.). Recursos genéticos e melhoramento, 2001. Porto Alegre: Gráfica Editora Pallotti, Fundação de Apoio à Pesquisa de Mato Grosso, Cap.2, p 29-55. NEVES, M. C. P.; COELHO, I. S.; ALMEIDA, D. L. Araruta; resgate de um cultivo tradicional. Rio de Janeiro: Seropédica – RJ: EMBRAPA, 2005, 4p (Seropédica – RJ: EMBRAPA Comunicado técnico 79). PALMER, J.D .Chloroplast DNA evolution and biosystematic uses of chloroplast DNA variation. The American Naturalist, v.130,n.1-2, p.6–29, 1987. PECKOLT, T.; PECKOLT, G. História das plantas medicinais e úteis do Brazil. Rio de Janeiro: Laemmert e C, 1893. (5º fascículo).
PINHEIRO, J. B.; ZUCCHI, M. I.; TELES, F. L.; ÁZARA, N. A.Molecular genetic diversity in accesses of saffron using RAPD markers. Acta Scientiarum - Agronomy, v. 25, n. 1, p. 195-199, 2003.
PIO CORRÊA, M. Dicionário das plantas úteis do Brasil e das exóticas cultivadas. Rio de Janeiro: Imprensa Nacional, 1984. v. 1. p. 150-151, 309. Editado pelo Serviço de Informação Agrícola.
PURSEGLOVE, J.W. Tropical crops: monocotyledons.England: Longman Group Limited, 1985. REVILLA, J. Plantas úteis da Bacia Amazônica. Manaus: INPA/SEBRAE, 2002. v.1. ROUT, G.R.; SENAPATI, S.K. and APARAJITA, S. Studies on the genetic relationship among 13 cultivars of Calathea (Marantaceae) using RAPD and ISSR markers. Advances in Horticultural Science, v. 21, n. 3, p. 147-155, 2007.
SHAW, J.; LICKEY, E.B.; SCHILLING, E.E.; SMALL, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: the tortoise and the hare III. American Journal of Botany, v. 94, n.3, p.275–288, 2007. SCHEINERT, P.; BEHRENS, B.; KAHLE, D. Optimizing DNA Amplification Protocols using the Eppendorf® Mastercycler®. Eppendorf North America, Apr. 8. 2003. Disponível em:<http://www.eppendorfna.com/applications/PCR_appl_protocolsMC.asp. Acesso em: 3 Nov. 2014. SHINOZAKI, K.; OHME, M.; TANAKA, M.; WAKASUGI, T.; HAYASHIDA, N.; MATSUBAYASHI, T.; ZAITA, N.; CHUNWONGSE, J.; OBOKATA, J.; YAMAGUCHI-SHINOZAKI, K.; OHTO, C.; TORAZAWA, K.; MENG, BY.; SUGITA, M.; DENO, H.; KAMOGASHIRA, T.; YAMADA, K.; KUSUDA, J.;

29
TAKAIWA, F.; KATO, A.; TOHDOH, N.; SHIMADA, H. e SUGIURA, M. The complete nucleotide sequence of the tobacco chloroplast genome: its gene organization and expression. EMBO J, v. 5, n.9, p. 2043–2049, 1986. SILVA, J.R. da, ASSUMPÇÃO, R., VEGRO, C.L.R. A inserção da fécula de mandioca no mercado de amido. Informações econômicas, v.30, n.7, p.31-41, jul.2000. SILVA, J. R. S.; MONTEIRO, D. A. Cultura da araruta industrial. O Agronômico, Campinas, v. 20, n. 11/12, p. 11- 21,1968. SOUZA, V. C.; LORENZI, H. Botânica Sistemática: guia ilustrado para identificação das famílias de Angiospermas da flora brasileira, baseado em APG II. Nova Odessa, Instituto Plantarum, 2005. 640p. STEANE*, D. A.; JONES, R. C. and VAILLANCOURT, R. E. A set of chloroplast microsatellite primers for Eucalyptus (Myrtaceae). Molecular Ecology, V. 5, n.3, p. 538–541, 2005. STOECKLE, M.Y.; GAMBLE, C.C.; KIRPEKAR, R.; YOUNG,G.;AHMED, S.; LITTLE, D.P. Commercial teas highlight plant DNA barcoding successes and challenges. Scientific Reports, v. 1, p. n.42, p.7, 2011. ŠTORCHOVÁ, H.; OLSON, M.S. The architecture of the chloroplast trnH-psbA noncoding region in angiosperms. Plant Systematics Evolution, v. 268, n.1-4, p. 235-256, 2007. SUGITA, M.; SHINOZAKI, K.; SUGIURA, M. Tobacco chloroplast tRNALys (UUU) gene contains a 2.5-kilobase-pair intron: an open reading frame and a conserved boundary sequence in the intron. Proceedings of the National Academy of Sciences, v.82, n.11, p.3557–3561, 1985. TAYLOR, C. et al. Determination of the order of substrate addition to MspI DNA methyltransferase using a novel mechanism-based inhibitor. The Biochemical Journal, v. 291, n.2, p. 493–504, 1993. TINGEY, S.V.; RAFALSKI, J.A. & WILLIAMS, J.G.K. Genetic analysis with rapd markers. in: SYMPOSIUM ON APPLICATIONS OF RAPD TECHNOLOGY TO PLANT BREEDING, 1, 1992. Anais. Minneapolis: Crop Science Society of America/American society for horticultural Science/American genetic association, 1992. p. 3-8. TURMEL, M., OTIS, C. and LEMIEUX, C. 2006. The chloroplast genome sequence of Chara vulgaris sheds new light into the closest green algal relatives of land plants. Molecular Biololy and Evoution,.v.23, n.6, p.1324-1338, 2006. VIJAYAN, K.; e TSOU CH. DNA barcoding in plants: taxonomy in a new perspective. Current Science, v.99, n.11 p. 1530-1541, 2010.

30
VOS, P.; HOGERS, R.; BLEEKER, M.; REIJANS, M.; van de LEE, T.; HORNES, M.; FRIJTERS, A.; POT, J.; PELEMAN, J.; KUIPER, M.; ZABEAU, M. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Research, v.23, n.21, p.4.407-4.414, 1995. WEEDEN, N.F. Inheritance and reliability of RAPD markers. In: SYMPOSIUM ON APPLICATIONS OF RAPD TECHNOLOGY TO PLANT BREEDING, 1.,1992, Minneapolis. Anais. Minneapolis: Crop Science Society of America/American Society for Horticultural Science/American Genetic Association, 1992. p. 12. WEISHING, K.; NYBOM, H.; WOLFF, K. and MEYER, W. DNA isolation and purification. In: DNA Fingerprinting in Plants and Fungi, CRC Press, 1995 p. 44–59. WHISTLER, R. L., TURNER, E. S. Fine structure of starch granule sections. Journal Polymer Science, v. 18, p. 153-156, 1955.
WILLIAMS, J.G.K.; KUBELIK, A.R.; LIAK, K.J.; RAFALSKI, J.A.; TINGEY, S.V. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research, v.18, n.22, p.6531-6535, 1990. WOLFF, K. ; MORGAN-RICHARDS, M. PCR markers distinguish Plantago major subspecies. Theoretical and Applied Genetics, v.96,n.2, p.282-286, 1998. ZHAO, J.; WISTLER, R. L. Spherical aggregates of starch granules as flavor carriers. Food Technology, v. 48, n. 7, p. 104-105, 1994. ZURAWSKI, G.; PERROT, B.; BOTTOMLEY, W.; WHITEFIELD, P.R. The structure of the gene for the large subunit of ribulose-1,5 -bisphosphate carboxylase from spinach chloroplast DNA. Nucleic Acids Research, v. 9, n.14, p. 3251–3270, 1981.

31
CAPÍTULO I: Caracterização dos genes matK e rbcL e da
variabilidade genética entre os acessos de Araruta (Maranta
arundinacea L.).

