diversidade, densidade e distribuiÇÃo altitudinal...
TRANSCRIPT

Ricardo Jannini Sawaya
DIVERSIDADE, DENSIDADE E DISTRIBUIÇÃO ALTITUDINAL
DA ANUROFAUNA DE SERAPILHEIRA
DA ILHA DE SÃO SEBASTIÃO, SP
São Paulo
1999

Ricardo Jannini Sawaya
DIVERSIDADE, DENSIDADE E DISTRIBUIÇÃO ALTITUDINAL
DA ANUROFAUNA DE SERAPILHEIRA
DA ILHA DE SÃO SEBASTIÃO, SP
Dissertação apresentada ao Instituto de Biociências da Universidade de São Paulo, para a obtenção de Título de Mestre em Ciências, na Área de Zoologia. Orientador: Ivan Sazima
São Paulo
1999

Sawaya, Ricardo Diversidade, densidade e distribuição altitudinal da anurofauna de serapilheira da Ilha de São Sebastião, SP 65 páginas Dissertação (Mestrado) - Instituto de Biociências da Universidade de São Paulo. Departamento de Zoologia. 1. diversidade 2. densidade 3. anurofauna I. Universidade de São Paulo. Instituto de Biociências. Departamento de Zoologia.
Comissão Julgadora:
Marcio R. C. Martins________ Otávio A. V. Marques________ Prof. Dr. Prof. Dr.
Miguel U. T. Rodrigues____ Prof. Dr. p/ Orientador

Aos meus pais,
por tudo.
Ao meu avô, Paulo,
pelo exemplo de uma vida dedicada à biologia.

“The biosphere is an intricate tapestry of interwoven life forms.”
E. O. Wilson, 1988

6
AGRADECIMENTOS
Este trabalho só foi possível a partir da ajuda de diversas pessoas. Àquelas que não
me recordo agora peço desculpas e agradeço da mesma forma.
Agradeço sinceramente:
a Glauco Machado, Renato Nunes, Carlos Henrique Vasconcelos, Luciano dos
Anjos e Angela Midori, pela amizade, profissionalismo e por formar a heróica e divertida
equipe que dividiu comigo os bons e maus momentos dos trabalhos de campo;
à querida Dani, por participar ativamente em todas as fases do trabalho, desde a
concepção do projeto até a conclusão da dissertação, por segurar todas as minhas barras
nos momentos mais difíceis e por estar sempre ao meu lado;
à Veridiana Vaccarelli, pela amizade e por integrar a equipe no momento em que
mais precisamos;
a Márcio Martins, pela amizade, orientação atuante em todas as fases do projeto e
por dividir comigo sua experiência de trabalho;
a Ivan Sazima, pela orientação e ajuda na fase final da dissertação;
a Ariovaldo Giaretta, por dividir comigo seus conhecimentos sobre anuros e
experiência de campo ao longo dos últimos anos, e pela ajuda na fase final da dissertação;
à Kátia Facure, pela ajuda na análise multivariada;
a José Pombal Jr., pela ajuda na taxonomia;
a Célio Haddad, pela ajuda na taxonomia;
à Fernanda Oliveira e Sandra Baptista pela ajuda na fase final da dissertação;

7
à Cristiane Leonel, por sempre apoiar o trabalho, desde a fase de coleta preliminar
de dados, pelo profissionalismo e exemplo de como dirigir um parque;
a Vini, Marquinhos, Hélio, Beto e demais empregados do Parque Estadual de
Ilhabela, que tornaram o trabalho de campo possível;
ao Instituto Florestal, pela hospedagem durante a coleta de dados, por testar minha
paciência e por me ensinar a lidar com as situações mais adversas;
a Vini, Daniel e Ildefonso, pelo companheirismo, por me ensinar muito sobre a Ilha
de São Sebastião e pela ajuda na “descoberta” e abertura de trilhas;
a Renato Nunes, Márcio Araújo, Glauco Machado, Angela Midori, Hermes
Fonseca, Sónia Andrade, Juliana Félix, Cristiana Damiano, Rodrigo Ribeiro, Ito, Rafael
Raimundo, Frank Barrientos e Joel Meyer, pela ajuda nos trabalhos de campo da coleta
preliminar de dados;
a Glauco Machado, Márcio Araújo, Hermes Fonseca e Renato Nunes pela amizade
e inúmeras discussões frutíferas ao longo de todo o trabalho;
à Vera Jannini e Rogério Sawaya, por todo o apoio, mesmo distantes da Ilha de São
Sebastião;
aos hotéis Petit Village e Devissé, pela hospedagem durante a coleta preliminar de
dados.

8
ÍNDICE
I. ÍNDICE DE TABELAS E FIGURAS............................................................................................... 10
II. INTRODUÇÃO............................................................................................................................. 11 III. OBJETIVOS................................................................................................................................ 14 IV. MATERIAL E MÉTODOS........................................................................................................... 15
1. Área de estudo............................................................................................................... 15
2. Coleta de dados............................................................................................................. 17
3. Análise de dados........................................................................................................... 23
4. Comentários taxonômicos........................................................................................... 25
5. Espécies regulares de serapilheira............................................................................... 27 V. RESULTADOS.............................................................................................................................. 29
1. Riqueza específica......................................................................................................... 29
2. Abundância relativa de espécies e dominância......................................................... 36
3. Densidade....................................................................................................................... 40
4. Relação entre a anurofauna de serapilheira e as variáveis ambientais................... 44
5. Modificação do método: uso de redes....................................................................... 46 VI. DISCUSSÃO................................................................................................................................ 47
1. Riqueza específica......................................................................................................... 47
2. Abundância relativa de espécies e dominância......................................................... 50
3. Densidade....................................................................................................................... 51
4. Relação entre a anurofauna de serapilheira e as variáveis ambientais................... 53
5. Comparação com outras localidades da Mata Atlântica......................................... 54
6. Método das parcelas amplas de 8 x 8 m.................................................................... 54 VII. CONCLUSÕES........................................................................................................................... 56 VIII. RESUMO.................................................................................................................................. 58

9
IX. ABSTRACT.................................................................................................................................. 60 X. APÊNDICE 1: Lagartos e serpentes encontrados nas parcelas............................................ 62 XI. REFERÊNCIAS BIBLIOGRÁFICAS............................................................................................. 63

10
ÍNDICE DE TABELAS E FIGURAS
Figura 1: Imagem de satélite da Ilha de São Sebastião.............................................................. 16 Figura 2: Aplicação do método das parcelas amplas de 8 x 8 m............................................. 20 Figura 3: Espécies de anuros de serapilheira da Ilha de São Sebastião.................................. 31 Figura 4: Curva média do coletor................................................................................................. 34 Figura 5: Riqueza específica ao longo do gradiente altitudinal................................................ 35 Figura 6: Avaliação da distribuição altitudinal das espécies..................................................... 37 Figura 7: Abundância relativa de espécies.................................................................................. 38 Figura 8: Espécies dominantes ao longo do gradiente altitudinal........................................... 39 Figura 9: Densidade de indivíduos por parcela ao longo do gradiente altitudinal................ 41 Figura 10: Densidade de indivíduos por parcela ao longo do gradiente altitudinal em cada transecto.............................................. 42
Figura 11: Densidade de indivíduos por parcela para as quatro espécies mais abundantes................................................................. 43
Figura 12: Relação entre os anuros de serapilheira e as variáveis ambientais, Análise de Correspondência Canônica (ACC).............. 45 Tabela 1: Pontos de amostragem, altitudes amostradas em cada transecto...................................................................... 18
Tabela 2: Lista de espécies de anuros de serapilheira da Ilha de São Sebastião, e coletadas nas 92 parcelas amostradas............................... 30
Tabela 3: Amostragens quantitativas de anuros de serapilheira na Mata Atlântica do Estado de São Paulo................................................................ 48

11
INTRODUÇÃO
O termo diversidade biológica inclui dois conceitos distintos: riqueza específica, ou
número de espécies, e heterogeneidade, ou abundância relativa de espécies (KREBS, 1989).
Diferentes índices têm sido propostos para expressar estes dois conceitos conjuntamente
(cf. MAGURRAN, 1988). Entretanto, vários autores discutem a relevância destes índices,
argumentando que medidas mais diretas de diversidade são mais compreensíveis (cf.
JANZEN, 1973; HURLBERT, 1971; SCOTT, 1976). MAY (1975) conclui que uma das medidas
de heterogeneidade mais satisfatórias disponíveis é a abundância da espécie dominante
(índice de Berger-Parker sensu MAGURRAN, 1988). Assim, a diversidade de espécies de uma
comunidade ou taxocenose pode ser medida pela riqueza específica e pela dominância da
espécie mais abundante expressa em porcentagem.
A comunidade científica tem reconhecido ultimamente a importância do estudo e
quantificação da diversidade biológica (cf. LOVEJOY, 1995), que está entre os objetivos
básicos da Estratégia Global para a Biodiversidade (WILSON, 1992). Estudos sobre
abundância e distribuição de espécies também têm sido enfatizados por fornecer
conhecimentos básicos para pesquisa nas áreas de ecologia, sistemática, biogeografia e
biologia da conservação (HEYER et al., 1994).
Existem diversas hipóteses para explicar o maior número de espécies existente nos
trópicos (cf. KREBS, 1985; PIANKA, 1966; RICKLEFS, 1990; PIANKA, 1994). A região
neotropical é a que possui a maior riqueza específica de anfíbios e répteis, sendo ideal para
o estudo dos padrões de diversidade destes grupos (CADLE & PATTON, 1988; DUELLMAN,
1988; HEYER, 1988; MARTINS, 1994). Entretanto, o uso de métodos padronizados é crítico
para comparações de dados coletados por diferentes pesquisadores em diferentes locais e
épocas (HEYER et al. 1994).

12
A serapilheira é a camada de folhas e outros detritos vegetais que se depositam
sobre o chão das florestas. A anurofauna associada à serapilheira em florestas tropicais é
caracterizada pela alta riqueza de espécies (e. g. HEYER et al., 1990; ZIMMERMAN &
RODRIGUES, 1990; VITT & CALDWELL, 1994). O método das parcelas amplas de 8 x 8 m
introduzido por LLOYD et al. (1968), e padronizado por JAEGER & INGER (1994) para a
amostragem de anuros de serapilheira, possibilitou a primeira comparação quantitativa das
faunas tropicais de vertebrados entre o velho e o novo mundo (SCOTT, 1976). Esse método
têm possibilitado o reconhecimento de padrões de diversidade e densidade de anuros de
serapilheira em diferentes localidades do planeta (veja revisões em INGER, 1980; ALLMON,
1991; GIARETTA, 1999). Estes padrões têm sido relacionados a fatores como altitude,
perturbação de florestas e variáveis ambientais e climáticas (SCOTT, 1976; INGER, 1980;
LIEBERMAN, 1986; FAUTH et al., 1989; HEINEN, 1992; RODRIGUEZ, 1992; GIARETTA et al.,
1997; GIARETTA, 1999; GIARETTA et al., no prelo; presente estudo). SCOTT (1976) compara
a herpetofauna de serapilheira de localidades da América Central e sudeste da Ásia,
sugerindo que haja diminuição no número de espécies com o aumento da altitude e
número de meses secos (pluviosidade menor do que 100 mm), e enfatiza a densidade de
indivíduos aproximadamente dez vezes maior na Costa Rica (América Central) em relação a
Bornéu (sudeste da Ásia). Esta grande diferença observada na densidade de indivíduos
entre localidades do novo e do velho mundo tem sido atribuída a fatores como
produtividade do ecossistema (SCOTT, 1976) ou composição de espécies vegetais (INGER,
1980), mas a questão é objeto de debates (MAY, 1980).
Embora anfíbios e escamados tenham sido analisados conjuntamente em alguns
estudos, a distinção entre os dois táxons deve ser considerada, pois parcelas amplas de 8 x 8
m não amostram eficientemente serpentes (cf. LLOYD, 1968; INGER, 1980; SCOTT, 1982;
LIEBERMAN, 1986; GIARETTA et al., 1997; GIARETTA, 1999; GIARETTA et al., no prelo;

