caracterizaÇÃo dos efeitos do fenol na cinÉtica ...livros01.livrosgratis.com.br/cp064074.pdf ·...
TRANSCRIPT

Instituto de Bioquímica Médica – CCS
Laboratório de Química Fisiológica da Contração Muscular
CARACTERIZAÇÃO DOS EFEITOS DO FENOL NACINÉTICA ENZIMÁTICA DA MIOSINA II
Raquel Guimarães Coelho
Rio de Janeiro
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
Raquel Guimarães Coelho
CARACTERIZAÇÃO DOS EFEITOS DO USO DO FENOL NACINÉTICA ENZIMÁTICA DA MIOSINA II
Profª Martha M. Sorenson
Orientadora
Professora Adjunta do Instituto de Bioquímica Médica
Profª Verônica Salerno Pinto
Co-orientadora
Professora Adjunta da Esco la de Educação Física e Desportos
Rio de Janeiro
2007
Dissertação de Mestrado apresentada ao
Programa de Pós-Graduação de Química
Biológica, Instituto de Bioquímica Médica,
Universidade Federal do Rio de Janeiro,
como parte dos requisitos necessários à
obtenção do título de Mestre em Ciências
(Química Biológica).

3
FICHA CATALOGRÁFICA
COELHO, Raquel Guimarães.
CARACTERIZAÇÃO DOS EFEITOS DO USO DO FENOL NA CINÉTICAENZIMÁTICA DA MIOSINA II
Rio de Janeiro, UFRJ, programa de Pós -Graduação em Química Biológica,2007.
N° de folhas: 108
Dissertação: Mestrado em Química Biológica
1 – Subfragmento 1 2 – Cinética Enzimática 3 - Fenol

4
FOLHA DE APROVAÇÃO
Raquel Guimarães Coelho
CARACTERIZAÇÃO DOS EFEITOS DO USO DO FENOL NA CINÉTICAENZIMÁTICA DA MIOSINA II
Rio de Janeiro,_______de_____________de 2007.
Banca Examinadora:
.........................................................................Profa. Martha M. Sorenson
Professora Adjunta do Instituto de Bioquímica Médica/ICB/UFRJOrientadora
................................................................ .........Profa. Verônica Salerno Pinto
Professora Adjunta da Escola de Educação Física e Desportos/CCS/UFRJCo-Orientadora
.........................................................................Prof. Antônio Galina Filho
Professor Adjunto do Instituto de Bioquímica Médica/CCS/UFRJ
.........................................................................Prof. Júlio Alberto Mignaco
Professor Adjunto do Instituto de Bioquímica Médica/CCS/UFRJ
............................................................ .............Prof. Marcelo Alves Ferreira
Professor Pesquisador da Fundação Oswaldo Cruz - FIOCRUZ
.........................................................................Prof. Marcelo Einicker Lamas
Suplente ExternoProfessor Adjunto do Instituto de B iofísica Carlos Chagas Filho /CCS/UFRJ
.........................................................................Prof. Hector Barrabin
Revisor e Suplente InternoProfessor Adjunto do Instituto de Bioquímica Médica/CCS/UFRJ

5
Dedico esta Tese primeiramente ao meu marido Tiago Costa Leite.
“Devido a minha admiração por você, iniciei todo es te processo. Através do seu
apoio e compreensão foi possível a minha dedicação na elaboração deste
trabalho, e através do nosso amor fomos companheir os neste e em todos os
trabalhos que hão de vir ”.
Sem dúvida, à família Coelho, por todo o amor e apoio de sempre, e:
A meu pai, Osório M. Coelho, por ter me ensinado a ter coragem e
perseverança.
A minha mãe, Joana Célia G. Coelho, por ter me ensinado a ser forte em todos
os momentos da minha vida.
A minha irmã, Camilla G. Coelho, por ter me dado a difícil missão de servir de
exemplo.
A vocês toda minha gratidão pelo incentivo e amor.

6
AGRADECIMENTOS
Ao Professor Héctor Barrabin pela revisão da tese e por gentilmente ter cedido
o Stopped-flow
Aos companheiros do Instituto pelo auxílio nos momentos de socorro.
À Professora Martha Sorenson pela orientação e amizade.
À Professora Verônica Salerno por me fazer parte integrante do Laboratório e
estar sempre me apoiando.
E aos amigos do Laboratório:
-Débora, nunca esquecerei de você.
-Marcelo e Daniel, amigos iguais a vocês é muito raro. Obrigada!
-Léo Nog, você é um exemplo.
-Cícero e Luciana, espero que a nossa experiência tenha sido tão proveitosa
para vocês como foi para mim.
-Renato, Zé Renato, Tiago, Leandro, Zé Henrique, Joyce e Jamila , todos vocês
contribuíram de alguma forma.

7
ABREVIAÇÕESA Actina
ADP Difosfato de adenosina
AlF4 Tetrafluor aluminato
ATP Trifosfato de adenosina
BSA Albumina sérica bovina
BeSO4 Sulfato de berílio
DTT Ditiotreitol
EDTA Ácido etilenodiamino tetracético
EGTA Ácido etileno-glicol-bis (β-aminoetil-éter)-N,N,N`,N`-tetracético
HEPES Ácido N-[2-hidroxietilpiperazina-N`-2 etanosulfônico]
HMM Meromiosina pesada
LC1 Cadeia leve 1 da miosina (ou essencial)
LC2 Cadeia leve 2 da miosina (ou regulatória)
LC3 Cadeia leve 3 da miosina (ou essencia l 2)
LMM Meromiosina leve
M Miosina
MESG 2-amino-6-mercapto-7-metilpurina ribosídeo
MHC Cadeia pesada da miosina
NaF Fluoreto de sódio
NaN3 Azida sódica
PEG Polietileno glicol
Pi Fosfato inorgânico
PNP Purina nucleosídeo fosforilase
rod Cauda da miosina
rpm Rotações por minuto
S1 Subfragmento-1 da miosina
S2 Subfragmento-2 da miosina
SDS Dodecilsulfato de sódio
SDS-PAGE Eletroforese em gel de poliacrilamida contendo dodecil sulfato
de sódio
TM Tropomiosina
TnC Troponina C

8
TnI Troponina I
TnT Troponina T

9
LISTA DE FIGURAS, TABELAS E ESQUEMAS
PágFigura 1: Constituição do músculo esquelético 20Figura 2: Modelo esquemático do filamento grosso e da miosina 22Figura 3: Fragmentos proteolíticos da miosina II 23Figura 4: Cristalografia do subfragmento -1 25Figura 5: Formas moleculares da actina 27Figura 6: Arranjo molecular do filamento fino 27Figura 7: Ciclo catalítico da miosina II 31Figura 8: Modelo da contração muscular 33Figura 9: Estrutura química do fenol 34Figura 10: Analise das purificações em gel de poliacrilamida 40Figura 11: Esquema representativo do Stopped-flow 48Figura 12: Gráfico representativo do sinal coletado no Stopped-flow 49Figura 13: Efeito do fenol na atividade Mg 2+ATPásica do S1 52Figura 14: Efeito do fenol na atividade Ca 2+ATPásica do S1 53Figura 15: Efeito do fenol na atividade K+EDTA-ATPásica do S1 54Figura 16: Efeito do tempo de exposição do S1 ao fenol sobreatividade Mg2+ATPásica
57
Figura 17: Efeito do tempo de exposição do S1 ao fenol sobreatividade actina ativada
58
Figura 18: Integridade do S1 na presença de fenol 60Figura 19: Efeito do fenol sobre a velocidade de hidrólise de ATP 63Figura 20: Efeito do fenol sobre a constante de ligação de ATP eisomerização do S1
64
Figura 21: Resultado obtido no experimento de single -turnover 68Figura 22: Esquema representativo do sistema MESG/PNP 69Figura 23: Liberação de Pi a partir do sistema MESG/PNP 71Figura 24: Resultado obtido no experimento de deslocamento deADP
74
Figura 25: Espectros de emissão de fluorescência do S1 napresença de fenol
81
Figura 26: Centro de massa espectral de fluorescência do S1 napresença de fenol
82
Figura 27: Efeito do fenol sobre o centro de massa espectral aolongo do tempo
84
Figura 28: Esquema representativo das mudanças conformacionaisna presença de análogos de ATP
86
Figura 29: Centro de massa espectral de fluorescência do S1complexados aos análogos na presença de fenol
89
Figura 30: Espectros de emissão de fluorescência do estadoS1*ADP na presença de fenol
90
Figura 31: Efeito do fenol sobre o centro de massa espectral nosestados S1 e S1*ADP
91
Figura 32: Resultado obtido do ensaio de co -sedimentação napresença de fenol por gel de SDS
95
Figura 33: Quantificação do S1 não ligado por co -sedimentação 96Figura 34: Reversibilidade do efeito do f enol na atividadeMg2+ATPásica
97

10
Figura 35: Reversibilidade do efeito do fenol na atividade K+EDTA -ATPásica
98
Tabela 1: Condições experimentais utilizadas para atividadesATPásicas do S1
44
Tabela 2: Efeito do fenol sobre atividade específica do S1 55Tabela 3: Resultado obtido das constantes cinéticas iniciais do ciclocatalítico na presença de fenol
65
Tabela 4: Resultado obtido das constantes cinéticas pós -hidrólise dociclo catalítico na presença de fenol
67
Esquemas 1: Ciclo catalítico da miosina II na ausência de actina 61Esquemas 2: Esquema das etapas de liberação de ADP 72

11
SUMÁRIO
Pág1 – Introdução 18
1.1 - Constituição e Organização do Músculo Esquelético 19
1.2 - O Filamento Grosso 21
1.3 - O Filamento Fino 25
1.4 - Mecanismo da Contração Muscular 28
1.5 - Mecanismo Cinético 29
1.6 - O Fenol 34
2 – Objetivos 37
3 – Materiais e Métodos 38
3.1 – Miosina 38
3.2 – Subfragmento-1 40
3.3 – Actina 41
3.4 – Polimerização de Actina 42
3.5 – Eletroforese 43
3.6 – Atividades ATPásicas 43
3.7 – Co-sedimentação 45
3.8 – Fluorescência em Estado Estacionário 46
3.9 – Cinética Rápida 47
3.10 – Reagentes 50
3.11 – Análise dos Dados 50
4 - Resultados: Parte I 51
4.1 – Atividade ATPasica com Diferentes Cátions na Presença de
Fenol
51
4.2 – Efeito do Tempo de Exposição do S1 ao Fenol sobre a
Atividade ATPasica
56
4.3 – Experimentos de Cinética Rápida Utilizando Stop ped-Flow 61
4.3.1 – Estados Pré-hidrólise 61
4.3.2 – Estados Pós-hidrólise 66
5 - Discussão: Parte I 75
6 - Resultados: Parte II 80
6.1 – Efeito do Fenol na Estrutura Terciária do S1 80

12
6.2 – Efeito do Fenol na Estrutura Terciária d o S1 na Presença de
Análogos de ATP
85
6.3 – Efeito do Fenol na Estrutura Terciária do S1 na Presença de
ADP
87
6.4 – Modificação Estrutural do S1 pelo Fenol Altera a Ligação com
Actina
92
6.5 – Reversibilidade dos Efeitos do Fenol 93
7 – Discussão: Parte II 99
8 – Conclusão 103
9 - Referências Bibliográficas 104

13
RESUMO
Coelho, Raquel Guimarães. Caracterização dos efeitos do uso do fenol na
cinética enzimática da miosina II. Rio de Janeiro, 2007. Dissertação (Mestrado
em Ciências Biológicas) – Instituto de Bioquímica Médica, Universidade
Federal do Rio de Janeiro.
Durante algumas décadas o fenol foi muito utilizado na clínica como
tratamento de algumas pato logias que apresentavam desordens
neuromusculares. Seu ponto de ação seria sobre o motoneurônio, reduzindo
sua atividade elétrica e conseqüentemente diminuindo a contração muscular. A
miosina é a enzima responsável pela conversão de energia química
proveniente da hidrólise do ATP em energia mecânic a durante a contração
muscular, e o fenol tem se mostrado um composto capaz de alterar o
comportamento enzimático da miosina II.
No estado estacionário, o fenol (0,25-0,5%) promove a ativação da
atividade Mg2+ATPásica do subfragmento -1 da miosina II, e concentrações
maiores deste promovem inibição. Entretanto, os efeitos do fenol são
específicos para o íon complexado ao ATP (Mg2+ ou Ca2+), pois a atividade
Ca2+ATPásica sofre uma pequena influê ncia do composto e a atividade
K+EDTA-ATPásica é inibida na presença d a droga. Ao analisarmos o ciclo
catalítico da miosina na presença de fenol por medidas de fluorescência
intrínseca (M*, M**), percebemos que este composto é capaz de diminuir as
constantes de ligação (passos 1 e 2) e a hidrólise do ATP (passo 3).

14
As constantes pós-hidrólise também foram analisadas na presença de
fenol, através de experimentos de single turnover e experimentos de
deslocamento de ADP. Nossos resultados sugerem que o efeito observado no
estado estacionário se deve ao aumento da liberação dos produtos do ciclo
catalítico (passo 4 e passo 6).
A actina promove um aumento da atividade Mg 2+ATPásica do S1 por
acelerar o mesmo passo (k +4) do ciclo. Entretanto, na presença de fenol, nós
observamos que o aumento na ativ idade enzimática não é tão pronunciado na
presença de actina quanto na sua ausência. Experimentos de co -sedimentação
mostraram que o fenol promove uma diminuição da ligação de actina à
miosina, alterando assim o estado de rigor. Uma aná lise estrutural por
fluorescência foi realizada para verificar se o fenol seria capaz de modular a
estrutura da proteína. Foi verificado que o fenol promove alteração tanto da
intensidade de fluorescência como também do centro de massa do S1 na
ausência de nucleotídeo, bem como na presença de ADP. Est es dados
sugerem que o solvente altera a estrutura terciária do S1 de forma a deixá -la
em uma conformação mais desenovelada, o que favorece a liberação dos
produtos. Entretanto, após experimentos de reversibilidade por diluição do
solvente, verificamos que os efeitos do fenol são revertidos completamente ,
tanto na atividade Mg2+ATPásica quanto na K+EDTA-ATPásica.
Estes dados indicam que o fenol possui grande influ ência na atividade
enzimática do S1, pois altera todos os passos do c iclo catalítico, acelerando o
passo limitante (k+4). As modificações estruturais promovidas pelo fenol podem
M+ATP M.ATP M*.ATP M**.ADP.Pi M*.ADP M.ADP M
ADPPi
k2 k5K1 K3 k6k4

15
contribuir para a liberação dos produtos, além de alterarem a ligação com a
actina. Sendo assim, o fenol se mostrou um composto específico apresent ando
efeitos bifásicos, dependendo da concentração e do tempo de ex posição à
proteína, sendo os efeitos reversíveis em baixas concentrações (até 0,5% de
fenol).

16
ABSTRACT
Coelho, Raquel Guimarães. Characterization of effects of phenol on the kinetics
of myosin II. Rio de Janeiro, 2007. Dissertação (Mestrado em Ciências
Biológicas) – Instituto de Bioquímica Médica, Universidade Federal do Rio de
Janeiro.
For a long time, phenol has been used to treat neuromuscular disorders
that are characterized by spasticity . Phenol would act on the motor neuron,
diminishing the electrical activity and consequently muscle contraction. Myosin
is the enzyme responsible for conver ting chemical energy derived from ATP
hydrolysis into the mechanical energy of muscle contraction and we show that
phenol can alter the enzymatic activity of myosin II.
In the steady state, phenol (0.25-0.5%) increases the Mg2+ATPase
activity of subfragment-1 (S1) of myosin II ; higher concentrations inhibit the
activity. However, phenol effects are specific to the cation-ATP complex (Mg2+
or Ca2+): the Ca2+ATPase is less activated, while the K+EDTA-ATPase activity is
markedly inhibited. Analysis of the catalytic cycle of S1 by intrinsic fluorescence
(M*, M**) shows that phenol reduces the binding constants (steps 1 and 2) and
the rate of ATP cleavage (step 3).
The post-hydrolysis rate constants were analyzed using single-turnover
and ATP chase experiments to estimate the rate of Pi and ADP release (step 4
and step 6). Our results suggest that the effects observed in the steady state
arise from enhanced rates of product release, particularly step 4.
M+ATP M.ATP M *.ATP M**.ADP.Pi M*.ADP M.ADP M
ADPPi
k2 k5K1 K3 k6k4

17
Actin promotes an increase in the Mg2+ATPase activity of S1 by
accelerating the same step (k+4). However, the increase in activity due to phenol
is not as pronounced in the presence of actin as in its absence. Co-
sedimentation assays showed that phenol decreases actin binding to S1 in the
rigor state. Intrinsic fluorescence was used to verify whether phenol modulates
the tertiary structure of S1. Phenol alters the fluorescence intensity and the
center of spectral mass of S1 in the absence of nucleotide and in the presence
of ADP. These results suggest that phenol promotes partial unfolding at S1,
which may facilitate the release of products . However, after reversibility
experiments by dilution of the solvent, the kinetic effects of phenol (0.25-0.5%)
were nevertheless completely reversed.

18
INTRODUÇÃO
Muitas lesões musculares possuem uma alta correlação com
deficiências ou disfunções do sistema neural, uma vez que ambos sistemas
(muscular e nervoso) interagem fisiologicamente. Uma alteração em qualquer
componente desses sistemas pode afetar sensivelmente a funcionalidade de
outros componentes. Muitas doen ças neurológicas levam o indivíduo a
desenvolver espasticidade ou rigidez acentuada da musculatura, provocando
um arco de movimento diminuído, dor e fadiga muscular (Sanger, 2003).
Durante anos utilizou-se o fenol como bloqueador químico para reduzir o
quadro de espasticidade muscular em pacientes que apresentavam esse
sintoma. Muitos estudos defendiam sua prática por reduzir a hipertonia
muscular e melhorar o quadro álgico (Koyama et al., 1992). Entretanto, pouco
se sabe sobre os efeitos colaterais do uso desse composto, principalmente no
que diz respeito a sua ação direta sobre as proteínas contr áteis.
Dentre as proteínas do músculo esquelético, a miosina II possui um ciclo
catalítico complexo, e a funcionalidade desse ciclo garante não só a tonicidade
muscular, mas também o processo contrátil. Portanto , esta tese visa
caracterizar a ação do fenol sobre a atividade ATPásica da miosina.

