comportamento diurnas em cativeiro na...
TRANSCRIPT

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
COMPORTAMENTO REPRODUTOR DE RAPINAS
DIURNAS EM CATIVEIRO NA COUDELARIA ALTER
REAL E EM VAIAMONTE
MANUEL ANTÓNIO PINTO SAPAGE
MESTRADO EM BIOLOGIA DA CONSERVAÇÃO
2011

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
COMPORTAMENTO REPRODUTOR DE RAPINAS
DIURNAS EM CATIVEIRO NA COUDELARIA ALTER
REAL E EM VAIAMONTE
MANUEL ANTÓNIO PINTO SAPAGE
Dissertação de Mestrado orientada por:
Prof. Doutor Luís Vicente
Dr. Carlos Manuel Crespo
MESTRADO EM BIOLOGIA DA CONSERVAÇÃO
2011


“In a desert land he found him, in a barren and howling waste. He shielded him and cared for
him; he guarded him as the apple of his eye, like an eagle that stirs up its nest and hovers over its
young, that spreads its wings to catch them and carries them on its pinions.”
Deuterenomy, 32:11-12
“I cannot here enter on the copious details which I have collected on this curious subject; but to show
how singular the laws are which determine the reproduction of animals under confinement, I may just
mention that carnivorous animals, even from the tropics, breed in this country pretty freely under
confinement, with the exception of the plantigrades or bear family; whereas, carnivorous birds, with the
rarest exceptions, hardly ever lay fertile eggs.”
Charles Darwin, On The Origin Of Species


Agradecimentos:
Este trabalho não seria de todo possível sem o contributo de inúmeras pessoas. Desta maneira quero agradecer:
Ao meu orientador, Prof. Doutor Luís Vicente, por ter aceite orientar a minha tese e por me ter dado a oportunidade de trabalhar com aves de rapina.
Ao meu orientador, Dr. Carlos Manuel Crespo, pelo seu apoio incondicional prestado ao longo de toda a tese e pelos seus valiosos conselhos que enriqueceram o meu conhecimento e a minha vontade de trabalhar neste tema.
À Dra. Susana Varela, a quem eu não devo só um, mas dois, três ou mais agradecimentos por ser a pessoa que teve a paciência e a simpatia de me corrigir a tese e de me auxiliar no tratamento estatístico dos dados, aumentando assim a sua qualidade científica do meu trabalho.
À Fundação Alter Real, por me ter acolhido de braços abertos, dando-me alojamento ao longo de todo período do trabalho.
Aos meus colegas da Falcoaria, Nuno Mimoso, José Maia e Pedro Barradas, que me acompanharam ao longo do trabalho e me deram muita informação acerca das aves estudadas.
Aos meus amigos de Alter do Chão, entre eles, Ricardo Alves, Ricardo Antunes, Duarte Nogueira, Cláudia Marques, Fábio Salgado e muitos outros, pela sua companhia nas horas vagas que seriam insuportáveis passá-las sozinho.
À Nini (Ana Rita Fragoso) por ter sido a pessoa que mais me pressionou para escrever a tese a tempo.
Ao pessoal “Show das Aves” do Jardim Zoológico de Lisboa onde sou voluntário, local onde comecei a trabalhar e a interessar-me pelas aves de rapina. Foram eles que me revelaram o prazer de trabalhar com aves de rapina e me deram muita informação acerca do maneio das mesmas. Principalmente a minha querida “chefa” “Clarinha” Ferreira e ao treinador Hugo Condez.
Um agradecimento muito especial ao Sr. Silvino Gomes, que embora já não esteja entre nós, que foi a pessoa que me introduziu ao mundo animal, aceitando-me como voluntário do Jardim Zoológico de Lisboa.
Ao Flávio Barros, orientando de Douturamento do Prof. Doutor Luís Vicente, que me convenceu ser orientando do mesmo, algo que nunca arrependerei.
Aos meus pais que foram aqueles que mais me apoiaram ao longo de todo o trabalho, quer em termos financeiros quer em termos morais, e que compreenderam o pouco tempo que tive dedicado a eles.
Às aves de rapina que manejo, que, embora sendo pouco ortodoxo o agradecimento aos próprios animais, não deixam de ser a essência, aquilo que me dá gosto e paciência para a concretização deste trabalho e possivelmente de muitos outros que virão no futuro.
E claro, a tantas outras pessoas que de uma maneira ou outra me ajudaram na concretização do trabalho, às vezes apenas com um simples desejo de “boa sorte na tese”. A estas pessoas manifesto do fundo do meu coração um “muito obrigado”.


ÍNDICE
RESUMO.......................................................................................................... 1
ABSTRACT…………………………………………………………………………..2
1. INTRODUÇÃO................................................................................................... 3 1.1. Taxonomia das aves de rapina................................................................... 3 1.2. Criação de aves de rapina diurnas em cativeiro para a falcoaria……........ 3 1.3. Métodos de reprodução de aves de rapina diurnas em cativeiro……….... 4 1.4. Biologia reprodutora das aves de rapina diurnas na natureza................... 6 1.5. Biologia das espécies estudadas no trabalho............................................. 8 1.6. Relevância deste estudo............................................................................. 11 1.7. Objectivos................................................................................................... 11
2. METODOLOGIA............................................................................................... 13 2.1. População e local de estudo....................................................................... 13 2.2. Recolha de dados....................................................................................... 14 2.3. Análises estatísticas....................................................................................16 2.4. Hipóteses estatísticas................................................................................ 18
3. RESULTADOS.................................................................................................. 20 3.1. Registos obtidos…………………………………………………………………20 3.2. As posturas dos ovos, o sucesso reprodutor e o comportamento............. 20 3.3. Esforço parental.......................................................................................... 21
4. DISCUSSÃO...................................................................................................... 29 4.1. As posturas dos ovos, o sucesso reprodutor e o comportamento.............. 29 4.2. Esforço parental.......................................................................................... 32
5. Considerações finais………………………………………………………………… 37 5.1. Perspectivas – Dados do Estudo em Relação ao Sucesso Reprodutor…. 37 5.2. Aspectos técnicos....................................................................................... 39
6. REFERÊNCIAS BIBLIOGRÁFICAS................................................................. 40

7. ANEXOS............................................................................................................ 43 7.1. Anexo I........................................................................................................ 44 7.2. Anexo II....................................................................................................... 45

1
RESUMO
Neste estudo foram analisados, entre Fevereiro e Junho de 2009, os comportamentos reprodutores de quatro casais de aves de rapina diurnas mantidas em cativeiro na Coudelaria Alter Real e em Vaiamonte, onde existem instalações de reprodução destas aves para a prática da falcoaria. O objectivo principal do estudo foi comparar o esforço parental dos casais entre duas posturas e seus períodos de pré-incubação, incubação e cuidados com as crias, utilizando diversas variáveis baseadas na frequência de visitas e proporção de tempo passado no ninho. A intenção era avaliar o efeito do insucesso da primeira postura (por causas naturais ou por remoção artificial de ovos) na produtividade das aves de rapina durante a segunda postura. Esperava-se que após insucesso da primeira postura os casais investissem na segunda com esforço igual ou superior ao da primeira, de modo a garantir o sucesso reprodutor.
As aves estudadas foram as seguintes: um casal de búteos de Harris (Parabuteo
unicinctus), um casal de falcões-peregrino (Falco peregrinus), um casal de falcões-sacre (F. cherrug) e um casal de falcões híbridos de gerifalte-sacre (F. rusticolus X F. cherrug) sendo o macho 75% falcão-gerifalte e a fêmea 50% falcão-gerifalte.
Nenhum destes casais conseguiu obter sucesso reprodutor. As causas desse insucesso
parecem ter sido diferentes para todos os casais e independentes umas das outras. Para além disso, apenas os búteos de Harris tiveram duas posturas, com investimento reduzido na segunda, ao contrário do esperado. Este resultado deve-se, provavelmente, ao comportamento do macho, cujo esforço parental foi menor na segunda postura em quase todas as variáveis.
Os resultados obtidos permitem inferir sobre algumas técnicas e condições de
reprodução das aves de rapina em cativeiro, mas não podem ser generalizados sem um estudo mais aprofundado e um esforço de amostragem maior – foram observados poucos casais e foi amostrada uma época reprodutiva apenas.
Palavras-chave: Aves de rapina diurnas; Comportamento reprodutor; Cativeiro; Falcoaria; Esforço parental, Segundas posturas.

2
ABSTRACT
In this work the reproductive behavior of four couples of diurnal birds of prey, maintained in captivity in Coudelaria Alter Real and in Vaiamonte, where they are bred for falconry purposes, were studied between February and June of 2009. The main goal of the study was to compare the parental effort of the couples between two clutches and their periods of pre-incubation, incubation and chicks’ rearing, using several variables based on the frequency of visits to the nest and on the proportion of time spent in the nest. The idea was to evaluate the failure effect of the first clutch (due to natural causes or to the artificial removal of the eggs) on the birds’ productivity in the second clutch. After the failure of the first clutch, a similar or greater investment was expected in the second clutch, in order to guarantee reproductive success.
The studied couples were as follows: one couple of Harris’ hawks (Parabuteo
unicinctus), one couple of peregrine falcons (Falco peregrinus), one couple of saker falcons (F. cherrug) and one couple of hybrids gyr-saker falcons (F. rusticolus X F. cherrug), where the male is 75% and the female 50% gyr falcon.
None of the couples studied did reproduce successfully. The causes of failure seem to
have been different for every couple and independent from each other. Moreover, only the Harris’ hawks laid two clutches, with less investment in the second, contradicting our predictions. This result reflects probably the behavior of the male that reduced his effort on almost all the variables during the second clutch.
The obtained results allow to infer on some techniques and breeding conditions of
these birds in captivity, but they can’t be generalized without a deeper study and a greater sampling effort – few couples were observed and only one breeding season was sampled.
Keywords: Diurnal birds of prey; Reproductive behavior; Captivity; Falconry; Parental Effort; Second Clutches.

3
1. INTRODUÇÃO
1.1. Taxonomia das aves de rapina
As aves de rapina consistem num grupo polifilético criado por Lineu no século XVIII para caracterizar espécies com certas características em comum, como o bico adunco ou as garras mais ou menos desenvolvidas (Heidenreich, 1997). Na segunda metade do século XIX este grupo foi dividido em dois, as aves de rapinas nocturnas e as aves de rapina diurnas, separação a que os estudos filogenéticos por hibridação do ADN e outras técnicas moleculares, realizados a partir dos anos 80 do século XX, vieram dar razão (e.g. Sibley & Ahlquist, 1990).
O grupo das aves de rapina diurnas possui uma taxonomia relativamente controversa, existindo várias classificações para o grupo (Amaddon & Bull, 1988; Sibley & Ahlquist, 1990; Heidenreich, 1997; Fergunson-Lees & Christie, 2001). Neste trabalho considero as aves de rapina diurnas como sendo as aves pertencentes à ordem Falconiformes seguindo a classificação de Amaddon & Bull (1988).
1.2. Criação de aves de rapina diurnas em cativeiro para a falcoaria
A falcoaria consiste numa modalidade cinegética em que o Homem utiliza algumas espécies de aves de rapina diurnas como instrumento de caça. Esta modalidade surgiu na Ásia Central no período do Neolítico (Crespo, 1999).
A Idade Média foi a época de ouro da falcoaria (Almaça, 1997) e esta modalidade manteve-se popular até ao final do século XVIII. Com a Revolução Francesa (Crespo, 1999) e a Revolução Industrial (Beebe, 1992) a prática da falcoaria decresceu rapidamente. Com o desenvolvimento das armas de fogo esta modalidade começou a perder muitos adeptos (Fox 1995; Crespo 1999). A caça com armas de fogo era de tal maneira eficaz que contribuiu para o extermínio de algumas aves de rapina (Fox, 1995).
As aves de rapina também sofreram ameaças como o DDT (Dicloro-Difenil-Tricloroetano), principalmente nas populações de falcão-peregrino (Falco peregrinus), que surgiram com o desenvolvimento da agricultura após a Segunda Guerra Mundial (Fox, 1995).
Com a diminuição das populações de aves de rapina, a legislação para a sua protecção começou a ficar cada vez mais exigente. Nos finais dos anos 70, os falcoeiros perceberam que a falcoaria não poderia ser auto-sustentável sem a reprodução em cativeiro (Fox, 1995) e a criação em cativeiro tornou-se imperativa na manutenção da actividade.
Apesar do primeiro caso de reprodução de aves de rapina diurnas com sucesso em cativeiro ter sido feito pelo falcoeiro alemão Waller, em 1942-43, com falcões-peregrino, os

4
primeiros dados consistentes da reprodução em cativeiro foram obtidas através da reprodução do peneireiro-comum e do peneireiro-americano (F. tinnunculus e F. sparverius) nos finais dos anos 60 (Cade et al., 1977).
Actualmente as técnicas de reprodução de aves de rapina foram desenvolvidas maioritariamente por falcoeiros. Mais de 90% dos esquemas de reprodução das aves de rapina foram iniciadas por pessoas com interesse em falcoaria. Foi igualmente graças à falcoaria que foi desenvolvido muito do conhecimento acerca das doenças destas aves (Perry-Jones, 2005).
As aves de rapina que são reproduzidas em cativeiro vivem em condições muito diferentes das encontradas na natureza. Estas aves são confinadas em câmaras de reprodução de tamanho limitado, o que limita igualmente a amplitude dos voos que as aves podem efectuar.
Um dos métodos de cortejamento das aves de rapina consiste em exibições de voos entre o casal. Nos falcões peregrinos, por exemplo, este comportamento vai aumentando de frequência até à postura dos ovos (Ratcliffe, 2000). Em cativeiro as exibições de voo são seriamente limitadas (Olendorf, 1972).
Outro comportamento que está também condicionado em cativeiro consiste na oferta de alimento para o parceiro (Park at al., 2003). Na natureza as aves caçam as suas próprias presas e oferecem-nas ao parceiro, enquanto que em cativeiro é o próprio criador que decide qual é o alimento a dar e quando é que o alimento é dado. Algumas vezes, como o macho não precisa de encontrar alimento para a fêmea, estes podem entrar em competição para incubar os ovos (Fox, 1995).
Em cativeiro alguns dos comportamentos reprodutores são manifestados de forma mais condensada (Fox, 1995), provavelmente porque o macho e a fêmea estão quase permanentemente em contacto visual, algo que não acontece na natureza.
1.3. Métodos de reprodução de aves de rapina diurnas em cativeiro
A criação em cativeiro de aves de rapina assume hoje uma grande importância, pois esta é necessária para a prática da falcoaria e até mesmo para a reintrodução de indivíduos na natureza.
Existem vários métodos para a reprodução de aves de rapina em cativeiro que vão desde a simulação do meio natural, até a métodos totalmente artificiais, que utilizam a inseminação artificial e a incubação de ovos em incubadora (Crespo, 1999).
O método mais natural consiste na colocação de um casal reprodutor em câmaras de reprodução, também conhecidas por “mudas”.