32
Caracterização dos genes matK e rbcL e da variabilidade genética
entre os acessos de Araruta (Maranta arundinacea L.).1
1 Parte da dissertação de mestrado da primeira autora
Ingrid Batista Pinto2; Carina Elisei de Oliveira 3 .
2 Aluna do mestrado em Biotecnologia da UCDB;
3 Professora Dra. responsável pelo laboratório de Biologia Molecular da UCDB;
.
Autor para correspondência: [email protected];

33
INTRODUÇÃO
A família Marantaceae é um grande grupo de plantas da ordem Zingiberales
(NEVES et al., 2005), apresenta distribuição de forma pantropical, são plantas
típicas de florestas úmidas tropicais, crescendo principalmente em margens de rios
e clareiras, são compostas por 530 espécies e 31 gêneros (KENNEDY, 1977;
ANDERSSON, 1986; KENNEDY,1997, 2000). De acordo com Souza e Lorenzi,
2005 o Brasil, possui 150 espécies, o gênero mais representativo no país é
Calathea e Maranta.
Os representantes desta família são utilizadas como plantas ornamentais e
algumas espécies são utilizados na alimentação humana tais como Maranta
arundinacea L. e Calathea allouia Lindl, (ARNS, 1997).
Maranta é um gênero exclusivamente neotropical com 34 espécies
distribuídas em toda a América Central e do Sul, mas com a maioria das espécies
concentrada na mata Atlântica, floresta Amazônica brasileira e no cerrado (VIEIRA e
SOUZA, 2008).
A araruta (Maranta arundinacea L.) é uma planta herbácea, com caule
articulado de 1,20 m de altura, rizoma fuziforme, casca brilhante, com escamas e
produzido em tufos aderentes aos rizomas (MONTEIRO e PERESSIN, 2002). Sua
principal importância esta relacionada às características do seu amido, o qual
confere leveza e alta digestibilidade aos confeitos (NEVES et al., 2005).
Devido à produção industrial de outras fontes de amido, tais como mandioca,
milho, aveia, cevada e trigo, o cultivo da araruta foi sendo gradativamente
substituído, chegando a quase extinção, sendo de grande importância a
caracterização de seu material genético através de marcadores moleculares, como
meio de preservação e conservação da sua diversidade e populações (FARALDO et
al., 2003, NASS, 2001).
Uma das técnicas utilizadas atualmente para a identificação de espécies é
baseada na técnica de DNA barcoding a qual realizada usando sequências de DNA
a partir de um pequeno fragmento do genoma, com o objetivo de contribuir para
uma ampla gama de estudos ecológicos e de conservação em que identificação
taxonômica tradicional não é viável (LAHAYE et al., 2008). Sete principais regiões
do DNA do cloroplasto foram analisadas como potencial barcoding, mais somente o

34
gene rbcL e matK estão entre as regiões de código de barras aprovados para as
plantas terrestres, segundo o CBOL (Consortium for the Barcode of Life).
A caracterização morfológica dos acessos de plantas visa basicamente à
diferenciação fenotípica entre os acessos, contribuindo para reduzirem as
duplicações. A caracterização e avaliação do germoplasma de algumas plantas são
fundamentais para a sua utilização mais eficiente nos trabalhos de melhoramento.
(FUKUDA et al., 1994).
Técnicas envolvendo marcadores moleculares permitem compreender e
organizar a variabilidade genética de um programa de melhoramento, acessando
variabilidade de DNA, que não é influenciado pelo ambiente como são, por exemplo,
os caracteres morfológicos e fenotípicos em geral de uma planta. Entre as técnicas
disponíveis, os marcadores RAPD (WILLIAMS et al., 1990), são utilizados por
alguns pesquisadores envolvidos no desenvolvimento de métodos de identificação
de cultivares (WEEDEN, 1992; MENEZES et al., 2002), principalmente por ser uma
técnica simples que não requer nenhuma informação prévia sobre sequências de
nucleotídeos do genoma da espécie. Além disso é bastante acessível e de custo
relativamente baixo que pode ser utilizada em organismos cujas informações
moleculares ainda são escassas.
Diante do exposto, este trabalho teve como objetivo identificar os acessos de
M. arundinacea através dos genes cloroplastidiais matK (maturase K) e o rbcL
(ribulose-1,5-bifosfato carboxilase/oxigenase), assim como determinar a
variabilidade genética entre os acessos de M. arundinacea (intraespecífica), por
meio de marcadores RAPD.
MATERIAL E MÉTODOS
AMOSTRAS
Obtenção das amostras
As amostras foram obtidas de plantas matrizes de M. arundinacea do acesso
denominado de Comum (adquirido do Centro de Tecnologia e Estudos do
Agronegócio da UCDB- CeTeAgro) e os rizomas dos acessos denominados de SC ,
Guadalupe e Seta adquiridos de banco de germoplasma CERNAGEM –EMBRAPA
Brasília e cultivados no CeTeAgro (Figura 1). Os acessos foram cultivados em

35
vasos no pátio lateral da casa de vegetação do CeTeAgro e posteriormente as
folhas foram coletadas para as análises.
Figura 1. Imagens dos quatro acessos da M. arundinacea. A) Acesso Comum-CeTeAgro, B) Acesso SC- CERNAGEM –EMBRAPA Brasília, C) Acesso Guadalupe- CERNAGEM –EMBRAPA Brasília, D) Acesso Seta- CERNAGEM –EMBRAPA Brasília.
AVALIACÃO DO PERFIL GENÉTICO
Para a análise molecular foram coletadas com aproximadamente três meses
de cultivo 10 folhas jovens, de cada acesso. O material colhido foi lavado com
hipoclorito e álcool 70%, e enxaguados com água destilada. Acondicionados em
placas de Petri e secos com toalha de papel esterilizada. Com auxílio de um pistilo
esterilizado, as folhas foram maceradas em cadinho de porcelana separadamente
com nitrogênio líquido, e transferidos para tubo tipo falcon de 50 mL e
imediatamente, acondicionados em ultrafreezer – 80°C.
EXTRAÇÃO DO MATERIAL GENÉTICO
A extração de DNA foi realizada a partir de folhas de acordo com Saghai-
Maroof et al. (1984), otimizado para extração de DNA de araruta.

36
Aproximadamente, 10g de tecido foliar fresco, macerado com nitrogênio
líquido, e ressuspendidas em 10 mL de tampão de extração CTAB 1% (70 μM de
NaCl; 50 μM de EDTA pH 8,0; 100 μM Tris-HCl pH 7,5; 1% de p/v de CTAB; 1% de
p/v de polivinil pyrrolidone, 140 μl de β- mercaptoetanol e água destilada
deionizada). A seguir, as amostras foram incubadas em banho-maria a 65ºC por
aproximadamente 60 minutos, agitadas levemente a cada cinco minutos. Após foi
adicionado 15 mL de fenol/clorofórmio (1:1), invertendo os tubos. Transferindo o
sobrenadante para tubos novos e o procedimento foi repetido mais uma vez. Em
seguida, foram adicionados 15 mL de clorofórmio/álcool isoamílico (CIA) na
proporção de 24:1 (v:v), efetuando-se a inversão dos tubos seguida de
centrifugação à 3400xg por 10 minutos. O sobrenadante foi coletado e transferido
para tubos novos e adicionado 20mL de isopropanol absoluto (v:v) a –20ºC,
promovendo a precipitação do DNA, o qual foi centrifugado, retirado o
sobrenadante, permanecendo o pellet. Após o DNA foi ressuspendido em 13mL de
TE (10 mM de Tris-HCl pH 8,0 e 1 mM de EDTA). Para precipitar o DNA foi
adicionado 0,5mL de NaCl 5M e 20 mL de etanol absoluto.Os tubos foram invertidos
e centrifugados. Em seguida foi acrescentado 10 ml de etanol 70% a –20ºC,
promovendo a lavagem do DNA, seco ao ar, e suspender novamente em 1mL de
tampão TE, adicionando 120 μL de RNAse (4 mg/mL), incubando a 37ºC por
30minutos. Em seguida, a suspensão de DNA foi armazenada a 20ºC até o
momento do uso.
A concentração do DNA foi estimada através de Biofotômetro (Eppendorff). A
integridade do DNA foi avaliada por eletroforese em gel de agarose 0,8 % em
tampão TBE 1x (89mM de Tris-base; 89mM de ácido bórico; 2 mM de EDTA pH 8,0
e água destilada e deionizada até completar o volume de 100 mL). Para a
visualização do DNA, o gel foi corado com brometo de etídio na proporção de 0,5
μg/mL de gel. Após a eletroforese, foi fotografado sob luz UV em sistema digital
L.Pix Image (Loccus Biotechnology).
VERIFICACÃO DA QUANTIDADE E QUALIDADE DO DNA
A concentração e pureza do DNA foram avaliadas através de
espectrofotômetro (modelo Epperdorf AG 2231 Homburg, Bio Photometer) pela
leitura de absorbância a 260nm e a 280nm, razão das leituras A260/ A280. Para cada
amostra foram realizadas três leituras e consideradas as médias.