19
1.1 – Constituição e Organização do Músculo Esquelético
O músculo esquelético é formado por um número variável de células.
Essas células, também chamadas de fibras musculares, se distribuem em
pequenos grupos separados por tecido conectivo, formando feixes musculares.
As fibras musculares possuem em seu interior proteínas que são responsáveis
pelo processo contráti l, formando as miofibrilas (Fig. 1A). Estas proteínas se
dispõem alternadamente de forma paralela , formando um esqueleto protéico.
Os sarcômeros são pequenas unidades contráteis das fibras musculares (Fig.
1B), compostos basicamente por estruturas filamentosas: o filamento grosso é
formado por moléculas de miosina , e o filamento fino constituído principalmente
por actina e proteínas regulatórias, que formam o s complexos troponina-
tropomiosina. Essa organização espacial permite que as essas proteínas
interajam entre si de maneira eficiente para produzir força e movimento
direcionados (Huxley, 1953).

20
Figura 1: Esquema representativo do nível de organização do músculoesquelético. (A) Área de seção transversa de músculo mostrando a s fibras
musculares agrupadas em feixes , separados por tecido conjuntivo, até a
microestrutura celular, indicando a fibra e seus constituintes. (B) Um arranjo
estrutural de um sarcômero, mostrando a disposição dos filamentos finos e
grossos por microscopia eletrônica (http:curlygirl.naturlink.pt/sarcomero.jpg).
B
ATendão
Fáscia
Músculo
Feixemuscular
Motoneurônio
Capilar
Retículosarcoplasmático
FilamentosNúcleo
Osso
Epimísio
PerimísioFibramuscularSarcolemaMiofibrila

21
1.2 – O filamento grosso
O filamento grosso é composto basicamente por moléculas de miosina
II, membro especializado de uma família denominada de motores moleculares,
encontrada em quase todas as células do corpo (Rayment et al., 1993;
Rayment, 1996). No músculo esquelético, essa proteína agrega -se em feixes,
com projeções chamadas de cabeça da miosina na superfície, o que permite
sua interação com os filamentos de actina (Huxley, 1953).
A miosina II é uma proteína formada por duas longas cadeias
polipeptídicas que se enovelam, compondo uma molécula com duas cabeças e
um corpo filamentoso (Fig. 2A). Essa conformação lhe confere um peso
molecular de aproximadamente 520.000 Da. A porção amino-terminal se
encontra na região da cabeça da cadeia pesada da miosina (MHC). Essa
região possui uma estrutura globular (por esse motivo recebe o nome de
cabeça), onde estão localizados o sítio de interação com o filamento fino e o
sítio catalítico da proteína. A porção C-terminal se localiza na região do
enovelamento de α-hélices de cada cadeia pesada formando um “coiled-coil”.
Entre a porção globular e a filamentosa, encontra -se a região do pescoço, onde
estão ancoradas as cadeias leves (LC) (Fig. 2B). As LCs s ão classificadas em
duas categorias: não essenciais ou regulatórias (LC 2) e essenciais (LC1 e LC3).
Essa classificação corresponde à perda da atividade ATPásica da miosina que
ocorre quando são removidas as LC 1 e LC3 (Mannherz & Goody, 1976) o que
sugere que essas subunidades realizam uma importante função estrutural
(Trybus, 1994). O mesmo não ocorre quando a LC 2 é removida. Cada MHC
está associada a uma LC essencial (LC 1 ou LC3) e uma LC não essencial
(Brooks et al., 2004).

22
Figura 2: Modelo esquemático do filamento grosso. (A) O filamento grosso
é formado pela agregação de diversas moléculas de miosina pelas caudas,
apresentando polaridades opostas de cada lado do filamento. (B)
Representação de uma única molécula de miosina II, mostrando as porções
globulares (cabeça), o pescoço, onde se encontram as cadeias leves
essenciais e não essenciais (representadas em preto) e a porção filamentosa
(cauda) (adaptado de Sellers & Goodson, 1995).
Uma ferramenta muito útil para o estud o da miosina e suas propriedades
é a utilização de pontos particulares em sua estrutura suscetíveis à proteólise
(Szent-Györgyi, 1951). Sendo assim, a miosina pode ser facilmente clivada em
fragmentos funcionais separados utilizando quimotripsina ou papaín a. Esse
tratamento ocasiona a separação da molécula em duas porções principais: a
meromiosina pesada (HMM) e meromiosina leve (LMM). A porção HMM pode
ser clivada gerando o subfragmento 1 (S1) e o subfragmento 2 (S2), como
mostrado na Figura 3.
B
A

23
Figura 3- Fragmentos proteolíticos da miosina II. Pela clivagem com
papaína obtém-se HMM (a) e LMM (b). Pela clivagem com quimotripsina na
presença de EDTA obtém-se S1 ligado às LC1 e LC3 (c) e a cauda (e), que
compreende o S2 (d) e o LMM (b) (Sellers & Goodson, 1995).
O fragmento globular da cabeça da miosina é chamado de domínio
motor (Rayment, 1996). Esta porção do S1 contém três subdomínios
estruturais distintos que podem ser divididos pela digestão limitada do S1 com
tripsina: o fragmento N-terminal de 25 kDa, o fragmento de 50 kDa e o
fragmento C-terminal de 20 kDa (Rayment et al., 1993). Através da estrutura
cristalográfica da proteína é possível identificar o sítio de ligação de
nucleotídeo, o qual se localiza entre os fragmentos de 25 kDa e 50 k Da (Fig. 4).
A interação com o filamento fino se dá pela ligação com actina , e esse ponto de
interação se localiza também no subfragmento de 50 KDa e na porção inicial
do subfragmento de 20 KDa (Geeves & Holmes, 1999; Sellers & Goodson,
1995). É importante observar que o local de interação com nucleotídeo e o sítio
de ligação com actina são fisicamente opostos. Essa forma estrutural é muito

24
importante para a compreensão do mecanismo enzimático do S1 , como
veremos adiante. O subfragmento de 20 kDa possui uma extensa α-hélice, com
uma pequena região chamada de região conversora. Essa região é
responsável por transmitir os movimentos rotacionais ocorridos durante a
hidrólise do ATP até a região do pescoço (Geeves & Holmes, 1999).
Durante a atividade enzimática desta molécula, algumas mudanças
conformacionais ocorrem (Hiratsuka, 1992). O monitoramento dessas
mudanças estruturais é possível porque o S1 possui 7 resíduos de triptofano
(dos 42 presentes na miosina). O Trp 510 é o resíduo que responde a
alterações estruturais, promovendo mudanças de fluorescência intrínseca
induzidas pela ligação e hidrólise de ATP (Park et al., 1996). Esse resíduo
localiza-se no domínio central de 50 kDa. Assim como a miosina , é possível
utilizar o S1 para o estudo da cinética enzimá tica da molécula, acompanhando
algumas atividades ATPásicas clássicas: Mg 2+ATPase e Mg2+ATPase actina-
ativada (consideradas fisiológicas) e também Ca 2+ ATPase e K+EDTA-ATPase.

25
Figura 4: Estrutura do S1 de músculo es quelético de galinha, obtida porcristalografia de raio-X (adaptado de Rayment et al., 1993). Observa-se a
formação basicamente de α-hélices, que vão da região globular até a cauda. A
porção superior e a inferior de 50 kDa estão representadas em vermelho, a
porção de 25 kDa em verde , e em azul a porção de 20 kDa. Estão
representadas também as cadeias leves essenciais (LCE - LC1 em amarelo) e
a cadeia leve regulatória (LCR-LC2 em rosa).
1.3 - O filamento fino
O principal constituinte do filamento fino é a actina, proteína altamente
conservada em diferentes células eucarióticas. A actina é formada pela
oligomerização de monômeros com baixo peso molecular (42.000 Da) (Geeves
& Holmes, 1999). Essa proteína pode assumir duas formas na célula: uma
monomérica (G-actina) e outra polimérica filamentosa (F-actina). No músculo
esquelético, a actina encontra -se quase que em totalidade sob a forma
filamentosa. Em condições fisiológicas (força iônica entre 0,15 e 0,2 M), a
actina se polimeriza, isto é, sofre um processo de agregação dos monômeros
Sítio de ligaçãodenucleotídeo
Sítio de ligaçãopara actina
LCE
LCR
S1 da Miosina Muscular Esquelética de Galinha

26
assumindo então a forma filamentosa (Fig. 5). Esse processo é dependente de
ATP, fazendo com que esta molécula de certa forma também seja considerada
uma ATPase (Huxley, 1953) . A formação do filamento fino é importante para
compor a célula muscular e ativar o processo contrátil, favorecendo a liberação
dos produtos da hidrólise de ATP pelo S1.
Além da actina, fazem parte do filamento fino duas outras proteínas, as
quais formam o complexo regulatório. São elas: a tropomiosina (Tm) e a
troponina (Tn) (Fig. 6). A Tn possui, por sua vez, três subunidades: a TnC,
capaz de ligar Ca2+ e funcionar como um sensor da contração muscular; a TnI,
que inibe a contração muscular em presença de ATP; e a TnT, que tem função
ancoradora, ligando o complexo à tropomiosina e/ou actina (Huxley, 1953;
Takeda et al., 2003). O complexo regulatório é assim chamado, pois ele regula
a interação acto-miosina. Este complexo circunda o filamento de actina a
intervalos de sete monômeros de actina, gerando um impedimento físico entre
os filamentos de actina e miosina (Farah & Reinach, 1995; Schiaffino &
Reggiani, 1996)

27
Figura 5: Esquema representativo das formas moleculares da actina. A
forma globular, G-actina (acima) e a forma filamentosa, F-actina (abaixo)
formado pelo enovelamento de doi s filamentos.
(http://ghr.nlm.nih.gov/handbook/illustrations/actin.jpg).
Figura 6- Modelo esquemático do arranjo molecular do filamento fino. O
complexo regulatório formado pela troponina (Tn) e pela tropomiosina (Tm)
sobre a actina no filamento fino e squelético. As subunidades do complexo
troponina, a TnC (vermelho), TnT (amarelo) , e TnI (verde), encontram-se sobre
as duas cadeias de tropomiosina, representadas em marrom e em laranja, que
se enroscam sobre o filamento de actina (cinza) num intervalo de sete
monômeros de G-actina (adaptado de Ebashi, 1974).
Actina
Monômero de actina
Forma filamentosa da actina
A c t i n a T r o p o m i o s i n a
S o b r e p o s i ç ã oc a b e ç a - c a u d a

28
1.4 – Mecanismo da Contração Muscular
O mecanismo da contração muscular vem sendo estudado por
pesquisadores do mundo todo e diversas controvérsias ainda persistem sobre
o funcionamento da maquinaria contrátil. A contração muscular não é um
evento exclusivo do músculo esquelético, pois o miocárdio e o tecido muscular
liso também desenvolvem força e moviment o através da interação protéica e
da conversão da energia química contida nas moléculas de ATP em trabalho
mecânico (Brooks et al., 2004). A miosina , juntamente com a actina responde
pelo processo da contração muscular através do trabalho físico -químico gerado
pela interação e deslizamento dessas proteínas. A “teoria dos filamentos
deslizantes” foi formulada por dois grupos ingleses (Huxley, 1957), baseados
em observações do encurtamento e estiramento do músculo. Os filamentos
finos deslizariam sobre os filamentos grossos , e esse movimento geraria a
tensão muscular. Apesar de basicamente correto, entretanto, hoje se sabe que
o mecanismo é muito mais complexo. É um consenso que ocorre interação
entre os filamentos grossos e finos, essa interação é dinâmica de forma a fazer
com que haja o deslizamento entre eles, e mais, todo esse processo é
dependente de ATP, ou melhor, da sua hidrólise pela porção catalítica da
miosina.
Juntamente com esses fenômenos moleculares existe também a
participação de um outro sistema e tecido. O processo de acoplamento
excitação-contração consiste na seqüência de eventos que dão origem à
contração muscular. Esses eventos envolvem a chegada de um impulso
nervoso à junção neuromuscular, a conseqüente despolarização da membrana
da célula muscular, seguida pela propagação dessa despolarização por toda a

29
fibra pelos túbulos “T”. Todos os efeitos posteriores gerados no interior da fibra
são ocasionados pelo influxo de Ca 2+ para dentro do sarcoplasma, liberado
pelo retículo sarcoplasmático. O aumento na concentração de Ca 2+ é percebido
pelo complexo regulatório, mais precisamente pela subunidade TnC. A TnC é a
primeira a sofrer uma mudança conformacional após a ligação de Ca2+. Essa
ligação favorece o deslocamento das subunidades inibitórias (TnI e TnT) de
forma a promover uma mudança no posicionamento da Tm, proteína que
realiza o impedimento físico para a interação entre actina e a miosina. No
músculo relaxado não há aumentos na concentração de Ca2+ e, portanto, o
complexo regulatório exerce seu papel até a chegada de algum estímulo
nervoso. Dessa forma, a ligação de Ca2+ na TnC permite a formação das
pontes cruzadas (ligação acto -S1). A formação dessas pontes e a hidrólise d e
ATP pela miosina promovem a contração muscular (Capitanio et al., 2006)
1.5 – Mecanismo cinético
Os primeiros estudos cinéticos da hidrólise do ATP pela miosina foram
realizados, no estado estacionário, entre as décadas de 40 e 50 (Szent -
Györgyi, 1951). As medidas realizadas no estado estacionário permitem a
obtenção de características enzimáticas importantes como as constantes de
afinidade e o comportamento da enzima em diferentes situações (Trybus,
2000). Entretanto, poucas informações sobre as constantes cinéticas e os
movimentos estruturais podem ser obtidas por esse método. Em 1971, Lymn e
Taylor propuseram um modelo para a interação acto -miosina e hidrólise de
ATP, baseado em medidas de cinética rápida usando proteínas purificadas.

30
Nesse modelo o ATP liga-se à miosina e é rapidamente hidrolisado em ADP e
Pi. A fase seguinte é a liberação de Pi , seguida pela liberação de ADP;
entretanto essa fase pós-hidrolise é muito lenta, limitando o andamento do ciclo
(Lymn & Taylor, 1971). Esse ciclo seria típico da fase de relaxamento muscular ,
onde vemos uma atividade enzimática basal, devido à lentidão do ciclo. Na
presença de actina, a atividade enzimática da m iosina pode ser acentuada em
mais de 10 vezes, dependendo da concentração de actina, de ATP , e da força
iônica do meio (Geeves, 1991). Na ausência de ATP e do complexo regulatório,
a miosina se liga fortemente à actina (K a ≈ 107 -108 M-1), caracterizando uma
ligação chamada de “estado de rigor”. A presença de ATP no meio reduz a
constante de afinidade, pois ele se liga rapidamente à miosina, diminuindo a
afinidade entre miosina e actina (K ≤ 104 M-1) (Geeves, 1991), levando à
formação de pontes fracas, de modo que a actina se dissocia da miosina. O
ATP ligado à miosina é então rapidamente hidrolisado , formando M.ADP.Pi. O
complexo ternário tem sua afinidade por actina recuperada , de modo que as
proteínas se ligam novamente , formando um complexo AM.ADP.Pi. Essa nova
ligação da actina acelera a liberação dos produtos da hidrólise do ATP,
principalmente a saída de Pi, descrita como a etapa limitante do ciclo (Lymn &
Taylor, 1971). Após a liberação de ADP, actina e miosina voltam a formar
pontes de rigor até que uma nova molécula de ATP desfaça novamente a
ligação AM. Dessa forma a presença de actina garante uma interação cíclica e
dinâmica (Fig. 7).

31
Figura 7: Ciclo catalítico da miosina (M) e actomiosina (AM) (Goldman &Brenner, 1987). Para alguns passos deste ciclo são apresentadas no texto as
constantes de associação entr e M e A. Essas constantes são dadas em M-1.
No modelo apresentado por Lymn & Taylor (1971), o evento gerador de
força está associado com a liberação dos produtos da hidrólise do ATP. Desse
modo, a actina promoveria uma mudança estrutural no complexo AM.ADP.Pi ,
de forma que a miosina desenvolva um movimento direcionado e orientado.
A Figura 8 correlaciona o modelo cinético proposto por Lymn & Taylor
(1971) com um modelo estrutural de interações entre actina e miosina,
mostrando que a comunicação entre o sítio de ligação de actina e o sítio de
ligação de ATP é coordenada através da abertura e fechame nto de uma fenda
existente na cabeça da miosina (localizado entre a porção superior e inferior de
50 kDa). No estado de rigor (A) a actina fecha a fenda e a miosina está
fortemente ligada a actina. A ligação de ATP ocorre em duas etapas:
inicialmente a ligação de ATP leva à abertura da fenda, diminuindo a afinidade
da miosina pela actina (B) e posteriormente o sítio de ligação de nucleotídeo
AM AM.ATP AM.ADP.Pi AM*ADP AM.ADP AM
M M.ATP M.ADP.Pi M*ADP M.ADP M
ATP ATP
ATP
Pi
Pi
ADP
ADP
1a 3a 5a 7a 9a 1a
1d 3d 5d 7d 9d 1d
ATP

32
fecha, promovendo uma isomerização da molécula de miosina (C). Essa
conformação induzida pelo ATP sofre novas modific ações após a hidrólise em
ADP e Pi, onde a miosina se liga novamente à actina (D). Em seguida, ocorre a
abertura do sítio para a liberação de Pi e a subseqüente liberação de ADP (E)
(Chalovich, 1992; Sellers & Goodson, 1995).

33
Figura 8: Modelo da contração muscular. (A) A miosina se liga a actina em
estado de rigor. (B) A molécula de ATP se liga à cabeça da miosina e esta se
desliga da actina. (C) O ATP é hidrolisa do em ADP + Pi. (D) Após a hidró lise a
miosina se liga novamente a actina. (E) A saída de Pi gera uma mudança
conformacional na cabeça da miosina ( “power stroke”), e a miosina puxa o
filamento de actina (Sellers & Goodson, 1995).

34
1.6 - O Fenol
O fenol é um composto químico classificado como hidrocarboneto
aromático, constituído por um anel de benzeno e uma hidroxila (Fig. 9). Esse
composto é usado terapeuticamente , desde 1966, no controle de seqüelas
neurológicas. O fenol é um importante bloqueador neuromuscular. Esse
composto é administrado por micro-injeções em determinado ponto motor com
o objetivo de bloquear a ação muscular nesta região, promovendo um
fenômeno chamado de neurólise química, o que leva ao estado de relaxamento
muscular. O objetivo da aplicação clínica do fenol em músculos é controlar os
estímulos que promovem o estado de hipertonia muscular (Khalili & Betts,
1967).
Figura 9: Representação da estrutura química de uma molécula de fenol.