5
Neste método o casal constrói o ninho ou arranja um ninho previamente construído, executa os comportamentos de corte, acasala, coloca os ovos no ninho, incuba-os e cuida das crias da forma mais natural possível, apesar de estar confinado ao espaço limitado da câmara de reprodução.
No método mais artificial é utilizado um macho e uma fêmea com imprinting sexual ao ser humano, em que é retirado sémen ao macho e colocado na cloaca da fêmea através do método de inseminação artificial. Os ovos assim produzidos são colocados numa incubadora artificial e posteriormente as crias são criadas pelo homem.
Os dois métodos acima descritos não são mutuamente exclusivos. As etapas podem ser intercaladas de modo a criar um método semi-natural ou semi-artificial, e.g., os ovos podem ser obtidos por inseminação artificial, mas colocados no ninho de um casal reprodutor para incubação e posteriormente retirados como pintos para a criação pelo ser humano. Todas as combinações são teoricamente possíveis, embora existam algumas etapas que sejam difíceis de passar do método artificial para o método natural, pois é necessária a aceitação do casal dos avos por parte do casal.
Ambos os métodos possuem aspectos positivos e negativos, de modo que se deve ponderar, com precaução, a importância do sucesso reprodutivo de cada ave e os meios disponíveis, antes de escolher o método a utilizar.
A reprodução artificial pode ser mais produtiva em termos do número de indivíduos reproduzidos e do número de crias que sobrevivem, embora possa criar um problema de imprinting com o ser humano. As aves podem igualmente rejeitar parceiros da sua própria espécie, o que dificulta a sua reintrodução na natureza quando criadas por este método.
A reprodução natural, pelo contrário, é um método menos produtivo, mas as crias crescem juntamente com os seus progenitores e adquirem comportamentos mais semelhantes àqueles ocorrentes no meio natural.
A época de reprodução, na maioria das aves de rapina, é anual, o que torna a
produtividade destas aves relativamente baixa. A técnica mais comum para o aumento dessa produtividade consiste na remoção dos primeiros ovos, de modo a provocar uma reposição da postura. Esta técnica é utilizada para o aumento da produtividade de várias espécies de aves (Fox, 1995).
Existem dois métodos para a remoção dos ovos do ninho. O primeiro método, conhecido como egg pulling, consiste em retirar os ovos à medida que a fêmea os vai depositando. Neste caso a fêmea tende sempre a colocar mais ovos de maneira a completar a postura (que varia consoante as espécies). Este método não é muito aconselhado, porque à medida que a fêmea vai depositando mais ovos, terá também que disponibilizar cada vez

6
mais nutrientes do seu organismo, esgotando-a fisiologicamente. A qualidade dos ovos é também afectada, pois à medida que estes vão sendo depositados, a quantidade de cálcio na casca vai diminuindo (Fox, 1995, Heidenreich, 1997).
O segundo método, mais aconselhável, consiste no retiro dos ovos alguns dias após a postura completa. Deste modo a fêmea apenas tentará depositar mais ovos após algumas semanas, o que permite preparação fisiológica para este encargo.
1.4. Biologia reprodutora das aves de rapina diurnas na natureza
O comportamento de reprodução na natureza é bastante diverso e relativamente complexo. Pensava-se, inicialmente, que muitas destas aves possuíam um sistema social exclusivamente monogâmico. Contudo, cada vez mais são encontrados dados de espécies que podem formar um sistema de poliginia (Korpimäki, 1988) e existem algumas espécies com o sistema de poliandria (e.g. Dawson & Mannan, 1991).
A maioria das aves de rapina possui um dimorfismo sexual invertido significativo, sendo a fêmea maior do que o macho, tornando-a dominante na relação (Amadon, 1975). Apesar disso, a fêmea não tem a capacidade de incubar os ovos e criar as crias na ausência do macho, o que torna importante para a fêmea a escolha de um macho com um grande território e uma boa capacidade de caça. Presume-se que muitos dos comportamentos de exibição do macho auxiliam a fêmea na procura de um parceiro (Olsen & Olsen, 1987).
Segundo Olsen & Olsen (1987), a maioria dos grupos poligâmicos nas aves de rapina acontecem em locais de reprodução com fracos recursos, fazendo com que a fêmea escolha mais do que um parceiro para aumentar a quantidade de comida trazida para o ninho. O sistema social não deve ser confundido com a reprodução cooperativa. Esta consiste em grupos onde vários indivíduos ajudam o casal a procurar alimento ou a cuidar das crias. Neste caso pode não existir cópula entre os indivíduos ajudantes e o casal reprodutor. Nas aves de rapina diurnas este comportamento ocorre em 29% dos géneros e em 14% das espécies (Kimball et al., 2003).
As aves de rapina diurnas possuem locais de nidificação muito distintos, mas estes podem ser divididos em dois grupos principais. Os falcões (família Falconidae) são aves que não constroem activamente o ninho. Os seus locais de nidificação consistem essencialmente em cavidades naturais ou apoderam-se dos ninhos de outras aves (Harrison & Castell, 2002). São aves bastante flexíveis em relação aos locais escolhidos para a nidificação, pois existem algumas espécies que nidificam em estruturas artificiais, como em pontes, em edifícios abandonados, ou em arranha-céus (Ellis et al., 1997; Herbert & Herbert, 1965), embora no seu habitat natural muitas espécies nidifiquem em escarpas ou falésias (Ratcliffe, 2000). As águias e os abutres (família Accpipitridae) são aves que constroem activamente o ninho,

7
normalmente em forma de grandes taças de paus e detritos, nas árvores, nas escarpas ou no chão (Harrison & Castell, 2002).
Algumas destas aves possuem vários ninhos no seu território (Ontiveros et al. 2008). Dependendo das aves de rapina, estas podem nidificar em colónias ou serem
solitárias, mas em geral as aves de rapina tornam-se mais agressivas durante o ciclo reprodutor, principalmente antes do período das posturas. Muitas aves de rapina diurnas defendem um território, impedindo a entrada de competidores (Wiącek, 2006).
O comportamento de cortejamento na natureza das aves de rapina diurnas são
diferentes de espécie para espécie e até mesmo de subespécie para subespécie (Wrege & Cade, 1977), mas existem algumas características gerais ocorrentes na maioria das espécies.
Antes da deposição dos ovos, Fyfe (1978) dividiu o comportamento do falcão-da-pradaria (Falco mexicanus) e do falcão-peregrino (F. peregrinus) em três fases que se podem aplicar à maioria destas aves, existindo algumas variantes e/ou ausência de alguns comportamentos consoante as espécies: (1) atracção de parceiros, (2) estabelecimento da relação e (3) preliminares da nidificação.
Na fase da atracção de parceiros, o macho ou a fêmea procuram poisos conspícuos e visitam locais potenciais de nidificação.
Na fase do estabelecimento da relação, o casal mantém-se no território; exibe voos de cortejamento; caça cooperativamente e poisa dentro do território ou no ninho. O macho continua a visitar os locais potenciais de nidificação; vocaliza para a fêmea; atrai a fêmea para o local de nidificação levando comida para o ninho; exibe-se para a fêmea no ninho e leva comida para a fêmea (transferência de comida). O casal come em conjunto e existe interacção entre o macho e a fêmea (arranjam as penas um do outro, bicam o bico um do outro, etc.).
Na fase dos preliminares da nidificação, a fêmea pede comida vocalizando e o casal começa a escavar o ninho (nas aves que constroem activamente o ninho, o casal começa a trazer paus para o ninho), exibindo-se nele, ocorrendo, por fim, o comportamento de cópula.
As aves de rapina copulam muito mais frequentemente do que o necessário para
fertilizar os ovos. Calcula-se que em média as aves de rapina copulam 100 vezes por postura (Negro & Grande, 2001).
Existem várias teorias para a elevada frequência de cópulas nas aves de rapina diurnas. Segundo Møller & Birkhead (1992), este comportamento tem como função (1) aumentar a probabilidade de fertilização dos ovos; (2) fortalecer ou manter a relação entre o casal; (3) aproveitar a vantagem de baixo risco predatório e/ou (4) diluir o esperma dos

8
competidores. Os acasalamentos extra-conjugais já foram descritos para muitas espécies de aves de rapina coloniais e solitárias (Mougeot, 2004; Penadés, 2007). Negro & Grande (2001) sugerem que a elevada frequência de cópulas tem também como função anunciar a ocupação de um determinado território.
Na maioria das aves de rapina são as fêmeas que asseguram a incubação dos ovos e os
machos têm como função a procura de alimento para as fêmeas. Em muitas espécies o macho auxilia na incubação dos ovos, mas por menos tempo que a fêmea (Heidenreich, 1997). Estas aves têm também a capacidade de renovar a sua postura após insucesso da primeira (Fox, 1995).
Nas aves de rapina diurnas são necessários entre 27 e 54 dias de incubação até à eclosão dos ovos, dependendo das espécies, mas já foram vistas aves a incubarem os ovos até 163% do tempo de incubação necessário quando estes são inviáveis. Acredita-se que a incubação prolongada tenha como função assegurar a certeza de insucesso, mas é provável que uma incubação excessivamente prolongada seja uma má adaptação dos indivíduos (Magalida et al., 2006).
As aves de rapina são altriciais, necessitando dos cuidados parentais nas primeiras
semanas de vida. Nos falcões existe uma assincronia na eclosão dos ovos e geralmente as aves mais jovens não conseguem sobreviver caso haja falta de alimento, mas quase nunca por fratricídio (Harrison & Castell, 2002).
Normalmente são ambos os progenitores que cuidam das crias, trazendo comida para o ninho. Existem estudos em algumas aves de rapina, como a águia-real (Aquila chrysaetos) (Collopy, 1984), o gavião (Accipiter nisus) (Newton, 1978) ou o gavião-do-mississipi (Ictinia mississippiensis) (Botelho et al., 1993) em que os progenitores vão levando cada vez mais alimento para as crias durante a primeira metade do seu desenvolvimento no ninho, quantidade que decrescerá na segunda metade do desenvolvimento até as crias terem capacidade para sair do ninho e caçar com os pais.
1.5. Biologia das espécies estudadas no trabalho
1.5.1. Búteo de Harris (Parabuteo unicinctus)
O búteo de Harris é a espécie mais popular da falcoaria moderna. A sua popularidade deve-se ao facto de ser uma das aves de rapina diurnas mais fáceis de treinar e de reproduzir em cativeiro (Perry-Jones, 2005; Glasier, 1998; Heidenreich, 1997). Esta espécie está

9
distribuída no sul da América do Norte e em algumas regiões da América do Sul (Fergunson-Lees & Christie, 2001).
Nesta espécie as fêmeas são, em média, 9% maiores e 40% mais pesadas do que os machos, embora haja um intervalo de sobreposição entre os pesos (Fergunson-Lees & Christie, 2001).
Na época reprodutora, estas aves normalmente têm comportamento cooperativo, i.e., vivem no grupo mais do que dois indivíduos e todos ajudam a cuidar das crias. Estes grupos podem chegar até sete indivíduos e exibir comportamento de poliandria, embora o número de cópulas e o acesso dos indivíduos ao ninho dependa da hierarquia dos mesmos (Dawson & Mannan, 1991).
O búteo de Harris é uma espécie normalmente sedentária. Na natureza, durante a época não reprodutora, podem formar grupos sociais que normalmente atingem 10 a 15 indivíduos (Fergunson-Lees & Christie, 2001), pois estas aves caçam em grupo. Na natureza estas aves fazem posturas que variam normalmente entre os 2 a 4 ovos (podendo variar entre os 1 a 5 ovos), sendo a sua incubação de 33 a 36 dias. As crias abandonam o ninho aos 45 a 50 dias após a eclosão. A época de reprodução do búteo de Harris varia consoante a região geográfica, podendo chegar a um ano na Venezuela (Fergunson-Lees & Christie, 2001).
1.5.2. Falcão-peregrino (Falco peregrinus)
O falcão-peregrino tem uma distribuição quase mundial, sendo uma das aves com a maior distribuição latitudinal (78ºN a 56ºS). Devido a esta grande distribuição, esta espécie está dividida entre 16 a 22 subespécies, dependendo da classificação dos autores (Fergunson-Lees & Christie, 2001; Dementiev, 1951; Brown & Amandon, 1968).
Dos poucos registos de reprodução cooperativa nesta espécie, apenas foram identificadas fêmeas imaturas como ajudantes do casal reprodutor. Presume-se que estes indivíduos sejam fêmeas com um ano que, sendo filopátricas, voltaram ao local de reprodução (Kimball et al., 2003).
A época reprodutora desta espécie ocorre dependendo das condições climáticas, da disponibilidade de presas e da localização geográfica. Nas regiões temperadas do hemisfério Norte, a época reprodutora começa em Fevereiro e termina entre Março e Junho. As fêmeas de falcão-peregrino são 12% a 18% maiores e 40% a 70% mais pesadas do que os machos, dependendo das subespécies (Fergunson-Lees & Christie, 2001).
Na natureza estas aves fazem posturas que variam normalmente entre 3 a 4 ovos, sendo o seu período de incubação de 29 a 33 dias. As crias abandonam o ninho aos 35 a 42 dias após a eclosão (Fergunson-Lees & Christie, 2001).