13
presente estudo) e lagartos em determinadas localidades (cf. GIARETTA et al., 1997;
GIARETTA, 1999; GIARETTA et al., no prelo; presente estudo).
ALLMON (1991) trabalhou especificamente com os anuros de serapilheira na
Amazônia Central, e analisou dados relativos a América Central, África e sudeste da Ásia,
propondo a riqueza de 20 espécies como relativamente constante para as florestas tropicais
de baixa altitude, e padrões regionais de densidade de indivíduos: sudeste da Ásia com 1 a 2
indivíduos/100 m2, América do Sul com 4 a 6, África com 9 a 10 e América Central com
14 a 15.
Desde o final da década de 80 tem sido observado o declínio de populações de
anfíbios anuros em todo o planeta (POUGH et al., 1998), inclusive em florestas protegidas (e.
g. DAPTF, 1999a). Para a criação de programas de conservação de anfíbios é necessário o
desenvolvimento de métodos de monitoramento ambiental (BEEBEE, 1996). Em 1991 foi
criado o programa “Declining Amphibian Populations Task Force (DAPTF)”, que têm
estabelecido métodos e protocolos para a amostragem e monitoramento de populações de
anfíbios (WAKE, 1994; veja também DAPTF, 1999b). Uma das maiores dificuldades
encontradas no início do programa foi a falta de informações básicas sobre anfíbios
(WAKE, 1994). Assim, o “DAPTF” tem como objetivo ainda a criação de grupos de
trabalho em todas as regiões do planeta onde vivem os anfíbios, especialmente aquelas para
as quais as informações são escassas (DAPTF, 1999c)
Para as regiões serranas do sudeste do Brasil compreendidas no Domínio
Morfoclimático da Floresta Atlântica (sensu AB’SABER, 1977), popularmente denominado
Mata Atlântica, e particularmente ricas em espécies de anfíbios, estudos de cunho ecológico
e/ou quantitativo sobre a anurofauna são praticamente inexistentes (HADDAD & SAZIMA,
1992). Exceções são os trabalhos recentes que utilizaram o método das parcelas amplas de
8 x 8 m no Estado de São Paulo, sudeste do Brasil (GIARETTA et al., 1997; GIARETTA,
1999; GIARETTA et al., no prelo; presente estudo). A partir destes trabalhos, começam a

14
tornar-se disponíveis informações necessárias para uma primeira abordagem sobre os
padrões de diversidade e densidade de anuros de serapilheira da Mata Atlântica do sudeste
do Brasil, e a comparação destas faunas com as de outras localidades.
A Ilha de São Sebastião, localizada no litoral norte do Estado de São Paulo, possui
extensa área de florestas protegidas por um parque estadual. O conhecimento sobre sua
anurofauna é escasso, baseado em poucos trabalhos de caráter qualitativo que foram
desenvolvidos na ilha (e. g. LUEDERWALDT, 1929; MÜLLER, 1968). Assim como em várias
regiões da Mata Atlântica, para a Ilha de São Sebastião inexistem dados ou trabalhos
quantitativos sobre a anurofauna.
OBJETIVOS
Este estudo teve como objetivos caracterizar:
1) a riqueza específica;
2) a abundância relativa e a dominância;
3) a densidade de indivíduos;
4) a relação com variáveis ambientais
da anurofauna de serapilheira, ao longo de um gradiente altitudinal de 0 a 900 m, na Ilha de
São Sebastião, Estado de São Paulo.

15
MATERIAL E MÉTODOS
Área de estudo
A Ilha de São Sebastião (Figura 1), município de Ilhabela, está localizada no litoral
norte do estado de São Paulo (23o47’ S; 45o24’ W). É a maior ilha marítima do Brasil, e está
incluída no Domínio Morfoclimático da Floresta Atlântica (sensu AB’SABER, 1977).
Sua cobertura vegetal está compreendida na região da Floresta Ombrófila Densa,
apresentando as seguintes formações: Floresta Submontana, Floresta Montana e Vegetação
Secundária (IBGE, 1983).
De relevo montanhoso, apenas aproximadamente 2% de sua superfície total
corresponde a áreas planas ou ligeiramente acidentadas (aqui designadas Florestas de
Baixada), aproximadamente 7% estão em cotas superiores a 900 m de altitude, e seu pico
mais alto atinge os 1.379 m (FRANÇA, 1951). Além de diminutas extensões planas, às vezes
representadas somente pelas praias, apenas duas áreas podem ser reconhecidas como
planícies que corresponderiam a Florestas de Baixada (FRANÇA, 1951): a “Planície do
Perequê”, localizada na face oeste da ilha, 4 km ao sul da “Vila de Ilhabela” (Figura 1), com
aproximadamente 260 ha de área e quase totalmente alterada e urbanizada (obs. pess.); e a
“Planície de Castelhanos”, localizada na face leste da ilha na “Bahia de Castelhanos”
(Figura 1), com aproximadamente 64 ha de área e vegetação secundária (obs. pess.).
Devido à precariedade, descontinuidade ou inexistência de dados disponíveis sobre
pluviosidade e temperatura para a Ilha de São Sebastião e cidade de São Sebastião, são
consideradas neste estudo as informações climáticas disponíveis para o município de
Ubatuba, localizado aproximadamente 70 km ao norte da ilha. O clima em Ubatuba é do
tipo tropical úmido, com chuvas bem distribuídas ao longo do ano, precipitações anuais ao
redor de 2,5 m e temperaturas médias de 21,2oC, sendo que de junho a setembro ocorrem

16
Figura 1 - Imagem de satélite modificada da Ilha de São Sebastião (Landsat; bandas 3,4 e 5). S.S. = cidade de São Sebastião; V.I. = "Vila de Ilhabela"; P.P. = "Planície do Perequê"; B.C. = "Bahia de Castelhanos"; T1 = Transecto 1 ("Estrada de Castelhanos"); T2 = Transecto 2 ("Morro do Pacuíba"); T3 = Transecto 3 ("Morro do Ramalho").

17
as menores temperaturas (Instituto Agronômico de Campinas, dados não publicados apud
GIARETTA, 1999). Na região ocorrem de um a dois meses com precipitação mensal inferior
a 100 mm, e janeiro é o mês que apresenta os maiores valores (GIARETTA, 1999).
Em 1958 a Ilha de São Sebastião tornou-se Área de Proteção Ambiental (GOMES &
RAMOS, 1994). O Parque Estadual de Ilhabela (PEIB) foi criado em 1977 com 27.025 ha de
área total (IBGE, 1983) e engloba cerca de 80% da ilha, assim como a totalidade das outras
ilhas, ilhotas e lajes que compõem o arquipélago (SMA, não publicado). Na maior parte da
face oeste da ilha, voltada para o canal de São Sebastião, o PEIB está restrito a altitudes
superiores a cota dos 200 m, incluindo em outras regiões cotas de 100 m e, mais raramente,
0 m de altitude.
Coleta de dados
A coleta da anurofauna de serapilheira foi realizada através do método das parcelas
amplas de 8 x 8 m (“Quadrat Sampling, Large Quadrats” sensu JAEGER & INGER, 1994),
modificado pelo uso de redes que cercaram as parcelas.
Entre 10 de dezembro de 1997 e 30 de janeiro de 1998, foram amostradas 92
parcelas em três diferentes transectos (Figura 1), o que correspondeu a área total de 5.888
m2 de amostragem da serapilheira. A coleta foi realizada nos meses mais quentes e úmidos
da região para evitar efeitos de sazonalidade climática.
Os três transectos amostrados (Figura 1; Tabela 1) correspondem a morros onde o
acesso foi possível. O Transecto 1, “Estrada de Castelhanos” (altitude máxima:
aproximadamente 750m), corresponde ao trecho da estrada de terra entre a “Bahia de
Castelhanos” e o ponto mais alto da encosta, na face leste da ilha. O Transecto 2, “Morro
do Pacuíba” (altitude máxima: aproximadamente 800m), corresponde a uma trilha, em
parte aberta pela equipe de trabalho, ao norte da ilha. O Transecto 3, “Morro do Ramalho”

Tabela 1 - Distribuição de altitudes dos pontos de amostragem em cada transecto amostrado na Ilha de São Sebastião. T1 = “Estrada de Castelhanos”; T2 = ”Morro do Pacuíba“; T3 = ”Morro do Ramalho”.
Atitude (m) T1 T2 T3
0
X
100
X X
200
X X X
300
X X X
400
X X X
500
X X X
600
X X X
700
X X X
800
X
900
X

19
(altitude máxima: aproximadamente 1.100 m), corresponde a uma trilha, aberta pela equipe
a partir dos 700m de altitude, na face oeste da ilha.
Em cada transecto, foi utilizado um delineamento amostral estratificado,
apropriado para amostragens ao longo de gradientes. Para cada transecto foram
determinados pontos de amostragem correspondentes à área de uma circunferência com 58
m de raio (aproximadamente 1,057 ha). Os pontos de amostragem foram determinados a
cada 100 m de altitude a partir do nível do mar, através de pelo menos três leituras de
altímetro (“Thommen Classic TX-12”; acurácia de 10m; Tabela 1). Nos três transectos
foram amostrados 23 pontos (aproximadamente 24,307ha).
Não foi possível a amostragem das mesmas faixas altitudinais nos três transectos
devido à pouca disponibilidade de ambientes preservados em baixas altitudes nos
transectos 2 e 3, e à altura máxima dos topos de morro acessíveis, entre 700 e 800m de
altitude nos transectos 1 e 2. Assim, a amostragem desigual em relação às faixas altitudinais
nos três transectos (Tabela 1) reflete, em parte, a disponibilidade de ambientes preservados
na Ilha de São Sebastião.
Em cada ponto de amostragem, foram realizadas quatro parcelas (totalizando 256
m2), determinadas por duas séries de aleatorizações. Na primeira, foi definida a direção a
partir do centro do ponto de amostragem, utilizando-se uma bússola (N, NE, E, SE, S, SO,
O ou NO). Na segunda, foi definida a distância a partir do centro (1 a 50m).
Cada parcela correspondeu a um quadrado de 8 m de lado (Figura 2A), e foi
amostrada por uma equipe previamente treinada, que variou de quatro a oito pessoas (na
maioria das vezes cinco ou seis pessoas). Após a determinação do local, uma pessoa
caminhou ao redor da área aproximada da parcela para homogeneizar a perturbação. A
parcela foi delimitada por quatro cordas com 8 m de comprimento. Em cada lado da
parcela, foi estendida uma rede de fibra sintética branca, com malha de aproximadamente 1

20
Figura 2 - Aplicação do método das parcelas amplas de 8 x 8 m no campo. A: parcela delimitada por cordas e cercada por redes; B: detalhe de um membro da equipe removendo a serapilheira com a ajuda de tridente manual de jardinagem; C: instalação e cobertura da porção inferior da rede com serapilheira; D: amostragem das redes.
A
B
C D

21
mm, 60 cm de altura e 9 m de comprimento, sustentada por 4 estacas de alumínio fixas à
mesma e enterradas no solo. Os primeiros 10 cm, na base de cada rede, foram dobrados
em direção ao interior da parcela e cobertos com serapilheira e pedras, ficando os 50 cm
restantes acima do solo (Figura 2C). As extremidades das redes foram entrelaçadas umas às
outras com as estacas externas e sobras de aproximadamente 1 m de cada rede, para evitar
frestas entre duas redes adjacentes.
Para cada parcela foram registradas, sempre pela mesma pessoa, as seguintes
informações: 1) número da parcela; 2) data; 3) local; 4) horário de início e término da
amostragem; 5) pressão atmosférica no início e término da amostragem (altímetro
“Thommen Classic TX-12”; acurácia de 1 mmHg); 6) condições gerais do clima (sol,
nublado, chuva, garoa ou neblina); 7) estimativa da distância de riachos ou corpos d’água;
8) altitude; 9) cobertura da copa dentro da parcela (densiômetro vertical “GRS
Densitometer”com 25 pontos de amostragem); 10) cobertura do solo por serapilheira
(estimativa de porcentagem entre 0 e 100%, com intervalos de 20%); 11) cobertura do solo
por pedras (estimativa de porcentagem entre 0 e 100%, com intervalos de 20%); 12)
cobertura do solo por arbustos (estimativa de porcentagem entre 0 e 100%, com intervalos
de 20%); 13) densidade do subbosque (estimativa de porcentagem entre 0 e 100%, com
intervalos de 20%); 14) cobertura do solo por bromeliáceas de chão (estimativa de
porcentagem entre 0 e 100%, com intervalos de 20%); 15) quantidade de bambu
(categorias: nada, pouco, médio ou muito); 16) quantidade de bromeliáceas em geral
(categorias: nada, pouco, médio ou muito); 17) quantidade de cipó (categorias: nada, pouco,
médio ou muito); 18) número de palmeiras; 19) altura máxima do dossel (clinômetro e
trena); 20) umidade relativa do ar na serapilheira em cinco pontos: a 1 m de cada vértice e
no centro quadrado (termohigrômetro digital “Protimeter”; acurácia de 2%); 21) umidade
relativa do solo em cinco pontos: a 1 m de cada vértice e no centro do quadrado (a 1 cm de
profundidade; medidor de condutividade do solo; acurácia de 10%); 22) altura da