37
DESENHO DE PRIMERS (INICIADORES) DOS GENES MATK E RBCL
Nas análises de (PCR), foram utilizados quatro primers desenhados,
utilizando o Custom Primers - OligoPerfect™ Designer da Invitrogem, balizados nas
sequências depositadas no GenBank ( Tabela 1).
PCR CONVENCIONAL
As reações de polimerase em cadeia (PCR) foram preparadas em volume de
50μ L contendo, tampão de PCR 1X, 1,65mM MgCl2, 0,1mM de cada dNTP, 20pM
de primer, 100ng de DNA genômico e 1U de TaqDNA polimerase (Invitrogen). A
amplificação foi realizada em termociclador PTC-100 (MJ Research) programado
como se segue: desnaturação inicial de 5min a 94ºC, 35 ciclos de 1min a 94ºC, 1
min a 50ºC e 1min a 72ºC, com extensão final de 10min a 72ºC, resfriamento a 4ºC.
Os produtos da PCR foram analisados em gel de agarose 2,0% e corados com
brometo de etídio a 0,5 μg/mL. A eletroforese foi realizada em cuba horizontal (Bio
Rad®) com tampão TBE 1X. As amostras foram submetidas à eletroforese a 100
Volts por duas horas, através de fonte Power Pac HV (Bio Rad®). O gel foi
fotografado sob luz UV em sistema digital L.Pix Image (Loccus Biotechnology). Após
o método de PCR as amostras foram purificadas, quantificadas, seguido de
sequenciamento. Ambas as cadeias de DNA foram geradas em um sequenciador
automatizado UP TO 950pb (Applied Biosystems Inc., Foster City, CA, EUA), e o
alinhamento foi determinado manualmente por meio do software BioEdit v.7.2.5
(HALL, 1999). As sequências obtidas foram comparadas nos bancos de dados
internacionais como o BOLD System do International Barcode of Life, e Genbank.
Tabela 1. Genes utilizados, primers e os números de acesso no GenBank
Primer Sequência 5’ 3’ GenBank Região de
Amplificação
rbcL forward AGTGTTGGATTTAAAGCTGGTGTT JQ592613.1 1-24
rbcL reverse TGGTTTAATAGTACATCCCAATAGGG JQ592613.1 503-476
matK forward GATCACGGTTTAACTAGTTCGATTTT JQ341325.1 100-125
matK reverse CTTGTTGCAATAGAAGAAAACCC JQ341325.1 1800-1778

38
ANÁLISE DAS SEQUÊNCIAS DOS GENES MATK E RBCL
Para se analisar acessos desconhecidos molecularmente, duas metodologias
foram propostas por Hebert et al.(2003). A primeira Kimura 2- parâmetros ou
K2P(KIMURA, 1980), sendo as árvores construídas utilizando Neighbor-Joining
(SAITOU N. e NEI M., 1987). A segunda é a utilização de busca nos bancos de
dados BLAST e BOLD Systems.
As sequências forward e reverse de cada par de primer foram feitas em
duplicata, editadas, e manualmente selecionado para eliminação de sequências de
baixa qualidade. e convertidas em formato FASTA no programa BioEdit (HALL,
1999). Alinhamentos múltiplos das sequências foram realizados através do software
Jalview (WATERHOUSE, 2009).
As sequências alinhadas foram analisadas através, da busca BLAST
(blast.ncbi.nlm.nih.gov/Blast.cgi) e BOLD Systems (www.barcodinglife.com), onde
um algoritmo procura trechos de similaridade local entre as sequências amostradas
e os resultados podem ser visualizados em ordem decrescente, das espécies já
identificadas, onde foi possível a comparação dos genótipos das plantas usadas
neste trabalho.
O modelo de substituição de nucleotídeos estabelecido para quantificar a
divergência entre indivíduos é o K2P, pois é o modelo mais indicado quando as
distâncias genéticas são pequenas (NEI e KUMAR, 2000). Os padrões de
divergência sugeridos pelas distâncias intra e interespecíficas foram graficamente
representados usando um dendrograma de Neighbor-Joining (NJ) com 1000
réplicas de bootstrap. As análises foram realizadas utilizando o programa MEGA 6
(TAMURA et al. 2013).
MARCADORES RAPD
Para as análises de diversidade genética, entre os acessos foi utilizado o
marcador molecular RAPD (Polimorfismos de DNA amplificados ao acaso), que se
baseia na amplificação arbitrária simultânea de fragmentos de DNA. Os 25 primers
utilizados e as suas respectivas sequências estão descritos na Tabela 2.

39
Tabela 2. Lista dos primers utilizados com suas respectivas sequências de bases.
Primer Sequência 5’ 3’ Primer Sequência 5’ 3’
OP 81 GGA GCG TAC T OPC 08 TGG ACC GGT G
OP 93 GTG CCG CACT OPD 01 ACC GCG AAG G
OP 95 GTG ACC AGA G OPD 08 GTG TGC CCC A
OPA 01 CAG GCC CTT C OPE 03 CCA GAT GCA C
OPA 02 TGC CGA GCT G OPE 06 AAG ACC CCT C
OPA 04 AAT CGG GCT G OPF 05 CCG AAT TCC C
OPA 05 AGG GGT CTT G OPM 05 GGG AAC GTG T
OPA 07 GAA ACG GGT G OPP 14 CCA GCC GAA C
OPA 17 GAC CGC TTG T OPAB 01 CCG TCG GTA G
OPA 18 AGG TGA CCG T OPAF 05 CCC GAT CAG A
OPA 20 GTT GCG ATC C OPAL 03 CCC ACC CTT G
OPC 04 CCG CAT CTA C OPBA 02 TGC TCGGCT C
OPC 07 GTC CCG ACG A - -
As reações de polimerase em cadeia (PCR) foram preparadas em volume de
25μ L contendo tampão de PCR 1X, 3mM MgCl2, 0,1mM de cada dNTP, 0,2μM
de primer, 30ng de DNA genômico e 1U/uL de TaqDNA polimerase (Invitrogen). A
amplificação foi realizada em termociclador PTC-100 (MJ Research) programado
como se segue: desnaturação inicial de 5min a 94ºC, seguido de 40 ciclos de 1min a
92ºC, 1min e 30 sec. a 37ºC, 1min e 30 sec. a 72ºC, com extensão final de 6min a
72ºC. Os produtos amplificados foram analisados em gel de agarose 1,0%. A
eletroforese foi realizada em cuba horizontal (Bio Rad®) com tampão TBE 1X a 100
Volts por duas horas, através de fonte Power Pac HV (Bio Rad). O gel foi corado
com brometo de etídio e foto documentado em grafado sob luz UV em sistema
digital L.Pix Image (Loccus Biotechnology).
Todas as etapas de preparação dos reagentes foram realizadas em capela
específica para PCR com prévia utilização de luz ultravioleta durante 15 minutos.
ANÁLISE DOS DADOS OBTIDOS ATRAVÉS DE MARCADORES RAPD
Com os géis obtidos gerou-se uma matriz binária em que os acessos foram
genotipados quanto à presença (1) e ausência (0) de bandas, com a matriz binária