10
1.5.3. Falcão-sacre (Falco cherrug)
O falcão-sacre é uma espécie que se encontra distribuída por partes da região Paleártica, Afro-Tropical e Indo-Malaia. Nesta espécie, as fêmeas são, em média, 15% maiores e 40% mais pesadas do que os machos. A época de reprodução ocorre de Março a Junho/Julho na região Oeste da distribuição e de Abril a Agosto na região Este da distribuição (Fergunson-Lees & Christie, 2001).
O comportamento cooperativo na reprodução não foi detectado nesta espécie (Kimball et al., 2003).
Na natureza estas aves fazem posturas que variam normalmente entre 3 a 5 ovos (podendo ser só de 2 ovos ou chegar aos 6) e o período de incubação é de cerca 30 dias. As crias abandonam o ninho aos 45 a 50 dias após a eclosão (Fergunson-Lees & Christie, 2001), semelhante ao búteo de Harris.
1.5.4. Falcão-gerifalte (Falco rusticolus)
O falcão-gerifalte é uma espécie que se encontra distribuída por todo o Círculo Polar Ártico. Nesta espécie as fêmeas são, em média, 7% maiores e 66% mais pesadas (Fergunson-Lees & Christie, 2001) do que os machos. Sendo o maior dos falcões, esta é uma das aves mais apreciadas da falcoaria. Na idade média era inclusivamente considerado para o uso exclusivo do rei em alguns países (Crespo, 1999; The Peregrine Fund, 2010). A época de reprodução ocorre de Abril a Agosto, fazendo posturas de 3 a 4 ovos (podendo variar entre os 2 a 7 ovos) e um período de incubação de 34 a 36 dias. As crias abandonam o ninho aos 46 a 50 dias (Fergunson-Lees & Christie, 2001).
1.5.5. Falcões híbridos de gerifalte-sacre (F. cherrug X F. rusticolus)
A hibridização de Falconiformes na natureza é relativamente rara (Oliphant, 1991), mas estas aves são muito hibridadas para a prática da falcoaria. Muitas das aves de rapina diurnas têm o potencial para a hibridação produzindo inclusivamente descendência fértil, mas esta prática tem levantado muitas questões a nível ético, principalmente devido à poluição genética que estes podem causar às populações naturais caso fujam do cativeiro (Fox, 1995; Fox, 2004). O risco de fuga é significativo, visto que estas aves são constantemente soltas para a prática da falcoaria.
Segundo Fox (2004), os híbridos de falcão-gerifalte e de falcão-sacre são muito utilizados na falcoaria porque são falcões maiores e mais velozes que o falcão-sacre puro e são mais tolerantes ao calor e mais resistentes às doenças do que o falcão-gerifalte. Este

11
mesmo autor afirma que as duas espécies podem ser consideradas uma super-espécie em que os híbridos aparentam reproduzir-se prontamente e sem limitações em cativeiro.
1.6. Relevância deste estudo
O estudo dos comportamentos reprodutores das aves de rapina em cativeiro é essencial para a optimização das técnicas de conservação ex-situ destas espécies, visto que a reprodução em cativeiro falha normalmente devido a problemas de comportamento (Sutherland, 1998; Nelson, 1972). Através da pesquisa bibliográfica efectuada, constatou-se a existência de poucos estudos científicos acerca da criação de aves de rapina em cativeiro, sendo a maioria dos trabalhos com recomendações para a reprodução destas aves baseada na observação empírica dos autores (e.g.: Todd, 1972; Cade et al., 1977; Harrell, 1972).
O desenvolvimento e divulgação de estudos científicos deste tema são relevantes para
melhorar a gestão da reprodução destes animais em cativeiro, principalmente por se tratar de espécies com apenas uma época reprodutiva por ano. A reprodução em cativeiro de aves de rapina, para além de produzir exemplares para a falcoaria, é, igualmente, uma ferramenta poderosa a favor da conservação (Saint-Jaime, 2002) e tem sido utilizado para a reintrodução de aves de rapina, como por exemplo do falcão-peregrino em Nova Iorque (The Peregrine Fund, 2010), ou do peneireiro-das-torres em Portugal (LPN, 2010).
1.7. Objectivos
Este trabalho consiste no estudo dos comportamentos reprodutores de vários casais de aves de rapina mantidas em cativeiro na Coudelaria Alter Real e em Vaiamonte. O objectivo principal é estudar os comportamentos reprodutores destas aves durante uma primeira e segunda posturas, de modo a medir possíveis diferenças no esforço parental do macho e da fêmea entre as duas posturas. A intenção é a de avaliar o efeito da perda natural de ovos e da eficácia da técnica da remoção artificial de ovos na produtividade das aves de rapina durante a segunda postura.
Prevê-se a ocorrência de uma segunda postura nos casais que tenham perdido a primeira, tal como acontece na natureza (Fox, 1995). Espera-se, nestes casos, que o esforço parental seja semelhante ou superior ao da primeira postura. Segundas posturas após o sucesso da primeira não deverão ocorrer dado o tempo de que os progenitores necessitam para cuidar das crias até à sua independência.

12
No caso de algum dos casais estudados não obter sucesso reprodutor em nenhuma das posturas pretende-se, como objectivo secundário, tentar relacionar esse insucesso com os comportamentos registados, de modo a identificar problemas para futuros melhoramentos das instalações e do maneio.
No caso de algum casal de aves de rapina estudada obter sucesso reprodutor, pretende-se, como objectivo secundário, comparar o tempo despendido pelo macho e pela fêmea a cuidar das crias e comparar o esforço de ambos com o dos casais que não obtiveram sucesso.
Tanto em casos de sucesso como de insucesso, espera-se que as fêmeas tenham um esforço parental superior ao dos machos a cuidar do ninho, dos ovos e das crias, comportamento comum à maior parte das aves de rapina, nas quais os machos exercem outras funções, como a procura de alimento (Heidenreich, 1997).

13
2. Metodologia
2.1. População e local de estudo
Este estudo foi realizado entre Fevereiro e Junho de 2009, em dois locais diferentes: na Coudelaria Alter Real, situada na freguesia de Alter do Chão (39°11’N, 7°39’W), e nas instalações da Caçamonte situadas na freguesia de Vaiamonte (39°5’N, 7°30’W), ambas as localidades situadas no Alto Alentejo.
Nestes dois locais foram estudados o comportamento de quatro casais de aves de rapinas diurnas: um casal de búteo de Harris (Parabuteo unicinctus); um casal de falcões-peregrino (Falco peregrinus); um casal de falcões-sacre (Falco cherrug) e um casal de híbridos de falcão-sacre e falcão-gerifalte (Falco cherrug x Falco rusticolus), sendo o macho 75% falcão-gerifalte e 25% falcão-sacre e a fêmea 50% falcão-gerifalte e 50% falcão-sacre (Tabela 1). As aves estudadas pertencem à empresa Caçamonte.
Todas as aves estudadas foram habituadas à presença humana, sendo aves já treinadas para a prática da falcoaria. Esta prática permite uma reprodução com maior sucesso em instalações mais artificiais (Fox, 1995) e também permite a diminuição do stress causado pela presença humana quando esta se mostra necessária.
As instalações da Coudelaria de Alter onde estão situados os casais de búteo de Harris e falcões-peregrino estudados, têm como dimensões 750cm de comprimento, 390cm de largura e 400cm de altura.
As instalações de Vaiamonte, onde estão situados os casais de falcões-sacre e híbridos de falcão-gerifalte e falcão-sacre, têm como dimensões 400cm de comprimento, 500cm de largura e 250cm de altura.
Todas as instalações possuem cerca de 25% de cobertura superior que permite a protecção da chuva e do sol, enquanto que o resto da instalação apenas possui uma rede metálica.
As paredes destas instalações são de tijolo e cimento, possuindo uma pequena abertura com vidro espelhado, de modo a permitir a observação das aves sem que estas sejam perturbadas e poleiros a diferentes alturas. O chão destas instalações é de cascalho, existindo no centro um recipiente com água para que as aves se possam banhar.
O local mais provável para a nidificação destes casais consiste em plataformas na zona coberta. Nos casais de búteos de Harris e de falcões-sacre, existe ainda em cima da plataforma um ninho artificial constituído por um cesto com paus.

14
Tabela 1: Características dos casais estudados
Casal Sexo Espécie Idade (anos)
Reprodução com sucesso em anos anteriores**
Observações
1 M Pu 15 Sim, com outra fêmea O macho é pai da fêmea.
F Pu 8 Não
2 M Fp 5 Não O casal foi criado junto desde pequeno,
com mais 6 crias nascidas numa incubadora.
F Fp 5 Não
3 M Fc 7 Sim, com a mesma fêmea F Fc 7 Sim, com o mesmo macho
4
M 75% Fr + 25% Fc
8 Sim, com a mesma fêmea* Macho com tamanho semelhante à fêmea.
F 50% Fr + 50% Fc
18 Sim, com o mesmo macho*
Legenda: Sexo: M – Macho, F – Fêmea; Espécie: Pu – Parabuteo unicinctus, Fp – Falco peregrinus, Fc – F. cherrug, Fr – Falco rusticolus; *Informação do criador anterior que não pôde ser verificada; **Entende-se reprodução com sucesso quando os progenitores criaram as suas crias até estas atingirem a sua independência.
Na instalação do casal de búteos de Harris espalharam-se por duas vezes alguns paus
no chão, antes do início do estudo e logo após o final da 1ª postura. Na plataforma do ninho dos falcões híbridos adicionou-se areia entre o dia 21 e 26 de Março.
As aves foram mantidas sempre dentro das instalações durante todo o período reprodutivo e de observações e com a menor perturbação humana possível, quer visual quer sonora.
As aves foram alimentadas diariamente através de uma abertura, que permitia colocar a comida no chão ou sobre uma plataforma. A comida fornecida consistia maioritariamente em carne de pombo e de codorniz, muitas vezes com um aditivo de vitaminas próprias para a reprodução destas aves. Antes de as aves colocarem os ovos, a alimentação consistia ocasionalmente em pintos do dia.
2.2. Recolha de dados
2.2.1. Método de gravação de imagens
A recolha de dados das aves de rapina foi feita através de gravações utilizando as câmaras de vídeo disponíveis nas instalações. As câmaras das instalações da Coudelaria de Alter captam imagens a preto e branco e são gravadas num gravador de VHS. As câmaras das

15
instalações de Vaiamonte captam imagens a cores que são enviadas através de um sistema sem fios a um gravador de DVD. As gravações foram feitas durante 20 semanas, tendo começado a 17 de Fevereiro de 2009 e terminado a 30 de Junho de 2009. Durante este período tentou-se obter pelo menos 8 horas de gravação por semana e por casal. Ficou estabelecido à partida que o término do estudo por casal aconteceria ou no final do período reprodutivo, assim que a sobrevivência das crias estivesse assegurada, ou, no caso de insucesso na reprodução, na ausência de posturas, ou no final da primeira ou da segunda posturas (caso ocorresse uma segunda), ou após o período de incubação.
2.2.2. Método de obtenção de dados
No presente trabalho considerou-se como indicadores do esforço parental variáveis relacionadas com a tendência da permanência do casal no ninho, pois estas variáveis mostram o desempenho e investimento do casal na preparação do ninho – fase de pré-incubação – e na incubação dos ovos.
Para a obtenção das variáveis a analisar, as gravações obtidas foram visualizadas de forma a registar, por amostragem contínua, a hora de entrada e de saída do macho ou da fêmea do ninho (Anexos I e II). Com estes dados, calcularam-se as seguintes variáveis:
• Frequência de visitas do macho/fêmea ao ninho: Número de vezes que o macho e a fêmea frequentaram o ninho por hora.
• Proporção do tempo de permanência do macho/fêmea no ninho: Número de horas em que o macho e a fêmea permaneceram no ninho a dividir pelo número de horas totais.
• Frequência de ambos os indivíduos no ninho: Número de vezes que ambos os indivíduos foram encontrados ao mesmo tempo no ninho por hora.
• Proporção do tempo de permanência de ambos os indivíduos no ninho: Número de horas em que ambos os indivíduos permaneceram ao mesmo tempo no ninho a dividir pelo número de horas totais.