22
serapilheira em cinco pontos: a 1 m de cada vértice e no centro do quadrado (régua
milimetrada); 23) altura da camada de raízes em cinco pontos: a 1 m de cada vértice e no
centro do quadrado (régua milimetrada); 24) temperatura da serapilheira no centro do
quadrado (termômetro de mercúrio; acurácia de 0,5oC e estimativa de 0,1oC); 25) número
de formigueiros; 26) número de cupinzeiros; 27) número de buracos; 28) comprimento e
perímetro de troncos caídos no solo (trena milimetrada); 29) perímetro a altura do peito de
árvores (diâmetro maior do que 3 cm; trena milimetrada); 30) número de árvores com
raízes tabulares e respectivos perímetros a altura do peito (trena milimetrada); 31)
inclinação do terreno nos dois lados paralelos mais inclinados da parcela (clinômetro e
trena; acurácia de 1o); 32) face de inclinação do terreno (N, NE, E, SE, S, SO, O ou NO);
33) número de anuros avistados e coletados na parcela; 34) número de anuros avistados e
coletados nas redes; 35) espécies de anuros avistadas e coletadas na parcela; e 36) espécies
de anuros avistadas e coletadas nas redes.
Após a instalação da parcela e das redes, a serapilheira foi cuidadosamente
removida pela equipe, que trabalhava de joelhos com a ajuda de tridentes manuais de
jardinagem (Figura 2B). Pelo menos uma pessoa se posicionou em cada lado da parcela,
sempre delimitando uma faixa de solo sem serapilheira de aproximadamente 40 cm, para
tornar visível a fuga de anuros. A serapilheira foi removida das periferias para o centro do
quadrado. Buracos ou frestas de troncos e pedras foram examinados com lanternas. Todos
os anuros encontrados na serapilheira ou vegetação até a altura de 50 cm foram coletados.
Após a amostragem da parcela, as quatro redes foram cuidadosamente examinadas (Figura
2D) e os anuros encontrados coletados. Os anuros foram acondicionados em sacos
plásticos umedecidos e marcados, separadamente em relação à coleta nas redes e na parcela
e/ou quando apresentavam tamanhos muito diferentes.
Todas as parcelas foram amostradas durante o dia, entre 06:20 e 18:30h, sendo que
a duração de cada uma variou entre 1h:18 min e 3h:11 min (em média, 2h:01min).

23
Em laboratório, os anuros foram identificados, medidos (comprimento rostro-
cloacal; paquímetro com acurácia de 0,05 mm), mortos em álcool a 10%, rotulados, fixados
em formol a 10%, e conservados em álcool a 70%. O tempo entre a coleta e a fixação
variou de 5 a 120h.
Análise de dados
Foram consideradas para as análises de dados as 92 parcelas amostradas na Ilha de
São Sebastião, e outros métodos, como procura visual, encontros ocasionais e procura em
sítios de reprodução, utilizados durante 13 meses de coleta preliminar na localidade (R. J.
SAWAYA, dados não publicados).
A riqueza específica de anuros de serapilheira foi determinada a partir das espécies
encontradas nas parcelas que foram consideradas como habitantes regulares da serapilheira
(veja abaixo, em “Espécies regulares de serapilheira”). Na lista de espécies de anuros de
serapilheira conhecidas para a localidade, foi incluída também uma espécie não coletada nas
parcelas (Tabela 2; Figura 3).
Para verificar se a maioria das espécies de serapilheira da localidade foram
encontradas nas parcelas, foi feita uma curva média do coletor. Esta curva foi feita com o
programa “EstimateS” (COLWELL, 1997), baseada em 200 curvas de acumulação de
espécies com aleatorização da ordem das parcelas.
Para estimar a riqueza específica de anuros de serapilheira da localidade, através do
número de espécies encontrado nas parcelas, foi escolhido um estimador de riqueza
baseado no método “Jacknife” (KUENOUILLE, 1956) que indica a ordem de “Jacknife” que
corresponde à melhor estimativa (BURNHAM & OVERTON, 1979). Assim, o número de
espécies mais ou menos o erro padrão da estimativa é expresso por N(JK), e a ordem do

24
“Jacknife”, que corresponde à melhor estimativa, por K (escolhida quando p>0,05; para
maiores detalhes veja BURNHAM & OVERTON, 1979).
A abundância relativa de cada espécie foi determinada através das porcentagens do
número de indivíduos de cada espécie em relação ao total de indivíduos avistados e
identificados nas parcelas.
A dominância foi determinada pelo índice de Berger-Parker (sensu MAGURRAN,
1988) expresso em porcentagem, que é simplesmente a porcentagem da espécie mais
abundante em relação ao total de indivíduos.
Nas análises de densidade, foram incluídos todos os anuros avistados nas parcelas:
os coletados, os avistados e identificados, e os avistados e não identificados. A densidade
de indivíduos foi expressa em média de indivíduos por 100 m2 apenas para a comparação
com trabalhos realizados em outras localidades. Para as outras análises, a densidade foi
expressa como mediana do número de indivíduos encontrados por parcela, por representar
uma medida de tendência central mais confiável, devido a inclinação, “outliers” e valores
extremos dos dados (ZAR, 1996). Para a confecção dos gráficos “Box-Whiskers”, e
determinação dos “outliers” e valores extremos, foi utilizado o programa “Statistica”
(STATSOFT, 1998).
Para testar as diferenças em relação ao tamanho rostro-cloacal dos indivíduos que
foram encontrados nas redes e nas parcelas, foi utilizado o teste não paramétrico U
(“Mann-Whitney”; ZAR, 1996) do programa “Statistica” (STATSOFT, 1998), considerando
como significativo p<0,05.
Para verificar a relação das variáveis ambientais com a anurofauna de serapilheira,
foi feita uma Análise de Correspondência Canônica (ACC), com o programa CANOCO,
versão 3.10 (TER BRAAK, 1987). Este método foi escolhido por permitir a análise dos dados
ambientais e biológicos conjuntamente, e a escolha das variáveis ambientais que melhor
explicam o padrão de distribuição das espécies. Para a construção da matriz de dados

25
biológicos (89 linhas e 9 colunas), foram consideradas as 89 parcelas que apresentaram pelo
menos um indivíduo, e as nove espécies que estiveram presentes em pelo menos cinco
parcelas. Para a construção da matriz de dados ambientais (89 linhas e 30 colunas), foram
consideradas 30 variáveis ambientais derivadas das informações medidas para cada parcela,
exceto as de número 1, 2, 3 e 32 (veja acima, em “Coleta de dados”). A relação de cada uma
das variáveis ambientais com a matriz de dados biológicos foi verificada pelo teste de
Monte Carlo (MANLY, 1991), com 999 permutações. Para a ordenação da ACC foram
consideradas apenas as nove variáveis significativas segundo este teste (p<0,05).
Comentários taxonômicos
Em virtude da grande riqueza de espécies de anuros da Mata Atlântica e da posição
taxonômica incerta de várias formas, não foi possível a denominação específica de todas as
populações amostradas neste estudo. A identificação e a denominação das espécies aqui
citadas foram baseadas na literatura corrente, consulta a coleções científicas e taxonomistas
que têm trabalhado em localidades correlatas, considerando apenas os caracteres
morfológicos. São comentadas a seguir as formas para as quais a denominação apresenta
problemas ou não foi possível a aplicação de um nome específico.
Brachycephalus sp. (Figura 3A) - população semelhante à de Boracéia, identificada
como Brachycephalus nodoterga por HEYER et al. (1990), mas com algumas diferenças
morfológicas. Brachycephalus nodoterga foi originalmente descrito por Miranda Ribeiro em
1920 para a Serra da Cantareira, localidade da Serra da Mantiqueira do Estado de São
Paulo, mas foi sinonimizado a Brachycephalus ephippium (FROST, 1985), sendo posteriormente
revalidada por HEYER et al. (1990) para Boracéia, Serra do Mar. Porém, a população de
Boracéia denominada B. nodoterga não confere com a diagnose da população descrita por
Miranda Ribeiro para a Serra da Cantareira (FROST, 1985). Assim, não foi atribuído um

26
nome específico à população da Ilha de São Sebastião, pelas diferenças morfológicas que
apresenta e pelo problema existente na denominação de B. nodoterga.
Hyla aff. arianae - trata-se de uma forma não descrita (J. P. POMBAL JR., com. pess.),
relacionada a Hyla arianae CRUZ E PEIXOTO, 1987, descrita e com distribuição conhecida
para o Estado de Santa Catarina.
Adenomera cf. marmorata (Figura 3F) - mais de uma espécie é tratada sob esse nome,
e a denominação para a Ilha de São Sebastião provavelmente inclui duas formas diferentes
(C. F. B. HADDAD, com. pess.), não sendo possível a separação com base em caracteres
morfológicos.
Eleutherodactylus guentheri (Figura 3H) - duas espécies estão sendo tratadas sob esse
nome no presente estudo, E. guentheri e E. aff. erythromerus, sendo o segundo possivelmente
uma forma não descrita (J. P. POMBAL JR., com. pess.). Por serem espécies estreitamente
relacionadas e pelos juvenis, que ocorreram em maior número, não serem distinguíveis
morfologicamente (A. A. GIARETTA, com. pess.), foi utilizado um único nome.
Eleutherodactylus aff. lacteus (Figura 3J) - Eleutherodactylus lacteus MIRANDA RIBEIRO,
1923, foi originalmente descrito de Iguape, Estado de São Paulo, mas o nome tem sido
atribuído também a espécimes do alto Itatiaia (J. P. POMBAL JR., com. pess.). Assim, no
momento só é possível afirmar que a forma da Ilha de São Sebastião pertence ao grupo.
Eleutherodactylus aff. parvus (Figura 3L) - apresenta diferenças morfológicas marcantes
em relação a Eleutherodactylus parvus, mas não há um nome disponível na literatura para esta
população. O grupo está sendo revisto por C. A. G. Cruz, que chama a atenção ao fato de
haver pelo menos quatro espécies distintas sob o nome E parvus (J. P. POMBAL JR., com.
pess.).
Physalaemus aff. signifer (Figura 3N) - possivelmente trata-se de uma forma não
descrita, pois não corresponde morfologicamente a nenhuma das espécies descritas do
grupo signifer.

27
Chiasmocleis cf. leucosticta (Figura 3P) - Chiasmocleis urbanae BOKERMANN, 1952,
descrito para a Ilha de São Sebastião, foi sinonimizado a Chiasmocleis leucosticta
(BOKERMANN, 1952), mas foi considerado como espécie válida por FROST (1985). CRUZ et
al. (1997) concordam com BOKERMANN (1952), considerando C. urbanae sinônimo de C.
leucosticta. O único indivíduo encontrado neste estudo apresenta diferenças morfológicas em
relação a C. leucosticta, mas a série tipo de C. urbanae não foi consultada, havendo, portanto
dúvidas quanto à identificação do espécime.
Espécies de anuros regulares de serapilheira
Não há um critério bem estabelecido para definir quais espécies de anuros são
habitantes regulares da serapilheira. Algumas definições têm sido propostas na literatura,
tais como: espécies que ocupam o chão da floresta pelo menos 25% do tempo (LLOYD et
al., 1968; SCOTT, 1976); que passam uma porção significativa de suas vidas ao nível da
serapilheira (ALLMON, 1991); que vivem diretamente no chão da floresta, ou alimentam-se
de organismos que vivem na serapilheira (HEINEN, 1992). Em alguns casos todas as
espécies coletadas na serapilheira são consideradas para as análises (LIEBERMAN, 1986), ou
o critério estabelecido não está claro (FAUTH, 1989). SCOTT (1982) define arbitrariamente
as espécies regulares de serapilheira, enfatizando que o conhecimento sobre o habitat das
espécies africanas é escasso. Apesar de LLOYD et al. (1968) proporem um critério claro,
incluem nas análises de diversidade espécies ocasionais de serapilheira (“refugee species”)
que foram coletadas nas parcelas, considerando-as como membros da taxocenose, e
enfatiza que a decisão é arbitrária mesmo quando as informações sobre o grupo são
abundantes.
Para a Mata Atlântica do sudeste do Brasil, informações sobre história natural de
anuros em geral são escassas, o que dificulta a definição das espécies regulares de

28
serapilheira. No presente estudo o critério também foi arbitrário, mas, sempre que possível,
baseado em observações realizadas durante 13 meses de coletas preliminares na Ilha de São
Sebastião, e a aplicação de outros métodos que não as parcelas (R. J. SAWAYA, dados não
publicados). Gêneros mais comumente considerados com regulares de serapilheira são
Eleutherodactylus, Brachycephalus, Adenomera, Bufo, Dendrophryniscus, Physalaemus, Proceratophrys,
Chiasmocleis e Myersiella (cf. SCOTT, 1976; TOFT, 1982; LIEBERMAN, 1986; FAUTH, 1989;
ALLMON, 1991; HEINEN, 1992; GIARETTA et al., 1997; GIARETTA, 1999; GIARETTA et al.,
no prelo). GIARETTA (1999) também considerou Scinax trapicheiroi como regular de
serapilheira. Outras espécies do mesmo grupo (catharinae) também podem ser encontradas
freqüentemente na serapilheira (A. A. GIARETTA, com. pess.). Assim, no presente estudo,
Scinax argyreornatus foi considerada como espécie regular de serapilheira. Apesar de
GIARETTA (1999) ter considerado uma espécie de Hyla do grupo circumdata como ocasional
de serapilheira, Hyla hylax, no presente estudo, foi considerada como espécie regular de
serapilheira, pois espécies do grupo podem ser encontradas freqüentemente neste ambiente
(Hyla hylax: obs. pess., e Hyla luctuosa: GIARETTA et al., 1997; GIARETTA et al., no prelo).
Espécies encontradas em parcelas no presente estudo, mas consideradas como
ocasionais de serapilheira, foram: Flectonotus fissilis, por estar associada a bromeliáceas, e
Hyla aff. arianae, por estar associada a poças temporárias (obs. pess.).