40
calculou-se a porcentagem de polimorfismo de cada primer utilizado através da
fórmula:
P = nbt
nbp
Sendo que:
P= porcentagem de polimorfismo (ou taxa de polimorfismo);
nbp=número de bandas polimórficas;
nbt=número de bandas total.
Com matriz binária calculou-se também o coeficiente de similaridade de
Jaccard entre todos os acessos conforme a formula abaixo:
Sij= a
a+b+c
Sendo que:
a = número de casos em que ocorre a presença da banda em os ambos acessos,
simultaneamente;
b = número de casos em que ocorre a presença da banda somente no indivíduo i;
c = número de casos em que ocorre a presença da banda somente no indivíduo j.
A análise de agrupamento foi realizada pelo método UPGMA (Unweighted
Pair-group Method with Arithmetical Average. Todas as análises foram rodadas no
programa R Studio, versão 0.98.1091.
RESULTADOS E DISCUSSÃO
O DNA extraído foi avaliado por eletroforese em gel de agarose,
demonstrando que o protocolo segundo Saghai-Marrof et al. (1984), utilizado nesta
pesquisa possibilitou a obtenção de um DNA genômico de boa quantidade e
qualidade. Isso foi possível devido à quantidade de reagentes (EDTA, PVP, 2-
Mercaptoetanol), que inibem a degradação do material genômico, inativando
enzimas específicas ou compostos secundários, promovendo a quelação de cátions

41
divalentes, tais como Mg2+ e Ca2+, mantendo o pH do tampão estável (Tris-HCl) e a
desproteinização gerada por diversos reagentes orgânicos utilizados nas etapas da
extração (Fenol, Clorofórmio, Alcool Isoamílico, Isopropanol), assim como a retirada
de RNA da amostra através do uso de RNAse (BONATO et al., 2002).
A utilização de todos os reagentes combinados em etapas determinadas da
extração, assim como a utilização do CTAB (Cationic hexadecyl trimethyl
ammonium bromide ou brometo de cetiltrimetilamônio), que faz com que
polissacarídeos e ácidos nucleicos possuam solubilidade diferenciada na presença
desse detergente, inibindo a ação de complexos polifenólicos que, segundo Couch
& Fritz (1990), aderem irreversivelmente ao DNA, inibindo a ação das DNA
polimerase e das endonucleases de restrição.
Os dados de quantificação do material genético dos quatro acessos
avaliados via espectrofotometria variou de 1,91 a 1,94 apresentando-se dentro dos
parâmetros desejados (Tabela 3). Segundo Barbosa (1998), um DNA puro tem
OD260/OD280 de 1,8 a 2,0 e uma razão menor que 1,8 indica possível
contaminação das amostras com proteínas e/ou outras substâncias em grande
quantidade na amostra, enquanto a razão maior que 2,0 indica que as amostras
devem estar contaminadas com compostos orgânicos.
Tabela 3. Dados de quantificação via espectrofotometria das extrações de material genético foliar de quatro acessos de araruta. A absorbância (Abs) foi medida no comprimento de onda de 260 a 280 nanômetros
Acesso Concentração
ng/µL
Qualidade
Abs: 260/280
Araruta Comum 3366 1.94
Araruta Seta 1961 1.91
Araruta Guadalupe 1375 1.94
Araruta SC 1936 1.92
A extração de DNA é apenas uma etapa que precede as técnicas aplicadas à
PCR, como a RAPD. A aplicação de um material genético de boa qualidade
empregado na técnica de PCR diminui as chances de alterações de resultados ou
inibição da mesma, ocasionado por algum contaminante que não tenha sido
removido no momento da extração.

42
As amplificações do gene matK e rbcL dos diferentes acesso de araruta
geraram fragmentos de 2000 pb e 500pb respectivamente, e não foi possível
verificar variação nos padrões de bandas amplificados entre os acessos.
Os produtos amplificados foram sequenciados em ambas as direções e estes
foram alinhados. As sequências e os fragmentos com baixa qualidade foram
eliminados manualmente, objetivando minimizar os erros.
O sequenciamento do gene matK, de todos os acessos estudados, produziu
sequências muito pequenas, e o acesso Seta foi excluído devido a má qualidade da
mesma. A falha de amplificação por PCR do matK em alguns grupos taxonômicos
foi relatada em alguns estudos, pois os iniciadores projetados dentro da região de
matK podem ser problemáticos para a amplificação de PCR e sequenciamento,
devido a ampla variabilidade na região do matK, o que leva à concepção de novos
primers taxon-específicos ou modificações dos iniciadores existentes (WOLF et al.,
1987; YU, et al., 2011). No entanto as sequências do gene matK, foram as que
obtiveram desempenho menos satisfatório no sequenciamento.
O matK é bastante útil em reconstruções filogenéticas em níveis taxonômicos
como a ordem ou a família, mas tem sido utilizado igualmente em níveis, como o
gênero ou a espécie (CHASE et al., 2007; LAHAYE et al., 2008), porém, por serem
muito similares, não houve distinção em nenhum dos níveis taxonômicos citados
acima para os acessos de Maranta arundinacea.
A região amplificada correspondente ao gene matK dos quatro acessos
(Comum, SC, Guadalupe e Seta), obteve-se um valor máximo de identidade de
99,66%, quando submetidas no programa, BOLD Systems. Houve uma diferença
quando a comparação ocorreu através do BLAST, onde as sequências deram um
valor de 100% de identidade entre os acessos. Estes resultados só confirmam que
os quatro acessos testados todos são da espécie M. arundinacea.
Já as sequências do gene rbcL, apresentaram boa qualidade no
sequenciamento. Os acessos Comum e JQ592613.1 (Genbank), apresentaram
maior similaridade entre si, diferenciando-se igualmente dos acessos SC,
Guadalupe e Seta. A partir do alinhamento das sequências foi possível gerar um
dendograma (Figura 2), possibilitando a construção de um grupo com subgrupo e o
outgroup. O subgrupo é constituído pelos acessos Comum e rbcL (JQ592613.1) de
M. arundinacea o valor de bootstrap, do ramo ficou em 89%, considerado
aceitável em relação a medida de confiabilidade dos ramos gerados pelas análises