35
Há uma variedade de estudos reportados sobre os efeitos do fenol.
Halpern & Meelhuysen (1966) testaram o fenol em pacientes com
espasticidade, obtendo bons resultados no alívio do tônus espástico causado
pela hipertonia muscular. Eles utilizaram a técnica de aplicação percutânea
direta no ponto motor do músculo. Khalili & Betts (1967) também testaram o
fenol na terapia de pacientes espásticos com aplicação diretamente na fibra
nervosa e também obtiveram bons resultados. A utilidade do bloqueador como
tratamento mostrou-se muito interessante; porém, estudos histol ógicos
realizados em animais, utilizando o mesmo procedimento metodológico
descrito por Halpern & Meelhuysen em 1966, revelaram que o uso desta
técnica de aplicação não garante que a solução de fenol atinja única e
exclusivamente o local desejado (Halpern, 1977), expondo áreas adjacentes
aos efeitos deletérios do composto. Se aplicado diretamente em tecido
muscular poderia estar trazendo danos às fibras musculares e ,
conseqüentemente, às proteínas contráteis. Lee & Downing (1981) estudaram
os efeitos do fenol sobre a atividade contrátil de músculos cardíacos isolados.
Seus resultados demonstraram ação inotrópica negativa do fenol, que afeta o
mecanismo excitação-contração. Machado (2003) investigou os efeitos do fenol
em fibras descascadas, encontrando dimin uição da tensão isométrica máxima
quando aplicado diretamente no músculo, provocando um relaxamento
muscular irreversível, independente do efeito neurolítico , em concentrações
iguais ou superiores a 1% de fenol. Entretanto, concentrações inferiores não
apresentaram danos aparentes às fibras musculares, e demonstrou ainda, que
quando aplicadas na presença de concentrações submáximas de Ca +2, tensão
isométrica aumentou, o que poderia estar sendo promovido pela ativação da

36
actomiosina. A ação direta do fenol s obre as proteínas contráteis desperta
interesse, uma vez que seus efeitos na contração muscular em fibras
descascadas são ambíguos e dose -dependentes.
Para entender o mecanismo pelo qual o fenol poderia estar agindo sobre
as pontes cruzadas, nossa proposta foi estudar os efeitos desse solvente sobre
a região catalítica da miosina, o S1, analisando os efeitos deste composto
sobre a atividade ATPásica da miosina II, e sua correlação com a ligação à
actina.

37
2 – OBJETIVOS
Geral:
• Estudar o comportamento enzimático e estrutural da miosina na
presença de fenol
Específicos:
• Caracterizar os passos cinéticos que envolvem as etapas de ligação de
nucleotídeo, clivagem e liberação dos produtos da atividade catalítica do
subfragmento 1 (S1) da miosina II muscular na presença de fenol.
• Determinar se o fenol afeta a interação acto -S1
• Avaliar se este composto é capaz de alterar a estrutura terciária do S1

38
3 - MATERIAIS E MÉTODOS
3.1 - Miosina
A miosina é purificada do músculo longissimus dorsi de coelho, seguindo
o protocolo de Bremel & Weber (1975) com algumas modificações. Este
protocolo é baseado na diferença de solubilidade entre as proteínas contráteis.
Enquanto a actina é insolúvel em alta força iônica (600 mM) a miosina
permanece em solução.
O coelho é sacrificado por traumatismo cervical seguido de sangria. Os
músculos dorsais são expostos, esfriados no gelo por 30 min e só então
removidos. Os músculos são limpos (para remoção de vasos, gorduras, tecidos
conectivos e nervos), lavados com água Milli-Q gelada e triturados três vezes,
usando moedor elétrico previamente gelado. Todas as etapas subseqüentes
são realizadas a 4 oC. Para cada 100 g de músculo moído, são adicionados
300 mL de tampão de extração (0 ,3 M KCl e 0,15 M KH2PO4, pH 6,5), e a
suspensão é submetida a agitação lenta por 10 min. A seguir, adiciona -se
1.200 mL de água Milli-Q gelada, sob agitação vigorosa durante 15 min , e a
suspensão é filtrada em gaze. A este filtrado adiciona -se 1.800 mL de água
Milli-Q gelada, seguido por centrifugação (30.000 x g , 30 min, 4 oC). A redução
da força iônica (30 mM) seguida de centrifugação leva à precipitação da
miosina; enquanto que nesta força iônica a actina fica solúvel. O sobrenadante
então é descartado e o pellet é ressuspenso em alta força iônica (600 mM),
homogenizado é acrescido do mesmo volume de água Milli -Q gelada. A
suspensão é centrifugada (30.000 x g, 30 min, 4 oC), para a retirada de actina e
dos complexos regulatórios. Após a centrifugação é adicionada ao
sobrenadante água Milli-Q gelada, o que leva à redução da força iônica, e a

39
suspensão é novamente centrifugada. O ciclo de purificação é repetido duas
vezes para eliminar contaminantes, principalmente actina. A análise da pure za
da miosina é feita através de gel de poliacrilamida (SDS-PAGE) (Fig.10) onde
se encontra a MHC (cadeia pesada de miosina) e LCs (cadeias leves 1, 2 e 3).
Uma pequena quantidade da proteína C (135 kDa), componente do filamento
grosso e sem atividade ATPásica, é co -purificada com a miosina. Os pelle ts de
miosina são ressuspensos em tampão de estocagem (600 mM KCl, 5mM
EDTA, 50 mM HEPES, pH 7 ,0 e 50% glicerol (v/v) a 4 oC). Para utilizarmos a
proteína o glicerol é retirado através de diluição em água gelada (4 oC) na
relação 13:1 (v/v). Após a diluição a proteína é centrifugada a 2 .500 rpm por 20
min a 4 oC. O pellet (miosina) é ressuspenso em 1 ml de 600 mM KCl, 50 mM
HEPES, pH 7,0, a 4 oC. A concentração de miosina é aferida pelo método de
biureto (Gornall et al., 1949) contra curva padrão de albumina de soro bovino
(BSA).

40
Figura 10: Análise de pureza das preparações de miosina, S1 e actina .
Eletroforese em gel de poliacrilamida (12%) na presença de SDS corado por
comassie. (a) miosina 5 g, (b) miosina 10 g, (c) S1 5 g, (d) S1 10 g, (e)
actina 2,5 g, (f) actina 5 g. MHC (cadeia pesada da miosina 200 KDa), S1
( 110 KDa), LC1 (cadeia leve 1 25 KDa), LC2 (cadeia leve 2 20 KDa), LC3
(cadeia leve 3 16 KDa), actina ( 42 KDa).
3.2 – Subfragmento 1
O subfragmento 1 é obtido segundo o método de Weeds & Taylor
(1975). O último precipitado da preparação da miosina é ressuspendido em
tampão de baixa força iônica (120 mM NaCl, 20 mM fosf ato de sódio, 1 mM
EDTA e pH 7,0). Através da dosagem da miosina, a concentração é ajustada
para 20 mg/mL e equilibrada a temperatura ambiente. Para a clivagem da
miosina, 50 g/ml de -quimotripsina (a partir do estoque a 0,5 mg/mL em 1
mM de HCl) são adicionados à miosina sob agitação lenta. Após 10 min a
reação é interrompida com 300 M de PMSF (estoque 100 mM em 70% de
Actina
a b c d e f
MHC
S1
LC2
LC1
LC3

41
etanol) e colocada em gelo. A solução é dialisada por 12 h a 4 oC contra 25 vol
de imidazol 50 mM (pH 7,0) e DTT 1 mM. Após a diálise, centrifuga-se por 90
min a 120.000 x g para a separação do precipitado (que contém a cauda e o
material não digerido) e do sobrenadante (que contém o S1). O S1 é então
dosado pelo método de biureto ou pelo coeficiente de extinção molar,
assumindo o seguinte valor: S1 110 kDa, E1%280 = 7,5 (Muhlrad &
Chaussepied, 1990). Em seguida o sobrenadante é concentrado e submetido a
uma cromatografia de troca iônica e as frações coletadas são analisadas em
espectrofotômetro (λ = 280 nm). Em seguida a prote ína é dialisada contra um
tampão contendo 10 mM de Pi, 100 mM de KCl, 20 mM de imidazol (pH 7.0),
0,2 mM de DTT e 0.2 mM de EDTA. Após a diálise acrescenta -se à solução
glicerol 50% (v/v) e a proteína é aliquotada (0 ,4 mL) e estocada em nitrogênio
líquido.
Para uso experimental a proteína é submetida a uma nova diálise de 12
h contra um tampão contendo 50 mM de Hepes (pH 7,0) e 50 mM de KCl,
seguida de centrifugação a 350.000 x g, a 4 ºC por 22 min. Após o processo a
proteína é novamente dosada (E 1%280 = 7,5)
3.3 - Actina
Preparação do Pó-Cetona (pó cetônico)
A actina é purificada pelo método de Pardee & Spudich (1982), a partir
dos músculos peitorais maior e menor de galinha, que se mostrou mais estável
do que a actina de músculo de coelho. O primeiro dia fornece a actina na forma
de pó cetônico.

42
A galinha é sacrificada por decapitação e os músculos peitora is maior e
menor são removidos e esfriados no gelo. Após cerca de 10 min, os vasos e a
gordura são retirados e o músculo é moído por duas vezes em um moedor de
carne previamente gelado. Os passos seguintes sã o feitos a 4 oC com todas as
soluções geladas. O músculo é agitado durante 10 min em 300 mL (para cada
100 g de músculo) de solução de fosfato (100 mM KCl, 137 mM KH 2PO4, 63
mM K2HPO4, a pH 6,5). O filtrado é desprezado e ao resíduo é adicionado 50
mM NaHCO3 (600 mL) sob agitação por 10 min a 4 oC, para a retirada da
troponina, miosina e outros contaminantes. A suspensão é filtrada em gaze,
desprezando o filtrado. Acrescenta -se ao que ficou retido (para cada 100 g de
músculo) 300 ml de 1 mM Na 2EDTA, sob agitação, durante 10 min a 4 ºC.
Novamente a suspensão é filtrada em gaze, desprezando o filtrado;
acrescenta-se 6 volumes de água Milli -Q ao que ficou retido (para cada 100 g
de músculo, 600 ml), sob agitação durante 5 min a 4 °C. A mistura é filtrada em
gaze, desprezando o filtrado e o material retido em gaze sofre esta etapa
novamente. Para a estocagem é feita desidratação com acetona (para cada
100 g de músculo, 300 ml de acetona), durante 10 min , a temperatura
ambiente, sob agitação. A mistura é filtrada em gaze e o material retido sofre
nova etapa de desidratação. A etapa de desidratação com acetona é feita 5
vezes e o pó cetônico é estocado a 4 oC.
3.4 - Polimerização da Actina
Para cada 3 g de pó acrescentam-se 50 mL de tampão (2,5 mM Tris-
HCl, pH 8,0, 0,2 mM MgCl2, 0,2 mM Na2ATP e 0,5 mM DTT) sob agitação por
10 min, 4 oC. Centrifuga-se a 15.000 x g durante 10 min, 4 oC. Após

43
centrifugação filtra-se o sobrenadante em gaze e guarda -se o filtrado. Esse
processo é repetido três vezes a 4 oC, juntando-se ao final os três filtrados.
Acrescenta-se ao filtrado, 50 mM KCl, 1 mM Na 2ATP, 2 mM MgCl2 e 10 mM
Tris, pH 7,5, sob agitação durante 30 min a temperatura ambiente. Cen trifuga-
se em alíquotas de 25 mL a 100,000 x g durante 2 h. Despreza-se o
sobrenadante e a superfície do precipitado (F -actina) é lavada três vezes com
tampão (20 mM de imidazol, 6,5 mM de KCl, 5 mM de MgCl 2, 0,5 mM de
EGTA, 1,5 mM de NaN3, 2 mM de Na2ATP e 1 mM de DTT, a pH 7,0). Após as
lavagens acrescenta-se 1 mL do tampão utilizado na lavagem e o precipitado é
mantido no gelo por até duas semanas. Depois de incubado em gelo por 12 h,
cada pellet é cuidadosamente homogeneizado com 1 m L deste tampão. A
suspensão é mantida no gelo por no máximo cinco dias. A análise da pureza
da actina é feita através de gel de SDS-poliacrilamida.
3.5 - Eletroforese
A eletroforese em presença de SDS é feita segundo Laemmli (1970) e
de acordo com as modificações in troduzidas por Giulian et al. ( 1983).
3.6 – Atividades ATPásicas
As diferentes atividades ATPásicas da miosina são determinadas através
da dosagem de Pi liberado, segundo Taussky & Shorr (1953), durante o tempo
determinado de hidrólise de ATP. A proteína é pré -incubada no meio de reação
na ausência de nucleotídeo, por 2 min – 10 min (com e sem fenol) e as reações

44
são iniciadas pela adição de ATP. As reações são realizadas a 25 °C, na
presença ou na ausência de fenol nos meios de reação descritos na Tabela 1.
Tabela 1: Meios de reação utilizados para medidas da atividade ATPásica do
S1
Atividade ATPásica
Mg2+ATP Ca2+ATP K+EDTA-ATP Mg2+ATP
Actina-ativada
Hepes 50 mM 50 mM 50 mM 50 mM
KCl 50 mM 50 mM 600 mM 50 mM
MgCl2 5 mM - - 5 mM
CaCl2 - 5 mM - -
K+EDTA - - 5 mM -
S1 0,05-0,1 mg/ml 0,1 mg/ml 0,1 mg/ml 0,05-0,1 mg/ml
Actina - - - 0,4 mg/ml
Fenol 0 – 2% 0 – 0,5% 0 – 0,5% 0 – 2%
ATP 3 mM 3 mM 3 mM 3 mM
pH 7,0 7,0 7,0 7,0
Os meios de reação são preparados duas vezes concentrados para permitir a adição dosolvente.
Os tempos de hidrólise variam de acordo com o experimento, dentro da
faixa linear de hidrólise de ATP. As reações são inter rompidas com ácido
perclórico 0,1 N contendo carvão ativado segundo modificação do método de
Grubmeyer & Penefsky (1981). O carvão é utilizado para a absorção do ATP
não hidrolisado, do fenol e para des naturação ácida da enzima. Os brancos
são feitos acrescentando-se carvão ativado para a desnaturação de proteína, e

45
em seguida acrescenta-se o ATP. Após a desnaturação da proteína, as
amostras são mantidas no gelo. As amostras são centrifugadas a 2 .800 x g em
centrifuga refrigerada (Beckman GS -6R) a 4 °C por 20 min.
A concentração de fosfato inorgâni co é medida em alíquotas de 1 mL do
sobrenadante pelo método colo rimétrico (Taussky & Shorr, 1953) comparando
com curva padrão de Pi (que é repetida a cada expe riência). Todos os
experimentos cinéticos reportados neste trabalho foram realizados em
duplicata e foram repetidos no mínimo com três preparações diferentes de
proteína.
3.7 – Co-sedimentação
A co-sedimentação foi realizada segundo o método de DasGupta &
Reisler (1991) com algumas modificações. O ensaio de co -sedimentação foi
realizado de duas formas: (1) primeiramente o S1 (9 µM) foi incubado com F -
actina (18 µM) em temperatura ambiente , na ausência de ATP, por 10 min, em
tampão contendo 50 mM de KCl, 50 mM de Hepes (pH 7,0) e 5 mM MgCl2.
Após esse tempo adicionamos 0 ,25 – 0,5% de fenol por mais 10 min. (2)
Incubamos o S1 com 0,25 – 0,5% de fenol no mesmo tampão descrito
anteriormente em temperatura ambiente na ausência de ATP por 10 min. Em
seguida adicionamos F-actina (18 µM) por mais 10 min. Após a segunda
incubação as amostras de 200 µl são centrifugadas a 100.000 x g durante 30
min a 4 ºC. Os pellets são ressuspendidos no mesmo tampão (200 µl) descrito
acima. Para determinar a quantidade de S1 ligada à actina, amostras de cada
sobrenadante e precipitado são aplicadas em um gel desnaturante (12%
poliacrilamida). O gel é corado com Coomassie Blue e digitalizado (ScanJet)

46
para realização de densitometria (Sigma Gel – versão 1.1 Jandel Scient ific).
Através da análise densitométrica a relação molar de S1 ligado à actina é
determinada. A quantidade de S1 ligado ou não à actina foi calculada através
dos valores obtidos na densitometria, utilizando um padrão com concentração
conhecida de S1 e actina.
3.8 – Fluorescência no Estado Estacionário
Para os experimentos de fluorescência intrínseca do S1, foi utilizado um
espectrofotômetro Jasco 6300 (Jasco Inc., M.D., EUA) e cubeta de quartzo de
1 mL com caminho ótico de 1 cm. As amostras foram excitadas a 295 nm e a
emissão de fluorescência da proteína foi varrida de 300 a 400 nm, a
temperatura ambiente (~ 20 ° C). Os espectros são analisados pelo programa
do espectrofluorímetro (FL 100) ou transferidos e analisados no programa
Sigma Plot 9.0. O centro de massa espe ctral foi calculado pela equação
(Lakowicz, 1999):
Centro de massa (nm) = Σ (λ . Іλ)/ Σ Іλ
Onde : Σ = somatório
I = intensidade de fluorescência
λ = comprimento de onda
Todos os espectros obtidos das amostras contendo proteínas foram subtraídos
do valor do espectro contendo tampão na ausência (controle) ou na presença
de fenol.

47
3.9 – Cinética Rápida
Nos experimentos de cinética rápida utilizamos um fluorímetro de
stopped-flow (Applied Photophysics SX17MV), gentilmente cedido pelos Prof s.
Héctor Barrabin e Helena Scofano (Lab. de Membranas Transportadoras,
IBqM) (Fig. 11). Este equipamento nos permite acompanhar mudanças
conformacionais do S1, traduzidas pelo aumento ou diminuição da
fluorescência intrínseca do triptofano. Essas alterações são muito rápidas, na
escala de milisegundos, e são induzidas pela ligação e hidrólise de Mg2+ATP
(Fig. 12). O tempo morto do aparelho (“ dead time”) é ~ 1 ms, o que permite
monitorar a maioria das mudanças estruturais do S1 que ocorrem durante o
ciclo catalítico. O aparelho é regul ado para excitar em 295 nm a amostra que
chega à câmara de mistura e a temperatura é ajustada em 21 ºC por um banho
circulante acoplado ao aparelho. A proteína é colocada em uma seringa e o
ATP em outra; em um dado momento as duas soluções são misturadas e m
uma câmara que sofre incidência de luz por uma janela de 1 ,5 mm de entrada
e a emissão é detectada a 90° através de uma fenda de 3, 0 mm (Fig. 11).
Neste instante o aparelho capta a emissão da fluorescência do triptofano com
um filtro de 320 nm. A este si nal adquirido em função do tempo , o programa
ajusta uma exponencial simples, fornecendo a velocidade da mudança
conformacional e a amplitude do sinal.
Cada ponto dos experimentos é a média de cinco ou mais aquisições de
sinal. Cada experimento é repetido no mínimo três vezes, com preparações
diferentes de proteína.