29
RESULTADOS
Em 92 parcelas amostradas na Ilha de São Sebastião, foram avistados 865 anuros de
serapilheira. Destes, 804 (92,9%) foram coletados e 61 (7,1%) foram avistados mas não
coletados. Dos 61 indivíduos avistados e não coletados, 42 puderam ser identificados com
segurança ao nível de espécie. Assim, para os 865 indivíduos avistados, foi possível a
identificação de 846 (97,8%), sendo que os 19 (2,2%) restantes não foram identificados e só
foram considerados para as análises de densidade.
Riqueza específica
Foram encontradas 17 espécies de anuros nas 92 parcelas amostradas, das quais 15,
de cinco famílias, foram consideradas como habitantes regulares da serapilheira (Tabela 2).
Para a Ilha de São Sebastião são conhecidas atualmente 16 espécies de anuros de
serapilheira (Tabela 2; Figura 3). O número de espécies encontrado nos transectos 1, 2 e 3
foi, respectivamente, nove, oito e quatorze (Tabela 2).
A curva média do coletor (Figura 4) não atingiu um platô, indicando que não foram
coletadas todas as espécies regulares de serapilheira existentes na localidade. O estimador
de riqueza escolhido, derivado do método “Jacknife” (BURNHAM & OVERTON, 1979),
indica que aproximadamente 20 espécies de serapilheira ocorrem na Ilha de São Sebastião
[N(JK)=20 ± 3,16; K=1; T(K)=1,74; p=0,08].
Em relação à altitude, entre 200 e 600 m a riqueza específica foi maior do que entre
0 e 100 m e 700 e 900 m (Figura 5).

30
Tabela 2 - Lista de espécies de anuros de serapilheira conhecidas para a Ilha de São Sebastião e espécies ocasionais de serapilheira coletadas nas 92 parcelas amostradas. A ocorrência das espécies regulares de serapilheira em cada transecto amostrado é indicada por um “x”; T1=Transecto 1; T2=Transecto 2; T3=Transecto 3.
FAMÍLIA ESPÉCIE T1 T2 T3 Brachycephalidae Brachycephalus sp. - - x Bufonidae Bufo crucifer x x x
Dendrophryniscus brevipollicatus x x x Hylidae Flectonotus fissilis 1 Hyla aff. arianeae 1
Hyla hylax x - - Scinax argyreornatus - - x
Leptodactylidae Adenomera cf. marmorata x x x Eleutherodactylus binotatus x x x Eleutherodactylus guentheri x x x Eleutherodactylus parvus x x x Eleutherodactylus aff. lacteus - - x Eleutherodactylus aff. parvus - - x Hylodes phyllodes x x x Physalaemus aff. signifer 2 Proceratophrys boiei - - x
Microhylidae Chiamoscleis cf. leucosticta - - x Myersiella microps x x x
1: Espécies ocasionais de serapilheira. 2: Espécie regular de serapilheira (R. J. Sawaya, dados não publicados) não encontrada nas 92 parcelas amostradas.

31
Figura 3 - Espécies de anuros de serapilheira conhecidas para a Ilha de São Sebastião. Família Brachycephalidae - A: Brachycephalus sp. Família Bufonidae - B: Bufo crucifer; C: Dendrophryniscus brevipollicatus Família Hylidae - D: Hyla hylax; E: Scinax argyreornatus Família Leptodactylidae - F: Adenomera cf. marmorata
B
C D
A
E F

32
Figura 3 - continuação Família Leptodactylidae - G: Eleutherodactylus binotatus; H: Eleutherodactylus guentheri; I: Eleutherodactylus parvus; J: Eleutherodactylus aff. lacteus; L: Eleutherodactylus aff. parvus; M: Hylodes phyllodes
G H
J
L
I
M
33
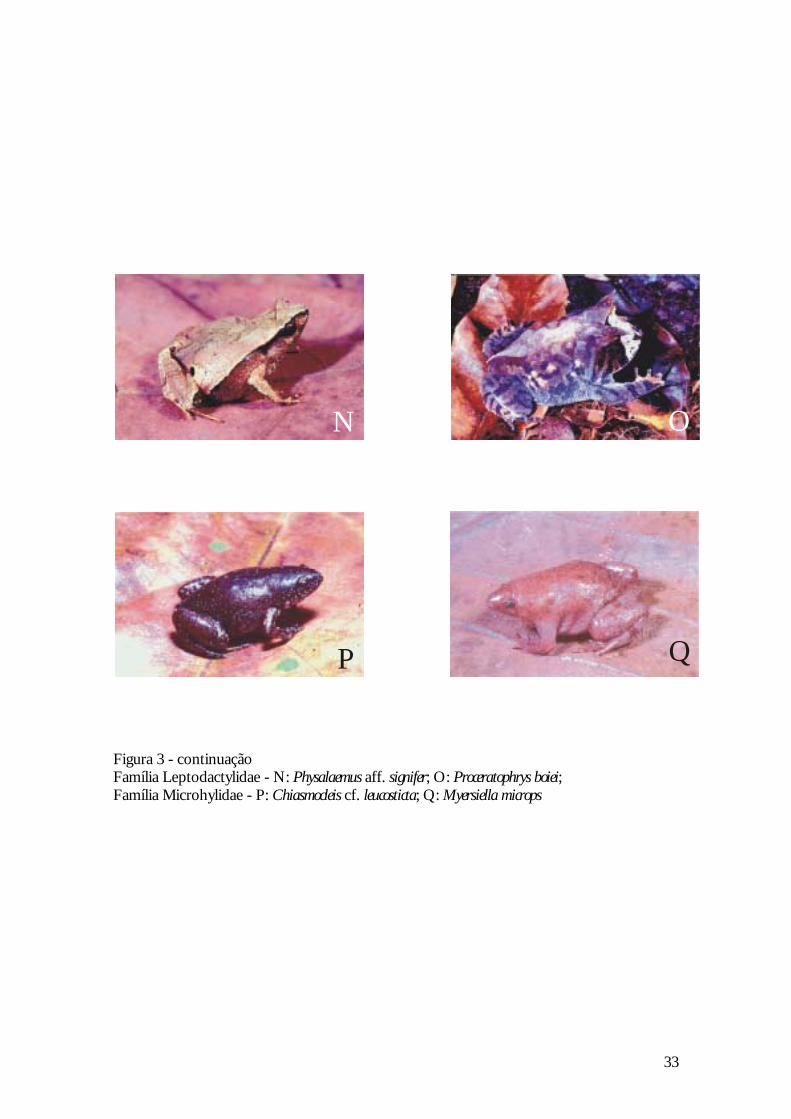
Figura 3 - continuação Família Leptodactylidae - N: Physalaemus aff. signifer; O: Proceratophrys boiei; Família Microhylidae - P: Chiasmocleis cf. leucosticta; Q: Myersiella microps
O
P
N
Q

34
Figura 4 - Curva média do coletor para as 92 parcelas amostradas na Ilha de São Sebastião, baseada em 200 curvas de acumulação de espécies com aleatorização da ordem das parcelas (para detalhes veja em “Material e métodos; Análise de dados”).
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
0 10 20 30 40 50 60 70 80 90
número cumulativo de parcelas
núm
ero
cum
ulat
ivo
de e
spéc
ies

35
Figura 5 - Variação da riqueza específica de anuros de serapilheira ao longo do gradiente altitudinal amostrado. Inclui Physalaemus aff. signifer, não coletado nas parcelas.
0
2
4
6
8
10
12
0 100 200 300 400 500 600 700 800 900
altitude (m)
rique
za e
spec
ífica

36
Em relação à distribuição altitudinal das 16 espécies de anuros de serapilheira
conhecidas para a Ilha de São Sebastião (baseada nas 92 parcelas amostradas e outros
métodos; R. J. SAWAYA, dados não publicados; Figura 6), nenhuma espécie foi restrita à
Floresta de Baixada. Physalaemus aff. signifer, Chiasmocleis cf. leucosticta e Proceratophrys boiei
foram restritos entre 0 e 300 m; apenas Eleutherodactylus parvus distribuiu-se ao longo de
todo o gradiente altitudinal; Brachycephalus sp. foi restrito entre 700 e 900 m; e
Eleutherodactylus aff. lacteus e Eleutherodactylus aff. parvus foram restritos aos 900 m de altitude
(Figura 6).
Abundância relativa de espécies e dominância
A anurofauna de serapilheira da Ilha de São Sebastião é composta por cinco
espécies relativamente comuns com contribuição percentual entre 5 e 35%, e dez espécies
raras com contribuição inferior a 2%, em relação ao total de indivíduos (Figura 7).
A espécie dominante foi Eleutherodactylus parvus, que representou 34,7% do total de
indivíduos avistados e identificados.
Ao longo do gradiente altitudinal, as quatro espécies mais abundantes,
Eleutherodactylus parvus, Eleutherodactylus binotatus, Brachycephalus sp. e Adenomera cf. marmorata,
foram dominantes em duas ou três diferentes faixas altitudinais (Figura 8). Adenomera cf.
marmorata foi dominante entre 0 e 200 m de altitude; E. parvus em 300, 600 e 700 m; E.
binotatus em 400 e 500 m; e Brachycephalus sp. em 800 e 900 m (Figura 8). Considerando
apenas a espécie que foi dominante em cada faixa altitudinal, não foi observado aumento
da porcentagem de dominância com o aumento da altitude (Figura 8).