43
filogenéticas, que estima o grau de suporte de cada ramo entre os acessos.
Entretanto, o ramo constituído pelos acessos SC, Guadalupe, Seta e rcbL
(3845927560) de M.arundinacea, retirado do GenbanK, por serem muito similares,
obtiveram valores < 50%, pois o bootstrap analisa a partir da criação de vários
conjuntos de sequências nas quais são escolhidas randomicamente colunas do
alinhamento múltiplo e são geradas novas análises para cada um dos novos
conjuntos gerados (LEPAGE,1992).
As diferenças de similaridade encontradas, neste estudo, provavelmente são
relacionadas, à distribuição geográfica dos diferentes acessos, ocorrendo
possivelmente alterações nas frequências alélicas adaptadas para as condições
ambientais ou simplesmente deriva genética.
Segundo Soltis e Soltis (1998), a maioria dos estudos filogenéticos sugere
que o gene rbcL é o mais adequado para reconstruir as relações em nível de gênero
entretanto, pouco utilizado em níveis específicos. Observou-se neste trabalho que a
região do gene rbcL foi facilmente amplificada e sequenciada para os acessos de M.
arundinacea, apresentando valores máximos de identidade e poder de
discriminação interespecífico satisfatórios.
Comparando - se o acesso Comum de M. arundinacea com as demais,
ocorreu diferença apenas por algumas regiões que apresentaram nucleotídeos
divergentes (Apêndice 1). O achado pode ajudar pesquisadores a distinguir plantas
distintas, mas que aparentemente pareçam estar relacionadas entre si.
As sequências obtidas do gene rbcL, foram analisadas e comparadas com
as sequências depositadas nos bancos de dados do BOLD Systems (Apêndice 1) e
os resultados apresentaram 100% de identidade dos quatro acessos testados. O
mesmo aconteceu quando as sequências foram submetidas à busca na base de
dados genômica BLAST. A região do gene matK, apesar do sequenciamento menos
satisfatório, apresentou valores de identidade através da busca BOLD Systems e
BLAST similares ao gene rcbL, ambos comprovam que os acessos são da espécie
M. arundinacea.
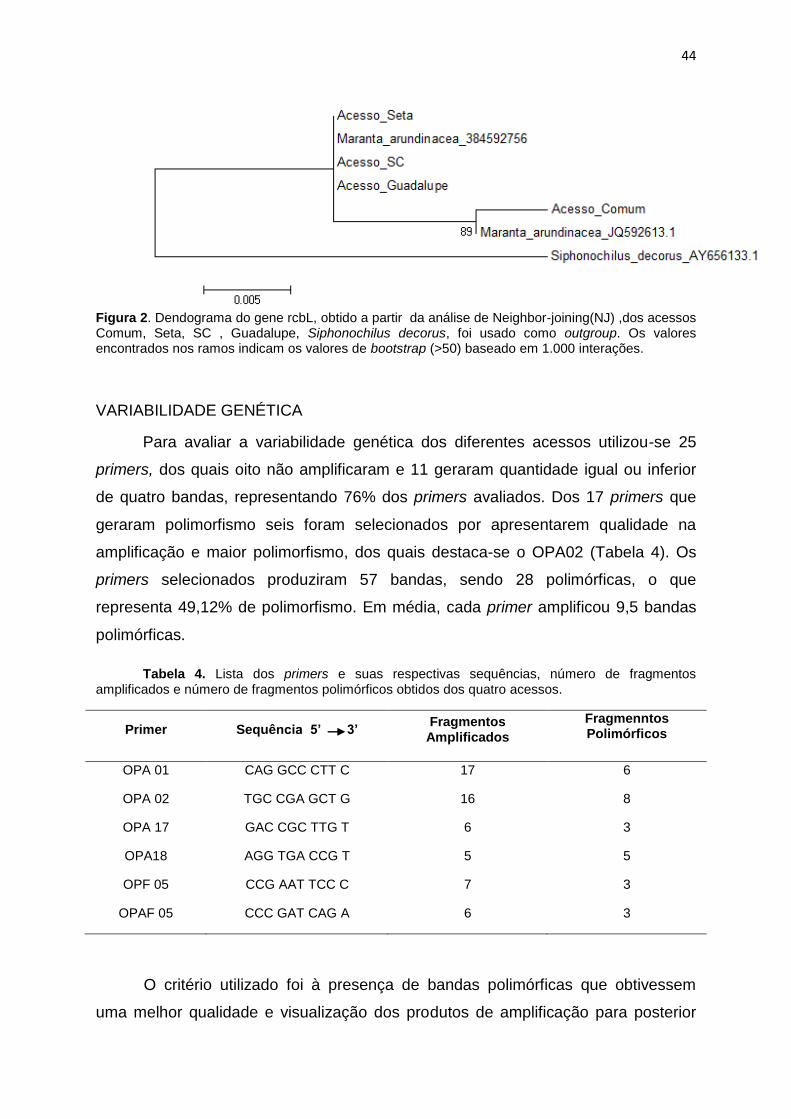
44
Figura 2. Dendograma do gene rcbL, obtido a partir da análise de Neighbor-joining(NJ) ,dos acessos Comum, Seta, SC , Guadalupe, Siphonochilus decorus, foi usado como outgroup. Os valores encontrados nos ramos indicam os valores de bootstrap (>50) baseado em 1.000 interações.
VARIABILIDADE GENÉTICA
Para avaliar a variabilidade genética dos diferentes acessos utilizou-se 25
primers, dos quais oito não amplificaram e 11 geraram quantidade igual ou inferior
de quatro bandas, representando 76% dos primers avaliados. Dos 17 primers que
geraram polimorfismo seis foram selecionados por apresentarem qualidade na
amplificação e maior polimorfismo, dos quais destaca-se o OPA02 (Tabela 4). Os
primers selecionados produziram 57 bandas, sendo 28 polimórficas, o que
representa 49,12% de polimorfismo. Em média, cada primer amplificou 9,5 bandas
polimórficas.
Tabela 4. Lista dos primers e suas respectivas sequências, número de fragmentos amplificados e número de fragmentos polimórficos obtidos dos quatro acessos.
Primer Sequência 5’ 3’ Fragmentos Amplificados
Fragmenntos Polimórficos
OPA 01 CAG GCC CTT C 17 6
OPA 02 TGC CGA GCT G 16 8
OPA 17 GAC CGC TTG T 6 3
OPA18 AGG TGA CCG T 5 5
OPF 05 CCG AAT TCC C 7 3
OPAF 05 CCC GAT CAG A 6 3
O critério utilizado foi à presença de bandas polimórficas que obtivessem
uma melhor qualidade e visualização dos produtos de amplificação para posterior

45
análise. No resultado da RAPD pode-se observar que os acessos SC, Guadalupe e
Seta, apresentaram similaridade genética pelo padrão de bandas evidenciado entre
esses genótipos, no entanto, o acesso denominado Comum se diferenciou por
apresentar bandas com padrões diferentes dos demais acessos.
Por meio do dendrograma formado (Figura 3), verificou-se a formação de
dois grupos em que o acesso denominado Comum, ficou posicionado em um ramo,
enquanto que os outros acessos (SC, Guadalupe e Seta), ficaram agrupados em
outro ramo. Também foi observado que os acessos SC, Guadalupe e Seta podem
ser considerados geneticamente iguais mediantes aos primers testados na análise
realizada pela técnica de RAPD.
Figura 3. Dendrograma de similaridade genética, obtido a partir dos produtos de amplificação por RAPD do primer OPA 02 para quatro acessos de araruta.
Com os resultados obtidos infere-se que o acesso Comum foi o mais
divergente, se diferenciando dos acessos SC, Guadalupe e Seta, supondo que uma

46
mudança evolutiva devido à distância geográfica, que segundo Slatkin (1987),
eventualmente restringe o processo evolutivo por prevenir a adaptação às
condições locais e promover a evolução espalhando novos genes e combinações
gênicas entre populações de uma determinada espécie. Esses acessos de Araruta
podem apresentar variabilidade genética ocasionada por mistura de sementes de
outras populações, por mutações genéticas ou cruzamentos naturais com plantas
de diferentes genótipos.
CONCLUSÃO
Observou-se através das análises de DNA barcoding, que todos os acessos
testados são da espécie M.arundinacea e através da técnica de RAPD conclui-se
que há divergência populacional entre o acesso Comum e os demais acessos.
O presente trabalho envolvendo os quatro acessos de araruta Comum, SC,
Guadalupe e Seta, nas técnicas apresentadas é pioneiro e possibilita a elaboração
de novos estudos para que no futuro ocorra a probabilidade da descoberta de genes
promissores, visando planejar os cruzamentos através do programa de
melhoramento genético de forma a aumentar as diversidades genéticas entre
genótipos de interesse.

47
REFERÊNCIAS BIBLIOGRÁFICAS
ANDERSSON L.. Revision of Maranta subgen. Maranta (Marantaceae). Nordic Journal of Botany,v. 6, n.6, p.729–756, 1986.
ARNS, Y.K.N. Estudo Taxonômico dos grupos Monotagma, Maranta e Myrosma (Marantaceae) em Pernambuco. 1997.105p. Dissertação de Mestrado – Biologia Vegetal. Universidade Federal de Pernambuco, Pernambuco,1997.
BARBOSA, M. M. Quantificação e controle da qualidade do DNA genômico. In: MILACH, S.ed. Marcadores moleculares em plantas. Porto Alegre, 1998. p.99-106.
BONATO, A. L. V.; VERZIGNASSI,J. R.;RESENDE, R.M.S.; FERNANDES,C.D.; LEGUIZAMON, G. O. de C. Extração de DNA genômico de Stylosanthes spp. Campo Grande: Embrapa Gado de Corte, 2002. 4p. (Comunicado técnico, 78).
CHASE, M.W.; COWAN, R.S.; HOLLINGSWORTH, P.M.; BERG, C.; MADRINAN, S.; PETERSEN, G.;SEBERG, O.; JORGSENSEN, T.; CAMERO, K.M.; CARINE, M.; PEDERSEN, N.; HEDDERSON, T.A.J.;CONRAD, F.;SALAZAR, G.A.; RICHARDSON, J.E.;HOLLINGSWORTH, M.L.; BARRACLOUGH, T.G.; KELLY,L. ; WILKINSON, M. A proposal for a standardized protocol to barcode all land plants. Taxon 56, v.2, n.2, p. 295–299, 2007.
COUCH, J. A; FRITZ, P. J. Isolation of DNA from plants high in polyphenolics. Plant Molecular Biology Reporter, v.8, p.8-12, 1990.
FARALDO, M.I.F. Marcadores moleculares em mandioca. In: CEREDA, M.P. Agricultura: Tuberosas Amiláceas Latino Americano, São Paulo: Fundação Cargill, 2003. v.2, p.101-117. FUKUDA, W. M. G.; CAVALCANTI J.; COSTA I.R.S. . Variabilidade genética e melhoramento da mandioca (Manihot esculenta Crantz). Recursos Genéticos e Melhoramento de Plantas para o Nordeste Brasileiro. 1994. Disponível em: <http://www.cpatsa.embrapa.br/catalogo/livrorg/mandioca.pdf>. Acesso: 03 de novembro de 2014. HALL, T. A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series: Oxford Journals, v. 41, p. 95-98, 1999.
HERBERT, P.D.N; RATNASINGHAM, S.; DeWAARD J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. The Royal Society, v.270, n.1, p. 96–99, 2003.