16
• Frequência de ocorrências do ninho vazio: Número de vezes que o ninho foi encontrado vazio por hora.
• Proporção do tempo em que o ninho se encontra vazio: Número de horas em que o ninho foi encontrado vazio a dividir pelo número de horas totais.
Nas fases em que os ninhos estavam com ovos, registou-se igualmente o tempo de incubação do macho e da fêmea, obtendo-se, deste modo, as seguintes variáveis.
• Frequência de incubação do macho/fêmea: Número de vezes que o macho e a fêmea incubaram os ovos por hora.
• Proporção do tempo de incubação do macho/fêmea: Número de horas em que o macho e a fêmea incubaram os ovos a dividir pelo número de horas totais.
A distinção entre o macho e a fêmea, no seio de cada casal, foi efectuada através de
diferentes características, algumas delas naturais – caracteres sexuais secundários e outros –, como o tamanho, a cor, o estado da plumagem e marcas várias na mesma, e outras artificiais, como a presença ou ausência de “almeyris” – uma tira de couro nas patas.
2.3. Análises estatísticas
De modo a comparar o esforço parental dos casais estudados durante a primeira e segunda posturas e na ausência e presença de ovos e crias, a época de reprodução foi dividida em seis períodos:
• Período 1: Período que decorre desde o início do estudo até à deposição do primeiro
ovo da primeira postura; corresponde ao primeiro período de pré-incubação.
• Período 2: Período que decorre desde a deposição do primeiro ovo da primeira postura até à ausência dos ovos da primeira postura; corresponde ao primeiro período de incubação.

17
• Período 3: Período que decorre desde o final da ausência de ovos da primeira postura, por eclosão dos mesmos, à independência, remoção ou morte das crias; corresponde ao período de cuidados com as crias da primeira postura; corresponde ao primeiro período de cuidados com as crias.
• Período 4: Período que decorre desde o final da ausência de ovos da primeira postura
(porque não eclodiram, foram retirados pelos tratadores ou partidos acidentalmente pelos progenitores, etc.) ou de crias também da primeira postura, à deposição do primeiro ovo da segunda postura; corresponde ao segundo período de pré-incubação.
• Período 5: Período que decorre desde a deposição do primeiro ovo da segunda postura até à ausência dos ovos da segunda postura (porque eclodiram, foram retirados ou partidos acidentalmente pelos progenitores, etc.); corresponde ao segundo período de incubação.
• Período 6: Período que decorre desde o final da ausência de ovos da segunda postura, por eclosão dos mesmos, à independência, remoção ou morte das crias; corresponde ao segundo período de cuidados com as crias.
Nos casais estudados em que ocorreu mais do que uma postura, decidiu-se comparar as diferentes variáveis consideradas entre os vários períodos, de modo a detectar diferenças entre a primeira e segunda posturas. Decidiu-se testar igualmente os dados dos quatro casais em conjunto e comparar o esforço parental entre os vários casais.
Foram utilizados diferentes testes estatísticos dependendo da natureza dos dados. Para escolher entre um teste paramétrico e um não paramétrico, testou-se a normalidade dos dados utilizando o teste de Shapiro-Wilk, onde todos os conjuntos de dados em que p<0,05 foram considerados provenientes de uma distribuição não normal. Os dados não normais foram
transformados utilizando a função �𝑥 + 0,5 para os dados referentes às frequências e
utilizando a função arcsin�√𝑥� para os dados referentes à proporção (Martin & Bateson, 1993), de modo a testar a sua normalidade após as transformações. Utilizaram-se testes paramétricos somente quando os dados originais ou transformados possuíam distribuição normal e o conjunto dos dados excedia trinta amostras.
Para as comparações entre casais, decidiu-se utilizar o teste paramétrico One-way ANOVA e o teste não-paramétrico Kruskal Wallis ANOVA.

18
Para as comparações dentro de cada casal, ou para o conjunto de todos os casais, de modo a testar a significância das diferenças das variáveis entre os vários períodos, escolheu-se o teste paramétrico One-way repeated measures ANOVA e o teste não paramétrico Friedman ANOVA. Para testar a significância das diferenças das variáveis entre os diferentes sexos, escolheu-se o teste paramétrico One-way ANOVA e o teste não-paramétrico de Mann-Whitney. No caso do macho e da fêmea apresentarem na mesma variável uma distribuição normal, optou-se por testar a influência das variáveis “sexo”, “período” e a interacção “sexo*período”, utilizando o teste Two-Way ANOVA.
Decidiu-se que quando as diferenças das variáveis entre os vários períodos se revelassem significativas, que se testaria a posteriori as diferenças entre combinações de períodos – os períodos 1+2+3 versus os períodos 4+5+6 – de modo a comparar o esforço parental entre a primeira e a segunda posturas.
Dentro de cada postura, decidiu-se comparar as diferenças de comportamento entre os vários períodos (1 vs 2 vs 3; 4 vs 5 vs 6) e entre posturas decidiu-se testar as diferenças entre os dois períodos com ausência de ovos (1 vs 4), as diferenças entre os dois períodos com ovos (2 vs 5) e as diferenças entre os dois períodos com crias (3 vs 6).
Para todos os testes a posteriori decidiu-se utilizar os testes paramétricos One-way repeated measures ANOVA e Paired samples t-test e os testes não-paramétricos Friedman Anova e Wilcoxon matched-pairs.
O registo de dados, a conversão de dados, a sua análise exploratória e a representação gráfica dos resultados foram efectuados no programa MICROSOFT EXCEL 2007. Todos os dados foram testados no programa STATISTICA 9 para WINDOWS. O nível de significância utilizado para todos os testes estatísticos foi de α=0,05.
O registo de dados, a conversão de dados, a sua análise exploratória e a representação gráfica dos resultados foi efectuada no programa MICROSOFT EXCEL 2007. Todos os dados foram testados no programa STATISTICA 9 para WINDOWS. O nível de significância utilizado para todos os testes estatísticos foi de α=0,05.
2.4. Hipóteses estatísticas
Com base nas hipóteses apresentadas na introdução e objectivos sobre as diferenças
entre o esforço parental do macho e da fêmea e as diferenças entre o esforço de ambos os progenitores numa segunda postura após o insucesso da primeira, as hipóteses estatísticas correspondentes são as seguintes:

19
H1: Investimento numa segunda postura após insucesso da primeira.
H01: Ausência de segunda postura após insucesso da primeira. H2: Esforço parental do casal na segunda postura igual ou superior ao da primeira.
H02: Esforço parental do casal na segunda postura inferior ao da primeira. H3: Aumento progressivo do investimento desde a pré-incubação aos cuidados com as crias.
H03: Igual investimento, ou redução progressiva desde a pré-incubação aos cuidados com as crias.
H4: Fêmeas com esforço parental superior ao dos machos a cuidar do ninho e dos ovos de ambas as posturas.
H04: Fêmeas com esforço parental igual ou inferior ao dos machos a cuidar do ninho e dos ovos de ambas as posturas.
H5: Fêmeas com esforço parental superior ao dos machos a cuidar das crias da primeira ou da segunda posturas.
H05: Fêmeas com esforço parental igual ou inferior ao dos machos a cuidar das crias da primeira ou da segunda posturas.
H6: Casais com sucesso com esforço parental superior aos casais com insucesso.
H06: Casais com sucesso com esforço parental igual ou inferior aos casais com insucesso.

20
3. RESULTADOS 3.1. Registos obtidos
Devido a problemas técnicos de gravação e algumas interferências no equipamento de gravação, alguns casais em algumas semanas possuem horas de gravação inferiores (Tabela 2). No total foram gravadas cerca de 487 horas de vídeo.
Tabela 2: Dados relativos à gravação de imagens dos casais
Casal Semanas Estudadas Horas totais
Búteo de Harris (Parabuteo unicinctus) 20 205 Falcão-peregrino (Falco peregrinus) 5 40 Falcão-sacre (Falco cherrug) 15 114 Falcão híbrido de gerifalte-sacre (F. rusticolus X F. cherrug)
16 126
3.2. As posturas dos ovos, o sucesso reprodutor e o comportamento
As datas das posturas dos ovos estão descritas na tabela 3. Nenhum dos quatro casais escolhidos para este estudo conseguiram reproduzir-se com
sucesso, tendo o insucesso ocorrido em diferentes períodos da época de reprodução. Globalmente, tendo em vista que não houve eclosão de ovos, ou que a morte das crias foi muito prematura, dos seis períodos reprodutores previamente estabelecidos sobraram quatro: o 1 e o 2 da primeira postura e o 4 e o 5 da segunda. Por essa razão, as hipóteses H5 e H6 sobre o esforço parental dos casais a cuidar das crias e sobre o esforço parental dos casais com e sem sucesso, não poderão ser testadas.
O casal de falcões-peregrino foi o único a não mostrar nenhum comportamento de cortejamento entre ambos os indivíduos. Durante as cinco semanas em que o casal foi estudado, sempre que o macho se aproximava da fêmea na plataforma, a fêmea apresentava uma reacção agressiva, atacando o macho e afastando-o da plataforma.
Os restantes casais apresentaram comportamentos reprodutores. Em todos estes casais ocorreu a deposição de ovos e a incubação dos ovos, tanto pelo macho como pela fêmea.
O casal de falcões híbridos demonstrou uma fraca capacidade na incubação dos ovos, apesar da fêmea ter depositado seis. Os dois primeiros ovos do casal foram encontrados

21
partidos no ninho dois dias depois de terem sido depositados e os restantes ovos, que foram introduzidos numa incubadora, estavam inférteis. Os ovos tinham uma variação de tamanho visivelmente significativa. Durante a incubação dos ovos o casal mostrou-se muito agitado, principalmente a fêmea que se levantava muito do ninho e rodava em torno dos ovos.
A fêmea do casal de falcões-sacre depositou cinco ovos, os quais, após cerca de um mês de incubação, se verificou estarem todos inférteis. Não foi possível identificar os dias da deposição dos ovos, embora os indivíduos tenham começado a sentar-se no ninho em posição de incubação após a quinta semana de estudo. Pensa-se que tenha sido este o momento da deposição dos ovos, mas também é possível que o casal estivesse simplesmente sentado, a descansar no ninho.
O casal de búteos de Harris foi o único casal que conseguiu obter crias. Dos sete ovos que foram depositados pela fêmea durante as duas posturas, apenas os três ovos provenientes da segunda postura eclodiram. Porém, todas as crias foram mortas e parcialmente ou totalmente consumidas pela progenitora. Os ovos provenientes da primeira postura foram incubados até ao dia 27 de Abril de 2009, data em que foram retirados do ninho.
Tabela 3: Datas das posturas dos ovos
Casal Búteo de Harris P. unicinctus
Falcão-peregrino F. peregrinus
Falcão-sacre F. cherrug
Falcão híbrido F. rusticolus X F.cherrug
1º ovo 16-03-20091* ** ?* 13-03-20091P
2º ovo 19-03-20091* ** ?* 17-03-20091P
3º ovo 21-03-20091* ** ?* 28-03-20091Inc*
4º ovo 24-03-20091* ** ?* 01-04-20091Inc*
5º ovo 24-05-20092E ** ?* 05-04-20091Inc*
6º ovo 26-05-20092E ** ** 08-04-20091Inc*
7º ovo 29-05-20092E ** ** ** Legenda: 1 1ª Postura; 2 2ª Postura; ? Não se pôde verificar as datas de postura; * Ovos inférteis; P Ovos partidos; E Ovos que eclodiram; Inc - Ovos colocados numa incubadora ** Nenhum ovo depositado.
3.3. Esforço parental
Dos quatro casais estudados, apenas o casal de búteos de Harris fez duas posturas e incubou os respectivos ovos, sendo portando o único a apresentar quatro períodos de reprodução, o 1 e o 2 da primeira postura e o 4 e o 5 da segunda. Por este motivo apenas os dados do casal de búteos de Harris foram analisados em separado para comparar o esforço