37
Figura 6 - Avaliação da distribuição altitudinal dos anuros de serapilheira conhecidos para a Ilha de São Sebastião. Inclui Physalaemus aff. signifer, não coletado nas parcelas.
faixas altitudinais (m) 0 100 200 300 400 500 600 700 800 900 Physalaemus aff. signifer xxxxxxxxxxxx Chiamoscleis cf. leucosticta xxx Proceratophrys boiei xxxxxxxx Adenomera cf. marmorata xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Hylodes phyllodes xxxxxxxxxxxxxxxxxxxxxxxxxxxx Myersiella microps xxxxxxxxxxxxxxxxxxxxxxx Hyla hylax xxxxxxxxxxxxxxxxxxxxxxx Bufo crucifer xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Dendrophryniscus brevipollicatus xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Eleutherodactylus parvus .xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Eleutherodactylus binotatus xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Scinax argyreornatus xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Eleutherodactylus guentheri xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx Brachycephalus sp. xxxxxxxxxxxx Eleutherodactylus aff. lacteus xxx Eleutherodactylus aff. parvus xxx
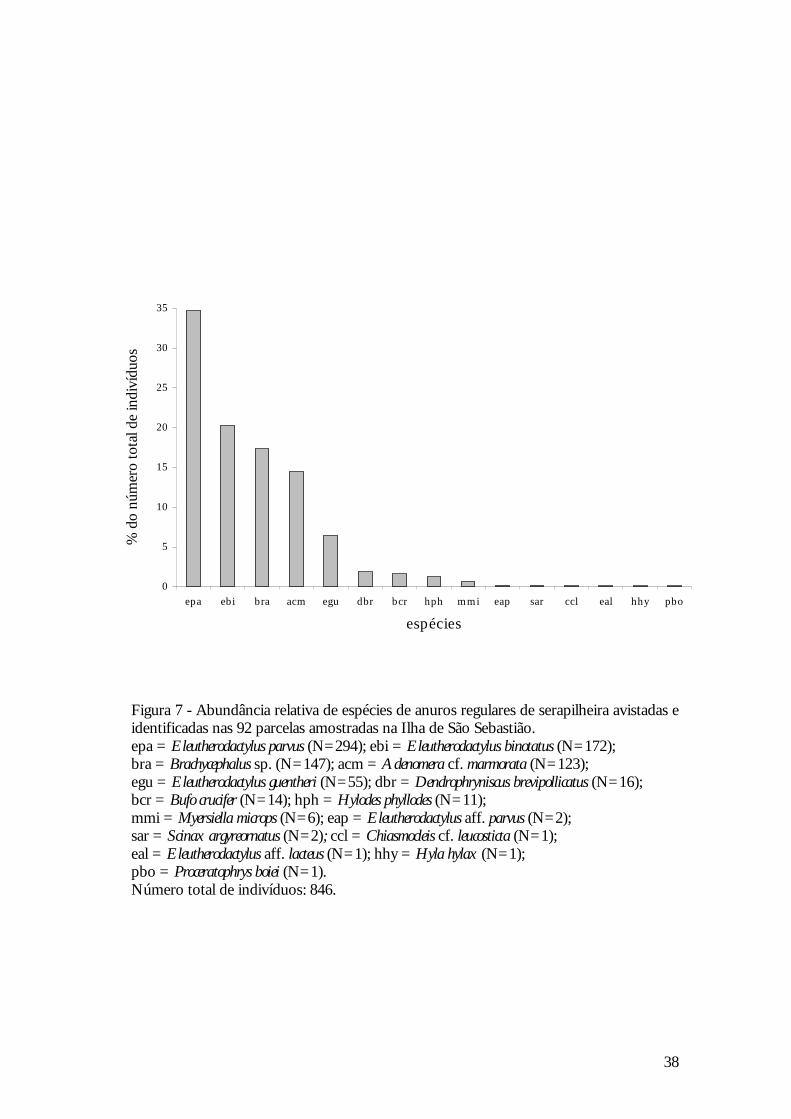
38
Figura 7 - Abundância relativa de espécies de anuros regulares de serapilheira avistadas e identificadas nas 92 parcelas amostradas na Ilha de São Sebastião. epa = Eleutherodactylus parvus (N=294); ebi = Eleutherodactylus binotatus (N=172); bra = Brachycephalus sp. (N=147); acm = Adenomera cf. marmorata (N=123); egu = Eleutherodactylus guentheri (N=55); dbr = Dendrophryniscus brevipollicatus (N=16); bcr = Bufo crucifer (N=14); hph = Hylodes phyllodes (N=11); mmi = Myersiella microps (N=6); eap = Eleutherodactylus aff. parvus (N=2); sar = Scinax argyreornatus (N=2); ccl = Chiasmocleis cf. leucosticta (N=1); eal = Eleutherodactylus aff. lacteus (N=1); hhy = Hyla hylax (N=1); pbo = Proceratophrys boiei (N=1). Número total de indivíduos: 846.
0
5
10
15
20
25
30
35
epa ebi bra acm egu dbr bcr hph m m i eap sar ccl eal hhy pbo
espécies
% d
o nú
mer
o to
tal d
e in
diví
duos

39
Figura 8 - Contribuição percentual das espécies dominantes ao longo do gradiente altitudinal amostrado. acm = Adenomera cf. marmorata; bra = Brachycephalus sp.; ebi = Eleutherodactylus binotatus; epa = Eleutherodactylus parvus.
0
10
20
30
40
50
60
0 100 200 300 400 500 600 700 800 900
altitude (m)
% d
o nú
mer
o to
tal d
e in
diví
duos
acmbraebiepa

40
Densidade
A densidade média de anuros de serapilheira da Ilha de São Sebastião foi de 14,7
ind./100 m2. O número de indivíduos avistados por parcela (ind./par.) variou entre 0 e 56,
com mediana 7,0, e primeiro e terceiro quartis 3,0 e 10,3 ind./par., respectivamente.
Ao longo do gradiente altitudinal, foi observada uma tendência de queda da
densidade (ind./par.) nas faixas entre 500 e 700 m de altitude, e aumento expressivo nas
faixas de 800 e 900 m (Figura 9).
Em função das diferentes altitudes mínimas e máximas amostradas nos três
transectos, o número de indivíduos avistados por parcela ao longo do gradiente altitudinal
foi analisado separadamente para cada transecto (Figura 10). Os transectos 1 e 2 são
semelhantes em relação à tendência de queda na densidade de indivíduos entre 400 e 700 m
de altitude, mas apresentam diferenças na faixa dos 300 m. O transecto 3 é semelhante aos
transectos 1 e 2, apresentando baixas densidades em 500 e 600 m de altitude, mas distinto
na faixa dos 700 m, onde a densidade é mais alta. Nas faixas de 800 e 900 m (altitudes não
amostradas nos transectos 1 e 2), o transecto 3 apresenta as maiores densidades observadas
em qualquer faixa altitudinal ou transecto.
Para as quatro espécies mais abundantes, que foram dominantes em duas ou três
faixas altitudinais (Figura 8) e representaram 87% do total de indivíduos avistados e
identificados, foi observada variação na densidade ao longo do gradiente altitudinal (Figura
11).
Eleutherodactylus parvus, que ocorreu entre 0 e 900 m de altitude, apresentou um
pequeno aumento de densidade em 400 e 500 m, e as mais altas densidades em 800 e 900
m.

41
Figura 9 - Variação do número de indivíduos avistados por parcela (“Box-Whiskers”) ao longo do gradiente altitudinal amostrado. Linhas horizontais = mediana; limites das caixas = 1o e 3o quartis (entre 25% e 75% dos valores observados); linhas verticais = valores mínimos e máximos excluindo-se “outliers” e extremos; o = “outliers”; + = valores extremos. O número de parcelas amostrado ao longo do gradiente está indicado acima dos valores máximos observados para cada faixa altitudinal.
altitude (m)
núm
ero
de in
diví
duos
por
par
cela
0
10
20
30
40
50
60
0 100 200 300 400 500 600 700 800 900
N=4
N=4
N=4
N=8N=12
N=12N=12
N=12
N=12
N=12

42
Figura 10 - Variação do número de indivíduos avistados por parcela (“Box-Whiskers”) ao longo do gradiente altitudinal amostrado em cada transecto. A: transecto 1; B: transecto 2; C: transecto 3. Em todas as faixas altitudinais foram amostradas quatro parcelas. Linhas horizontais = mediana; limites das caixas = 1o e 3o quartis (entre 25% e 75% dos valores observados); linhas verticais = valores mínimos e máximos observados.
TRANSECTO 1
altitude (m)
núm
ero
de in
diví
duos
por
par
cela
0
10
20
30
40
50
60
0 100 200 300 400 500 600 700 800 900
TRANSECTO 2
altitude (m)
núm
ero
de in
diví
duos
por
par
cela
0
10
20
30
40
50
60
0 100 200 300 400 500 600 700 800 900
TRANSECTO 3
altitude (m)
núm
ero
de in
diví
duos
por
par
cela
0
10
20
30
40
50
60
0 100 200 300 400 500 600 700 800 900
A
B
C

Figura 11 - Variação dencontradas nas parcelasA: = Eleutherodactylus pB: = Eleutherodactylus bLinhas horizontais = mevalores observados); linhe extremos; o = “outlierPara o número de parce
núm
ero
de in
diví
duos
por
par
cela
0
5
10
15
0 100
núm
ero
de in
diví
duos
por
par
cela
0
10
20
30
0 100
s
Eleutheroactylus parvu
Aaltitude (m)200 300 400 500 600 700 800 900
Brachycephalus sp.
B
o número de indivíduos das quat (“Box-Whiskers”) ao longo do gradiarvus; = Brachycephalus sp. inotatus; = Adenomera cf. marmorata diana; limites das caixas = 1o e 3o quaas verticais = valores mínimos e máx
s”; + = valores extremos. las amostradas em cada faixa altitudin
altitude (m)
200 300 400 500 600
Adenomera cf. marmorata
r
Eleutherodactylus binotatus
43
o espécies mais abundantes ente altitudinal amostrado.
rtis (entre 25% e 75% dos imos excluindo-se “outliers”
al veja acima Figura 10.
700 800 900

44
Eleutherodactylus binotatus, que ocorreu entre 100 e 900 m de altitude, apresentou
aumento de densidade em 200 m, queda em 300 m, maior aumento em 400 m, e queda
progressiva de densidade entre 400 e 900m.
Brachycephalus sp., restrito ao transecto 3 entre 700 e 900m de altitude, apresentou
aumento progressivo de densidade entre 700 e 900m.
Adenomera cf. marmorata, que ocorreu entre 0 e 600m de altitude, apresentou queda
progressiva de densidade entre 0 e 600m.
Relação entre a anurofauna de serapilheira e as variáveis ambientais
Foi testada a relação de 30 variáveis ambientais com a distribuição das nove
espécies de anuros de serapilheira que estiveram presentes em pelo menos cinco parcelas
(veja em “Material e Métodos; Análise de Dados”). Nove variáveis (altitude, altura da
camada de raízes, quantidade de bambu, inclinação do terreno, temperatura da serapilheira,
umidade do solo, distância de riachos, altura da camada de serapilheira e variação da
pressão atmosférica) foram significativamente correlacionadas com a abundância das
espécies, sendo utilizadas para a ordenação das parcelas e das espécies na Análise de
Correspondência Canônica (ACC). A Figura 12 ilustra a ordenação das parcelas e das
espécies ao longo do 1o e 2o eixos da ACC, e os vetores das variáveis ambientais sobre as
quais a análise foi gerada. A variância total explicada por estas nove variáveis foi de 42%.
O eixo 1 da ACC (autovalor: 0,55; p<0,01) ilustra basicamente a distribuição de
espécies ao longo do gradiente altitudinal. A localização de cada espécie em relação a este
eixo indica em que posição do gradiente cada uma é mais abundante. Da menor para a
maior altitude, as espécies distribuem-se ao longo do eixo na seguinte ordem: Bufo crucifer,
Adenomera cf. marmorata, Myersiella microps, Hylodes phyllodes, Eleutherodactylus binotatus,
Dendrophryniscus brevipollicatus, Eleutherodactylus parvus, Eleutherodactylus guentheri e
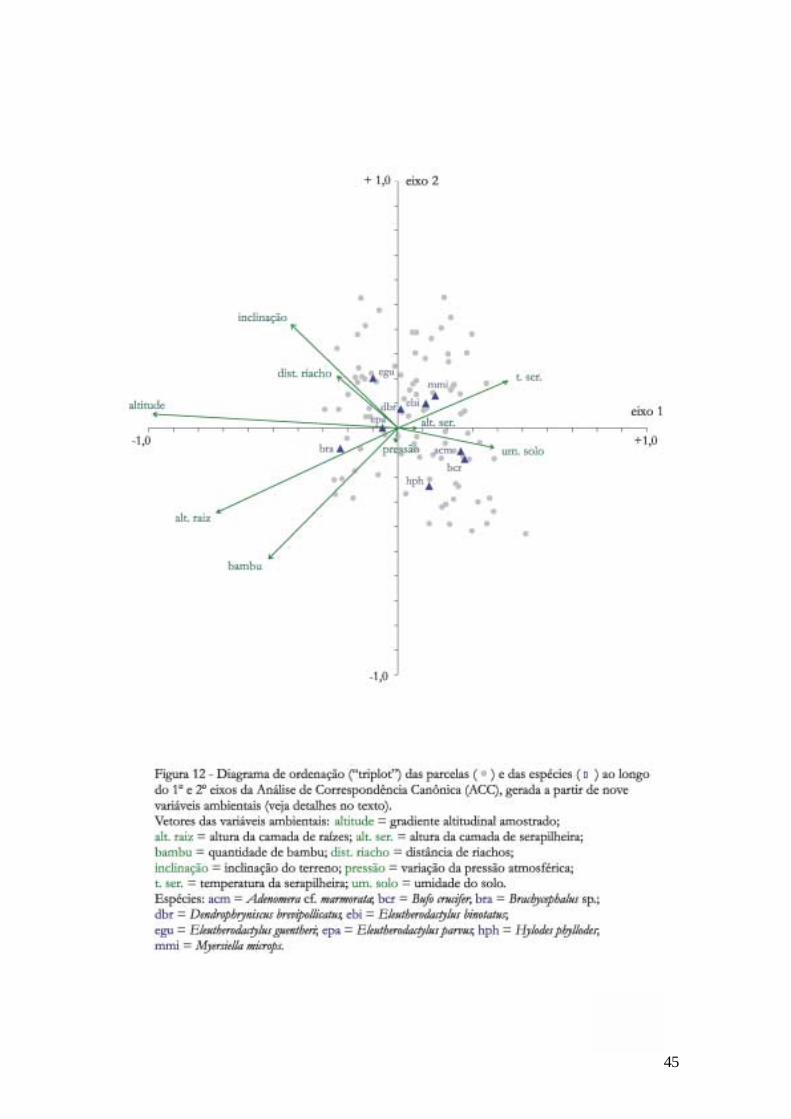
45