48
KENNEDY, H. Systematics and pollination of the ‘closedflowered’ species of Calathea (Marantaceae). University of California Publications in Botany, 1977. v.71, p.1–90. KENNEDY, H. New species of Calathea (Marantaceae) endemic to Costa Rica. Canadian Journal of Botany, v.75, n.8, p.1356-1362, 1997. KENNEDY, H. Diversification in pollination mechanisms in the Marantaceae. In: Wilson, KL, Morrisson DA, (ed). Monocots – Systematics and evolution. Melbourne: CSIRO, 2000, p.335–343. KIMURA, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, v.16, p.111-120, 1980. LAHAYE, R.; van der BANK, M.; BOGARIN, D.; WARNER, J.; PUPULIN, F.; GIGOT, G.; MAURIN, O.; DUTHOIT, S.; BARRACLOUGH, T. G.; SAVOLAINEN, V. DNA Barcoding the floras of biodiversity hotspots. Proceedings of the National Academy of Sciences, v. 105, n. 8, p. 2923-2928, 2008. LEPAGE, R.; BILLARD, L. Exploring the limits of bootstrap. New York: John Wiley and Sons, 1992. p.632. LIU, K.J.; MUSE, S.V. PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics, v.21, n.9, p. 2128-2129, 2005.
MENEZES, C.C.E. et al. Análise da pureza genética e discriminação de cultvares de vinca (Catharanthus roseus(L.) G. Don) usando “Randon Amplified Polymorphic DNA” em DNA extraído de sementes e folhas. Revista Brasileira de Sementes, v. 24, n. 1, p. 279-285, 2002. MONTEIRO, D. A.; PERESSIN, V. A. Cultura da araruta. In:CEREDA, M. P. (Coord.). Agricultura: tuberosas amiláceas Latino Americanas. São Paulo: Fundação Cargill, 2002, v. 2, p. 440-447.
NASS, L.L. Utilização de recursos genéticos vegetais no melhoramento. In: NASS,L.L.; VALOIS, A.C.C.; VALADARES- INGLIS, M.C.(Ed.). Recursos genéticos e melhoramento, 2001. Porto Alegre: Gráfica Editora Pallotti, Fundação de Apoio à Pesquisa de Mato Grosso, Cap.2, p 29-55. NEI M. e KUMAR S. Molecular Evolution and Phylogenetics. Oxford University Press, 2000.
NEVES, M. C. P.; COELHO, I. S.; ALMEIDA, D. L. Araruta; resgate de um cultivo tradicional. Rio de Janeiro: Seropédica – RJ: EMBRAPA, 2005, 4p (Seropédica – RJ: EMBRAPA Comunicado técnico 79).

49
RStudio (2012). RStudio: Integrated development environment for R (Version 0.98.1091 – © 2009-2014 RStudio, Inc). Computer software. Boston, MA., 2012.
SAGHAI-MAROOF, M.A.; SOLIMAN, K.M.; JORGENSEN, R.A.; ALLARD, R.W. Ribosomal DNA spacer length polymorphism in barley: Mendelian inheritance, chromosome location and population dynamics. Proceedings of the National Academy of Sciences of the U.S.A, v.89, n.2, p.1477-1481, 1984.
SAITOU N. e NEI M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, v. 4, n.4, p.406-425, 1987.
. SLATKIN, M. Gene Flow and the Geographic Structure of Natural Populations. Science, v. 236, n.4803, p.787-792, 1987.
SOLTIS D.E. e SOLTIS P.S. Choosing an approach and an appropriate gene for phylogenetic analysis. In: Soltis D.E.; Soltis P.M.; Doyle J.J. (ed) Molecular systematics of plants. II. DNA sequencing. Kluwer Academic Publishers, Dordrecht, 1998, p. 21–24 SOUZA, V. C.; LORENZI, H. Botânica Sistemática: guia ilustrado para identificação das famílias de Angiospermas da flora brasileira, baseado em APG II. Nova Odessa, Instituto Plantarum, 2005. 640p.
. TAUTZ, D.; ARCTANDER, P.; MINELLI, A.; THOMAS, R. H.; VOGLER, A. P. Aplea for DNA taxonomy. Trends in Ecology e Evolution, v.18, n.2, p.70–74, 2003. TAMURA K.; STECHER G.; PETERSON D.; FILIPSKI A.; and KUMAR S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0.Molecular Biology and Evolution, v.30, p. 2725-2729, 2013.
VIEIRA, S. e SOUZA, V.C. Four new species of Maranta L. (Marantaceae) from Brazil. Botanical Journal of the Linnean Society, v.158, n.1, p.131–139, 2008.
WATERHOUSE, A.M.; PROCTER, J.B.; MARTIN, D.M.A; CLAMP, M. and BARTON, G. J. "Jalview Version 2 - a multiple sequence alignment editor and analysis workbench" Bioinformatics, v.25 n.9 p.1189-1191,2009. WEEDEN, N.F. Inheritance and reliability of RAPD markers. In: SYMPOSIUM ON APPLICATIONS OF RAPD TECHNOLOGY TO PLANT BREEDING, 1.,1992, Minneapolis. Anais. Minneapolis: Crop Science Society of America/American Society for Horticultural Science/American Genetic Association, 1992. p. 12.