48
Figura 11: Esquema representativo do fluorímetro de stopped-flow. As
soluções são colocadas nas respectivas seringas. A cada aquisição, são
ejetadas 50 μl de cada seringa simultâneamente à câmara de mistura. O
experimento é repetido no mínimo cinco vezes para cada condição , e os sinais
são coletados e ajustados matematicamente pelo software.
S1
ATP
Câmara de
Mistura
295 nm

49
Figura 12: Gráfico representativo do sinal coletado n o fluorímetro de
“stopped-flow”. As constantes cinéticas são obtidas pelo ajuste matemático
da curva por uma exponencial simples, no pró prio aparelho. Nesta condição
temos 2 μM de S1 com 15 μM ATP ( concentrações finais na câmara de
mistura). Kobs = 36,4 s-1. O aumento de fluorescência é de 23%.
Tempo (s)0,0 0,1 0,2 0,3 0,4 0,5
F
(AU
)

50
3.10 - Reagentes
O fenol, ATP, ADP, BeSO 4, AlCl3, e NaF, PMSF. α-quimiotripsina (Nº
C7762, type 1-S, 47 unidades/mg -1), membranas de diálise (retenção acima de
12 KDa) são adquiridos da Sigma Chemical Company. O fenol é preparado em
concentração estoque de 5%. O BeSO 4, o AlCl3 e o NaF foram gentilmente
cedidos pela Profa. Mônica Ferreira Moreira, do Instituto de Química - UFRJ.
Todas as soluções são feitas com água Milli -Q.
A resina DE-52 é adquirida da Whatman. O concentrador de proteína e a
membrana YM 30 (retenção acima de 30 KDa) utilizada no concentrador são
adquiridos da Amicon.
3.11 - Análise dos Dados
Todos os resultados são expressos como média + erro padrão do
número de experimentos com proteínas diferentes.
Quando necessário os resultados experimentais foram analisados
através do teste t de Student no programa Sigma Plot 9.0 (Jandel Scientific),
usando P <0,05 como critério para diferença significativa.

51
4 – RESULTADOS: Parte I
4.1 -Atividade ATPásica com diferentes cátions na presença de fenol.
Para estudarmos os efeitos do fenol no ciclo catalítico da miosina,
comparamos inicialmente a hidrólise do ATP pelo S 1 em diferentes
concentrações de fenol e cátions complexados ao ATP. Dados anteriores do
nosso laboratório (manuscrito em preparação) mostraram que o fenol
potencializa a atividade Mg2+ATPásica do S1. Entretanto, o fenol apresenta
efeitos diversificados dependendo do íon complexado ao ATP. Quando o íon
complexado ao ATP é o Mg 2+, o fenol em baixas concentrações (0 ,25 – 0,5%)
aumenta a atividade ATPásica (~6 e 11 vezes, respectivamente) quando
comparado à atividade obtida na ausência de fenol (Fig. 13). Qua ndo o íon
complexado ao ATP é o Ca 2+, o fenol apresenta um pequeno efeito
estatisticamente significativo somente na concentração de 0 ,5% (Fig. 14). A
atividade K+EDTA-ATPásica apresenta um efeito diferente das demais na
presença de fenol (Fig. 15). Nesta c ondição o fenol (0,25–0,5%) promove uma
inibição de 8 a 9 vezes em relação ao controle. Os resultados das atividades
específicas do S1 nas diferentes condições acima estão resumidos na Tabela
2.
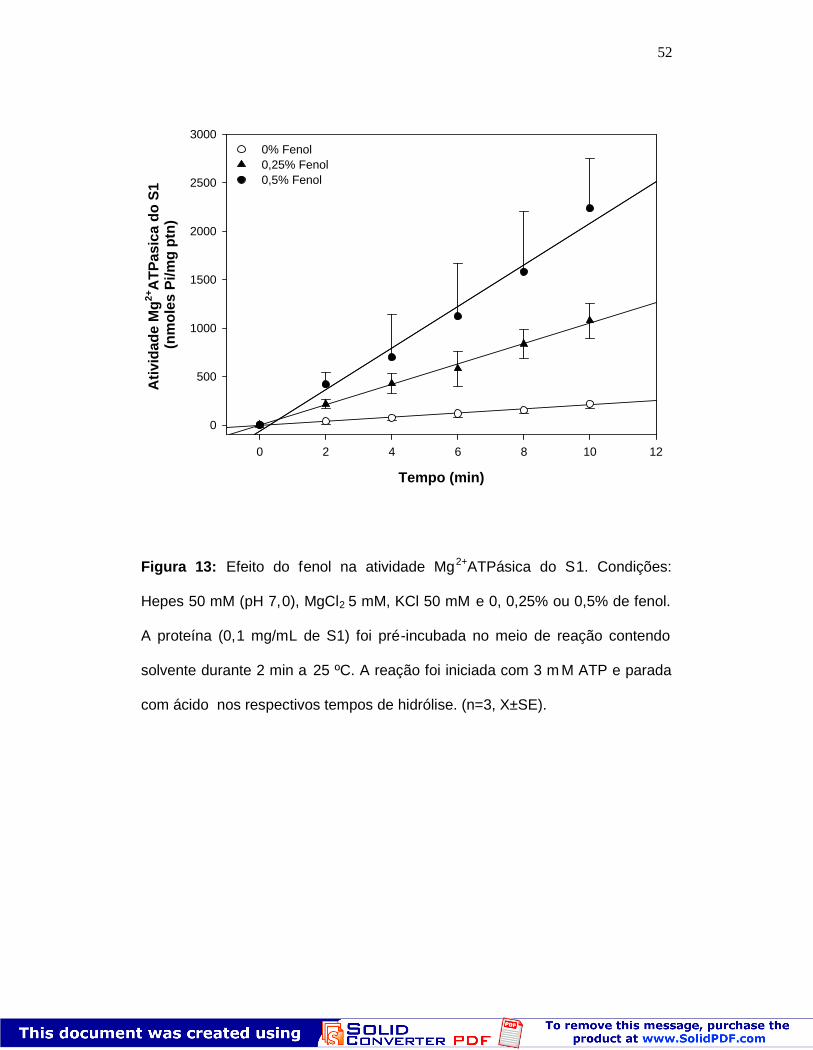
52
Tempo (min)
0 2 4 6 8 10 12
Ativ
idad
e M
g2+A
TPas
ica
do S
1(n
mol
es P
i/mg
ptn)
0
500
1000
1500
2000
2500
30000% Fenol0,25% Fenol0,5% Fenol
Figura 13: Efeito do fenol na atividade Mg2+ATPásica do S1. Condições:
Hepes 50 mM (pH 7,0), MgCl2 5 mM, KCl 50 mM e 0, 0,25% ou 0,5% de fenol.
A proteína (0,1 mg/mL de S1) foi pré-incubada no meio de reação contendo
solvente durante 2 min a 25 ºC. A reação foi iniciada com 3 m M ATP e parada
com ácido nos respectivos tempos de hidrólise. (n=3, X±SE).

53
Tempo (min)
0 2 4 6 8 10
Ativ
idad
e C
a2+A
TPas
ica
do
S1(n
mol
s Pi
/mg
ptn)
0
2000
4000
6000
8000
10000
0% Fenol0,25% Fenol0,5% Fenol
Figura 14: Efeito do fenol na atividade Ca 2+ATPásica do S1. Condições: Hepes
50 mM (pH 7,0), CaCl2 5 mM, KCl 50 mM e 0, 0,25% ou 0, 5% de fenol. A
proteína (0,1 mg/mL de S1) foi pré-incubada no meio de reação cont endo
solvente durante 2 min a 25 ºC. A reação foi iniciada com 3 mM ATP e parada
com ácido nos respectivos tempos de hidrólise. (n=3, X±SE).

54
Tempo (min)
0 2 4 6 8 10 12
Ativ
idad
e K
+ EDTA
ATP
asic
a do
S1
(nm
oles
Pi /
mg
ptn)
0
2000
4000
6000
80000% Fenol0,25% Fenol0,5% Fenol
Figura 15: Efeito do fenol na atividade K +(EDTA)-ATPásica do S1. Condições:
Hepes 50 mM (pH 7,0), KCl 600 mM, 5 mM K+EDTA e 0, 0,25% ou 0,5% de
fenol. A proteína (0,1 mg/mL de S1) foi pré-incubada no meio de reação
contendo solvente durante 2 min a 25 ºC. A reação foi iniciada com 3 mM ATP
e parada com ácido nos respectivos tempos de hidrólise. (n=3, X±SE).

55
Tabela 2: Efeitos do fenol sobre a atividade específica do S1 com diferentes
cátions complexados ao ATP. Valores obtidos da média pa ra um n = 3, ± SE. *
Teste t não pareado p < 0,05, para concentração de fenol em relação ao
controle. **Teste t pareado p < 0, 05, em relação ao controle.
Atividade Específica do S1 (nmol Pi.min -1. mg-1)
Fenol
0% 0,25% 0,5%
Mg2+ATPásica 19 4 111 12* 241 20*
Ca2+ATPásica 667 185 1075 278 1096 255**
K+EDTA-ATPásica 570 2 68 11* 31 6*

56
4.2 – Efeito do tempo de exposição do S1 ao fenol sobre a atividade
Mg2+ATPásica.
Os dados anteriores (Fig. 13) demonstram que o fenol promove uma
ação potencializadora sobre a atividade Mg 2+ATPásica do S1. Para avaliar se
este efeito é dependente da exposição da proteína ao fenol na ausência de
ATP, experimentos de pré-incubação do S1 com o fenol foram realizados.
Quando a proteína foi exposta ao fenol durante diferentes tempos de pré -
incubação, sem nucleotídeo, observamos que os efeitos do solvente são
dependentes do tempo de exposição e da concentraçã o (Fig. 16). Em
concentrações de 0,25 e 0,5%, o fenol manteve seu efeito estimulador sobre a
atividade ATPasica do S1. Com 1% de fenol, entretanto, a atividade
Mg2+ATPásica foi aumentada em 7 vezes, mas esse perfil de ativação se
perdeu a medida em que o tempo de pré-incubação da proteína com o solvente
foi aumentado e, em concentrações maiores (2%) o fenol inibiu a atividade
Mg2+ATPásica (Fig. 16).
A atividade enzimática do S1 é acentuada na presença de actina . Como
já é sabido, a interação do S1 com actina acelera o passo limitante do ciclo
catalítico do S1, o que aumenta a atividade ATPásica. Visando avaliar se a
presença de actina durante a pré -incubação do S1 com o fenol tem efeito
protetor contra as ações do fenol sobre o S1 observadas na Figura 16,
experimentos utilizando a atividade act ina-ativada foram realizados. Pode ser
observado na Figura 17 que na presença de actina os efeitos de ativação do
solvente são mantidos por mais tempo, mesmo com 1% de fenol. Contudo,
quando comparamos as atividades Mg 2+ATPásicas do S1 e a atividade

57
Mg2+ATPase actina-ativada percebemos que o efeito de ativação que o fenol
promove é menor na presença de actina. Na maior concentração de fenol
utilizada (2%), a actina não exerce nenhum efeito protetor contra a inibição
promovida pelo solvente (Fig. 17).
Tempo pré-incubação (min)
2 4 6 8 10
Ativ
idad
e M
g2+A
TPás
ica
do S
1(n
mol
es/m
g.m
in)
-200
0
200
400
600
800
1000
1200
1400
Figura 16: Efeito do tempo de exposição do S1 ao fenol sobre a atividade
Mg2+ATPasica do S1. (●) controle, (○) 0,25% de fenol, (▼) 0, 5% de fenol, (∆) 1
% de fenol e (■) 2 % de fenol. Condições: Hepes 50 mM (pH 7,0), MgCl2 5 mM,
KCl 50 mM. A proteína (0,1 mg/mL de S1) foi pré-incubada no meio de reação
contendo ou não solvente em d iferentes tempos (2-10 min) a 25 ºC. A reação
foi iniciada com 3 mM ATP e a hidrólise acompanhada por 2 min (n=3, X±SE).

58
Figura 17: Efeito do tempo de exposição do S1 ao fen ol sobre a atividade
actina ativada do S1. (●) controle, (○) 0,25% de fenol, (▼) 0, 5% de fenol, (∆) 1
% de fenol e (■) 2 % de fenol. Condições: Hepes 50 mM (pH 7,0), MgCl2 5 mM,
KCl 50 mM e 0, 0,25% ou 0,5% de fenol. O S1 (0,1 mg/m L) e actina (0,4
mg/mL) foram pré-incubados por 2 min no meio de reação. Em seguida foi
adicionada solvente e após diferentes tempos de pré-incubação (2-10 min) a 25
ºC, a reação foi iniciada com 3 mM ATP e a hidrólise acompanhada por 2 min
(n=3, X±SE).
Tempo de pré-incubação (min)
0 2 4 6 8 10 12
Ativ
idad
e M
g2+A
TPás
ica
Act
o-S1
(nm
oles
/mg.
min
)
0
200
400
600
800
1000
1200

59
Devido à redução na atividade Mg 2+ATPásica do S1 quando exposto ao
fenol por longos períodos de pré -incubação na ausência de ATP (Fig. 16),
experimentos foram realizados para avaliar se o fenol promove degradação
proteíca. O S1 foi exposto a diferentes tempos de pré -incubação com solvente,
e diferentes tempos de hidrólise. Cada condição testada foi analisada através
de um gel de poliacrilamida 12 %.
O fenol não apresentou nenhum efeito de degradação do S1 que
pudesse ser percebido por essa técnica, mesmo em tempos maiores que 10
min na ausência de ATP (Fig . 18).

60
Figura 18: Integridade do S1 na presença de fenol visto em SDS-PAGE
12%. Foram aplicadas amostras de ensaios com S1 (0,1 mg/ml) + fenol (0,5%)
após diferentes tempos de hidrólise de ATP (10 e 15 min). Os slots são
representados por: (1) S1+ 5 min pré -incubação, (2) S1+ 5 min pré-incubação +
10 min hidrólise, (3) S1+ 5 min pré -incubação + 15 min hidrólise, (4) S1+ 5 min
pré-incubação com fenol), (5) S1+ 5 min pré-incubação com fenol + 10 min
hidrólise, (6) S1+ 5 min pré-incubação com fenol + 15 min hidrólise, (7) S1+ 10
min pré-incubação, (8) S1+ 10 min pré -incubação + 10 min hidrólise, (9) S1+ 10
min pré-incubação + 15 min hidrólise, (10) S1 + 10 min pré -incubação com
fenol, (11) S1+ 10 min pré-incubação com fenol + 10 min hidrólise, (12) S1+ 10
min pré-incubação com fenol + 15 min hidrólise. Os experimentos foram feitos
em temperatura ambiente em tampão contendo Hepes 50 mM (pH 7,0), MgCl2
5 mM, KCl 50 mM e ATP 3 mM. Foi utilizado coloração por nitra to de prata
segundo Blum et al. (1987). O gel é representativo de 3 experimentos
independentes.
1 2 3 4 5 6 7 8 9 10 11 12
110 KDa

61
4.3 Experimentos de cinética rápida usando stopped -flow
4.3.1 Estados pré-hidrólise
Para tentar identificar as possíveis ações do fenol sobre o ciclo catalíti co
utilizamos uma outra abordagem, além da análise do Pi liberado na reação de
hidrólise pelo estado estacionário. O espectrofluorímetro de cinética rápida nos
permite acompanhar passos do ciclo relacionados à ligação e clivagem do ATP
e à liberação dos produtos, através da fluorescência intrínseca do triptofano
(Johnson & Taylor, 1978). A Figura 19 mostra a velocidade de ligação de ATP
na presença e na ausência de fenol a 21 °C. A velocidade máxima de aumento
de fluorescência do S1 em altas concentrações de ATP (Fig. 19) nos fornece a
velocidade de clivagem do ATP correspondente à constante K3 (k+3 + k-3)
(Tabela 3) e através da análise do início da curva, obtemos a constante de
velocidade de segunda ordem da ligação de ATP (K1k+2) (Fig. 20) segundo o
esquema 1:
Pode ser observado que a presença de fenol promove a redução tanto
do kobs máximo (Fig. 19) quanto do k obs entre as concentrações de 1 -4 µM de
ATP (Fig. 20). A ligação e a hidrólise de ATP na presença de fenol foram
analisadas utilizando a fluorescência intrínseca em função da concentração de
ATP. A constante cinética é registrada em cada concentração de ATP através
do ajuste de uma exponencial simples. Em baixas concentrações de ATP, é
M+ATP M.ATP M *.ATP M**.ADP.Pi M*.ADP M.ADP M
ADPPi
k2 k5K1 K3 k6k4

62
possível acompanhar o aumento inicial de fluorescência correspondente à
ligação de nucleotídeo e isomerização do S1 (etapas 1 e 2). Os dados
experimentais dessas constantes de velocidade medidas em baixas
concentrações de ATP foram ajustadas por regressão linear, e nos permitiram
calcular o valor de K1k+2 (inclinação da reta). A velocidade máxima de hidrólise
de ATP é fornecida pelo aumento máximo de fluorescência obtido no estado
M**ADP.Pi (etapa 3), e corresponde à constante de clivagem de ATP (k+3 + k-
3). As condições experimentais e os valores obtidos estão resumidos na Tabela
3. Os valores encontrados para o controle estão de acordo com os descritos na
literatura, os quais variam entre 1 e 9 x 10 6 M-1 s-1 para K1.k+2 assim como 60 a
150 s-1 para k+3 +k-3 (Johnson & Taylor, 1978; Milla r & Geeves, 1988; Shaw et
al., 2003). Pode ser observado na T abela 3 que a concentração de 0, 25% de
fenol já é suficiente para diminuir as velocidades de ligação e hidróli se do ATP
em aproximadamente 50 a 30% respectivamente .