22
parental do macho e da fêmea nestes mesmos períodos. A hipótese H01 de que não há segunda postura após insucesso da primeira foi, portanto, apenas rejeitada no caso dos búteos.
Devido à maior abundância de dados no casal de búteos de Harris e à ausência da maior parte dos períodos de reprodução nos outros casais, os resultados dos testes de todos os casais em conjunto são muito semelhantes aos dos búteos. Por este motivo, apenas se apresentam os resultados dos testes de todos casais em conjunto nos poucos casos em que estes são diferentes dos resultados do casal de búteos.
Note-se que devido ao número de dias de gravação efectuadas entre o início do estudo e a deposição dos primeiros ovos (período 1) no casal de búteos ter sido inferior aos dias de gravação nos períodos 2, 4 e 5 (porque o tempo de gravação total no período 1 foi dividido entre os quatro casais, enquanto que nos períodos subsequentes o casal de búteos de Harris obteve um maior número de horas e dias de gravação, devido ao insucesso prematuro e consequente redução progressiva do tempo de amostragem para os outros casais), o teste de Wilcoxon na comparação entre os períodos 1 e 2 e entre os períodos 1 e 4 apresentou sempre um nível de significância acima de 0,05 mesmo quando T=0 (em que N=4 e p=0,06789). Estes valores indicam uma amostra demasiado pequena para o período 1, o que torna a sua comparação com os outros períodos inconclusiva, pois a hipótese nula nunca poderá ser rejeitada devido a um erro de tipo II.
3.3.1. Frequência de visitas do macho/fêmea ao ninho
Após a transformação das frequências das visitas do macho e da fêmea búteos de Harris ao ninho estes apresentam uma distribuição normal (Shapiro-Wilk W=0,97599; p=0,21280), de maneira que foi possível efectuar um teste de Two-way ANOVA. A influência dos períodos (1, 2, 4 e 5) mostrou-se significativa (Df=3; F=5,115; p=0,003229), enquanto que a influência do sexo (Df=1; F=0,357; p=0,552689) e a interacção entre o sexo e o período (Df=3; F=1,769; p=0,162758) não (Figura 1a). Não se rejeita H04.
A frequência do macho e da fêmea no ninho não é significativamente diferente nos períodos 1+2 (primeira postura) em relação aos períodos 4+5 (segunda postura) (Wilcoxon matched-pairs: N=15; T=0; p=0,00065). Rejeita-se H02.
As frequências não são significativamente diferentes entre os períodos 1 e 4 que se referem à ausência de ovos (N=4; T=0; p=0,06789; provavelmente erro tipo II, tal como explicado acima), ou entre os períodos 2 e 5 que se referem à incubação (N=11; T=11; p=0,050461), rejeitando-se H02. Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes (N=8; T=14: p=0,0575403). O mesmo acontece na segunda postura (período 4 vs período 5) (N=16;T=47; p=0,277531). Não se rejeita H03.
23
Figura 1: Frequência do macho e da fêmea no ninho (a) nos diferentes períodos e (b) nos períodos 1+2 e 4+5. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas em a) e b) são significativas (p<0,05).
Considerando todos os casais em conjunto, verificou-se que os machos e as fêmeas não apresentam diferenças significativas de comportamento entre os quatro períodos (Friedman ANOVA, macho: χ2(N=4, Df=3)=6,0; p=0,11161; fêmea: χ 2(N=4, Df=3)=3,9; p=0,27247).
3.3.2. Proporção do tempo de permanência do macho/fêmea no ninho
O macho e a fêmea permaneceram no ninho numa proporção significativamente diferente, estando o macho menos tempo no ninho em relação à fêmea (Mann-whitney: Df=1; U=264; p=0,00012), rejeitando-se H04. As diferenças do macho (Friedman ANOVA: χ2(N=4; Df=3)=9,3 p=0,02556) e da fêmea (χ2(N=4; Df=3)=10,8; p=0,01286) relativamente aos diferentes períodos também são significativas (Figura 2a).
No macho, a proporção de tempo passado no ninho no período 1+2 é significativamente superior ao tempo passado no ninho no período 4+5 (Wilcoxon matched-pairs: N=15; T=11; p=0,005386), não se rejeitando H02. A fêmea não apresenta diferenças significativas nestes períodos (N=15; T=32; p=0,11177) (figura 2b), rejeitando-se H02.
Ambos os indivíduos apresentam diferenças não significativas entre os períodos 1 e 4 (macho: N=4 T=0; p=0,06789; fêmea: N=4; T=0; p=0,06789), rejeitando-se H02. Existem diferenças significativas na permanência do macho no ninho no período 2 em relação ao período 5 (N=11; T=10; p=0,040861), sendo esta menor no período 5, não se rejeitando H02. Na fêmea esta diferença não é significativa (N=11; T=21; p=0,286004), rejeitando-se H02.
Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes tanto para o macho como para a fêmea (macho: N=4, T=0, p=0,067; fêmea: N=4, T=0, p=0,067; provavelmente erro tipo II), mas a amostra obtida é demasiada pequena. Na segunda postura (período 4 vs período 5) as frequências apenas são
0,00,51,01,52,02,53,03,54,0
1 2 4 5
Freq
uênc
ia
Períodos
Frequência do macho e da fêmea no ninho
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
1+2 4+5
Freq
uênc
ia
Períodos
Frequência do macho e da fêmea no ninhoa) b)

24
significativamente diferentes na fêmea (macho: N=8, T=11, p=0,32699; fêmea: N=8; T=0; p=0,011719). Não se pode rejeitar H03 para o primeiro caso, mas rejeita-se no segundo.
Figura 2: Proporção do tempo do macho e da fêmea no ninho (a) nos diferentes períodos e (b) nos períodos 1+2 e 4+5. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas em a) são significativas (p<0,05). As diferenças apresentadas em b) apenas são significativas para o macho (p<0,05).
Considerando todos os casais em conjunto, verificou-se que os machos não apresentam diferenças significativas de comportamento entre os quatro períodos (Friedman ANOVA: χ2(N=4; Df=3)=7,5; p=0,5756). O resultado para as fêmeas foi semelhante ao do casal de búteos de Harris.
3.3.3. Frequência de ambos os indivíduos no ninho
A frequência da presença de ambos os indivíduos do casal de búteos de Harris no ninho apresentou diferenças significativas nos diferentes períodos (Friedman ANOVA: χ2(N=4; Df=3)=9,3; p=0,02556) (Figura 3a). Nos períodos 1+2 e 4+5, as vezes em que o casal se junta no ninho é significativamente diferente (Wilcoxon matched-paires: N=15; T=0; p=0,000655), estando mais vezes juntos no período 1+2 (Figura 3b). No período 2 e no período 5 as diferenças também são significativas (N=11; T=6; p=0,016369), estando o casal mais vezes junto no período 2. Nos dois casos não se pode rejeitar H02. As diferenças entre os períodos 1 e 4 não são significativas (N=4; T=0; p=0,06789), rejeitando-se H02 neste caso. Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes (N=4; T=0; p=0,06789; provavelmente erro tipo II). O mesmo acontece na segunda postura (período 4 vs período 5) (N=8; T=16; p=0,779435). Não se rejeita, nos dois casos, H03. Considerando todos os casais em conjunto, o resultado foi semelhante a este.
0,00,10,20,30,40,50,60,70,80,91,0
1 2 4 5
Prop
orçã
o
Períodos
Proporção do macho e da fêmea no ninho
0,00,10,20,30,40,50,60,70,8
1+2 4+5
Prop
orçã
o
Períodos
Proporção do macho e da fêmea no ninhoa) b)

25
Figura 3: Frequência dos ambos os indivíduos no ninho (a) nos diferentes períodos e (b) nos períodos 1+2 e 4+5. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas em a) e b) são significativas (p<0,05).
3.3.4. Proporção do tempo de permanência de ambos os indivíduos no ninho
A proporção de tempo de permanência de ambos os indivíduos do casal de búteos de Harris no ninho é significativamente diferente nos vários períodos (Friedman ANOVA: χ2(N=4; Df=3)=9,3; p=0,02556) (Figura 4a). O período 1+2 e o período 4+5 também apresentam diferenças significativas para esta variável (Wilcoxon matched-pairs: N=15; T=12; p=0,006407), estando o casal menos tempo juntos no período 4+5 (figura 4b), não se podendo rejeitar H02. Quando se compara o período 1 com o período 4 (N=4; T=0; p=0,06789) ou o período 2 com o período 5 (N=11; T=15; p=0,109512), as diferenças não são significativas, pelo que se pode rejeitar H02. Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes (N=4; T=0; p=0,06789; provavelmente erro tipo II). O mesmo acontece na segunda postura (período 4 vs período 5) (N=8; T=10; p=0,262619). Não se rejeita, nos dois casos, H03. Considerando todos os casais em conjunto, o resultado foi semelhante a este.
Figura 4: Proporção do tempo em que ambos os indivíduos permanecem no ninho (a) nos diferentes períodos e (b) nos períodos 1+2 e 4+5. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas em a) e b) são significativas (p<0,05).
0,0
1,0
2,0
3,0
4,0
5,0
6,0
1 2 4 5
Freq
uênc
ia
Períodos
Frequência de ambos os indivíduos no ninho
0,00,51,01,52,02,53,03,54,04,5
1+2 4+5
Freq
uênc
ia
Períodos
Frequência de ambos os indivíduos no ninho
0,000,020,040,060,080,100,120,140,16
1 2 4 5
Prop
orçã
o
Períodos
Proporção de ambos os indivíduos no ninho
0,000,020,040,060,080,100,120,14
1+2 4+5
Prop
orçã
o
Períodos
Proporção de ambos os indivíduos no ninho
a) b)
a) b)

26
3.3.5. Frequência de ocorrências do ninho vazio
A frequência de ocorrências do ninho vazio no casal de búteos de Harris é significativamente diferente entre os vários períodos (Friedman ANOVA: χ2(N=4; Df=3)=9,9; p=0,01944) (Figura 5). As diferenças não são significativas no período 1+2 em comparação com o período 4+5 (Wilcoxon matched-pairs: N=15; T=33; p=0,125154), nem entre os períodos 1 e 4 (N=4; T=1; p=0,144128). Rejeita-se, nos dois casos, H02. Porém, foram encontradas diferenças significativas entre o período 2 e o período 5 (N=11; T=0; p=0,003346), estando o ninho mais vezes vazio no período 5, o que não permite rejeitar H02. Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes (N=4; T=0; p=0,06789; provavelmente erro tipo II). Na segunda postura (período 4 vs período 5), as frequências são significativas, sendo menores durante a incubação (N=8; T=3; p=0,035693). Sendo assim, rejeita-se H03 apenas no segundo caso. Considerando todos os casais em conjunto, o resultado foi semelhante a este.
Figura 5: Frequência de ocorrência do ninho vazio nos diferentes períodos. As barras de erro representam o erro padrão associado à média. As diferenças entre os períodos são significativas (p<0,05).
3.3.6. Proporção do tempo em que o ninho se encontra vazio
A proporção do tempo em que o ninho se encontra vazio no casal de búteos de Harris é significativamente diferente entre os vários períodos (Friedman ANOVA: χ2(N=4, Df=3)=11,1; p=0,01120) (Figura 6a). As diferenças são significativas entre os períodos 1+2 e 4+5 (Wilcoxon matched-pairs: N=15; T=8; p=0,003143), estando o ninho vazio durante mais tempo no período 4+5 (Figura 6b). Entre os períodos 2 e 5 também existem diferenças significativas (N=11; T=0; p=0,003346), estando o ninho vazio durante mais tempo no período 5. Em ambos os casos não se pode rejeitar H02. As diferenças entre os períodos 1 e 4
0,0
1,0
2,0
3,0
4,0
5,0
6,0
1 2 4 5
Freq
uênc
ia
Períodos
Frequência do ninho vazio

