chuva de sementes como indicador de restauração ... · chuva de sementes como indicador de...
TRANSCRIPT

Chuva de sementes como indicador de restauração ecológica em matas
ripárias do Distrito Federal
Barbara Silva Pachêco
Orientador: José Francisco Gonçalves Junior
Co-orientadoras: Fabiana de Gois Aquino
Lidiamar Barbosa de Albuquerque
Montes Claros, Minas Gerais
2014

Bárbara Silva Pachêco
Chuva de sementes como indicador de restauração ecológica em matas
ripárias do Distrito Federal
Dissertação apresentada ao Programa de Pós-
Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros como requisito necessário
para a conclusão do curso de mestrado em
Ciências Biológicas.
Montes Claros, Minas Gerais
2014

Bárbara Silva Pachêco
Chuva de sementes como indicador de restauração ecológica em matas
ripárias do Distrito Federal
Dissertação apresentada ao Programa de Pós-
Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros como requisito necessário
para a conclusão do curso de mestrado em
Ciências Biológicas.
Aprovada em: 27 de março de 2014
Montes Claros, Minas Gerais
2014

Agradecimentos
A vida nos reserva grandes surpresas, grandes pessoas e grandes momentos!
Agradeço àquilo que ela me designou neste período tão intenso que foi o Mestrado. Um
momento de muitas descobertas, estudos, dúvidas, afirmações, crescimento profissional
e principalmente pessoal. Ao final desta etapa, não saio somente com um diploma,
sinceramente, de todos os ganhos, esse é o que menos pesa para mim. Levo, desse
processo, pessoas, histórias, bons momentos, sorrisos, gargalhadas, viagens: levo você!
Tenho certeza de que esses foram meus maiores diplomas, os maiores e melhores
títulos!
Essa etapa não seria nada sem as dezenas de pessoas que agora estão gravadas em mim.
Agradeço veementemente a elas e mesmo assim sei que tudo o que estas palavras dizem
é insuficiente.
Obrigada aos que tiveram alguma participação neste processo chamado Mestrado! Você
sorriu para mim? Você, em silêncio, ficou ali ao meu lado? Você me ensinou das
pequenas às grandes coisas? Você caminhou comigo? Você me ajudou a triar milhares
de minúsculas sementes? Você foi a campo para coletar durante horas aquela grande
quantidade de material? Você me levou para ver um filme na tentativa de me distrair um
“tiquim”? Você entendeu e escutou minhas lamúrias raivosas? Você entendeu minhas
ausências e distâncias? Você me agraciou com sua experiência? O que tenho a dizer a
você é um sincero e carinhoso: muito obrigada!
Digamos que desde o início sou uma “área em processo de restauração”. Obrigada você,
que foi poleiro e serviu de facilitador. Obrigada você, que serviu como uma ilha de
nucleação permitindo que tudo tivesse um início. Obrigada você, que com um trator
passou para revolver meu solo e deixar que o banco de boas sementes voltasse a existir.
Obrigada você, que afastou de mim os fatores de degradação. Obrigada você, que
cuidou para que pensamentos invasores não tomassem de conta e abafassem meus
regenerantes. Obrigada vocês, que fizeram de mim uma “área nova”, com processos
restaurados e disposta a continuar mantendo, bem, vários serviços ecológicos, sociais e
pessoais.
E quem seria esse “você”?
Sei bem que vocês sabem! Rs!
Juntamente com o título, levo pouco de mim e muito de vocês. Serei eternamente grata.
Agradeço especial e imensamente aos meus pais e meus irmãos pelo apoio e orgulho
incondicional.
Agradeço à Embrapa Cerrados pela estrutura e corpo funcional, especialmente pela
ajuda dos funcionários Natália, Valdeci, Paixão e Nelsinho. Aprendi com vocês
ensinamentos que levarei pelo resto da vida.
Agradeço ao funcionário Juaci Malaquias pelo apoio nas análises estatísticas.
Agradeço aos proprietários das áreas experimentais pelo apoio e pela viabilização do
experimento: Sr. Emanuel e Sr. Cupertino.
Agradeço aos pesquisadores da Embrapa: Dr. Bruno Machado Telles Walter, Dr.
Luciano de Bem Bianchetti, Dra. Antonieta Nassif Salomão e Dr. José Francisco
Montenegro Valls e a doutoranda Priscila Alves dos Reis pela grande ajuda na
identificação, preliminar, dos diásporos.
Agradeço à equipe do projeto “Aquaripária” por tudo, principalmente à Aline Cristina e
Simone Rodrigues que foram minhas companheiras e meu braço direito no processo.

Obrigada também aos estagiários do projeto que tanto me apoiram: sem vocês, eu não
teria conseguido!
Agradeço à Unimontes pela oportunidade, pelos amigos maravilhosos e inesquecíveis
que me apresentou, pelo “nuuuu”, pelo “mooooço”, pelo “treeem” e pelo “uaaai” que
aprendi durante esta inesquecível vivência! Rsrs!
Obrigada ao meu trio de orientadores Dr. José Francisco Junior, Dra. Lidiamar
Albuquerque e Dra. Fabiana Aquino que abriram portas, me direcionaram e orientaram
com tamanha responsabilidade, carinho e dedicação.
Meu eterno e muito obrigada!
“Mar calmo nunca fez bom marinheiro”

Resumo
PACHÊCO, B.S. Chuva de sementes como indicador de restauração ecológica em
matas ripárias no Distrito federal. (Dissertação Mestrado em Ciências Biológicas -
UNIMONTES), 2014.
O conhecimento da variação da chuva de sementes contribui para a compreensão dos
processos reprodutivos e da dinâmica dos ecossistemas podendo subsidiar ações de
restauração de áreas degradadas. Projetos que visam restaurar ambientes degradados
necessitam de ferramentas para avaliar o progresso da restauração. Os indicadores
ecológicos compreendem essas ferramentas que permite interpretar a situação da área
para intervir. O objetivo deste trabalho foi avaliar a chuva de sementes como indicador
de restauração ecológica em áreas de zonas ripárias no Distrito Federal. Foram
instalados 132 coletores de sementes em duas áreas de matas ripárias que diferem
quanto ao histórico de uso da terra e paisagem de entorno (Rio Jardim e Córrego Capão
Comprido). Nas duas área foram instalados tratamentos, com três repetições (parcelas),
para avaliação do aporte de chuva de sementes tanto na mata remanescentes (MR) como
na área em processo de restauração (APR). Na APR foram instalados tratamentos com
as técnicas de restauração ecológica: nucleação de Anderson, linha de recobrimento e
diversidade, poleiro, nucleação Anderson + poleiro, controle. Em cada área foram
instalados diferentes tratamentos (de acordo com o tamanho da área). Em cada parcela
foi distribuído quatro coletores de chuva de sementes (50 x 50 cm, a 15 cm do solo,
malha de nylon de 1 mm²). Como cada tratamento possuía três repetições, foram
avaliados, portanto 12 coletores/tratamento/mês. O material depositado nos coletores foi
recolhido mensalmente por um ano (março de 2012 a fevereiro de 2013), em sacos de
papel e triado no Laboratório de Biologia Vegetal da Embrapa Cerrados e Laboratório
de Biologia da conservação da UNIMONTES. As sementes foram separadas, com
auxilio de estereomicroscópio, sendo quantificadas e identificadas quando possível em
espécie e modo de dispersão. Na área do Jardim foi amostrado no total 100.040
diásporos (8337sem/m2), sendo 4053 (1351m
2) MR e 96987(10777sem/m
2) na APR. No
Coité (Capão Comprido) foi 74.316 diásporos (3539sem/m2), sendo 5718 (1906sem/m
2)
MR e 68598 (3811sem/m2). Sendo as maiores quantidades representadas pela espécie
invasora Urochloa decumbens (Stapf) R.D. Nas duas áreas a síndrome de dispersão
predominante na área em processo de restauração (APR) foi a anemocórica e na mata
remanescente (MR) a “não anemocória”. Em ambas as áreas as sementes de tamanhos
pequenos (<2mm) foram mais abundantes. No aporte de chuva de sementes houve
variação entre os meses, sendo o pico em abril para o Rio Jardim e fevereiro para o
Coité. Em ambas as áreas registrou-se diferenças entre o aporte de chuva de sementes
na APR e na MR. No Jardim os tratamentos que apresentaram os maiores aportes de
chuva de sementes quanto à abundância e riqueza foram T3 (controle) e T4 (mata
remanescente), respectivamente (p<0,001,GL:8). Na área Coité foram, respectivamente,
T5 (linha de diversidade e recobrimento) e T7 (mata remanescente) (p<0,001, GL:14).
Com os resultados obtidos para ambas as áreas é possível observar que a chuva de
sementes reflete as situações distintas de cada área respondendo a pequenas variações,
refletindo o estado de conservação de cada ambiente e permitindo ter conclusões
também no âmbito da paisagem. Devido a essa acurácia, sensibilidade e
mensurabilidade a chuva de sementes pode ser utilizada como indicador de restauração
ecológica em áreas ripárias, permitindo extrair do estudo informações sobre a

capacidade de regeneração do ecossistema. Podendo ser utilizado como indicador do
potencial de regeneração ou como indicador de automanutenção para subsidiar as
atividades de manejo e/ou a escolha de melhores metodologias para a restauração.
Abstract
Knowledge of the variation of seed rain contributes to the understanding of reproductive
processes and dynamics of ecosystems can support programs to restore degraded areas .
Projects to restore degraded environments require tools to assess the progress of the
restoration. Ecological indicators that comprise these tools allows us to interpret the
situation of the area to intervene . The objective of this study was to evaluate the seed
rain as an indicator of ecological restoration in areas of riparian zones in the Federal
District . 132 seed collectors were installed in two areas of riparian forests that differ in
history of land use and landscape around ( Rio Jardim and Capão Comprido) . In each
area plots to evaluate the contribution of seed rain in different ecological restoration
techniques were demarcated. Each technique or combination of more than one was
considered as treatment (nucleação de Anderson , linha de recobrimento e diversidade,
poleiros, nucleação de Anderson + poleiro, and mata remanescentes). In each area
different treatments were installed (according to the size of the area) , with three
replicates (plots) . Diagonal of each plot of restoration treatments was distributed four
collectors of seed rain (50 x 50 cm , 15 cm of soil , nylon mesh of 1 mm ²) . As each
treatment had three repetitions, therefore 12 collectors / treatment / month. The material
deposited in the traps was collected monthly for one year (March 2012 to February
2013 ) , in paper bags and screened at the Laboratory of Plant Biology (EMBRAPA
Cerrados) and Laboratory of Conservation Biology (UNIMONTES). The seeds were
separated with the aid of stereomicroscope and identified and quantified when possible
in cash and dispersal mode . In Rio Jardim area was sampled in total 100.040
propagules (8337sem/m2) and 4053 (1351m
2) and MR 96987 (10777sem/m
2) in the
APR. In Coité (Capão Comprido) was 74,316 propagules (3539sem/m2) and 5718
(1906sem/m2) and MR 68598 (3811sem/m
2). With the largest quantities represented by
invasive species Urochloa decumbens (Stapf)RD. In areas syndrome dispersion
predominant in the area in the restoration process (APR) was anemocoric and remnant
forest "non anemochory " was prevalent . Predominated seed sizes small (<2mm) in
both areas . Variation occurred in the supply of seed rain between the months being the
peak observed for the Jardim area in April and the Coité was in February . In both areas
the differences between the supply of seed rain in APR (area being restored) and MR
(remaining forest) were observed . In Jardim in the treatments with highest intakes of
seed rain in abundance and richness were T3 (control ) and T4 (remaining forest) ,
respectively (p<0.001) . In Coité area were, respectively, T5 (diversity and coating line)
and T7 (remaining forest) (p < 0.001). With the results obtained for both areas is
possible to observe that the seed rain reflect the different situations in each area
responding to small variations reflecting the conservation status of each environment
and allowing to draw conclusions within the landscape . Because of this accuracy,
sensitivity and measurability seed rain can be used as an indicator of ecological
restoration in riparian areas , allowing the study to extract information about the
regeneration capacity of the ecosystem . Can be used as an indicator of the potential for
regeneration or as an indicator of self-maintenance to support interventions and best
choice of methodologies for restoration .

Sumário
1. INTRODUÇÃO GERAL _________________________________________________ 9
2. METODOLOGIA GERAL _______________________________________________ 13
2.1. Caracterização das áreas de estudo ________________________________________ 13
2.2. Instalação das técnicas de restauração ecológica _____________________________ 14
2.3. Avaliação de indicador ecológico __________________________________________ 17
3. REFERÊNCIAS BIBLIOGRÁFICAS ________________________________________ 18
4. Capítulo 1: CARACTERIZAÇÃO DA CHUVA DE SEMENTES EM TRECHO DE MATA RIPÁRIA EM RESTAURAÇÃO ECOLÓGICA NO CERRADO- DF. ___________________ 20
4.1. Introdução ___________________________________________________________ 22
4.2. Material e métodos ____________________________________________________ 24 4.2.1 Área de estudo ______________________________________________________________ 24 4.2.2. Delineamento experimental ___________________________________________________ 26
4.3. Análise dos dados ______________________________________________________ 28
4.4. Resultados ___________________________________________________________ 28
4.5. Discussão ____________________________________________________________ 37
4.6. Referências Bibliográficas _______________________________________________ 42
5. Capítulo 2 – A CHUVA DE SEMENTES PODE CONTRIBUIR PARA A RESTAURAÇÃO ECOLÓGICA DE MATA RIPÁRIA? _________________________________________ 45
5.1. Introdução ___________________________________________________________ 47
5.2. Material e métodos ____________________________________________________ 49 5.2.1. Área de estudo _____________________________________________________________ 49 5.2.2. Delineamento experimental ___________________________________________________ 51
5.3. Análise dos dados ______________________________________________________ 52
5.4. Resultados ___________________________________________________________ 52
5.5. Discussão ____________________________________________________________ 57
5.6. Referências Bibliográficas _______________________________________________ 62
6. CONCLUSÃO GERAL _________________________________________________ 65
ANEXO 1 ____________________________________________________________ 66
ANEXO 2 ____________________________________________________________ 67
ANEXO 3 ____________________________________________________________ 72

9
1. INTRODUÇÃO GERAL
As matas ripárias são faixas de vegetação que estão sob interferência direta da
presença de água, sobretudo no período chuvoso (RODRIGUES, 1991). Estas matas
exercem influencia sobre a bacia hidrográfica na qual estiver inserida, interferindo no
fluxo de água, conferindo resistência mecânica ao solo, retendo sedimentos, exercendo
estabilidade ao leito e influenciando na morfologia do canal (HICKIN, 1984), sendo
também relevante para a manutenção de ecossistemas aquáticos (GREGORY et al,
1991; GONÇALVES & CALLISTO, 2013).
No bioma Cerrado as matas ripárias vêm passando por modificações
consideráveis devido aos diversos impactos antrópicos, causando diferentes níveis de
degradação, impactando negativamente o ambiente ribeirinho (ALBUQUERQUE et al,
2010). Os impactos podem gerar alterações na diversidade das espécies da fauna e da
flora aquática e terrestre, no fluxo natural de escoamento, nos níveis das águas dos rios,
na recarga do aquífero até erosão dos taludes e encostas (SOUSA & KOBIYAMA,
2003). Essas inúmeras modificações, segundo os mesmos autores, prejudicam o fluxo
hidrológico, difundindo os problemas dentro de toda a estrutura que compõe o
ecossistema, causando uma série de perturbações a esse meio.
A conversão de áreas naturais para fins agropecuários ou urbanos transformam a
paisagem e geram níveis de degradações. De acordo com o nível da perturbação, a mata
ripária perde sua capacidade de resiliência, ou seja, capacidade de retornar através da
regeneração natural ao estado anterior à degradação.
A capacidade de regeneração natural está diretamente relacionada à
oportunidade de colonização e estabelecimento de indivíduos novos recrutados a partir
do banco de sementes e plântulas do solo, da chuva de sementes e/ou de rebrotas
(HUBBELL et al, 1986; ALVES & METZER, 2006). Outro fator chave na capacidade
de regeneração é o nível de fragmentação da paisagem. Com a fragmentação a paisagem
fica composta de manchas de vegetação natural inseridas em matriz antropogênica
(plantações, pastagens, zona urbana). Esta fragmentação acarreta isolamento de áreas de
vegetação natural, podendo influenciar, por exemplo, a dispersão de sementes e por
consequência a colonização (SEOANE, 2005; PIVELLO et al, 2006; JORDANO et al,
2006; STAGGEMEIER & GALETTI, 2007).

10
Em áreas degradadas a presença de fragmentos próximos facilita a chuva de
sementes (entrada de propágulos) e aumentam as chances de restauração da área. A
chuva de sementes representa as sementes ou propágulos (diásporos) que chegam ao
solo através de mecanismos de dispersão (bióticos e abióticos), podendo ser originadas
do próprio fragmento (autóctones) ou de fragmentos afastados da área (alóctones)
(ARAÚJO et al, 2002). Portanto, perturbações nas florestas podem causar perda direta
de animais dispersores ou alterações na estrutura da vegetação. Estes fatos apresentam-
se como limitações ao processo de dispersão.
A limitação de dispersão ocorre quando, independente da quantidade de
sementes produzidas, a quantidade dispersa é limitada pela baixa atividade ou ausência
do dispersor, fazendo com que as sementes alcancem poucos sítios de recrutamento
(BARBOSA et al, 2006). Esses fatores limitantes acabam por restringir a chuva de
sementes, alterando os processos de auto-manutenção, regeneração natural e expansão
da floresta (PIVELLO, 2006), consequentemente o futuro da comunidade vegetal.
A regeneração natural é dependente da produção de sementes viáveis (FENNER,
1985) e a dispersão destas sementes é uma questão-chave na dinâmica da vegetação
(HARPER, 1977). Swaine & Whitmore (1988) observaram que as características das
sementes como o tamanho, tipo de dispersão e dormência podem moldar os padrões da
história de vida das árvores tropicais, determinando onde e como a germinação e o
estabelecimento podem ocorrer.
Para que a regeneração natural ocorra são necessários que os propágulos se
dispersem e tenham capacidade de germinarem e se estabelecerem (DUNCAN &
CHAPMAN, 2002; ALVES & METZER, 2006). Isso pressupõe que o ambiente
degradado tenha condições mínimas viáveis a oferecer ao propágulo para que este
participe efetivamente da dinâmica sucessional do ecossistema (JORDANO, 2006;
PIVELLO et al, 2006).
A dispersão de sementes consiste no processo de remoção da semente da planta-
mãe para locais favoráveis onde a predação e a competição são mais baixas (JANZEN,
1970; JORDANO, 2006). Esse processo é fundamental na dinâmica florestal e no
desenvolvimento da sucessão ecológica. As mudanças na comunidade serão
direcionadas pela chegada e estabelecimento dos diásporos (HARDESTY & PARKER
2002). Portanto, para que uma área perturbada tenha capacidade de retornar as
condições do passado ou para se aproximar das funções ecológicas originais é
necessário adotar estratégias de restauração ecológica (ALBUQUERQUE et al, 2010).