50
WILLIAMS, J.G.K.; KUBELIK, A.R.; LIAK, K.J.; RAFALSKI, J.A.; TINGEY, S.V. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research, v.18, n.22, p.6531-6535, 1990. WOLFE, K.H.; LI, W.H. e SHARP, P.M. (1987) Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast and nuclear DNAs. Proceedings of the National Academy of Sciences of the USA, v.84, n.24, p. 9054–9058, 1987. YU, J.; XUE, J. H.; ZHOU, S. L. New universal matK primers for DNA Barcoding angiosperms. Journal of Systematics and Evolution, v.49, n.3, p.176-181, 2011.
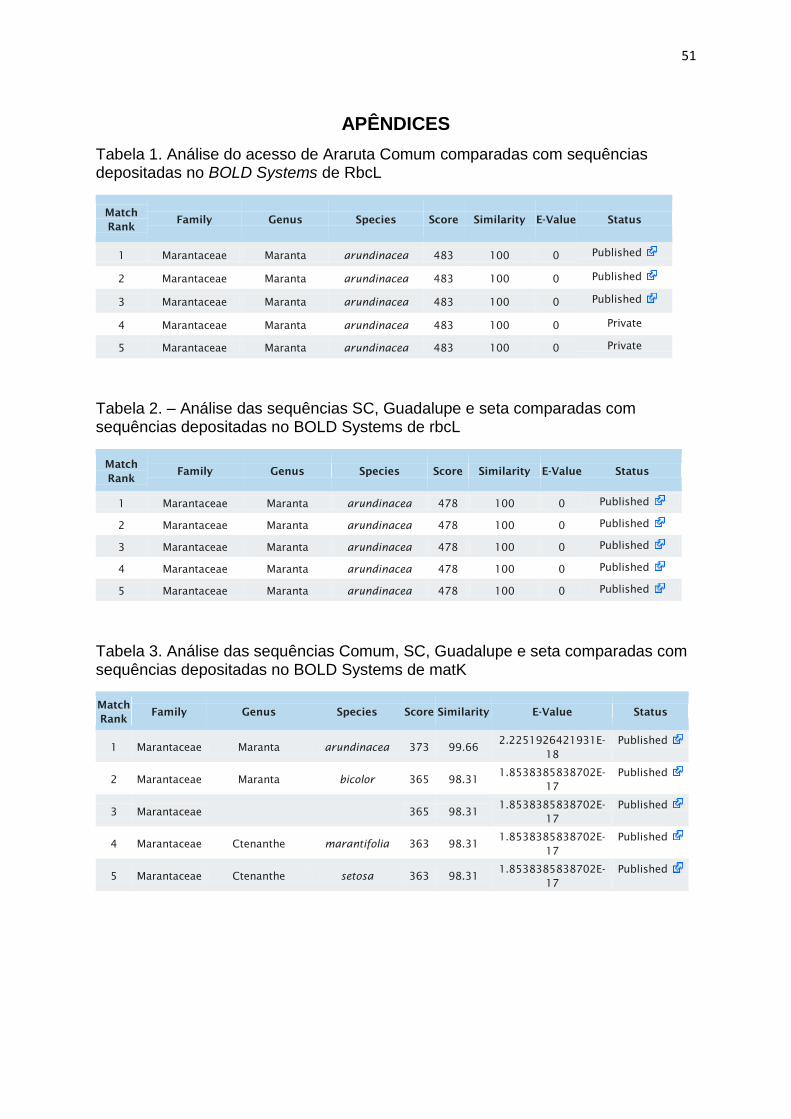
51
APÊNDICES
Tabela 1. Análise do acesso de Araruta Comum comparadas com sequências depositadas no BOLD Systems de RbcL
Match
Rank Family Genus Species Score Similarity E-Value Status
1 Marantaceae Maranta arundinacea 483 100 0 Published
2 Marantaceae Maranta arundinacea 483 100 0 Published
3 Marantaceae Maranta arundinacea 483 100 0 Published
4 Marantaceae Maranta arundinacea 483 100 0 Private
5 Marantaceae Maranta arundinacea 483 100 0 Private
Tabela 2. – Análise das sequências SC, Guadalupe e seta comparadas com sequências depositadas no BOLD Systems de rbcL
Match
Rank Family Genus Species Score Similarity E-Value Status
1 Marantaceae Maranta arundinacea 478 100 0 Published
2 Marantaceae Maranta arundinacea 478 100 0 Published
3 Marantaceae Maranta arundinacea 478 100 0 Published
4 Marantaceae Maranta arundinacea 478 100 0 Published
5 Marantaceae Maranta arundinacea 478 100 0 Published
Tabela 3. Análise das sequências Comum, SC, Guadalupe e seta comparadas com sequências depositadas no BOLD Systems de matK
Match
Rank Family Genus Species Score Similarity E-Value Status
1 Marantaceae Maranta arundinacea 373 99.66 2.2251926421931E-
18
Published
2 Marantaceae Maranta bicolor 365 98.31 1.8538385838702E-
17
Published
3 Marantaceae
365 98.31 1.8538385838702E-
17
Published
4 Marantaceae Ctenanthe marantifolia 363 98.31 1.8538385838702E-
17
Published
5 Marantaceae Ctenanthe setosa 363 98.31 1.8538385838702E-
17
Published
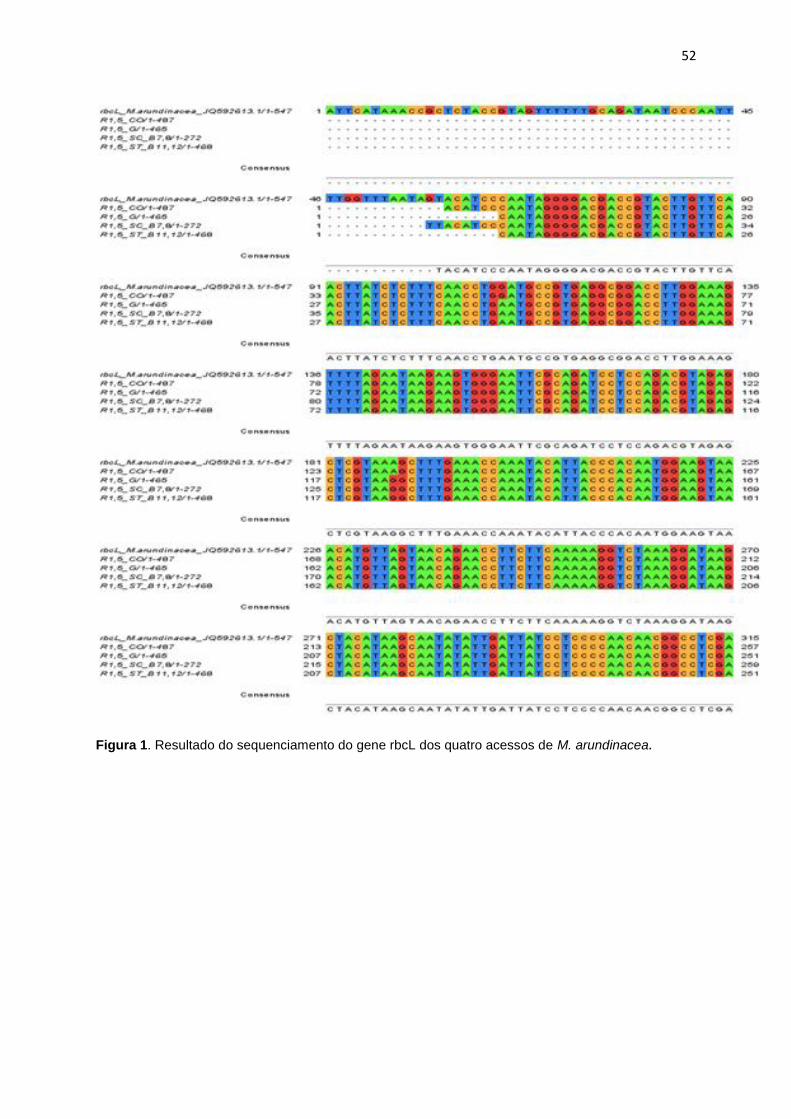
52
Figura 1. Resultado do sequenciamento do gene rbcL dos quatro acessos de M. arundinacea.