27
não são significativas (N=4; T=4; p=0,715001), rejeitando-se H02 neste caso. Relativamente à primeira postura (período 1 vs período 2), as frequências não são significativamente diferentes (N=4; T=0; p=0,06789; provavelmente erro tipo II). O mesmo não acontece na segunda postura, onde o ninho fica menos tempo vazio durante a incubação (período 4 vs período 5) (N=8; T=0; p=0,011719), rejeitando-se H03 apenas no segundo caso. Considerando todos os casais em conjunto, o resultado foi semelhante a este.
Figura 6: Proporção do tempo em que o ninho se encontra vazio (a) nos diferentes períodos e (b) nos períodos 1+2 e 4+5. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas em a) e b) são significativas (p<0,05).
3.3.7. Frequência de incubação do macho e da fêmea
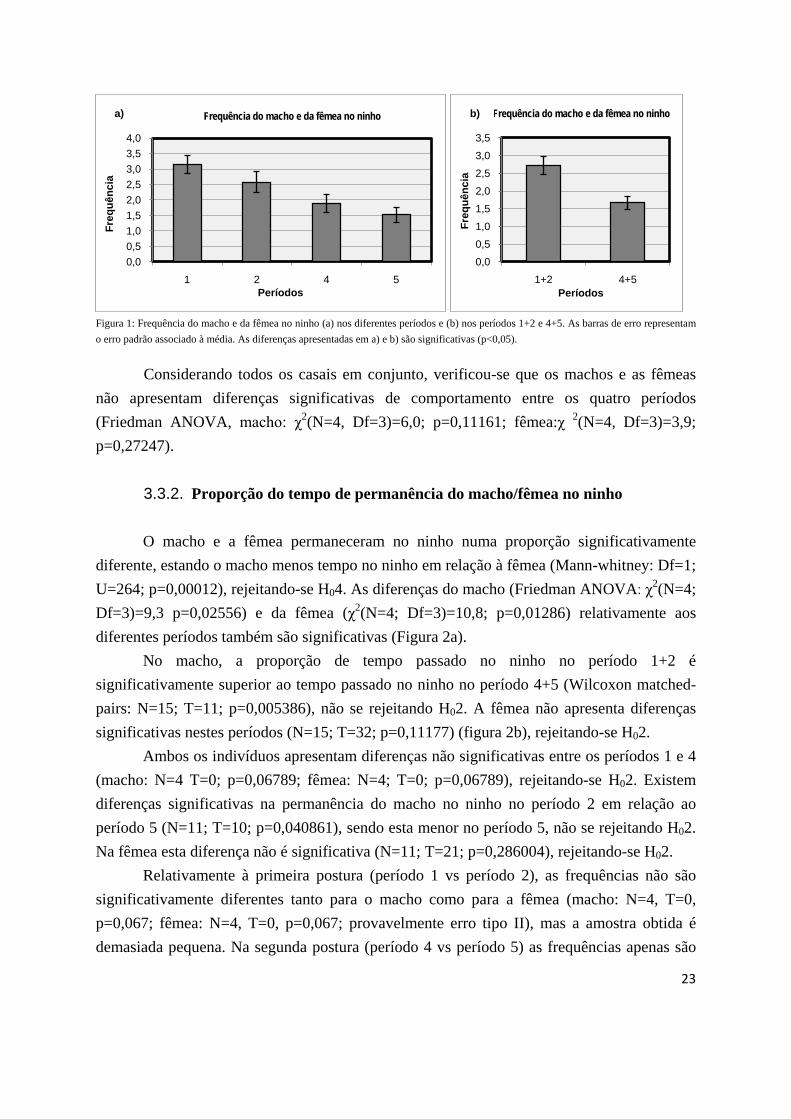
As frequências de incubação do macho e da fêmea búteos de Harris apresentam uma distribuição normal (Shapiro-Wilk: W=0,95205, p=0,6561), de maneira que foi possível efectuar um teste de Two-way ANOVA. A frequência de incubação do casal é significativamente diferente entre os períodos 2 e 5 (Df=1; F=22,749; p=0,000025), sendo maior no período 2. Analisando o macho e a fêmea independentemente, apenas o macho incubou significativamente mais vezes na primeira postura (Wilcoxon matched-pairs: macho: N=11; T=1; p=0,004439; fêmea: N=11; T=11; p=0,050461). Ambos os casos não permitem rejeitar H02. Analisando o efeito do sexo, a fêmea e o macho incubaram com uma frequência significativamente diferente (Df=1; F=16,851; p=0,000193), tendo a fêmea incubado mais vezes do que o macho, rejeitando-se H04. A interacção entre o sexo e o período (Df=1; F=1,329; p=0,25576) não se mostrou significativa (Figura 7). Considerando todos os casais em conjunto, o resultado foi semelhante a este.
0,0
0,2
0,4
0,6
0,8
1,0
1 2 4 5
Prop
orçã
o
Período
Proporção do ninho vazio
0,0
0,1
0,2
0,3
0,4
0,5
0,6
1+2 4+5Pr
opor
ção
Períodos
Proporção do ninho vazioa) b)
28
Figura 7: Frequência de incubação do macho e da fêmea . As barras de erro representam o erro padrão associado à média. As diferenças apresentadas são significativas (p<0,05).
3.3.8. Proporção de tempo de incubação do macho e da fêmea
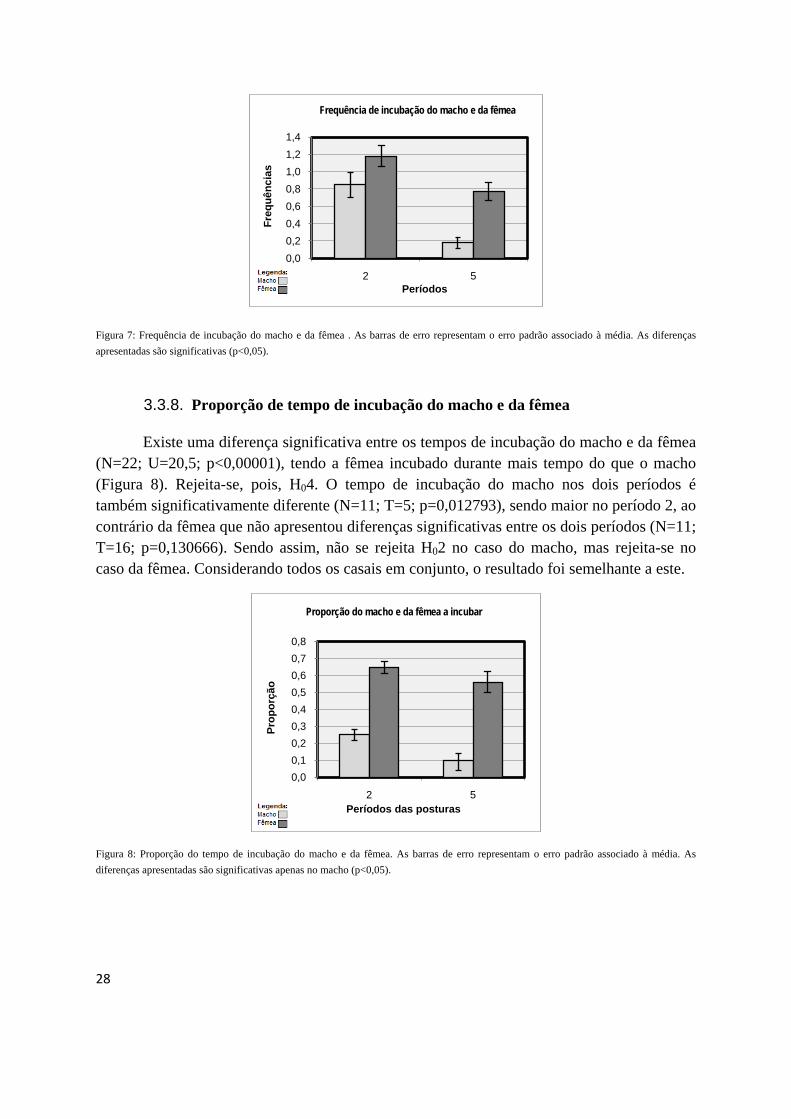
Existe uma diferença significativa entre os tempos de incubação do macho e da fêmea (N=22; U=20,5; p<0,00001), tendo a fêmea incubado durante mais tempo do que o macho (Figura 8). Rejeita-se, pois, H04. O tempo de incubação do macho nos dois períodos é também significativamente diferente (N=11; T=5; p=0,012793), sendo maior no período 2, ao contrário da fêmea que não apresentou diferenças significativas entre os dois períodos (N=11; T=16; p=0,130666). Sendo assim, não se rejeita H02 no caso do macho, mas rejeita-se no caso da fêmea. Considerando todos os casais em conjunto, o resultado foi semelhante a este.
Figura 8: Proporção do tempo de incubação do macho e da fêmea. As barras de erro representam o erro padrão associado à média. As diferenças apresentadas são significativas apenas no macho (p<0,05).
0,00,20,40,60,81,01,21,4
2 5Fr
equê
ncia
s
Períodos
Frequência de incubação do macho e da fêmea
0,00,10,20,30,40,50,60,70,8
2 5
Prop
orçã
o
Períodos das posturas
Proporção do macho e da fêmea a incubar

29
4. DISCUSSÃO 4.1. As posturas dos ovos, o sucesso reprodutor e o comportamento
4.1.1. Casal de búteos de Harris
A primeira postura deste casal consistiu em quatro ovos inférteis e a segunda em três ovos férteis, cujas crias foram mortas e consumidas pela progenitora pouco tempo depois da eclosão.
A infertilidade da primeira postura poder-se-á explicar pelo facto de o macho ser pai da fêmea, hipótese que foi posteriormente descartada, visto que na segunda postura a fêmea depositou três ovos férteis. O fenómeno de inbreeding em aves de rapina mantidas em cativeiro já foi detectado em casais de búteo de Harris, resultando, por vezes, em crias com deficiência (Fox, 1995). A reprodução em cativeiro de casais com elevados níveis de consanguinidade deve, pois, ser evitada ou, então, utilizada como último recurso.
Outra hipótese consiste no stress induzido pela colocação tardia do casal na câmara de reprodução. O casal foi lá colocado em finais de Janeiro, cerca de mês e meio apenas antes de a fêmea colocar o seu primeiro ovo. Os machos nas aves de rapina podem copular sem ejacular esperma quando são muito jovens ou no início da época reprodutora (Fox, 1995). Em futuras reproduções de aves de rapina em cativeiro, será necessário ter em conta os rituais de reprodução de modo a que o macho e a fêmea se sincronizem melhor. A morte e consumo das crias da segunda ninhada por parte da fêmea é, muito provavelmente, um comportamento deslocado, visto que não foi encontrado nenhum registo deste comportamento na natureza. Este comportamento poderá, por isso, dever-se às condições de vida em cativeiro, à inexperiência da fêmea enquanto reprodutora, visto que era a sua primeira reprodução, e/ou ao facto de a fêmea estar habituada a consumir pintos do dia, tendo-os confundido com as suas próprias crias.
Os pintos de frango com um dia são um dos alimentos mais utilizados na falcoaria. Este tem o benefício de ser barato, poder ser facilmente dividido em bocados pequenos para o treino das aves e conter pouco valor calórico, o que permite um maior consumo sem comprometer o peso da ave (Hall, 2003). Apesar disso, não é um alimento que seja recomendado para aves em reprodução devido à sua fraca quantidade de cálcio e à sua baixa componente nutricional (Hall, 2003; Fox, 1995).
Na reprodução em cativeiro os pintos podem constituir outro problema: sendo estes muito parecidos às crias das aves de rapina, as aves habituadas a comê-los podem confundir as crias com alimento (Carlos Crespo, com. pess.). Embora o casal não tivesse sido alimentado com pintos do dia durante o período da incubação, é possível que a fêmea,

30
inexperiente, tenha associado os pintos de galinha aos seus. No ano seguinte ao estudo, a fêmea, após acasalamento com o mesmo macho, depositou três ovos que foram recolhidos numa incubadora e que deram origem a três crias saudáveis.
4.1.2. Casal de falcões-sacre
Todos os ovos que a fêmea colocou mostraram-se inférteis, apesar de este casal já se ter reproduzido com sucesso durante várias épocas reprodutoras anteriores e de se ter reproduzido com sucesso no ano seguinte ao estudo. É provável que o insucesso se tenha devido ao stress induzido por um novo ambiente (novas câmaras reprodutoras), tal como aconteceu com o casal de búteo de Harris. O casal de falcões-sacre foi colocado na instalação em finais de Janeiro, sendo esta uma instalação desconhecida para ambos os indivíduos.
4.1.3. Casal de falcões-peregrinos A fêmea de falcão-peregrino utilizada neste estudo possui claramente um imprinting
sexual ao ser humano, o que poderá ser a principal causa da rejeição do macho de falcão-peregrino como parceiro sexual.
Após a época reprodutora, o casal foi utilizado para fins de exibição ao público onde era manejada diariamente. Notou-se que durante os maneios a fêmea mostrava sinais de cortejamento direccionados aos tratadores, principalmente durante a época reprodutora. O comportamento mais comum consiste na exibição da inclinação da cabeça que é utilizado pelos falcões-peregrinos na proximidade do parceiro sexual (Wrege & Cade, 1977) (Anexo III). O incorrecto imprinting das aves foi uma das principais causas de insucesso reprodutor nas primeiras tentativas de reprodução de aves de rapina em cativeiro (Heidenreich, 1997). Segundo Fox (1995), o imprinting sexual nestas aves é desenvolvido à nascença e dirigida à espécie provedora de alimento, apesar de existirem outras variáveis que influenciam este comportamento. Outra razão possível para a rejeição do macho por parte da fêmea neste casal de falcões-peregrinos é o facto de ambos os indivíduos terem sido criados em conjunto, com mais seis outras crias. Fox (1995) sugere que alguns indivíduos não aceitam parceiros sexuais quando estes são criados em conjunto, como estratégia de redução da probabilidade de inbreeding.

31
4.1.4. Casal de falcões híbridos Existem duas causas plausíveis para a infertilidade dos seis ovos neste casal: a
hibridação e a idade avançada da fêmea. Os falcões híbridos apesar de serem férteis têm, por regra geral, maior dificuldade em
obter sucesso reprodutor devido ao fenómeno de outbreeding (Fox, 1995). Nestas aves as posturas podem sofrer várias anormalidades, como a variação significativa de tamanho e de peso dos ovos (Heidenreich, 1997). Este fenómeno foi detectado no presente estudo, apesar de se considerar que os falcões sacre e de gerifalte pertencerem a uma super-espécie (Fox, 2004), cujo grau de fertilidade nos híbridos é, geralmente, elevado.
O casal em causa foi obtido no ano do estudo por um criador que garante um histórico de sucesso reprodutor que não pôde ser confirmado.
Além da hibridação, a fêmea é uma ave com uma idade relativamente avançada (18 anos), o que poderá ter diminuído o seu sucesso.
Este casal teve igualmente uma postura fora do comum, pois o intervalo entre a deposição do primeiro ovo e do último foi de 26 dias, não havendo nenhuma distinção clara entre diferentes posturas, o que sugere um fenómeno de egg-pulling. É provável que o facto de os primeiros dois ovos deste casal se terem partido poucos dias depois de terem sido depositados, tenha despoletado na fêmea um comportamento de egg-pulling, o que provavelmente aliado à sua idade avançada, terá levado à produção de ovos de fraca qualidade.
As razões pelas quais os primeiros dois ovos deste casal se partiram durante a incubação poderão ser a espessura dos ovos e o comportamento de incubação da fêmea. Segundo Castilla et al. (2009), que estudaram a resistência da casca dos ovos de várias espécies do género Falco, a hibridação não é uma variável que influencie nem a espessura nem a dureza da casca, mas uma idade avançada da fêmea sim.
A fêmea, durante a incubação, demonstrou um comportamento muito mais agitado do que o macho. Enquanto o macho incubava sentado, a fêmea levantava-se muito frequentemente e mudava de posição, semelhante ao comportamento de escavação do ninho mas sem efectivamente empurrar a areia para trás. Não foi possível determinar qual dos indivíduos partiu os ovos, mas suspeita-se que tenha sido a fêmea por causa deste comportamento. Embora não se conheça a origem do comportamento, este pode estar relacionado com alterações hormonais associadas à idade da fêmea, as quais poderão ter influenciado, por sua vez, o comportamento de incubação (Margalida et al., 2006). Contudo, teria sido necessário um estudo completar para validar esta hipótese.