11
Desta forma, o repovoamento com espécies nativas, que tenham capacidade de
maximizar as interações biológicas (e.g. polinização e dispersão de sementes), configura
uma estratégia viável para restaurar ambientes degradados (REIS & KAGEYAMA,
2003;). As matas ripárias na região do Cerrado são pouco conhecidas em muitos
aspectos incluindo na restauração (FELFILI et al, 2000; MODNA et al, 2010), o que
dificulta a definição de metodologias e escolhas de espécies.
Os fragmentos com vegetação natural contribuem com a disponibilidade de
propágulos que poderão ser levados e, eventualmente, recrutados nas áreas degradadas
(TRES et al, 2007), sendo que a distância entre os fragmentos é o fator limitante para a
dispersão de propágulos (TERBORGH, 2013). Este fator influencia a chegada dos
propágulos nas áreas perturbadas e pode afetar a capacidade de regeneração natural
(RODRIGUES et al, 2004). Além da distância dos fragmentos, deve-se levar em conta a
presença de dispersores de sementes que são indispensáveis para acelerarem o processo
de “cicatrização” de áreas degradadas.
A chegada de diásporos e o posterior estabelecimento das sementes dependem
de fatores como o tipo de dispersão, tamanho e qualidade das sementes (SWAINE &
WHITMORE, 1988). As síndromes de dispersão de sementes predominantes na
comunidade permitem inferir sobre a estrutura da vegetação, seu estádio sucessional e
seu grau de conservação (HARPER, 1977). Em florestas tardias observa-se o
predomínio de sementes zoocóricas, enquanto que em áreas abertas com algum grau de
perturbação há predomínio da anemocoria (OLIVEIRA & MOREIRA, 1992;
PINHEIRO & RIBEIRO, 2001). Da mesma maneira, a proporção de zoocoria diminui
de áreas úmidas para áreas secas (GENTRY, 1982).
A variação da chuva de sementes nos ajuda na compreensão da dinâmica
florestal e dos processos reprodutivos envolvidos na manutenção da comunidade
(HOFGAARD, 1993). Os padrões sazonais de frutificação e a variação temporal no
fluxo de propágulos nas florestas tropicais (WHITE, 1994; MORELLATO, 1995)
podem ser fundamentais para determinar a ocupação de uma população em um habitat
(HARPER, 1977).
Para compreender a dinâmica de uma área em processo de restauração, natural
ou induzida, são necessárias avaliações de indicadores ecológicos. Os indicadores
ecológicos são parâmetros biológicos que por suas características qualitativas ou
quantitativas representam o estado do sistema ecológico permitindo inferir sobre suas
características e eventuais mudanças ao longo do tempo (DALE & BEYELER, 2001),

12
devendo ser adequados a escala temporal e espacial (MONOLIADIS, 2002). Os
indicadores ecológicos podem estar relacionados com o clima, aspectos edáficos,
ecológicos, fenológicos, fisiológicos (POGIANNI & GONÇALVES, 1998).
A utilização de indicadores baseia-se na hipótese de que a presença ou ausência,
tal como as flutuações, demonstram as mudanças que ocorrem nos vários níveis
hierárquicos, desde as espécies até ecossistemas (NOON ET AL, 1999). Diante da
dificuldade em trabalhar em vários níveis hierárquicos, um dos principais desafios no
uso de indicadores é encontrar um conjunto que forneça informações interpretáveis
podendo ser usados para compreender as condições ecológicas em seu amplo espectro
de variação (DALE & BEYELER, 2001). Apesar dos desafios, o uso dos indicadores
pode auxiliar na interpretação de mudanças nos estágios iniciais de sucessão, por
exemplo, auxiliando na avaliação da eficiência dos métodos de restauração ecológica e,
consequentemente, subsidiando o aprimoramento dos métodos utilizados (VAN
STRAALEN, 1998).
A chuva de sementes, bem como o banco de sementes e plântulas, acúmulo de
serapilheira e regenerantes representa um indicador ecológico com resultados
mensuráveis e sensíveis às mudanças decorrentes de degradações e de ações de
restauração (SIQUEIRA, 2002; ARAÚJO, 2002; TRES et al, 2007). Neste contexto, a
chuva de sementes pode representar um indicador do potencial de resiliência do
ambiente degradado, podendo sinalizar as inúmeras trajetórias de colonização possíveis
para a área, norteando assim ações corretivas e direcionando melhor os trabalhos de
restauração ecológica.
Diante deste contexto, este trabalho tem por objetivo geral avaliar a chuva de
sementes como indicador de restauração ecológica em áreas de zonas ripárias no
Distrito Federal para subsidiar ações e nortear a escolha de metodologias mais
adequadas para a restauração ecológica. O objetivo específico foi analisar quali-
quantitativamente a chuva de sementes nas áreas remanescentes e nas áreas em processo
de restauração.
As hipóteses que nortearam o trabalho foram: i) os padrões quali-quantitativos
variam da área remanescente para a área degradada, ii) a chuva de sementes indica a
capacidade de regeneração natural caracterizando a quantidade de diásporos que
chegam à área degradada sendo, com isso, um bom indicador de restauração ecológica.

13
2. METODOLOGIA GERAL
2.1. Caracterização das áreas de estudo
Esta dissertação foi desenvolvida dentro do projeto: AQUARIPÁRIA: Restauração
ecológica de ambientes ripários sob influência de atividades agrícolas e urbanas em
mananciais de três bacias hidrográficas, iniciado em novembro de 2011, com o objetivo
de testar metodologias para a restauração ecológica de matas ripárias. Para esta
dissertação foram selecionadas duas destas bacias, sendo uma no rio Preto e outra no rio
Descoberto, Distrito Federal.
As bacias selecionadas estão localizadas em zona rural, altitude de
aproximadamente 1160 m, clima tropical semi-úmido (Aw), segundo classificação de
Köppen, com dois períodos bem definidos: chuvoso e seco. A área da bacia do rio Preto
está localizada a leste do Distrito Federal e às margens do rio Jardim, no Núcleo Rural
Tabatinga, Planaltina, DF (15°44’54,9”S e 47°35’07,7”W). O trecho de mata ripária
desmatado possuía cerca de 130 metros de extensão; A área da bacia do Descoberto está
localizada a oeste do Distrito Federal às margens do córrego Capão Cumprido, no
INCRA 8, Brazlândia , DF (15°44’32,79”S e 48°08’59,81”W). O trecho de mata ripária
desmatada possuía cerca de 330 metros (Figura 1).
Figura 1: Localização das bacias hidrográficas onde as áreas experimentais do Rio Jardim
(Bacia do Rio Jardim) e do Córrego Capão Cumprido (Bacia do Descoberto) estão inseridas,
Distrito Federal.

14
2.2. Instalação das técnicas de restauração ecológica
Para instalação dos experimentos do projeto Aquaripária houve a seleção das áreas
com base em análise multicriterial no contexto da paisagem (OLIVEIRA et al, 2011).
Entre os critérios estabelecidos cabem ressaltar que se levou em consideração a
presença de remanescentes florestais próximos às áreas a serem restauradas, bem como
o interesse do proprietário em restaurar sua área.
Em cada área, rio Jardim e Capão Comprido, ao se selecionar o trecho a ser
restaurado, foi escolhido a mata remanescente mais próxima para servir de ecossistema
referência. Portanto, em cada local havia duas situações distintas: mata remanescente
(MR), considerada como o tratamento referência, e área em processo de restauração
(APR), onde foram instalados tratamentos com as técnicas de restauração ecológica.
Cada tratamento teve três repetições (parcelas), assim na mata remanescente foram
instaladas três parcelas (repetições A, B e C), bem como em cada tratamento da área em
processo de restauração (Tabela 1 e Figura 2).
No rio Jardim e no Capão Comprido foram instalados, respectivamente, quatro
tratamentos (12 parcelas) e sete tratamentos (21 parcelas). No rio Jardim foram
plantadas 522 mudas (15 sp) e no Capão comprido 1089 (16sp) (Anexo 1). Os
tratamentos foram:
a. Mata remanescente
A mata remanescente, adjacente à área em processo de restauração, representou o
controle positivo das condições naturais.
b. Nucleação de Anderson
Consiste no plantio de ilhas de vegetação com o objetivo principal de atração de
fauna. Este método admite distribuição das mudas em núcleos podendo ter diversas
combinações (ANDERSON, 1953).
Neste trabalho o plantio consistiu em núcleos de cinco plantas sendo distribuídas
no formato de cruz (+). Foram utilizadas nas extremidades quatro mudas de arbustivas
da família Melastomataceae, com potencial para a restauração (ALBUQUERQUE et al,
2013) e no centro do núcleo uma muda de arbórea (Figura 2a).

15
c. Nucleação Anderson + Poleiro
Este tratamento combina as metodologias de Nucleação de Anderson e poleiro.
A distribuição dos poleiros na parcela foi sistematizada, sendo que entre núcleos foram
instalados os poleiros artificiais (Figura 2b).
d. Poleiros
Neste tratamento foram instalados poleiros artificiais de eucalipto com três
hastes equidistantes para pouso das aves. Desta forma, podem exercer a função
nucleadora ao atrair aves dispersoras que defecam sementes de espécies presentes nos
remanescentes de matas ripárias próximos a área experimental (Figura 2c).
e. Linhas de recobrimento e diversidade
As mudas foram plantadas em linhas combinando espécies de grupos ecológicos
distintos, boa cobertura, capacidade para atração de fauna e diferentes formas de vida.
Na linha de recobrimento utilizaram-se espécies de crescimento rápido e boa cobertura
de copa e na linha de diversidade plantou-se mudas arbóreas e arbustivas,
preferencialmente com boa capacidade para atração de fauna e espécies de rápido
crescimento, mas que não têm boa cobertura, bem como as secundárias. De tal forma,
que o primeiro grupo pode favorecer o crescimento do grupo da diversidade e
desfavorecem o desenvolvimento de espécies competidoras através do sombreamento
da área de recuperação (NAVE, 2005) (Figura 2d).
f. Controle
Neste tratamento não foi feito nenhum tipo de manejo.

16
Tabela 1: Tratamentos instalados em cada uma das áreas de zonas ripárias, Distrito
Federal.
Jardim Capão Comprido
T1: Nucleação de Anderson 3x3m T1: Nucleação Anderson 3x3m
T2: Linhas de recobrimento e diversidade T2: Nucleação Anderson 5x5m
T3: Controle T3: Poleiro
T4: Mata remanescente T4: Nucleação Anderson +
Poleiro
T5: Linha de recobrimento e
diversidade
T6: Controle
T7: Mata remanescente
Figura 2: Técnicas de restauração ecológica avaliadas
para matas ripárias, Distrito Federal. Técnicas
nucleadoras: nucleação de Anderson (A), nucleação de
Anderson + Poleiro (B), poleiro (C). Linha de
recobrimento e diversidade (D). Círculos verdes: Espécies
arbustivas. Círculos marrons: Espécies arbóreas. Círculos
azuis: Linha de diversidade. Círculos amarelos: Espécies
de recobrimento. Cruz: Poleiros artificiais.

17
2.3. Avaliação de indicador ecológico
Para avaliar o potencial da chuva de sementes como indicador ecológico foi
utilizado o enquadramento dos resultados em alguns aspectos. A partir da avaliação será
empregada a escala de Likert em cinco níveis (SCORIZA et al, 2009), do menos
eficiente (0) para o mais eficiente (5) seguindo os aspectos descritos na tabela 2.
Tabela 2: Aspectos e descritores atribuídos aos indicadores ecológicos para determinar sua eficiência em
avaliar o estado de uma área e monitorar tendências. Adaptado de Segip (1995), Metzger (2002).
ASPECTO DESCRITORES
Sensibilidade Capaz de detectar alterações no ambiente
e diferenças entre eles
Resultabilidade Produz resultados mensuráveis
Custo Baixo custo de implantação
Compreensão e interpretação Capaz de ser compreendido e interpretado
com facilidade
Previsibilidade ou tendência
Indicam claramente tendências em
resposta a alterações no ambiente ao
longo do tempo, permitindo a avaliação e
monitoramento
Escala Apresentam valores e parâmetros claros
que podem ser aplicados como referência
para avaliar o estado de cada área e suas
alterações
Síntese Permite a formulação de critérios que
reúnem vários atributos do ecossistema
ou área, possibilitando uma análise única
de várias características ou estados das
áreas foco.
Este trabalho de dissertação constitui-se de dois capítulos que vão gerar dois
artigos científicos. A metodologia mais detalhada esta descrita para cada capítulo, sendo
que aspectos que versam sobre a metodologia geral serão direcionados com os dizeres
“ver metodologia geral”.

18
3. REFERÊNCIAS BIBLIOGRÁFICAS ALBUQUERQUE, L.B.; AQUINO, F.G.; ALONSO, A.M. ; LIMA, J.E.W. ; BRAGA; SOUSA-SILVA,
J.C. Restauração Ecológica de Matas Ripárias: uma questão de sustentabilidade. Série Documentos.
Brasília: Embrapa Cerrados, 75p, 2010.
ALVES, L.F. & METZGER, J.P. A regeneração florestal em áreas de floresta secundária na reserva
Florestal do Morro Grande, Cotia, SP. Biota Neotropica, 2006.
ANDERSON, M.L. Spaced-group planting. Unasylva. v. 7, n. 2, p. 12-18, 1953.
ARAÚJO, R.S de. Chuva de sementes e deposição de serapilheira em três sistemas de revegetação de
áreas degradadas na Reserva Biológica de Poço das Antas (RJ). 2002. 92f. Dissertação (Mestrado em
Ciências Ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro, Seropédica, 2002.
BARBOSA, K.C.; PIZO, M.A. Seed rain and seed limitation in a planted Gallery Forest in Brazil.
Restoration Ecology, v.14, n. 4, p. 504-515, 2006.
ALBUQUERQUE, L.B.; AQUINO, F.G.; COSTA, L.C.; MIRANDA, Z.J.; & SOUSA, S.R. Espécies de
Melastomataceae JUSS. com potencial para restauração ecológica de mata ripária no Cerrado.
Polibotánica, n. 35, p. 1-19, 2013.
DALE, V.H.; BEYELER, S.C. Challenges in the development and use of ecological
indicators. Ecological indicators, n. 1.1, p.3-10, 2001.
DUNCAN, R. S., & CHAPMAN, C. A. Seed dispersal and potential forest succession in abandoned
agriculture in tropical Africa. Ecological Applications, v. 9, n. 3, p. 998-1008, 1999.
DURIGAN, G. . O uso de indicadores para monitoramento de áreas em recuperação. Cadernos da Mata
Ciliar, v. 4, p. 11-13, 2011.
FELFILI, J. M., RIBEIRO, J. F., FAGG, C. W., MACHADO, J. W. B. Recuperação de matas de galeria.
Embrapa Cerrados. n. 21, p. 1-45, 2000.
FENNER, M. Seed ecology. Chapman and Hall. 1985.
FREIRE, M. Chuva de sementes, banco de sementes do solo e deposição de serapilheira como
bioindicadores ambientais no bioma Mata Atlântica, Teresópolis, RJ. 2006. 81 f. Dissertação
(Mestrado em Recursos Ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro,
Seropédica. 2006.
GENTRY, A.H. Patterns of neotropical plant species diversity. Evolution Biology. n.15, p. 1-84, 1982.
GONÇALVES JR, J. F., & CALLISTO, M. Organic-matter dynamics in the riparian zone of a tropical
headwater stream in Southern Brasil. Aquatic Botany, n. 109, p. 8-13, 2013.
GREGORY, S.V.; SWANSON, F.J.; MCKEE, W.A.; CUMMINS, K.W. An ecosystem perspective of
riparian zones. Focus on links between land and water. BioScience, v. 41, p. 540-551, 1991.
HARDESTY, B.D. & PARKER, V.T. Community seed rain patterns and a comparison to adult
community structure in a West African tropical forest. Plant Ecology. n. 164, p. 49–64, 2002.
HARPER, J. L. Population biology of plants. Population biology of plants. 1977.
HICKIN, E.J. Vegetation and river channel dynamics. The Canadian Geographer, v. 28, p. 111-126,
1984.
HOFGAARD, A. Seed rain quantity and quality, 1984-1992, in a high altitude old-growth spruce forest,
northern Sweden. New Phytologist, n. 125, p. 635-640. 1993.
HUBBELL, S.P.; FOSTER, R.B. Biology, chance, and history and the structure of tropical rain forest tree
communities. Community ecology, p. 314-330, 1986.
JANZEN, D. H. Herbivores and the number of tree species in tropical forests. American naturalist, p.
501-528, 1970.
JORDANO, P.; GALETTI, M.; PIZO, M. A.; & SILVA, W. R. Ligando frugivoria e dispersão de
sementes à biologia da conservação. In: Duarte, C.F.; Bergallo, H.G.; Dos Santos, M.A.; and V a,
A.E. (eds.). Biologia da conservação: essências. Editorial Rima, São Paulo, Brasil. 2006, p. 1-436.
MANOLIADIS, O.G. Development of ecological indicators a methodological framework using
compromise programming. Ecological indicators, n. 2, p. 169-176, 2002.
METZGER, J. P.; CASATTI, L.; VERDADE, L. Indicadores de conservação e avaliação do
conhecimento para conservação. In: I Workshop de Síntese do Programa BIOTA/FAPESP na
Universidade de São Carlos, SP, 2002. Relatório do grupo de trabalho. Disponível em:
<http://www.biota.org.br/info/historico/simp2002/gt2.pdf>. Acesso em: 21 set. 2013.
MODNA, D.; DURIGAN, G.; & VITAL, M. V. C. Pinus elliottii Engelm como facilitadora da
regeneração natural da mata ciliar em região de Cerrado, Assis, SP, Brasil. Scientia Forestalis, v. 38,
n. 85, p. 73-83, 2010.

19
MORELLATO, P.C. As estações do ano na floresta. In: H.F. Leitão Filho & L.P. Morellato (eds.).
Ecologia e preservação de uma floresta tropical urbana - Reserva de Santa Genebra. Campinas,
Editora da UNICAMP. 1995.
NAVE, A. G. Banco de sementes autóctone e alóctone, resgate de plantas e plantio de vegetação nativa na
Fazenda Intermontes, município de Ribeirão Grande, SP. 2005. 218 f. Tese (Doutorado) - Escola
Superior de Agricultura Luiz de Queiroz. Universidade de São Paulo. Piracicaba, 2005.
NOON, B.R.; SPIES, T.A.; RAPHAEL, M.G. Conceptual basis for designing an effectiveness monitoring
program. In: Mulder, B.S., et al. (Eds.). The Strategy and Design of the Effectiveness Monitoring
Program for the Northwest Forest Plan. US Department of Agriculture, Forest Service, Gen. Tech.
Rep. PNW-GTR-437, Portland, OR, 1999, p. 21-48
OLIVEIRA, C. H.; SAITO, C. H; FETTER, R. http://www.ecoa.unb.br/aquariparia/. Acessado em
outubro de 2011.
OLIVEIRA, P.E.A.M.; MOREIRA, AG. Anemocoria em espécies de cerrado e Mata de Galeria de
Brasilia, DF. Revista Brasileira de Botânica, São Paulo, v. 15, n. 2, p. 163-174, 1992.
PIJL, LEENDERT. Principles of dispersal in higher plants. Springer-Verlag. 1982.
PINHEIRO, F.; RIBEIRO, F.R. Síndromes de dispersão de sementes de matas de galeria do Distrito
Federal. In: Ribeiro, F.R.; Fonseca, C.E.L.; Sousa-Silva, J. C. Cerrado: Caracterização e
recuperação de matas de galeria. Embrapa Cerrados. Brasília. p. 899, 2001.
PIVELLO, V. R.; PETENON, D.; JESUS, F. M. D.; MEIRELLES, S. T.; VIDAL, M. M.; ALONSO, R.
D. A. S.; FRANCO, G. A. D. C. & METZGER, J. P. Chuva de sementes em fragmentos de Floresta
Atlântica (São Paulo, SP, Brasil), sob diferentes situações de conectividade, estrutura florestal e
proximidade da borda. Acta Botanica Brasilica, v. 20, n. 4, p. 845-859. 2006.
POGGIANI, F.; STAPE, J. L.; DE MORAES GONÇALVES, J. L. Indicadores de sustentabilidade das
plantações florestais. Série técnica IPEF, v. 12, n. 31, p. 33-44. 1998.
REIS, A.; KAGEYAMA, P.Y. Restauração de áreas degradadas utilizando interações interespecíficas. In:
Kageyama, P.Y.; Oliveira, R.E.; Moraes, L.F.D.; Engel, V.L & Gandara, F.B. Restauração ecológica
de ecossistemas naturais. Botucatu, FEPAF, p. 91-110. 2003.
RODRIGUES, R.R. Análise de um remanescente de vegetação natural às margens do Rio Passa Cinco,
Ipeúna, SP. 1991. 325f. Tese de doutorado, Universidade Estadual de Campinas, Campinas. 1991.
RODRIGUES, R. R.; MARTINS, S. V.; BARROS, L. C. Tropical rain forest regeneration in an area
degraded by mining in Mato Grosso state, Brazil. Forest Ecology and Management, v. 190, p. 323-
333, 2004.
SCORIZA, R.N. Serrapilheira como indicador ambiental aplicado na avaliação de fragmentos florestais
em sorocaba, 87f. 2009. Monografia de conclusão de curso (Universidade Federal de São Carlos)-
Sorocaba. 2009. SEGIP. The state enviromental Goals and Indicators Project. Evaluation criteria. In: BERGQUIST, G. et
al. Prospective indicators for state use in performance agreements. Florida: SEGIP/Florida Center
for public Management, 1995. p. B6-B7.
SEOANE, C.E.S; KAGEYAMA, P.Y.; RIBEIRO, A.; MATIAS, R.; REIS, M.S.; BAWA, K. Efeitos da
fragmentação florestal sobre a imigração de sementes e a estrutura genética temporal de populações de
Euterpe edulis Mart. Rev. Inst. Flor. São Paulo, v. 17, n. 1, p. 25-43, 2005.
SIQUEIRA, L.P. Monitoramento de áreas restauradas no interior do Estado de São Paulo, Brasil.
2002. 116 f. Dissertação (Mestrado em Recursos Florestais) - Escola Superior de Agricultura Luiz de
Queiroz, Universidade de São Paulo, Piracicaba. 2002.
SOUZA, D. P; KOBIYAMA, M. Ecoengenharia em zona ripária: renaturalização de rios e recuperação de
vegetação ripária. I Seminário de Hidrologia Florestal, p. 121, 2003.
STAGGEMEIER, V. G., & GALETTI, M. Impacto humano afeta negativamente a dispersão de sementes
de frutos ornitocóricos: uma perspectiva global. Revista Brasileira de Ornitologia, v.15, n. 2, p. 281-
287. 2007.
SWAINE, M.D.; & WHITMORE, T.C. On the definition of ecological species groups in tropical rain
forests. Vegetatio, v. 75, n. 1-2, p. 81-86, 1988.
TERBORGH, J. Using Janzen–Connell to predict the consequences of defaunation and other disturbances
of tropical forests. Biological Conservation, n.163, p.7-12. 2013
TRES, D.R.; ANNA. C.S.S.; BASSOS,. S.; LANGA, R.; JUNIOR, U. R.; REIS, A. Banco e Chuva de
Sementes como Indicadores para a Restauração Ecológica de Matas Ciliares. Revista Brasileira de
Biociências, Porto Alegre, v. 5, n. 1, p. 309-311, jul, 2007.
VAN STRAALEN, N.M. Evaluation of bioindicator systems derived from soil arthropod
communities. Applied Soil Ecology. v. 9, n. 1, p. 429-437. 1998.
WHITE, L.J.T. Patterns of fruit-fall phenology in the Lopé Reserve, Gabon. Journal of Tropical
Ecology, n. 10, p. 289-312, 1994.