53
Continuação -Figura 1. Resultado do sequenciamento do gene rbcL dos quatro acessos de M.
arundinacea.

54
Figura 2. Resultado do sequenciamento do gene matK, dos quatro acessos de M. arundinacea.
.

55
Continuação- Figura 2. Resultado do sequenciamento do gene matK, dos quatro acessos de M.
arundinacea.
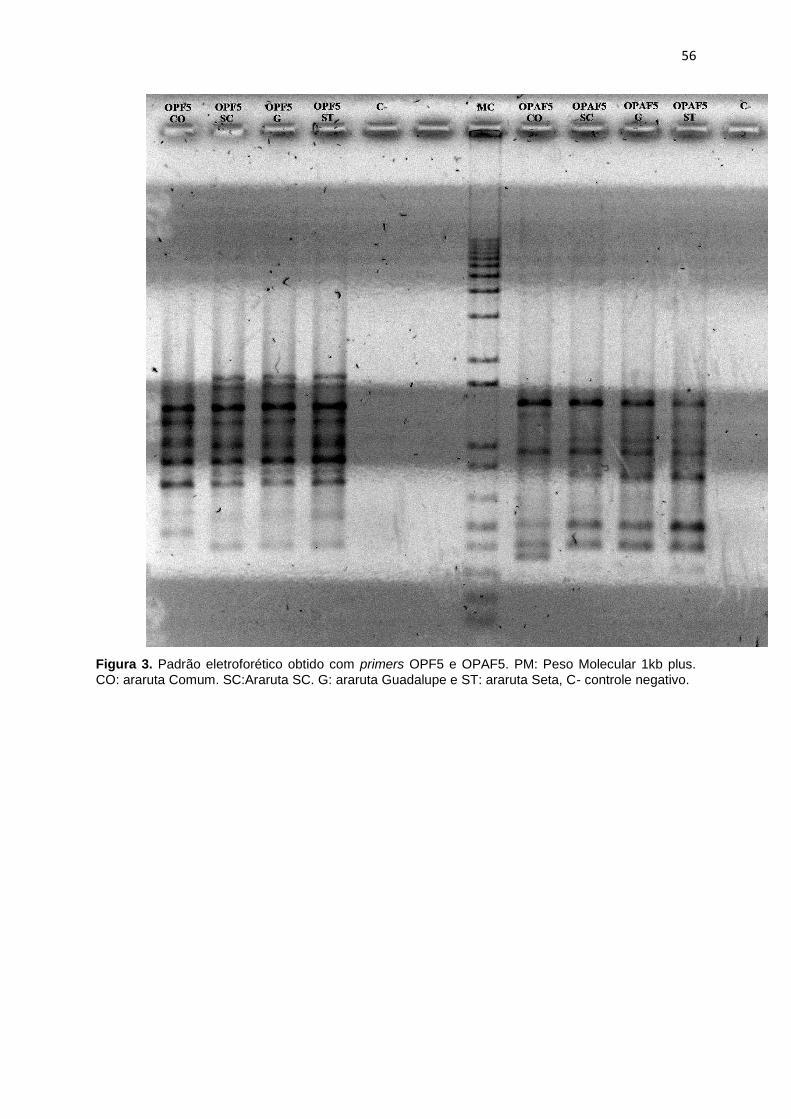
56
Figura 3. Padrão eletroforético obtido com primers OPF5 e OPAF5. PM: Peso Molecular 1kb plus.
CO: araruta Comum. SC:Araruta SC. G: araruta Guadalupe e ST: araruta Seta, C- controle negativo.
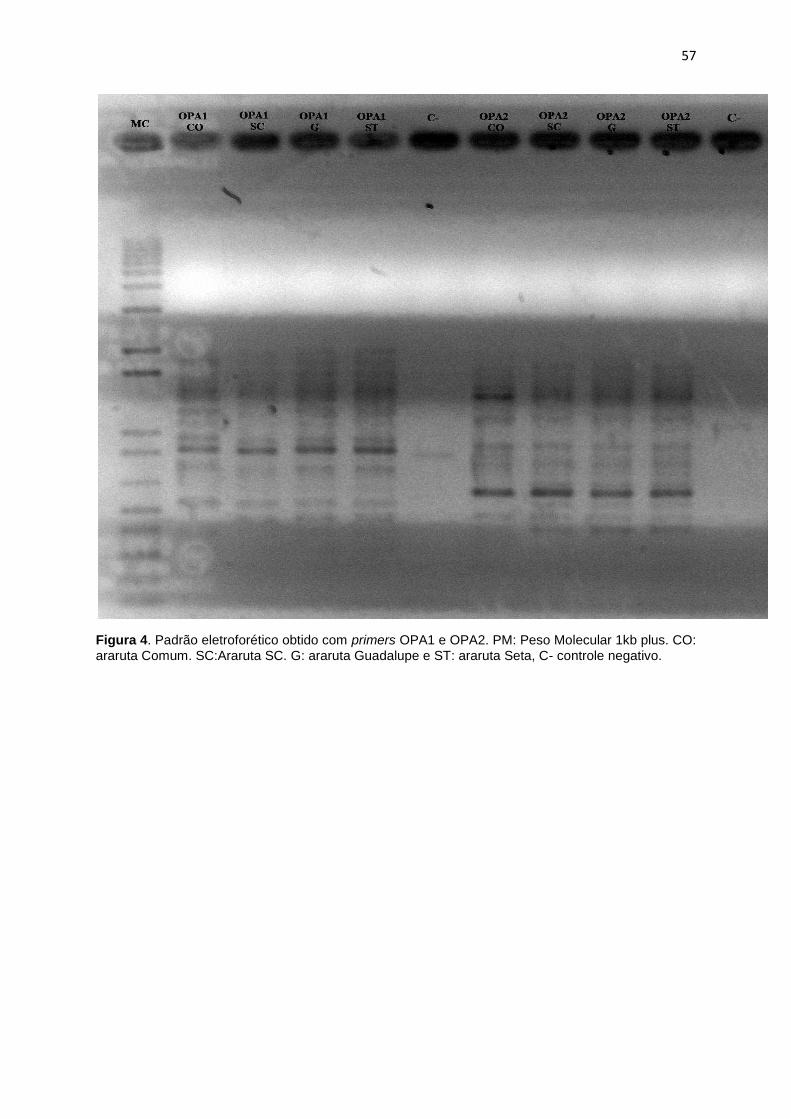
57
Figura 4. Padrão eletroforético obtido com primers OPA1 e OPA2. PM: Peso Molecular 1kb plus. CO:
araruta Comum. SC:Araruta SC. G: araruta Guadalupe e ST: araruta Seta, C- controle negativo.
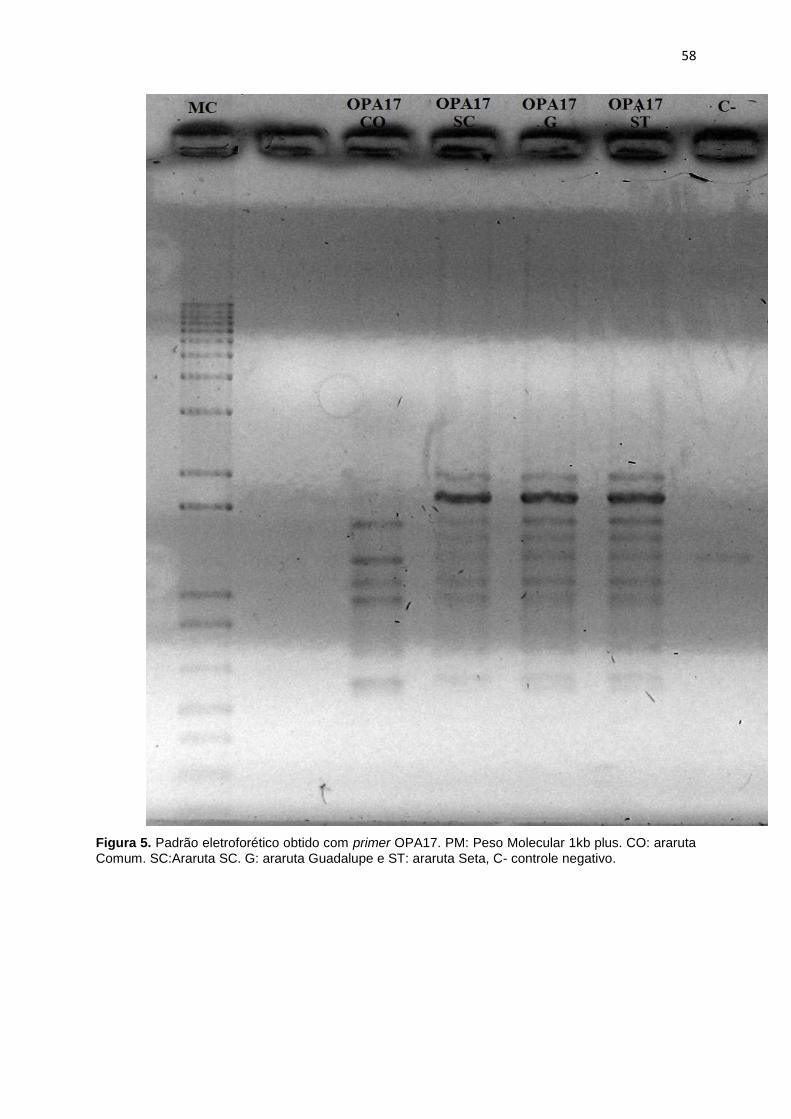
58
Figura 5. Padrão eletroforético obtido com primer OPA17. PM: Peso Molecular 1kb plus. CO: araruta
Comum. SC:Araruta SC. G: araruta Guadalupe e ST: araruta Seta, C- controle negativo.