32
4.2. Esforço Parental
4.2.1. Frequência e proporção de tempo do macho e da fêmea no ninho
A frequência e a proporção de tempo do macho e da fêmea no ninho são indicadores do esforço parental do casal, com significados diferentes nos períodos de pré-incubação e incubação.
No período de pré-incubação, tanto a frequência de visitas ao ninho, como o tempo passado no ninho pelo macho e pela fêmea são duas variáveis que se relacionam com o esforço parental, indicando a energia e tempo que ambos despendem a preparar o ninho para a reprodução.
Durante o período de incubação estas duas variáveis relacionam-se também com o esforço parental, indicando, desta vez, a energia e tempo despendidos pelos progenitores a incubar e a proteger os ovos.
Nos quatro casais estudados e, em particular, no casal de búteos de Harris, a frequência de visitas do macho e da fêmea ao ninho diminuiu ao longo do tempo, independentemente de terem existido dois períodos de incubação (Figura 1). Tendo em conta o insucesso da primeira postura, seria de esperar um aumento ou igual frequência de visitas ao ninho na segunda postura, sobretudo no período de incubação, tendo sido essa a previsão apresentada em H2. Os resultados obtidos contrariam, no entanto, essa previsão. A causa desta diminuição pode ser explicada por três hipóteses, não mutuamente exclusivas entre si.
A primeira hipótese relaciona-se com a construção activa do ninho pelo casal. Nos primeiros dias em que o ninho foi fornecido ao casal, este foi visto a levar paus para o ninho e a arranjá-lo com muita frequência. Esta frequência foi diminuindo ao longo do tempo. Durante o segundo período de pré-incubação e de incubação, o ninho utilizado foi o mesmo, o que diminuiu a necessidade de nele depositar muitos mais paus.
A segunda hipótese consiste no esforço parental do casal, em termos de frequência de visitas, ter, de facto, diminuído ao longo dos dois períodos de pré-incubação e de incubação. Dado que o casal investiu muito tempo e energia na preparação do ninho e na incubação dos ovos da primeira postura, é possível que os dois progenitores ou, pelo menos, um não tenha conseguido recuperar fisiologicamente, tendo diminuído o seu esforço parental na segunda postura.
Por fim, a frequência de visitas do macho e da fêmea ao ninho é uma variável que, na verdade, não pode ser interpretada sem o tempo de permanência dos mesmos no ninho, pois a sua diminuição pode dever-se simplesmente a uma presença mais prolongada dos progenitores no ninho.

33
Sendo assim, relativamente à proporção de tempo passado no ninho (Figura 2), a fêmea de búteo de Harris permanece mais tempo no ninho do que o macho, principalmente durante os períodos de incubação. Este fenómeno é comum dentro da espécie, em que o macho apenas incuba 30% do tempo (Wingfield et al., 2000) e está de acordo com as previsões apresentadas em H4 (rejeita-se H04).
Comparando a permanência no ninho entre os diferentes períodos, observa-se que dentro da mesma postura, os progenitores não aumentaram o seu esforço parental da pré-incubação para a incubação, ao contrário do que se previa em H3, à excepção da fêmea na segunda postura. Este resultado deve-se, provavelmente, à falta de dados na primeira postura (erro tipo II).
Entre a primeira e segunda posturas, o macho diminuiu o tempo despendido com a segunda postura (períodos 1+2 vs 4+5), nomeadamente na incubação (períodos 2 vs 5), algo que não aconteceu na fêmea, a qual manteve o mesmo nível de esforço. Estes resultados, juntamente com os dados da frequência do macho e da fêmea no ninho, sugerem uma diminuição do esforço parental do macho em relação ao cuidado com os ovos, o que rejeita – no caso do macho – a hipótese de que a perda da primeira postura aumenta ou, pelo menos, mantém o nível de esforço na segunda (H2).
4.2.2. Frequência e proporção de tempo de ambos os indivíduos no ninho
Os dados relativos à frequência e à proporção de tempo de ambos os indivíduos no ninho, no caso do casal de búteos de Harris, podem indicar a coesão do casal nos períodos de pré-incubação e incubação, sendo mais relevantes durante a incubação (Figuras 3 e 4).
Foi observado que ambos os indivíduos incubam os ovos, embora em proporções diferentes. A troca de incubação entre os dois indivíduos é normalmente feita quando um dos progenitores chega ao ninho e tenta com a cabeça empurrar o ventre do parceiro, de modo a que este último se levante do ninho e deixe o primeiro incubar. Deste modo, seria de esperar uma maior frequência e tempo despendidos por ambos no ninho na presença de ovos em ambas as posturas (Figuras 3a e 4a), tal como tinha sido previsto em H3, algo que não aconteceu provavelmente devido à falta de dados na primeira postura (erro tipo II) e à diminuição, por parte do macho, da frequência e do tempo passado no ninho durante a segunda postura.
Os indivíduos permaneceram mais vezes juntos e durante significativamente mais tempo durante a primeira postura do que na segunda (períodos 1+2 vs 4+5; Figuras 3b e 4b) e estão mais vezes juntos durante a primeira incubação do que na segunda (períodos 2 vs 5; Figura 3a), ao contrário do que seria de esperar, segundo a hipótese H2 do aumento ou igual

34
esforço parental após o insucesso da primeira postura (não se rejeita H02). Este resultado pode indicar uma menor sincronização do casal, assim como uma menor motivação para permanecer no ninho por parte, pelo menos, de um dos progenitores. Conjuntamente com os dados analisados anteriormente, pode-se inferir que estas diferenças nos dois períodos de incubação se devem, provavelmente, ao macho.
4.2.3. Frequência e proporção de tempo em que o ninho se encontra
vazio
A frequência do ninho vazio é uma variável que deve ser considerada juntamente com a proporção de tempo do ninho vazio, pois a primeira indica a tendência para qualquer um dos membros do casal abandonar o ninho, enquanto a segunda indica a tendência para qualquer um dos membros do casal regressar ao ninho quando este se encontra vazio. Estas duas variáveis tornam-se mais importantes na medição do esforço parental na presença de ovos no ninho, altura em que estes precisam de ser incubados.
Note-se que a frequência e a proporção de ausência do casal no ninho não têm o mesmo significado em cativeiro e na natureza, pois em cativeiro o casal encontra-se confinado num espaço fechado, de maneira que nunca poderá abandonar as imediações do ninho.
Segundo os resultados obtidos, a frequência e a proporção de tempo em que os indivíduos se ausentam do ninho são significativos em relação aos diferentes períodos, o que coincide com os resultados esperados, dado que existem períodos com presença de ovos e períodos com ausência de ovos. Apenas na segunda postura existiu, claramente, da pré-incubação para a incubação, uma redução da frequência e do tempo em que o ninho é deixado vazio (períodos 4 vs 5; Figuras 5a e 6a), tal como previsto em H3 (rejeita-se H03). Na primeira postura tal não aconteceu provavelmente devido à falta de amostras (erro tipo II).
O ninho encontra-se vazio durante mais tempo na segunda postura (períodos 1+2 vs 4+5; Figura 6b) e, considerando apenas os períodos em que existem ovos em ambas as posturas, o ninho encontra-se mais vezes vazio e durante mais tempo no período 5 em relação ao período 2. Ambos os resultados indicam um menor esforço parental do casal em relação aos ovos da segunda postura, o que não suporta, mais uma vez, a hipótese H2 do aumento ou igual esforço parental após a perda de uma primeira postura (não se rejeita H02).
4.2.4. Frequência e proporção de tempo de incubação
A frequência e a proporção de tempo de incubação do macho e da fêmea (Figuras 7 e 8) são duas variáveis que se podem considerar muito semelhantes à frequência e à proporção

35
de tempo do macho e da fêmea no ninho durante os períodos de incubação (períodos 2 e 5; Figuras 1 e 2). Existe, contudo, uma diferença: as Figuras 7 e 8 dizem respeito à incubação propriamente dita e as Figuras 1 e 2 à presença dos progenitores no ninho, podendo incluir, neste último caso, comportamentos de incubação e de não incubação.
Observou-se que é a fêmea que incuba os ovos com mais frequência a maior parte do tempo, o que condiz com o comportamento esperado (H4), pois a fêmea é maior e dominante no casal, tal como acontece na natureza (rejeita-se H04). Contudo, independentemente desta diferença, que é natural, observa-se no macho uma redução significativa do tempo passado a incubar da primeira para a segunda posturas. Os resultados assim obtidos não suportam, mais uma vez no caso do macho, a hipótese H2 do aumento ou igual esforço parental durante a segunda incubação após o insucesso da primeira postura (não se rejeita H02).
No caso da fêmea, observou-se igualmente uma redução da frequência de incubação da primeira para a segunda postura, mas, em contrapartida, um igual esforço parental no tempo total de incubação, o que suporta a H2 (rejeita-se H02).
4.2.5. Geral
Os dados obtidos no presente estudo, apesar de serem escassos e de não amostrarem sucesso reprodutor em nenhum dos casais, permitem responder com algum detalhe a quatro das hipóteses apresentadas.
O investimento numa segunda postura após insucesso da primeira (H1) aconteceu em apenas um dos quatro casais amostrados. Este resultado não rejeita H1 em absoluto, visto que, na natureza, só acontecem segundas posturas após o insucesso de tentativas prévias, mas sugere que esse investimento não é condição obrigatória, dependendo provavelmente de múltiplos factores, como condições ambientais, coesão entre os membros do casal, etc. Dado que as aves estudadas se estavam a reproduzir em cativeiro e nalguns casos em instalações novas e/ou com parceiros desconhecidos, parceiros da mesma criação ou parceiros consanguíneos, as condições para o investimento na segunda postura podem ter ficado comprometidas para alguns destes casais.
Relativamente ao casal de búteos de Harris, que foi o único a fazer duas posturas, os resultados sugerem um esforço parental dos machos inferior ao das fêmeas a cuidar do ninho e dos ovos de ambas as posturas, em termos de tempo passado no ninho (Figura 2) e de frequência e tempo passados a incubar os ovos (Figuras 7 e 8), tal como previsto em H4, visto que é este o comportamento típico na natureza, na qual os machos se dedicam sobretudo à caça (Heidenreich, 1997).

36
Devido à falta de dados na primeira postura, nunca se observou um maior investimento parental entre o período da pré-incubação e o período de incubação na primeira postura, ao contrário do que se previa em H3. Contudo, pela análise das Figuras 2 a 6, a tendência para um maior esforço parental durante a incubação da primeira postura (período 1 vs 2) é bem visível, o que sugere um erro estatístico de tipo II. Relativamente à segunda postura, o aumento do esforço parental durante a incubação só se verifica na fêmea (na proporção de tempo que esta passa no ninho; Figura 2), e na frequência e proporção de tempo em que o ninho é deixado vazio pelo casal (Figuras 5 e 6). O comportamento do macho é inverso, no qual há uma tendência para a diminuição do seu esforço parental durante a segunda incubação (Figura 2). Sendo assim, rejeita-se H03 somente no caso da fêmea.
Relativamente a H2, segundo a qual se previa um igual ou maior esforço parental de ambos os progenitores da primeira para a segunda postura, os resultados sugerem um igual esforço por parte da fêmea nas duas posturas, relativamente ao tempo passado no ninho na pré-incubação e incubação (Figura 2), assim como na incubação apenas (Figuras 2a e 8). Em termos de casal, tanto a frequência como o tempo passado no ninho são menores na segunda postura (Figuras 3 e 4), e o tempo de ninho vazio é também maior na segunda postura (Figura 6), assim como a frequência e o tempo na segunda incubação (Figuras 5 e 6). Estes resultados, que não suportam H2, podem dever-se, tal como nos resultados observados para H, ao comportamento do macho (Figuras 2, 7 e 8). Rejeita-se, então, H02 somente no caso da fêmea.

37
5. Considerações finais
Do presente estudo sobre o comportamento reprodutor de quatro casais de aves de rapina diurnas mantidas em cativeiro na Coudelaria Alter Real e em Vaiamonte, realizado entre Fevereiro e Junho de 2009, no qual o objectivo principal foi medir e comparar o esforço reprodutor dos progenitores numa segunda postura após insucesso da primeira, pode-se inferir o seguinte: (H1) o investimento numa segunda postura não acontece sempre que há insucesso da primeira, devendo depender de múltiplos factores ambientais que devem ser estudados; (H4) as diferenças no comportamento reprodutor do macho e da fêmea encontradas na natureza, com a fêmea investindo mais tempo e energia a cuidar do ninho e dos ovos, mantêm-se em cativeiro; há, por parte da fêmea, (H3) um aumento do esforço parental da pré-incubação para a incubação e (H2) um investimento global semelhante entre a primeira e a segunda posturas, ao contrário do que se observa no macho, no qual há uma redução do investimento em ambos os casos.
Os resultados obtidos com H2 sugerem que a prática da remoção artificial dos ovos da primeira postura para induzir uma segunda, de modo a aumentar a produtividade das aves em cativeiro, nem sempre resulta num maior investimento por parte dos progenitores e consequente sucesso.
Estas afirmações devem, contudo, ser feitas com muitas reservas, pois os dados correspondem apenas a um casal e a uma única época reprodutora, podendo, para além disso, ter sido influenciados por outras variáveis que não foi possível controlar.
Deve-se igualmente salientar que embora sendo o macho a diminuir o seu esforço parental através de uma menor presença/incubação dos ovos durante o período 5, foi a fêmea que matou as suas crias quando estas nasceram. Isto indica que um maior esforço parental numa etapa da reprodução por parte da fêmea não indica maior esforço na etapa seguinte.
5.1. Perspectivas – Dados do Estudo em Relação ao Sucesso Reprodutor
O presente estudo contemplava, inicialmente, tal como exposto nos objectivos, uma análise ao comportamento parental dos progenitores relativamente às crias, assim como análises comparativas entre os casais e entre os vários períodos, desde a pré-incubação aos cuidados com as crias, objectivos que não puderam ser concretizados devido ao insucesso reprodutor de todos os casais estudados e ao fim prematuro do esforço reprodutor de três dos quatro casais. Curiosamente, desses quatro casais estudados, todos possuíram um motivo aparentemente diferente como causa desse insucesso, o que demonstra a complexidade da reprodução de aves de rapina diurnas em cativeiro.