20
4. Capítulo 1: CARACTERIZAÇÃO DA CHUVA DE SEMENTES EM TRECHO
DE MATA RIPÁRIA EM RESTAURAÇÃO ECOLÓGICA NO CERRADO-
DF.
Resumo
A dispersão de sementes é um processo fundamental na dinâmica florestal. A chegada e
provável estabelecimento de diásporos de diferentes espécies irão direcionar o
desenvolvimento sucessional e as mudanças na comunidade. O conhecimento da
variação da chuva de sementes é importante, pois contribui para a compreensão dos
processos reprodutivos e da dinâmica dos ecossistemas, além de subsidiar ações de
restauração em ambientes degradados. O objetivo deste trabalho foi analisar a chuva de
sementes como indicador de restauração ecológica em trecho de mata ripária do rio
Jardim, DF-Brasil. Para tal foi analisado quali-quantitativamente a chuva de sementes
nas áreas remanescentes e nas áreas em restauração, assim como caracterizado o aporte
da chuva de sementes em cada técnica de restauração. A área experimental localiza-se a
leste do Distrito Federal às margens do rio Jardim. O trecho de mata ripária a ser
restaurada possui 0,7ha. Este trecho foi utilizado durante muito tempo para agricultura,
após este período foram plantadas mudas na área visando sua recuperação, por iniciativa
do proprietário. A área experimental encontra-se inserida em uma matriz agrícola,
porém adjacente a área experimental ocorrem vegetações nativas distintas (mata de
galeria e vereda descaracterizada). Foram instalados 48 coletores no total, com 0,25 m²
e malha de 1 mm² a 15 cm do solo, sendo 12 instalados na mata remanescente (MR) e
12 em cada tratamentos (T1: Nucleação de Anderson, T2: Linha de recobrimento e
diversidade e T3: Controle) da área em processo de restauração (APR). O material
depositado nos coletores foi recolhido mensalmente, de fevereiro de 2012 a janeiro de
2013, em sacos de papel e triado com auxílio de estereomicroscópio no Laboratório de
Biologia Vegetal da Embrapa Cerrados. Do material foram separadas as sementes,
sendo estas quantificadas e identificadas em morfo-espécie. Nos 12 meses avaliados
foram coletados 100.040 propágulos no total, sendo 4.053 (338sementes/m2) na MR e
96.987 (8.082sementes/m2) na APR. A família mais abundante na APR foi a Poaceae. A
espécie exótica Uruchloa decumbens foi a mais abundante na área em restauração
(64%). Ocorreu variação no aporte de chuva de sementes entre os meses, sendo o pico
foi no mês de abril. Nos meses de maio e agosto foram registradas as maiores riquezas
de diásporos não anemocóricos e anemocóricos na APR, respectivamente 54 e 40%. Na
MR ambos os picos foram registrados no mês de agosto. Dos diáporos coletados 21%
foram menores que 2mm e as sementes representaram 92% dos diásporos coletados. O
tratamento que apresentou maior aporte (abundância) de diásporos foi o T3 (controle)
(4936sem/m2) (p<0,001, GL:8), em relação à riqueza o tratamento que apresentou maior
aporte foi a MR (T4) (49 morfoespécies), seguido de T2 com 35 morfoespécies. Os
dados mostram que houve diferenças na abundância, riqueza, síndrome de dispersão
entre as áreas APR e MR. Ao se analisar a chuva de sementes como indicador de

21
restauração ecológica, verificou-se que ela reflete as pequenas diferenças existente entre
tratamentos, representando fidedignamente as condições do meio e permitindo inferir
conclusões no âmbito da paisagem. Devido a essa acurácia, sensibilidade,
mensurabilidade a chuva de sementes pode ser utilizada como indicador de restauração
ecológica em áreas ripárias, permitindo extrair do estudo informações sobre a
capacidade de regeneração do ecossistema.
Abstract
Seed dispersal is a key process in forest dynamics . The likely arrival and establishment
of propagules of different species will drive the development and successional changes
in the community . Knowledge of the variation of seed rain is important because it
contributes to the understanding of reproductive processes and dynamics of ecosystems
and subsidize restoration in degraded environments . The goal was to analyze
qualitative and quantitative seed rain in the remaining areas and in restoration areas and
characterize the contribution of seed rain in each restoration technique located on the
banks of the Rio Jardim (Distrito Federal, Brazil). The stretch of riparian forest to be
restored has 0.7 ha , this passage was used for farming long after this period seedlings
were planted in seeking recovery area , initiated by the owner. The experimental area is
embedded in an agricultural matrix, but adjacent to the experimental area distinct native
vegetation ( gallery forest and uncharacterized palm swamp ) occur . 48 collectors were
installed overall, with 0.25 m² and mesh size of 1 mm ² to 15 cm of soil , with 12
installed in the remaining forest ( MR ) and 12 in each treatment ( T1 : nucleation
Anderson , T2 : Line coverage and diversity and T3 : control ) of the restoration site (
APR ) . The material deposited in the traps was collected monthly from February 2012
to January 2013, in paper bags and screened with the stereomicroscope at the
Laboratory of Plant Biology EMBRAPA Cerrados . The thin material seeds were
separated , quantified and identified to the level of morpho- species. In the 12 months
reviews 100.040 seedlings were collected in total, with 4.053 ( 338 seeds/m2 ) in the
forest and 96.987 ( 8082seeds/m2 ) at the restoration site . The most abundant family in
the APR was the Poaceae . The exotic species Uruchloa decumbens was the most
abundant in the restoration site ( 64 % ). Variation occurred in the supply of seed rain
between the months , with the peak in April . In the months of May and August were
recorded greater riches and not anemocorics diasporas in APR, respectively 54 and 40
% . MR in both peaks were recorded in August . 21 % of diaporos collected were
smaller than 2mm and seeds accounted for 92 % of the seeds collected. The treatment
that showed higher uptake ( abundance) of diaspore was the " 3 " ( 4936seeds/m2 ) ( p <
0.001 ) , compared to the wealth treatment showed the greatest contribution was the MR
( T4 ) ( 49 morphospecies ), then T2 with 35 morphospecies . The data evaluated
differences in the abundance, richness , dispersal syndrome between APR and MR areas
. From the study it was noted that the seed rain reflect even the smallest differences
between existing treatments, faithfully representing the environmental conditions and
allowing to draw conclusions even within the landscape. Because of this accuracy,
sensitivity, measurability, seed rain can be used as an indicator of ecological restoration
in riparian areas, allowing the study to extract information about the regeneration
capacity of the ecosystem .

22
4.1. Introdução
As matas ripárias são vegetações florestais que acompanham os cursos de água e
desempenhando funções ecológicas indispensáveis a manutenção da biodiversidade
(RIBEIRO & WALTER, 1998; RODRIGUES et al, 2000; ALBUQUERQUE, et al
2010). Estas matas destacam-se por desempenhar funções ecológicas, como auxiliar na
manutenção dos corpos de água, proteção de nascentes, conservação do solo,
sustentação de taludes, contensões de erosões, manutenção de condições a fauna
(ATTANASIO et al, 2012; AQUINO & VILELA, 2008).
As matas ripárias são protegidas por legislação vigente específica (Lei nº 12.651,
de 25 de maio de 2012), porém continuam sendo alvo de degradações e são reduzidas
pelas ações antrópicas. Dependendo do nível de degradação as matas necessitam de
técnicas restauradoras (FELFILI et al, 2000) para auxiliar na maximização de interação
biológicas como polinização e dispersão (REIS & KAGEYAMA, 2003).
A chuva de sementes é o processo pelo qual ocorre entrada de propágulos no
sistema. Esta entrada depende de alguns fatores tais como: distância e qualidade das
fontes (fragmentos remanescentes) e presença de agentes dispersores, os quais
determinarão o fluxo de diásporos ao longo do ano (PIVELLO et al, 2006; JORDANO,
2006). Paralelamente, as características das sementes como o tamanho, tipo de dispersão
e dormência podem moldar os padrões da história de vida das árvores tropicais,
determinando o futuro da comunidade vegetal (SWAINE & WHITMORE, 1988). Estes
fatores por sua vez irão influenciar no processo de restauração ecológica das matas.
A fragmentação de ecossistemas naturais pode ser um dos motivos de limitação
à chuva de sementes podendo impossibilitar os processos de movimentação de animais
frugívoros, influenciando a relação entre plantas e seus dispersores de sementes (PIZO
& SANTOS, 2011). A fragmentação decorre, sobretudo, da ampliação de áreas
agrícolas, resultando em isolamento dos habitats remanescentes, comprometendo a
dispersão de sementes e a regeneração da vegetação (MAGNAGO et al, 2012). Alguns
fatores como árvores isoladas na área a ser restaurada possibilitam a conexão da
paisagem diminuindo a limitação à dispersão, possibilitando a visitação da fauna
dispersora à área (HOLL, 1998). Do ponto de vista de restauração, árvores isoladas

23
podem atrair dispersores de sementes de fragmentos florestais próximos, assim agindo
como focos de recrutamento de plantas florestais e contribuindo para a expansão da
floresta em pastagens abandonadas (GUEVARA & LABORDE, 1993; SLOCUM &
HORVITZ, 2000; HERRERA & GARCIA, 2009).
Neste contexto, o estudo da chuva de sementes é importante para se entender o
processo de entrada dos futuros indivíduos de uma comunidade vegetal sendo, portanto
indicador do potencial de regeneração das florestas tropicais (HOWE &
SMALLWOOD, 1982; JORDANO et al, 2006; PIVELLO et al, 2006; TRES et al,
2007). A sustentabilidade de ecossistemas florestais depende da manutenção dos
processos de regeneração natural das espécies vegetais (DARONCO et al, 2013).
O ecossistema restaurado é aquele que atingiu o nível esperado de
biodiversidade e sustentabilidade, sendo possível representar tais parâmetros através de
alguns atributos mensuráveis, como por exemplo, aporte de serapilheira, taxa de
recrutamento e mortalidade (DURIGAN et al, 2011). Para se avaliar a possibilidade de
um ecossistema em restauração pelo menos aproximar-se de um ecossistema de
referência é necessário realizar o monitoramento dos indicadores ecológicos.
Os indicadores ecológicos são aqueles que permitem interpretar as condições do
meio, determinando as melhores medidas de intervenção na área e possibilitando
maiores chances de restauração de processos ecológicos (DURIGAN, 2011). Segundo
Rodrigues & Gandolfi (1998) a biodiversidade, equabilidade, fluxo e ciclagem de
nutrientes, fauna de solo e outros elementos, são indicadores ambientais promissores
para a avaliação de áreas naturais e restauradas.
A chuva de sementes tem sido estudada por alguns autores para entendimento do
aporte em diferentes sistemas (ARAUJO et al, 2002; RUDGE et al, 2008 entre outros),
porém poucos estudos levantam a questão do uso da chuva de sementes como indicador
de restauração ecológica (FREIRE, 2006; TRES et al, 2007). Tendo em vista a
importância da chuva de sementes para a manutenção dos ecossistemas e o seu
potencial de indicador ecológico, este trabalho visa caracterizar a chuva de sementes em
área em processo de restauração e área de mata remanescente com o objetivo de buscar
informações que permitam avaliar a chuva de sementes como um indicador de
restauração ecológica em zona ripária do Distrito Federal.

24
4.2. Material e métodos
4.2.1 Área de estudo
A área experimental localiza-se a leste do Distrito Federal às margens do rio
Jardim, no Núcleo Rural Tabatinga, Planaltina-DF (15°44’54,9”S e 47°35’07,7”W). O
trecho de mata ripária a ser restaurada possui 0,7 ha. Neste trabalho a área foi
denominada “Jardim”.
A sub-bacia do Rio Jardim apresenta solos dos tipos latossolos, nitossolos,
cambissolos, plitossolos, gleissolos e neossolos (REATTO et al, 2000) e relevo
variando entre plano e fortemente-ondulado (SPERA et al, 2002). Avaliação realizada
na Bacia do Rio Jardim detectou que na área predominam solos com aptidão para
lavouras anuais e perenes (SPERA et al, 2002). Na área experimental a classe de solo
presente foi o Plintossolo Háplico (REATTO, 2014 com. pessoal).
Esta área do Jardim encontra-se numa matriz agrícola composta de solo exposto
em grandes proporções e apenas uma pequena área é representada por remanescentes da
vegetação nativa. Este panorama indica baixa oferta de ambientes naturais com notável
histórico de conversão de áreas naturais em agrícolas (HENKE, 2014 com. pessoal).
A leste da área em processo de restauração (APR) encontra-se a mata
remanescente (MR) sendo caracterizada pela predominância de espécies arbóreas
formando um dossel continuo, com sub bosque pouco denso, com poucos regenerantes,
geralmente, de pequeno porte. A MR apresenta sinais de perturbação como presença de
lianas e solo exposto em alguns pontos. As matas ripárias do Rio Jardim são
classificadas como mal-drenadas (SILVA JUNIOR, 2001). A oeste da APR ocorre uma
pequena faixa de vegetação que parece uma vereda descaracterizada apresentando
indivíduos isolados de Mauritia flexuosa. Esta descaracterização provavelmente foi
devido à instalação de drenos a oeste da APR, quando há alguns anos atrás a área era
utilizada para fins agrícolas. Na propriedade do Rio Jardim, adjacente a APR, existe
área que passou por processo de restauração (plantio de mudas) no passado. Além disso
a propriedade apresenta pomar comercial constituído principalmente de fruteiras
cítricas, a sudoeste da APR. (Figura 1.1).

25
Figura 1.1: Localização da área do Jardim, Distrito Federal, Brasil. No detalhe. A) área em processo de
restauração as margens do Rio Jardim. B) mata remanescente, Brasil, Distrito Federal, Planaltina. (Fonte:
Google Earth, imagem de: 10/12/2013. Acesso em: 10/02/2014).
A APR é um ambiente heterogêneo caracterizando-se pela presença de
indivíduos arbóreos isolados que foram mantidos na área experimental. A área apresenta
arbustos esparsos, porém é dominada por herbáceas. Este trecho foi utilizado durante
muito tempo para agricultura. Posteriormente, por iniciativa do proprietário, foram
feitas algumas tentativas para a recuperação da área com plantios de mudas de espécies
nativas e o controle da braquiária era feito com herbicidas (Figura 1.2).
Figura 1.2: Área em processo de recuperação (APR) do rio
Jardim, Distrito Federal, antes da instalação do experimento.

26
4.2.2. Delineamento experimental
A chuva de sementes foi avaliada com uso de coletores de diásporos (frutos e
sementes) confeccionados com madeira, tela de nylon de 1mm2
com 0,25 m2
de área
(Figura 1.3). Na diagonal de cada parcela foram instalados quatro coletores a 15 cm do
solo (Figura 1.4A). Sendo portanto 12 coletores/tratamento. Cada parcela foi
considerada uma unidade amostral, ou seja, o conjunto de quatro coletores,
representando um total de 1 m2
de amostragem por parcela.
Figura 1.3: Coletor de sementes instalados nas áreas
experimentais para coleta da chuva de sementes.
Na mata remanescente, por ser uma faixa estreita ao longo do rio, as parcelas
instaladas foram retangulares, mas com as mesmas dimensões da área em processo de
restauração. Portanto, os coletores foram dispostos equidistantes, respeitando a
disposição espacial destas parcelas e de forma a amostrar toda a variação da chuva de
sementes na parcela seguindo a mesma lógica da área em processo de restauração
(Figura 1.4B), mantendo desta forma o mesmo esforço amostral.
Figura 1.4: Desenho esquemático mostrando a disposição dos coletores de sementes nos tratamentos
instalados na APR (A) e no tratamento instalado no fragmento de mata ripária (B).

27
O material contido nos coletores foi recolhido mensalmente de fevereiro de 2012
a janeiro de 2013, sendo armazenado em sacos de papel e, posteriormente, triado em
laboratório. O material foi secado à sombra e à temperatura ambiente, pois esta
metodologia não danifica a morfologia das sementes facilitando a triagem e a
identificação.
A triagem ocorreu em duas etapas: grossa e fina. Pela triagem grossa foram
separados, utilizando-se peneiras (malha 2 mm), materiais grossos (folhas, galhos e
outros), dos materiais finos (pó que passou pela peneira). Este “pó” foi triado no
estereomicroscópio (triagem fina) separando-se sementes e frutos. Quando foram
observadas sementes e/ou frutos maiores, que não passaram pela malha da peneira, estes
foram separados manualmente. Os diásporos (sementes e frutos) foram contados e
morfoespeciados para posterior identificação. As sementes e os frutos também foram
desenhados, fotografados, medidos e separados em dois grupos: anemocóricas ou não
anemocóricas. Para identificação das sementes utilizou-se literatura especializada,
consulta à carpoteca do Herbário da Embrapa Cenargen (DF), comparação com material
coletado em estudo florístico realizado na mesma área de estudo e auxilio de
especialistas. A nomenclatura científica das espécies foi revisada seguindo o Mobot
(Missouri Botanical Garden).
Para facilitar a contagem, as sementes de braquiária (Urochloa
decumbens (Stapf R.D.) foram processadas no aparelho Seed sorter para separação das
cascas e sementes. Após este processamento, a quantidade de sementes em cada amostra
foi estimada por média de sementes em proveta de 5 ml. Toda a triagem foi realizada no
Laboratório de Biologia vegetal da Embrapa Cerrados, Distrito Federal.
A avaliação do indicador ecológico foi realizada através da análise de aspectos
sugeridos por SEGIP (1995) e Metzger (2002) (ver item 2.3, metodologia geral).

28
4.3. Análise dos dados
Para a análise os dados obtidos foram submetidos à construção de modelos
lineares generalizados (GLMs) utilizando-se o software R versão 3.0 (R Development
Core Team, 2008). O aporte da chuva de sementes, riqueza e abundância (variáveis
respostas), foi comparado entre os diferentes tratamentos (variável explicativa) de
restauração ecológica. Todos os GLMs foram submetidos à análise residual para avaliar
a adequação da distribuição dos erros (CRAWLEY, 2007). Também através do software
R foram calculados o coeficiente de equidade de Pielou (J), para analisar a diversidade
da comunidade estudada (BROWER & ZAR, 1984), e o índice Shanon-Weaver (H’).
4.4. Resultados
Na avaliação da chuva de sementes durante um ano foram coletados no total
100.040 diásporos, representado uma densidade de 8.336,50 sem/m², pertencentes a 158
morfoespécies (Figura 1.5). Destas, 29 foram identificadas em espécie, 25 em gênero,
18 em família e 86 não identificados (Anexo 2). Na área estudada o índice de
diversidade Shannon-Weaver (H’) e de equidade de Pielou (J) foram, respectivamente,
1,67 e 0,33.