46
Brachycephalus sp. As espécies próximas à origem do eixo (Eleutherodactylus parvus e
Dendrophryniscus brevipollicatus) foram encontradas praticamente ao longo de todo o gradiente
(veja também Figura 6). Com o aumento da altitude, também aumenta a altura da camada
de raízes, a quantidade de bambu, a inclinação do terreno e a distância de riachos, e
diminuem a umidade do solo e a temperatura da serapilheira. As variáveis altura da camada
de serapilheira e variação da pressão estão pouco relacionadas com este eixo, sendo
representadas por vetores relativamente curtos.
O eixo 2 da ACC (autovalor: 0,19; p<0,01) está relacionado, por ordem de
importância, às variáveis: quantidade bambu, inclinação do terreno, altura da camada de
raízes, distância de riachos e temperatura da serapilheira.
Hylodes phyllodes, Adenomera cf. marmorata e Bufo crucifer estão ordenados em sentido
contrário a inclinação do terreno e distância de riachos, e no mesmo sentido de umidade do
solo.
Modificação do método das parcelas amplas: uso de redes
A modificação do método das parcelas amplas de 8 x 8 m, pelo uso de redes de
fibra sintética que cercaram as parcelas, aumentou consideravelmente o número de anuros
avistados. Dos 865 anuros avistados, 396 estavam nas redes, o que corresponde a 45,8% do
total de indivíduos.
Todas as espécies avistadas nas redes também foram avistadas nas parcelas. Em
relação ao tamanho (comprimento rostro-cloacal) dos indivíduos avistados nas redes e nas
parcelas, foi observada diferença significativa (p<0,01; U=65.412,5; Nrede=366;
Nparcela=457), sendo que na rede os indivíduos foram menores (mediana 12,8; primeiro e
terceiro quartis 10,0 e 17,6, respectivamente) do que nas parcelas (mediana 14,2; primeiro e
terceiro quartis 11,5 e 22,7, respectivamente).

47
DISCUSSÃO
Riqueza específica
Conhecimentos sobre a anurofauna da Ilha de São Sebastião estão limitados a
poucos estudos (e. g. LUEDERWALDT, 1929; MÜLLER, 1968; CRUZ et al., 1997; R. J. SAWAYA,
dados não publicados; presente estudo). No presente estudo, é estimado para a Ilha de São
Sebastião ao redor de 20 espécies de anuros de serapilheira. Em Ubatuba, a maioria das
espécies de serapilheira é conhecida (cf. GIARETTA, 1999), pois vários pesquisadores têm
trabalhado na localidade nos últimos anos. Assim, GIARETTA (1999) não utilizou qualquer
estimador de riqueza baseado nas parcelas amostradas, e apresenta uma lista de anuros de
serapilheira com 21 espécies. ALLMON (1991) propõe o número de aproximadamente 20
espécies para florestas tropicais de baixa altitude, o que parece se aplicar às localidades
estudadas na Serra do Mar do Estado de São Paulo, Ilha de São Sebastião e Ubatuba.
Para a Serra da Mantiqueira, no Estado de São Paulo, estudos quantitativos e
conhecimentos prévios sobre as localidades estudadas (GIARETTA et al., 1997; GIARETTA et
al., no prelo) apontam um menor número de espécies de anuros de serapilheira, entre 11 e
12 (veja também Tabela 3). As altitudes amostradas na Serra da Mantiqueira, entretanto,
limitaram-se às faixa entre 850 e 1.250m. SCOTT (1976) sugere haver diminuição do
número de espécies com o aumento da altitude e número de meses secos (pluviosidade
menor do que 100 mm). Assim, além das maiores altitudes, as menores pluviosidades e a
ocorrência de maior número de meses secos (Tabela 3) poderiam explicar o menor número
de espécies observados na Serra da Mantiqueira em relação à Serra do Mar. Entretanto, as
localidades da Serra da Mantiqueira correspondem a reservas menores e com maior grau de
perturbação antrópica, o que dificulta a comparação entre as duas serras.

48

49
Em Ubatuba, uma diminuição abrupta da riqueza específica foi observada a partir
dos 550m de altitude, o que correspondeu a uma relação não linear de declínio do número
de espécies com o aumento da altitude (GIARETTA, 1999). Apesar de já ter sido observada
para localidades da América Central (e. g. SCOTT, 1976; FAUTH et al., 1989), a diminuição da
riqueza com o aumento da altitude tem sido tratada na literatura como uma relação linear.
Nas Florestas de Baixada de Ubatuba foi observado um maior número de espécies, com
seis espécies que ocorreram somente aí (GIARETTA, 1999). Na Ilha de São Sebastião, o
padrão de riqueza em relação à altitude é semelhante ao observado para Ubatuba, com
exceção das faixas altitudinais entre 0 e 200m, que apresentam baixo número de espécies
(Figura 5). A menor riqueza observada em menores altitudes na Ilha de São Sebastião
provavelmente está associada à escassez de planícies litorâneas na localidade (veja acima
“Material e métodos; Área de estudo”) e, praticamente, à inexistência de Florestas de
Baixada, que devem representar um ambiente particular.
A teoria da biogeografia de ilhas propõe um menor número de espécies em áreas
menores e mais isoladas (MACARTHUR & WILSON, 1967; veja revisões em PIANKA, 1994;
BEGON et al., 1986). Segundo a teoria, para a Ilha de São Sebastião não seria esperado um
decréscimo importante no número de espécies por sua grande dimensão e proximidade do
continente. Para as altitudes maiores da ilha, entretanto, a diminuição da área deve ser um
fator importante, que poderia explicar o menor número de espécies observado em regiões
mais altas. Brachycephalus sp. está restrito na Ilha de São Sebastião a altitudes acima dos 700
m, tendo sido observado em apenas um dos três transectos amostrados, justamente o que
apresenta as maiores altitudes (aproximadamente 1.100 m; veja também Tabela 2). Os
outros dois transectos apresentam altitudes máximas de aproximadamente 750 e 800m, e
provavelmente áreas muito reduzidas acima dos 700m que suportem populações de
Brachycephalus sp. Além disso, estes transectos apresentam um menor número de espécies
em relação ao transecto 3 (veja Tabela 2).

50
Na Ilha de São Sebastião, assim como em Ubatuba (GIARETTA, 1999), foi
observada alta sobreposição na ocorrência de espécies ao longo do gradiente altitudinal
amostrado (Figura 6). Entretanto uma análise quantitativa (ACC) evidenciou substituição
das nove espécies mais abundantes ao longo do gradiente altitudinal, o que deve estar
relacionado a diferentes preferências de habitat entre as espécies (veja discussão abaixo em
“Densidade de indivíduos” e “Relação entre a anurofauna de serapilheira e as variáveis
ambientais”).
Abundância relativa de espécies e dominância
Apesar de ser esperado para florestas tropicais alta equitatividade na abundância
relativa de espécies (LLOYD et al., 1968), ou seja, baixos valores de dominância, LIEBERMAN
(1986) não considera a equitatividade como uma característica distintiva de florestas
tropicais. Como esperado para a maioria das comunidades ou taxocenoses (KREBS, 1989), a
Ilha de São Sebastião apresenta um maior número de espécies raras, em relação a espécies
comuns (Figura 7). Considerando os valores de dominância para as amostragens
quantitativas das localidades de Mata Atlântica do Estado de São Paulo (Tabela 3), a
variação é considerável. Para as localidades da Serra do Mar a espécie dominante é a
mesma, Eleutherodactylus parvus, mas com valores diferentes no verão em Ubatuba, 51,9%
(GIARETTA, 1999), e Ilha de São Sebastião, 34,7% (presente estudo). Para a única localidade
da Serra da Mantiqueira amostrada no verão, Atibaia, a espécie dominante é diferente,
Brachycephalus ephippium, e o valor de dominância é de 54,3% (GIARETTA et al., no prelo;
Tabela 3). Considerando que Eleutherodactylus parvus também ocorre em Atibaia,
provavelmente a diferença entre as duas serras, em relação à espécie dominante no verão,
está relacionada às diferentes condições climáticas (e. g. pluviosidade média anual e número
de meses secos) e, conseqüentemente, às diferentes tolerâncias ambientais das espécie

51
dominantes, à altitude e/ou à diferente composição de espécies das localidades (mais
parecidas entre si nas localidades da Serra do Mar). Considerando apenas o inverno (Tabela
3), estação não amostrada na Ilha de São Sebastião, a Serra do Mar caracteriza-se por
menores valores de dominância em Ubatuba (43,2%) em relação à Serra da Mantiqueira em
Atibaia (78,5%) e no Japi (71,5%). Nesse caso, os diferentes valores de dominância entre as
duas serras não estão relacionados às espécies dominantes, que são diferentes em Atibaia e
Japi, mas sim à linha de neblina que ocorre em regiões mais altas da Serra da Mantiqueira,
possivelmente aumentando a umidade e diminuindo os efeitos da sazonalidade climática
nessas regiões (GIARETTA et al., 1997; GIARETTA et al., no prelo).
Para a Ilha de São Sebastião, foi observada substituição das espécies dominantes ao
longo do gradiente altitudinal, como também observado para as duas altitudes amostradas
na Serra do Japi (GIARETTA et al., 1997) e no gradiente altitudinal amostrado em Ubatuba
(GIARETTA et al., no prelo). Entretanto, quando considerada a espécie dominante em cada
faixa altitudinal, não foi observado aumento da dominância com o aumento da altitude,
como proposto por SCOTT (1976). É possível que o padrão observado por SCOTT (1976)
seja resultado da comparação de diferentes altitudes em diferentes localidades na América
Central, e não limitada a um gradiente altitudinal contínuo como o analisado no presente
estudo.
Densidade
A densidade média de indivíduos observada para a Ilha de São Sebastião, 14,7
ind./100 m2, não corrobora o padrão regional de ALLMON (1991) proposto para a América
do Sul, de 4 a 6 ind./100 m2. As generalizações de ALLMON (1991) sobre padrões regionais
de densidade apresentam como problemas diferenças dos métodos utilizados nos estudos
que são comparados (e. g. parcelas de 5 x 5m e de 8 x 8 m), esforço amostral diferente nas

52
localidades e falta de replicação em algumas regiões. Para a proposta do padrão regional de
densidade da América do Sul, ALLMON (1991) considerou apenas dados relativos à região
de Manaus, Estado do Amazonas. Sendo a própria Amazônia uma região variável em
relação à abundância de anuros de serapilheira (M. MARTINS, com. pess.), provavelmente a
América do Sul apresenta variação ainda maior. Em relação à Mata Atlântica do Estado de
São Paulo, a Serra do Mar apresenta densidades maiores em relação à Mantiqueira no
verão, a primeira com 14 a 17 ind./100 m2, e a segunda ao redor de 6 ind./100 m2 (dados
disponíveis apenas para Atibaia; Tabela 3).
Ao longo do gradiente altitudinal, na Ilha de São Sebastião, foi observada uma
tendência de queda na densidade de indivíduos nas faixas altitudinais de 500 a 700m, e
aumento expressivo em 800 e 900m (Figura 9). Quando analisados os três transectos
separadamente (Figura 10), os transectos 1 e 2 (limitados entre 0 e 700m de altitude)
apresentam tendências bastante diferentes do transecto 3 (limitado entre 200 e 900m de
altitude) na faixa dos 700 m. Os transectos 1 e 2, para os quais as altitudes máximas dos
morros são de aproximadamente 750 e 800m, respectivamente, apresentam uma queda
progressiva na densidade de indivíduos entre os 400 e 700m. É possível que a baixa
densidade observada nestes transectos nas maiores altitudes esteja relacionada à
proximidade destas com os topos de morro, que podem representar um ambiente distinto
daqueles de altitudes intermediárias. O Transecto 3, com altitude máxima de
aproximadamente 1.100 m, também apresenta menores densidades em 500 e 600m, mas
maiores densidades em 700m, que apenas neste caso está distante do topo de morro. As
grandes densidades observadas em 800 e 900m de altitude no transecto 3, podem estar
relacionadas às grandes distâncias destas faixas altitudinais em relação ao topo do morro,
possibilitando a presença de Brachycephalus sp. e a ocorrência de Eleutherodactylus parvus em
alta densidade. (Figura 11A).