63
(ATP µM)
0 75 150 225 300 375 450 525
k obs (
S-1)
-15
0
15
30
45
60
75
90
Controle0,25% Fenol
Figura 19: Efeito do fenol sobre a velocidade de ligação e hidrólise de ATP.
Cada ponto é a média de 3 experimentos, onde as constantes foram obtidas
por ajuste de uma exponencial simples (materiais e métodos) ao curso
temporal do aumento de fluorescência intrínseca do S1 (2 μM) induzida por
ATP. As amostras foram excitadas em 295 nm e a emissão coletada com um
filtro de 320 nm (X ± SE, n=3), 21 ºC. O tampão utilizado e os valores obtidos
estão listados na Tabela 3.
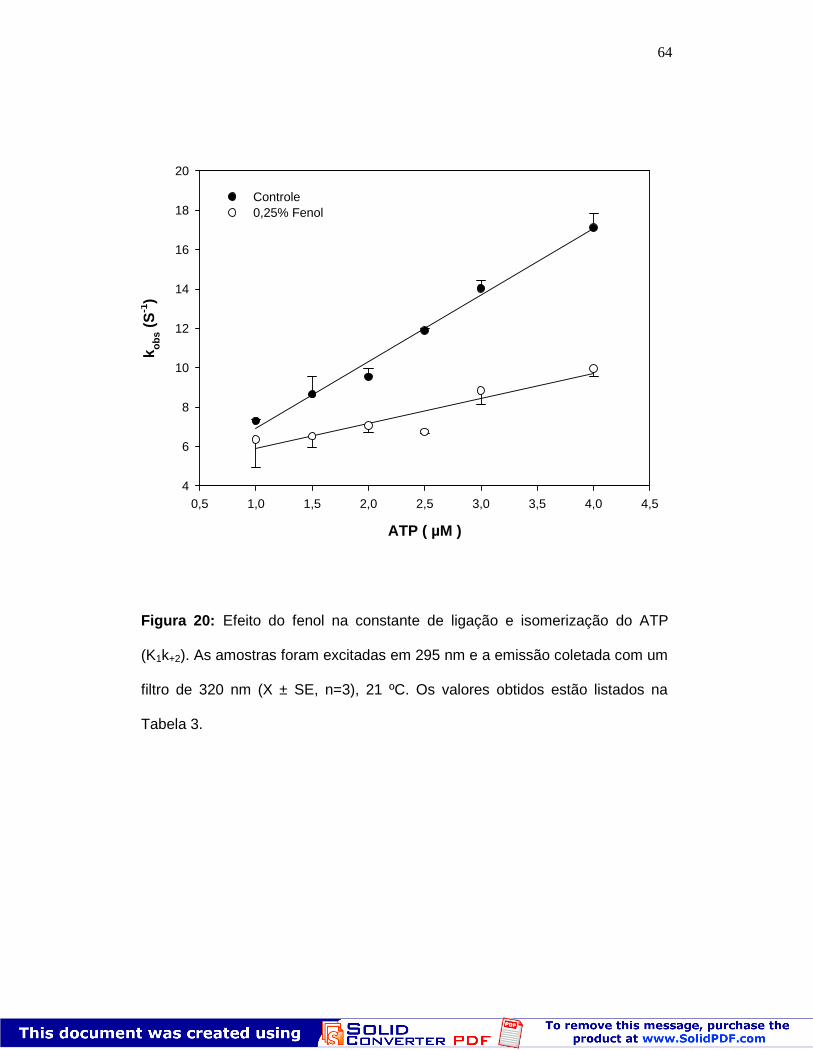
64
ATP ( µM )
0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 4,5
k obs (
S-1)
4
6
8
10
12
14
16
18
20
Controle0,25% Fenol
Figura 20: Efeito do fenol na constante de ligação e isomerização do ATP
(K1k+2). As amostras foram excitadas em 295 nm e a emissão coletada com um
filtro de 320 nm (X ± SE, n=3), 21 ºC. Os valores obtidos estão listados na
Tabela 3.

65
Tabela 3: As soluções utilizadas nos experimentos e os valores obtidos nas
constantes iniciais do ciclo catalítico (esquema 1). Todos os valores estão
descritos com médias e erro padrão adquirido de 3 -6 preparações protéicas
diferentes. *teste t pareado p < 0,05, para 0,25% de fenol em relação ao
controle.
Constantes Cinéticas
Controle 0.25% fenol
Hepes 50 mM 50 mM
KCl 50 mM 50 mM
MgCl2 5 mM 5 mM
S1 2 μM 2 μM
ATP 0.5-500 μM 0.5-500 μM
pH 7.0 7.0
K1.k+2 3.4 ± 0.3 μM-1 s-1 1.52 ± 0.4 μM-1 s-1 *
k+3 + k-3 72 ± 7 s-1 48 ± 2 s-1 *

66
4.3.2 Estados pós-hidrólise
Experimentos de single-turnover
Os dados anteriores mostram que o fenol foi capaz de alterar os passos
cinéticos iniciais do ciclo catalítico. Entretanto essa modulação não explic a o
efeito de ativação observado na atividade Mg 2+ATPásica. Pelo contrário,
embora a diminuição promovida pelo solvente na velocidade dos estados
iniciais do ciclo catalítico seja pequena (~ 30%), o efeito é antagônico ao visto
no estado estacionário. Sendo assim, passamos às etapas de liberação de
produtos. O primeiro produto da hidrólise do ATP a ser liberado é o Pi.
Experimentos de single-turnover foram realizados visando medir a velocidade
do passo de liberação de Pi (k +4, esquema 1). Este experimento é feito com
excesso de proteína em relação à concentração de ATP (2:1), acompanhando
a queda de fluorescência após o aumento transiente que reflete a formação de
M**ADP.Pi (Shaw et al., 2003).
Pode ser observado na Figura 21 que o fenol aumenta a velocida de de
liberação de Pi. Mesmo no controle, a liberação do Pi ocorre em duas fases:
uma fase rápida seguida de uma fase lenta (Fig. 21 A). As curvas foram
ajustadas e os valores obtidos para k obs são mostrados na Tabela 4. Os valores
encontrados para a fase rápida do controle se assemelham com o que é
descrito na literatura (0,055 s-1, 21 °C, pH 8,0) (Bagshaw & Trentham, 1974).
Além disso, a fase rápida corresponde a 40-50% da amplitude do sinal obtido
em todas as condições experimentais , tanto no controle quanto na presença de
fenol (Fig. 21B). Dessa forma, os valores obtidos para kobs na fase rápida (Fig.
21C) correspondem à liberação de Pi, sendo este passo catalítico o principal
responsável pelo efeito ativador promovido pelo fenol na atividade

67
Mg2+ATPásica. Todos os meios de reação experimentais e os valores das
constantes relacionadas à liberação dos produtos estão descritos na Tabela 4.
Tabela 4: Efeito do fenol sobre as etapas pós -hidrolise.
Constantes Cinéticas
Controle 0,25% fenol 0,5% fenol
Hepes 50 mM 50 mM 50 mM
KCl 50 mM 50 mM 50 mM
MgCl2 5 mM 5 mM 5 mM
S1 2 μM 2 μM 2 μM
ATP 1 μM 1 μM 1 μM
pH 7,0 7,0 7,0
kcat1 0,026 ± 0.005 s-1 0,15 ± 0.02 s-1 * -
kobs rápida 0,053 ± 0.004 s-1 0,33 ± 0.02 s-1 * 0,35 ± 0.02 s-1 *
kobs lenta 0,008 ± 0.0004 s-1 0,006 ± 0.0004 s-1 0,006 ± 0.0004 s-1
ATPa 400 μM 400 μM 400 μM
ADPa 20 μM 20 μM 20 μM
k-b 1,93 ± 0,6 s-1 5,57 ± 0,4 s-1 * 6,44 ± 0,5 s-1 *
Valores de kobs em pós-hidrólise do controle e na presença de fenol (esquema
1), bem como as condições experimentais. kobs rápida representa a constante de
liberação de Pi (k+4) e k-b representa a constante de liberação de ADP. *
Diferença estatística entre controle e testados com fenol (p < 0 ,05). a Condição
utilizada para os experimentos de deslocamento de ADP. Todos os valores
estão descritos com médias e erro padrão adquirido de 3 -6 preparações
protéicas diferentes. 1 Calculado a partir dos valores em estado estacionário da
Figura 13, corrigidos para 21ºC (Barouch & Moss, 1971).
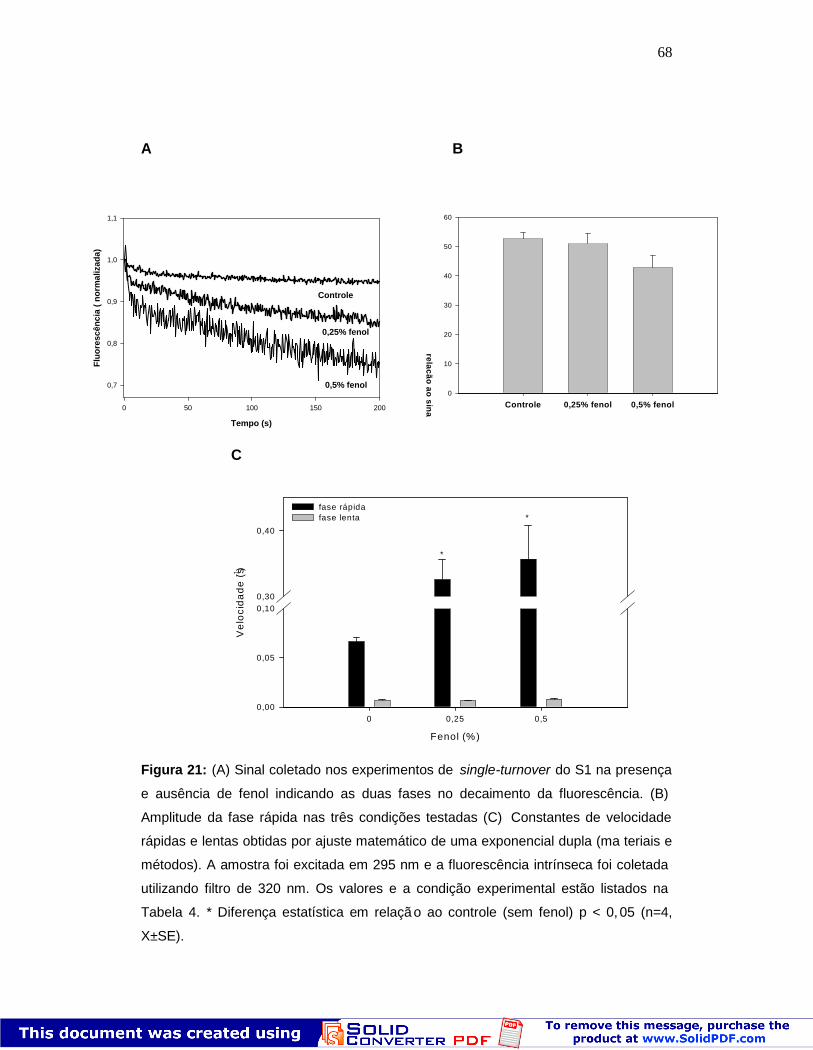
68
A B
Tempo (s)
0 50 100 150 200
Fluo
resc
ênci
a ( n
orm
aliz
ada)
0,7
0,8
0,9
1,0
1,1
Controle
0,25% fenol
0,5% fenol
Controle 0,25% fenol 0,5% fenolA
mplitude da fase rápida em
relação ao sinal total (%)
0
10
20
30
40
50
60
C
Fenol (%)
0 0,25 0,5
Vel
ocid
ade
(s-1)
0,00
0,05
0,100,30
0,40
fase rápidafase lenta *
*
Figura 21: (A) Sinal coletado nos experimentos de single-turnover do S1 na presença
e ausência de fenol indicando as duas fases no decaimento da fluorescência. (B)
Amplitude da fase rápida nas três condições testadas (C) Constantes de velocidade
rápidas e lentas obtidas por ajuste matemático de uma exponencial dupla (ma teriais e
métodos). A amostra foi excitada em 295 nm e a fluorescência intrínseca foi coletada
utilizando filtro de 320 nm. Os valores e a condição experimental estão listados na
Tabela 4. * Diferença estatística em relaçã o ao controle (sem fenol) p < 0, 05 (n=4,
X±SE).

69
Como observado na Figura 21 o fenol aumentou significativamente a
liberação de Pi. Entretanto, a constante que nos fornece essa informação é
adquirida com base na mudança de fluorescência intrínseca do S1. Como o
sinal coletado muda o seu perfil na presença de fenol , fomos investigar se o
fenol estaria realmente modulando a liberação de Pi ou se o solvente poderia
estar promovendo uma alteração no perfil de fluorescência. Como estratégia
para a nossa pergunta utilizamos um kit comercial (EnzCheck, da Molecular
Probes) contendo purina nucleosídeo fosforilase (PNP), a qual cataliza a
reação de fosforólise por Pi em 2-amino-6-mercapto-7-metilpurina ribosídeo
(MESG), gerando 2-amino-6-mercapto-7-metilpurina e ribose-1-fosfato (Figura
22). A análise é baseada na diferença na absorbância entre MESG e a base
purínica formada como produto (Webb, 1992).
Figura 22: Esquema representativo da reação do sistema MESG/PNP
(Webb, 1992)
PurinaNucleosídeoFosforilase Ribose
1 - fosfatoFosfato

70
Foi realizado um experimento de single-turnover onde as concentrações
de S1 e ATP eram equivalentes (1:1) juntamente com o sistema acoplado de
fosforilação MESG/PNP. Após a ligação do ATP ao S1 e posterior hidrólise, o
fosfato liberado no meio é utilizado no sistema MESG/PNP para formação de
ribose-1-fosfato (Fig. 22). O experimento foi realizado na aus ência e na
presença de fenol (0,5%) no meio e as constantes de velocidade foram
adquiridas acompanhando o aumento da absorbância do 2-amino-6-mercapto-
7-metilpurina à medida que ribose-1-fosfato fosse sendo formado. Dessa forma
adquirimos a constante de velocidade de liberação e consumo de Pi,
denominada de kapp.
Na Figura 23 pode ser observado que na presença de fenol há um
aumento na velocidade de formação de ribose -1-fosfato, sugerindo pelo valor
de kapp = 0,09 ± 0,002 s-1 maior em relação ao controle (kapp = 0,04 ± 0,003 s-1).
Este resultado confirma assim a ação do fenol sobre o passo de liberação de Pi
(k+4, esquema 1). Isso significa que o fenol induz uma liberação de Pi da
hidrólise de ATP pelo S1 de forma rápida, sendo o fosfato liberado utilizado
mais rapidamente pela PNP na formação de ribose -1-fosfato (Fig. 22). Como
mostrado na Figura 23, o fenol é capaz de induzir a uma maior velocidade de
formação de ribose-1-fosfato a partir do acoplamento dessas rea ções. Em
contra partida, o fenol não foi capaz de alterar a atividade da PNP quando
testada isoladamente do sistema S1 -ATP (não mostrado). Além disso, as
concentrações dos substratos do sistema MESG/PNP foram saturantes não
limitando a velocidade de formação de ribose-1-fosfato.
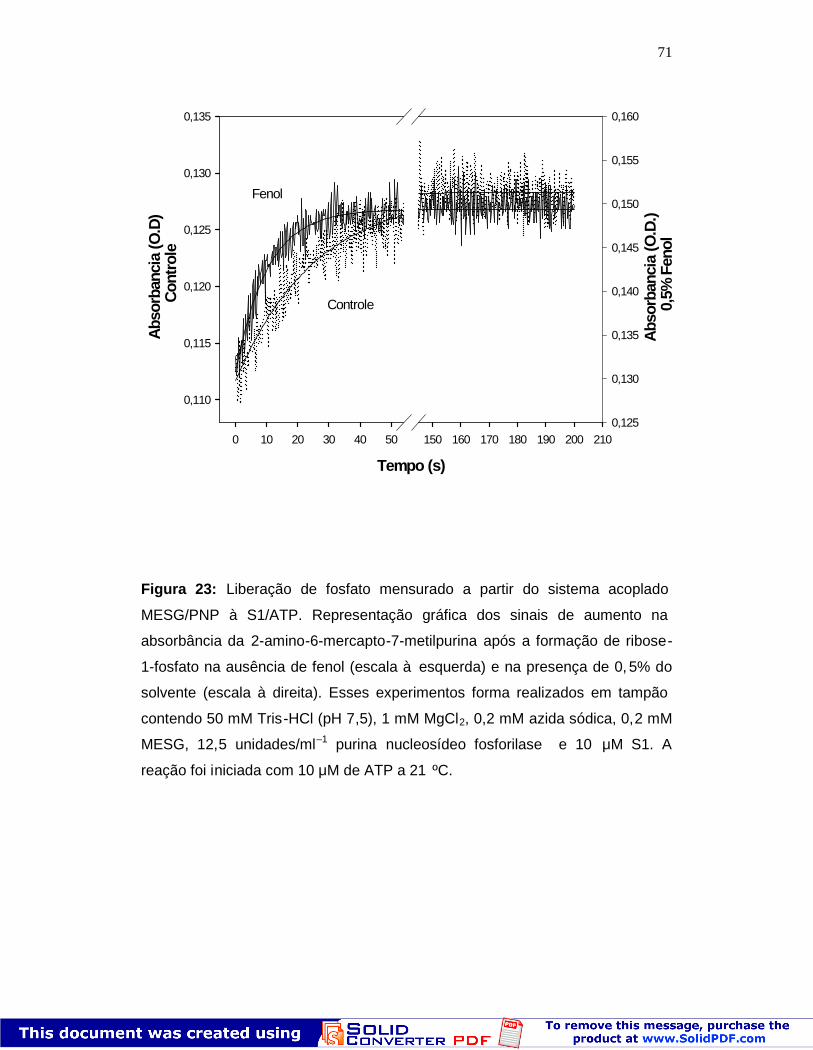
71
Tempo (s)
0 10 20 30 40 50 150 160 170 180 190 200 210
Abso
rban
cia
(O.D
)Co
ntro
le
0,110
0,115
0,120
0,125
0,130
0,135
Abso
rban
cia
(O.D
.)0,
5% F
enol
0,125
0,130
0,135
0,140
0,145
0,150
0,155
0,160
Fenol
Controle
Figura 23: Liberação de fosfato mensurado a partir do sistema acoplado
MESG/PNP à S1/ATP. Representação gráfica dos sinais de aumento na
absorbância da 2-amino-6-mercapto-7-metilpurina após a formação de ribose-
1-fosfato na ausência de fenol (escala à esquerda) e na presença de 0, 5% do
solvente (escala à direita). Esses experimentos forma realizados em tampão
contendo 50 mM Tris-HCl (pH 7,5), 1 mM MgCl2, 0,2 mM azida sódica, 0,2 mM
MESG, 12,5 unidades/ml–1 purina nucleosídeo fosforilase e 10 μM S1. A
reação foi iniciada com 10 μM de ATP a 21 ºC.