29
Figura 1.5: Algumas das espécies coletadas na chuva de sementes na área Jardim, Distrito Federal.
a) Ageratum conyzoides L. (Asteraceae), b) Cenchrus polystachios (L.) Morrone (Poaceae), c) Borreria
alata (Aubl.) DC. (Rubiaceae), d) Cecropia polystachya Trécul (Urticaceae), e) Terminalia
glabrescens Mart. (Combretaceae), f) Crotalaria incana L. (Fabaceae), g) Schefflera grandiflora (A.C.
Sm.) Frodin (Araliaceae), h) Spermacoce latifolia Aubl. (Rubiaceae), i) Hyptis lophantha Mart. ex
Benth. (Lamiaceae), j) Miconia cuspidata Mart. ex Naudin (Melastomataceae), k) Stylosanthes
viscosa (L.) Sw. (Fabaceae), l) Fimbristylis miliacea (L.) Vahl (Cyperaceae).
Na área em processo de restauração (APR) foram coletados 96.987 diásporos
(10.777sem/m2). A família mais abundante foi a Poaceae sendo representada
principalmente pela espécie Urochloa decumbens (Stapf) R.D. Webster (Syn.
braquiária) com 64% do total de diásporos coletados. Excluindo-se a braquiária, do total
amostrado para a APR, foram coletados 2.902,5 sem/m². Na mata remanescente (MR)
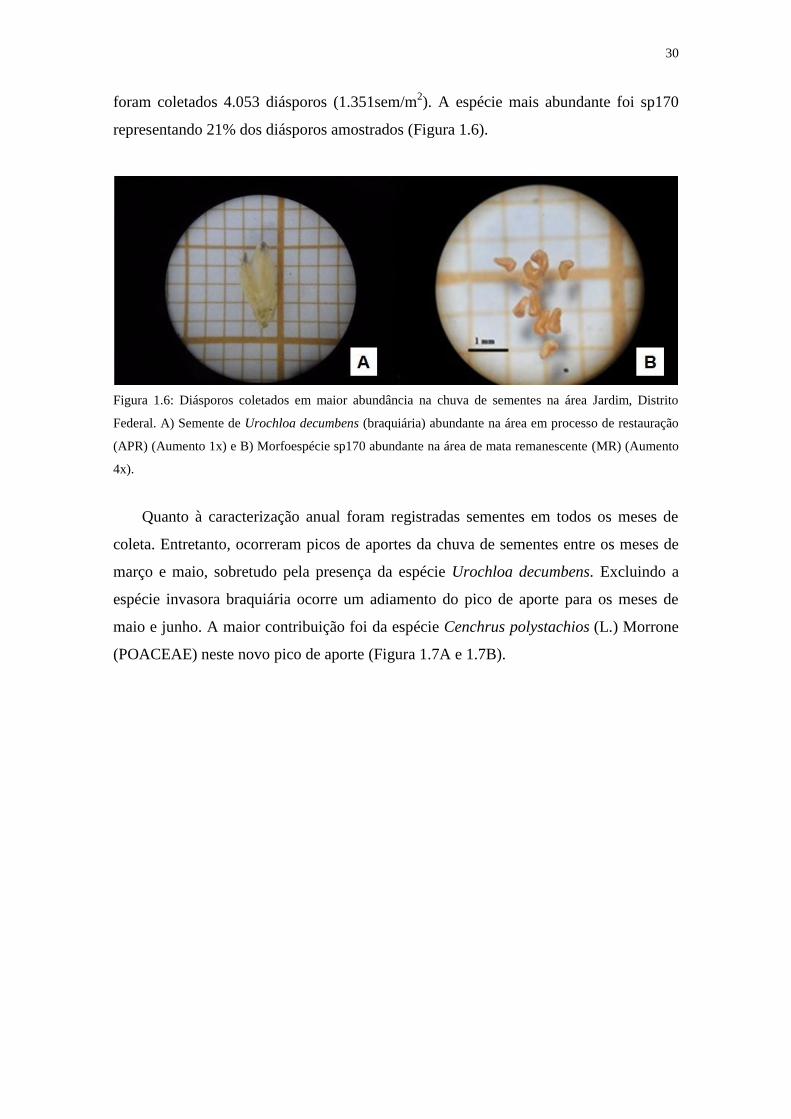
30
foram coletados 4.053 diásporos (1.351sem/m2). A espécie mais abundante foi sp170
representando 21% dos diásporos amostrados (Figura 1.6).
Figura 1.6: Diásporos coletados em maior abundância na chuva de sementes na área Jardim, Distrito
Federal. A) Semente de Urochloa decumbens (braquiária) abundante na área em processo de restauração
(APR) (Aumento 1x) e B) Morfoespécie sp170 abundante na área de mata remanescente (MR) (Aumento
4x).
Quanto à caracterização anual foram registradas sementes em todos os meses de
coleta. Entretanto, ocorreram picos de aportes da chuva de sementes entre os meses de
março e maio, sobretudo pela presença da espécie Urochloa decumbens. Excluindo a
espécie invasora braquiária ocorre um adiamento do pico de aporte para os meses de
maio e junho. A maior contribuição foi da espécie Cenchrus polystachios (L.) Morrone
(POACEAE) neste novo pico de aporte (Figura 1.7A e 1.7B).

31
Figura 1.7: Abundância total de diásporos amostrados mensalmente na área do Jardim, Distrito Federal.
Considerando e desconsiderando a braquiária da amostra.
Considerando-se o total de diásporos a chuva de sementes foi representada por 8%
de frutos (n=7.674) e 92% de sementes (n=92.366). Os diásporos apresentaram
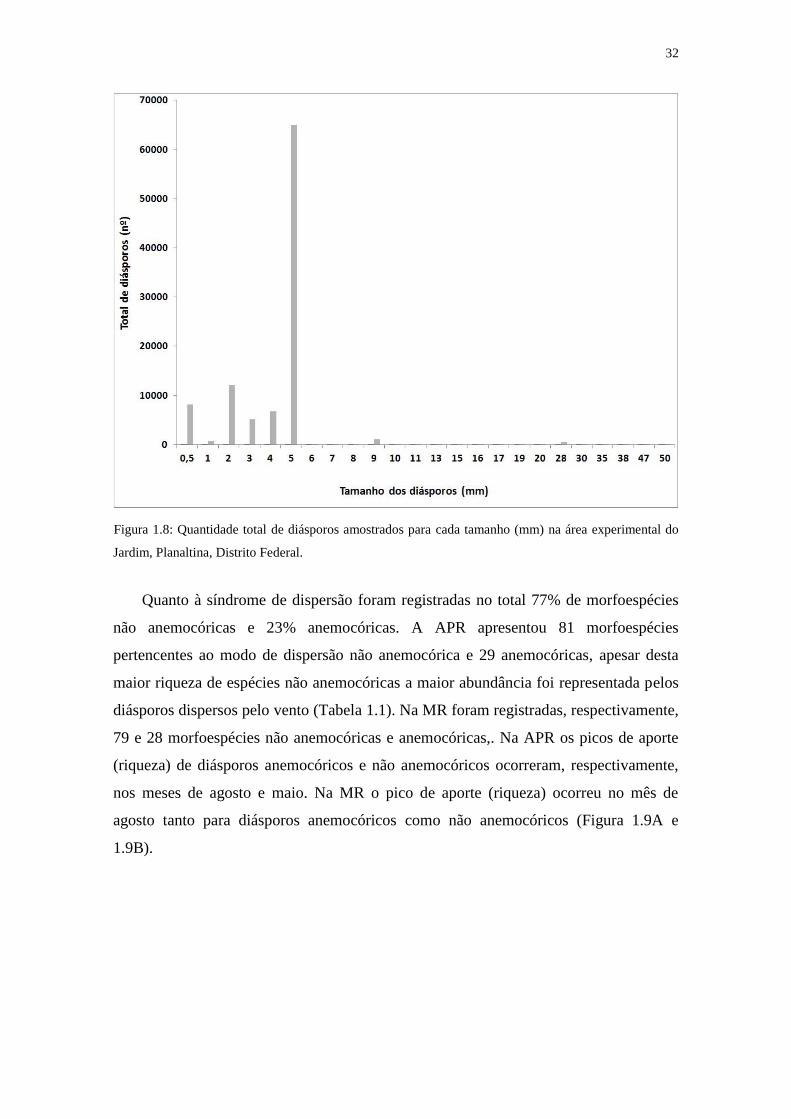
tamanhos diversos entre menor que 1 mm e 5cm. Foram coletados 21% de diásporos
menores (ou igual) a 2 mm e 0,7% maiores que 1 cm (Figura 1.8).
32
Figura 1.8: Quantidade total de diásporos amostrados para cada tamanho (mm) na área experimental do
Jardim, Planaltina, Distrito Federal.
Quanto à síndrome de dispersão foram registradas no total 77% de morfoespécies
não anemocóricas e 23% anemocóricas. A APR apresentou 81 morfoespécies
pertencentes ao modo de dispersão não anemocórica e 29 anemocóricas, apesar desta
maior riqueza de espécies não anemocóricas a maior abundância foi representada pelos
diásporos dispersos pelo vento (Tabela 1.1). Na MR foram registradas, respectivamente,
79 e 28 morfoespécies não anemocóricas e anemocóricas,. Na APR os picos de aporte
(riqueza) de diásporos anemocóricos e não anemocóricos ocorreram, respectivamente,
nos meses de agosto e maio. Na MR o pico de aporte (riqueza) ocorreu no mês de
agosto tanto para diásporos anemocóricos como não anemocóricos (Figura 1.9A e
1.9B).

33
Ao se analisar os tratamentos, na área do rio Jardim, verificou-se que houve
diferenças significativas na abundância e na riqueza da chuva de sementes entre todos
os tratamentos (Figura 1.10 e 1.11). O tratamento 3 apresentou a maior abundância de
sementes (4936sem/m2) (p<0,001, GL:8) sendo que 59% foi representado pela espécie
invasora Urochloa decumbens. O tratamento que apresentou maior riqueza foi T4
(49spp), seguido pelo tratamento T2 (35spp), linha de recobrimento e diversidade
(p<0,001, GL: 8).
Tabela 1.1: Riqueza e abundância (total e porcentagem) dos modos de dispersão não anemocórica (NA) e
anemocórica (A) para a área em processo de restauração (APR) e mata remanescente (MR) na área
experimental Jardim, DF.
APR MR
NA A Total NA A Total
Riqueza 81 (73,6%) 29 (26,4%) 110 79 (73,8%) 28 (26,6%) 107
Abundância 15608 (16,2%) 80553 (83,8%) 96161 2020 (49,8%) 2034 (50,2%) 4054

34
Figura 1.9: Riqueza total de diásporos anemocóricos e não anemocóricos nos 12 meses de coleta na
área do Rio Jardim, Distrito Federal. A) Riqueza total de diásporos amostrados na área em processo de
restauração (APR). B) Riqueza total de diásporos amostrados na área de mata remanescente (MR).
A
B

35
Figura 1.10: Abundância de diásporos em cada tratamento (T1: Nucleação de Anderson, T2:
linha de recobrimento e diversidade, T3: Controle, T4: mata remanescente) analisado na área
do Rio Jardim, Distrito Federal, entre os meses de fevereiro de 2012 a janeiro de 2013
(p<0,001, Df: 8, distribuição: Poisson). Barras representam o erro-padrão. As letras
minúsculas diferentes, acima das colunas (tratamentos), significam que os tratamentos são
significativamente diferentes.

36
Figura 1.11: Riqueza de diásporos em cada tratamento (T1: Nucleação de Anderson, T2:
linha de recobrimento e diversidade, T3: Controle, T4: mata remanescente) analisado na
área do Rio Jardim, Distrito Federal, entre os meses de fevereiro de 2012 e janeiro de 2013.
(p<0,001; Df: 8 distribuição: poisson). Barras representam o erro-padrão. As letras
minúsculas diferentes, acima das colunas (tratamentos) , significam que os tratamentos são
significativamente diferentes.
Na avaliação da chuva de sementes como indicador ecológico a partir da analise
dos dados coletados e dos descritores verificou-se que quanto aos aspectos
sensibilidade, resultabilidade, compreensão e interpretação, previsibilidade, escala e
síntese o valor obtido foi 5 (mais eficiente) para cada um, para o aspecto custo o valor
foi 3. A soma final dos valores foi 33, a valor máximo possível 35.

37
4.5. Discussão
A densidade de sementes da área do rio Jardim foi alta (8.336,50 sem/m²),
principalmente pelo volume de sementes de braquiária proveniente da área em processo
de restauração (APR). No entanto, ao retirar-se a braquiária e considerar-se apenas a
APR a densidade de diásporos continua alta (3.518 sem/m²) quando comparada a outros
estudos realizados no Brasil. Penhalber & Mantovani (1996) registraram 1.804, 2
sem/m² em mata secundária na Mata Atlântica. Araújo (2002) em estudo realizado em
floresta ombrófila densa obteve 7.456,1 sem/m²; Siqueira (2002) relatou 591,3 sem/m²
em áreas restauradas na Mata Atlântica e. Lagos & Marimon (2012) registraram 1.164
sem/m² em Cerrado. A alta quantidade de diásporos de braquiária coletados na área
Jardim representa o alto poder regenerativo desta espécie, sua alta capacidade
reprodutiva e de dispersão, o que, segundo Pivello (2011) pode prejudicar o
recrutamento e estabelecimento das espécies nativas. Por outro lado, em função da
densidade de diásporos permanecer alta, mesmo ao retirar-se a braquiária, mostra que a
área do Jardim tem um alto potencial de regeneração natural em função da
complexidade da paisagem.
A paisagem do entorno da área a ser restaurada tem influencia direta no aporte
de chuva de sementes. A área do Jardim encontra-se inserida numa matriz agrícola
(pastagem, soja e fruteiras) com poucas áreas de vegetação nativa. Este panorama leva
em conta um aspecto mais amplo da paisagem, porém adjacentes à área experimental
ocorrem áreas nativas. A proximidade destas áreas nativas intensifica o efeito de borda e
as interações positivas, que facilitam o aporte de chuva de sementes e aumenta o
potencial de regeneração natural da área em processo de restauração (APR). A chuva de
sementes tem importante papel na regeneração natural de áreas em processo de
restauração, sendo responsável pela base da dinâmica florestal que é a chegada de
sementes, autóctones e alóctones, que serão responsáveis pelo enriquecimento da área
degradada (SIQUEIRA, 2002; COLE et al, 2010). Para que o processo de regeneração e
cicatrização de áreas abertas ocorra é necessário que haja, nas proximidades das áreas a
serem restauradas, fragmentos ou áreas florestais íntegras que servirão de fonte de
propágulos (JORDANO, 2006).
A presença de fragmentos próximos a áreas degradadas facilita, mas não
necessariamente implica na chegada e recrutamento de propágulos. Em áreas isoladas o
aporte de chuva de sementes é menor (HOWE, 1993, JORDANO, 2006). Para que esse

38
grau de isolamento seja reduzido são necessários elementos na paisagem, como árvores
isoladas por exemplo, que facilitem o fluxo de propágulos entre a matriz florestal e a
área aberta, sendo esta comunicação facilitada pelos agentes dispersores.
A presença de fontes de propágulos na paisagem do Jardim como os fragmentos
de mata ripária e vereda podem ter facilitado o aporte da chuva de sementes, sendo que
árvores e arbustos isolados podem ter exercido a conexão entre a vegetação circundante
e a APR. Árvores isoladas em pastos abandonados podem funcionar como facilitadoras
do processo de recrutamento atuando como pontos de nucleação em áreas degradadas
(GUEVARA & LABORDE, 1993; PIZO & DOS SANTOS, 2011) podendo
desempenhar a função de ‘trampolim’ facilitando a movimentação de animais através da
matriz (FISCHER & LINDENMAYER, 2002). Bocchese et al (2008) em estudo no
Mato Grosso do Sul, em área de pastagem dominada por espécies invasoras, concluiu
que árvores isoladas e poleiros artificiais contribuíram para o aporte de sementes
permitindo uma diversidade local de dispersores.
A chuva de sementes apresentou variação de aporte entre os meses. A variação
anual na produção de frutos e sementes influencia na dinâmica e no recrutamento das
populações representando um componente importante a ser entendido para estimar o
potencial de regeneração de determinada comunidade vegetal (PENHALBER &
MANTOVANI, 1997). Nas florestas tropicais as espécies florescem e frutificam o ano
todo (FREIRE, 2006). Porém, os picos de produção variam de acordo com o local, a
comunidade vegetal estudada e com as condições ambientais (MORELLATO et al,
1989; ARRIAGA & MERCADO, 2004). Neste estudo a variação de aporte representa
que a oferta de recursos para a fauna dispersora de sementes está bem distribuída ao
longo do ano, indicando que a fauna dispersora estará desempenhando seus serviços
ecológicos auxiliando assim no enriquecimento da APR, bem como da MR.
O pico de deposição de sementes, desconsiderando-se a braquiária da análise, foi
nos meses de maio e junho. Nestes meses 49% dos diásporos foram representados pela
espécie Cenchrus polystachios. Esta espécie pertence à família Poaceae e é
anemocórica, por isso foi mais amostrada nos meses de seca que é a época mais
favorável para este tipo de dispersão (MORELLATO & LEITÃO-FILHO, 1996).
A chuva de sementes do Jardim foi representada, em sua grande maioria, por
diásporos menores que 2 mm. Essa alta proporção de diásporos pequenos deveu-se a
malha (<1 mm) utilizada nos coletores, a acurácia no processo de triagem fina das
amostras e ao estagio inicial de sucessão que se encontra a área. Espécies colonizadoras

39
de estágios iniciais de sucessão possuem sementes pequenas que são produzidas em
grandes quantidades para facilitar o alcance de locais adequados para posterior
estabelecimento (SALISBURY, 1974; PINÃ-RODRIGUES et al, 1990).
Diásporos não anemocóricos representaram alta riqueza na APR. Esta riqueza,
provavelmente, está relacionada à variedade de ambientes que circunda a área. Tal fato
contribui para o incremento do aporte da chuva de sementes, principalmente com
relação às espécies zoocóricas e autocóricas que compuseram o modo de dispersão
denominado “não anemocórica”. A paisagem circundante a área em processo de
restauração influencia diretamente na quantidade e qualidade da chuva de sementes
(JORDANO, 2006; PIVELLO et al, 2006). O contexto de paisagem está incluso na
análise de indicador no aspecto síntese (SEGIP 1995, METZGER et al, 2002), sendo,
portanto, importante levantar o entendimento da chuva de sementes neste contexto.
Através das síndromes de dispersão é possível a avaliação do estado de
conservação do meio (PIÑA-RODRIGUES, 1994). A medida que a perturbação cresce,
e a área é levada a um panorama de degradação mais intenso, o número de espécies
zoocóricas tende a diminuir e a anemocoria a aumentar, portanto em áreas perturbadas é
de se esperar uma proximidade do número de espécies zoocóricas e anemocóricas
(SIQUEIRA, 2002). A mata remanescente (MR) do rio Jardim apresenta abundância
equilibrada entre diásporos anemocóricos e não anemocóricos, sendo portanto uma área
remanescente com certo grau de perturbação, tendo espécies de estádio sucessional mais
tardio que a área aberta.
Na APR a riqueza de diásporos não anemocóricos foi semelhante à amostrada na
MR, porém a maior discrepância ocorreu em relação à abundância. Na APR a maior
abundância corresponde a diásporos anemocóricos. A abundância de diásporos
anemocóricos é maior em áreas abertas principalmente áreas com presença de
gramíneas invasoras (PENHALBER & MANTOVANI, 1997; PIVELLO, 2011).
Como o plantio na APR foi instalado em 2011, as espécies plantadas ainda não
compuseram a chuva de sementes da área. Este trabalho representa o “tempo 0” do
processo de restauração, podendo servir de comparação para os próximos anos de
estudos na área e de base para o acompanhamento da chuva de sementes em área sob
processo de restauração.
O tratamento T3 (controle) apresentou a maior abundância de sementes. Este
aporte foi devido à ausência de manejo, à presença da braquiária e ao maior número de
árvoretas remanescentes isoladas, representadas no total das três réplicas por 15

40
indivíduos com tamanho médio de 3,30m, bem como pelo visível aumento do número
de regenerantes de espécies arbustivas. A presença de arbustos e árvores isoladas na
área em processo de restauração é considerada facilitadora de colonização e
recrutamento de espécies florestais (DOS SANTOS et al, 2011) Estudo de Nave et al
(2005) constatou que pasto formado por braquiária com regeneração arbustiva ou
arbórea apresentaram maior média de germinação de banco de sementes quando
comparada a pasto sem regeneração natural. A composição do banco de sementes esta
diretamente ligada à chuva de sementes autóctones e alóctones, portanto a maior média
encontrada no estudo de Nave (2005) reflete o maior aporte de chuva de sementes em
áreas com o contexto de presença de regenerantes (arbustivos e arbóreos).
O tratamento T2 (linha de recobrimento e diversidade) apresentou o maior
aporte de chuva de sementes (riqueza), indicando que este pode estar cumprindo melhor
o papel de atração de dispersores e mantendo uma conexão com a paisagem. Porém,
provavelmente, pelo pouco tempo da implantação dos experimentos não foi possível
afirmar tal questão. As parcelas de T2 estão de forma aleatória na área, duas localizadas
próximas à mata remanescente e uma próxima a área de vereda degradada, sendo
observado o mesmo padrão para as parcelas T1. Nas parcelas de T2 foram mapeados
menos indivíduos arbóreos isolados (árvores remanescentes) quando comparadas ao
tratamento T3. Os tratamentos diferem quanto a forma de plantio e manejo, porém por
estes aspectos e pelas observações de localização das parcelas não é possível afirmar,
ou mesmo supor, o porquê da maior riqueza aportada no T2, sendo necessário maior
tempo de estudo para acompanhar o crescimento das mudas, a cobertura da área, o
manejo e assim entender melhor o que esta influenciando o aporte de chuva de sementes
no tratamento em questão. O diferente manejo de áreas a serem restauradas, ou em
processo de restauração, pode incitar comportamentos diferentes em espécies e seus
respectivos grupos (BARBOSA, 2004; NAVE et al, 2005).
A chuva de sementes como indicador ecológico agrega informações mensuráveis
de maneira que sua presença, e a qualidade, possibilitam afirmar sobre as possíveis
trajetórias da restauração. Um indicador ecológico deve ter três funções básicas:
permitir o conhecimento da situação ecológica de determinado local, levando-se em
consideração sua evolução espaço-temporal; servir de comparação e criar padrões;
subsidiar a tomada de decisões futuras e a elaboração de estratégias para o manejo da
área (CLAUDE & PIZARRO, 1996). Através dos dados apresentados neste estudo foi
possível determinar a composição da chuva de sementes e entender seus componentes

41
(tamanho de sementes e síndromes de dispersão), determinando assim o processo como
um todo. Tendo em vista esse aspecto mais sistêmico da chuva de sementes, esta
representa um indicador ecológico estando contida no grupo de indicadores com
características de “funcionamento” (Durigan, 2011).
A chuva de sementes apresentou sensibilidade, pois detectou diferenças entre
ambientes distintos, APR e MR; através dos modos de dispersão, tamanho das
sementes, índice de equabilidade foi possível afirmar e discutir sobre o grau de
conservação da área. Os dados coletados foram mensuráveis (quantitativamente e
qualitativamente) ao longo de um ano de avaliação, gerando, com isso, resultabilidade.
Observando-se os resultados é possível afirmar que a área Jardim tem um potencial de
regeneração natural, sendo possível, e mantendo afastados os fatores de degradação,
prever o futuro da área. A metodologia de coleta e a triagem são possíveis de ser
replicadas em diferentes áreas e pode-se chegar a uma conclusão clara para nortear
tomadas de decisões futuras sobre a restauração através dos resultados da chuva de
sementes.
Com base nos aspectos e descritores sobre indicadores ecológicos (SEGIP, 1995;
METZGER et al, 2002) e levando-se em conta os resultados obtidos com este estudo, a
nota atribuída a chuva de sementes foi 33. Partindo-se do pressuposto de que a nota
máxima seria 35 a chuva de sementes apresentou-se acima da média, sendo, portanto,
um bom indicador ecológico. O único aspecto negativo para a chuva de sementes foi
apresentar um custo maior, em função dos gastos com a confecção dos coletores, equipe
grande para coleta de material em campo e triagem, bem como de pessoas capacitadas
para a identificação das sementes. Porém estes custos não inviabilizam o uso da chuva
de sementes como indicador ecológico.
Com base nos resultados deste estudo pode-se concluir que a chuva de sementes
é um indicador ecológico eficiente, mesmo tendo um alto custo de implantação e
manutenção, pois expressa de maneira fidedigna as diferenças locais no aporte de
sementes, ou seja, nas chances diferenciais de regeneração de uma determinada área.
Além disso, a chuva de sementes reflete a presença dos processos ecológicos envolvidos
permitindo, consequentemente, o entendimento da dinâmica florestal.

42
4.6. Referências Bibliográficas ALBUQUERQUE, L.B.; AQUINO, F.G.; ALONSO, A.M.; LIMA, J. E. W.; BRAGA; SOUSA-SILVA,
J.C. Restauração Ecológica de Matas Ripárias: uma questão de sustentabilidade. Série Documentos.
Brasília: Embrapa Cerrados, p.75. 2010.
AQUINO, F.G.; VILELA, M.F. Importância das matas ripárias. 2008. Artigo em Hypertexto.
Disponível em: http://www.infobibos.com/Artigos/2008_4/matas/index.htm. Acesso em: 17 jan.2014
ARAÚJO, R.S de. Chuva de sementes e deposição de serapilheira em três sistemas de revegetação de
áreas degradadas na Reserva Biológica de Poço das Antas (RJ). 2002. 92f. Dissertação (Mestrado em
Ciências Ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro, Seropédica, 2002.
ARRIAGA, L., & MERCADO, C. Seed bank dynamics and tree‐fall gaps in a northwestern Mexican
Quercus‐Pinus forest. Journal of Vegetation Science, v. 15, n. 5, p. 661-668, 2004.
ATTANASIO, C.M.; GANDOLFI, S.; ZAKIA, M.J.B; VENIZIANI, J.C.T.J.; LIMA, W.P. A
importância das áreas ripárias para a sustentabilidade hidrológica do uso da terra em microbacias
hidrográficas. Bragantia, Campinas, v. 71, n. 4, p. 493-501, 2012.
BARBOSA, L.M. Considerações gerais e modelos de recuperação de formações ciliares. In:
RODRIGUES, R.R.; LEITÃO-FILHO,H.F. (Ed.). Matas Ciliares: Conservação e recuperação. 3
ed. São Paulo: EDUSP, 2004, v. 1, p. 235-247, 2004.
BOCCHESE, R.A.; OLIVEIRA, A.C.M.; FEVERO, S.; GARNÉS, S.J.S & LAURA, V.A. Chuva de
sementes e estabelecimento de plântulas a partir da utilização de árvores isoladas e poleiros artificiais
por aves dispersoras de sementes, em área de Cerra, Mato Grosso do Sul. Revista Brasileira de
Ornitologia, v. 16, n. 3, p. 207-213, 2008.
BROWER, J.E.; ZAR, J.H.; & VON ENDE, C.N. Field and laboratory methods for general ecology. Wm.
C. C. Brown, Dubuque, Iowa, 1984, 161p.
CAUGHLAN, L.; & OAKLEY, K.L. Cost considerations for long-term ecological
monitoring. Ecological indicators, v. 1, n. 2, p. 123-134, 2001.
COLE, R. J.; HOLL, K. D.; & ZAHAWI, R. A. Seed rain under tree islands planted to restore degraded
lands in a tropical agricultural landscape. Ecological Applications, v. 20, n. 5, p.1255-1269, 2010.
CRAWLEY, M. J. The R book. John Wiley & Sons Inc, p. 942, 2007.
DARONCO, C.; MELO, A.C.G.D.; & DURIGAN, G. Restored versus reference ecosystem: case study of
plant community at a riparian forest in the Cerrado region, Assis, São Paulo State, Brazil. Hoehnea, v.
40, n. 3, p. 485-498, 2013.
DOS SANTOS, M.M.G.; OLIVEIRA, J.M.; MÜLLER, S. C.; & DE PATTA PILLAR, V. Chuva de
sementes de espécies lenhosas florestais em mosaicos de floresta com Araucária e campos no Sul do
Brasil. Acta Botanica Brasilica, v. 25, n. 1, p. 160-167, 2011.
DURIGAN, G. O uso de indicadores para monitoramento de áreas em recuperação. Cadernos da Mata
Ciliar, v. 4, p. 11-13, 2011.
FELFILI, J. M.; RIBEIRO, J. F.; FAGG, C. W.; MACHADO, J.W.B. Recuperação de matas de galeria.
Embrapa Cerrados. n. 21, p. 1-45, 2000.
FISCHER,J.; LINDENMAYER,D.B. The conservation value of paddock trees for birds in a variegated
landscape in southern New South Wales. 2. Paddock trees as stepping stones. Biodiversity and
Conservation, v. 11, p. 833-849, 2002.
FREIRE, M. Chuva de sementes, banco de sementes do solo e deposição de serapilheira como
bioindicadores ambientais no bioma Mata Atlântica, Teresópolis, RJ. 2006. 81 f. Dissertação
(Mestrado em Recursos Ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro,
Seropédica. 2006.
GUEVARA, S.; & LABORDE, J. Monitoring seed dispersal at isolated standing trees in tropical pastures:
consequences for local species availability. In: Fleming, T. H.; & Estrada, A. Frugivory and seed
dispersal: ecological and evolutionary aspects . Springer Netherlands. 1993, p. 319-338.
HERRERA, J.M.; GARCIA, D. The role of remnant trees in seed dispersal through the matrix: being
alone is not always so sad. Biological Conservation, v. 1452, p. 149-158, 2009.
HOLL, K.D. Do bird perching structures elevate seed rain and seedling establishment in abandoned
tropical pasture?. Restoration Ecology, v. 6, p. 253-261, 1998.
HOWE, H.F.; SMALLWOOD, J. Ecology of seed dispersal. Annual Review of Ecology and
Systematics, Palo Alto, CA, v. 13, p. 201-228, 1982.
HOWE, H.F. Aspects of variation in a neotropical seed dispersal system. Vegetatio, v. 107, n. 108, p.
149-162, 1993.
JORDANO, P.; GALETTI, M.; PIZO, M.A.; & SILVA, W.R. Ligando frugivoria e dispersão de sementes
à biologia da conservação. In: Duarte, C.F.; Bergallo, H.G.; Dos Santos, M.A.; and V a, A.E. (eds.).
Biologia da conservação: essências. Editorial Rima, São Paulo, Brasil. 2006, p. 1-436.

43
LAGOS, M.D.C.C.; & MARIMON, B.S. Chuva de sementes em uma floresta de galeria no Parque do
Bacaba, em Nova Xavantina, Mato Grosso, Brasil. Revista Árvore, n. 36, v. 2, p. 311-320, 2012.
MAGNAGO, L.F.S.; MARTINS, S.V.; VENZKE, T.S. & IVANAUSKAS, N.M. Os processos e estágios
sucessionais da Mata Atlântica como referência para restauração florestal. In: S.V. Martins (org.).
Restauração ecológica de ecossistemas degradados. Editora UFV, Viçosa, p. 69-100, 2012.
METZGER, J.P.; CASATTI, L.; VERDADE, L. Indicadores de conservação e avaliação do
conhecimento para conservação. In: I Workshop de Síntese do Programa BIOTA/FAPESP na
Universidade de São Carlos, SP, 2002. Relatório do grupo de trabalho. Disponível em:
<http://www.biota.org.br/info/historico/simp2002/gt2.pdf>. Acesso em: 21 set. 2013.
MCGEOCH, M.A.; & CHOWN, S. L. Scaling up the value of bioindicators. Trends in Ecology &
Evolution, v. 13, n. 2, p. 46-47, 1998.
MORELLATO, L.P.C.; RODRIGUES, R.R.; LEITÃO-FILHO, H.F.; JOLY, C.A. Estudo fenológico
comparativo de espécies arbóreas de florestas de altitude e floresta mesófila semidecídua na Serra do
Japi, Jundiaí, SP. Revista Brasileira de Botância, n. 12, p. 85-98, 1989.
MORELLATO, L.P.C.; LEITÃO-FILHO, H.F. Reproductive phenology of climbers in a South-easthern
Brazilian Forest. Biotropica, n. 28, p. 180-191, 1996.
NAVE, A. G. Banco de sementes autóctone e alóctone, resgate de plantas e plantio de vegetação nativa na
Fazenda Intermontes, município de Ribeirão Grande, SP. 2005. 218 f. Tese (Doutorado) - Escola
Superior de Agricultura Luiz de Queiroz. Universidade de São Paulo. Piracicaba, 2005.
PENHALBER, E.D.F.; & MANTOVANI, W. Floração e chuva de sementes em mata secundária em São
Paulo, SP. Brazilian Journal of Botany, v. 20, n. 2, p. 205-220, 1997.
PIÑA-RODRIGUES, F.C.M.; COSTA, L.G.S. & REIS, A. Estratégias de estabelecimento de espécies
arbóreas e o manejo de florestas tropicais. In: CONGRESSO FLORESTAL BRASILEIRO, 6,
Anais... Campos do Jordão, SP, v. 3, 1990, p. 676-684.
PIÑA-RODRIGUES, F.C.M. Aplicação de conceitos ecológicos para o diagnóstico e recuperação de
áreas naturais. Floresta Ambiente, v. 1, p. 49-58, 1994.
PIVELLO, V.R.; PETENON, D.; JESUS, F.M.D.; MEIRELLES, S.T.; VIDAL, M.M.; ALONSO, R.D.
A. S.; FRANCO, G.A.D.C. & METZGER, J.P. Chuva de sementes em fragmentos de Floresta
Atlântica (São Paulo, SP, Brasil), sob diferentes situações de conectividade, estrutura florestal e
proximidade da borda. Acta Botanica Brasilica, v. 20, n. 4, p. 845-859. 2006.
PIVELLO, V. R. Invasões biológicas no cerrado brasileiro: efeitos da introdução de espécies exóticas
sobre a biodiversidade. Ecologia. info, 33. 2011. Disponível em:
http://www.ecologia.info/cerrado.htm. Acesso em: 23 dez. 2013.
PIZO, M.A.; DOS SANTOS, B.T.P. Frugivory, post‐feeding flights of frugivorous birds and the
movement of seeds in a brazilian fragmented landscape. Biotropica, v. 43, n. 3, p. 335-342, 2011.
REATTO, A.; CORREIA, J.R.; SPERA, S.T.; CHAGAS, C.S.; MARTINS, E.S.; ANDAHUR, J.P.; &
ASSAD, M.L.C.L. Levantamento semi-detalhado dos solos da Bacia do Rio Jardim-DF, escala 1:
50.000. Planaltina: Embrapa Cerrados, n. 18, p. 1-63, 2000.
REIS, A.; KAGEYAMA, P.Y. Restauração de áreas degradadas utilizando interações interespecíficas. In:
Kageyama, P.Y.; Oliveira, R.E.; Moraes, L.F.D.; Engel, V.L & Gandara, F.B. Restauração ecológica
de ecossistemas naturais. Botucatu, FEPAF, p. 91-110. 2003.
RODRIGUES, R.R.; GANDOLFI, S. Restauração de florestas tropicais: subsídios para uma definição
metodológica e indicadores de avaliação e monitoramento. In: DIAS, L.E.; MELLO, J.W.V. (Ed.).
Recuperação de áreas degradadas. Viçosa: UFV; SOBRADE,1998. p. 203-215.
RODRIGUES, R.R. Uma discussão nomenclatural das formações ciliares. Matas ciliares: conservação e
recuperação, n. 2, p. 91-99, 2000.
RIBEIRO, J. F.; WALTER, B.M.T.; SANO, S.M.; & ALMEIDA, S.D. (1998). Fitofisionomias do bioma
Cerrado. Cerrado: ambiente e flora, p. 89-166.
RUDGE, A. D. C. Contribuição da chuva de sementes na recuperação de áreas e do uso de poleiros como
técnica catalisadora da sucessão natural. 2008. 105f. Dissertação (Mestrado em Ciencias ambientais e
florestais) - Universidade Rural do Rio de Janeiro. 2008.
SALISBURY E. Seed size and mass in relation to environment. Proceedings of the Royal Society,
London, Series B. n. 186, p. 83-88, 1974.
SEGIP. The state enviromental Goals and Indicators Project. Evaluation criteria. In: BERGQUIST, G. et
al. Prospective indicators for state use in performance agreements. Florida: SEGIP/Florida Center
for public Management, 1995. p. B6-B7.
SILVA JÚNIOR, M.D.; FELFILI, J.M.; WALTER, B.M.T.; NOGUEIRA, P.E.; REZENDE, A.V.;
MORAIS, R.D.O.; NÓBREGA, M.G.G.; SOUZA-SILVA, J.C. Análise da flora arbórea de Matas de
Galeria no Distrito Federal: 21 levantamentos. In: Ribeiro, F.R.; Fonseca, C.E.L., Sousa-Silva, J. C.

44
Cerrado: caracterização e recuperação de Matas de Galeria. Brasília: EMBRAPA, 2001, p. 143-
208.
SIQUEIRA, L.P.D. Monitoramento de áreas restauradas no interior do Estado de São Paulo, Brasil.
2002. 116f. Dissertação (Mestrado em Recursos florestais com opção em conservação de recursos
florestais) – Escola Superior de agricultura Luiz de Queiroz. Universidade de São Paulo, Piracicaba,
2002.
SLOCUM, M.G.; & HORVITZ, C.C. Seed arrival under different genera of trees in a neotropical
pasture. Plant Ecology, v. 149, n. 1, p. 51-62, 2000.
SPERA, S. T.; REATTO, A.; MARTINS, E. D. S.; CORREIA, J. R.; BLOISE, G. L. F.; & SILVA, A. D.
Aptidao agricola das terras da Bacia do Rio Jardim, DF. Embrapa Cerrados. 2002.
SWAINE, M. D.; & WHITMORE, T. C. On the definition of ecological species groups in tropical rain
forests. Vegetatio, v. 75, n. 1-2, p. 81-86, 1988.
TRES, D. R.; ANNA. C.S.S.; BASSOS, S.; LANGA, R.; JUNIOR, U.R.; REIS, A. Banco e Chuva de
Sementes como Indicadores para a Restauração Ecológica de Matas Ciliares. Revista Brasileira de
Biociências, Porto Alegre, v. 5, n. 1, p. 309-311, jul, 2007.

45
5. Capítulo 2 – A CHUVA DE SEMENTES PODE CONTRIBUIR PARA A
RESTAURAÇÃO ECOLÓGICA DE MATA RIPÁRIA?
Resumo
A chuva de sementes é o processo pela qual ocorre a chegada de diásporos a uma área
sendo dependente dos agentes de dispersão, dos elementos da paisagem, da distância
dos fragmentos, entre outros. Entender o funcionamento da chuva de sementes em
matas ripária é de suma importância para subsidiar políticas públicas e indicar melhores
metodologias de restauração. O objetivo deste trabalho foi analisar a chuva de sementes
como indicador de restauração ecológica em trecho de mata ripária do rio Capão
Comprido, DF-Brasil. Para tal foi analisado quali-quantitativamente a chuva de
sementes nas áreas remanescentes (MR) e nas áreas em processo de restauração (APR)
localizadas às margens do córrego Capão Comprido (Distrito Federal - Brasil). A área a
ser restaurada (2,6ha) foi utilizada durante 30 anos em atividades de pecuária tendo a
atividade sido encerrada em 2008. A área pertence a uma paisagem basicamente
agrícola com poucos remanescentes de vegetação nativa nas adjacências. Foram
instalados 84 coletores no total, com 0,25 m² e malha de 1 mm² a 15 cm do solo, sendo
12 instalados em cada tratamento (T1: Nucleação Anderson 3x3m, T2: Nucleação
Anderson 5x5m, T3: Poleiro, T4: Nucleação Anderson + Poleiro, T5: Linha de
recobrimento e diversidade, T6: Controle, T7: Mata remanescente). O material
depositado nos coletores foi recolhido mensalmente, de março de 2012 a fevereiro de
2013, em sacos de papel e triado com auxilio de estereomicroscópio no Laboratório de
Biologia Vegetal da Embrapa Cerrados. Do material foram separadas as sementes,
sendo estas quantificadas e identificadas em morfo-espécie. Nos 12 meses avaliados
foram coletados 74.316 propágulos no total, sendo 5.718 (272,28 sementes/m2) na mata
e 68.598 (3.266 sementes/m2) na área em restauração. As famílias mais abundantes na
APR e MR foram, respectivamente, Poaceae e Asteraceae. A espécie braquiária
(Uruchloa decumbens) foi a mais abundante na área em restauração (87%). Ocorreu
variação no aporte de chuva de sementes entre os meses, mas o pico foi no mês de
fevereiro. Quanto à síndrome de dispersão foram registradas, no total, 71% de
morfoespécies não anemocóricas e 29% anemocóricas. O tratamento que apresentou
maior aporte (abundância) de diásporos foi o T5 (linha de recobrimento e diversidade)
(p<0,001, GL:14), em relação a riqueza o tratamento que apresentou maior aporte foi o
T7 (mata remanescente) não havendo diferença estatística entre os demais tratamentos.
Através dos dados é possível avaliar diferenças quanto à abundância, riqueza, síndrome
de dispersão entre as áreas APR e MR. A partir do estudo é possível observar que a
chuva de sementes reflete mesmo as menores diferenças apresentando aspectos
relevantes a um indicador ecológico como, sensibilidade, resultabilidade,
previsibilidade, síntese, , Representando fidedignamente as condições do meio e
permitindo concluir que a paisagem influencia no aporte da chuva de sementes. Por
estes motivos a chuva de sementes pode ser utilizada como indicador de restauração
ecológica. Este trabalho propõe o uso da chuva de sementes como dois parâmetros:
indicador do potencial de restauração e indicador de automanutenção do sistema.

46
Abstract
The riparian forests are woods accompanying waterways. The protection and
restoration of these are relevant to the maintenance of biodiversity and hydrological
cycles. Seed rain is the process in which the arrival of propagules happens to an area,
depending on scattering factors, elements of landscape fragmentation, among others .
Understanding the workings of the seed rain in riparian forests is of paramount
importance to support public policies and to indicate better restoration methodologies.
The objective of this study was to analyze the seed rain as an indicator of ecological
restoration in riparian stretch of the river Capão Comprido, DF , Brazil . For such was
analyzed qualitative and quantitatively the seed rain in the remaining areas and in areas
where restoration located on the banks of the stream Capão Comprido (Distrito Federal
- Brazil) . The area to be restored (2.6 ha) was used for 30 years in the cattle-raising
activity having been closed in 2008. The area belongs to a primarily agricultural
landscape with few remaining native vegetation in the vicinity . 84 collectors were
installed overall, with 0.25 m² and mesh size of 1 mm ² to 15 cm of soil , with 12
installed in each treatment ( T1 to T7 ). The material deposited in the traps was
collected monthly from March 2012 to February 2013, in paper bags and screened with
the aid of a stereomicroscope at the Laboratory of Plant Biology (EMBRAPA Cerrados)
and Laboratory of Conservation Biology (UNIMONTES). The material seeds were
separated, being quantified and identified to the level of morpho- species. In the 12
months evaluated 74.316 seedlings were collected in total, with 5.718 (272.28 seeds/m2)
in the forest and 68.598 (3266 seeds/m2) the restoration site . The most abundant
families in the APR and MR were, respectively, Poaceae and Asteraceae. The exotic
species braquiária (Uruchloa decumbens) was the most abundant in the restoration site
(87 %) . Variation occurred in the supply of seed rain between the months, with the
peak in February. Regarding dispersion syndrome, it was recorded in total, 71 % of non
anemochoric morphospecies and 29 % anemochoric. The treatment that showed higher
uptake (abundance) of diaspore was the T5 (line coverage and diversity) (p < 0.001, Df:
14), the relative wealth treatment showed the greatest contribution was the T7
(remaining forest) with no significant differences between the other treatments. From
this data it is possible to assess differences in the abundance, richness , dispersal
syndrome between APR and MR areas . From the study it is observed that the seed rain
reflect even the smallest differences presenting relevant aspects as an ecological
indicator, sensitivity, resultabilidade, predictability, synthesis, faithfully. Representing
the environment conditions and allowing to conclude that the landscape influences the
contribution seed rain. For these reasons the seed rain can be used as an indicator of
ecological restoration. This work proposes the use of seed rain as two parameters: an
indicator of the potential for restoration and display of self-maintenance system.

47
5.1. Introdução
A restauração ecológica baseia-se em iniciar ou acelerar o processo de sucessão
em ambientes degradados (SER, 2004). A restauração contemporânea visa recuperar
ambientes através da restauração de processos ecológicos que, em equilíbrio dinâmico,
vão manter as funções do ecossistema, possibilitando a manutenção da biodiversidade e
de serviços ecossistêmicos (BRANCALION et al, 2013).
A restauração tem o objetivo principal de fornecer o “gatilho” para iniciar a
sucessão ecológica em áreas degradadas. Para entender o processo de restauração são
necessárias ferramentas de monitoramento e avaliação. Os indicadores ecológicos são
ferramentas que vem ao encontro desta necessidade, estes, portanto, são parâmetros
mensuráveis e de fácil interpretação que permitem unir o monitoramento e o futuro
manejo de áreas em processo de restauração (SEGIP, 1995; CLAUDE & PIZARRO,
1996; DALE & BEYELER, 2001; METZGER, 2002).
A chuva de sementes é o processo pelo qual ocorre a entrada de propágulos, no
sistema (MARTINS, 2007), sendo dependente dos agentes dispersores (animais, vento),
fragmentos fontes, distância e qualidade destas fontes (PIVELLO et al, 2006;
JORDANO, 2006). Os diásporos ao alcançarem o solo pode abastecer o banco de
sementes que constituirá as plântulas e os indivíduos adultos da comunidade vegetal
futura (HUBBELL et al, 1986).
Na comunidade vegetal os mecanismos envolvidos nos propágulos podem
favorecer seu estabelecimento e sobrevivência (LABOURIAU, 1973; DARONCO et al,
2013). Características como síndrome de dispersão, tamanho, qualidade, refletem a
estrutura da vegetação, estádio sucessional e grau de conservação (HARPER, 1977;
GENTRY, 1982; PINHEIRO & RIBEIRO, 2001). A zoocoria é a síndrome de dispersão
predominante em fitofisionomias florestais (PINHEIRO & RIBEIRO, 2001;
STEFANELLO et a1, 2010), um dos fatores que contribui para este fato é a presença de
dossel continuo, que diminui a incidência luminosa fazendo com que o crescimento
inicial do indivíduo seja dependente, em parte, de fontes do material de reserva das
sementes (PIJL, 1982). Propágulos zoocóricos apresentam maior quantidadede material
de reserva que é utilizado pela fauna dispersora como fonte de alimento (MORELLATO
& LEITÃO-FILHO, 1992).
A movimentação de fauna dispersora é dificultada em paisagens com a matriz
dominada por áreas abertas, destinadas a atividades antrópicas (SANT’ANNA et al,

48
2011). A fragmentação e perda de habitats é um desses fatores que limitam a dispersão
tendo implicações diretas na biodiversidade (FAHRIG, 2003). No entanto, elementos
como árvores isoladas remanescentes em áreas de pastos abandonados e áreas agrícolas
podem facilitar e servir como trampolim na movimentação da fauna dispersora
(ALDRICH & HAMRICK, 1998; FISCHER & LINDENMAYER, 2002; PIZO &
SANTOS, 2011), potencializando, assim, o processo de cicatrização de áreas
degradadas.
A proteção e restauração das matas ripárias são salvaguardas por legislação
específica, porém continuam a ser degradadas. Esse processo de degradação, em uma
mesma área, pode fazer com que a situação de degradação piore pela redução da
capacidade de regeneração natural e fertilidade e compactação do solo devido a pisoteio
ou uso de máquinas agrícolas (RODRIGUES, 2013). As matas ripárias pelo fato de
ocorrerem como extensões dendríticas ao longo dos corpos de água são ecossistemas
que permitem uma conexão entre ambientes diversos, servindo como verdadeiros
corredores ecológicos (OLIVEIRA FILHO & RATTER, 1995; RODRIGUES, 2005).
Para avaliar áreas florestais muitas vezes são utilizados parâmetros estruturais
como classes de altura e área basal ao invés dos parâmetros funcionais. Tendo em vista
que o intuito de projetos de restauração é restituir os processos ecológicos é necessária
avaliação de parâmetros funcionais do ecossistema (ARAÚJO et al, 2002). Neste
contexto que entra em questão o uso de indicadores ecológicos, como por exemplo, a
chuva de sementes. Tendo em vista a importância da chuva de sementes para a
manutenção dos ecossistemas este trabalho visa caracterizar a chuva de sementes em
área em processo de restauração e área de mata remanescente com o objetivo de obter
informações para se avaliar a chuva de sementes como indicador de restauração
ecológica em zona ripária do Distrito Federal.

49
5.2. Material e métodos
5.2.1. Área de estudo
O experimento foi instalado à margem direita do córrego Capão Comprido,
Brazlândia-DF (15°44’32,79”S e 48°08’59,81”W), em trecho de mata ripária de 2,6 ha.
Para este trabalho a área foi denominada de “Coité”.
O córrego está inserido na Bacia Hidrográfica do Rio Descoberto que ocupa uma
área de 896 Km2
sendo 84% da área esta inserida no DF e 16% no Estado do Goiás
(CARMO et al, 2003). Os solos são do tipo latossolo vermelho-escuro, cambissolos e
latossolos vermelho-amarelo, sendo que os latossolos ocupam mais de 50% da Bacia
(EMBRAPA, 1978; REATTO et al, 2004). A cobertura vegetal compreende várias
fitofisionomias do Cerrado como campo limpo (33%), cerrado (20,5%), mata ciliar
(5%) e áreas de reflorestamento (1,5%) (RÊGO et al, 1997; FERNANDES et al, 2005).
A Sub-bacia do Capão Comprido apresenta área total de 15,89km2,
76% das
propriedades ocorrentes em sua área correspondem a chácaras (FERREIRA et al, 1992).
Adjacente à APR (área em processo de restauração) ocorre a mata de galeria
com faixa de 0 a 5m de largura. A mata remanescente (MR) está a oeste da APR a 238
m de distância, sendo caracterizada pela presença de espécies herbáceas, arbustivas e
arbóreas, possuindo área total de 3,6ha (Figura 2.1).
A APR é um ambiente heterogêneo com poucas espécies arbóreas e arbustivas,
sendo caracterizada pela presença abundante de espécies herbáceas, principalmente, a
gramínea invasora Urochloa decumbens (Stapf) R.D (Syn. braquiária) (Figura 2.2). Esta
área foi utilizada durante 30 anos como pastagem para bovinos, mas no ano de 2008
foram encerradas as atividades de pecuária na área.

50
Figura 2.1: Localização da área do Coité, Distrito Federal. A) Mata remanescente. B) Área em processo
de restauração as margens do córrego Capão Comprido, Brasil, Distrito Federal, Brazlândia. (Fonte:
Google Earth, imagem de: 10/12/2013. Acesso em: 10/02/2014).
Figura 2.2: Área em processo de restauração (APR) no Coité com estacas de
demarcação dos locais de plantio das mudas, Brazlândia, Distrito Federal
No Coité foram instaladas parcelas aleatórias em área degradada sob processo de
restauração (APR) com diferentes tratamentos, três repetições (parcelas), sendo que
cada tratamento representa uma técnica de restauração (ver metodologia geral), com
manejo diferenciado. Na MR foi instalado o tratamento de referência também com três
parcelas. A escolha dessa mata remanescente baseou-se na proximidade espacial com a
área em processo de restauração.

51
5.2.2. Delineamento experimental
Para a avaliação da chuva de sementes utilizou-se coletores de diásporos (frutos
e sementes) confeccionados com madeira e tela de nylon de 1mm2, cada coletor com
área de 0,25m2.
Na diagonal de cada parcela foram instalados quatro coletores a 15 cm
do solo, totalizando12 coletores/tratamento.
Na área em processo de restauração (APR) os coletores foram instalados na
diagonal das parcelas quadradas. Na mata remanescente (MR), em função de ter apenas
uma faixa estreita, os coletores foram instalados nas parcelas retangulares em linhas
paralelas ao leito do rio, porém com o mesmo esforço amostral. Para as análises, cada
parcela foi considerada uma unidade amostral, ou seja, o conjunto de quatro coletores,
representando um total de 1m2
de amostragem por parcela.
De março de 2012 a fevereiro de 2013 o material retido nos coletores foi
coletado mensalmente. Este material foi acondicionado e triado em laboratório. Quando
necessário o material foi seco a sombra à temperatura ambiente para minimizar os danos
à morfologia das sementes facilitando a sua triagem e identificação.
Na triagem foram separados os materiais grossos (p.ex. folhas, galhos) dos finos
(pó que passou pela peneira) utilizando peneira (malha 2 mm). O “pó” foi triado no
estereomicroscópio separando-se os diásporos, os quais foram morfoespeciados,
ilustrados, fotografados, medidos e classificados em modo de dispersão (não
anemocórica e anemocórica). Quando foram observadas sementes e/ou frutos maiores,
que não passaram pela malha da peneira, estes foram separados manualmente. Para a
identificação foram utilizados literatura especializada, consulta à carpoteca do Herbário
da Embrapa Cenargen (DF) exsicatas coletadas em estudo florístico da área, assim
como o auxilio de especialistas.
As sementes de braquiária (Urochloa decumbens) foram processadas no
aparelho Seed sorter para separação das cascas e sementes. Após este processamento, a
quantidade de sementes em cada amostra foi estimada por média de sementes em
proveta de 5 ml. Toda a triagem foi feita no Laboratório de Biologia vegetal da
Embrapa Cerrados, DF e no Laboratório de Biologia da conservação da Unimontes,
Montes Claros, MG
Para a avaliação da chuva de sementes como indicador ecológico foi analisada
com base na metodologia proposta ppor SEGIP (1995) e Metzger (2002), (ver item 2.3,
metodologia geral).

52
5.3. Análise dos dados
Os dados foram analisados construindo-se modelos lineares generalizados
(GLMs) utilizando-se o software R versão 3.0 (R Development Core Team, 2008). O
aporte da chuva de sementes, riqueza e abundância, foram comparados entre os
tratamentos de restauração ecológica, sendo as variáveis respostas riqueza e abundância
de diásporos e variável explicativa, os tratamentos. Todos os GLMs foram submetidos à
análise residual para se avaliar a adequação da distribuição dos erros (CRAWLEY,
2007). Para se analisar a diversidade da comunidade utilizou-se também o software R
para se calcular o índice Shanon-Weaver (H’) e o coeficiente de equidade de Pielou (J).
5.4. Resultados
A quantidade de diásporos coletados na chuva de sementes no período de um
ano no Coité foi no total de 74.316 diásporos, representado uma densidade de 3.538
sem/m², pertencentes a 107 morfoespécies. Destes, 14 foram identificados em espécie,
17 gênero, 18 família e 58 não identificados (Anexo 3). Na área estudada o índice de
diversidade Shannon-Weaver e de equidade de Pielou foram H’ = 0,95 e J = 0,20.
Na área em processo de restauração (APR) foram coletados no total 68.598
diásporos (3.811 sem/m2). A família mais abundante foi a Poaceae com dominância de
Urochloa decumbens (Stapf) R.D. Webster (Syn. braquiária), representando 87% dos
diásporos coletados. Na mata remanescente (MR) coletou-se 5.718 diásporos (1906
sem/m2), sendo a espécie mais abundante a sp125 (55%), pertencente à família
Asteraceae (Figura 2.3). Excluindo-se a braquiária do total foram amostrados 698
sem/m².
Figura 2.3: Foto da espécie mais abundante
na área de mata remanescente no Capão
comprido: morfoespécie sp125
(Asteraceae). (Aumento 1x).

53
Na caracterização anual houve pico de aporte da chuva de sementes no mês de
fevereiro, porém neste mês 77% do aporte foi em função da espécie Urochloa
decumbens. Porém ao retirar-se a braquiária, o pico ocorreu em setembro (Figura 2.4),
sendo que 92% do total de diásporos amostrados nesse mês foi representado pela
morfoespécie sp 125 pertencente a família Asteraceae.
Figura 2.4: Abundância de diásporos, considerando e desconsiderando a braquiária na coleta, amostrados
mensalmente na área do Córrego Capão comprido, Distrito Federal.
A composição da chuva de sementes foi de aproximadamente 1% de frutos (n=621)
e 99% de sementes (n=73.695). Os diásporos foram de tamanhos diversos variando
entre 1 mm e 6,5 cm, sendo 7% menores que 2 mm e os maiores que 1cm 0,7%. (Figura
2.5).

54
Figura 2.5: Quantidade total de diásporos amostrados em cada tamanho (mm) na área experimental do
Coite, Brazlândia, Distrito Federal.
Em relação à síndrome de dispersão foram caracterizadas 71% de morfoespécies
não anemocóricas e 29% anemocóricas. A APR apresentou 43 morfoespécies não
anemocórica e 24 anemocóricas; na MR foram 52 morfoespécies não anemocóricas e 21
anemocóricas (Tabela 21).
Tabela 2.1: Riqueza e abundância (total e porcentagem) dos grupos de dispersão não anemocórica (NA) e
anemocórica (A) para a área em processo de restauração (APR) e mata remanescente (MR) na área
experimental Capão Comprido, Distrito Federal.
APR MR
NA A Total NA A Total
Riqueza 43 (72%) 24 (38%) 60 52 (75%) 21 (30%) 69
Abundância 7632 (12%) 53268 (87%) 60900 1712 (30%) 3995 (70%) 5707
O tratamento que apresentou maior abundância de diásporos foi T5, linha de
recobrimento e diversidade, sendo que 79% foi de braquiária (Urochloa decumbens),

55
todos os tratamento apresentaram diferenças significativas (p<0,001, GL:14) (Figura
2.6). Avaliando-se a riqueza de espécies o tratamento que teve maior riqueza foi T7
(p<0,001, GL:14), mata remanescente, não havendo diferença significativa entre os
demais tratamentos (Figura 2.7).
Figura 2.6: Abundância de diásporos em cada tratamento analisado (T1: Nucleação
Anderson 3x3m, T2: Nucleação Anderson 5x5m, T3: Poleiro, T4: Nucleação Anderson +
Poleiro, T5: Linha de recobrimento e diversidade, T6: Controle, T7: Mata remanescente)
na área experimental Coité, Distrito Federal, entre março de 2012 e fevereiro de 2013.
(p<0,001; Df: 14, distribuição: poisson). Barras representam o erro-padrão. As colunas
(tratamentos) com a mesma letra não são significativamente diferentes.

56
Figura 2.7: Riqueza de diásporos em cada tratamento analisado (T1: Nucleação Anderson 3x3m,
T2: Nucleação Anderson 5x5m, T3: Poleiro, T4: Nucleação Anderson + Poleiro, T5: Linha de
recobrimento e diversidade, T6: Controle, T7: Mata remanescente) na área experimental Coité,
Distrito Federal, entre março de 2012 e fevereiro de 2013. (p<0,001; Df: 14, distribuição:
quasipoisson). Barras representam o erro-padrão. As colunas (tratamentos) com a mesma letra não
são significativamente diferentes.
Na avaliação de indicador ecológico seguindo os níveis da escala de Likert os
aspectos sensibilidade, resultabilidade, compreensão e interpretação, previsibilidade,
escala e síntese obteve nota cinco. Para o aspecto “custo” a nota atribuída foi dois
resultando assim num somatório de 32.

57
5.5. Discussão
O aporte de sementes total na área do Coité foi representado por uma alta
densidade (3.538 sem/m²) comparado aos trabalhos realizados em áreas degradadas e
em matas remanescentes, por exemplo Quintela (1996) registrou 3.215,3 sem/m² em
Floresta de Tabuleiro; Godim et al (2005) registrou 1.131,25 sem/m² em quatro
fragmentos de diferentes tamanhos sob domínio da Mata atlântica e Lagos e Marimon
(2012) 1.164 sem/m² em estudo realizado em mata de galeria no bioma Cerrado. A alta
densidade da chuva de sementes no Coité foi devido à presença da braquiária. Este fato
deve-se a dificuldade no controle desta espécie invasora e principalmente por ser a
espécie mais abundante ao redor dos coletores.
Em estudo realizado em Juiz de Fora (SOARES et al, 2009) em clareira
antrópica dominada pela invasora Melinis minutiflora (capim-gordura) verificou que
93% da chuva de sementes foi representado por esta espécie. A alta dominância do
capim-gordura diminuiu a equidade da área, que apresentou 0,19 de índice de Pielou.
No Coité 64% do aporte de chuva de sementes foi devido a presença de diásporos de
braquiária. Esta alta dominância, como no estudo de Soares (2009), foi responsável pelo
baixo valor do índice de Pielou que foi de 0,20. A presença de gramíneas invasoras em
áreas em processo de restauração no Cerrado prejudica sua capacidade de resiliência,
dificultando o recrutamento de espécies nativas (HOLANDA et al, 2010), seja pela
capacidade alelopática ou pela grande capacidade competidora dessas espécies (SOUZA
FILHO et al, 2005; SOUZA et al, 2006).
A densidade de diásporos, retirando-se a braquiária e considerando apenas os
diásporos coletados na APR, foi baixa (426,57sem/m²). Esta densidade para o Coite
revela que sua resiliência é baixa. Portanto, a compreensão da chuva de sementes é
importante para o conhecimento do potencial de regeneração natural (COLE et al, 2010;
CUBIÑA & AIDE, 2001; HOWE et al, 2010). Com isso a área do Coite apresenta uma
limitação para a chegada e recrutamento de diásporos, podendo assim necessitar de mais
intervenções para cicatrização da área degradada.
Na mata remanescente (MR) o aporte de chuva de sementes foi diferente da
APR, não sendo dominada por braquiária. A família Asteraceae foi a mais abundante na
MR. Resultado semelhante também foi encontrado em mata ciliar na região do rio

58
Itajaí-Açu, Mata Atlântica, onde foram coletados na chuva de sementes sob poleiros
artificiais seis espécies dessa família (TOMAZI et al, 2010).
Houve variação no aporte da chuva de sementes na área Coité conforme o
esperado. A variação anual na produção de frutos e sementes influencia na dinâmica e
no recrutamento das populações representando um componente importante a ser
entendido para estimar o potencial de regeneração de determinada comunidade
(PENHALBER & MANTOVANI, 1997). Esta variação é dependente, principalmente,
da fenologia das espécies, que para se entender o padrão seriam necessários estudos
específicos com repetições temporais.
A alta proporção de sementes pequenas na área experimental deve-se
principalmente a malha (<1 mm) utilizada nos coletores, a acurácia no processo de
triagem e a altura na qual os coletores foram instalados que permitiu coletar sementes de
vários estratos. O tamanho das sementes reflete o grau de preservação da área, ou seja, o
estágio sucessional. Em áreas abertas a alta incidência de luz favorece a proliferação de
espécies pioneiras, situação que se assemelha a das áreas degradadas (LAURANCE,
2001; OLIVEIRA et al , 2004). Mudanças nos padrões de distribuição espacial da
comunidade vegetal, como a que acontece em áreas abertas, resulta na escassez de
certos tipos morfológicos de sementes e domínio de outros (MELO et al, 2006).
A paisagem com domínio da pecuária nas propriedades rurais do Coité evidencia
as consequências da fragmentação na chuva de sementes. O processo de fragmentação
de paisagens naturais tem implicação direta na biodiversidade (FAHRIG, 2003),
influenciando na comunidade dentro dos remanescentes e na matriz circundante
(SAUDERS, 1991; JORDANO et al, 2006). Fragmentos remanescentes resultantes do
processo de fragmentação são afetados por problemas tais como isolamento, distância
de outros fragmentos, efeito de borda, tipo da matriz circundante dentre outros. Para um
grande número de espécies a fragmentação é prejudicial (CERQUEIRA et al, 2005).
A grande riqueza e abundância de espécies anemocóricas na APR revelam o
estagio sucessional inicial em que se encontra a área do Coité. Na MR foi amostrado
maior riqueza das espécies “não anemocórica”, ou seja, diásporos zoocóricos e
autocóricos contribuem mais para a chuva de sementes em áreas de matas, revelando o
estado mais avançado de conservação da mata remanescente, quando comparada a APR.
Na área de mata o dossel continuo facilita o recrutamento de plântulas lenhosas
favorecendo. Esta facilitação aumenta a atividade de aves, mamíferos e outros

59
dispersores o que aumenta, proporcionalmente, a deposição de sementes zoocoricas
(HOWE, 1990; HOLL et al, 2013).
Embora a MR apresente diásporos zoocóricos e autocóricos, ainda há limitação
para a chegada de diásporos zoocóricos de maior tamanho (>1cm) tanto na mata como
na APR. Na borda, esta fato pode ser explicado, segundo Holl (1999) e Cubiña & Aide
(2001) porque há alguns metros da borda da mata ocorre uma queda na deposição de
sementes zoocóricas. Para refinar esta análise, neste trabalho, seria necessário
desagrupar o grupo “não anemocorica” e diferenciar as síndromes de cada espécie. No
entanto, à dificuldade na identificação das espécies e a perda de características
morfológicas de algumas sementes ainda não foi possível essa caracterização e
quantificação das síndromes autocóricas e zoocoricas.
Sementes de espécies zoocóricas de tamanho médio como Schefflera grandiflora
(A.C. Sm.) Frodin e pequenas como Piper sp. e Cecropia polystachya Trécul ocorreram
na APR. Isto pode revelar que apesar do histórico da área, da presença de espécies
invasoras e da distância do fragmento mais próximo, tais fatores não representaram
filtros tão restritivos a chegada dessas espécies na APR, indicando que há
movimentação de fauna dispersora de sementes na área. A chegada de espécies
pioneiras e zoocóricas, como a do gênero Cecropia, ao sítio de restauração pode dar
início a sucessão secundária local (DOMINIQUE et al, 1986).
A presença de Cecropia polystachya e Piper sp. na APR evidencia a presença de
morcegos dispersores de sementes na área a ser restaurada. As espécies do gênero
Cecropia têm pelo menos cinco espécies de morcegos que dispersam suas sementes
(FLEMING & WILLIAMS, 1990; SATO et al, 2008). Estudos de Mello et al (2006)
sugeriram que a fenologia do gênero Piper na Mata Atlântica é o fator mais importante
na determinação das estações reprodutivas dos morcegos. Salazar et al (2013) estudaram
as consequências da dispersão não aleatória de Piper por morcegos da espécie Carollia
perspicillata (Phyllostomidae) e mostraram que a presença do dispersor e as
características da dispersão causam efeitos sobre a comunidade de Piper, influenciando
nas taxas de folivoria na espécie.
Relações entre morcegos e plantas através da quiropterocoria são essenciais para
a aceleração da sucessão secundária em áreas degradadas (MEDELLÍN & GAONA,
1999) e para a manutenção de processos ecológicos. O comportamento do animal de
dispersar as sementes é um auxilio essencial, um serviço ecossistêmico sem custo, na
restauração de ambientes degradados (REIS et al, 1999).

60
Na APR, mesmo com o pouco tempo de implantação do experimento, foram
encontradas diferenças no aporte entre os tratamentos. O tratamento que apresentou a
maior abundância de diásporos foi o tratamento T5 (linhas de diversidade e
recobrimento). As parcelas deste tratamento foram instaladas, aleatoriamente mais
próximas da área de mata de galeria ao longo do rio. Este fato pode ter contribuído para
a maior abundância de diásporos. Além disso, duas das parcelas do tratamento T5 foram
instaladas, aleatoriamente, em áreas mais planas, este fato pode ter facilitado à ação dos
ventos e com isso a coleta predominante de braquiária. As diferenças significativas
encontradas entre os tratamentos mostra a sensibilidade da chuva de sementes mesmo a
pequenas diferenças estruturais e de composição da vegetação. A MR apresentou a
maior riqueza confirmando que matas remanescentes são, ainda, a principal fonte de
biodiversidade em paisagens afetadas com atividades antrópicas.
Estabelecer plantios em linhas e em núcleos facilita a chegada de sementes de
espécies arbóreas no inicio da sucessão e representa uma estratégia aplicável para
aumentar a chuva de sementes em áreas agropastoris abandonadas (COLE et al, 2010).
Neste estudo ainda não foram observados tais diferenças mais refinadas devido ao
pouco tempo entre o plantio e a avaliação (12 meses), porém a longo prazo espera-se
que a chuva de sementes seja mais diversa nos tratamentos que combinaram mais
técnicas de restauração (T4: Nucleação + poleiros, por exemplo), pois aumenta a
heterogeneidade que é diretamente proporcional ao aporte de chuva de sementes.
Espera-se que com o passar do tempo e a cada monitoramento subsequente haja
incrementos gradativos de mais elementos na paisagem da área em processo de
restauração (APR) e reflita em um aumento no aporte da riqueza da chuva de sementes,
principalmente de sementes zoocóricas. Porém serão necessários estudos a longo prazo
para confirmar tal hipótese. A importância de estudos longos referentes ao processo de
restauração e o aporte da chuva de sementes também foi discutido por outros autores
como Cole (2010), Del Castillo e Rios (2008) e Holl e colaboradores (2013).
Embora o período de doze meses seja ainda pequeno em termos de restauração
ecológica os dados apresentados mostram que o processo foi desencadeado e pode ser
evidenciado ao se analisar qualitativamente a chuva de sementes como indicadora de
restauração ecológica. Dos aspectos analisados a eficiência do indicador chuva de
sementes fica evidente através dos resultados de diferenças entre a APR e a MR, bem
como pelas análises feitas entre os tratamentos. Desta forma foi possível se mensurar e
qualificar o aporte de sementes, podendo ser replicado em outros estudos, dando escala

61
ao experimento. Com o conhecimento da composição, síndrome de dispersão, tamanho
das sementes foi possível inferir sobre estado de conservação, presença de espécies
pioneiras e tardias e ausência de elementos na paisagem, isso reflete que os resultados
foram passiveis de compreensão e interpretação tornando previsíveis as tendências da
área. Este indicador reuniu sinteticamente várias características da área e das espécies
que compõem estes ambientes, o que possibilita saber o estado de conservação tanto da
APR como da MR, bem como a partir dos dados será possível traçar novas estratégias
para implementação dos experimentos no intuito de acelerar o processo de restauração.
No aspecto custo o indicador chuva de sementes mostrou-se não ser econômico,
tendo um custo médio em função dos gastos com a confecção de coletores, bem como a
demanda de uma grande equipe para coleta, triagem e pessoal treinado para a
identificação.
O aporte da chuva de sementes é um indicador abrangente, quantitativo e
qualitativo, porém para ter um entendimento mais refinado é necessário agrupar estudos
de chuva de sementes, recrutamento e sobrevivência de plântulas e indivíduos adultos,
bem como, entender quais são os filtros ecológicos envolvidos durante todo o processo,
desde a dispersão à sobrevivência do individuo arbóreo adulto, por exemplo. Estudos de
demografia de plantas indicam que uma comunidade vegetal é fruto da chegada de
sementes, chuva de sementes, e da sobrevivência destas no ecossistema (SCHUPP et al,
1989; REID & HOLL, 2012).
Aspectos sistêmicos como tamanho e a conectividade entre fragmentos
influenciam diretamente na comunidade vegetal local e consequentemente no aporte de
chuva de sementes (PIVELLO et al, 2006). Devido a esta importância do contexto de
paisagem na restauração ecológica e as diferenças apresentadas neste estudo a chuva de
sementes poderá servir em dois panoramas: como indicador do potencial de
restauração ou como indicador de automanutenção que consistem avaliar o estado de
conservação do ecossistema. Desta forma, a chuva de sementes é um parâmetro
importante para se avaliar tanto ambientes degradados como fragmentos remanescentes.

62
5.6. Referências Bibliográficas ALDRICH, P.R.; HAMRICK, J.L.; CHAVARRIAGA, P.; & KOCHERT, G. Microsatellite analysis of
demographic genetic structure in fragmented populations of the tropical tree Symphonia
globulifera. Molecular Ecology, v. 7, n. 8, p. 933-944, 1998.
ARAÚJO, R.S de. Chuva de sementes e deposição de serapilheira em três sistemas de revegetação de
áreas degradadas na Reserva Biológica de Poço das Antas (RJ). 2002. 92f. Dissertação (Mestrado em
Ciências Ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro, Seropédica, 2002.
BRANCALION, P. H.; MELO, F. P.; TABARELLI, M.; & RODRIGUES, R. R. Biodiversity persistence
in highly human-modified tropical landscapes depends on ecological restoration. Tropical
Conservation Science, v. 6, n. 6, p. 705-710, nov-dec. 2013.
BROWER, J. E.; ZAR, J. H.; & VON ENDE, C. N. Field and laboratory methods for general ecology.
Wm. C. C. Brown, Dubuque, Iowa, 1984, 161p.
CARMO, M. S. D.; BOAVENTURA, G. R.; & ANGÉLICA, R. S. Estudo geoquímico de sedimentos de
corrente da bacia hidrográfica do Rio Descoberto (BHRD), Brasília/DF. Geochimica Brasiliensis, v.
17, n. 2, p. 106-120, 2003.
CERQUEIRA, R.; BRANT, A.; NASCIMENTO, M.T.; PARDINI, R. Fragmentação: alguns conceitos.
In: RAMBALDI, D. M. O., & OLIVEIRA, D. A. S. D. O (orgs.). Fragmentação de ecossistemas:
causas, efeitos sobre a biodiversidade e recomendações de políticas públicas. 2ªed. 2005, 510p.
CLAUDE, M.; & PIZARRO, R. Indicadores de sustentabilidad y contabilidad ambiental para el caso
chileno. Sustentabilidad ambiental del crecimiento económico chileno. Santiago: Universidad de
Chile. 1996.
COLE, R. J.; HOLL, K. D.; & ZAHAWI, R. A. Seed rain under tree islands planted to restore degraded
lands in a tropical agricultural landscape. Ecological Applications, v. 20, n. 5, p. 1255-1269, 2010.
CRAWLEY, M. J. The R book. John Wiley & Sons Inc, p. 942, 2007.
CUBIÑA, A.; & AIDE, T. M. The effect of distance from forest edge on seed rain and soil seed bank in a
tropical pasture1. Biotropica, v. 33, n. 2, p. 260-267, 2001.
DALE, V.H.; BEYELER, S.C. Challenges in the development and use of ecological
indicators. Ecological indicators, n. 1.1, p. 3-10, 2001.
DEL CASTILLO, R.F., AND RIOS, M.A.P. Changes in seed rain during secondary succession in a
tropical montane cloud forest region in Oaxaca, Mexico. Journal of Tropical Ecology, n. 24, p. 433–
444, 2008.
DOMINIQUE, P.C. Inter-relations between frugivorous vertebrates and pioneer plants: Cecropia, birds
and bats in French Guyana in frugivores and seed dispersal. Springer Netherlands, p. 119-135, 1986.
EMBRAPA. Serviço Nacional de Levantamento e Conservação de Solos (Rio de Janeiro, RJ).
Levantamento de reconhecimento dos solos do Distrito Federal. Rio de Janeiro, EMBRAPA-SNLCS.
Boletim Técnico, v. 53. 1978. p. 455.
FAHRIG, L. Effects of habitat fragmentation on biodiversity. Annual review of ecology, evolution, and
systematics, p. 487-515, 2003.
FELFILI, J.M.; RIBEIRO, J.F.; FAGG, C.W.; MACHADO, J.W.B. Recuperação de matas de galeria.
Embrapa Cerrados. n. 21, p. 1-45, 2000. FERNANDES, S. C. Avaliação de imagens de satélite de alta e muito alta resolução espacial para a
discriminação de classes de uso e ocupação do solo: aplicação ao caso da sub-bacia do córrego
Capão Comprido no Distrito Federal. 2005. 106p. Dissertação (Mestrado em Tecnologia Ambiental
e Recursos Hídricos,) - Departamento de Engenharia Civil e Ambiental, Universidade de Brasília,
Brasília, DF. 2005 .
FERREIRA, V.A.P.; BORGES, M.N.; & ANJOS, E.F.S. Determinação dos aportes totais e carga por
unidade de área de nitrogênio e fósforo com base na caracterização do uso do solo nas bacias dos
Lagos Descoberto e Santa Maria, Brasília-DF. Acta Limnol, v. 4, p. 163-174, 1992.
FISCHER,J.; LINDENMAYER,D.B. The conservation value of paddock trees for birds in a variegated
landscape in southern New South Wales. 2. Paddock trees as stepping stones. Biodiversity and
Conservation, vol. 11, p. 833-849, 2002.
FLEMING, T.H. & WILLIAMS, C.F. Phenology, seed dispersal, and recruitment in Cecropia peltata
(Moracea) in Costa Rica tropical dry forest. Journal of Tropical Ecology, n. 6, p. 163-178, 1990.
GENTRY, A.H. Patterns of neotropical plant species diversity. Evolution Biology, n. 15, p. 1-84, 1982.
GONDIM, F.R. Aporte de serrapilheira e chuva de sementes como bioindicadores de recuperação
ambiental em fragmentos da floresta atlântica. 2005. 83p. Dissertação (Mestrado em Ciências
ambientais e Florestais) - Universidade Federal Rural do Rio de Janeiro. 2005.
HARPER, J.L. Population biology of plants. Population biology of plants. 1977.

63
HUBBELL, S.P.; FOSTER, R.B. Biology, chance, and history and the structure of tropical rain forest tree
communities. Community ecology, p. 314-330, 1986.
HOLANDA, F.S.R.; GOMES, L.G.N.; DA ROCHA, I.P.; SANTOS, T.T.; DE ARAÚJO FILHO, R.N.;
VIEIRA, T.R.S.; & MESQUITA, J.B. Crescimento inicial de espécies florestais na recomposição da
mata ciliar em taludes submetidos à técnica da bioengenharia de solos. Ciência Florestal, v. 20, n. 1,
2010.
HOLL, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: seed rain, seed
germination, microclimate, and soil. Biotropica, n. 31, p. 229-242, 1999.
HOLL, K.D. Restoring Tropical Forest. Nature Education Knowledge, v. 4, n. 4, p. 4, 2013.
HOWE, H.F. Survival and growth of juvenile Virola surinamensis in Planama: effects of herbivory and
canopy closure. Oecologia, n. 6, p. 259-280, 1990.
HOWE, H.; URINCHO-PANTALEON, Y.; DE LA PEÑA-DOMENE, M. & MART´INEZ-GARZA, C.
Early seed fall and seedling emergence: precursors to tropical restoration. Oecologia, v. 164, p. 731–
740, 2010.
JORDANO, P.; GALETTI, M.; PIZO, M. A.; & SILVA, W. R. Ligando frugivoria e dispersão de
sementes à biologia da conservação. In: Duarte, C.F.; Bergallo, H.G.; Dos Santos, M.A.; and V a,
A.E. (eds.). Biologia da conservação: essências. Editorial Rima, São Paulo, Brasil. 2006, p. 1-436.
LABOURIAU, M.L.S. A semente de Magonia pubescens St. Hil. - Morfologia e germinação. Academia
Brasileira de Ciências, n. 45, p. 501-537, 1973.
LAGOS, M.D.C.C.; & MARIMON, B.S. Chuva de sementes em uma floresta de galeria no Parque do
Bacaba, em Nova Xavantina, Mato Grosso, Brasil. Revista Árvore, n. 36, v. 2, p. 311-320, 2012.
LAURANCE, W.F. The hyper-diverse flora of the central Amazon: an overview. In: Bierregaard Jr,
R.O.; Gascon, C.; Lovejoy, T.E.; Mesquita, R. Lessons from Amazonia: ecology and conservation
of a fragmented forest. Yale University Press, New Haven, Connecticut. p. 47-53, 2001.
MARTINS, S.V. Recuperação de matas ciliares. 2 ed. Viçosa, MG. Aprenda Fácil Editora. 2007. 255p.
MELLO, M.A.R. Interações entre o morcego Carollia perspicillata (Linnaeus, 1758) (Chiroptera:
Phyllostomidae) e plantas do gênero Piper (Linnaeus, 1737)(Piperales: Piperaceae) em uma área de
Mata Atlântica. 2002.
MELO, F.P.L.; DIRZO, R.; & TABARELLI, M. Biased seed rain in forest edges: evidence from the
Brazilian Atlantic forest. Biological Conservation, v. 132, n. 1, p. 50-60, 2006.
METZGER, J.P.; CASATTI, L.; VERDADE, L. Indicadores de conservação e avaliação do
conhecimento para conservação. In: I Workshop de Síntese do Programa BIOTA/FAPESP na
Universidade de São Carlos, SP, 2002. Relatório do grupo de trabalho. Disponível em:
<http://www.biota.org.br/info/historico/simp2002/gt2.pdf>. Acesso em: 21 set. 2013.
MORELLATO, L.P.C.; LEITÃO-FILHO, H.F. Padrões de frutificação e dispersão na Serra do
Japi. História natural da Serra do Japi: ecologia e preservação de uma área florestal no Sudeste do
Brasil.(LPC Morellato, org.). Editora da UNICAMP/FAPESP, Campinas, p. 112-140, 1992.
NAVE, A. G. Banco de sementes autóctone e alóctone, resgate de plantas e plantio de vegetação nativa na
Fazenda Intermontes, município de Ribeirão Grande, SP. 2005. 218 f. Tese (Doutorado) - Escola
Superior de Agricultura Luiz de Queiroz. Universidade de São Paulo. Piracicaba, 2005.
OLIVEIRA FILHO, A.T. & RATTER, J.A. A study of the origin of central Brazilian forests by the
analysis of plant species distribution patterns. Edinburgh Journal of Botany, n. 52, p. 141-194,
1995.
OLIVEIRA, M.A.; GRILLO, A.S.; & TABARELLI, M. Forest edge in the Brazilian Atlantic forest:
drastic changes in tree species assemblages. Oryx, v. 38, n. 4, p. 389-394, 2004.
PIJL, LEENDERT. Principles of dispersal in higher plants. Springer-Verlag.1982.
PINHEIRO, F.; RIBEIRO, F.R. Síndromes de dispersão de sementes de matas de galeria do Distrito
Federal. In: Ribeiro, F.R.; Fonseca, C.E.L., Sousa-Silva, J. C. Cerrado: Caracterização e
recuperação de matas de galeria. Embrapa Cerrados. Brasilia. p. 899, 2001.
PIVELLO, V. R.; PETENON, D.; JESUS, F. M. D.; MEIRELLES, S. T.; VIDAL, M. M.; ALONSO, R.
D. A. S.; FRANCO, G.A.D.C. & METZGER, J.P. Chuva de sementes em fragmentos de Floresta
Atlântica (São Paulo, SP, Brasil), sob diferentes situações de conectividade, estrutura florestal e
proximidade da borda. Acta Botanica Brasilica, v. 20, n. 4, p. 845-859. 2006.
QUINTELA, M.F. Estudo do potencial de regeneração das espécies de uma floresta tropical de
Tabuleiros, Linhares, ES. São Carlos, 1996, 334f. Tese (Doutorado em Ecologia e Recursos
Naturais) – Centro de Ciencias Biologicas e da Saude, Universidade Federal de São Carlos. 1996.
REATTO, A.; MARTINS, E.D.S.; FARIAS, M.F.R.; SILVA, A.V.; CARVALHO JÚNIOR, O.D.;
OLIVEIRA JUNIOR, R.C.; ... & VALENTE, M. Mapa pedológico digital: SIG atualizado do
Distrito Federal escala 1: 100.000 e uma síntese do texto explicativo. Embrapa Cerrados, 2004. p.
31.

64
RÊGO, M.C.A.M. Mapeamento geotécnico da porção sul da área de proteção ambiental da bacia do
rio Descoberto.1997. 118p. (Dissertação de Mestrado em Geotecnia) - Universidade de Brasília.
1997.
REID, J.L.; & HOLL, K.D. Arrival≠ survival. Restoration Ecology, v. 21, n. 2, p. 153-155, 2013.
REIS, A.; KAGEYAMA, P.Y. Restauração de áreas degradadas utilizando interações interespecíficas. In:
Kageyama, P.Y.; Oliveira, R.E.; Moraes, L.F.D.; Engel, V.L & Gandara, F.B. Restauração ecológica
de ecossistemas naturais. Botucatu, FEPAF, p. 91-110. 2003.
REIS, A.; ZAMBONIM, R.M.; NAKAZONO, E.M. Recuperação de áreas florestais degradadas
utilizando a sucessão e as interações planta-animal. Série Cadernos da Biosfera, São Paulo, n. 14, p.
1-42, 1999.
RIBEIRO, J.F. Cerrado: matas de galeria. 1ª Ed .Planaltina: EMBRAPA-CPAC, 1998, p. 164.
RODRIGUES, E. Ecologia da restauração. 1ª Ed. Londrina: Editora Planta, 2013.
RODRIGUES, M.T. A biodiversidade dos Cerrados: conhecimento atual e perspectivas, com uma
hipótese sobre o papel das matas galerias na troca faunística durante ciclos climáticos. In: SCARIOT,
A.; SOUSA SILVA, J.C.; FELFILI, J.M. (Org.). Cerrado: Ecologia, biodiversidade e conservação.
Brasilia, DF: Ministério do Meio Ambiente, 2005, p. 235-246.
SALAZAR, D.; KELM, D.H.; & MARQUIS, R.J. Directed seed dispersal of Piper by Carollia
perspicillata and its effect on understory plant diversity and folivory. Ecology, v. 94, n. 11, p. 2444-
2453, 2013.
SANT’ANNNA, C.S.; TRES, D.R.; & REIS, A. Restauração ecológica: sistemas de nucleação. São
Paulo: SMA. p. 63, 2010.
SATO, T.M.; PASSOS, F.D.C.; & NOGUEIRA, A.C. Frugivoria de morcegos (Mammalia, Chiroptera)
em Cecropia pachystachya (Urticaceae) e seus efeitos na germinação das sementes. Papéis Avulsos
de Zoologia (São Paulo), v. 48, n. 3, p. 19-26, 2008.
SAUNDERS, D.A.; HOBBS, R.J. & MARGULES, C.R. Biological consequences of ecosystem
fragmentation: a review. Conservation Biology, v. 5, n. 1, p. 18-32, 1991.
SCHUPP, E.W.; HOWE, H.F.; AUGSPURGER, C.K. & DOUGLAS, J.L. Arrival and survival in tropical
treefall gaps. Ecology, v. 70, n. 3, p. 562-564, 1989.
SEGIP. The state enviromental Goals and Indicators Project. Evaluation criteria. In: BERGQUIST, G. et
al. Prospective indicators for state use in performance agreements. Florida: SEGIP/Florida Center
for public Management, 1995. p. B6-B7.
SER. Society for Ecological Restoration International and IUCN Commission on Ecosystem
Management. In: Ecological Restoration, a means of conserving biodiversity and sustainin
livelihoods (org.). Society for Ecological Restoration International, Tucson, Arizona, USA and IUCN,
Gland, Switzerland. 2004. p. 8.
SOARES, S. Banco de sementes, chuva de sementes eo uso de técnicas de nucleação na restauração
ecológica de uma clareira dominada por Melinis minutiflora P. Beauv. 2009 (Dissertação de
mestrado apresentada ao programa de pós-graduação em Ecologia da Universidade Federal de Juiz de
Fora).2009.
SOUZA FILHO, A.P.S.; PEREIRA, A.A.G. & BAYMA, J.C. Aleloquímico produzido pela gramínea
forrageira Brachiaria humidicola. Planta Daninha, v. 23, n. 1, p. 25-32, 2005.
SOUZA, L.S.; VELINI, E.D.; MARTINS, D.; & ROSOLEM, C.A. Efeito alelopático de capim-
braquiária (Brachiaria decumbens) sobre o crescimento inicial de sete espécies de plantas
cultivadas. Planta Daninha, n. 24, 2006.
STEFANELLO, D.; IVANAUSKAS, N.M.; MARTINS, S.V.; SILVA,E.; & KUNZ, S.H. Síndromes de
dispersão de diásporos das espécies de trechos de vegetação ciliar do rio das Pacas, Querência-
MT. Acta Amazonica, v. 40, n. 1, p. 141-150, 2010.
TOMAZI, A.L.; ZIMMERMANN, C.E.; & LAPS, R.R. Poleiros artificiais como modelo de nucleação
para restauração de ambientes ciliares: caracterização da chuva de sementes e regeneração
natural. Revista Biotemas, v. 23, n. 3, 2010.

65
6. CONCLUSÃO GERAL
Com os resultados foi possível interpretar as condições em que se encontram as
duas áreas: Jardim e Coité. Por exemplo, através do índice de diversidade, foi possível
perceber que estas se diferenciam quanto ao aporte de chuva de sementes. No Jardim o
índice é quase duas vezes maior que no Coité, permitindo concluir que elementos da
paisagem contribuíram para um aporte mais diversificado nesta área. Além disso, as
áreas diferem quanto ao histórico de uso da terra. Tais fatos influenciaram no aporte de
chuva de sementes e tiveram papel fundamental nas características dos diásporos da
chuva de sementes tanto da mata remanescente como da área em processo de
restauração. Dentro deste contexto, a avaliação da chuva de sementes como indicador de
restauração permitiu perceber essas diferenças entre as áreas.
Neste trabalho verificou-se que os padrões de riqueza e abundância da chuva de
sementes variam da área remanescente para a área degradada, sendo que a área
remanescente tem maior riqueza de espécies e menor abundância de diásporos do que a
área degradada, bem como a chuva de sementes foi capaz de indicar a capacidade de
regeneração natural caracterizando a quantidade de diásporos que chegam à área
degradada sendo, com isso, um bom indicador de restauração ecológica, confirmando
assim as hipóteses formuladas neste trabalho.
Como a chuva de sementes indica a capacidade de regeneração natural de uma
área degradada, por representar tanto as sementes autóctones como as alóctones, neste
estudo foi possível fazer uma análise quali-quantitativa do potencial de restauração de
cada área, desta forma conclui-se que a chuva de sementes é sensível as mudanças do
meio, apresenta resultabilidade, é facilmente interpretada e compreendida, permite
previsibilidade, apresenta parâmetros claros para se avaliar a capacidade de resiliência
de cada área. Portanto, estes resultados poderão nortear ações de manejo necessárias
para acelerar o processo de restauração da APR.
Em síntese a chuva de sementes é um ótimo indicador ecológico, podendo ser
aplicada como indicador do potencial de restauração, assim como indicador de
automanutenção. Porém o processo de restauração ecológico é muito mais lento no
Cerrado e requer mais alguns anos de avaliação para poder determinar as diferenças
mais precisas entre os métodos utilizados.

66
ANEXO 1
Tabela1: Espécies utilizadas no plantio nas áreas em processo de restauração do Rio Jardim e do Córrego Capão Comprido,
Distrito Federal.
Nome científico Família Nome popular
Garcinia gardneriana (Planch. & Triana) Zappi Clusiaceae Bacupari-da-mata
Copaiba langsdorfii (Desf.) Kuntze Fabaceae Copaíba
Calophyllum brasiliense Cambess. Calophyllaceae Guanandi
Aspidosperma parvifolium A. DC. Apocynaceae Guatambu-da-mata
Inga laurina (Sw.) Willd. Fabaceae Ingá-da-mata
Handroanthus impetiginosus (Mart. ex DC.)
Mattos
Bignoniaceae
Ipê-roxo
Handroanthus serratifolius (Vahl) S. O. Grose
Bignoniaceae Ipê-amarelo-da-mata
Cybistax antisyphilitica (Mart.) Mart. Bignoniaceae Ipê-verde
Genipa americana L. Rubiaceae Jenipapo
Alibertia sessilis (Vell.) K. Schum. Rubiaceae Marmelada-de-cachorro
Miconia chamissois Naudin Melastomataceae Miconia
Lafoensia glyptocarpa Koehne Lythraceae Mirindiba
Maclura tinctoria (L.) Don ex Steud. Moraceae Moreira
Guazuma ulmifolia Lam. Malvaceae Mutamba
Tapirira guianensis Aubl. Anacardiaceae Pau-pombo
Rapanea gardneriana (A. DC.) Mez Primulaceae Pororoca
Croton urucurana Baill. Euphorbiaceae Sangra-d’água
Tibouchina stenocarpa (DC.) Cogn. Melastomataceae Tibouchina
Tococa formicaria Mart. Melastomataceae Tococa

67
ANEXO 2 Tabela 2: Diásporos coletados na área do Rio Jardim (Distrito Federal) identificados ao menor nível taxonômico possível, juntamente com o grupo de dispersão, tipo
de diásporo, tamanho (mm) e quantidade total de diásporos para cada morfo coletada. Siglas: NA: não- anemocórica, A: anemocórica, S: Semente, F: Fruto.
Família Identificação Grupo de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade total
de diásporos
APOCYNACEAE Odontadenia sp. NA S 9 1
Matalea sp. A S 17 4
ARALIACEAE Schefflera grandiflora (A.C. Sm.) Frodin NA S 7 8
ASTERACEAE Ageratum conyzoides L. A S 2 7387
Clibadium armanii (Balb.) Sch. Bip. ex O.E. Schulz NA S 4 3
Bidens sp. NA S 8 11
Chromolaena maximilianii (generic SCHRADER ex DC.) R.M. King & H. Rob. A S 4 108
Eremanthus sp. A S 4 10
Blainvillea rhomboidea Cass. NA S 7 1
Erechtites sp. NA S 2 11
Vernonia polyanthes (Spreng.) Less. A S 3 1925
sp37 A S 2 102
sp65 A S 3 67
sp99 A S 2 38
sp110 A S 2 33
sp125 A S 4 290
A5 A S 3 17
A38 A S 2 6
BORAGINACEAE Cordia sp. NA S 1 40
BURSERACEAE Protium sp. NA S 9 354
COMBRETACEAE Terminalia glabrescens Mart. A S 9 681
Terminalia sp. A S 47 5
CYPERACEAE Fimbristylis miliacea (L.) Vahl NA S <1 1
DILLENIACEAE Davilla nitida (Vahl) Kubitzki NA F 6 164
EUPHORBIACEAE Chamaesyce hirta (L.) Millsp. NA S <1 1306
sp35 NA S 3 7
FABACEAE Zornia sp. NA S 2 9
Anadenanthera sp. NA S 13 4
Inga sp. NA F 50 10

68
Família Identificação Grupo de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade total
de diásporos
Crotalaria incana L. NA S 3 23
Stylosanthes viscosa (L.) Sw. NA S 3 2
Ormosia sp. NA S 5 1
Chamaecrista nictitans (L.) Moench NA S 3 1
Desmodium adscendens (Sw.) DC. NA S 3 6
A37 A S 11 1
A51 NA F 6 1
LAMIACEAE Hyptis sp. NA S 2 4
Hyptis suaveolens (L.) Poit. NA S 2 169
Hyptis lophantha Mart. ex Benth. NA S 1 270
LECYTHIDACEAE sp140 NA F 16 1
LYTHRACEAE Lafoensia sp. NA F 15 1
Cuphea sp. NA S <1 2482
MALVACEAE Sidastrum micranthum (A. St.-Hil.) Fryxell NA S 2 3226
Triumfetta semitriloba Jacq. NA F 5 376
Luehea sp. A S 5 194
MALPIGIACEAE sp135 A S 28 122
MELASTOMATACEAE Miconia cuspidata Mart. ex Naudin NA S 1 62
sp24 NA S 1 15
MYRTACEAE sp158 NA S 4 5
PHYLLANTHACEAE Phyllanthus tenellus Roxb. NA S 1 58
PIPERACEAE Piper sp. NA S 1 116
POACEAE Urochloa decumbens (Stapf) R.D. Webster A S 5 64370
Cenchrus sp. NA F 5 7
Rhynchelytrum repens (Willd.) C.E. Hubb. A F 3 673
Brachiaria mutica (Forssk.) Stapf NA S 3 2102
Cenchrus polystachios (L.) Morrone A F 4 6048
Isachne sp. NA S 1 38
Andropogon sp. A F 4 12
Pennisetum purpureum Schumach. A F 6 4
Paspalum sp. NA S 2 13
Melinis sp. NA F 2 192
sp44 A F 2 3

69
Família Identificação Grupo de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade total
de diásporos
sp70 NA S 1 1
sp78 NA F 1 2
POLYGONACEAE Securidaca sp. A S 35 7
RUBIACEAE Borreria alata (Aubl.) DC. NA S 2 542
Spermacoce latifolia Aubl. NA S <1 239
sp23 NA S 2 2
SAPINDACEAE Serjania sp. A S 28 337
Serjania erecta Radlk. A S 19 8
URTICACEAE Cecropia polystachya Trécul NA S 2 182
VOCHISIACEAE Vochysia pyramidalis Mart. A S 10 18
INDETERMINADAS sp7 NA S 3 31
sp14 NA F 15 2
sp15 NA S 2 3
sp17 NA F 5 10
sp21 NA F 7 3
sp32 NA S 2 74
sp34 NA S 3 77
sp36 NA S 4 284
sp38 NA S 3 9
sp40 A S 1 58
sp43 NA S <1 3
sp45 NA S 4 3
sp46 NA F 7 49
sp47 NA S 3 2
sp49 NA S 6 9
sp50 NA F 20 2
sp51 NA F 7 10
sp52 NA S <1 2
sp53 NA S 1 1
sp54 NA S <1 9
sp56 NA S 1 1
sp57 NA S 5 2
sp58 NA F 3 7

70
Família Identificação Grupo de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade total
de diásporos
sp59 NA S 2 3
sp60 NA F 13 4
sp61 NA F 3 13
sp62 NA S 3 4
sp63 NA S 15 162
sp64 NA S 3 2
sp72 NA S <1 85
sp75 NA S 3 118
sp82 NA F <1 2
sp83 NA S <1 342
sp85 NA F 2 3
sp86 NA S 4 19
sp89 NA F 3 16
sp93 NA S 2 100
sp97 NA S <1 2602
sp98 NA S <1 5
sp101 A S 2 8
sp106 NA S 1 5
sp108 NA S <1 7
sp109 NA F 5 3
sp111 NA S 1 114
sp112 NA S 2 1
sp115 NA F 8 2
sp118 NA F 10 3
sp119 A S 8 1
sp126 NA F 9 2
sp127 NA S 5 3
sp129 A S 28 2
sp132 NA S 3 1
sp134 NA F 8 3
sp136 NA S 6 2
sp137 NA F 5 3
sp138 NA F 5 2

71
Família Identificação Grupo de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade total
de diásporos
sp139 NA F 15 4
sp141 A S 38 1
sp142 NA F 9 1
sp147 NA F 5 24
sp154 NA S 1 5
sp156 NA F 13 5
sp157 NA S 7 2
sp159 NA S 7 3
sp160 NA S 5 1
sp161 NA S 2 2
sp168 A S 30 4
sp169 NA S 7 1
sp170 NA S <1 1068
sp172 NA F 4 1
sp173 NA F 8 1
sp174 NA S 1 4
sp175 NA S 2 4
sp183 NA S 3 10
sp184 NA S 7 1
A1 NA S 7 4
A2 A S 8 4
A3 NA S 2 33
A6 NA S 2 1
A17 NA S 4 1
A23 A S 5 2
A33 NA S 2 5
A34 A S 9 9
A36 NA S 5 1
A57 NA F 6 5
A58 NA S 3 1

72
ANEXO 3
Tabela 3: Diásporos coletados na área do Coité (Distrito Federal) identificados ao menor nível taxonômico possível, juntamente com o grupo de dispersão, tipo de
diásporo, tamanho (mm) e quantidade total de diásporos para cada morfo coletada.
Família Identificação Guilda de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade de
diásporos
APOCYNACEAE Matalea sp. NA F 17 2
ASTERACEAE Vernonia polyanthes (Spreng.) Less. A S 3 465
Ageratum conyzoides L. A S 2 787
Chromolaena maximilianii (generic SCHRADER ex DC.) R.M. King & H. Rob. A S 4 266
Eremanthus sp. A S 4 24
sp110 A S 3 1
sp125 A S 4 3225
sp65 A S 3 7
sp37 A S 3 2
A5 A S 3 2
A38 A S 2 1
A45 NA S 4 1
BORAGINACEAE Cordia sp. NA S 1 14
BURSERACEAE Protium sp. NA S 9 19
COMBRETACEAE Terminalia sp. A S 47 17
CYPERACEAE sp28 NA S 5 9
EUPHORBIACEAE sp25 NA F 7 2
sp92 NA S 3 14
FABACEAE Crotalaria incana L. NA S 3 1
Stylosanthes viscosa (L.) Sw. NA S 3 2
Senegalia sp. NA S 8 2
Lonchocarpus sp. NA S 8 4
Zornia sp. NA S 2 237
sp149 NA S 10 112
A24 NA S 3 3
LYTRACEAE Cuphea sp. NA S <1 4
MALPIGUIACEAE Tetrapterys sp. NA S 35 12
MELASTOMATACEAE Miconia cuspidata Mart. ex Naudin NA S 1 95
sp24 NA S 1 62

73
Família Identificação Guilda de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade de
diásporos
MORACEAE sp71 NA S <1 75
URTICACEAE Cecropia polystachya Trécul NA F 2 200
PIPERACEAE Piper sp. NA S 1 580
POACEAE Urochloa decumbens (Stapf) R.D. Webster A S 5 59631
Rhynchelytrum repens (Willd.) C.E. Hubb. A F 3 3
Brachiaria mutica (Forssk.) Stapf NA S 3 3937
Cenchrus polystachios (L.) Morrone A F 2 40
Olyra latifolia L. NA S 6 23
Isachne sp. NA S 1 2960
Melinis sp. NA F 2 1
Andropogon sp. A F 4 202
Pennisetum purpureum Schumach. A F 6 38
Paspalum sp. NA S 2 40
Oplismenus hirtellus (L.) P. Beauv. NA F 7 24
sp44 A F 2 6
sp70 NA S 1 1
POLYGONACEAE Securidaca sp. A S 35 3
PHYLLANTHACEAE Phyllanthus tenellus Roxb. NA S 1 2
RUBIACEAE sp23 NA S 2 1
Faramea cyanea Müll. Arg. NA F 6 9
Borreria alata (Aubl.) DC. NA S 2 2
SAPINDACEAE Serjania sp. A S 28 163
INDETERMINADAS sp7 NA S 3 39
sp21 NA F 7 2
sp32 NA S 2 16
sp40 A S 1 1
sp42 A S 11 1
sp45 NA S 4 496
sp46 NA F 7 28
sp54 NA S <1 12
sp61 NA F 3 4
sp66 NA S 2 5
sp68 NA F 6 4

74
Família Identificação Guilda de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade de
diásporos
sp72 NA S <1 3
sp75 NA S 3 8
sp87 NA F 7 4
sp89 NA F 3 1
sp94 NA F 3 8
sp101 A S 2 5
sp106 NA S 1 5
sp109 NA F 5 9
sp115 NA F 8 11
sp129 A S 28 3
sp134 NA F 8 2
sp139 NA F 15 1
sp146 NA F 9 1
sp147 NA F 5 1
sp150 NA S 9 9
sp151 NA S 9 1
sp153 NA S 1 1
sp154 NA S 1 22
sp161 NA S 2 2
sp163 NA S 1 2
sp165 A F 65 14
sp166 NA S 5 19
sp167 NA S 15 2
sp170 NA S <1 11
sp174 NA S 1 22
sp182 NA S 3 2
A6 NA S 2 7
A17 NA S 4 4
A20 NA S 5 34
A21 NA S <1 75
A27 NA S 3 5
A36 NA S 5 11
A40 NA S 5 1

75
Família Identificação Guilda de
dispersão
Tipo de
diásporo
Tamanho
(mm)
Quantidade de
diásporos
A47 A S 37 1
A48 NA S 2 1
A66 NA S 6 1
A75 A S 2 1
A76 A S 1 1
A79 NA F 30 1
A80 NA F 7 1
A81 A S 35 1
A82 A S 20 58
A84 NA F 2 1
A85 NA F 13 1
A86 A S 2 1