38
Através dos dados obtidos neste trabalho, sugere-se tomar algumas medidas antes e durante a reprodução das aves de rapina diurnas para poder obter maior sucesso reprodutor devendo-se evitar:
1) A colocação do casal reprodutor nas câmaras de reprodução imediatamente antes da época reprodutora – Os ovos férteis do casal de búteo de Harris na segunda postura e os ovos férteis do casal de falcões-sacre no ano seguinte ao do estudo, levam a acreditar que as posturas inférteis se devem principalmente ao stress induzido pela colocação do casal num novo ambiente.
2) A criação de uma primeira postura por algum progenitor inexperiente – No caso de existir algum progenitor que esteja no primeiro ano de reprodução, caso se queira uma maior garantia na produção de crias, sugere-se a recolha dos ovos da primeira postura para uma incubadora, deixando aos progenitores a criação da segunda postura. Deste modo garante-se que algumas das crias não sejam mortas pelos progenitores e ajuda-se a que os progenitores ganhem experiência na criação da segunda postura, pois o sucesso na incubação dos ovos não significa sucesso na criação da prole.
3) A alimentação dos casais reprodutores com pintos do dia – Sugere-se que seja
reduzido ao máximo possível o consumo de pintos do dia pelos animais reprodutores, de modo a que estes não os confundam com as próprias crias, principalmente nos sujeitos inexperientes.
4) A formação de casais em que pelo menos um dos indivíduos esteja com imprinting sexual ao ser humano – Embora convenha que as aves estejam habituadas ao ser humano, as aves que manifestem um comportamento de cortejamento dirigido ao homem, provavelmente não irão aceitar um conspecífico como parceiro sexual. Neste caso poderá optar-se por inseminação artificial.
5) A formação de casais com indivíduos criados juntos numa incubadora – Os indivíduos que são criados juntos é provável que se identifiquem como sendo irmãos e podem rejeitar-se um ao outro enquanto parceiros sexuais como prevenção natural para o fenómeno de inbreeding.

39
5.2. Aspectos técnicos
A metodologia deste estudo não permitiu uma análise estatística robusta aos dados em questão, pois apenas foram estudados quatro casais reprodutores de diferentes espécies ou híbridos por apenas um ano, o que pode não reflectir um padrão de comportamento reprodutor para as espécies em questão em cativeiro. Em Portugal não existem muitos centros de reprodução de aves de rapina que permitam a observação de muitos casais em reprodução num único ano, estando por isso este estudo restringido aos casais reprodutores do centro.
Previamente ao estudo, não se conheciam em detalhe os sujeitos, nem estes estavam muito familiarizados entre si, gastando-se por isso tempo com alguns casais sem nenhum ou com pouco comportamento reprodutor.
A qualidade das gravações neste estudo, tanto em relação à resolução da imagem como à posição das câmaras, não permitiu fazer um estudo mais elaborado acerca do comportamento destas aves.

40
6. REFERÊNCIAS BIBLIOGRÁFICAS • ALMAÇA, C. 1997. Falconers: the first Portuguese naturalists. Archives of Natural Histories, 24, 175-187.
• AMADON, D. 1975. Why are female birds of prey larger than males? Journal of Raptor Research, 9, 1–11.
• AMADON, D. & BULL, J. 1988. Hawks and owls of the world: a distributional and taxonomic list. Proceedings of the
Western Foundation of Vertebrate Zoology, 3(4), 295-357 in HEIDENREICH, M. 1997. Birds of Prey Medicine and Management. Blackwell Science. 284 pp.
• BEEBE, F. L. 1992. The Compleat Falconer. Hancock House Publishers. 334 pp.
• BROWN, L.H. & AMADON, D. 1968. Eagles, Hawks, and Falcons of the World. McGraw-Hill Book Co.. 945 pp. in RATCLIFFE, D. 2000. The Peregrine Falcon. T & AD Poyser. 454 pp.
• BOTELHO, E. S.; GENNARO, A. L.; ARROWOD, P. C. 1993. Parental care, nestling behaviors and nestling interactions in a Mississippi kite (Ictinia mississippiensis) nest. Journal of Raptor Research, 27(1), 16-20.
• CADE, T. M.; WEAVER, J. D.; PLATT, J. B. & BURNHAM W. A. 1977. The Propagation of Large Falcons in Captivity. Raptor Research, 11(1/2), 28-48
• CASTILLA, A. M.; HERREL, A.; van DONGEN, S.; FURIO, N.; NEGRO, J. J. 2009. Determinants of Eggshell Strength in Endangered Raptors. Journal of Experimental Zoology, 311A, 303-311.
• COLLOPY, M. W. 1984. Parental care and feeding ecology of golden eagle nestlings. The Auk, 101, 753-760.
• CRESPO, C. 1999. A Arte da Falcoaria. Edições Inapa, Lisboa. 140 pp.
• DAWSON, J. W. & MANNAN R. W. 1991. Dominance hierarchies and helper contributions in Harris’ Hawks. The Auk, 108, 649-660.
• DEMENTIEV G.P. 1951. The Peregrine Falcon in the URSS. In: DEMENTIEV G.P. & GALDKOV N.A. (eds.). The
birds of Soviet Union, vol I, pp. 80-100. Soviet Science, Moscow (Em Russo) in RATCLIFFE, D. 2000. The Peregrine Falcon. T & AD Poyser. 454 pp.
• ELLIS, D. H.; ELLIS, M. H.; TSENGEG, P. 1997. Remarkable saker falcon (Falco cherrug) breeding records for Mongolia. Journal of Raptor Research, 31(3), 234-240.
• FERGUSON-LEES, J & CHRISTIE, D. A. 2001. Raptors of the world. Houghton Mifflin Company. 992 pp.
• FOX, N. 1995. Understanding the Bird of Prey. Hancock House Publishers. 368 pp.
• FOX, N. (Ed) 2004. The Use of Exotic and Hybrid Raptors in Falconry – Information arising from the International Committee on Hybrids. Revisto. 59 pp.

41
• FYFE, R. 1978. Breeding behavior of captive and wild praire and peregrine falcons. in NELSON, R. W. (Ed) 1972. Special Conference on Captivity Breeding of Raptors – A Report: Part C. Behavioral Considerations to Egg-laying. Raptor Research, 6, C37-C68.
• GLASIER, P. 1998. Falconry & Hawking. Batsford. 2ªed. 352 pp.
• HALL, T. 2003. Falconry Basics – A Handbook for Beginners. Swan Hill Press. 250 pp.
• HARRELL B. E. Special Conference on Captivity Breeding of Raptors – A Report: Part A. Introduction. Raptor Research, 6(Supp.), A1-A6.
• HARRISON, C. & CASTELL, P. 2002. Collins Field Guide – Bird nests, eggs and nestlings – Britain and Europe. Harper Collins Publishers. Revisto. 474 pp.
• HEIDENREICH, M. 1997. Birds of Prey Medicine and Management. Blackwell Science. 284 pp.
• HERBERT, R. A. & HERBERT, K. G. S. 1965. Behavior of peregrine falcons in the New York City Region. The Auk, 82, 62-64.
• KIMBALL, R. T.; PARKER, P. G.; BEDNARZ, J. C. 2003. Ocurrence and evolution of cooperative breeding among
the diurnal raptors (Accipitridae and Falconidae). The Auk, 120(3), 717-729.
• KORPIMÄKI, E. 1988. Factors promoting polygyny in European birds of prey – a hypothesis. Oecologia, 77, 278-285.
• LPN – LIGA PARA A PROTECÇÃO DA NATUREZA. http://www.lpn.pt/, acedido a 27 de Setembro de 2010.
• MARGALIDA, A; AROOYO, B. E.; BORTOLOTTI G. R.; BERTRAN, J. 2006. Prolongued Incubation in Raptors: Adaptative or Nonadaptative Behavior? Journal of Raptor Research, 40(2), 159-163.
• MARTIN, P; BATESON, P, 1993. Measuring Behaviour: An Introductory Guide. Cambridge University Press, 238 pp.
• MØLLER, A. P. & BIRKHEAD, T. R. 1992. Frequent copulations and mate guarding as alternative paternity guards
in birds: a comparative study. Behaviour, 118, 170-186.
• MOUGEOT, F. 2004. Breeding density, cuckoldry risk and copulation behavior during the fertile period in raptors: a comparative analysis. Animal Behaviour, 67, 1067-1076
• NEGRO, J. J. & GRANDE, J. N. 2001. Territorial signalling: a new hypothesis to explain frequent copulation in raptorial birds. Animal Behaviour, 62, 803-809.
• NELSON, R. W. (Ed) 1972. Special Conference on Captivity Breeding of Raptors – A Report: Part C. Behavioral Considerations to Egg-laying. Raptor Research, 6, C37-C68.
• NEWTON, I. 1978. Feeding and development of Sparrowhawk (Accipiter nisus) nestling. Journal of Zoology, 184, 465-487 in BOTELHO, E. S.; GENNARO, A. L.; ARROWOD, P. C. 1993. Parental care, nestling behaviors and nestling interactions in a Mississippi kite (Ictinia mississippiensis) nest. Journal of Raptor Research, 27(1), 16-20.

42
• OLENDORF, R. 1972. The need for a change in direction – Toward the same goal. in NELSON, R. W. (Ed) 1972. Special Conference on Captivity Breeding of Raptors – A Report: Part C. Behavioral Considerations to Egg-laying. Raptor Research, 6, C37-C68.
• OLIPHANT, L. W. 1991. Hybridization between a Peregrine Falcon and a Praire Falcon in the wild. Journal of Raptor Research, 25(2), 36-39.
• OLSEN, P. D. & OLSEN, J. 1987. Sexual Size Dimorphism in Raptors: Intrasexual Competition in the Larger Sex for a Scarce Breeding Resource, the Smaller Sex. Emu, 87, 59-62.
• ONTIVEROS, D.; CARO, J.; PLEGUEZUELOS, J. M. 2008. Possible functions of alternative nests in raptors: the case of Bonelli’s Eagle. Journal of Ornithology, 149, 253-259.
• PARK, F.; BVSc; MACVS 2003. Behavior and Behavioral Problems of Australian Raptors in Captivity. Seminars in Avian and Exotic Pet Medicine, 12(4), 232-241.
• PENADÉS, G. C. 2007. Female lesser kestrel (falco naumanni) also accepts extra-pair copulation. Journal of Ornithology, 148, 563-564.
• PERRY-JONES, J. 2005. Falconry: Care, Captive Breeding and Conservation. David & Charles. 216 pp.
• RATCLIFFE, D. 2000. The Peregrine Falcon. T & AD Poyser. 454 pp.
• SAINT-JALME, M. 2002. Endangered avian species captive propagation: an overview of functions and techniques. Avian and Poultry Biology Reviews, 13(3), 187-202.
• SIBLEY, C.G. & AHLQUIST, J.E. 1990. Phylogeny and Classification of Birds. Yale University Press, New Haven,
London. 1080 pp.
• SUTHERLAND, W. J. 1998. The importance of behavioural studies in conservation biology. Animal Behaviour, 56, 801-809.
• THE PEREGRINE FUND. http://www.peregrinefund.org/, acedido a 27 de Setembro de 2010.
• TODD, F. S. 1972. Captive Breeding of Harpy Eagles. Raptor Research, 6(4), 137-143.
• WIĄCEK, J. 2006. Aggressive behavior in Montagu’s harrier Circus pygarcus during the courtship period. Biologia, Bratislava, 61(5), 593-595.
• WINGFIELD, J.C.; JACOBS, J.D.; TRAMONTIN, A.D.; PERFITO, N.; MEDDLE, S.; MANEY, D.L.; SOMA, K. 2000. Toward an ecological basis of hormone behavior interactions in reproduction of birds, in: WALLEN, K. & SCHNEIDER, J.E. (eds.), Reproduction in Context. Social and Environmental Influences on Reproduction, A Bradford Book, MIT Press, Cambridge MA, London, pp. 85-128.
• WREGE, P. H. & CADE, T. J. 1977. Courtship Behavior of Large Falcons in Captivity. Raptor Research, 11(1/2), 1-46.

43
7. ANEXOS

44
Anexo I: Folha de Registo da Entrada e Saída do Ninho/ Folha de registo de estados comportamentais
Código da Gravação:
Data:____/____/ 2009
Casal:
Observações:
Hora do Início da Gravação: Hora do Final da Gravação:
Hora de Vídeo/ Hora Real ♂ ♀ ? Observações: Início Fim

45
Anexo II: Folha de Registo de eventos comportamentais
Código da Gravação:
Data:____/____/ 2009
Casal:
Observações:
Hora do Início da Gravação: Hora do Final da Gravação: Legenda:
Tempo Comportamentos ♂ ♀ ? Observações