72
Experimentos de deslocamento de ADP
O próximo produto da hidrólise de ATP a ser liberado pelo ciclo catalítico
da miosina é o ADP. Para obter informações sobre a dis sociação da ADP nós
utilizamos um ensaio de deslocamento de ADP pelo ATP (Trentham et al.,
1972; Trentham et al., 1976). Para tal, o S1 foi incubado com ADP em uma
seringa e em seguida adicionamos um excesso de ATP. À medida em que o
ADP é liberado do sítio catalítico pelo S1, o ATP se liga à proteína. Como o
ATP possui uma afinidade maior pelo S1 comparado ao ADP, este se liga
preferencialmente ao S1. O processo foi acompanhado pelo aumento de
fluorescência induzido pela formação do complexo M**ADP.Pi, res ultado da
ligação e clivagem do ATP. A Figura 24 mostra o efeito no deslocamento do
ADP promovido pelo ATP em diferentes concentrações de fenol. Segundo
dados da literatura (Trentham et al., 1972; Trentham et al., 1976; Shaw et al.,
2003), a liberação de ADP ocorre em duas etapas, como podemos observar no
esquema 2:
A etapa limitante neste caso é k -b, que corresponde à mudança conformacional
de uma ligação mais forte entre a miosina e o ADP para uma ligação mais
fraca, com a posterior liberação de ADP para o meio. O valor obtido para k -b do
controle (Tabela 4) se assemelha com o que é descrito na literatura (1,7 s-1, pH
7,0 a 21 ºC) (Shaw et al., 2003; Kovacs et al., 2004). Como pode ser observado
na Figura 24 o deslocamento de ADP na presença de fenol é mais rápido. Este
M + A D Pk + a
k -a
M .A D P M *A D Pk + b
k -b
M + A D Pk + a
k -a
M .A D P M *A D Pk + b
k -b

73
dado indica que o fenol aumenta a constante de velocidade de liberação de
ADP do sítio catalítico.

74
A
Tempo (seg)
0 2 4 6 8 10 12
Intensidade de Fluorescência (U.A
.)
-0,1
0,0
0,1
0,2
0,3
0,4
Controle
FenolA
Fenol (%)0,0 0,2 0,4 0,6 0,8
k -b(s
-1)
2
4
6
8
10B
Figura 24: (A) Exemplo de um sinal obtido no deslocamento do ADP induzido
pelo ATP na ausência e na presença de fenol (0, 25%). Condições: 2 µM de S1,
20 µM de ADP, 50 mM Hepes (pH 7,0), 50 mM KCl, 5 mM de MgCl 2, e 400 µM
de ATP, temp. 21 ºC. A linha contínua mostra o ajuste de uma exponencia l
simples aos dados obtidos com kobs = 1,7 s-1 para controle e 5,2 s-1 para S1
tratado com fenol. (B) Deslocamento de ADP na presença de concent rações
crescentes de fenol (0-0,75%) (n=3, X ± SE).

75
5 – DISCUSSÃO: Parte I
Existem muitos trabalhos que estud am as propriedades enzimáticas da
miosina II, utilizando agentes perturbadores da estrutura da proteína (Klibanov,
1989; Salerno et al., 1997; Mariano et al., 2001). Este trabalho caracteriza o
fenol como um agente perturbador e estimulador da atividade Mg2+ATPásica do
subfragmento 1 da miosina II (Fig. 13). No entanto, o fenol apresenta algumas
particularidades. Da mesma forma que este composto apresenta um efeito
dose-dependente para a estimulação da atividade Mg2+ATPásica, o mesmo
efeito dose-resposta também é encontrado para a inibição da atividade
K+EDTA-ATPásica do S1 (Fig. 15).
Já é sabido que, dependendo do cátion complexado ao ATP, a proteína
pode assumir diferentes conformações, alterando o seu ciclo catalítico. Na
presença de Mg2+ ou Ca2+, o passo limitante do ciclo catalítico é a liberação de
Pi do sítio, sendo este passo mais rápido na presença de Ca 2+ do que na
presença de Mg2+ (Lymn & Taylor, 1970). Na atividade K+EDTA-ATPásica o
passo limitante não é a liberação de Pi , mas a clivagem do ATP (Lymn &
Taylor, 1970). Dessa forma, na presença de fenol, essa diferença entre as
atividades enzimáticas do S1 é mais acentuada.
Nossos resultados mostram que o fenol promove diferentes efeitos no
ciclo catalítico, dentre eles, o decréscimo na constante de velocidade da
hidrólise (k3 + k-3 - esquema 1) em aproximadamente ~ 30% (Fig. 19), e uma
redução na constante de ligação do ATP ao S1 (K1k2) ~ 50%, indicando que o
fenol inibe as etapas iniciais do ciclo catalítico (Fig. 20). Essa inibição poderia
explicar o efeito negativo observado na atividade K +EDTA-ATPásica, uma vez

76
que o passo limitante desta atividade é justamente a clivagem de ATP. Por
outro lado, na presença de Ca 2+ ou Mg2+, o complexo ternário M.ADP.Pi é
estabilizado, o que retarda a liberação de Pi (Lymn & Taylor, 1970). Entretanto,
o fenol possui diferentes efeitos para ambos cátions divalentes, indicando que
os efeitos promovidos pelo fenol são específicos para os complexos formados
na presença desses diferentes cátions complexados ao ATP. Essa
especificidade pode ser observada comparando o efeito do solvente nas
atividades Ca2+ e Mg2+ATPásicas, onde o fenol mostrou um pequeno efeito
sobre a atividade Ca2+ATPásica comparada com a atividade Mg 2+ATPásica, na
qual esse solvente aumentou mais de 400% a atividade (Fig. 13 e 14).
Salerno et al. (1997), utilizando 2,4-dinitrofenol (DNP), mostraram efeitos
similares aos encontrados aqui na presença de fenol. O DNP também é um
potente estimulador da atividade Mg 2+ATPásica e depressor da atividade
K+EDTA-ATPásica. Entretanto, o DNP promove um aumento na atividade
Ca2+ATPásica maior, diferentemente do encontrado com fenol, on de somente
na concentração de 0,5% de fenol houve um aumento significativo. Nós
acreditamos que as diferenças encontradas entre as ati vidades Mg2+ATPásicas
e Ca2+ATPásicas podem ser atribu ídas principalmente às diferenças
estruturais existentes entre os substratos . A afinidade pelo Mg2+ATP é muito
maior do que para Ca2+ATP ou K+ATP (Salerno, et al., 1997). A presença de K +
também promove uma diferença na estabilidade química do substrato K+ATP
em relação aos outros, além de poder se ligar a diferentes sítios da proteína ,
modificando sua conformação (Weeds & Taylor, 1975; Salerno, et al. 1997).
Todas essas evidências podem estar sendo potencializadas pelo solvente
tornando seus efeitos mais específicos.

77
Alguns autores têm descrito que as alterações na atividade enzimática
do S1 podem ser devidos à desnaturação protéica causada pela presença de
solventes orgânicos (Rayment, 1996; Komatsu e t al., 2004). No entanto,
Komatsu et al. (2004) mostraram um aumento na atividade Mg 2+ATPásica
promovida pela adição de álcool, independentemente de um processo
desnaturante. Em nossos dados, um longo tempo de exposição ao fenol ( ≥ 15
min) promove um decréscimo da atividade Mg 2+ATPásica do S1, e a
concentração de solvente utilizada é um fator determinante para este efeito
(Figs. 16 e 17).
Quando testamos as atividades ATPásicas em concentrações
crescentes de fenol ao longo de 10 min de pré -incubação na ausência de
nucleotídeo, observamos que concentrações menores (0,25 e 0,5%) do
solvente promovem um aumento da atividade que é independente do tempo
em que a proteína ficou exposta a ele (Fig. 16 e 17). Todavia, em altas
concentrações (1 e 2%) o efeito estimu lador promovido pelo fenol é perdido ou
a atividade é completamente inibida (Fig. 15 e 16). Esse dado é similar ao
descrito por Machado (2003), onde concentrações até 0, 8% de fenol
prejudicavam de forma irreversível o desenvolvimento da tensão isométrica
máxima em fibras musculares descascadas.
A análise quantitativa do efeito de cada um dos fatores que influenciam a
atividade enzimática é avaliada através do aumento ou redução da velocidade
da reação catalisada. Através dos experimentos de “cinética rápid a” podemos
obter as constantes cinéticas das etapas intermediárias do ciclo. As constantes
cinéticas encontradas para a etapa de ligação ( K1k2) de ATP são 3,4 μM-1s-1 no
controle e 1,5 μM-1s-1 na presença de 0,25% fenol (Fig. 19; Tabela 3). Além

78
disso, a constante k3 + k-3 , responsável pela clivagem do ATP, também sofre
um decréscimo de 72 ± 4 s -1 na ausência de fenol para 47,7 ± 2 s-1 na presença
de 0,25% de fenol (Fig. 19; Tabela 2). Esses resultados indicam que o fenol
altera os passos iniciais do ciclo catalítico de forma a retardar ou desacelerar
os estágios pré-hidrólise.
Embora o fenol altere os passos iniciais do ciclo catalítico, esses efeitos
são pequenos, e não explicam o efeito ativador observado no estado
estacionário para a atividade Mg2+ATPásica. Pelo contrário, chegam a ser
contraditórios. Entretanto, uma estimulação das etapas pós -hidrólise poderia
justificar o aumento observado, pois o passo limitante do ciclo catalítico na
atividade Mg2+ATPásica é justamente a liberação dos produtos (Bags haw &
Trentham, 1974). Sendo o fenol um ativador na faixa de concentração abaixo
de 0,8%, uma grande possibilidade seria o fenol estar alterando as etapas pós -
hidrólise do ciclo.
O Pi é primeiro produto da hidrólise do ATP a ser liberado do sítio. A
constante cinética que reflete a velocidade de liberação de Pi é k4. Na
presença de fenol, observamos uma grande diferença no perfil do sinal obtido
na medida de single-turnover em relação ao controle, corroborando a hipótese
de que o fenol poderia estar modul ando a atividade Mg2+ATPásica, acelerando
o passo limitante do ciclo catalítico (Fig. 21; Tabela 4). A utilização do sistema
acoplado MESG/PNP nos fornece um outro indício de que seja justamente na
liberação de produtos que o fenol exerça seu efeito ativador. O sistema
acoplado MESG/PNP, na presença de fenol, mostrou que a produção de
ribose-1-fosfato a partir do Pi no meio é mais acentuada. Isso ocorre devido a
uma maior disponibilidade de Pi no meio gerado pela atividade ATPásica do S1

79
(Figs. 22 e 23). Como K3 é dependente da concentração de M*ATP (Johnson &
Taylor, 1978), é esperado que esse passo catalítico sofra uma redução na
presença de fenol. No entanto, o aumento promovido pelo fenol em k4 é em
torno de 5 vezes, o que supera os efeitos inibitório s sobre as etapas iniciais do
ciclo.
Em adição ao efeito em k4, a liberação de ADP, o segundo produto a ser
liberado pelo ciclo, também é alterad a na presença de fenol. Os resultados
obtidos nos experimentos de deslocamento de ADP por ATP mostram que a
transição do estado M*ADP para M.ADP na presença do solvente ocorre de
forma mais rápida e conseqüentemente gera uma maior velocidade na
liberação de ADP pelo ciclo (Esquema 2 e Fig. 24). Esses dados indicam que o
principal efeito ativador desse solvente se encontra sobre as etapas pós-
hidrólise do ciclo catalítico.

80
6 – Resultados Parte II
6.1 - Efeito do Fenol na Estrutura Terciária do S1
Afim de observar o comportamento estrutural da proteína (S1) na
presença de fenol, utilizamos a fluorescência para analisar a estrutura terciária
do S1. Na Figura 25, são mostrados os espectros de fluorescência intrínseca
do S1 na presença de concentrações crescentes de fenol. Na presença do
solvente, tanto a intensidade de emissão quanto o perfil do pico de emiss ão
são alterados. O fenol promove um decréscimo na intensidade de emissão de
fluorescência intrínseca do S1 e desloca o seu pico de emissão para
comprimentos de onda maiores, correspondentes à região do vermelho (Fig.
25). Estes resultados indicam que os r esíduos de triptofanos do S1 ficam mais
expostos à água com adição de fenol, característico de uma conformação
menos enovelada. Isso fica mais evidente quando analisamos o centro de
massa espectral (materiais e métodos) na presença e na ausência de fenol.
Nessa análise o fenol promove um deslocamento no centro de massa de ~ 3
nm para regiões de menor energia (Fig. 26). Cabe ressaltar que na pre sença
de 0,25 e 0,5% do solvente, a atividade Mg 2+ATPásica encontra-se muito
ativada (Fig. 13), sugerindo que as al terações promovidas pelo fenol na
estrutura do S1 ocorrem sem causar um completo desenovelamento.

81
Figura 25: Espectros típicos de emissão de fluorescência do S1 (0, 05 mg/ml)
na ausência e na presença de fenol. Os experimentos foram realizados em
tampão contendo 50 mM Hepes (pH 7,0), 50 mM KCl, 5 mM MgCl 2 e 0,25 e
0,5% fenol. Os experimentos foram realizados à temperatura ambiente e a
solução de fenol foi adicionada à proteína segundos antes da leitura espectral.
Comprimento de onda (nm)
280 300 320 340 360 380 400 420
Inte
nsid
ade
Fluo
resc
enci
a (U
.A.)
-20
0
20
40
60
80
100
120
140
S1S1 + 0,25% FenolS1 + 0,5% Fenol

82
Figura 26: Centro de massa do espectro de fluorescência intrínseca de S1, na
ausência e presença de concentrações crescentes de fenol (0, 25 - 1%). Os
experimentos foram realizados em tampão contendo 50 mM Hepes (pH 7,0), 50
mM KCl, 5 mM MgCl2 e 0,25 ou 0,5 ou 1% fenol, à temperatura ambiente.
* teste t para p < 0,05 em relação ao controle (n=3, X±SE).
Cen
ter o
f mas
s (n
m)
Fenol (%)
0 0,25 0,5 1
Cen
tro
de m
assa
(nm
)
340
342
344
346
348
350
352
* * *

83
Como podemos observar nas atividades ATPásicas (Figs. 16 e 17), o
tempo de exposição da proteína e a concentração do solvente são fatores
importantes no efeito do fenol sobre o S1 . Diante disso, fomos verificar se o
efeito promovido pelo fenol na estrutura do S1 se mantinha de acordo com o
tempo de análise, uma vez que em ensaios de atividade ATPásica, a proteína
fica exposta ao solvente por períodos longos (2-15 min). Submetemos o S1 a
0,25% de fenol e acompanhamos o perfil de fluorescência e o centro de massa,
de 60 até 600 s na presença de solvente (Fig. 27). Podemos observar que o
efeito do solvente sobre o espectro de fluorescência da proteína perdura por
todo tempo de análise, deslocando o centro de massa para regiões do
vermelho. Isso sugere que os efeitos estruturais provocados pelo fenol no S1
contribuam para o aumento da atividade Mg 2+ATPásica.
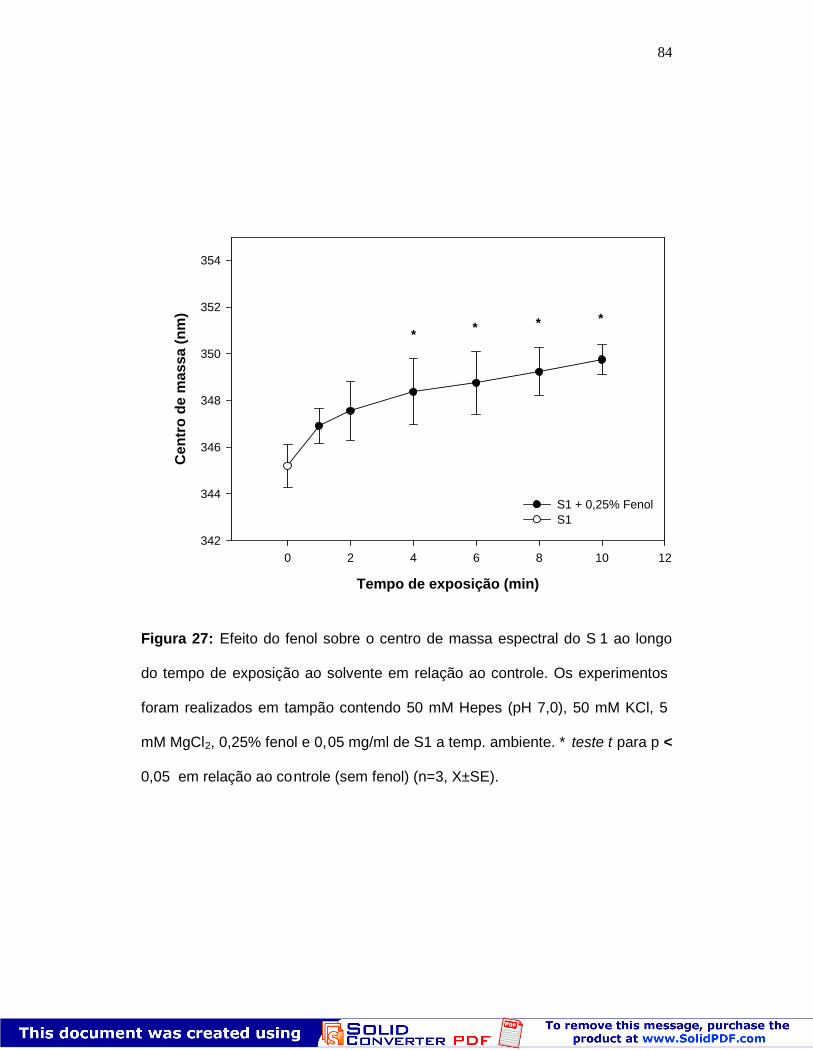
84
Figura 27: Efeito do fenol sobre o centro de massa espectral do S 1 ao longo
do tempo de exposição ao solvente em relação ao controle. Os experimentos
foram realizados em tampão contendo 50 mM Hepes (pH 7,0), 50 mM KCl, 5
mM MgCl2, 0,25% fenol e 0,05 mg/ml de S1 a temp. ambiente. * teste t para p <
0,05 em relação ao controle (sem fenol) (n=3, X±SE).
Tempo de exposição (min)
0 2 4 6 8 10 12
Cen
tro
de m
assa
(nm
)
342
344
346
348
350
352
354
S1 + 0,25% FenolS1
* * * *

85
6.2 - Efeito do Fenol na Estrutura Terciária do S1 na Presença de
Análogos de ATP
A fim de quantificar o comportamento estrutural do S1 na presença de
nucleotídeo e fenol, submetemos o S1 às mesmas condições descritas
anteriormente, mas na presença de análogos de ATP. É sabido que o ATP é
capaz de induzir um aumento de fluorescência em conseqüência da sua
ligação à proteína. Entretanto, a ligação de ATP e a sua posterior hidrólise
ocorrem de forma muito rápida, o que dificulta a análise espectral.
Como estratégia para acompanhar as mudanças estruturais sofridas
pelo S1 durante o ciclo catalítico, utilizamos o fluoreto de berílio (BeF 3-) e o
tetrafluor aluminato (AlF4-). Estes compostos “complexados” ao ADP,
pertencem a uma classe de análogos (Robinson et al., 1986; Fisher et al.,
1995) que mimetizam os estados M*ATP e M**ADP.Pi (esquema 1),
respectivamente. Os complexos se ligam fortemente ao sítio catalítico da
proteína, e uma vez ligados, não são hidrolisáveis como ATP (Robinson et al.,
1986; Fisher et al., 1995), mas promove m mudanças conformacionais
semelhantes às induzidas por ATP. Maruta et al. (1998) mostraram que
complexos formados a partir de AlF 4- e BeF3
- em HMM são potentes inibidores
da atividade HMM-ATPásica tanto em músculo esquelético quanto em músculo
liso, tornando os complexos ternários inativos e portanto inibindo a atividade
enzimática. A ligação de BeF 3- e AlF4
- ao S1 é mediada pela presença de
fluoreto de sódio (NaF) e ADP, formando os complexos M.ADP .BeF3-,
M.ADP.AlF4- (Fisher et al., 1995; Phan et al., 1997; Maruta et al., 1998) . A
Figura 28 mostra um esquema estrutural do S1 complexado aos análogos e
suas respectivas modificações conformacionais.