53
Relação entre a anurofauna de serapilheira e as variáveis ambientais
Nove variáveis ambientais apresentaram relação significativa com as abundâncias
das nove espécies de anuros de serapilheira mais amostradas na Ilha de São Sebastião. A
“altitude”, entretanto, não pode ser considerada estritamente como uma variável ambiental,
e sim como uma variável de posição. No diagrama de ordenação da Análise de
Correspondência Canônica (ACC), o vetor “altitude” corresponde praticamente ao eixo 1,
que ilustra a substituição das espécies ao longo do gradiente altitudinal (Figura 12; veja
também figuras 8 e 11). O número relativamente alto de variáveis ambientais relacionadas
às abundâncias das espécies, indica diferentes preferências de habitat entre cada uma delas.
As espécies que estão ordenadas em sentido contrário à altitude e distância de riachos, e em
mesmo sentido de umidade do solo, Adenomera cf. marmorata, Bufo crucifer e Hylodes phyllodes,
são mais abundantes em terrenos menos inclinados e mais próximos a riachos, que
apresentam maior umidade do solo. Bufo crucifer e A. cf. marmorata são espécies oportunistas
que normalmente invadem áreas degradadas (obs. pess.), tendo ocorrido em altas
densidades na única Floresta de Baixada amostrada na ilha, que se encontra mais degradada
que as demais regiões. Adenomera cf. marmorata também está associada a regiões de menor
altitude, onde apresenta as maiores densidades (Figura 11B). Hylodes phyllodes, uma espécie
nitidamente associada a riachos, provavelmente está ordenada em sentido contrário à
altitude em função das parcelas amostradas em regiões mais altas apresentarem maior
distância de riachos (veja na figura 12 a associação entre os vetores altitude e distância de
riachos).

54
Comparação com outras localidades da Mata Atlântica
Apesar de apenas quatro estudos quantitativos sobre a anurofauna de serapilheira
terem sido realizados na Mata Atlântica do Estado de São Paulo, e em reservas com
características diferentes, torna-se possível uma primeira abordagem sobre padrões de
diversidade e densidade para o grupo nas Serras da Mantiqueira e do Mar. Em relação à
Serra da Mantiqueira, a Serra do Mar parece caracterizar-se pela maior riqueza específica e
densidade de indivíduos. Em grande parte, a maior riqueza observada na Serra do Mar
pode estar associada a presença de florestas em menores altitudes. Apenas amostragens
mais detalhadas em altitudes superiores a 850m na Serra do Mar, poderiam confirmar a
hipótese. Para verificar os padrões observados e propostos para cada serra, e possibilitar
uma maior compreensão dos padrões de distribuição altitudinal da anurofauna de
serapilheira, seriam úteis estudos quantitativos em regiões mais preservadas e com
gradientes altitudinais mais longos na Serra da Mantiqueira (e. g. Serra do Itatiaia).
Método das parcelas amplas de 8 x 8 m
O método das parcelas amplas de 8 x 8 m possibilita boas estimativas de
abundância relativa e densidade de anuros de serapilheira (HEYER et al., 1994). Em relação à
riqueza específica, nas duas localidades da Serra do Mar do Estado de São Paulo onde o
método foi utilizado, Ubatuba (GIARETTA, 1999) e Ilha de São Sebastião (presente estudo),
foram coletadas aproximadamente 75% das espécies de serapilheira que provavelmente
ocorrem em cada localidade. Assim, para uma melhor amostragem da riqueza específica de
anuros de serapilheira, seria apropriado combinar dois ou três métodos diferentes dentre
aqueles propostos em HEYER et al. (1994). Apesar de não amostrar toda a fauna de anuros
de serapilheira, as parcelas amplas também podem ser indicadas para a amostragem de

55
espécies dificilmente coletadas através de outros métodos. Em Ubatuba (GIARETTA, 1999),
onde a anurofauna é relativamente bem conhecida, foi coletada uma nova espécie,
Psyllophryne hermogenesi, que ocorre desde o nível do mar até os 700m de altitude (GIARETTA
& SAWAYA, 1998). Na Ilha de São Sebastião (presente estudo), apesar da anurofauna ser
relativamente pouco conhecida, foram coletadas três formas do gênero Eleutherodactylus sem
um nome disponível (E. aff. erythromerus, E. aff. lacteus e E. aff. parvus).
Embora o método das parcelas amplas tenha sido utilizado em outras localidades
do planeta para a amostragem de répteis, parece ser limitado para a amostragem de
serpentes e lagartos. Nas 92 parcelas amostradas na Ilha de São Sebastião foram avistados e
coletados apenas dez lagartos de três espécies, e oito serpentes de cinco espécies (Apêndice
1). Assim, como proposto por JAEGER & INGER (1994), o método deve ser utilizado
apenas para a amostragem de anuros de serapilheira.
A modificação do método pelo uso de redes que cercam as parcelas (GIARETTA,
1999; presente estudo; Figura 2) é de grande importância, por aumentar em até 45% o
número de indivíduos coletados.

56
CONCLUSÕES
A Ilha de São Sebastião apresenta aproximadamente 20 espécies de anuros de
serapilheira, assim como florestas tropicais de baixa altitude em diferentes localidades dos
planeta. Maior número de espécies ocorre entre 200 e 600 m de altitude.
A taxocenose é composta por um maior número de espécies raras, que representam
menos de 5% do total de indivíduos.
Diferentes espécies são dominantes ao longo do gradiente altitudinal, mas não há
um aumento no valor de dominância em maiores altitudes.
A densidade média de indivíduos na Ilha de São Sebastião, 14,7 ind./100 m2, está
entre as maiores densidades observadas em outras florestas tropicais do planeta, sendo
semelhante a de Ubatuba, SP, sudeste do Brasil, e de localidades da América Central.
Ao longo do gradiente altitudinal, a densidade de indivíduos foi menor entre os 500
e 700m de altitude, e maior nos 800 e 900m. Densidades maiores observadas nos 800 e
900m estão limitadas a apenas um dos três transectos amostrados e estão relacionadas à
maior altitude máxima de morros, possibilitando a ocorrência de Brachycephalus sp. e
Eleutherodactylus parvus em alta densidade.
Apesar da alta sobreposição na ocorrência de espécies, análises quantitativas
demonstram substituição de espécies ao longo do gradiente altitudinal amostrado.
Provavelmente esta substituição de espécies nas faixas altitudinais reflete as diferentes
preferências de habitat de cada uma, evidenciadas pela relação de nove variáveis ambientais
com a sua distribuição.
Em relação à Serra da Mantiqueira, a Serra do Mar apresenta maior riqueza
específica e maior densidade de anuros de serapilheira, provavelmente associadas à altitude
e pluviosidade.

57
O método das parcelas amplas de 8 x 8 m não amostra todas as espécies de anuros
de serapilheira, mas permite a coleta de espécies dificilmente amostradas por outros
métodos. Uma importante modificação do método das parcelas amplas é o uso de redes,
que podem aumentar o número de indivíduos coletados em até 45%.

58
RESUMO
Foram caracterizadas a riqueza específica, a abundância relativa e a densidade de
anuros de serapilheira ao longo de um gradiente altitudinal de 0 a 900m, em uma localidade
de Mata Atlântica do sudeste do Brasil. O estudo foi realizado na Ilha de São Sebastião,
litoral norte do Estado de São Paulo, em três diferentes transectos. Foram amostradas 92
parcelas amplas de 8 x 8 m cercadas por redes, durante o dia, entre 10 de dezembro de
1997 e 30 de janeiro de 1998. Foram avistados 865 indivíduos, dos quais 846 de 15
diferentes espécies foram identificados, e 804 foram coletados. Foi estimado para a
localidade um total de 20 espécies de serapilheira, sendo que atualmente são conhecidas 16.
Foi observado um maior número de espécies entre os 200 e 600 m de altitude, sendo que
apenas três foram restritas entre 0 e 300 m, e três entre 700 e 900 m. Uma única Floresta de
Baixada amostrada apresentou baixa riqueza e nenhuma espécie exclusiva, provavelmente
pela escassez deste tipo florestal na localidade. Apesar da alta sobreposição na ocorrência
de espécies, análises quantitativas evidenciaram substituição de espécies ao longo do
gradiente altitudinal amostrado, especialmente para as quatro espécies mais abundantes,
provavelmente devido a diferentes preferências de habitat de cada uma. A taxocenose de
anuros de serapilheira da Ilha de São Sebastião é composta por cinco espécies
relativamente comuns, e pelo menos dez raras com contribuição percentual em relação ao
número total de indivíduos menor do que 5%. As quatro espécies mais abundantes foram
dominantes em duas ou três faixas altitudinais, mas os valores gerais de dominância não
aumentaram em maiores altitudes. A densidade média de indivíduos para a localidade foi de
14,7 ind./100 m2. Foram observadas as menores densidades de indivíduos entre 500 e
700m de altitude. As maiores densidades observadas em 800 e 900m de altitude, faixas
amostradas em apenas um transecto, foram relacionadas a duas espécies, Eleutherodactylus
parvus e Brachycephalus sp. O método das parcelas amplas de 8 x 8 m amostrou

59
aproximadamente 75% do número total de espécies estimado para a localidade e poucas
espécies de lagartos e serpentes, indicando que é apropriado apenas para a amostragem de
anuros de serapilheira, mas deve ser combinado a outros métodos para melhor
caracterização da riqueza total de espécies. Através dos quatro estudos quantitativos
realizados na Mata Atlântica do Estado de São Paulo, em relação à Serra da Mantiqueira a
Serra do Mar caracteriza-se pela maior riqueza e maior densidade de anuros de serapilheira,
o que provavelmente está relacionado a altitude e características pluviométricas.

60
ABSTRACT
The species richness, relative abundance and density of litter frogs were sampled in
a rain forest locality, in Southeast Brazil, along an altitudinal gradient between 0 and 900
meters. The study was carried out in “Ilha de São Sebastião”, São Paulo State, in three
different transects. Ninety-two 8 x 8 m plots enclosed by plastic fences were sampled
during the day between December 10, 1997 and January 30, 1998. Eight hundred and sixty
five individuals were found, of which 846 were identified as belonging to fifteen different
species; 804 were collected. Twenty species of litter frogs were estimated for the locality, of
which sixteen are currently known. Between 200 and 700 meters of altitude a great number
of species was observed and only three were restricted between 0-300m and 700-900m. In
a single lowland forest sampled there was low richness and no restricted species, probably
due to the scarcity of this forest type in the locality. Along the altitudinal gradient there was
high overlap of species occurrence but quantitative analyses illustrated species replacement,
especially for the four more abundant species, probably due to different habitat
preferences. The litter frog assemblage of “Ilha de São Sebastião” has five relatively
common species and at least ten rare species that were represented by fewer than 5% of
the total number of individuals. The four abundant species were dominant in two or three
altitudinal bands, but the general values of dominance were not increased in higher
altitudes. The mean density of individuals observed for the locality was 14,7 ind./100 m2.
The lowest densities were observed between 500 and 700 meters of altitude. The highest
densities observed between 800 and 900 meters, bands sampled in only one transect, were
due to two species, Eleutherodactylus parvus and Brachycephalus sp. The 8 x 8 meter plot
method has sampled approximately 75% of the total number of species estimated for the
locality and a few species of lizards and snakes, indicating that it is a good method only for
litter frog assemblage samples but should be combined with other methods to sample the

61
total species richness. Based on the four quantitative studies of litter frogs in the Atlantic
Rain Forest of southeastern Brazil, “Serra do Mar” could be characterized by a higher
species richness and density of individuals when compared to “Serra da Mantiqueira”,
probably due to altitudinal and pluviometric characteristics.

62
Apêndice 1 - Espécies de lagartos e serpentes, e número de indivíduos coletados em cada faixa altitudinal, nas 92 parcelas amostradas na Ilha de São Sebastião.
subordem família espécie N altitude
Lacertilia
Polychrotidae Enyalius iheringii 7 400 (N=1), 500 (N=4) e
600m (N=2)
Gekkonidae Gymnodactylus darwini 2 200m
Gymnophtalmidae Placosoma glabellum 1 300m
Serpentes
Viperidae Bothrops jararaca 3 200, 500 e 600m
Bothrops jararacussu 1 400m
Colubridae Clelia plumbea 1 400m
Dipsas petersi 1 0m
Echinantera cephalostriata 2 400 e 700m

63
REFERÊNCIAS BIBLIOGRÁFICAS AB’SABER, A. N. 1977. Os domínios morfoclimáticos na América do Sul. Primeira
aproximação. Geomorfologia, 52:1-21. ALLMON, W. D. 1991. A plot study of forest litter frogs, Central Amazon, Brazil.
Journal of Tropical Ecology 7:503-522. BEEBEE, T. J. C. 1996. Ecology and Conservation of Amphibians. Chapman and Hall,
London. BEGON, M., J. L. HARPER & C. R. TOWNSEND. 1986. Ecology, Individuals, Populations and
Communities. Third ed. Blackwell Science, London. BOKERMANN, W. C. A. 1952. Microhylidae da coleção do Departamento de Zoologia
(Amphibia – Anura). Papéis Avulsos do Departamento de Zoologia, São Paulo, 10(16): 271-292. BURNHAM, K. P., & W. S. OVERTON. 1979. Robust estimation of population size
when capture probabilities vary amoung animals. Ecology 60: 927-936. CADLE, J. E. & J. L. PATTON. 1988. Distribution patterns of some amphibians, reptiles
and mammals of the eastern andean and southern Peru. Pp. 225-244 in VANZOLINI, P. E. & W. R. HEYER (eds.). Proceedings of a Workshop on Neotropical Distribution Patterns. Academia Brasileira de Ciências, Rio de Janeiro.
COLWELL, R. K. 1997. EstimateS: Statistical estimation of species richness and shared species from samples. Version 5. User’s Guide and application published at: http://viceroy.eeb.uconn.edu/estimates.
CRUZ, C. A. G.; U. CARAMASCHI & E. IZECKSOHN. 1997. The genus Chisamocleis Méhely, 1904 (Anura, Microhylidae) in the Atlantic Rain Forest of Brazil, with a description of three new species. Alytes 15: 49-71.
DAPTF. 1999a. Declining Amphibian Populations Task Force (DAPTF) Media Briefing 54th March 1998. http://www2.open.ac.uk/Ecology/J_Baker/DAPTF.Press.B5.html Visitado em junho de 1999.
DAPTF. 1999b. The Declining Amphibian Populations Task Force (DAPTF) Home Pages. http://www2.open.ac.uk/Ecology/J_Baker/JBtxt.html Visitado em junho de 1999.
DAPTF. 1999c. Declining Amphibian Populations Task Force (DAPTF) Future Goals. http://www2.open.ac.uk/Ecology/J_Baker/DAPTF.Future.html Visitado em junho de 1999.
DUELLMAN, W. E. 1988. Patterns of species diversity in anuran amphibians in the american tropics. Ann. Missouri Bot. Gard. 75:79-104.
FAUTH, E. F., C. I. BRIAN, and J. B. SLOWINSKI. 1989. Elevational patterns of species richness, evenness and abundance of the Costa Rican leaf-litter herpetofauna. Biotropica 21:178-185.
FRANÇA, A. 1951. A Ilha de São Sebastião. Estudo de Geografia Humana. Univ. Press. FFLCH - USP, São Paulo.
FROST D. R. (ed.) 1985. Amphibian Species of the World. Assoc. Coll. Lawrence, Kansas. GIARETTA, A. A. 1999. Diversidade e densidade de anuros de serapilheira num gradiente
altitudinal na Mata Atlântica costeira. Universidade Estadual de Campinas. Tese de doutoramento.
GIARETTA, A. A & R. J. SAWAYA. 1998. A second species of Psyllophryne (Anura-Brachycephalidae). Copeia, 1998(4): 985-987.
GIARETTA, A. A.; R. J. SAWAYA; G. MACHADO; M. S. ARAÚJO; K. G. FACURE; H. F. MEDEIROS & R. NUNES. 1997. Diversity and abundance of litter frogs at altitudinal sites at Serra do Japi, Southeastern Brazil. Revta. Bras. Zool. 14(2): 341-346.

64
GIARETTA, A. A.; K. G. FACURE; R. J. SAWAYA; J. MEYER & N. CHEMIN. Biotropica, no prelo. Diversity and abundance of litter frogs of a montane forest in Southeastern Brazil: seazonal and altitudinal changes.
GOMES, M. J. & M. J. RAMOS. 1994. Ilhabela. Ibla Gráfica e Editora, São Paulo. GROMBONE, M.T.; L.C. BERNACCI ; J. A. A. MEIRA-NETO ; J. Y. TAMASHIRO
H. F. LEITÃO-FILHO. 1990. Estrutura fitossociológica da floresta semidecídua de altitude do Parque Municipal da Grota Funda (Atibaia-Estado de São Paulo). Acta bot. bras., 4 (2): 47-64.
HADDAD, C. F. B. & I. SAZIMA. 1992. Anfíbios anuros da Serra do Japi. Pp. 188-210 in L. P. C. MORELLATO (org.). História Natural da Serra do Japi: Ecologia e Preservação de uma Área Florestal no Sudeste do Brasil. Editora da Unicamp/FAPESP, Campinas.
HEINEN, J. T. 1992. Comparisons of the leaf litter herpetofauna in abandoned cacao plantations and primary rain forest in Costa Rica: some implications for faunal restoration. Biotropica 24:43-439.
HEYER, W. R. 1988. On frog distribution patterns east of the Andes. Pp. 245-273 in VANZOLINI, P. E. & W. R. HEYER (eds.). Proceedings of a Workshop on neotropical distribution patterns. Academia Brasileira de Ciências, Rio de Janeiro.
HEYER, W. R.; A. S. RAND; C. A. G. CRUZ; O. L. PEIXOTO & C. E. NELSON. 1990. Frogs of Boraceia. Arq. Zool., São Paulo, 31:231-410.
HEYER, W. R.; M. A. DONNELLY; R. W. MCDIARMID; L. C. HAYEK & M. S. FOSTER (eds). 1994. Measuring and Monitoring Biological Diversity. Standard Methods for Amphibians. Smithsonian Institution Press, Washington.
HURLBERT, H. S. 1971. The nonconcept of species diversity: a critique and alternative parameters. Ecology 52: 577-586.
IBGE. 1983. Projeto Radambrasil. Folhas SF 23/24 Rio de Janeiro/Vitória. Ministério das Minas e Energia. Secretaria-Geral, Rio de Janeiro.
INGER, R. F. 1980. Densities of floor-dwelling frogs and lizards in lowland forests of Southeast Asia and Central America. American Naturalist 115:761-770.
JAEGER, R. G. & R. F. INGER. 1994. Standard techniques for inventory and monitoring: Quadrat Sampling. Pp. 97-102 in HEYER, W. R.; M. A. DONNELLY; R. W. MCDIARMID; L. C. HAYEK & M. S. FOSTER (eds), Measuring and Monitoring Biological Diversity. Standard Methods for Amphibians. Smithsonian Institution Press, Washington.
JANZEN, D. H. 1973. Sweep samples of tropical foliage insects: Description of study sites, with data on species abundances and size distributions. Ecology 54: 659-686.
KREBS, C. J. 1985. Ecology: The Experimental Analysis of Distribution and Abundance. Third ed. Harper & Row, New York.
KREBS, C. J. 1989. Ecological Methodology. Harper and Row, Publishers, New York. KUENOUILLE, M. H. 1956. Notes on bias in estimation. Biometrika 43:353-360. LIEBERMAN, S. S. 1986. Ecology of the leaf litter herpetofauna of a neotropical rain
forest: La Selva, Costa Rica. Acta Zoológica Mexicana 15:1-72. LLOYD, M., R. F. INGER, and F. W. KING. 1968. On the diversity of reptile and
amphibian species in a Bornean rain forest. American Naturalist 102:497-515. LOVEJOY, T. E. 1995. The quantification of biodiversity: an esoteric quest or a vital
component of susteinable development?. Pp. 81-87 in HAWKSWORTH, D. L. Biodiversity. Measurements and Estimation. Chapman and Hall, London.
LUEDERWALDT, H. 1929. Resultados de uma excursão scientífica a Ilha de São Sebastião no littoral do Estado de São Paulo em 1925. Rev. Mus. Paul. 16: 1-81.
MACARTHUR, R. H. & E. O. WILSON. 1967. The Theory of Island Biogeography. Princeton Univ. Press., Princeton, New Jersey.

65
MANLY, B. F. J. 1991. Randomization, Bootstrap and Monte Carlo Methods in Biology. Second ed. Chapman & Hall. London.
MARTINS, M. R. C. 1994. História natural e ecologia de uma taxocenose de serpentes de mata na região de Manaus, Amazônia Central, Brasil. Universidade Estadual de Campinas. Tese de doutoramento.
MAGURRAN, A. E. 1988. Ecological Diversity and its Measurements. Princeton University Press., New Jersey.
MAY, R. M. 1975. Patterns of species abundance and diversity. Pp. 81-120 in CODY, M. L. and J. M. DIAMOND (eds.), Ecology and Evolution of Communities. Harvard University Press., Cambridge.
MAY, R. M. 1980. Why are fewer frogs and lizards in Southeast Asia than in Central America? Nature 287. September 11th, 1980.
MÜLLER, P. 1968. Die Herpetofauna der Insel von São Sebastião (Brasilien). Saarbrücker Zeitung, Verlag und Druckerei GmbH, Germany.
PIANKA, E. R. 1966. Latitudinal gradients in species diversity: a review of concepts. The American Naturalist 100: 33-46.
PIANKA, E. R. 1994. Evoloutionary Ecology. Fifth ed. Harper Collins College Publishers. Austin, Texas.
POUGH, F. H.; R. M. ANDREWS; J. E. CADLE; M. L. CRUMP; A. H. SAVITZKY; K. D. WELLS. 1998. Herpetology. Prentice-Hall, New Jersey.
RICKLEFS, R. E. 1990. Ecology. Third ed. Freeman and Company, New York. RODRIGUEZ, L. O. 1992. Structure et organization du peuplement d’anoures de Cocha
Cashu, Parc National Manu, Amazonie Péruvienne. Rev. Ecol. (Terre Vie) 47: 151-197. SCOTT, N. J., Jr. 1976. The abundance and diversity of the herpetofaunas of tropical
forest litter. Biotropica 8:41-58. SMA. Projeto de Preservação da Floresta Tropical (Mata Atlântica)
SMA - KFW. Componente B: Consolidação das Unidades de Conservação. Projeto Piloto: Subcomponente B5. Versão 2 - 07/11/1994. Não Publicado.
STATSOFT. 1998. STATISTICA for Windows, Release 5.1 J. StatSoft, Inc., Tulsa. TER BRAAK, C. F. 1987. CANOCO - a Fortran program for canonical community
ordination by partial detrended canonical correspondence analisys, principal component analisys (version 2.1). Wageningen, ITI-TNO.
TOFT, C. A. 1982. Community structure of litter anurans in a tropical forest, Makokou, Gabon: a preliminary analysis in the minor dry season. Rev. Ecol. (Terre Vie) 36: 223-232.
VITT, L. J. & J. P. CALDWELL. 1994. Resource utilization and guild structure of small vertebrates in the Amazon forest leaf litter. J. Zool. 234: 463-476.
WAKE, D. B. 1994. Foreword. in HEYER, W. R.; M. A. DONNELLY; R. W. MCDIARMID; L. C. HAYEK & M. S. FOSTER (eds), Measuring and Monitoring Biological Diversity. Standard Methods for Amphibians. Smithsonian Institution Press, Washington.
WILSON, E. O. 1992. Estrategia de conservación de la biodiversidad. Pp. 19-36 in Estrategia Global para la Biodiversidad: Pautas de Acción para Salvar, Estudiar y Usar en Forma Sostenible y Equitativa la Riqueza Biótica de la Terra. WRI/UICN/PNUMA.
ZAR, J. H. 1996. Biostatistical Analysis. Third ed. Prentice-Hall, Upper Saddle River, New Jersey.
ZIMMERMANN, B. L. e M. T. RODRIGUES. 1990. Frogs, Snakes, and Lizards of the INPA/WWF reserves near Manaus, Brazil. Pp. 426-454. in A. H. GENTRY (ed.), Four Neotropical Rainforests. Yale University Press, New Haven.