86
Figura 28: Esquema representativo das mudanças conformacionais na região
globular da miosina (S1), nos estados de transição S1 *ATP seguido pelo
S1.**ADP.Pi (A e B respectivamente). Neste modelo, as mudanças
conformacionais estão particularmente relacionadas ao movimento da porção
inferior do fragmento de 50 kDa em resposta à ligação de ATP e ADP.Pi. O
sítio de ligação de nucleotídeo se encontra fechado (A e B) (Adaptado de
Fisher et al., 1995).

87
Para verificar se o fenol modula a estrutura do S1 no estado M*ATP,
incubamos o S1 com ADP previamente (2 min) e adicionamos 5 mM de NaF e
2 μM de BeSO4- por mais 3 min para formação do complexo, e o analisamos na
presença e na ausência de fenol (0, 25 – 1%). O mesmo procedimento foi
realizado no ensaio para a formação do complexo M**A DP.Pi utilizando ADP e
AlF4- .
A Figura 29 mostra que, diferentemente do que ocorre na estrutura do
S1 na ausência de nucleotídeo, na presença desses complexos o fenol não foi
capaz de alterar significativamente a estrutura terciária do S1.
6.3 - Efeito do Fenol na Estrutura Terciária do S1 na Presença de ADP.
O ADP também se liga ao sítio catalítico do S1, induzindo a uma
mudança conformacional que pode ser acompanhada por um aumento na
fluorescência intrínseca do S1. Entretanto, esse aumento de flu orescência é
bem menor comparado ao aumento de fluorescência induzido pela ligação de
ATP. Para verificar a estrutura do estado M*ADP, nós incubamos o S1 com
ADP na presença e na ausência de difer entes concentrações de fenol (0, 25 –
1%). Nesse estado, ao contrário dos estados analisados anteriormente, a
proteína encontra-se em um estado transitório de abertura do sítio catalítico , e
esta conformação permite uma exposição parcial do sítio catalítico ao meio
aquoso (Geeves & Holmes, 1999). Analisando o perfil de fluorescência e o
centro de massa na presença de fenol (Figs. 30 e 31), também se observa um
importante decréscimo da fluorescência e o deslocamento do pico de emissão
para regiões de menor energia. Este resultado foi similar ao encontrado para o
S1 na ausência de nucleotídeo (Fig. 25 e 26). Es tes dados sugerem que a ação

88
do fenol sobre a estrutura terciária do S1 seria mais pronunciada nos estados
conformacionais onde o sítio de interação com o nucleotídeo encontra -se
exposto ou parcialmente exposto. D essa forma, o fenol estaria modulando a
estrutura terciária do S1, favorecendo a saída de Pi e A DP do sítio, como
evidenciado pelos resultados das constantes cinéticas obtidas para liberação
dos produtos (Tabela 4).

89
Fenol (%)
0 0,25 0,5 1
Cen
tro
de m
assa
(nm
)
334
336
338
340
342
344
346
348 S1*ATPS1**.ADP.Pi
Figura 29: Centro de massa dos espectros de fluorescência intrínseca do S1
(0.05 mg/ml) nos estados que mimetizam S1*ATP (M.ADP.BeF 3-) e S1**ADP.Pi
(S1.ADP.AlF4-), na presença e na ausência de fenol (0, 25 – 1%). Os
experimentos foram realizados em tampão contendo 50 mM Hepes (pH 7,0), 50
mM KCl, e 5 mM MgCl2. A proteína (0,05 mg/ml) foi previamente incubada com
2 mM de ADP (2 min), seguida pela adição de 5 mM de NaF e 2 µM de BeSO 4-
ou 150 µM de AlCl3 por mais 3 min antes da aná lise. Após a formação do
complexo era adicionado 0,25 -1% fenol, segundos antes da leitura do espectro
à temperatura ambiente (n=4; X±S.E.).

90
Figura 30: Espectros típicos de emissão da fluorescência intrínseca do S1
complexado ao ADP formando M*ADP na presença e na ausência de fenol
(0,25 - 1%). Os experimentos foram realizados em tampão contendo 50 mM
Hepes (pH 7,0), 50 mM KCl, 5 mM MgCl 2, e 0,25 ou 0,5 ou 1% de fenol, 0,05
mg/mL de S1 e 2 mM ADP a temperatura ambiente.
Comprimento de Onda (nm)
280 300 320 340 360 380 400 420
Inte
nsid
ade
de F
luor
escê
ncia
(U.A
.)
-20
0
20
40
60
80
100
S1
S1.ADP
S1.ADP + 0.25% fenol
S1.ADP + 1% fenol
S1.ADP + 0.5% fenol

91
Fenol (%)
0 0,25 0,5 1
Cen
tro
de m
assa
(nm
)
342
344
346
348
350
S1S1.ADP *
**
* *
**
**
Figura 31: Efeito do fenol (0,25 - 1%) sobre o centro de massa do S1 e da
S1.ADP. Os experimentos foram realizados em tampão contendo 50 mM
Hepes (pH 7,0), 50 mM KCl, 5 mM MgCl 2, 0,05 mg/ml de S1 e 2 mM ADP. *
teste t para p 0,05 em relação ao controle sem fenol; ** teste t para p 0,05
em relação ao controle sem fenol, mas com ADP, a temperatura ambiente
(n=5, X±SE).

92
6.4 – Modificação Estrutural do S1 pelo Fenol Altera a Ligação com Actina
Ao analisar a estrutura cristalografica do S 1 (Fig. 4), podemos observar
que o sítio de ligação para o nucleotíd eo está localizado entre as porções de 25
kDa e 50 kDa. Entre a porção de 20 kDa e 50 kDa encontra -se o sítio de
ligação para actina. Pode ser observado também (Fig. 4) que o sítio de liga ção
e hidrólise de ATP e o sítio de ligação de actina são geometricamente opostos.
Essa disposição espacial permite uma relativa “comunicação” entre esses
sítios. Isso fica bem evidente quando analisamos o ciclo catalítico (Fig. 7), e
verificamos que as mudanças estruturais promovidas pela ligação do ATP ao
complexo A.M geram uma perda da afinidade da miosina pela actina,
conhecida como ponte fraca, levando à dissociação do complexo A ~M.ATP.
Por outro lado, a re-associação da actina ao complexo ternário M.A DP.Pi
também modifica o ambiente estrutural, favorecendo a abertura do sítio
catalítico e a liberação dos produtos da hidrólise (Lymn & Taylor, 1971; Geeves
& Holmes, 1999) (Fig. 7).
Os resultados anteriores mostram que o fenol é capaz de alterar a
estrutura terciária do S1 na ausência de nucleotídeo e na presença de ADP.
Esses estados são fortemente influenciados pela ligação com actina, formando
o complexo de rigor (A.M) e/ou formando ponte forte com o complexo M*.ADP,
gerando A.M*.ADP. Para verificar se o fenol, uma vez modulando esses
estados intermediários, afetaria a ligação da actina, utilizamos um ensaio de
co-sedimentação tanto no rigor (A.M) quanto em ponte forte (A.M*.ADP). A co -
sedimentação é baseada na diferença de solubilidade entre o S1 e a a ctina e
foi realizada em duas etapas de incubação: (1) Primeiramente as proteínas

93
(actina e S1) são incubadas por 10 min seguido da adição de fenol e (2) Nós
incubamos o S1 com fenol por 10 min e em seguida adicionamos actina. As
amostras então são centrif ugadas, e o S1 livre, pela sua solubilidade em baixa
força iônica, permanece no sobrenadante. A actina, em sua forma filamentosa,
possui baixa solubilidade e sedimenta formando um precipitado. Será
encontrado no pellet o S1 que se ligar à actina no período de pré-incubação ou
durante a centrifugação, formando o complexo de rigor ou ponte forte. As
Figuras 32 e 33 mostram que o fenol (0 ,5%) quando incubado previamente
com S1, altera a ligação de actina. Entretanto, quando o rigor é formado
previamente o fenol não mostra o mesmo efeito. Isso indica que durante o
período de pré-incubação com o solvente, o S1 sofre modificações estruturais
provocadas pelo fenol que diminuem a interação acto -S1. O mesmo efeito não
é observado na presença de ADP, sugerindo que as modificações estruturais
nos estados S1 e S1.ADP são diferentes.
6.5 – Reversibilidade dos Efeitos do Fenol
O efeito do fenol na diminuição da tensão isométrica máxima em fibras
musculares descascadas é irreversível em concentrações acima de 0,8%
(Machado, 2003). Entretanto, a atividade Mg 2+ATPásica do S1, medida com a
proteína em solução, é aumentada em concentrações abaixo de 0,8%, como
mostrado na Figura 13. Contudo, ao aumentarmos a concentração para 1 e 2%
de fenol esse efeito estimulador é perdido (Fig. 16).
Para verificar se o efeito do fenol poderia ser reversível, pré -incubamos
0,5 mg/ml de S1 com fenol 0 ,5% em meio de reação durante 2 min a 30 ºC.

94
Logo após retiramos uma alíquota dessa solução e transferimos para um outro
tubo onde a concentração tanto de proteína quanto de fenol foi diluída 10
vezes. Após a diluição a concentração final de fenol foi reduzida para 0,05%.
Em seguida verificamos a atividade ATPásica da amostra diluída. Como
mostrado na Figura 34, o efeito do fenol em solução pode ser revertido
completamente, tanto para a atividade Mg2+ATPásica quanto para a atividade
K+EDTA ATPásica (Fig. 35). É válido ressaltar que, o fenol altera
significativamente ambas as atividades, estimulando na presença de Mg 2+ e
inibindo na presença de K +EDTA, e ambos os efeitos foram abolidos. Es te
resultado mostra que o efeito do fenol sobre a atividade enzimática é passível
de reversão.

95
Figura 32: Ensaios de co-sedimentação em estados de rigor (A.S1) e ponte
forte (A.S1.ADP) na presença de fenol (0,25% e 0,5%). Os números
representam as amostras de sobrenadante e precipitados onde (1) A.S1; (2)
A.S1 pré-incubado por 10 min + 0,25% fenol; (3) A.S1 pré -incubado por 10 min
+ 0,5% fenol; (4) S1.Fenol (0,25%) pré -incubado por 10 min + Actina; (5)
A.S1.Fenol (0,5%) pré-incubado por 10 min + Actina; (6) A.S1.ADP; (7)
A.S1.ADP pré-incubado por 10 min + 0,25% fenol; (8) A.S1.ADP pré -incubado
por 10 min + 0,5% fenol; (9) S1.ADP.Fenol (0,25%) pré -incubado por 10 min +
Actina; (10) S1.ADP.Fenol (0,5%) pré -incubado por 10 min + Actina. O gel
usado era desnaturante com SDS-poliacrilamida 12%.
Sobrenan
Pellet
S1
110 kDakDa
42 kDa
110 kDakDaAct
Rigor
Ponte Forteforte
1 2 3 4 5 6 7 8 9 10
1 2 3 4 5 6 7 8 9 10
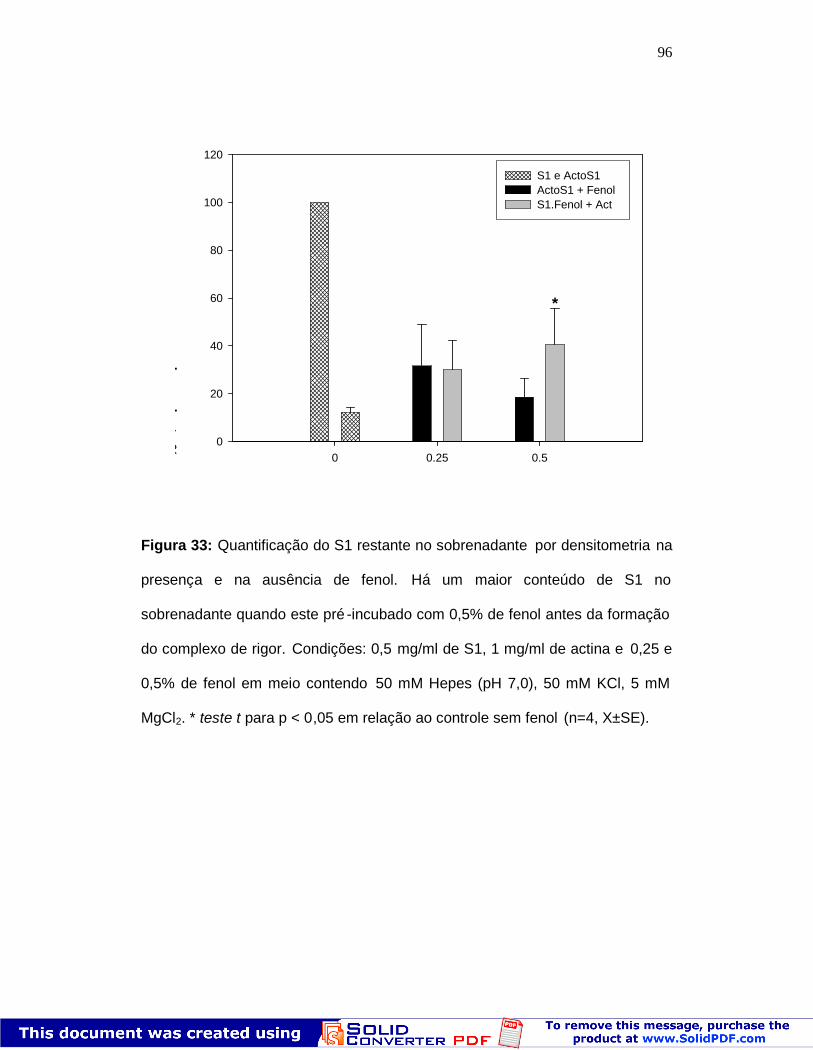
96
Figura 33: Quantificação do S1 restante no sobrenadante por densitometria na
presença e na ausência de fenol. Há um maior conteúdo de S1 no
sobrenadante quando este pré -incubado com 0,5% de fenol antes da formação
do complexo de rigor. Condições: 0,5 mg/ml de S1, 1 mg/ml de actina e 0,25 e
0,5% de fenol em meio contendo 50 mM Hepes (pH 7,0), 50 mM KCl, 5 mM
MgCl2. * teste t para p < 0,05 em relação ao controle sem fenol (n=4, X±SE).
0 0.25 0.5
sobrenadante % 0
20
40
60
80
100
120
S1 e ActoS1ActoS1 + FenolS1.Fenol + Act
*

97
Figura 34: Reversibilidade do efeito do fenol na atividade Mg 2+ATPásica.
Condições: 0,5 mg/mL de S1 foi previamente incubada com 0, 5% de fenol
durante 2 min em meio de reação contendo 50 mM Hepes (pH 7,0), 50 mM
KCl, 5 mM MgCl2 a 25 ºC. Após esse período a proteína e o fenol foram
diluídos para 0,05 mg/ml e 0,05%, respectivamente. A essa nova so lução é
adicionada 3 mM de ATP. As curvas controle (0% de fenol) e controle positivo
para 0,05% de fenol, foram realizados com 0, 05 mg/mL de S1 mantendo o
tempo de pré-incubação de 2 min a 25 °C antes da adição de ATP (n=3,
X±SE).
Tempo (min)
0 2 4 6 8 10 12
nmol
s Pi
.mg
ptn
0
1000
2000
3000
4000
5000
6000ControleControle 0,05% fenol0,5% de fenolReversão

98
Tempo (min)
0 2 4 6 8 10 12
nmols Pi.m
g ptn
0
2000
4000
6000
8000
10000
12000ControleControle 0,05% fenol0,5% fenolReversão
Figura 35: Reversibilidade do efeito do fenol na atividade K+EDTA ATPásica.
Condições: 0,5 mg/mL de S1 foi previamente incubada com 0.5% de fenol
durante 2 min em meio de reação contendo 50 mM Hepes (pH 7,0), 600 mM
KCl e 5 mM K2EDTA a 25 ºC. Após esse período a proteína e o fenol foram
diluídos para 0,05 mg/ml e 0,05%, respectivamente. A essa nova sol ução foi
adicionada 3 mM de ATP. As curvas controle (0% de fenol) e controle positivo
para 0.05% de fenol foram realizados com 0,05 mg/mL de S1 mantendo o
tempo de pré-incubação de 2 min a 25 °C antes da adição de ATP (n=3,
X±SE).

99
7. DISCUSSÃO: Parte II
A relação entre a estrutura e função das proteínas , de um modo geral, já
é bem estabelecidana literatura (Dobson, 2000). Estudos acerca da
estabilidade de proteínas, incluindo experimentos de desnaturação e
renaturação protéica, têm sido bastante úteis na investigação da estrutura e
função de diversas proteínas , incluindo a miosina. A miosina é uma proteína
que possui uma estrutura altamente assimétrica (Rayment et al., 1993). A
estrutura tridimensional do S1 de mú sculo esquelético de galinha (Fig. 3)
mostra uma série de fendas e domínios que dividem a proteína em partes
distintas, cada uma com uma participação fundamental para atividade
enzimática (Rayment et al., 1993).
Solutos perturbadores de estrutura de prot eínas, como ureía e guanidina
(Yancey et al., 1982), agem de uma forma muito peculiar, aumentando a área
de superfície das proteínas e gerando seu desenovelamento. Diversos
trabalhos mostram que o S1 é uma proteína altamente suscetível à ação de
solventes e solutos (Muhlrad et al., 1986; Muhlrad, 1989; Nozais et al., 1992;
Ortiz-Costa et al., 2002). Embora os dados obtidos neste trabalho não mostrem
diretamente a estrutura cristalográfica do S1, os experimentos mostram que a
estrutura do S1 é influenciada pela ação do fenol.
Os espectros de emissão de fluorescência intrínseca do S1 na ausência
de nucleotídeo mostraram que o fenol tem um efeito sobre a estrutura terciár ia
da proteína. Do mesmo modo, essas concentrações modularam a atividade
catalítica do S1. Esses resultados sugerem que o efeito sobre a estrutura do S1
poderia estar modulando a função catalítica da proteína. Entretanto, o fenol

100
apresenta efeitos bifásicos no que se refere à concentração do solvente sobre
a atividade ATPásica. Como observado na s Figuras 16 e 17, concentrações
maiores de fenol mostram uma completa inativação da enzima. A estrutura
protéica, no entanto, sofre algumas modificações sem necessariamente
apresentar uma alta taxa de desenovelamento. Isto pode ser percebido através
dos centros de massa onde para as concentrações de 0,25 e 1% o
deslocamento é o mesmo, não havendo diferença significativa ao aumentar a
concentração (Figs. 26 e 31). Os espectros de emissão na presença de
análogos de ATP corroboram a idéia de que o fenol não p romove desnaturação
severa da proteína uma vez que o fenol não foi capaz de modular a estrutura
terciária do S1 na presença dos análogos de ATP (Fig. 29) nas concentrações
utilizadas. Além disso, nós podemos observar , nas Figuras 34 e 35, que após a
diluição do fenol a atividade Mg 2+ATPásica e K+EDTA ATPásica do S1
retornam aos níveis obtidos na presença da menor concentração de fenol. Isso
indica que qualquer efeito de desenovelamento promovido pelo fenol é parcial
e reversível em baixas concentrações. A exposição de superfícies hidrofóbicas
favorece o desenovelamento, assim como a formação de agregados da
molécula de S1, como mostrado por Muhlrad et al. (1986). Yamam oto et al.
(1994) também observaram que o processo de formação de agregados pela
miosina é acompanhado pelo aumento na turbidez das soluções protéicas após
serem submetidas a altas pressões. Curiosamente, o tratamento do S1 com
fenol também acarretou um aumento da turbidez em concentrações elevadas
(1-2%) ou quando incubadas por longos períodos (>10 min), mesmo em
concentrações menores que 1%. Esses dados indicam que o fenol pode
promover diferentes alterações na estrutura terciária do S1 , gerando estados

101
intermediários com alta atividade enzimática em baixas concentrações do
solvente. Em contrapartida, em concentrações maiores que 0 ,8%, o fenol pode
estar levando ao desenovelamento e desnaturação, como sugerido pela perda
da atividade.
A presença de nucleotídeo e a sua interação com o sítio de ligação é
capaz de promover diferentes estados confo rmacionais transitórios no S1
(Johnson & Taylor, 1978). Nesse sentido, a mudança conformacional
promovida durante o fechamento do sítio catalítico consegue reverter ou
superar a ação do fenol sobre a estrutura do S1. Dessa forma, durante os
estados em que o sítio catalítico encontra-se fechado, o fenol não foi capaz de
modular a estrutura do S1, como mostrado na Figura 29. Em contrapartida, na
presença de ADP, nos observamos novamente uma alteração estrutural
provocada pelo fenol. Esses achados nos mostram mais uma vez que o efeito
do fenol é mais pronunciado quando o sítio catalítico se encontra na forma
aberta ou expondo suas superfícies hidrofóbicas , favorecendo a liberação dos
produtos da hidrólise de ATP. Essa hipótese é baseada na observação do
efeito da ligação da actina ao S1, onde a interação acto-S1 é capaz de acelerar
a saída dos produtos de hidrólise modulando a abertura do sítio catalítico
(Lymn & Taylor, 1971), e conseqüentemente estimulando a atividade ATPásica.
Nossos dados sugerem que, na ausência de actina, o fenol estaria assumindo
o papel desta, acelerando a liberação dos produtos.
Diante disso, se adicionássemos fenol ao complexo acto -S1, obteríamos
um aumento parecido ou maior da atividade ATPásica da miosina. Entretanto,
esta hipótese não se confirmou nos experimentos na presença de actina. Ao
analisarmos a Figura 17, observamos que , apesar da atividade ATPásica ser

102
estimulada, a magnitude do efeito do fenol na presença de actina é bem menor
quando comparado à sua ausência (Fig. 16). Na presença de actina em
concentrações saturantes, a enzima estaria trabalhando aproximadamente com
70% da sua capacidade máxima e nessa condição outros moduladores
positivos (temperatura, força iônica, etc) aumentariam muito pouco a
capacidade enzimática. Desta forma, o efeito que o fenol promove sobre a
atividade Mg2+ATPásica do S1 pode estar induzindo efeito similar.
Interessantemente, os ensaios de co -sedimentação mostram que o fenol
interfere na ligação acto-S1, diminuindo a fração de S1 ligada à actina quando
tratada previamente pelo solvente. Isso nos leva a crer que o fenol não só
mimetize o papel da actina no ciclo catalítico como também a ação deste
solvente sobre a estrutura terciária do S1 altera direta ou indiretamente a
região de ligação com actina.

103
8. CONCLUSÕES
STEADY-STATE
• O efeito causado pelo fenol na atividade ATPásica é dependente da
concentração e do tempo de exposição da proteína ao solvente;
• Os efeitos do fenol são reversíveis em baixas concentrações.
• Os efeitos do fenol são específicos e dependentes do íon complexado
ao ATP.
• Os efeitos do fenol são menos pronunciados na presença de actina.
CICLO CATALÍTICO
• O fenol altera todos os passos cinéticos do ciclo catalítico.
• Nos estados pré-hidrólise o fenol promove um efeito inib itório, reduzindo
a velocidade de ligação e isomerização correspondente às constantes
K1k2.
• O fenol diminui a constante de clivagem de ATP que corresponde a k3 +
k-3
• O fenol acelera as constantes k4 e k-b, correspondentes à liberação dos
produtos, e isso é refletido no aumento da atividade ATPásica do S1
(kcat).
MUDANÇAS CONFORMACIONAIS
• O fenol induz mudanças conformacionais na estrutura do S1 de forma a
favorecer a exposição e liberação dos produtos. Entretanto , essa
mudança promove uma diminuição da liga ção da actina ao S1.

104
9 - REFERÊNCIAS BIBLIOGRAFICAS
Bagshaw CR, Trentham DR (1974) The characterization of myosin -product complexes and of product -release steps during the magnesiumion-dependent adenosine triphosphatase reaction. Biochem J 141: 331 -349.
Barouch WW, Moss C (1971) Effect of temperature on actin activationon heavy meromyosin ATPase. Biochim Biophys Acta 234: 183 -189.
Blum H., Beier H., and Gross H. J. (1987) Improved silver staining ofplant protein, RNA and DNA in polyacrylamide g els. Eletrophoresis 8, 93-99.
Bremel RD, Weber A (1975) Calcium binding to rabbit skeletal myosinunder physiological conditions. Biochim Biophys Acta 376: 366 -374.
Brooks, G. A., Fahey, T. D., and Baldwin, K. M. (2004) ExercisePhysiology Human Bioenergetics and his Applications. 4º edition. NewYork.
Capitanio M, Canepari M, Cacciafesta P, Lombardi V, Cicchi R, Maffei M,Pavone FS, Bottinelli R (2006) Two independent mechanical events inthe interaction cycle of skeletal muscle myosin with actin . Proc Natl AcadSci U S A 103: 87-92.
Chalovich JM (1992) Actin mediated regulation of muscle contraction.Pharmacol Ther 55: 95-148.
DasGupta G, Reisler E (1991) Nucleotide -induced changes in theinteraction of myosin subfragment 1 with actin: detect ion by antibodiesagainst the N-terminal segment of actin. Biochemistry 30: 9961 -9966.
Ebashi S (1974) Regulatory mechanism of muscle contraction withspecial reference to the Ca -troponin-tropomyosin system. EssaysBiochem 10: 1-36.
Farah CS, Reinach FC (1995) The troponin complex and regulation ofmuscle contraction. FASEB J 9: 755 -767.
Fisher AJ, Smith CA, Thoden J, Smith R, Sutoh K, Holden HM, RaymentI (1995) Structural studies of myosin:nucleotide complexes: a revisedmodel for the molecular basis of muscle contraction. Biophys J 68: 19S -26S.
Geeves MA (1991) The dynamics of actin and myosin association andthe crossbridge model of muscle contraction. Biochem J 274 ( Pt 1): 1 -14.
Geeves MA, Holmes KC (1999) Structural mechanism of musclecontraction. Annu Rev Biochem 68: 687 -728.

105
Giulian GG, Moss RL, Greaser M (1983) Improved methodology foranalysis and quantitation of proteins on one -dimensional silver-stainedslab gels. Anal Biochem 129: 277 -287.
Goldman YE, Brenner B (1987) Specia l topic: molecular mechanism ofmuscle contraction. General introduction. Annu Rev Physiol 49: 629 -636.
Gornall AG, Bardawill CJ, David MM (1949) Determination of s erumproteins by means of the biuret reaction. J Biol Chem 177: 751-766.
Grubmeyer C, Penefsky HS (1981) Cooperativity between catalytic sitesin the mechanism of action of beef heart mitochondrial adenosinetriphosphatase. J Biol Chem 256: 3728 -3734.
Halpern D (1977) Histologic studies in animals after intramuscularneurolysis with phenol. Arch Phys Med Rehabil 58: 438 -443.
Halpern D, Meelhuysen FE (1966) Phenol motor point block in themanagement of muscular hypertonia. Arch Phys Med Rehabil 47: 659 -664.
Hiratsuka T (1992) Spatial proximity of ATP -sensitive tryptophanylresidue(s) and Cys-697 in myosin ATPase. J Biol Chem 267: 14949 -14954.
Huxley HE (1953) X-ray analysis and the problem of muscle. Proc R SocLond B Biol Sci 141: 59-62.
Huxley HE (1957) The double array of filaments in cross -striated muscle.J Biophys Biochem Cytol 3: 631-648.
Johnson KA, Taylor EW (1978) Intermediate states of subfragment 1 andactosubfragment 1 ATPase: reevaluation of the mechanism.Biochemistry 17: 3432-3442.
Khalili AA, Betts HB (1967) Peripheral nerve block with phenol in themanagement of spasticity. Indications and complications. JAMA 200:1155-1157.
Klibanov AM (1989) Enzymatic catalysis in anhydrous organic solvents.Trends Biochem Sci 14: 141-144.
Komatsu H, Shigeoka T, Ohno T, Kaseda K, Kanno T, Matsumoto Y,Suzuki M, Kodama T (2004) Modulation of actomyosin motor function by1-hexanol. J Muscle Res Cell Motil 25: 77 -85.
Kovacs M, Toth J, Hetenyi C, Malnasi -Csizmadia A, Sellers JR (2004)Mechanism of blebbistatin inhibition of myosin II. J Biol Chem 279:35557-35563.

106
Koyama H, Murakami K, Suzuki T, Suzaki K (1992) Phenol block for hipflexor muscle spasticity under ultrasonic monitoring. Arch Phys MedRehabil 73: 1040-1043.
Laemmli UK (1970) Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature 227: 680-685.
Lakowicz, J. R. (1999) Principles of Fluorescence Spectroscopy. KluwerAcademic/Plenum Publisher, New York.
Lee JC, Downing SE (1981) Negative inotropic effects of phenol onisolated cardiac muscle. Am J Pathol 102: 36 7-372.
Lymn RW, Taylor EW (1970) Transient state phosphate production in thehydrolysis of nucleoside triphosphates by myosin. Biochemistry 9: 2975 -2983.
Lymn RW, Taylor EW (1971) Mechanism of adenosine triphosphatehydrolysis by actomyosin. Biochemistry 10: 4617-4624.
Machado C.J.V. (2003) Caracterização dos efeitos do neurobloqueadorquímico fenol nas proteínas contráteis e regulatórias de músculoesquelético. Rio de Janeiro, 109p. Tese (Mestrado em CiênciasBiológicas) - Medicina Física e Reabili tação da Universidade Federal doRio de Janeiro.
Mannherz HG, Goody RS (1976) Proteins of contractile systems. AnnuRev Biochem 45: 427-465.
Mariano AC, Alexandre GM, Silva LC, Romeiro A, Cameron LC, Chen Y,Chase PB, Sorenson MM (2001) Dimethyl sulphoxide enhances theeffects of P(i) in myofibrils and inhibits the activity of rabbit skeletalmuscle contractile proteins. Biochem J 358: 627 -636.
Maruta S, Homma K, Ohki T (1998) Conformational changes at thehighly reactive cystein and lysine reg ions of skeletal muscle myosininduced by formation of transition state analogues. J Biochem (Tokyo)124: 578-584.
Millar NC, Geeves MA (1988) Protein fluorescence changes associatedwith ATP and adenosine 5' -[gamma-thio]triphosphate binding to skeletalmuscle myosin subfragment 1 and actomyosin subfragment 1. BiochemJ 249: 735-743.
Muhlrad A (1989) Isolation and characterization of the N-terminal 23-kilodalton fragment of myosin subfragment-1. Biochemistry 28: 4002-4010.

107
Muhlrad A, Chaussepied N (1990) Effect of nucleotides, actin andtemperature on thermolysin digestion of myosin subfragment -1. Eur JBiochem 192: 663-668.
Muhlrad A, Kasprzak AA, Ue K, Ajtai K, Burghardt TP (1986)Characterization of the isolated 20 KDa and 50 KDa fragments of themyosin head. Biochim Biophys Acta 869: 128-140.
Nozais M, Bechet JJ, Houadjeto M (1992) Inactivation, subunitdissociation, aggregation, and unfolding of myosin subfragment -1 Duringguanidine denaturation. Biochemistry 31: 1210 -1215.
Ortiz-Costa S, Sorenson MM, Sola-Penna M (2002) Counteractingeffects of urea and methylamines in function and structure of skeletalmuscle myosin. Arch Biochem Biophys 408: 272 -278.
Pardee JD, Spudich JA (1982) Purification of muscle actin. MethodsEnzymol 85 Pt B: 164-181.
Park S, Ajtai K, Burghardt TP (1996) Optical activity of a nucleotide -sensitive tryptophan in myosin subfragment 1 during ATP hydrolysis.Biophys Chem 63: 67-80.
Phan BC, Peyser YM, Reisler E, Muhlrad A (1997) Effect of comp lexesof ADP and phosphate analogs on the conformation of the Cys707 -Cys697 region of myosin subfragment 1. Eur J Biochem 243: 636 -642.
Rayment I (1996) The structural basis of the myosin ATPase activity. JBiol Chem 271: 15850-15853.
Rayment I, Rypniewski WR, Schmidt-Base K, Smith R, Tomchick DR,Benning MM, Winkelmann DA, Wesenberg G, Holden HM (1993) Three -dimensional structure of myosin subfragment -1: a molecular motor.Science 261: 50-58.
Robinson JD, Davis RL, Steinberg M (1986) Fluoride and berylliuminteract with the (Na + K)-dependent ATPase as analogs of phosphate. JBioenerg Biomembr 18: 521-531.
Salerno VP, Ribeiro AS, Dinucci AN, Mignaco JA, Sorenson MM (1997)Specificity and kinetic effects of nitrophenol analogues that activatemyosin subfragment 1. Biochem J 324 ( Pt 3): 877 -884.
Sanger TD (2003) Childhood onset generalised dystonia can bemodelled by increased gain in the indirect basal ganglia pathway. JNeurol Neurosurg Psychiatry 74: 1509 -1515.
Schiaffino S, Reggiani C (1996) Molecular diversity of myofibrillarproteins: gene regulation and functional significance. Physiol Rev 76:371-423.

108
Sellers JR, Goodson HV (1995) Motor proteins 2: myosin. Protein Profile2: 1323-1423.
Shaw MA, Ostap EM, Goldman YE (2003) Mechan ism of inhibition ofskeletal muscle actomyosin by N -benzyl-p-toluenesulfonamide.Biochemistry 42: 6128-6135.
Szent-Györgyi A (1951) The Chemistry and Chemical Mechanism ofMuscular Contraction. Transactions of the New York Academy ofSciences 162 pg.
Takeda S, Yamashita A, Maeda K, Maeda Y (2003) Structure of the coredomain of human cardiac troponin in the Ca(2+) -saturated form. Nature424: 35-41.
Taussky HH, Shorr E (1953) A microcolorimetric method for thedetermination of inorganic phosphorus. J Biol Chem 202: 675-685.
Trentham DR, Bardsley RG, Eccleston JF, Weeds AG (1972) Elementaryprocesses of the magnesium ion -dependent adenosine triphosphataseactivity of heavy meromyosin. A transient kinetic approach to the study ofkinases and adenosine triphosphatases and a colorimetric inorganicphosphate assay in situ. Biochem J 126: 635 -644.
Trentham DR, Eccleston JF, Bagshaw CR (1976) Kinetic analysis ofATPase mechanisms. Q Rev Biophys 9: 217 -281.
Trybus KM (1994) Regulation of e xpressed truncated smooth-musclemyosins - role of the essential light-chain and tail length. Journal of BiolChem 269: 20819-20822.
Trybus KM (2000) Biochemical studies of myosin. Methods 22: 327 -335.
Webb MR (1992) A continuous spectrophotometric assay for i norganicphosphate and for measuring phosphate release kinetics in biologicalsystems. Proc Natl Acad Sci U S A 89: 4884 -4887.
Weeds AG, Taylor RS (1975) Separation of subfragment -1 isoenzymesfrom rabbit skeletal muscle myosin. Nature 257: 54 -56.
Yamamoto K, Yoshida Y, Morita J, Yasui T (1994) Morphological andphysicochemical changes in the myosin molecules induced byhydrostatic pressure. J Biochem (Tokyo) 116: 215 -220.
Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN (1982) Livingwith water-stress - evolution of osmolyte systems. Science 217: 1214-1222.

109
ANEXO I
CURRICULUM VITAE
NOME: Raquel Guimarães CoelhoNASCIMENTO: 26/01/1981.NATURALIDADE: Rio de Janeiro
FORMAÇÃO ACADÊMICA
- Bacharel em Educação Física – Escola de Educação Física e Desportos daUniversidade Federal do Rio de Janeiro, março de 1998 a março de 2002.
ORIENTAÇÃO DE ESTUDANTE:
1. Cícero Figueiredo Freitas – iniciação científica de novembro de 2005 anovembro de 2006 (atualmente aluno de pós -graduação do Instituto deBioquímica Médica da Universidade Federal do Rio de Janeiro).
2. Luciana Elena de Souza Fraga Machado - iniciação científica de setembro de2005 a junho de 2007 (atualmente aluna de pós -graduação do Instituto deBioquímica Médica da Universidade Federal do Rio de Janeiro).
COMUNICAÇÃO EM CONGRESSO
- 4 comunicações em congressos naciona is
PUBLICAÇÕES:
Leite, T.C., Da Silva, D., Coelho, R.G., Zancan, P., & Sola-Penna, M. (2007). Lactatefavors the dissociation of skeletal muscle 6 -phophofructo-1-kinase tetramers down-regulating the enzyme and muscle glycolysis. Biochem J. 2007 Aug 1; [Epub ahead ofprint]PMID: 17666012 [PubMed - as supplied by publisher]
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo