caracterização de bactérias produtoras de substâncias...
TRANSCRIPT

Olinda Cabral da Silva Santos
Caracterização de bactérias produtoras de substâncias
antimicrobianas isoladas de esponjas marinhas
Orientada por:
Marinella Silva Laport Marcia Giambiagi-deMarval
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO INSTITUTO DE MICROBIOLOGIA PROF PAULO DE GÓES
RIO DE JANEIRO AGOSTO DE 2008
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ciências Biológicas (Microbiologia)

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Santos, Olinda Cabral da Silva Caracterização de bactérias produtoras de substâncias antimicrobianas isoladas de esponjas marinhas / Olinda Cabral da Silva Santos – Rio de Janeiro, 2008. xiii, 117p.:il.;29,7cm. Dissertação (Mestrado em Ciências Biológicas) Universidade Federal do Rio de Janeiro/ Instituto de Microbiologia Prof. Paulo de Góes, 2008. Orientadores: Marinella Silva Laport e Marcia Giambiagi-deMarval Referências bibliográficas: f. 99-113. 1. litoral do Rio de Janeiro 2. esponjas marinhas 3. bactérias associadas 4. substâncias antimicrobianas 5. 16S rRNA 6. antimicrobianos. I. Laport, Marinella Silva e Giambiagi de-Marval, Marcia. II. Universidade Federal do Rio de Janeiro, Instituto de Microbiologia Prof. Paulo de Góes, Mestrado em Ciências Biológicas (Microbiologia). III. Caracterização de bactérias produtoras de substâncias antimicrobianas isoladas de esponjas marinhas.

iii
Olinda Cabral da Silva Santos
Caracterização de bactérias produtoras de substâncias antimicrobianas isoladas de esponjas marinhas
Rio de Janeiro, 07 de agosto de 2008.
(Marinella Silva Laport, Ph.D, UFRJ – Professor Orientador)
(Marcia Giambiagi-deMarval, Ph.D, UFRJ – Professor Orientador)
(Lucy Seldin, Ph.D, UFRJ – Presidente Banca)
(Sergio Eduardo Longo Fracalanzza, Ph.D, UFRJ – Membro Banca)
(Geraldo Renato de Paula, Ph.D, UFF – Membro Banca)
(Rafael Silva Duarte, Ph.D, UFRJ – Revisor Banca)

iv
O presente trabalho foi realizado no Laboratório de Microbiologia Molecular, Departamento de Microbiologia Médica, Instituto de Microbiologia Prof. Paulo de Góes, Centro de Ciências da Saúde (CCS), Universidade Federal do Rio de Janeiro, sob a orientação da Prof.as Marinella Silva Laport e Marcia Giambiagi-deMarval.

v
AGRADECIMENTOS
A Deus, por ter me dado uma família linda, amigos verdadeiros, força para lutar pelo
que anseio, coragem para enfrentar os desafios e proteção para sempre seguir a diante.
À minha avó, uma pessoa inesquecível, que mesmo diante de uma vida extremamente
sofrida sempre confiou em Deus e nos seus desígnios.
À minha irmã agradeço pela amizade, pelo afeto, por ter me dado um afilhado
encantador e um cunhado que não me pede dinheiro emprestado!
À Profª. Marinella Silva Laport agradeço pela orientação, incentivo, apoio, confiança e
acima de tudo, pelos conhecimentos adquiridos e pela amizade.
À Profª. Marcia Giambiagi-deMarval agradeço por ter me ajudado nos momentos mais
difíceis. Se não fosse por seu apoio, talvez não estivesse escrevendo estes agradecimentos
hoje.
À Profª. Kátia Regina Netto dos Santos, a quem devo eterna gratidão.
Ao Prof. Rafael agradeço pela compreensão e paciência.
Aos amigos Bruno, Cristina, Danielle, Naira, Elaine, Milena e Paula pelas
contribuições, carinho e agradável convívio.
Aos amigos e colegas do laboratório de Microbiologia Molecular: Paula, Naira,
Elaine, André, Orlando, Nathália, Juliana, Cleyton, Palloma, Luana e Lorayne pelo
companheirismo, apoio e momentos divertidos e agradáveis proporcionados ao longo desse
período.
Ao Instituto de Microbiologia Prof. Paulo de Góes, nas pessoas da Profª. Agnes Marie
Sá Figueiredo, Diretora, e Profª. Thais Cristina Baeta Soares Souto Padron, Coordenadora de
Pós-Graduação.

vi
Aos meus pais, Ivete e Antonio, Pelos ensinamentos que desde cedo guiam meus passos, Pela confiança e companheirismo em todos os momentos,
Pela amizade, respeito e carinho, Por todas as oportunidades oferecidas, Por estarmos unidos nesta caminhada,
dedico.
Ao meu marido, Silvestre, companheiro e amigo alguém que estará sempre a meu
lado, com amor, ofereço.

vii
RESUMO
Olinda Cabral da Silva Santos
Caracterização de bactérias produtoras de substâncias antimicrobianas isoladas de esponjas marinhas
Orientadores: Marinella Silva Laport e Marcia Giambiagi-deMarval
Resumo da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Ciências Biológicas.
O uso extensivo de antibióticos tem acelerado o surgimento e seleção de bactérias resistentes justificando a intensa busca por novos produtos com atividade antimicrobiana. As esponjas apresentam uma diversidade notável de metabólitos secundários, muitos deles de grande interesse para a pesquisa biomédica e farmacológica, como agentes antibióticos, antitumorais e antivirais. Vários estudos sugerem que as bactérias associadas às esponjas podem ser as verdadeiras fontes de alguns destes compostos. As bactérias, por produzirem rapidamente uma grande biomassa, podem tornar possível a produção dos compostos em larga escala sem a necessidade de coletar constantemente ou cultivar as esponjas. Este trabalho teve por objetivo caracterizar bactérias produtoras de substâncias antimicrobianas a partir das esponjas Arenosclera brasiliensis, Clathrina aurea, Dragmacidon reticulatus, Geodia corticostylifera, Haliclona spn., Mycale microsigmatosa, Paraleucilla magna, Petromica citrina, Polymastia janeirensis e Tedania ignis, encontradas na costa da cidade do Rio de Janeiro. Para esta finalidade, as esponjas foram coletadas, trituradas em água destilada estéril e o homogeneizado semeado em triplicata em três diferentes meios (BHI, Marine e Marine-mar). As colônias crescidas foram analisadas diariamente por sete dias quanto às características morfológicas e, assim, de um total de 168 UFC (unidades formadoras de colônias), 149 mantiveram-se viáveis após novas passagens. Todas as estirpes viáveis foram submetidas ao teste de detecção de produção de substâncias antimicrobianas (SAM). Dezesseis estirpes (11%) apresentaram atividade inibitória contra a bactéria Corynebacterium fimi. Dentre as estirpes SAM-positivas, nove apresentaram um amplo espectro de ação contra estirpes de importância médica e foram submetidas à identificação fenotípica e molecular. Pela coloração de Gram foram observados três bacilos Gram-positivos e seis bacilos Gram-negativos. Todas as estirpes foram caracterizadas como móveis, catalase e oxidase positivas. As estirpes foram, então, identificadas através do seqüenciamento parcial do gene que codifica para o 16S rRNA e dos sistemas bioquímicos API 50CH ou API 20NE. Dentre as estirpes Gram-negativas, 3 foram identificadas como Pseudovibrio sp., 2 como Pseudomonas fluorescens e 1 como Pseudomonas putida. As três estirpes Gram-positivas foram identificadas como Bacillus pumilis. A seguir, foram avaliadas as condições ótimas de temperatura e salinidade para o crescimento celular e para a produção das substâncias antimicrobianas. Neste contexto, as estirpes SAM-positivas apresentaram um ótimo de crescimento e de produção das substâncias antimicrobianas em uma faixa entre 15-42°C e na concentração de 0,5-2% de NaCl. Dentre as estirpes SAM-positivas identificadas, as mais promissoras como fontes de substâncias antimicrobianas foram aquelas isoladas das esponjas Haliclona spn., P. magna, C. aurea e M. microsigmatosa. Estes resultados sugerem que as bactérias identificadas podem ser potenciais fontes de produção de substâncias antimicrobianas contra infecções bacterianas de importância médica. Palavras-chave: litoral do Rio de Janeiro, esponjas marinhas, bactérias associadas, substâncias antimicrobianas, 16S rRNA, antimicrobianos.
Rio de Janeiro Agosto de 2008

viii
ABSTRACT
Olinda Cabral da Silva Santos
Characterization of bacteria producing antimicrobial substances isolated from marine sponges
Orientadores: Marinella Silva Laport e Marcia Giambiagi-deMarval
Abstract da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Ciências Biológicas.
The extensive use of antibiotics has accelerated the emergence of resistant bacteria and therefore has been strengthening the search for new products with antimicrobial activity. The sponges present a remarkable diversity of secondary metabolites, many of them of great interest for biomedical research and pharmacology, such as antibiotics, antivirals and antitumorais agents. Several studies suggest that bacteria associated with sponges could be the sources of some of these compounds. As bacteria quickly produce a large biomass, they could serve as source for the production of compounds on a large scale without constantly collecting or cultivating the sponges. Thus, the aim of this work was to phenotypically and phylogenetically characterize bacteria producing antimicrobial substance from the sponges Arenosclera brasiliensis, Clathrina aurea, Dragmacidon reticulatus, Geodia corticostylifera, Haliclona spn., Mycale microsigmatosa, Paraleucilla magna, Petromica citrina, Polymastia janeirensis and Tedania ignis, encountered at the coast of Rio de Janeiro. The sponges were collected, triturated in distilled water and the homogenized was plated in triplicate on three different media (BHI, Marine and Marine-Mar). The colonies growth was analyzed for 7 days out of 168 CFU (colony forming units), 149 have remained viable after new passages. All the viable isolates had been submitted to the test for production of antimicrobial substances (SAM). Sixteen isolates (detection rate of 11%) expressed inhibitory activity against the bacteria Corynebacterium fimi. Among the SAM-positive isolates, nine showed a broad range activity against strains of medical importance and were subjected to phenotypic and molecular identification. Three Gram-positive and six Gram-negative rods were observed by the Gram stain procedure. All isolates were characterized as motile, oxidase and catalase positive. Thus, the isolates were identified through the partial sequencing of the gene coding for the 16S rRNA and biochemical systems API 50CH or API 20NE. Among the Gram-negative isolates, 3 were identified as Pseudovibrio sp., 2 as Pseudomonas fluorescens and 1 as Pseudomonas putida. The Gram-positive isolates were identified as Bacillus pumilis. Moreover, the optimum conditions of temperature and salinity for the growth and antimicrobial substances production were evaluated. The SAM-positive isolates had a intensive growth and antimicrobial substance production in a range between 15-42°C and 0.5-2% NaCl concentration. Among the identified SAM-positive isolates, the most promising sources of antimicrobial substances were those isolated from sponges Haliclona spn., P. magna, C. aurea and M. microsigmatosa. These results suggest that the identified bacteria can be potential sources of antimicrobial substances production against bacterial infections of medical relevance. Keywords: coast of Rio de Janeiro, marine sponges, associated bacteria, antimicrobial substance, 16S rRNA, antimicrobial.
Rio de Janeiro Agosto de 2008

ix
SUMÁRIO
Página
1 INTRODUÇÃO 1
1.1 Poríferos: Aspectos Gerais ------------------------------------------------------------- 1
1.2 Microrganismos Associados aos Poríferos ------------------------------------------- 6
1.2.1 Bactérias Associadas aos Poríferos ----------------------------------------------- 8
1.3 Potencial Biotecnológico das Bactérias Associadas aos Poríferos ---------------- 11
1.3.1 Principais Gêneros Bacterianos Produtores de Substâncias
Antimicrobianas ---------------------------------------------------------------------------
11
1.3.2 Produção de Substâncias Bioativas ----------------------------------------------- 13
1.4 A Problemática das Bactérias Multirresistentes aos Antimicrobianos ----------- 22
2 OBJETIVOS 27
2.1 Objetivo Geral --------------------------------------------------------------------------- 27
2.2 Objetivos Específicos ------------------------------------------------------------------- 27
3 MATERIAIS E MÉTODOS 28
3.1 Meios de cultura -------------------------------------------------------------------------- 28
3.2 Espécies de esponjas --------------------------------------------------------------------- 29
3.3 Coleta dos espécimes e preparação dos macerados brutos das esponjas ---------- 29
3.4 Isolamento de estirpes bacterianas associadas às esponjas e condições de
cultura ------------------------------------------------------------------------------------------
30
3.5 Ensaios de produção de substância antimicrobiana (SAM) ------------------------ 30
3.5.1 Triagem de estirpes produtoras de SAM associadas às esponjas ------------- 31
3.5.2 Determinação do espectro de ação das SAM ------------------------------------ 31
3.5.3 Teste de inibição cruzada ---------------------------------------------------------- 33
3.6 Caracterização fenotípica das estirpes marinhas SAM-positivas ------------------ 33
3.6.1 Aspectos Fenotípicos --------------------------------------------------------------- 33
3.6.2 Aspecto morfo-tintorial ------------------------------------------------------------- 33
3.6.3 Produção da enzima catalase ------------------------------------------------------ 34
3.6.4 Produção da enzima citocromo-oxidase ------------------------------------------ 34
3.6.5 Teste de motilidade ----------------------------------------------------------------- 34
3.6.6 Análise das condições de temperatura e salinidade para o crescimento das
estirpes marinhas SAM-positivas --------------------------------------------------------
35

x
3.6.7 Identificação das estirpes marinhas SAM-positivas através do sistema
comercial miniaturizado ------------------------------------------------------------------
35
3.6.8 Susceptibilidade aos antimicrobianos -------------------------------------------- 36
3.7 Identificação molecular das estirpes marinhas SAM-positivas -------------------- 37
3.7.1 Obtenção de DNA total com guanidina ------------------------------------------ 37
3.7.2 Reação de polimerase em cadeia (PCR) ----------------------------------------- 38
3.7.3 Eletroforese de DNA em gel de agarose ----------------------------------------- 39
3.7.4 Seqüenciamento parcial do gene que codifica para o 16S rRNA ------------- 39
3.8 Caracterização parcial da(s) substância(s) antimicrobiana(s) quanto à natureza
química -----------------------------------------------------------------------------------------
40
3.9 Análise das condições de temperatura e salinidade para a produção da(s)
substância(s) antimicrobianas(s) ------------------------------------------------------------
40
3.10 Organograma das atividades----------------------------------------------------------- 41
4 RESULTADOS 43
4.1 Isolamento de bactérias associadas às esponjas -------------------------------------- 43
4.2 Atividade antimicrobiana das estirpes associadas às esponjas --------------------- 47
4.3 Caracterização fenotípica das estirpes marinhas SAM-positivas ------------------ 55
4.3.1 Caracterização das estirpes marinhas SAM-positivas -------------------------- 55
4.3.2 Análise das condições de temperatura e salinidade para o crescimento das
estirpes marinhas SAM-positivas --------------------------------------------------------
58
4.3.3 Identificação das estirpes marinhas SAM-positivas através do sistema
comercial miniaturizado ------------------------------------------------------------------
61
4.3.4 Susceptibilidade aos antimicrobianos -------------------------------------------- 61
4.4 Identificação molecular das estirpes marinhas SAM-positivas -------------------- 64
4.5 Caracterização da(s) substância(s) antimicrobiana(s) quanto à natureza química
---------------------------------------------------------------------------------------------------
66
4.6 Análise das condições de temperatura e salinidade para a produção da(s)
substância(s) antimicrobianas(s) -----------------------------------------------------
69
4.7 Identificação e caracterização de bactérias produtoras de substâncias
antimicrobianas isoladas de esponjas marinhas -------------------------------------------
72
5 DISCUSSÃO 74
6 BIBLIOGRAFIA 99
7 ANEXO 114

xi
LISTA DE ABREVIATURAS Ab = Arenosclera brasiliensis
ATCC = “American Type Culture Collection”
ATM = aztreonam
MTM = “Motility Test Medium” (Meio de cultura para teste de motilidade)
BHI = “Brain Heart Infusion” (Infusão de coração e cérebro)
Ca = Chlatrina aurea
CA-MRSA = “community-associated methicillin-resistant Staphylococcus aureus”
(Staphylococcus aureus resistentes à meticilina associados a comunidade)
CAZ = ceftazidima
CCM = “Czechoslovak Collection of Microorganisms”
CDC = “Centers for Disease Control and Prevention”
CFE = cefalexina
CIP = ciprofloxacina
CLO = cloranfenicol
CLSI = “Clinical and Laboratory Standards Institute”
CMI = Concentração mínima inibitória
DGGE = “Denaturing gradient gel electrophoresis” (Eletroforese em gel com gradiente
desnaturante)
DMSO = “Dimethyl sulfoxide” (Dimetilsulfóxido)
DNA = ácido desoxirribonucléico
dNTPs = desoxirribonucleotídeos 5’-trifosfatados
Dr = Dragmacidon reticulatus
EDTA = ácido etilenodiaminotetracético
ERI = eritromicina

xii
ESBL = “Extended-Spectrum Beta-Lactamase” (Beta-lactamases de espectro estendido)
f = “forward” (direto)
G = Geodia corticostylifera
GEN = gentamicina
H = Haliclona spn.
HPLC/MS = “High-performance liquid chromatography tandem mass spectrometer”
(Espectrometria de massa acoplada à cromatografia líquida de alta eficiência)
kb = quilobase(s), 1.000 pb
M = meio de cultura Marine
MM = Meio de cultura Marine-mar
Mm = M. microsigmatosa
mRNA = RNA mensageiro
MRSA = “methicillin-resistant Staphylococcus aureus” (Staphylococcus aureus resistentes à
meticilina)
NCTC = “National Collection of Type Cultures”
ND = não determinado
NRPs = “nonribosomal peptides” (peptídeos não-ribossômicos)
NRPSs = “nonribosomal peptides synthetases” (sintetases de peptídeos não-ribossômicos)
NT = não testado
Pc = Petromica citrina
PCR = “Polymerase Chain Reaction” (Reação de polimerase em cadeia)
PEN = penicilina G
Pj = Polymastia janeirensis
PKs = “polyketides” (policetídeo)
PKSs = “polyketides synthases” (sintases de policetídeos)

xiii
Pm = Paraleucilla magna
PVL = “ Panton-Valentine leukocidin” (Leucocidina Panton-Valentine)
r = “ reverse” (reverso)
RNA = ácido ribonucléico
rRNA = RNA ribossomal
SAM = substância(s) antimicrobiana(s)
SCCmec = “Staphylococcal cassette chromosome mec” (Cassete cromossômico de estafilococos mec) spn = nova espécie
SUT = sulfazotrim (sulfametoxazol – trimetoprim)
T.A. = temperatura ambiente
TAE = Tris – Ácido acético – EDTA
Taq = Thermus aquaticus
TE = Tris – EDTA
TET = tetraciclina
Ti = Tedania ignis
TIGR = “The Institute for Genomic Research”
TRIS = Hidroximetil – aminometano
UFC = Unidade(s) formadora(s) de colônia(s)

1
1 INTRODUÇÃO
1.1 Poríferos: Aspectos Gerais
Os poríferos constituem o filo mais primitivo dos metazoários: embora sejam
pluricelulares, suas células possuem um certo grau de independência e não se organizam em
tecidos (SIMPSON, 1984). O filo é subdividido em três classes, chamadas de Hexactinellida,
Calcarea e Demospongiae. As esponjas pertencentes à classe Hexactinellida são caracterizadas
por possuírem um esqueleto de espículas de sílica (dióxido de silício), e por isso, são comumente
chamadas de esponjas-de-vidro. As esponjas-de-vidro estão distribuídas em todo o mundo e são
os poríferos predominantes nas águas profundas da Antártida. Calcarea é uma classe que se
distingue dos outros poríferos pelos seus indivíduos serem constituídos por espículas de calcáreo
(carbonato de cálcio). Existem esponjas calcáreas em todos os oceanos do mundo, a maioria
restrita a águas costeiras relativamente rasas. Desmospongiae é a classe que inclui a maioria das
espécies de esponjas já descritas. Diferenciam-se por possuirem um esqueleto de fibras de
espongina (rede de fibras protéicas elásticas), uma substância exclusiva destes animais. São
encontradas em ambientes marinhos desde a zona de maré até profundidades de cerca de 9.000m,
assim como, em ambientes transicionais e aquáticos de águas doces (HOOPER & VAN SOEST,
2002).
As esponjas são organismos sésseis e filtradores, que, apesar de sua morfologia simples,
são especialmente eficientes em obter alimento da água circundante (REISWIG, 1971). Os
poríferos se nutrem, na maioria das vezes, das menores frações orgânicas, tais como matéria
orgânica dissolvida e picoplâncton (onde se incluem as bactérias e também algas e vários tipos de
vírus), podendo ser altamente eficientes na execução de tal tarefa (ingestão de até 100% das

2
bactérias presentes na água filtrada) (PILE, PATTERSON & WITMAN, 1996). É estimado que
cerca de 24.000 litros de água podem ser filtrados por 1kg de esponja em um único dia (VOGEL,
1977).
A arquitetura das esponjas é diferente de qualquer outro táxon, apresentando uma
fisiologia bastante simples na sua organização (Figura 1) (SIMPSON, 1984).
Figura 1. Representação esquemática de uma esponja. Visualização das estruturas e do mecanismo de filtração (fonte: http://leodavici.tripod.com/id8.html).
A superfície de uma esponja apresenta uma série de poros (ou óstios) formados por
células tubulares chamadas de porócitos, pelos quais a água penetra, trazendo alimento e
oxigênio. No interior do animal, há uma cavidade central, o átrio (ou espongiocele) revestido por
células chamadas de coanócitos e, na parte superior, uma abertura maior, o ósculo, pelo qual a
água sai (SIMPSON, 1984).

3
A epiderme é formada por uma fina camada de células achatadas, os pinacócitos, as quais
são interrompidas pelos porócitos. Os pinacócitos também podem se diferenciar em miócitos,
células que controlam o tamanho do ósculo, a abertura dos poros e, conseqüentemente, o fluxo de
água dentro da esponja (SIMPSON, 1984).
O mesênquima (interno à camada dos pinacócitos) é uma camada gelatinosa que preenche
o corpo das esponjas e é composto por células móveis e indiferenciadas, chamadas de amebócitos
(ou arqueócitos). Os amebócitos têm papel fundamental na distribuição de nutrientes, na
produção de gametas, na regeneração celular e na sustentação da esponja. Por serem totipotentes,
os amebócitos podem dar origem a células chamadas de espongioblastos, que produzem a
espongina, responsável pela sustentação da esponja; e escleroblastos, que produzem espículas de
calcário ou de sílica, para a sustentação da parede do corpo e manutenção da esponja ereta
(SIMPSON, 1984). A espongina junto com as espículas permitem o desenvolvimento de
indivíduos bastante largos, tais como os encontrados entre as espécies tropicais (Figura 2C)
(HOOPER & VAN SOEST, 2002; TAYLOR et al., 2007).
(A) (B) (C) Figura 2. Espécies de esponjas. A esponja incrustante Tedania digitata (A), a espécie pedunculada Axinella cannabina (B) e a gigante Xestospongia testudinaria (C) (TAYLOR et al., 2007).

4
No átrio, encontram-se os coanócitos, também chamados de células de colarinho, porque
possuem um flagelo rodeado por uma coroa de cílios. Estas células funcionam como uma espécie
de sistema digestivo e sistema respiratório combinados, uma vez que os flagelos criam uma
corrente que renova a água que as cobre, da qual elas retiram o oxigênio e as partículas de
alimentos e transferem para os amebócitos distribuírem para todas as células da esponja
(SIMPSON, 1984).
Também presentes no mesênquima de muitas esponjas, estão densas comunidades de
microrganismos (VACELET & DONADEY, 1977). A existência destes supostos simbiontes
junto com os amebócitos é um tanto paradoxal e implica no reconhecimento de diferentes tipos
microbianos pelas células da esponja ou na utilização de mecanismos pelos simbiontes para evitar
a fagocitose (WILKINSON, 1984). Wehrl, Steinert & Hentschel (2007) observaram que bactérias
derivadas da água do mar foram consumidas duas ordens de magnitude mais rapidamente pelas
esponjas do que um consórcio de bactérias derivadas de esponjas.
A maioria das esponjas é incrustante (Figura 2A), formando camadas de espessura variada
em substratos duros, em geral rochas, mas também em qualquer outro, como madeiras, metal ou
cimento (RUPPERT & BARNES, 1994). As poucas espécies encontradas em substratos móveis
como lama, areia ou cascalho são geralmente eretas e finas para evitar o soterramento pelo
sedimento, podendo ter um pedúnculo ou raiz para ancoragem no substrato, como, por exemplo,
a espécie Axinella cannabina (Figura 2B). As esponjas estão entre os mais belos e admirados
invertebrados marinhos, com uma enorme diversidade de tamanhos, formas e cores nas mais
diferentes tonalidades (RUPPERT & BARNES, 1994).
Vários animais se alimentam de esponjas, embora o dano causado por estes predadores
seja geralmente pequeno. Alguns moluscos, ouriços e estrelas-do-mar, além de peixes tropicais
(donzelas, peixes-borboleta) e tartarugas, comem esponjas. Para se protegerem da predação

5
excessiva, as esponjas apresentam alguns mecanismos de defesa (MÜLLER & MÜLLER, 2003).
O mecanismo primário de defesa das esponjas é de natureza química. As esponjas produzem uma
ampla gama de substâncias tóxicas, alguns bastante potentes. Espécies de alguns gêneros como
Tedania e Neofibularia podem causar dermatites dolorosas em seres humanos (ISBISTER &
HOOPER, 2005). Muitas espécies produzem substâncias com atividade antimicrobiana
(antibacteriana, antifúngica e antiviral) (MÜLLER & MÜLLER, 2003). Além de defesas
antipredação e contra infecções microbianas, as toxinas das esponjas servem também para a
competição por espaço com outros invertebrados, como ascídias, briozoários, corais e até mesmo
outras esponjas. Isto permite a algumas esponjas crescerem rapidamente e recobrirem a fauna e a
flora adjacentes (BAVESTRELLO et al., 2000).
O conhecimento acerca da espongofauna marinha brasileira é tipicamente fragmentário,
restringindo-se a uma lista de espécies para algumas poucas localidades no litoral. Hadju & Van
Soest (1992) realizaram uma revisão do gênero Asteropus (filo Demospongiae, ordem
Astrophorida, família Ancorinidae) para o oceano Atlântico, incluindo a descrição de três novas
espécies e a revisão da família Coppatiidae. Posteriormente, Hadju & Desqueyroux-faundez
(1994) apresentaram uma sinopse de três novas espécies de Mycale na América do Sul: M.
beatrizae, M. darwini e M. thielei (Demospongiae, Poecilosclerida, Mycalidae). A composição e
a distribuição da fauna de esponjas do litoral de Pernambuco foram publicadas por Muricy &
Moraes (1999) quando demonstraram a existência de um grande número de espécies, tendo
assinalado nove novos registros para a costa brasileira. Mothes, Hadju & Van Soest (2000)
realizaram algumas observações sobre o gênero Tedania no sudoeste do Atlântico tropical, dando
ênfase à análise de diversos morfotipos depositados em diferentes instituições. A revisão do
gênero Aplysina, com a descrição de novas espécies coletadas no canal de São Sebastião (SP) foi
publicada por Pinheiro & Hajdu (2001).

6
Apesar de serem conhecidas cerca de 300 espécies para a costa brasileira, esta, junto com
a costa pacífica da América do Sul, estão dentre as menos estudadas com referência a sua
espongofauna marinha (MURICY & SILVA, 1999). Apenas recentemente, iniciaram-se esforços
intensivos no sentido de se obter uma lista completa das espécies de esponjas, em especial da
Classe Demospongiae, ocorrendo no Canal de São Sebastião e adjacências (SARMENTO &
CORREIA, 2002). As coletas foram feitas no canal de São Sebastião e na costa de Ilhabela, no
litoral norte de São Paulo. Foram encontradas 140 espécies de esponjas, o equivalente a quase
metade das cerca de 300 já registradas para a costa brasileira (LANNA et al., 2007). Em função
do pouco conhecimento de aspectos biológicos básicos e dos padrões de distribuição espacial dos
poríferos, não é possível ainda classificar as espécies como endêmicas, invasoras, raras ou em
extinção, para a costa brasileira (SARMENTO & CORREIA, 2002).
1.2 Microrganismos Associados aos Poríferos
As interações entre esponjas e microrganismos ocorrem de várias formas. Para as
esponjas, diferentes microrganismos podem representar fontes de alimentos (PILE,
PATTERSON & WITMAN, 1996), patógenos/parasitas (BAVESTRELLO et al., 2000), ou
simbiontes (WILKINSON, 1983). Na última década, a aplicação de ferramentas moleculares
permitiu demonstrar que cada um dos três domínios da vida (Bacteria, Archaea e Eukarya) são
residentes nas esponjas (PRESTON et al., 1996; GARSON et al., 1998; HENTSCHEL et al.,
2002).
Muitas esponjas contêm elevadas quantidades de bactérias em seus tecidos, chegando a
ocupar 40 a 60% do total de sua biomassa (o equivalente a 108/1010 células por grama de esponja)
(HENTSCHEL et al., 2002). Através de análises de seqüenciamento do gene que codifica para o

7
RNA ribossomal 16S (16S rRNA) diversos filos bacterianos foram descritos. Entre os filos
representantes encontram-se: Acidobacteria, Actinobacteria, Bacteroidetes, Chloroflexi,
Cyanobacteria, Deinococcus-Thermus, Firmicutes, Gemmatimonadetes, Nitrospira,
Planctomycetes, Proteobacteria (Alpha, Beta, Delta e Gamaproteobacteria), Spirochaetes e
Verrucomicrobia (LOPEZ et al., 1999; HENTSCHEL et al., 2002; HILL et al., 2006). Membros
dos filos Actinobacteria, Bacteroidetes, Cyanobacteria, Planctomycetes, Proteobacteria e
Verrucomicrobia também já foram isolados em cultura pura (BURJA & HILL, 2001; CHELOSSI
et al., 2004; LAFI, GARSON & FUERST, 2005). Além disso, um novo filo, Poribacteria, foi
isolado de várias esponjas (FIESELER et al., 2004).
Contrastando com as esponjas marinhas, evidências disponíveis para espécies de água
doce sugerem que a diversidade e a abundância microbianas são muito menores. Através da
pesquisa do gene que codifica para o 16S rRNA, foi verificado que a esponja de água doce
Spongilla lacustris apresentava uma microbiota com menor diversidade em comparação àquela
apresentada pelas esponjas marinhas (GERNERT et al., 2005).
Embora freqüentemente investigadas através da aplicação de diferentes metodologias,
indícios de arquéas associadas a esponjas continuam sendo raramente encontrados (MARGOT et
al., 2002). Com poucas exceções em Euryarchaeota, as arquéas encontradas em esponjas
marinhas são membros do filo Crenarchaeota (WEBSTER & HILL, 2001; HOLMES &
BLANCH, 2006). Em um estudo sobre arquéas associadas a esponjas, verificou-se que a espécie
de Crenarchaeota “Candidatus Cenarchaeum symbiosum”, intimamente relacionada com outras
Crenarchaeotes não-termofílicas que habitam diversos ambientes marinhos e terrestres,
compreendia mais de 65% do total de células procarióticas presentes em Axinella mexicana, uma
esponja da Califórnia (PRESTON et al., 1996).

8
Microrganismos eucarióticos também residem em esponjas. Dinoflagelados
(SCHÖNBERG & LOH, 2005) e diatomáceas (BAVESTRELLO et al., 2000) já foram descritos,
com as diatomáceas sendo mais prevalentes em regiões polares. Esponjas de água doce
geralmente apresentam microalgas endossimbiontes, principalmente Zoochlorellae (BIL et al.,
1999). Fungos derivados de esponjas têm recebido uma atenção crescente devido ao seu potencial
biotecnológico (HÖLLER et al., 2000). Interessantemente, das 681 estirpes fúngicas isoladas de
16 espécies de esponjas, a maioria pertencia aos gêneros ubiquitários em habitats terrestres
(Aspergillus e Penicillium) (HÖLLER et al., 2000). Porém, não foi esclarecido se tais fungos
eram simbiontes verdadeiros ou provenientes da dieta alimentar da esponja. Por outro lado, um
estudo demonstrou, através de análises microscópicas, evidências da simbiose de uma levedura
com uma esponja do gênero Chondrilla, tanto a partir do tecido adulto da esponja, quanto das
estruturas reprodutivas, com fortes indicações de transmissão vertical da levedura simbionte
(MALDONADO et al., 2005).
Pouco se sabe sobre vírus em esponjas, embora partículas semelhantes a vírus tenham
sido observadas no núcleo celular da esponja Aplysina cavernicola. Foi sugerido que tais
partículas pudessem estar envolvidas na patologia da célula (VACELET & GALLISSIAN, 1978).
Atualmente, estudos indicam que as esponjas contêm uma mistura de microrganismos
generalistas e específicos (TAYLOR et al., 2004) e que as comunidades microbianas a elas
associadas são estáveis no espaço e no tempo (FRIEDRICH et al., 2001; TAYLOR et al., 2005).
1.2.1 Bactérias Associadas aos Poríferos
Supõe-se que há 600 milhões de anos atrás estabeleceu-se entre esponjas e bactérias uma
relação de mutualismo (WILKINSON, 1984), através da qual supostos benefícios para a esponja
incluem a rigidez estrutural (devido ao muco produzido pela bactéria) (WILKINSON, 1978c),

9
incorporação direta de matéria orgânica dissolvida na água do mar (YAHEL et al., 2003),
digestão e reciclagem de colágeno insolúvel produzido pela esponja (WILKINSON, 1979) e
produção de metábolitos secundários que são utilizados pelo hospedeiro para proteção contra
patógenos e predadores (HENTSCHEL et al., 2001). Em alguns casos, a relação entre esponja e
seu simbionte foi experimentalmente demonstrada. Por exemplo, Webster & Hill (2001)
sugeriram que um simbionte dominante na cultura de Rhopaloeides odorabile auxiliava na
nutrição da esponja, baseados na localização desta Alphaproteobacteria ao redor dos coanócitos e
na habilidade desta estirpe de crescer em meios onde aminoácidos e pequenos polipeptídeos eram
as fontes primárias de carbono.
O modo pelo qual as esponjas adquirem seus simbiontes ainda é desconhecido, mas há
duas possibilidades: aquisição por filtração da água do mar ou transmissão vertical dos
simbiontes. As bactérias podem resistir à digestão ou ao sistema imune da esponja através (1) da
formação de cápsula ou de biofilme, (2) crescimento no microambiente formado pelo
mesênquima da esponja e (3) subseqüente estabelecimento como simbionte (FRIEDRICH et al,
1999). A segunda possibilidade sugere que os simbiontes podem ser transmitidos verticalmente
através das larvas (SHARP et al., 2007), ou durante o processo de reprodução assexuada, onde
gêmulas, botões ou ramos dão origem a uma esponja adulta (GAINO et al., 2006). A transmissão
vertical via reprodução assexuada permite a transferência de uma diversidade maior de bactérias
do que a transmissão através da linhagem germinativa, que é mais restrita (HENTSCHEL et al.,
2002).
A noção de que esponjas marinhas poderiam conter uma microbiota específica surgiu há
três décadas atrás com trabalhos realizados por Vacelet (1975), Vacelet & Donadey (1977) e
Wilkinson (1978a-c, 1984, 1987). Baseados em microscopia eletrônica e em estudos de cultivo
bacteriano, estes autores propuseram três tipos de comunidades bacterianas associadas às

10
esponjas: (1) numerosas populações de bactérias específicas presentes no mesênquima da
esponja, (2) pequenas populações de bactérias específicas ocorrendo intracelularmente e (3)
populações de bactérias não-específicas, semelhantes às encontradas no ambiente marinho
adjacente (VACELET, 1975; WILKINSON, 1978b).
Outro significante avanço veio em 2002, quando Hentschel e colaboradores integraram
estes conceitos de comunidades em estudos moleculares. Eles definiram “clusters” esponja-
específicos como grupos bacterianos derivados de esponjas que apresentavam seqüências do gene
que codifica para o 16S rRNA que (1) eram mais similares entre si do que com seqüências
recuperadas de outras fontes; (2) eram encontradas em pelo menos duas espécies de esponjas e/ou
na mesma espécie, mas coletadas em diferentes regiões geográficas; e (3) agrupavam-se
independentemente do método de inferência de filogenia utilizado (HENTSCHEL et al., 2002).
Além disso, estudos subseqüentes dão suporte a esta teoria, com numerosos relatos de
estirpes similares (e em alguns casos esponja-específicas) encontradas em diferentes espécies de
esponjas. A maioria das seqüências do gene que codifica para o 16S rRNA obtidas de estirpes
bacterianas coletadas de dois espécimes, sendo um de esponja não identificada e o outro da
espécie Discodermia dissoluta, foi intimamente relacionada às seqüências bacterianas derivadas
de outras esponjas (HILL, 2004; SCHIRMER et al., 2005). Estas incluem membros de
Acidobacteria, Bacteroidetes, Nitrospira e Proteobacteria. Fieseler e colaboradores (2004)
encontraram membros de um novo filo chamado de Poribacteria, que é relacionado com
Chlamydiae Planctomycetes e Verrucomicrobia (WAGNER & HORN, 2006), em esponjas de
diversas regiões geográficas, mas nunca nas amostras adjacentes de água do mar ou de
sedimentos. Juntos, estes resultados indicam que as comunidades bacterianas associadas são
únicas e específicas às esponjas.

11
1.3 Potencial Biotecnológico das Bactérias Associadas aos Poríferos
1.3.1 Principais Gêneros Bacterianos Produtores de Substâncias Antimicrobianas
Na natureza, antibióticos e outros metabólitos secundários produzidos por
microrganismos têm por função básica antagonizar o crescimento de outros microrganismos,
afetando sua sobrevivência e reprodução. Os antibióticos são conhecidos por conferirem
vantagem seletiva quando há competição entre microrganismos pelo mesmo nicho ecológico
(VINING, 1990). Sua produção depende de um complexo sistema regulador e geralmente é
induzido na fase estacionária de crescimento microbiano (MARTIN & LIRAS, 1989).
Conforme dados apresentados por Hentschel e colaboradores (2001), Pabel e
colaboradores (2003), Chelossi e colaboradores (2004), Selvin e colaboradores (2004), Thakur e
colaboradores (2005), Anand e colaboradores (2006) e Muscholl-Silberhorn, Thiel & Imhoff
(2008), os gêneros bacterianos que se destacam na produção de substâncias antimicrobianas
incluem Flavobacter, Pseudoalteromonas e Vibrio, que são comumente associados às esponjas;
Bacillus, Corynebacteria, Micrococcus e Pseudomonas, que são numericamente bem
representados nas esponjas; e Actinomyces, Aeromonas, Enterobacteriaceae e Streptomyces, que
são raramente isolados de esponjas.
O gênero Flavobacter é ubiquitário, e estima-se que esteja associado a mais de 40% da
microbiota marinha viável. A maioria das espécies de Flavobacter é altamente proteolítica e é
conhecida por produzir também lipases extracelulares (ALONSO et al., 2007).
Pseudoalteromonas é um gênero bacteriano comumente associado a hospedeiros
eucarióticos em comunidades marinhas incrustantes. É capaz de produzir substâncias bioativas
que auxiliam na competição por nutrientes e na colonização de superfícies (BOWMAN, 2007).

12
Substâncias produzidas pelas espécies de Pseudoalteromonas podem inibir ou facilitar as
incrustações de outros organismos e, por isso, este gênero tem um forte potencial para a
estruturação da comunidade (BOWMAN, 2007).
Bactérias do gênero Vibrio são abundantes no ambiente aquático, onde colonizam tecidos
e/ou órgãos de vários animais marinhos, como corais, peixes, moluscos, crustáceos e esponjas.
Por vezes, atuam como agentes patogênicos nestes organismos (THOMPSON, IIDA & SWINGS,
2004). Em esponjas, foram encontradas estirpes de Vibrio capazes de produzir um peptídeo
antibiótico chamado de andrimide com atividade contra espécies do gênero Bacillus (OCLARIT
et al., 1994).
Os gêneros Actinomyces, Corynebacterium e Streptomyces estão amplamente
distribuídos no meio marinho. Eles são considerados importantes produtores de agentes
bioativos, exibindo um arsenal de estratégias para competição por espaço e nutrientes. Várias
espécies dos gêneros citados, associadas às plantas e aos animais marinhos, têm sido relatadas
por expressarem propriedades antibacterianas, antifúngicas e antitumorais (ZHENG et al., 2000).
As bactérias pertencentes ao gênero Aeromonas são móveis e predominantemente de vida
livre. Estirpes de Aeromonas também podem colonizar animais aquáticos, por vezes atuando
como patógenos de peixes e crustáceos (STARLIPER & MORRISON, 2000). Em um trabalho
realizado por Castillo e colaboradores (2001), das 27 estirpes produtoras de substâncias
antimicrobianas isoladas de corais e esponjas, 51,8% eram pertencentes a este gênero.
As espécies marinhas de Bacillus são normalmente isoladas de sedimentos e
invertebrados marinhos (PABEL et al., 2003). As espécies deste gênero são conhecidas por gerar
esporos sob condições adversas, tais como as encontradas nos ecossistemas marinhos
(HENTSCHEL et al., 2001). Interessantemente, a formação de esporos é co-regulada com a

13
produção de antibióticos (MARAHIEL et al., 1993). No ambiente marinho, este gênero tem sido
considerado um dos maiores produtores de metabólitos bioativos (PABEL et al., 2003).
Micrococcus são amplamente distribuídos no meio marinho; eles são normalmente de
vida livre e ficam dispersos na coluna d’água. Eventualmente, podem ser encontrados em
superfícies submersas (MONTALVO et al., 2005). Há poucos relatos sobre seu isolamento em
esponjas, destacando-se uma estirpe de Micrococcus sp. isolada da esponja Tedania ignis,
responsável por produzir peptídeos cíclicos biologicamente ativos (STIERLE, CARDELLINA &
SINGLETON, 1988).
As espécies pertencentes ao gênero Pseudomonas são encontradas em invertebrados
marinhos, sedimentos e na água do mar. São caracterizadas por apresentarem uma ampla
diversidade metabólica, o que permite a sua ocorrência em diferentes ambientes (ROMANENKO
et al., 2005). Nos últimos anos, este gênero tem atraído atenção devido a grande capacidade de
produzir metabólitos secundários de interesse biotecnológico, tais como, antibióticos, enzimas,
fitohormônios e sideróforos (ISNANSETYO & KAMEI, 2003). Em esponjas, já foram descritas
espécies produtoras de substâncias antimicrobianas (JAYATILAKE et al., 1996; DEBITUS et
al., 1998; BULTEL-PONCÉ et al., 1998; ZHENG et al., 2000; SINGH et al., 2003) e novas
espécies, como exemplo, a espécie chamada de Pseudomonas pachastrellae, isolada da esponja
Pachastrella sp. (ROMANENKO et al., 2005).
1.3.2 Produção de Substâncias Bioativas
As esponjas (e suas bactérias associadas) são as maiores produtoras marinhas de novas
substâncias, com mais de 200 novos metabólitos descritos a cada ano (BLUNT et al., 2006).
Além disso, as substâncias derivadas de esponjas estão em triagem clínica e pré-clínica (como
agentes anticâncer e antiinflamatório), mais do que as substâncias de qualquer outro filo marinho

14
(BLUNT et al., 2005). A ocorrência de substâncias estruturalmente similares em esponjas não
relacionadas, particularmente aqueles que são conhecidos por serem produzidos exclusivamente
por microrganismos, sugere que tais substâncias (incluindo algumas que já estão em teste) sejam
produzidas pelas bactérias associadas às esponjas (PIEL, 2004). De fato, vários metabólitos
derivados de esponjas se assemelham a produtos naturais bacterianos ou pertencem às classes de
substâncias típicas destes microrganismos, como, por exemplo, complexos de policetídeos e
peptídeos não-ribossômicos (PIEL, 2004).
Policetídeos (PKs) são metabólitos secundários de bactérias, fungos, plantas e animais.
São encontrados em abundância na natureza e se destacam por apresentarem propriedades
antibiótica, anticancerígena e imunossupressora (Figura 3) (STAUNTON & WEISSMAN, 2001).
Estas substâncias são sintetizadas por enzimas chamadas de sintases de policetídeos (PKSs) que
catalisam a polimerização de subunidades de acetilcoenzima A e propionilcoenzima A, em um
processo similar a síntese dos ácidos graxos. As sintases de policetídeos são classificadas, com
base em sua arquitetura protéica, em três famílias distintas (I, II e III). As sintases do tipo I e II
são proteínas multifuncionais somente encontradas em microrganismos. Contrariamente, as
sintases do tipo III são proteínas homodiméricas pequenas e são exclusivamente encontradas em
plantas (HOPWOOD, 1997). As sintases de policetídeos do tipo I têm atraído especial interesse
devido a sua maior capacidade de sintetizar policetídeos biologicamente ativos (MCDANIEL,
WELCH & HUTCHINSON, 2005).

15
Figura 3. Exemplos de policetídeos. O tipo de policetídeo e sua atividade biológica primária estão indicados na figura (Adaptado de STAUNTON & WEISSMAN, 2001).
A maior parte dos metabólitos secundários bioativos é um peptídeo ou possui subestrutura
peptídica. A maioria desses oligopeptídeos (4 a 10 aminoácidos) supõe-se ser sintetizada pelas
sintetases de peptídeos não-ribossômicos (NRPSs) (WELKER & VON DÖHREN, 2006). Na
síntese ribossômica de peptídeo, a formação das ligações peptídicas é direcionada pelos
ribossomos e o RNA mensageiro (mRNA) funciona como molde determinando a seqüência do
aminoácido do produto. O número de aminoácidos incorporados naturalmente nos peptídeos
ribossômicos está restrito a 20, codificados pelos códons de mRNA. Na síntese não ribossômica
de peptídeos, proteínas modulares com múltiplas atividades catalisam o agrupamento dos
peptídeos e simultaneamente servem como molde. A síntese não-ribossômica utiliza uma grande

16
variedade de substratos, como aminoácidos não-protéicos, hidroxiácidos e substâncias
policetídicas, especialmente elaboradas para serem incorporadas na estrutura destes peptídeos
(SIEBER & MARAHIEL, 2005).
Peptídeos não-ribossômicos (NRPs) formam uma família muito diversificada de produtos
naturais, com uma ampla gama de atividades biológicas e farmacológicas, tais como, antibióticos,
citostáticos, imunossupressores e sideróforos (Figura 4) (WALSH, 2004). Diferentemente dos
peptídeos ribossômicos, os peptídeos não-ribossômicos geralmente apresentam uma estrutura
cíclica extremamente resistente às enzimas proteolíticas (SIEBER & MARAHIEL, 2005).
Figura 4. Exemplos de peptídeos não-ribossômicos. O tipo de peptídeo não-ribossômico e sua atividade biológica primária estão indicados na figura (Adaptado de WALSH, 2004).
Enter obactina Sideróforo
Ciclosporina Agente imunossupressor
Epotilone Agente citostático
Bacitracina Antibiótico polipeptídico
Vancomicina Antibiótico glicopeptídico

17
Um exemplo de substância antimicrobiana produzida por bactérias associadas a esponjas
(Tabela 1) é o antibiótico natural andrimide, uma substância policetídica anti-Bacillus. Oclarit e
colaboradores (1994) relataram sobre o isolamento deste antibiótico, tanto da esponja Hyatella
sp., quanto da espécie de Vibrio sp. isolada deste animal.
Outro exemplo é o isolamento de uma estirpe de Micrococcus sp. da esponja Tedania
ignis, responsável por produzir as mesmas dicetopiperazinas, cyclo-(L-Pro-L-Val), cyclo-(L-Pro-L-
Leu) e cyclo-(L-Pro-L-Ala) (STIERLE, CARDELLINA & SINGLETON, 1988), previamente
isoladas do extrato desta espécie de esponja (SCHMITZ et al., 1983). Dicetopiperazinas (ou
dipeptídeos cíclicos) pertencem à classe dos peptídeos não-ribossômicos e são conhecidas por
apresentarem propriedades antimicrobianas, antitumorais e antivirais (RHEE, 2004). É uma
classe relativamente inexplorada de peptídeos bioativos, que podem vir a apresentar grande
potencial biotecnológico. Jayatilake e colaboradores (1996) fracionaram o meio de cultura de
uma estirpe de Pseudomonas aeruginosa isolada da esponja Isodictya setifera (THORNTON,
1995), e encontraram seis novas dicetopiperazinas e duas fenazinas já descritas.
Surpreendentemente, em seus ensaios, as seis dicetopiperazinas foram inativas como substâncias
antimicrobianas, mas as duas fenazinas foram ativas contra Bacillus cereus (concentração
mínima inibitória (CMI) < 0.5 µg/mL). As substâncias foram menos ativas contra Micrococcus
luteus e Staphylococcus aureus (CMI > 5 µg/mL). Fenazinas pertencem à classe dos alcalóides
(moléculas heterocíclicas contendo nitrogênio), e mais de 60 diferentes derivados desta
substância têm sido identificados. A maioria das fenazinas exibe uma variedade de propriedades
antimicrobianas, especialmente contra bactérias Gram-positivas e fungos patogênicos de plantas
(GIDDENS & BEAN, 2007; LIU et al., 2007).

18
Tabela 1. Exemplos de substâncias antimicrobianas produzidas por bactérias associadas a esponjas.
Classe química da substância
Substância antimicrobiana
Espécie bacteriana produtora
Espécie de esponja Espectro de ação Referência
policetídeos andrimide Vibrio sp. Hyatella sp. Bacillus sp. OCLARIT et al., 1994 peptídeos não-ribossômicos
dicetopiperazinas Micrococcus sp. Tedania ignis Atividade antimicrobiana STIERLE, CARDELLINA
& SINGLETON, 1988 alcalóides fenazinas Pseudomonas aeruginosa Isodictya setifera B. cereus, M. luteus e S. aureus JAYATILAKE et al., 1996
peptídeos não-ribossômicos
tiopeptídeos Bacillus cereus Halichondria
japonica Enterococcus e Staphylococcus
multirresistentes NAGAI et al., 2003;
SUZUMURA et al., 2003
peptídeos não-ribossômicos
lipopeptídeos (fengicinas, iturinas e
surfactinas) Bacillus subtilis
Aplysina aerophoba
A. tumefaciens, B. megaterium, C. michiganensis, E. coli, P. vulgaris, S. aureus, S. epidermidis, S. lentus
e Vibrio sp.
PABEL et al., 2003
piranos alfa-pirano I Pseudomonas sp. Esponja marinha B. subtilis, E. coli, E. faecalis, E.
faecium, M. catarrhalis, S. aureus, e S. pneumoniae
SINGH et al., 2003
policetídeos rifamicina Salinispora sp. Pseudoceratina
clavata Bactérias Gram-positivas
HEWAVITHARANA et al., 2007
Adaptada de LAPORT, SANTOS & MURICY, in press.

19
A família de antibióticos tiopeptídicos pertence à classe de peptídeos não-ribossômicos
que contém uma molécula de enxofre associada a sua estrutura (BAGLEY et al., 2005). Muitos
tiopeptídeos inibem a síntese protéica em bactérias e compartilham modos de ação comuns,
agindo diretamente sobre o ribossomo ou em fatores de alongamento de proteínas (BAGLEY et
al., 2005). Dois tiopeptídeos cíclicos, contendo resíduos de tiazol e piridina e vários aminoácidos
não-usuais foram obtidos a partir de uma cultura de B. cereus isolados da esponja Halichondria
japonica (NAGAI et al., 2003; SUZUMURA et al., 2003). Estas substâncias exibiram potente
atividade antibacteriana contra estirpes de Enterococcus e Staphylococcus resistentes a drogas.
Lipopeptídeos cíclicos (CLPs) pertence à classe de peptídeos não-ribossômicos que
contém uma molécula de lipídeo associada a sua estrutura. São produzidos por diferentes grupos
de bactérias, tanto Gram-positivas quanto Gram-negativas (MOFFITT & NEILAN, 2000) e
representam uma classe única de metabólitos secundários bioativos (HSIEH et al., 2008).
Espécies de Bacillus freqüentemente produzem uma grande variedade destes antimicrobianos,
incluindo fengicinas, iturinas e surfactinas. O interesse em tais substâncias resulta do seu
excepcional poder surfactante e de suas propriedades antibacteriana, antifúngica, anti-
Mycoplasma e antitumoral (BONMATIN, LAPREVOTE & PEYPOUX, 2003). Duas estirpes de
Bacillus pumilus e três estirpes de Bacillus subtilis isolados da esponja A. aerophoba (PABEL et
al., 2003) produziram metabólitos bioativos que foram identificados como lipopeptídeos. Para
caracterizar a atividade antimicrobiana, estas estirpes foram testadas contra um painel de
microrganismos, incluindo estirpes-padrão de bactérias Gram-positivas (S. aureus) e Gram-
negativas (E. coli), estirpes sensíveis e resistentes de S. aureus, Staphylococcus epidermidis e
Staphylococcus lentus, um isolado de esponja (Vibrio sp. SB1), e quatro estirpes ambientais de
referência (Agrobacterium tumefaciens, Bacillus megaterium, Clavibacter michiganensis e
Proteus vulgaris). Dentre as cinco estirpes de Bacillus sp. isoladas, a estirpe A184 de B. subtilis

20
foi a mais versátil na produção de lipopeptídeos (fengicinas, iturinas e surfactinas) e foi ativa
contra todos os microrganismos testados.
Compostos aromáticos e heterocíclicos biologicamente ativos contendo anéis de pirano
anexados estão difundidos na natureza, sendo encontrados em uma ampla variedade de
organismos. Pirano é um anel composto de cinco átomos de carbono e um átomo de oxigênio,
contendo duas duplas ligações. Apesar dos piranos terem pouco significado em química, uma
variedade de seus derivados são moléculas biológicas importantes (DONNER, GILL &
TEWIERIK, 2004). Uma nova espécie de Pseudomonas, isolada de uma esponja marinha das
ilhas Fiji (SINGH et al., 2003), ganhou destaque por produzir dois novos alfa-piranos. Estas
substâncias apresentaram potente atividade antimicrobiana contra bactérias Gram-positivas,
incluindo várias estirpes resistentes a drogas, e fraca atividade contra bactérias Gram-negativas.
Alfa-pirano I, o maior e mais ativo composto produzido por este microrganismo, apresentou
CMIs de 2 a 4 µg/mL contra Enterococcus faecium, Moraxella catarrhalis e S. aureus; 16-64
µg/mL contra Enterococcus faecalis e Streptococcus pneumoniae; e > 128 µg/mL contra E. coli.
A rifamicina faz parte de um grupo de antibióticos chamado de ansamicina (policetídeos),
que é produzido por diferentes actinomicetos (ou actinobacteria) e apresenta destacada atividade
antibacteriana contra bactérias Gram-positivas (DEWICK, 2002). As rifamicinas são conhecidas
por serem produzidas pela espécie Amycolatopsis mediterranei, presente no solo (SCHUPP et al.,
1998). Hewavitharana e colaboradores (2007) utilizaram a técnica de espectrometria de massa
acoplada à cromatografia líquida de alta eficiência (HPLC/MS) para a identificação e
caracterização de rifamicina B e rifamicina SV em várias estirpes da actinobactéria Salinispora
isolada da esponja Pseudoceratina clavata. Todas as linhagens de Salinispora testadas foram
positivas para a produção de rifamicina, estabelecendo, assim, uma fonte alternativa deste
antibiótico.

21
Como a maioria das bactérias simbiontes apresenta difícil cultivo pelas técnicas
tradicionais, é amplamente aceito que as técnicas baseadas em culturas são inadequadas para
estudar a diversidade bacteriana de amostras ambientais (AMANN, LUDWIG & SCHLEIFER,
1995). De fato, a percentagem da comunidade microbiana da esponja cultivável varia entre
espécies e representa 3-11, 0,1-0,23 e 0,15% do total da população bacteriana presente em
Ceratoporella nicholsoni (SANTAVY & COLWELL, 1990), R. odorabile (WEBSTER & HILL,
2001) e A. aerophoba (FRIEDRICH et al., 2001), respectivamente. Estas observações são, em
geral, consistentes com a estimativa de que apenas 1% dos microrganismos é cultivável, no
contexto dos ecossistemas naturais (HENTSCHEL et al., 2003). A utilização de métodos
moleculares para descrever a diversidade microbiana tem fornecido um grande reforço ao
conhecimento sobre comunidades microbianas e também tem sido útil na manipulação
experimental das condições de cultura a fim de fornecer alternativas para o crescimento de
bactérias ambientais (WEBSTER & HILL, 2001; HENTSCHEL et al., 2003). No entanto, tendo
em vista o fato da aquacultura de esponjas ainda ser precoce e poucos avanços terem sido
alcançados em relação a cultura de células de esponjas (MUSCHOLL-SILBERHORN, THIEL &
IMHOFF, 2008), o cultivo bacteriano ainda é essencial para se pesquisar novas substâncias
bioativas. Isto porque as bactérias produzem rapidamente grandes quantidades de biomassa e,
conseqüentemente, grandes quantidades de produtos secundários biologicamente ativos poderiam
facilmente ser produzidas em uma escala biotecnológica, sem a necessidade de se coletar ou
cultivar a esponja (MUSCHOLL-SILBERHORN, THIEL & IMHOFF, 2008).

22
1.4 A Problemática das Bactérias Multirresistentes aos Antimicrobianos
Em 1929, ao descrever o primeiro antibiótico, a penicilina, Fleming foi, também, o
primeiro autor a descrever o fenômeno da resistência bacteriana aos antibióticos, ao demonstrar
que as bactérias do grupo colitifóide (enterobactérias) e o bacilo piociânico (P. aeruginosa) não
sofriam a ação inibitória da penicilina. Esta resistência própria de determinadas espécies
bacterianas a um determinado antibiótico é conhecida como resistência natural e constitui uma
característica biológica das bactérias, não tendo maior importância na terapêutica antimicrobiana,
por ser previsível. Assim, os bacilos Gram-negativos são naturalmente resistentes à penicilina G,
assim como, os cocos Gram-positivos são naturalmente resistentes às polimixinas (DRAGO,
2007).
A grande dificuldade que a terapêutica antimicrobiana encontra nos dias de hoje, e cada
vez mais difundida, é a chamada resistência adquirida, isto é, mudanças na sensibilidade dos
microrganismos a antimicrobianos devido à aquisição de genes de resistência. Esta aquisição de
genes pode ocorrer tanto através de mutações como através de mecanismos que envolvem a troca
genética entre os microrganismos (DRAGO, 2007).
O primeiro notável exemplo da importância e difusão da resistência adquirida entre as
bactérias está relacionado à espécie S. aureus, que, ao início da era da terapia antibiótica,
mostrava-se bastante sensível à ação da penicilina. Com a ampla utilização deste antibiótico,
estes microrganismos adquiriram resistência à sua ação. Atualmente, a resistência do estafilococo
à penicilina G atinge quase 100% das estirpes hospitalares na maioria dos países, inclusive no
Brasil (MARTINS & CUNHA, 2007). Além disso, observou-se que, no Brasil, cerca de 80% dos
estafilococos encontrados na comunidade apresentam elevada resistência a este antibiótico. Tal

23
fato não mais permite a utilização da penicilina no tratamento das infecções estafilocócicas, ainda
que originadas fora do ambiente hospitalar (RIBEIRO et al., 2007).
A terapêutica alternativa para os estafilococos resistentes à penicilina surgiu com a síntese
das penicilinas sintéticas, meticilina e oxacilina, introduzidas na clínica a partir da década de
1960. Contudo, a ampla utilização destas drogas no meio hospitalar trouxe como conseqüência o
surgimento e seleção de estirpes de estafilococos também resistentes a estas penicilinas,
encontrados nos hospitais brasileiros com variações entre 20% e 60% (MARTINS & CUNHA,
2007). Os estudos sobre a sensibilidade dos estafilococos em nosso país demonstraram que, em
grande parte, os S. aureus comunitários mantêm a sensibilidade às cefalosporinas de primeira
geração e à oxacilina, o que permite o uso destas drogas no tratamento de pacientes ambulatoriais
com infecções estafilocócicas (DE MIRANDA et al., 2007). No entanto, já existem registros
indicando o aumento da freqüência dos estafilococos resistentes à oxacilina também no meio
extra-hospitalar, como relatado recentemente por Martins & Cunha (2007).
Ressalta-se que os estafilococos resistentes à oxacilina sejam também resistentes às
cefalosporinas de primeira geração (as cefalosporinas de 2ª, 3ª e 4ª gerações são menos ativas
sobre bactérias Gram-positivas), restando como alternativa terapêutica para as infecções graves
causadas por estas bactérias o uso dos antibióticos glicopeptídicos (teicoplanina e vancomicina),
das estreptograminas e das oxazolidinonas (JOHN & HARVIN, 2007). No entanto, já se
descrevem, desde 1996, estirpes de S. aureus com sensibilidade diminuída à vancomicina (CDC,
1997) e, mais recentemente (2002), com ampla resistência aos glicopeptídeos em uso clínico
(CDC, 2002), restando como alternativa terapêutica atual as estreptograminas e as
oxazolidinonas (KUTI et al., 2008). Como agente complicador, outras espécies de estafilococos,
como S. epidermidis, S. haemolyticus e S. hominis, também são resistentes à meticilina e atuam

24
como reservatórios de genes de resistência, transferindo-os para S. aureus e tornando-o
multirresistente às drogas (MARTINS & CUNHA, 2007).
Ainda entre as bactérias Gram-positivas, consistem em bactérias com elevado perfil de
resistência os Enterococcus e os pneumococos resistentes às penicilinas, que por vezes exigem o
emprego da vancomicina em situações de gravidade. No entanto, foi registrada em diferentes
países a ocorrência de infecções por enterococos também resistentes aos glicopeptídeos, o que
exigiu alternativas terapêuticas como as oxazolidinonas e a pesquisa envolvendo novas drogas,
tais como a dalbavancina, a daptomicina e a oritavancina (BRANDILEONE et al., 2006;
VILELA et al., 2006; POULAKOU & GIAMARELLOU, 2008).
A resistência dos bacilos Gram-negativos aos antibióticos beta-lactâmicos e
aminoglicosídeos estimulou a pesquisa sobre novos antimicrobianos, resultando no
desenvolvimento das carbapenemas, de fluoroquinolonas e de associações de drogas beta-
lactâmicas com substâncias inibidoras de beta-lactamases ativas sobre estes microrganismos
(RICE, 2007). Observou-se, porém, que o uso implementado intra-hospitalar destes novos
antimicrobianos resultou no surgimento e seleção de microrganismos resistentes também à sua
ação, com a descrição de estirpes de Acinetobacter baumanii/calcoaceticus, Enterobacter,
Klebsiella e P. aeruginosa com altos níveis de resistência para todos os antibióticos em uso
clínico (FALAGAS, 2007).
Entre os bacilos Gram-negativos entéricos e os não-fermentadores, a resistência às drogas
antimicrobianas constitui um problema grave em todos os países. Como por exemplo,
freqüentemente observa-se a resistência de E. coli à ampicilina tanto no ambiente hospitalar
quanto na comunidade (FALAGAS & KARAGEORGOPOULOS, 2008). Já a resistência desta
bactéria aos aminoglicosídeos, às carbapenemas e às fluoroquinolonas sofre variações conforme a
origem hospitalar ou comunitária do microrganismo e, mesmo numa determinada região, existe

25
variabilidade na sensibilidade entre as estirpes hospitalares (BRISTIANOU et al., 2008). Estas
diferenças são resultantes de uma maior ou menor pressão de seleção das estirpes resistentes pelo
emprego dos antimicrobianos no ambiente hospitalar (DRAGO, 2007).
A pressão de seleção de microrganismos resistentes exercida pelos antimicrobianos é
conseqüência não só do uso terapêutico ou profilático destas drogas em medicina e odontologia
humanas, como também de seu emprego ou de derivados ou análogos na conservação de
alimentos, no combate a elementos biológicos daninhos aos seres humanos e na engorda e
tratamento de animais destinados à alimentação (MOULIN et al., in press). Exemplo desta última
assertiva é referido por Oliveira e colaboradores (2002), que verificaram que estirpes de
Salmonella isoladas de porcos apresentaram alta resistência ao ácido nalidíxico, estreptomicina e
tetraciclina. Foi observado que estes antimicrobianos eram comumente utilizados para o
tratamento de infecções nestes animais (OLIVEIRA et al., 2002).
O tratamento de infecções por microrganismos resistentes a drogas pode ser realizado por
diversos modos, destacando-se nos hospitais o conhecimento da microbiota do paciente, do sítio
de infecção e o uso adequado e prudente de antibióticos (CEGELSKI et al., 2008).
Paralelamente, a pesquisa de novos antimicrobianos vem se tornando imprescindível, pois
amplia as perspectivas quanto ao desenvolvimento de novos agentes para compor o arsenal
terapêutico contra microrganismos multirresistentes (BUTLER & BUSS, 2006; LUZHETSKYY,
PELZER & BECHTHOLD, 2007). A biodiversidade brasileira fornece os subsídios para a busca
dessas substâncias na natureza, incluindo fontes como as esponjas marinhas e suas bactérias
associadas, onde espécies raras ou novas podem apresentar grande potencial para o
desenvolvimento de novas gerações de agentes antibióticos (HADJU & LOPES, 2007).

26
Neste contexto, estirpes bacterianas isoladas de esponjas marinhas coletadas no litoral do
Rio de Janeiro foram analisadas quanto a capacidade de produzir substâncias antimicrobianas
que inibissem o crescimento de bactérias de importância médica.

27
2 OBJETIVOS
2.1 Objetivo Geral
O objetivo geral deste trabalho foi caracterizar bactérias produtoras de substâncias
antimicrobianas isoladas a partir de 10 espécies de esponjas coletadas na costa da cidade do Rio
de Janeiro.
2.2 Objetivos Específicos
1) Isolar bactérias cultiváveis a partir das esponjas pertencentes às espécies Arenosclera
brasiliensis, Clathrina aurea, Dragmacidon reticulatus, Geodia corticostylifera, Haliclona spn.,
Mycale microsigmatosa, Paraleucilla magna, Petromica citrina, Polymastia janeirensis e
Tedania ignis;
2) Verificar estirpes bacterianas associadas às esponjas com potencial produção de substância(s)
antimicrobiana(s);
3) Identificar e caracterizar por técnicas convencionais e moleculares as estirpes bacterianas
produtoras de substância(s) antimicrobiana(s);
4) Caracterizar parcialmente a(s) substância(s) antimicrobiana(s) quanto à natureza química;
5) Analisar as condições ótimas de temperatura e salinidade para a produção da(s) substância(s)
antimicrobiana(s).

28
3 MATERIAIS E MÉTODOS
3.1 Meios de cultura
Neste trabalho foram utilizados os seguintes meios de cultura:
• Meio BHI: Brain Heart Infusion (Difco, Detroit, Michigan, USA). Para estoque bacteriano, o
meio BHI foi utilizado em caldo acrescido de glicerol a 30% (v/v) e sólido inclinado; para os
testes de produção de substância antimicrobiana (SAM), foi utilizado em caldo, sólido [acrescido
de ágar a 1,5% (p/v)] e semi-sólido [acrescido de ágar a 0,7% (p/v)]; para testar as condições
ótimas de crescimento e produção das substâncias antimicrobianas foi utilizado em caldo e sólido
acrescidos de NaCl numa concentração final de 1, 1,5 e 2%.
• Meio Marine: Marine Broth 2216 (Difco). Para estoque bacteriano, o meio Marine foi
utilizado em caldo acrescido de glicerol a 30% (v/v) e sólido inclinado; para os testes de
produção de SAM foi utilizado em caldo, sólido [acrescido de ágar a 1,5% (p/v)] e semi-sólido
[acrescido de ágar a 0,7% (p/v)]; para testar as condições ótimas de crescimento e produção das
substâncias antimicrobianas foi utilizado em sua forma sólida. Este meio também foi utilizado em
sua forma sólida nos testes de susceptibilidade aos antimicrobianos, devido ao fato das estirpes
marinhas isoladas a partir deste meio não terem crescido no meio de cultura padronizado para o
antibiograma. Neste trabalho, o meio chamado de Marine foi dissolvido em água destilada e o
meio chamado de Marine-mar foi dissolvido em água do mar estéril. Este último foi padronizado
por nosso grupo.
• Meio Müeller-Hinton Ágar (Difco): Este meio foi utilizado nos testes de susceptibilidade aos
antimicrobianos para as estirpes marinhas isoladas a partir do meio BHI.

29
• Meio de Motilidade: MTM (Motility Test Medium – Difco) a 1,6% (p/v); caldo nutriente
(Nutrient Broth – Difco) a 0,4% (p/v); NaCl a 0,1% (p/v). Este meio foi utilizado nos testes de
motilidade das estirpes marinhas SAM-positivas.
• Meio Ágar Cetrimide (Merck, Whitehouse Station, New Jersey, USA): Este meio foi
utilizado para o isolamento de estirpes marinhas pertencentes ao gênero Pseudomonas.
3.2 Espécies de esponjas
As espécies das esponjas Arenosclera brasiliensis, Clathrina aurea, Dragmacidon
reticulatus, Geodia corticostylifera, Haliclona spn., Mycale microsigmatosa, Paraleucilla
magna, Petromica citrina, Polymastia janeirensis e Tedania ignis foram coletadas em abril de
2006 no Arquipélago das Cagarras (23° 01’ S – 43° 11’ L), e em janeiro e março de 2007 na
Praia Vermelha (22° 57’ S – 43° 09’ L), Rio de Janeiro. A coleta e identificação dos espécimes
foram realizadas pelo professor Guilherme Muricy, do laboratório de Poríferas, Departamento de
Invertebrados, Museu Nacional – UFRJ.
3.3 Coleta dos espécimes e preparação dos macerados brutos das esponjas
A coleta das espécies de esponjas foi realizada através de mergulhos em locais não muito
profundos, em torno de 1-6m de profundidade. Os espécimes foram retirados manualmente das
esponjas associadas a sedimentos (principalmente rochas) e colocados em sacos plásticos,
geralmente um espécime por saco. Ao chegar no barco, os espécimes foram lavados rapidamente
em água destilada estéril e cerca de 3-10g foram colocados em triplicata em tubos falcon
contendo 10ml dos meios BHI, Marine e Marine-mar acrescidos de anfotericina B (1mg/ml,
preparada em DMSO, Sigma, Saint Louis, Missouri, USA). No laboratório, os espécimes foram

30
triturados em seus respectivos caldos com o auxílio de um bastão de vidro estéril a fim de se
obter o macerado bruto.
3.4 Isolamento de estirpes bacterianas associadas às esponjas e condições de
cultura
Cerca de 20µl dos macerados brutos foram inoculados em duplicata em placas contendo
três diferentes meios de cultura: BHI, Marine e Marine-mar, após 24h, 48h e 72h da coleta.
Adicionalmente, na 5ª coleta utilizou-se o meio Ágar Cetrimide visando favorecer o isolamento
de estirpes marinhas pertencentes ao gênero Pseudomonas sp. As culturas foram mantidas por até
sete dias à temperatura ambiente (em torno de 25-30°C) e examinadas diariamente, observando-
se o tempo de crescimento e a morfologia colonial. As colônias foram isoladas de acordo com os
aspectos morfológicos e, quando purificadas, foram estocadas nos seus respectivos meios de
isolamento (BHI, Marine e Marine-mar), nas formas sólida e inclinada a 4°C e líquida adicionada
de glicerol a -20°C.
3.5 Ensaios de produção de substância antimicrobiana (SAM):
Os ensaios foram realizados como descrito por Giambiagi-deMarval e colaboradores
(1990), com modificações.
As estirpes marinhas isoladas foram cultivadas em caldos de seus respectivos meios de
isolamento (BHI, Marine e Marine-mar) à temperatura ambiente, com tempos de crescimento
variando entre 24 e 48h. Com o auxílio de uma micropipeta, dez microlitros de cada cultura
foram inoculados sob a forma de pontos sobre a superfície de seus respectivos meios e crescidos
à temperatura ambiente por 48h. A seguir, as estirpes marinhas foram mortas por exposição, por

31
cerca de 30min, a vapores de clorofórmio. As placas foram mantidas a 37oC por 30min, a fim de
que todo clorofórmio fosse evaporado, e a seguir, foram vertidos, sobre a superfície do mesmo
meio, 3ml de BHI semi-sólido contendo as estirpes indicadoras (30µl de cultura em fase
estacionária de crescimento; cada estirpe indicadora foi crescida previamente em caldo BHI a
37°C por 18h). As placas foram incubadas a 37oC por 18h e, posteriormente, a produção de SAM
foi verificada através da medição dos halos de inibição de crescimento da estirpe indicadora.
Somente foram consideradas estirpes marinhas SAM-positivas, isto é, produtoras de substâncias
antimicrobianas, aquelas que apresentaram halos com diâmetros maiores do que 10mm.
3.5.1 Triagem de estirpes produtoras de SAM associadas às esponjas
Para o rastreamento inicial das estirpes marinhas SAM-positivas, foi utilizada a estirpe
Corynebacterium fimi NCTC 7547. Após o rastreamento, as estirpes marinhas SAM-positivas
foram testadas contra estirpes-padrão representativas de microrganismos Gram-positivos
(Staphylococcus aureus ATCC 29213 e Enterococcus faecium ATCC 19434) e Gram-negativos
(Escherichia coli ATCC 25922 e Pseudomonas aeruginosa ATCC 27853).
3.5.2 Determinação do espectro de ação das SAM
Baseado nos resultados dos testes contra estirpes-padrão representativas de
microrganismos Gram-positivos e Gram-negativos, as metodologias descritas no item 3.5 foram
repetidas com um número maior de estirpes indicadoras como descrito no subitem 3.5.1. As
estirpes usadas como indicadoras (Tabela 2) foram estirpes-padrão e de origem clínica, as quais
foram previamente caracterizadas quanto aos perfis de susceptibilidade aos antimicrobianos pelos
laboratórios de origem.

32
Tabela 2. Estirpes bacterianas que foram utilizadas como indicadoras na determinação do
espectro de ação das estirpes marinhas SAM-positivas.
Estirpes-padrão Características Relevantes Referências Staphylococcus epidermidis
ATCC 12228 ----- ATCC
Staphylococcus haemolyticus CCM 2737 ----- CCM
Staphylococcus hominis ATCC 27844
----- ATCC
Acinetobacter sp. ATCC 14293
----- ATCC
Enterobacter cloacae ATCC 23355
Produz cefalosporinase (beta-lactamase II)
ATCC
Klebsiella pneumoniae ATCC 700603
Resistente à ampicilina, aztreonam, cefoxitina, cefpodoxima, ceftazidima,
cloranfenicol, piperaciclina, tetraciclina; ESBL+
ATCC
Estirpes clínicas Enterococcus faecalis V583 Resistente à vancomicina 2 Enterococcus faecalis 5AE Resistente à vancomicina 1
Staphylococcus aureus 42AE Resistente à ampicilina, meticilina,
penicilina (MRSA) 1
Staphylococcus aureus 559a SCCmecIV, PVL+ (CA-MRSA) 1
Staphylococcus epidermidis 57s Resistente à ampicilina,
ciprofloxacina, penicilina, tetraciclina 1
Staphylococcus haemolyticus 109s Resistente à ampicilina, canamicina, gentamicina, oxacilina, penicilina
1
Staphylococcus hominis 79s Resistente à ampicilina, penicilina 1 Acinetobacter baumanii 76AE ---- 1
Acinetobacter calcoaceticus 56AE ---- 1 Citrobacter freundii 27AE ---- 1
Enterobacter cloacae 43AE Resistente à ampicilina, cefalotina,
cefoxitina, tetraciclina 1
Enterobacter hafniae 30AE ---- 1
Escherichia coli 54AE Resistente à ampicilina, cloranfenicol,
sulfametoxazol/trimetoprim, tetraciclina
1
Klebsiella pneumoniae 52AE ---- 1 Klebsiella pneumoniae 19AE ESBL+ 1
Pseudomonas aeruginosa 3AE Resistente à aztreonam, piperacilina/tazobactam
1
Salmonella 4AE ---- 1 Serratia marcescens 38AE ---- 1
Stenotrophomona maltophilia 17AE
---- 1
MRSA: Staphylococcus aureus resistentes à meticilina; CA-MRSA: Staphylococcus aureus resistentes à meticilina associados a comunidade; SCCmecIV: cassete cromossômico de estafilococos mec tipo IV; PVL: Panton-Valentine leucocidina; ESBL: beta-lactamases de espectro estendido; ----: o perfil de susceptibilidade aos antimicrobianos não foi analisado pelo laboratório de origem; 1. Coleção do Laboratório de Infecção Hospitalar, IMPPG, UFRJ; 2. Estirpe seqüenciada pelo “The Institute for Genomic Research (TIGR)” (http://cmr.tigr.org/cgi-bin/CMR/GenomePage.cgi?org=gef).

33
3.5.3 Teste de inibição cruzada
O teste de inibição cruzada foi realizado com o objetivo de observar os tipos de interações
que ocorrem entre estirpes marinhas produtoras de substâncias antimicrobianas pertencentes ao
mesmo nicho ecológico. Este experimento foi feito conforme descrito no item 3.5, sendo que, as
estirpes marinhas SAM-positivas foram utilizadas como produtoras e/ou indicadoras.
3.6 Caracterização fenotípica das estirpes marinhas SAM-positivas
Os testes para caracterização das estirpes marinhas que apresentaram atividade contra
estirpes de importância médica foram realizados de acordo com as metodologias descritas por
MacFaddin (2000) e Bannerman & Peacock (2007). Os testes utilizados foram:
3.6.1 Aspectos fenotípicos
O crescimento das estirpes marinhas SAM-positivas foi acompanhado por sete dias à
temperatura ambiente e examinado diariamente, observando-se os aspectos fenotípicos como
tempo de crescimento, coloração, tipo de colônia (mucóide ou rugosa) e produção de pigmento
no meio.
3.6.2 Aspecto morfo-tintorial
Uma colônia bacteriana foi transferida para uma lâmina de vidro e o seu aspecto morfo-
tintorial foi analisado através da coloração de Gram. A interpretação foi realizada através de
microscopia óptica, observando-se coloração, morfologia, arranjo das células bacterianas e
presença de esporos.

34
3.6.3 Produção da enzima catalase
Uma colônia bacteriana foi transferida para uma lâmina de microscopia e recoberta com
uma gota de peróxido de hidrogênio (Proquímios, Rio de Janeiro, Brasil) a 3% (v/v). A formação
de bolhas foi indicativa de reação positiva. As estirpes S. epidermidis ATCC 14990 e E. faecalis
ATCC 29212 foram utilizadas como controles positivo e negativo, respectivamente.
3.6.4 Produção da enzima citocromo-oxidase
A determinação da produção da enzima citocromo-oxidase foi realizada seguindo as
recomendações do fabricante do sistema Bactident oxidase (Merck, Darmstadt, Alemanha). Uma
colônia bacteriana, crescida em seu respectivo meio, foi aplicada na fita teste. No caso de estirpes
citocromo-oxidase positivas, a zona de reação apresentou uma coloração azul-violeta. Como
controle negativo foi utilizada a estirpe E. coli ATCC 35218 e como controle positivo a estirpe P.
aeruginosa ATCC 27853.
3.6.5 Teste de motilidade
Uma colônia bacteriana foi repicada horizontalmente com auxílio de uma agulha
bacteriológica em até ¾ do tubo contendo o meio de motilidade. A motilidade bacteriana foi
observada diretamente a partir do exame dos tubos após o período de incubação, que variou entre
24 a 48h. As estirpes móveis apresentaram um crescimento difuso que se estendeu lateralmente a
partir da linha de inoculação. As estirpes E. coli ATCC 25922 e K. pneumoniae ATCC 700603
foram utilizadas como controle positivo e negativo, respectivamente.

35
3.6.6 Análise das condições de temperatura e salinidade para o crescimento das estirpes
marinhas SAM-positivas
As estirpes marinhas SAM-positivas foram analisadas quanto à capacidade de crescimento
a 15°C, 37ºC e 42°C, nos seus respectivos meios de cultivo.
As estirpes marinhas SAM-positivas isoladas em meio BHI foram testadas quanto à
capacidade de crescimento em concentrações de 1, 1,5 e 2% de NaCl. A concentração de 2% de
NaCl adicionado ao meio BHI correspondeu a condição de salinidade do meio Marine.
3.6.7 Identificação das estirpes marinhas SAM-positivas através do sistema comercial
miniaturizado
O sistema comercial API (bioMérieux, Marcy l'Etoile, France) foi utilizado de acordo
com as determinações do fabricante. Para identificação das estirpes marinhas Gram-positivas foi
utilizado o sistema API 50CH, constituído por 49 testes bioquímicos utilizados para o estudo da
fermentação de substratos, pertencentes à família dos hidratos de carbono e derivados (ácidos
urônicos, heterosídeos e polialcoois). O sistema miniaturizado foi inoculado com uma suspensão
bacteriana de turvação equivalente a 0,5 de McFarland em meio fornecido pelo fabricante. Após
o período de incubação a 32°C por 24-48h foram feitas as leituras e interpretação. A identificação
foi feita com o auxílio de um algoritmo de identificação fornecido pelo fabricante.
Para identificação das estirpes marinhas Gram-negativas não fermentadoras foi utilizado
o sistema API 20NE, que combina oito testes convencionais e 12 testes de assimilação. O sistema
foi inoculado com uma suspensão bacteriana de turvação equivalente a 0,5 de McFarland em
meios fornecidos pelo fabricante. Após o período de incubação à temperatura ambiente por 24-
48h foram feitas as leituras. A identificação foi feita com o auxílio do catálogo analítico
fornecido pelo fabricante.

36
3.6.8 Susceptibilidade aos antimicrobianos
O critério usado para a escolha dos antibióticos foi em função dos aspectos morfo-
tintoriais observados pela coloração de Gram. Para as estirpes Gram-positivas, foram escolhidos
discos contendo os seguintes antibióticos: cefalexina (30µg), ciprofloxacina (5µg), cloranfenicol
(30µg), eritromicina (15µg), gentamicina (10µg), penicilina G (10 UI), sulfazotrim
(sulfametoxazol-trimetoprim) (25µg) e tetraciclina (30µg). Para as estirpes Gram-negativas,
foram escolhidos os discos contendo os seguintes antibióticos: aztreonam (30µg), cefalexina
(30µg), ceftazidima (30µg), ciprofloxacina (5µg), cloranfenicol (30µg), gentamicina (10µg),
sulfazotrim (25µg) e tetraciclina (30µg).
Os procedimentos foram realizados conforme indicado pelo “Clinical and Laboratory
Standards Institute” (CLSI, 2008) com modificações. Uma suspensão bacteriana com uma
turbidez equivalente a 0,5 da escala de McFarland foi preparada em salina 0,85% e semeada
confluentemente com o auxílio de um “swab” em ágar Müeller-Hinton. Posteriormente, foi
depositado sobre o meio discos impregnados com os antibióticos. Os diâmetros das zonas de
inibição foram medidos após 24h de incubação à temperatura ambiente.
Excepcionalmente, para as estirpes marinhas isoladas em meio Marine, este teste foi
realizado neste mesmo meio de cultura, com 48h de incubação à temperatura ambiente.
Embora os diâmetros das zonas de inibição de crescimento das estirpes marinhas tenham
sido medidos, o manual do CLSI não dispõe de dados para comparação, pois estas estirpes não
pertencem às espécies que são mais prevalentes na clínica. Foram consideradas resistentes a um
determinado antibiótico, aquelas estirpes capazes de crescer ao redor do disco (sem halo visível).

37
3.7 Identificação molecular das estirpes marinhas SAM-positivas
As estirpes marinhas SAM-positivas foram identificadas molecularmente baseando-se no
seqüenciamento parcial do gene que codifica para o 16S rRNA (WEISBURG et al., 1991).
3.7.1 Obtenção de DNA total com guanidina
O DNA total das estirpes marinhas SAM-positivas foi extraído através da técnica de
extração com guanidina, conforme descrito por Pitcher, Saunders e Owen (1989), com
modificações. As estirpes marinhas foram inoculadas em 3ml de caldo BHI, Marine ou Marine-
mar e foram incubadas à temperatura ambiente em um período de 24 a 48h, dependendo do
tempo de crescimento da estirpe. Do caldo de cultura foi retirado 1ml que foi transferido para um
microtubo. Essa suspensão bacteriana foi centrifugada a 3000xg por 5min (microcentrífuga
Fanem, modelo 213), o sobrenadante foi desprezado e o sedimento suspenso em 1ml de tampão
TE 1X (Tris-HCl 10mM, pH 7,8; EDTA 1mM, pH 7,8). A cultura foi novamente centrifugada a
3000xg por 5min, o sobrenadante foi desprezado e o sedimento ressuspendido em 100µl de
tampão TE 1X contendo lisozima (1mg/ml – Amersham Pharmacia Biotech, Stockholm,
Sweden), RNAse (1mg/ml – Sigma) e Brij 35 [0,5% (v/v) – Sigma]. Esta suspensão foi incubada
a 37°C por 1h. Decorrido este tempo, foram adicionados 500µl de tiocianato de guanidina (5M –
Promega, Madison, Wisconsin, USA). O tubo foi agitado manualmente e, posteriormente,
incubado à temperatura ambiente por 7min. Esta etapa foi seguida por um resfriamento do lisado
em gelo por 10min e adição de 250µl de acetato de amônio (7,5M – Reagen, Milpitas, Califórnia,
USA). O tubo foi invertido cinco vezes e incubado no gelo por 10min. A este lisado foram
adicionados 500µl de clorofórmio-álcool isoamílico [24:1 (v/v) – Merck]. O tubo foi agitado
manualmente e, posteriormente, centrifugado a 12.500xg por 12min. A fase aquosa foi, então,

38
transferida para um novo microtubo e o DNA precipitado com 0,54 volumes de isopropanol
(Merck) gelado. A solução foi invertida gentilmente até a visualização de um precipitado branco
fibroso. Depois, foi feita uma centrifugação a 12.500xg por 5min e o sobrenadante foi, então,
desprezado. O sedimento foi lavado 5 vezes com 100µl de etanol [70% (v/v) – Merck] gelado. O
sobrenadante foi, então, desprezado e o sedimento dissolvido em 50µl de tampão TE 1X. O DNA
obtido foi mantido a 4°C até o momento de uso.
3.7.2 Reação de polimerase em cadeia (PCR)
As reações de PCR ocorreram em um termociclador com controle de temperatura
programável (PTC-100; MJ Ressearch, Inc.). A mistura da reação (volume final de 50µl) foi
constituída por uma solução contendo os quatro desoxinucleotídeos 5’-trifosfatados (2,5mM de
cada dNTP) (Biotools, Valle de Tobalina, Madrid, Spain), tampão 10X da enzima (KCl 50mM;
Tris-HCl 20mM, pH 8,4 – Biotools), 1,5mM de MgCl2 (Biotools), 100ng de DNA (extraído
conforme descrito no subitem 3.7.1), 1,5U de Taq DNA-polimerase (Biotools) e 20pmol dos
iniciadores universais 9f [5’-GAGTTTGATCCTGGCTCAG-3’] e 1492r [5’-
GGYTACCTTGTTAACGACTT-3’], descritos por Weisburg e colaboradores (1991).
O protocolo para a amplificação do gene que codifica para o 16S rRNA foi padronizado
em nosso laboratório e compreendeu uma etapa inicial de aquecimento a 95oC por 4min, para a
desnaturação do DNA, seguida por 40 ciclos de 95oC por 45seg (desnaturação), 57oC por 45seg
(anelamento) e 72oC por 1min (extensão), e uma extensão final a 72oC durante 5min. O produto
da PCR, com tamanho em torno de 1,4kb, foi purificado empregando-se o sistema comercial
QIAquick – PCR Purification Kit (Qiagen, Germantown, Maryland, USA).

39
3.7.3 Eletroforese de DNA em gel de agarose
O produto de amplificação da PCR foi visualizado através de eletroforese em gel de
agarose de acordo com Sambrook e Russell (2001). O gel de agarose 0,8% (p/v) foi preparado em
tampão TAE 1X (Tris 40mM, pH 7,8; ácido acético 20mM; EDTA 1mM, pH 7,8). Às
preparações de DNA, foram adicionados 1/5 do volume de tampão de amostra [sacarose 60%
(p/v); EDTA 0,1M, pH 7,8; azul de bromofenol 0,2% (p/v)], e estas foram submetidas à
eletroforese em cuba horizontal (Gibco, modelo Horizon 11–14) a 60V, em tampão TAE 1X, por
aproximadamente 40min. Após a corrida, o gel foi corado com uma solução de brometo de etídio
(0,5µg/ml) durante 30min. Posteriormente, o gel foi observado através do emprego de um
transiluminador de luz UV e a imagem foi documentada pelo Sistema de Fotodocumentação Doc-
PrintII (Vilber Lourmat). O padrão de tamanho molecular “1-kb DNA ladder” (Invitrogen,
Carlsbad, California, USA) foi utilizado para mensuração comparativa do tamanho do produto
amplificado.
3.7.4 Seqüenciamento parcial do gene que codifica para o 16S rRNA
O seqüenciamento foi realizado na Unidade de Genômica do Instituto de Biofísica da
Universidade Federal do Rio de Janeiro utilizando-se o seqüenciador “ABI PRISM 3.100 Genetic
Analyzer” (Applied Biosystems, Foster City, USA), o iniciador universal 9f (descrito no subitem
3.7.2) e o kit de reação “Big Dye Terminator 3.1 Cycle Sequencing Ready Reaction” (Applied
Biosystems, Foster City, USA). As seqüências obtidas, com tamanhos em torno de 700pb, foram
identificadas através da comparação com seqüências homólogas presentes no banco de dados do
GenBank, pela ferramenta de procura “BLAST search”, disponível no endereço eletrônico
www.ncbi.nlm.nih.gov/BLAST.

40
3.8 Caracterização parcial da(s) substância(s) antimicrobiana(s) quanto à
natureza química
Para se identificar preliminarmente a natureza química das substâncias antimicrobianas
produzidas pelas estirpes marinhas, as enzimas fosfolipase C, pronase, protease, proteinase K e
tripsina (Sigma, 1mg/ml, preparadas em Tris-HCl 50mM, pH 8,0; CaCl2 10mM), e NaOH 0,2N
(preparado em água) foram utilizados conforme descrito por Giambiagi-deMarval e
colaboradores (1990). A caracterização química foi realizada como no teste da produção de
SAM, detalhado anteriormente no item 3.5, sendo que, antes de se verter a estirpe indicadora C.
fimi NCTC 7547, foram adicionados 40µl da solução para a qual a sensibilidade da SAM foi
analisada, ao redor da região de crescimento da estirpe marinha SAM-positiva. Posteriormente, as
placas foram incubadas a 37°C por 4h e a estirpe indicadora foi vertida, dando continuidade ao
procedimento. A redução de 50% ou ausência de halo de inibição com a SAM tratada com
protease, pronase, proteinase K ou tripsina indicou uma natureza protéica. Quando tratada com a
fosfolipase C, indicou uma natureza lipídica e, quando tratada com NaOH, indicou que a SAM
poderia ter natureza ácida.
Em todos os experimentos descritos para a caracterização da substância antimicrobiana,
como controle foi mantida na mesma placa a estirpe produtora sem tratamento enzimático ou
adição de solução de NaOH.
3.9 Análise das condições de temperatura e salinidade para a produção da(s) substância(s)
antimicrobianas(s).
As estirpes marinhas SAM-positivas foram analisadas quanto à capacidade de produzir a
substância antimicrobiana a 15°C, 37ºC e 42°C, nos seus respectivos meios de cultivo. Essas

41
análises foram realizadas de acordo com o protocolo descrito no item 3.5. Os tamanhos dos halos
de inibição de crescimento da estirpe indicadora C. fimi obtidos nestes testes foram medidos e
comparados com aqueles observados à temperatura ambiente.
As estirpes marinhas SAM-positivas isoladas em meio BHI foram testadas quanto à
capacidade de produzir a substância antimicrobiana em concentrações de 1, 1,5 e 2% de NaCl.
Essas análises foram realizadas de acordo com o protocolo descrito no item 3.5. Os tamanhos dos
halos de inibição de crescimento da estirpe indicadora C. fimi obtidos nestes testes foram
medidos e comparados com aqueles obtidos a partir de uma concentração de 0,5% de NaCl
(condição de salinidade do meio BHI).
3.10 Organograma das atividades
O trabalho experimental do presente estudo foi desenvolvido conforme organograma
apresentado na Figura 5.

42
Coleta das esponjas
Incubação à temperatura ambiente por 24h, 48h ou 72h.
Rastreamento com C. fimi
Estirpes-padrão S. aureus, E. faecium, E. coli, P.areuginosa
Determinação do espectro de ação
Inibição cruzada
Caracterização Fenotípica
Identificação pelo seqüenciamento do gene que codifica para o 16S
rRNA
Natureza química Temperatura Salinidade
Morfologia Gram
Catalase Oxidase
Motilidade Temperatura Salinidade
Antibiograma
API 50 CH API 20 NE
Caracterização da SAM
Seleção das estirpes marinhas que foram ativas contra estirpes de importância médica
Figura 5. Organograma das atividades realizadas no presente estudo.
Semeadura em ágar-BHI, ágar-Marine, ágar-Marine-mar e ágar Cetrimide
Observação das placas por 7 dias à temperatura ambiente
Isolamento baseado na morfologia colonial, purificação e estoque
Teste de SAM
Espécimes triturados em 10ml de caldo BHI, Marine e Marine-mar (macerados brutos)

43
4 RESULTADOS
4.1 Isolamento de bactérias associadas às esponjas
Um total de 168 unidades formadoras de colônias (UFC) foram isoladas dos macerados
brutos das esponjas. Dentre estas, 26 UFC foram isoladas a partir de Haliclona spn., 25 UFC
de D. reticulatus, 24 UFC de M. microsigmatosa, 22 UFC de T. ignis, 16 UFC de P.
janeirensis, 15 UFC de P. magna, 12 UFC de P. citrina, 10 UFC de A. brasiliensis, 10 UFC
de C. aurea e 8 UFC de G. corticostylifera (Figura 6).
Figura 6. Total de UFC isoladas das espécies de esponjas avaliadas neste trabalho.
Total de UFC isoladas de esponjas
26 25 24
22
16 15
12 10
10
8
0
5
10
15
20
25
30
Espécies de Esponjas
Haliclona spn.
D. reticulatus
M. microsigmatosa T. ignis
P. janeirensis
P. magna
P. citrina
A. brasiliensis C. aurea
G. corticostylifera
N° d
e U
FC
isol
adas

44
Quanto aos três principais meios (BHI, Marine e Marine-mar) de cultura utilizados,
não houve diferença significativa entre o número total de UFC isoladas, pois 60 UFC foram
obtidas a partir do meio BHI, 55 UFC do meio Marine e 53 UFC do meio Marine-mar. Dentre
as 168 UFC isoladas, 149 permaneceram viáveis após a segunda passagem em meio de cultura
(Figura 7).
Figura 7. Isolamento de estirpes associadas às esponjas. Total de UFC isoladas (azul escuro) e de UFC que se mantiveram viáveis (azul claro) após a segunda passagem em meios de cultura. M – meio Marine; MM – meio Marine-mar.
UFC isoladas do total de esponjas por meio de cultura utilizado
53
55
60
168
51
47
51
149
0 50 100 150 200
TOTAL
BHI
M
MM
Mei
os
de c
ultiv
o d
as
est
irpes
Número de UFC isoladas
Total Viáveis

45
Em relação ao meio Cetrimide, 10 UFC foram isoladas. Dentre estas, 5 UFC foram
isoladas a partir dos macerados brutos das esponjas A. brasilienses, G. corticostylifera,
Haliclona spn. (Figura 8) e T. ignis preparados em meio BHI. Três UFC foram isoladas a
partir do macerado bruto da esponja Haliclona spn. preparado em meio Marine, enquanto que
2 UFC foram isoladas a partir do macerado bruto da mesma esponja preparado em meio
Marine-mar.
Figura 8. Foto da estirpe H51 isolada em meio Cetrimide a partir do macerado bruto preparado com a esponja Haliclona spn. em meio BHI.
O crescimento das colônias, nas placas de isolamento primário, foi observado entre um
e sete dias a partir da semeadura do macerado bruto da esponja. A formação de colônias
mucóides e pigmentadas foi uma característica que prevaleceu dentre as estirpes isoladas, nos
três diferentes meios de cultivo. Foram observadas colônias nas cores branca, creme, amarela,
laranja, vermelha e marrom. Na Figura 9 pode-se observar alguns exemplos de colônias
isoladas nos meios BHI, Marine e Marine-mar.

46
(A) (B)
(C) Figura 9. Foto representativa da pigmentação observada nas estirpes marinhas G51 (vermelha) e Ab42 (amarela) após cultivo e isolamento no meio BHI (A); em sentido horário nas estirpes Pm41 (marrom), Ti41 e Pj 32 (brancas), Dr32 (amarela), Mm52 (branca), Pc34 (amarela) e Ca33 (branca) após cultivo e isolamento no meio Marine (B), e na estirpe Pc42 após cultivo e isolamento no meio Marine-mar (C). Estirpes marinhas isoladas de: Ab – A. brasiliensis; Ca – C. aurea; Dr – D. reticulatus; G – G. corticostylifera; Mm – M. microsigmatosa; Pc – P. citrina; Pj – P. janeirensis; Pm – P. magna; Ti – T. ignis.

47
Como critério de identificação, as estirpes foram nomeadas com as iniciais do nome da
espécie da esponja da qual foram isoladas, seguidas do número da coleta e da ordem de
isolamento da UFC a partir do meio de cultura. Exemplo: Mm33BHI – esta estirpe foi a
terceira colônia isolada da placa de ágar-BHI semeada com o macerado bruto da esponja
Mycalle microsigmatosa coletada na 3ª coleta.
4.2 Atividade antimicrobiana das estirpes associadas às esponjas
Entre as 149 estirpes marinhas que permaneceram viáveis, 16 (11%) apresentaram
atividade inibitória contra a estirpe indicadora C. fimi NCTC 7547 (Tabelas 3 e 4). Dentre
estas, encontram-se 9 isoladas em meio BHI (H40, H41, H51, Pc31, Pc32, Dr31, Mm33, Ti41
e Pm52). Três estirpes foram isoladas do meio Marine (Pm31, Ca31 e Mm37) e 4 do meio
Marine-mar (Mm33, H31, H42 e H43).
Dentre as 16 estirpes marinhas, 9 (56%) também apresentaram atividade inibitória
contra estirpes-padrão e clínicas, sendo que as isoladas em meio BHI (H40, H41, H51, Pc31,
Pc32 e Dr31) constituíram 67% (6/9) das estirpes classificadas como SAM-positivas
(produtoras de substâncias antimicrobianas). As três estirpes restantes foram isoladas do meio
Marine (Pm31, Ca31 e Mm37).
As estirpes marinhas H40, H41 e H51 apresentaram um maior espectro de ação, onde
se mostraram ativas contra cerca de 90% das estirpes indicadoras testadas (Tabela 3).
Outro destaque entre as estirpes SAM-positivas pode ser dado para a Pm31, Ca31 e
Mm37 (Tabela 4). Estas inibiram cerca de 60% das indicadoras testadas, sendo, em sua
maioria, estirpes Gram-positivas. O mesmo ocorreu para as estirpes Pc31 e Pc32 que foram
menos ativas contra estirpes Gram-negativas (Tabela 3).

48
A estirpe Dr31 foi a que apresentou um menor espectro de ação (13,4%), sendo ativa
somente contra as estirpes-padrão S. aureus, S. epidermidis e E. faecium (Tabela 3). Três
estirpes isoladas em meio BHI (Mm33, Ti41 e Pm52) e 4 estirpes isoladas em meio Marine-
mar (Mm33, H31, H42 e H43) inibiram a indicadora C. fimi, porém não apresentaram
atividade inibitória contra as estirpes-padrão S. aureus, E. faecium, E. coli e P. aeruginosa
(Tabelas 3 e 4). Sendo assim, estas estirpes não foram analisadas quanto à capacidade de
inibição das outras estirpes indicadoras.

49
Tabela 3. Testes de produção de SAM realizados com estirpes marinhas isoladas em meio
BHI contra estirpes de importância médica.
Os valores apresentados correspondem ao tamanho do halo de inibição do crescimento medido em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. Os valores apresentados na última linha correspondem ao percentual de inibição das estirpes marinhas em relação às estirpes indicadoras. (-): Resultado negativo; NT: não testado; Estirpes marinhas isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Ti – T. ignis; Pm – P. magna.
Estirpes Indicadoras Estirpes Marinhas (BHI) Estirpes-padrão H40 H41 H51 Pc31 Pc32 Dr31 Mm33 Ti 41 Pm 52 C. fimi NCTC 7547 23 26 43 46 42 20 18 18 15 S. aureus ATCC 29213 23 20 30 40 40 19 - - - S. epidermidis ATCC 12228 30 30 28 45 38 20 NT NT NT S. hominis ATCC 27844 31 28 37 41 43 - NT NT NT S. haemolyticus CCM 2737 27 27 35 40 40 - NT NT NT E. faecium ATCC 19434 18 21 30 23 20 20 - - - E. coli ATCC 25922 25 15 22 18 16 - - - - P. aeruginosa ATCC 27853 23 20 30 35 35 - - - - K. pneumoniae ATCC 700603 25 24 - - - - NT NT NT Acinetobacter sp. ATCC 14293 - - - 45 45 - NT NT NT E. cloacae ATCC 23355 22 25 18 - - - NT NT NT Estirpes clínicas S. aureus 42AE 23 27 17 - - - NT NT NT S. aureus 559a 20 22 43 40 40 - NT NT NT S. epidermidis 57s 25 25 33 30 30 - NT NT NT S. hominis 79s 25 27 20 35 30 - NT NT NT S. haemolyticus 109s 19 15 35 31 36 - NT NT NT E. faecalis V583 19 17 32 - - - NT NT NT E. faecalis 5AE - 20 - - - - NT NT NT E. coli 54AE 20 17 20 35 39 - NT NT NT P. aeruginosa 3AE 20 - 32 - - - NT NT NT E. cloacae 43AE 19 12 23 20 20 - NT NT NT E. hafniae 30AE 28 21 23 18 - - NT NT NT A. baumanii 76AE 20 20 - - - - NT NT NT A. calcoaceticus 56AE 19 18 30 35 30 - NT NT NT K. pneumoniae 52AE 18 15 21 - - - NT NT NT K. pneumoniae 19AE 16 20 32 - - - NT NT NT Salmonella 4AE 21 17 25 - - - NT NT NT S. marcescens 38AE 18 17 20 - - - NT NT NT S. maltophilia 17AE - - 16 - - - NT NT NT C. freundii 27AE 19 16 26 - - - NT NT NT Inibição (%) 90% 90% 86,7% 56,7% 53,4% 13,4% 20% 20% 20%

50
Tabela 4. Testes de produção de SAM realizados com estirpes marinhas isoladas nos meios Marine e Marine-mar.
Estirpes Indicadoras Estirpes Marinhas (M) Estirpes Marinhas (MM) Estirpes-padrão Pm31 Ca31 Mm37 Mm33 H31 H42 H43 C. fimi NCTC 7547 24 31 34 14 17 21 21 S. aureus ATCC 29213 27 28 20 - - - - S. epidermidis ATCC 12228 35 25 30 NT NT NT NT S. hominis ATCC 27844 25 23 24 NT NT NT NT S. haemolyticus CCM 2737 40 38 43 NT NT NT NT E. faecium ATCC 19434 20 22 15 - - - - E. coli ATCC 25922 - - 12 - - - - P. aeruginosa ATCC 27853 22 30 30 - - - - K. pneumoniae ATCC 700603 - - - NT NT NT NT Acinetobacter sp. ATCC 14293 - - - NT NT NT NT E. cloacae ATCC 23355 - - - NT NT NT NT Estirpes clínicas S. aureus 42AE - - - NT NT NT NT S. aureus 559a 25 17 20 NT NT NT NT S. epidermidis 57s 17 15 16 NT NT NT NT S. hominis 79s 25 25 26 NT NT NT NT S. haemolyticus 109s 30 20 20 NT NT NT NT E. faecalis V583 12 11 14 NT NT NT NT E. faecalis 5AE 16 12 15 NT NT NT NT E. coli 54AE 21 15 20 NT NT NT NT P. aeruginosa 3AE 12 14 15 NT NT NT NT E. cloacae 43AE - - - NT NT NT NT E. hafniae 30AE - - - NT NT NT NT A. baumanii 76AE - - - NT NT NT NT A. calcoaceticus 56AE 23 18 23 NT NT NT NT K. pneumoniae 52AE - - - NT NT NT NT K. pneumoniae 19AE - - - NT NT NT NT Salmonella 4AE 12 14 14 NT NT NT NT S. marcescens 38AE - - - NT NT NT NT S. maltophilia 17AE 13 13 15 NT NT NT NT C. freundii 27AE 10 - - NT NT NT NT Inibição (%) 63,4% 60% 60% 20% 20% 20% 20% Os valores apresentados correspondem ao tamanho do halo de inibição do crescimento medido em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. Os valores apresentados na última linha correspondem ao percentual de inibição das estirpes marinhas em relação às estirpes indicadoras. (-): Resultado negativo; NT: não testado; Estirpes marinhas isoladas de: Pm – P. magna; Ca – C. aurea; Mm – M. microsigmatosa; H – Haliclona spn.; M: meio Marine; MM: meio Marine-mar.

51
Com o objetivo de verificar a capacidade de inibição entre as estirpes marinhas SAM-
positivas, foi realizado o experimento de inibição cruzada (Tabela 5). Neste experimento,
nenhuma das estirpes marinhas testadas apresentou auto-inibição. As estirpes H40, H41 e
H51, isoladas da mesma esponja e coletadas em épocas diferentes (01/2007 e 03/2007), não
apresentaram inibição umas contra as outras (Tabela 5). Quando utilizadas como estirpes
produtoras apresentaram perfis de inibição distintos. As estirpes H40 e H41 inibiram o
crescimento de 7 e de 10 das 13 estirpes indicadoras testadas, respectivamente. Já a estirpe
H51 só inibiu o crescimento da estirpe H43, isolada em meio Marine-mar. Contrariamente,
quando utilizadas como indicadoras, estas estirpes não tiveram o seu crescimento inibido por
nenhuma das estirpes marinhas produtoras.
As estirpes Pc31 e Pc32 isoladas da mesma esponja coletada na mesma época
apresentaram perfis de inibição bastante semelhantes (Tabela 5), onde 4 das 13 estirpes
indicadoras testadas tiveram o seu crescimento inibido por estas estirpes. A estirpe Dr31 foi a
indicadora que teve seu crescimento inibido pela maioria das estirpes produtoras (56%). Além
disso, como estirpe produtora, só inibiu o crescimento das estirpes Pc32 e Ti41.
As estirpes Mm33, Pm52 e Ti41 não inibiram nenhuma das indicadoras marinhas
testadas (Tabela 5). Em relação a estirpe Ti41, esta não cresceu como indicadora sobre as
estirpes isoladas em meio Marine-mar.
As estirpes marinhas Pm31, Ca31 e Mm37, isoladas de esponjas de espécies diferentes,
coletadas na mesma época (04/2006), também não mostraram inibição umas contra as outras
(Tabela 5). Além disso, estas estirpes foram as que apresentaram um espectro de ação mais
amplo, pois inibiram 10 das 16 estirpes indicadoras testadas. Por outro lado, como

52
indicadoras, não cresceram sobre as estirpes isoladas em meio BHI e só tiveram seu
crescimento inibido pelas estirpes H42 e H43, isoladas em meio Marine-mar.
Quanto às estirpes Mm33, H31, H42 e H43 isoladas em meio Marine-mar, estas não
mostraram inibição umas contra as outras e se mostraram bastante ativas contra estirpes
marinhas, onde juntas inibiram 7 das 15 estirpes indicadoras testadas.

53
Tabela 5. Testes de inibição cruzada realizados com as estirpes marinhas SAM-positivas.
Os valores apresentados correspondem ao tamanho do halo de inibição do crescimento medido em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. Os valores apresentados na última linha correspondem ao percentual de inibição das estirpes marinhas utilizadas como indicadoras. (-): Resultado negativo; ND: não determinado; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ti – T. ignis; Ca – C. aurea; M: meio Marine; MM: meio Marine-mar.
Estirpes Indicadoras BHI M MM
Mei
os
Estirpes Produtoras
H40 H41 H51 Pc31 Pc32 Dr31 Mm33 Pm52 Ti41 Pm31 Ca31 Mm37 Mm33 H31 H42 H43 H40 - - - 18 - - 17 25 24 ND ND ND 17 - 14 20 H41 - - - 19 17 21 19 25 26 ND ND ND 20 17 16 18 H51 - - - - - - - - - ND ND ND - - - 45 Pc31 - - - - - 13 - - 23 ND ND ND - - 13 14 Pc32 - - - - - 12 - - 23 ND ND ND - - 13 13 Dr31 - - - - 10 - - - 16 ND ND ND - - - -
Mm33 - - - - - - - - - ND ND ND - - - - Pm52 - - - - - - - - - ND ND ND - - - -
BH
I
Ti41 - - - - - - - - - ND ND ND - - - - Pm31 - - - 17 19 17 13 20 21 - - - 17 15 13 13 Ca31 - - - 20 19 17 12 20 24 - - - 18 18 15 14 M
Mm37 - - - 18 19 18 14 20 21 - - - 14 19 15 14 Mm33 - - - - 13 19 32 - ND - - - - - - - H31 - - - - 15 12 - - ND - - - - - - - H42 - - - 14 - 16 - - ND 30 30 25 - - - - H43 - - - - - - - - ND 33 31 25 - - - - M
M
Inibição (%) 0% 0% 0% 37% 44% 56% 37% 31% 50% 12% 12% 12% 31% 25% 44% 50%

54
Foi observado que o tamanho dos halos de inibição de crescimento variaram entre 12 e
46mm, como exemplificado na Figura 10. Não foi observada discrepância entre os tamanhos
dos halos quando foi feita a duplicata do experimento.
(A) (A) (B) (C)
(D) (E) (F) Figura 10. Ensaios de produção de SAM com as estirpes marinhas Pm31 contra S. haemolyticus CCM 2737 (A), H51 contra E. faecalis V583 (B), Mm37 contra E. coli 54AE (C), Pc31 (direita) e Pc32 (esquerda) contra A. calcoaceticus (D), Pc31 (direita) e Pc32 (esquerda) contra a estirpe marinha Ca31 (E) e H40 contra a estirpe marinha H31 (F). A estirpe Pm31 foi isolada a partir da esponja P. magna em meio Marine, as estirpes H51 e H40 foram isoladas a partir da esponja Haliclona spn. em meio BHI, a estirpe Mm37 foi isolada a partir da esponja M. microsigmatosa em meio Marine, as estirpes Pc31 e Pc32 foram isoladas a partir da esponja P. citrina em meio BHI, a estirpe Ca31 foi isolada da esponja C. aurea em meio Marine e a estirpe H31 foi isolada a partir da esponja Haliclona spn. em meio Marine-mar.
37mm 30mm 25mm
30mm 35mm 17mm 19mm
20mm

55
De acordo com os resultados obtidos nos testes de produção de SAM, as estirpes
marinhas Mm33, Ti41 e Pm52 isoladas em meio BHI e as estirpes marinhas Mm33, H31,
H42, H43 isoladas em meio Marine-mar apresentaram atividade somente contra a estirpe
indicadora C. fimi. Como o objetivo deste trabalho era caracterizar estirpes marinhas que
inibissem o crescimento de estirpes de importância médica, somente as estirpes H40, H41,
H51, Pc31, Pc32 e Dr31 isoladas em meio BHI e as estirpes Pm31, Ca31 e Mm37 isoladas em
meio Marine foram selecionadas para as etapas seguintes (itens 3.6 ao 3.9 do Materiais e
Métodos).
4.3 Caracterização fenotípica das estirpes marinhas SAM-positivas
4.3.1 Caracterização das estirpes marinhas SAM-positivas
As 9 estirpes SAM-positivas foram, então, caracterizadas fenotipicamente conforme
descrito no item 3.6 do Materiais e Métodos. Os resultados dos testes realizados a partir do
crescimento em temperatura ambiente estão apresentados na Tabela 6.
As estirpes SAM-positivas apresentaram morfologia colonial marrom e mucóide, bege
e mucóide com produção de pigmento, ou creme com aspecto rugoso.
O crescimento das estirpes SAM-positivas foi verificado à temperatura ambiente,
sendo que as estirpes isoladas em meio BHI formaram uma massa celular visível em 24h e as
estirpes isoladas em meio Marine apresentaram uma massa equivalente em 48h de
crescimento.
Morfotintorialmente, 6 estirpes foram caracterizadas como bacilos Gram-negativos e 3
apresentaram-se como bacilos Gram-positivos, com formação de esporos (Figura 11).

56
Além disso, todas as estirpes SAM-positivas foram positivas para o teste da catalase e
da oxidase e foram caracterizadas como móveis (Tabela 6).
(A) (B) Figura 11. Coloração de Gram das estirpes Pc31, isolada da esponja P. citrina em meio BHI, caracterizada como bacilo Gram-positivo (A) e Pm31, isolada da esponja P. magna em meio Marine, caracterizada como bacilo Gram-negativo (B).

57
Tabela 6. Características fenotípicas das estirpes marinhas SAM-positivas crescidas em temperatura ambiente.
(+): Resultado positivo; (-): Resultado negativo; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ca – C. aurea; M: meio Marine.
Mei
os
Amostra Características Morfológicas Produção
de Pigmento
Coloração de Gram Presença
de Esporos Catalase Oxidase Motilidade
Tempo de
Crescimento
H 40 Colônia bege e mucóide + Bacilo Gram-negativo - + + + 24h H 41 Colônia bege e mucóide + Bacilo Gram-negativo - + + + 24h H 51 Colônia bege e mucóide + Bacilo Gram-negativo - + + + 24h Pc 31 Colônia creme e rugosa - Bacilo Gram-positivo + + + + 24h Pc 32 Colônia creme e rugosa - Bacilo Gram-positivo + + + + 24h
BH
I
Dr 31 Colônia creme e rugosa - Bacilo Gram-positivo + + + + 24h Pm 31 Colônia marrom e mucóide - Bacilo Gram-negativo - + + + 48h Ca 31 Colônia marrom e mucóide - Bacilo Gram-negativo - + + + 48h M
Mm 37 Colônia marrom e mucóide - Bacilo Gram-negativo - + + + 48h

58
4.3.2 Análise das condições de temperatura e salinidade para o crescimento das estirpes
marinhas SAM-positivas
As estirpes marinhas SAM-positivas foram testadas quanto à capacidade de
crescimento a 15°C, 37ºC e 42°C. De acordo com os resultados obtidos observou-se que a
15°C as estirpes H51, Pc31, Pc32 e Dr31 isoladas em meio BHI e as estirpes Pm31, Ca31 e
Mm37 isoladas em meio Marine apresentaram um tempo de crescimento maior do que aquele
observado à temperatura ambiente. Somente as estirpes H40 e H41 apresentaram um tempo de
crescimento equivalente ao observado à temperatura ambiente (Tabela 7).
Na temperatura de 37°C (Tabela 7), as estirpes isoladas em meio BHI apresentaram um
tempo de crescimento equivalente ao observado à temperatura ambiente. Quanto às estirpes
isoladas em meio Marine, somente a Mm37 apresentou um tempo de crescimento equivalente
ao observado à temperatura ambiente. As estirpes Ca31 e Pm31 não cresceram nesta
temperatura.
Em relação ao teste de crescimento a 42°C (Tabela 7), as estirpes H51, Pc31, Pc32 e
Dr31 isoladas em meio BHI apresentaram um tempo de crescimento equivalente ao observado
à temperatura ambiente. Em contrapartida, as estirpes H40 e H41 isoladas em meio BHI e as
estirpes Pm31, Ca31 e Mm37 isoladas em meio Marine não foram capazes de crescer nesta
temperatura.

59
Tabela 7. Análise da capacidade de crescimento das estirpes marinhas SAM-positivas nas
temperaturas de 15°C, 37°C e 42°C.
Estes dados correspondem aos resultados de dois experimentos e discrepâncias entre os resultados não foram observadas. (+): Presença de crescimento; (-): Ausência de crescimento; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ca – C. aurea; M: meio Marine.
Mei
os
Estirpes Controle T.A.
Tempo cresc. 15°C Tempo
cresc. 37°C Tempo cresc. 42°C Tempo
cresc.
H40 + 24h + 24h + 24h - - H41 + 24h + 24h + 24h - - H51 + 24h + 48h + 24h + 24h Pc31 + 24h + 48h + 24h + 24h Pc32 + 24h + 48h + 24h + 24h
BH
I
Dr31 + 24h + 48h + 24h + 24h Ca31 + 48h + 72h - - - - Pm31 + 48h + 72h - - - - M
Mm37 + 48h + 72h + 48h - -

60
As estirpes marinhas SAM-positivas isoladas em meio BHI foram testadas quanto à
capacidade de crescimento em concentrações de 1, 1,5 e 2% de NaCl. Foi observado que as
estirpes H40, H41 e Dr31 apresentaram um tempo de crescimento equivalente ao observado
em uma concentração de 0,5% de NaCl (condição de salinidade do meio BHI). Enquanto
que, as estirpes H51, Pc31 e Pc32 apresentaram um tempo de crescimento maior do que
aquele observado em uma concentração de 0,5% de NaCl (Tabela 8).
Tabela 8. Análise da capacidade de crescimento das estirpes marinhas SAM-positivas
isoladas em meio BHI em concentrações de 1, 1,5 e 2% de NaCl.
Estes dados correspondem aos resultados de dois experimentos e discrepâncias entre os resultados não foram observadas. (+): Presença de crescimento; (-): Ausência de crescimento; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus.
Mei
os
Estirpes Controle
0,5% NaCl
Tempo cresc.
1% NaCl
Tempo cresc.
1,5% NaCl
Tempo cresc.
2% NaCl
Tempo cresc.
H40 + 24h + 24h + 24h + 24h H41 + 24h + 24h + 24h + 24h H51 + 24h + 48h + 48h + 48h Pc31 + 24h + 48h + 48h + 48h Pc32 + 24h + 48h + 48h + 48h
BH
I
Dr31 + 24h + 24h + 24h + 24h

61
4.3.3 Identificação das estirpes marinhas SAM-positivas através do sistema comercial
miniaturizado
Através do API 50CH, as estirpes Pc31, Pc32 e Dr31 foram identificadas como
Bacillus pumilus com 99,9% de identidade. A partir do API 20NE, as estirpes H40 e H41
foram identificadas como Pseudomonas fluorescens com 98% de identidade e a estirpe H51
foi identificada como Pseudomonas putida com 99,6% de identidade. As estirpes Pm31,
Ca31 e Mm37 isoladas em meio Marine não foram identificadas por este método.
4.3.4 Susceptibilidade aos antimicrobianos
Os resultados dos testes de susceptibilidade aos antimicrobianos estão apresentados
na Tabela 9. De acordo com a ausência dos halos de inibição de crescimento, as estirpes
H40, H41 e H51 isoladas em meio BHI foram consideradas resistentes aos antibióticos
cefalexina e sulfazotrim. Além destes antibióticos, a estirpe H51 foi considerada resistente
ao cloranfenicol (Figura 12) e as estirpes H40 e H41, resistentes ao aztreonam (Tabela 9).
Figura 12. Foto do antibiograma da estirpe H51 (Pseudomonas putida). Antibióticos testados: ATM – aztreonam; CAZ – ceftazidima; CFE – cefalexina; CIP – ciprofloxacina; CLO – cloranfenicol; GEN – gentamicina; SUT – sulfazotrim; TET – tetraciclina. H – Haliclona spn.
CFE CLO
SUT
GEN
CIP
TET
CAZ
ATM

62
As estirpes Pm31, Ca31 e Mm37 isoladas em meio Marine foram resistentes aos
antibióticos gentamicina e ciprofloxacina. Além destes antibióticos, as estirpes Pm31 e
Mm37 foram resistentes ao sulfazotrim, sendo esta última também resistente à ceftazidima
(Tabela 9).
As estirpes Pc31, Pc32 e Dr31 isoladas em meio BHI tiveram o seu crescimento
inibido em todos os testes (o menor halo de inibição de crescimento foi de 22mm de
diâmetro) (Tabela 9).

63
Tabela 9. Testes de susceptibilidade aos antimicrobianos das estirpes marinhas SAM-positivas.
Os valores apresentados correspondem aos tamanhos dos halos de inibição do crescimento medidos em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. (-): Não houve inibição de crescimento; NT: antibiótico não testado; Antibióticos testados: ATM – aztreonam; CAZ – ceftazidima; CFE – cefalexina; CIP – ciprofloxacina; CLO – cloranfenicol; ERI – eritromicina; GEN – gentamicina; PEN – penicilina G; SUT – sulfazotrim; TET – tetraciclina. Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ca – C. aurea; M: meio Marine.
Meios Estirpes CFE TET CLO SUT GEN CIP ATM CAZ ERI PEN H 40 - 27 9 - 18 35 - 18 NT NT H 41 - 28 10 - 20 37 - 20 NT NT H 51 - 20 - - 17 30 23 22 NT NT Pc31 35 40 22 45 26 31 NT NT 35 42 Pc32 37 40 24 45 25 31 NT NT 36 40
BH
I Dr31 38 40 25 45 28 35 NT NT 39 45 Pm 31 44 23 44 - - - 25 16 NT NT Ca 31 45 28 54 25 - - 23 14 NT NT M
Mm 37 28 19 44 - - - 16 - NT NT

64
4.4 Identificação molecular das estirpes marinhas SAM-positivas
Através do seqüenciamento parcial do gene que codifica para o 16S rRNA e da
comparação, pela ferramenta BLAST, das seqüências obtidas com seqüências homólogas
depositadas no banco de dados GenBank, todas as estirpes marinhas SAM-positivas foram
identificadas em gênero ou espécie.
As estirpes H40 e H41 isoladas em meio BHI tiveram suas seqüências alinhadas
com seqüências obtidas a partir do gene que codifica para o 16S rRNA do gênero
Pseudomonas. Dentre estas seqüências, destacaram-se os alinhamentos das seqüências das
estirpes H40 e H41 com as seqüências das estirpes depositadas como P. fluorescens e P.
gessardii com 96% de homologia.
A estirpe H51 isolada em meio BHI teve sua seqüência alinhada com seqüências
obtidas a partir do gene que codifica para o 16S rRNA do gênero Pseudomonas com 96%
de homologia entre elas.
As estirpes Ca31 e Pm31 isoladas em meio Marine tiveram suas seqüências
alinhadas com seqüências obtidas a partir do gene que codifica para o 16S rRNA do gênero
Pseudovibrio. Dentre estas seqüências, destacaram-se os alinhamentos das seqüências das
estirpes Ca31 e Pm31 com as seqüências das estirpes depositadas como P. ascidiaceicola e
P. japonicus com 96% de homologia.
A estirpe Mm37 isolada em meio Marine teve sua seqüência alinhada com
seqüências obtidas a partir do gene que codifica para o 16S rRNA do gênero Pseudovibrio.
Dentre estas seqüências, destacou-se o alinhamento da seqüência da estirpe Mm37 com a
seqüência da estirpe depositada como P. denitrificans com 96% de homologia.

65
A estirpe Pc31 isolada em meio BHI teve sua seqüência alinhada com seqüências
obtidas a partir do gene que codifica para o 16S rRNA do gênero Bacillus. Dentre estas
seqüências, destacou-se o alinhamento da seqüência da estirpe Pc31 com as seqüências das
estirpes depositadas como B. pumilus e B. subtilis com 99% de homologia.
As estirpes Pc32 e Dr31 isoladas em meio BHI tiveram suas seqüências alinhadas
com seqüências obtidas a partir do gene que codifica para o 16S rRNA do gênero Bacillus
sp. Dentre estas seqüências, destacaram-se os alinhamentos das seqüências das estirpes
Pc32 e Dr31 com as seqüências das estirpes depositadas como B. pumilus com homologias
de 99% e 96%, respectivamente.
Tabela 10. Resultados da identificação das estirpes marinhas SAM-positivas através do
sistema bioquímico API e do seqüenciamento parcial do gene que codifica para o 16S
rRNA.
Meio Estirpes marinhas SAM-positivas
Identificação Molecular Identificação Bioquímica
H40 Pseudomonas fluorescens ou P. gessardii (97%) P. fluorescens (98%) H41 P. fluorescens ou P. gessardii (97%) P. fluorescens (98%) H51 Pseudomonas sp. (96%) P. putida (99,6%) Pc31 Bacillus pumilus ou B. subtilis (99%) B. pumilus (99,9%) Pc32 B. pumilus (99%) B. pumilus (99,9%)
BHI
Dr31 B. pumilus (96%) B. pumilus (99,9%) Pm31 Pseudovibrio ascidiaceicola ou P. japonicus (96%) - Ca31 P. ascidiaceicola ou P. japonicus (96%) - M
Mm37 P. denitrificans (96%) -

66
4.5 Caracterização da(s) substância(s) antimicrobiana(s) quanto à
natureza química
De acordo com os valores observados na Tabela 11, as SAM produzidas pelas
estirpes H41 (P. fluorescens), H51 (P. putida), Pc31 (Figura 13) e Pc32 (B. pumilus),
isoladas do meio BHI, e Pm31 (Pseudovibrio sp.), isolada do meio Marine, foram
resistentes à ação de todas as enzimas proteolíticas testadas, assim como, resistentes à ação
da fosfolipase C e do NaOH.
Figura 13. Caracterização da SAM produzida pela estirpe Pc31 quanto à natureza química. Cada um dos 4 pontos representa a estirpe Pc31 crescida à temperatura ambiente por 48h. A estirpe C. fimi foi utilizada como indicadora neste experimento.
A SAM produzida pela estirpe H40 (P. fluorescens) isolada do meio BHI se
mostrou sensível (redução de 50% do halo de inibição) à ação da fosfolipase C (Tabela 11).
Em relação a SAM produzida pela estirpe Dr31 (B. pumilus) isolada do meio BHI,
pode-se observar que a SAM foi sensível à ação de enzimas proteolíticas (pronase e
Controle
Protease
Pronase Proteinase K
Controle
Tripsina
Fosfolipase C
NaOH

67
proteinase K). Além dessa característica, a SAM também se mostrou sensível à ação do
NAOH (Tabela 11).
A SAM produzida pela estirpe Ca31 (Pseudovibrio sp.) isolada do meio Marine foi
sensível à ação de apenas uma enzima proteolítica (pronase) (Figura 14) com redução de
50% do halo de inibição quando comparado ao controle. Além disso, esta SAM também foi
sensível ao NaOH (Tabela 11).
E finalmente, a SAM produzida pela estirpe Mm37 (Pseudovibrio sp.) isolada do
meio Marine apresentou sensibilidade à fosfolipase C e ao NaOH (Tabela 11).
Figura 14. Sensibilidade da SAM produzida pela estirpe Ca31 à enzima proteolítica pronase. Cada um dos 4 pontos representa a estirpe Ca31 crescida à temperatura ambiente por 48h. A estirpe C. fimi foi utilizada como indicadora neste experimento.
Controle
Pronase
Protease
Proteinase K

68
Tabela 11. Testes realizados para a caracterização da natureza química das substâncias antimicrobianas produzidas pelas estirpes
marinhas.
Meios Estirpes Controle Protease Pronase ProteinaseK Tripsina FosfolipaseC NaOH H40 24 19 21 21 20 12 23 H41 25 25 24 25 24 25 25 H51 45 45 45 45 44 45 45 Pc31 44 43 44 44 44 43 44 Pc32 40 40 40 39 40 39 40
BH
I
Dr31 20 15 - - 20 20 - Ca31 33 28 10 27 33 32 - Pm31 24 20 21 20 20 21 20 M
Mm37 35 30 35 35 31 11 - Os valores apresentados correspondem aos tamanhos dos halos de inibição do crescimento medidos em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. A coluna controle representa as estirpes marinhas não tratadas com as enzimas e com NaOH. A estirpe C. fimi foi utilizada como indicadora neste teste. (-) ausência de halo. Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ca – C. aurea; M: meio Marine.

69
4.6 Análise das condições de temperatura e salinidade para a produção
da(s) substância(s) antimicrobianas(s).
As estirpes marinhas SAM-positivas foram analisadas quanto à capacidade de
produzir as substâncias antimicrobianas a 15°C, 37ºC e 42°C. Portanto, foram analisadas
apenas as estirpes capazes de crescer em cada uma destas temperaturas, conforme
resultados apresentados no item 4.3.2 e na Tabela 7.
De acordo com os resultados apresentados na Tabela 12 observou-se que a 15°C as
estirpes H40 e H41 (P. fluorescens) isoladas do meio BHI produziram as SAM e
mantiveram o mesmo halo de inibição do crescimento da estirpe indicadora C. fimi, quando
comparado com aquele observado à temperatura ambiente. A estirpe H51 (P. putida)
isolada do meio BHI e as estirpes Pm31, Ca31, Mm37 (Pseudovibrio sp.) isoladas do meio
Marine produziram as SAM a 15°C, porém apresentaram uma redução no halo de inibição
do crescimento da estirpe indicadora. Já as estirpes Pc31, Pc32 e Dr31 (B. pumilus) isoladas
do meio BHI não produziram as SAM nesta temperatura.
Na temperatura de 37°C (Tabela 12), as estirpes H51 (P. putida) e Dr31 (B.
pumilus) isoladas do meio BHI e a estirpe Mm37 (Pseudovibrio sp.) isolada do meio
Marine produziram as SAM e mantiveram o mesmo halo de inibição do crescimento da
estirpe indicadora C. fimi, quando comparado com aquele observado à temperatura
ambiente. As estirpes Pc31 e Pc32 (B. pumilus) isoladas do meio BHI produziram as SAM
a 37°C, porém apresentaram uma redução no halo de inibição do crescimento da estirpe
indicadora. As estirpes H40 e H41 (P. fluorescens) isoladas do meio BHI não produziram
as SAM, enquanto que as estirpes Pm31 e Ca31 (Pseudovibrio sp.) isoladas do meio
Marine não cresceram nesta temperatura.
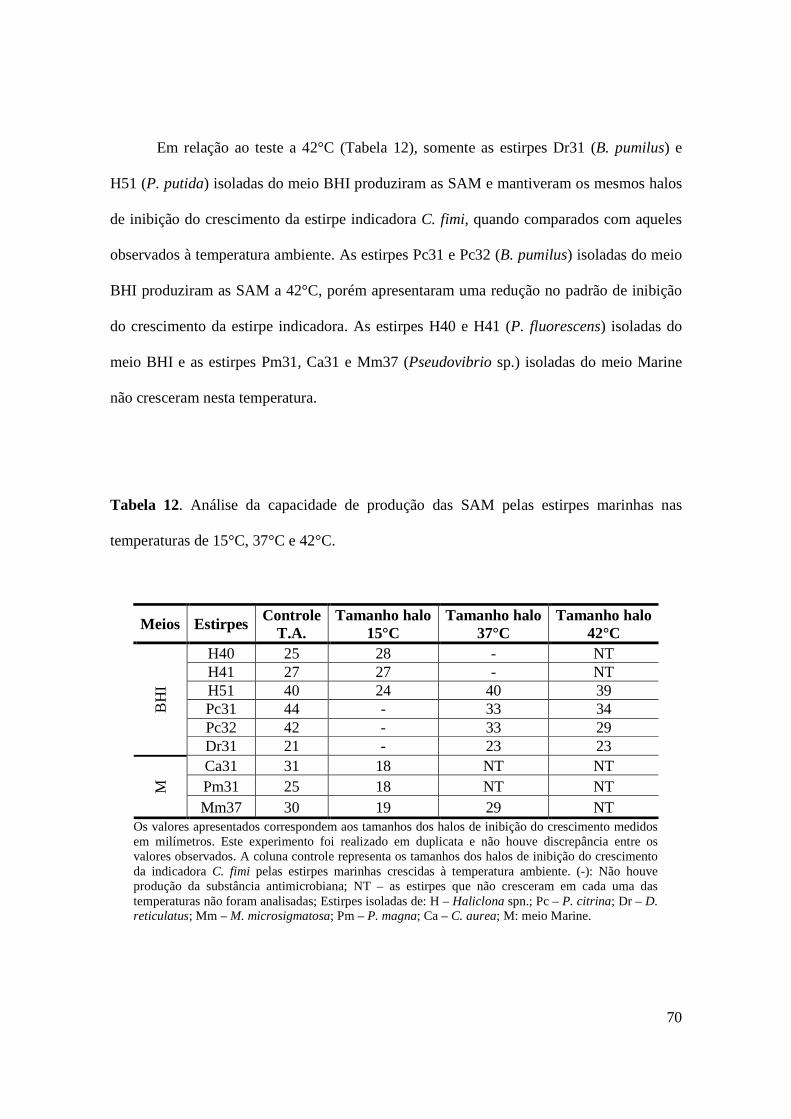
70
Em relação ao teste a 42°C (Tabela 12), somente as estirpes Dr31 (B. pumilus) e
H51 (P. putida) isoladas do meio BHI produziram as SAM e mantiveram os mesmos halos
de inibição do crescimento da estirpe indicadora C. fimi, quando comparados com aqueles
observados à temperatura ambiente. As estirpes Pc31 e Pc32 (B. pumilus) isoladas do meio
BHI produziram as SAM a 42°C, porém apresentaram uma redução no padrão de inibição
do crescimento da estirpe indicadora. As estirpes H40 e H41 (P. fluorescens) isoladas do
meio BHI e as estirpes Pm31, Ca31 e Mm37 (Pseudovibrio sp.) isoladas do meio Marine
não cresceram nesta temperatura.
Tabela 12. Análise da capacidade de produção das SAM pelas estirpes marinhas nas
temperaturas de 15°C, 37°C e 42°C.
Meios Estirpes Controle T.A.
Tamanho halo 15°C
Tamanho halo 37°C
Tamanho halo 42°C
H40 25 28 - NT H41 27 27 - NT H51 40 24 40 39 Pc31 44 - 33 34 Pc32 42 - 33 29
BH
I
Dr31 21 - 23 23 Ca31 31 18 NT NT Pm31 25 18 NT NT M
Mm37 30 19 29 NT Os valores apresentados correspondem aos tamanhos dos halos de inibição do crescimento medidos em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. A coluna controle representa os tamanhos dos halos de inibição do crescimento da indicadora C. fimi pelas estirpes marinhas crescidas à temperatura ambiente. (-): Não houve produção da substância antimicrobiana; NT – as estirpes que não cresceram em cada uma das temperaturas não foram analisadas; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus; Mm – M. microsigmatosa; Pm – P. magna; Ca – C. aurea; M: meio Marine.

71
As estirpes marinhas SAM-positivas isoladas em meio BHI foram testadas quanto à
capacidade de produzir a substância antimicrobiana em concentrações de 1, 1,5 e 2% de
NaCl.
De acordo com os resultados observados na Tabela 13, nas três concentrações de
NaCl analisadas, somente as estirpes H41 (P. fluorescens), H51 (P. putida) e Pc31 (B.
pumilus) produziram as SAM. Sendo que, a estirpe Pc31 produziu a SAM e manteve o
mesmo halo de inibição do crescimento da estirpe indicadora C. fimi, quando comparado
com aquele observado em uma concentração de 0,5% de NaCl (Figura 15). Embora as
estirpes H41 e H51 tenham produzido as SAM, estas apresentaram uma redução no halo de
inibição do crescimento da estirpe indicadora, quando comparado com aquele observado
em uma concentração de 0,5% de NaCl
(A) (B) (C) Figura 15. Produção da SAM pela estirpe Pc31 nas concentrações de 1% (A), 1,5% (B) e 2% (C) de NaCl contra a estirpe indicadora C. fimi.

72
Tabela 13. Análise da capacidade de produção das SAM pelas estirpes marinhas isoladas
em meio BHI em concentrações de 1, 1,5 e 2% de NaCl.
Estirpes Tamanho halo Controle
Tamanho halo NaCl 1%
Tamanho halo NaCl 1,5%
Tamanho halo NaCl 2%
H40 25 - - - H41 27 18 16 17 H51 40 18 20 19 Pc31 44 41 45 40 Pc32 42 - - - Dr31 21 - - -
Os valores apresentados correspondem aos tamanhos dos halos de inibição do crescimento medidos em milímetros. Este experimento foi realizado em duplicata e não houve discrepância entre os valores observados. A coluna controle representa a produção de SAM pelas estirpes marinhas crescidas em meio BHI sem adição de NaCl, isto é, cresceram nas condições de salinidade do próprio meio. (-): Não houve produção da substância antimicrobiana; Estirpes isoladas de: H – Haliclona spn.; Pc – P. citrina; Dr – D. reticulatus.
4.7 Identificação e caracterização de bactérias produtoras de substâncias
antimicrobianas isoladas de esponjas marinhas
Os resultados do presente estudo estão descritos em resumo na Tabela 14.

73
Tabela 14. Resultados conclusivos obtidos pelas estirpes marinhas SAM-positivas
analisadas no presente estudo.
O meio BHI possui em sua composição 0,5% de NaCl; T.A. – temperatura ambiente (em torno de 25-30°C).
Meios Estirpes Marinhas Identificação
Condições ótimas para produção da SAM
(Temperatura e Salinidade) H40 Pseudomonas fluorescens 15°C-T.A., 0,5% de NaCl H41 Pseudomonas fluorescens 15°C-T.A., 0,5% de NaCl H51 Pseudomonas putida T.A.-42°C, 0,5% de NaCl Pc31 Bacillus pumilus T.A., 0,5-2% de NaCl Pc32 Bacillus pumilus T.A., 0,5% de NaCl
BH
I
Dr31 Bacillus pumilus T.A.-42°C, 0,5% de NaCl Pm31 Pseudovibrio sp. T.A., 2% de NaCl Ca31 Pseudovibrio sp. T.A., 2% de NaCl M
Mm37 Pseudovibrio sp. T.A.-37°C, 2% de NaCl

74
5 DISCUSSÃO
As doenças infecciosas causadas por bactérias, fungos, vírus e parasitas ainda
constituem o principal problema de saúde pública. Seu impacto é particularmente maior em
países em desenvolvimento, devido à precariedade dos sistemas de saúde e emergência de
microrganismos multirresistentes (OKEKE et al., 2005).
Os produtos naturais continuam sendo a principal fonte de novos agentes
terapêuticos para várias doenças, incluindo as infecciosas. Somente uma pequena porção da
diversidade disponível de fungos, da fauna e flora marinhas, de bactérias e de plantas têm
sido explorada, e sugere-se que recursos ilimitados podem ser obtidos através do estudo
destes organismos (CLARDY & WALSH, 2004).
Neste contexto, o presente trabalho teve por objetivos caracterizar e identificar
bactérias produtoras de substâncias antimicrobianas associadas a esponjas marinhas
coletadas na costa da cidade do Rio de Janeiro. O primeiro passo para atingir tais objetivos
foi isolar bactérias associadas às esponjas A. brasiliensis, C. aurea, D. reticulatus, G.
corticostylifera, Haliclona spn., M. microsigmatosa, P. magna, P. citrina, P. janeirensis e
T. ignis a partir de três diferentes meios de cultura. No decorrer das coletas, as
metodologias de isolamento foram sendo otimizadas a fim de se obter um maior número de
espécies bacterianas isoladas. Desta forma, podemos citar o emprego do meio Cetrimide na
5ª coleta, visando o isolamento de espécies do gênero Pseudomonas, espécies estas
comprovadamente produtoras de substâncias antimicrobianas (JAYATILAKE et al., 1996;
ZHENG et al., 2000; SINGH et al., 2003).

75
Neste trabalho, um total de 168 UFC foram isoladas a partir 10 espécies de esponjas
cultivadas nos meios BHI, Marine e Marine-mar. Anand e colaboradores (2006) isolaram
75 UFC a partir de quatro espécies de esponjas marinhas provenientes da Índia, utilizando
apenas um meio de cultura. Em contrapartida, Graeber e colaboradores (2003) isolaram 378
UFC a partir de 12 espécies de esponjas marinhas provenientes da Noruega, utilizando
quatro diferentes meios de cultura. Por sua vez, Dey e colaboradores (2003) isolaram 133
UFC a partir de cinco espécies de esponjas marinhas provenientes da Austrália, utilizando
cinco diferentes meios de cultura. A diferença observada entre as taxas de isolamento
bacteriano foi, em grande parte, atribuída aos diferentes meios de cultura utilizados.
Segundo Muscholl-Silberhorn, Thiel e Imhoff (2008) o crescimento e a diversidade das
colônias bacterianas são amplamente influenciados pelo meio de cultivo. No presente
trabalho, não houve muita diferença entre o número de UFC isoladas quanto aos três
principais meios (BHI, Marine e Marine-mar) de isolamento utilizados. Nos outros
trabalhos, embora tenham sido utilizados diferentes meios de cultivo para aumentar as
chances de isolamento de UFC associadas a esponjas, o meio Marine foi o que
proporcionou maior número de morfotipos diferentes, provavelmente porque mimetiza as
condições do ambiente marinho. Curiosamente, a maioria das colônias apresentou
pigmentação amarela, laranja, vermelha ou marrom, independente do meio de cultura
utilizado, como observado também em nosso trabalho.
O segundo passo foi detectar quais estirpes marinhas eram produtoras de
substâncias antimicrobianas. Segundo Cos e colaboradores (2006), a escolha e a validação
das técnicas de triagem são essenciais para garantir uma seleção confiável de substâncias
com ações farmacológicas relevantes. Neste contexto, a estirpe C. fimi foi selecionada
como indicadora para os testes de rastreamento inicial por dois motivos: (1) as bactérias

76
Gram-positivas são muito mais sensíveis à ação das substâncias antimicrobianas do que as
bactérias Gram-negativas (CLARDY & WALSH, 2004); (2) a estirpe C. fimi é conhecida
por ser altamente sensível às bacteriocinas estafilocócicas (GIAMBIAGI-DEMARVAL et
al., 1990). Assim, das 149 estirpes marinhas ou UFCs que permaneceram viáveis, 16 (11%)
apresentaram atividade inibitória contra a estirpe indicadora C. fimi.
Ainda segundo Cos e colaboradores (2006), a escolha dos microrganismos a serem
testados depende do objetivo da investigação. Em uma triagem inicial, estirpes-padrão
devem ser utilizadas e devem representar as espécies patogênicas mais comuns. Várias
combinações são possíveis, mas devem conter pelo menos uma estirpe Gram-positiva e
uma Gram-negativa. Neste trabalho, foram utilizadas as estirpes-padrão S. aureus e E.
faecium (Gram-positivas) e E. coli e P. aeruginosa (Gram-negativas). As estirpes que
apresentarem atividade contra as estirpes-padrão utilizadas na triagem inicial devem ser
testadas contra outras estirpes-padrão de relevância médica e estirpes clínicas
multirresistentes. Desta forma, nove das 16 estirpes marinhas foram capazes de inibir o
crescimento de estirpes de importância médica utilizadas como indicadoras no presente
estudo.
A porcentagem (11%) de estirpes produtoras de substâncias antimicrobianas
observada no presente trabalho corresponde àquela descrita por Hentschel e colaboradores
(2001), onde observaram que 27 (11%) das 238 estirpes testadas foram produtoras de
substâncias antimicrobianas. Estas estirpes foram testadas contra estirpes-padrão de E. coli
e S. aureus e estirpes clínicas de S. aureus e S. epidermidis. O maior halo de inibição de
crescimento da estirpe-padrão S. aureus foi de 18mm. Em nosso trabalho, o menor halo de
inibição de crescimento desta estirpe foi de 19mm. Quanto à estirpe-padrão E. coli, apenas
quatro (14,8%) das 27 estirpes marinhas apresentaram atividade contra esta estirpe, com

77
halos de inibição de crescimento variando entre 12 e 14mm (Hentschel et al., 2001).
Enquanto que, em nosso trabalho, seis (37,5%) das 16 estirpes marinhas apresentaram
atividade, com halos de inibição de crescimento variando entre 12 e 25mm. Ainda neste
trabalho, Hentschel e colaboradores observaram um determinado padrão de inibição, onde
as estirpes marinhas Gram-positivas foram ativas contra estirpes indicadoras Gram-
positivas e estirpes marinhas Gram-negativas foram ativas contra estirpes indicadoras
Gram-negativas. Neste caso, foi sugerido que este padrão poderia ser provocado pela ação
de bacteriocinas. As bacteriocinas são polipeptídeos que atuam inibindo espécies
intimamente relacionadas e mantendo a estirpe produtora ilesa (BABA & SCHNEEWIND,
1998). Assim, estirpes Gram-negativas produzem bacteriocinas contra estirpes Gram-
negativas, e estirpes Gram-positivas produzem bacteriocinas contra estirpes Gram-
positivas. O padrão de inibição citado acima não ocorreu em nosso trabalho, onde se
observou um espectro de ação maior contra estirpes indicadoras Gram-positivas, porém, as
estirpes marinhas Gram-positivas e Gram-negativas foram ativas tanto contra estirpes
indicadoras Gram-negativas quanto Gram-positivas.
Em um outro trabalho, realizado por Muscholl-Silberhorn, Thiel e Imhoff (2008),
283 (15,2%) das 1.866 estirpes marinhas testadas foram produtoras de substâncias
antimicrobianas contra estirpes-padrão de E. coli, B. subtilis e Staphylococcus lentus. Esta
taxa só foi válida para o primeiro teste realizado a partir da transferência direta da colônia
da placa mãe para a placa de teste da atividade. As colônias foram, então, purificadas e
estocadas, e a seguir foi realizado o segundo teste de atividade. Observou-se que somente
163 das 283 estirpes marinhas apresentaram atividade antimicrobiana contra as mesmas
estirpes indicadoras testadas anteriormente. No terceiro teste, a queda foi mais acentuada,
onde apenas 42 das 283 estirpes mantiveram a atividade. Neste caso, foi sugerido que a

78
atividade foi perdida no decorrer das transferências das estirpes para o meio de cultura
fresco. Em nosso trabalho, a produção da substância antimicrobiana e a atividade
antimicrobiana subseqüente mostraram-se bastante estáveis no decorrer dos experimentos,
inclusive não apresentaram discrepâncias nos valores dos halos de inibição de crescimento
obtidos nos testes realizados em duplicata.
A ecologia microbiana presente no mesênquima das esponjas, considerando o
grande número de bactérias taxonomicamente diversas, fornece um ambiente rico em
interações ainda pouco conhecidas (IMHOFF & STÖHR, 2003). Poucos trabalhos têm se
dedicado a estudar como tais interações poderiam afetar a secreção de substâncias
antimicrobianas produzidas pelas estipes marinhas dentro das esponjas (ANGELL et al.,
2006). Em nosso trabalho, um determinado padrão foi observado entre estirpes marinhas
intimamente relacionadas: estas não apresentaram inibição umas contra as outras.
Inicialmente, levantou-se a hipótese de que a não inibição era devido ao fato de terem sido
isoladas da mesma esponja. Podemos citar como exemplo desta assertiva, as estirpes H40 e
H41 identificadas como Pseudomonas fluorescens e a estirpe H51 identificada como
Pseudomonas putida isoladas em meio BHI a partir da mesma espécie de esponja, porém
em coletas diferentes. No entanto, observou-se que estirpes isoladas de esponjas diferentes
também não inibiram umas as outras. Como foi o caso das estirpes Pm31, Ca31 e Mm37
(Pseudovibrio sp.) isoladas em meio Marine a partir das esponjas Paraleucilla magna,
Clathrina aurea e Mycale microsigmatosa, respectivamente.
Atualmente, sabe-se que, as estirpes produtoras de antibióticos desenvolveram
várias estratégias para evitar à ação de seus antibióticos sobre suas células
(YANAGISAWA & KAWAKAMI, 2003). Um exemplo desta assertiva é o antibiótico
mupirocina, isolado pela primeira vez a partir de uma estirpe de P. fluorescens (FULLER et

79
al., 1971). Sabe-se que este antibiótico é um potente inibidor de várias isoleucil-tRNA
sintetases (HUGHES & MELLOWS, 1980). Da mesma forma que outras estirpes
produtoras de antibióticos, a P. fluorescens é totalmente resistente à ação da mupirocina.
Neste caso, uma alteração estrutural da isoleucil-tRNA sintetases permitiu que as células de
P. fluorescens se tornassem resistentes à sua ação (HUGHES, MELLOWS &
SOUGHTON, 1980). Provavelmente, por serem intimamente relacionadas, as estirpes H40,
H41 e H51 isoladas em meio BHI, as estirpes Pm31, Ca31 e Mm37 isoladas em meio
Marine e as estirpes Mm33, H31, H42 e H43 isoladas em meio Marine-mar, desenvolveram
o mesmo mecanismo de defesa apresentado pela estirpe produtora intimamente relacionada.
Além desta, outras interações puderam ser observadas. As estirpes intimamente
relacionadas, Pc31, Pc32 e Dr31 (Bacillus pumilus), isoladas em meio BHI, apresentaram
perfis de inibição distintos. As estirpes Pc31 e Pc32 foram ativas contra quatro das 16
estirpes indicadoras testadas. Em contrapartida, a estirpe Dr31 foi ativa somente contra
duas estirpes indicadoras. Além disso, quando utilizada como indicadora, a estirpe Dr31
teve o seu crescimento inibido pela maioria das estirpes produtoras. Ainda entre as estirpes
isoladas em meio BHI, as estirpes H40 e H41 (P. fluorescens) inibiram o crescimento de
sete e de dez das 16 estirpes indicadoras testadas, respectivamente. Enquanto que, a estirpe
H51 (P. putida) só inibiu o crescimento de uma estirpe indicadora. Contrariamente, quando
utilizadas como indicadoras, estas estirpes não tiveram o seu crescimento inibido por
nenhuma das estirpes marinhas produtoras. As diferenças observadas nestes padrões de
inibição podem estar relacionadas com o tipo de substância antimicrobiana produzida.
Sabe-se, atualmente, que uma única espécie pode produzir substâncias distintas com ação
em diferentes alvos (BRINKHOFF et al., 2004). Análises recentes confirmam estes dados,
mostrando que a estrutura da população bacteriana presente na esponja é extremamente

80
organizada, e que esta organização depende das estirpes mais ativas (LO GIUDICE et al.,
2007).
As estirpes Pm31, Ca31 e Mm37 (Pseudovibrio sp.) isoladas em meio Marine
quando utilizadas como produtoras foram as estirpes que apresentaram o espectro de ação
mais amplo. Este tipo de comportamento corrobora com a hipótese de dominância dos
membros do gênero Pseudovibrio sobre a comunidade bacteriana das esponjas. Neste caso,
as substâncias antimicrobianas produzidas por estas estirpes seriam capazes de inibir
fortemente o crescimento de estirpes marinhas competidoras, além de serem efetivas na
proteção de seu hospedeiro (THIEL & IMHOFF, 2003). Uma outra particularidade foi
observada para estas estirpes. As estirpes Pm31, Ca31 e Mm37 não cresceram como
indicadoras sobre estirpes isoladas em meio BHI, mostrando-se pouco tolerantes a
concentrações de NaCl menores do que aquela encontrada no ambiente marinho.
As estirpes Mm33, H31, H42 e H43 isoladas em meio Marine-mar se mostraram
mais ativas contra bactérias marinhas do que contra bactérias terrestres, onde juntas
inibiram 9 das 16 estirpes indicadoras testadas. Em relação às estirpes terrestres, somente a
indicadora C. fimi teve seu crescimento inibido por estas estirpes marinhas. Nos testes de
produção de substâncias antimicrobianas, somente as estirpes que produzem
constitutivamente tais substâncias são rastreadas com sucesso. Entretanto, sabe-se
atualmente, que há estirpes marinhas que só produzem substâncias antimicrobianas em
resposta a sinais químicos enviados por estirpes marinhas competidoras (MEARNS-
SPRAGG et al., 1998).
O terceiro passo do trabalho foi identificar as estirpes produtoras de substâncias
antimicrobianas que apresentaram um espectro de ação mais abrangente. A identificação
molecular foi realizada em paralelo com a caracterização fenotípica. A partir do

81
seqüenciamento parcial do gene que codifica para o 16S rRNA, as nove estirpes puderam
ser organizadas em três grupos. O primeiro grupo, composto por três estirpes Gram-
positivas móveis, de morfologia colonial creme e rugosa e formadoras de esporos, foi
identificado como pertencente ao filo Firmicutes, classe Bacilli, família Bacillaceae e
gênero Bacillus. A estirpe Pc31 foi identificada através do seqüenciamento parcial do gene
que codifica para o 16S rRNA como Bacillus pumilus ou Bacillus subtilis com 99% de
homologia cada. Com o auxílio do sistema comercial miniaturizado API 50CH foi possível
identificar esta estirpe como B. pumilus com 99,9% de identidade. Enquanto que, as
estirpes Pc32 e Dr31 foram identificadas através do seqüenciamento como Bacillus pumilus
com 99% e 96% de homologia, respectivamente. Com o auxílio do API, foi possível
identificar estas estirpes como B. pumilus com 99,9% de identidade.
O segundo grupo, composto de três estirpes Gram-negativas móveis, de morfologia
colonial bege e mucóide e produtoras de pigmento no meio, foi identificado como
pertencente ao filo Proteobacteria, classe Gammaproteobacteria, família
Pseudomonadaceae e gênero Pseudomonas. As estirpes H40 e H41 foram identificadas
através do seqüenciamento parcial do gene que codifica para o 16S rRNA como
Pseudomonas fluorescens ou Pseudomonas gessarddi com 97% de homologia cada. Com o
auxílio do API 20NE, foi possível identificar estas estirpes como P. fluorescens com 98%
de identidade. Enquanto que, a estirpe H51 foi identificada através do seqüenciamento
como membro do gênero Pseudomonas com 96% de homologia. Com o auxílio do API, foi
possível identificar esta estirpe como Pseudomonas putida com 99,6% de identidade.
O terceiro grupo, composto por três estirpes Gram-negativas móveis e de
morfologia colonial marrom e mucóide, foi identificado como pertencente ao filo
Proteobacteria, classe Alphaproteobacteria, família Rhodobacteraceae e gênero

82
Pseudovibrio. As estirpes Pm31 e Ca31 foram identificadas através do seqüenciamento
parcial do gene que codifica para o 16S rRNA como Pseudovibrio ascidiaceicola ou
Pseudovibrio japonicus com 96% de homologia cada. Enquanto que, a estirpe Mm37 foi
identificada através do seqüenciamento como Pseudovibrio denitrificans com 96% de
homologia. Estas estirpes não foram identificadas pelo API, porque este método não possui
um sistema de identificação específico para o gênero Pseudovibrio.
Estes resultados estão de acordo com aqueles descritos por Muscholl-Silberhorn,
Thiel e Imnhoff (2008). Através do seqüenciamento do gene que codifica para o 16S rRNA
foram identificadas 39 estirpes produtoras de substâncias antimicrobianas isoladas de dez
espécies de esponjas coletadas no mar Mediterrâneo. Estas estirpes puderam ser agrupadas
em três filos: Proteobacteria (Alpha e Gammaproteobacteria), Firmicutes e
Actinobacteria. Interessantemente, 21 (54%) das 39 estirpes foram identificadas como
membros do gênero Pseudovibrio (Alphaproteobacteria). Além disso, quatro estirpes
(10%) foram identificadas como integrantes do gênero Pseudomonas
(Gammaproteobacteria) e 11 estirpes (28%) foram identificadas como integrantes do
gênero Bacillus (Firmicutes). Em um trabalho realizado por Anand e coloboradores (2005),
os gêneros Vibrio, Pseudomonas e Bacillus representaram a maioria das estirpes produtoras
de substâncias antimicrobianas isoladas de quatro espécies de esponjas coletadas na Índia.
Hentschel e colaboradores (2001) identificaram 27 estirpes produtoras de substâncias
antimicrobianas, as quais foram predominantemente agrupadas como membros dos gêneros
Pseudoalteromonas (Gammaproteobacteria), Pseudovibrio e Bacillus. Desta forma,
podemos concluir que independentemente da localização geográfica ou da espécie de
esponja coletada, os filos Proteobacteria, Firmicutes e Actinobacteria são
predominantemente produtores de substâncias antimicrobianas nas esponjas marinhas.

83
O gênero Pseudovibrio foi descrito em 2004 por Shieh, Lin e Jean e continha uma
única espécie, Pseudovibrio denitrificans, isolada da água do mar em Taiwan. Uma
segunda espécie, Pseudovibrio ascidiaceicola, isolada de uma ascídia (um invertebrado
marinho), foi descrita posteriormente por Fukunaga e colaboradores (2006). Em 2007, uma
nova espécie foi descrita por Hosoya e Yokota, Pseudovibrio japonicus, isolada da água do
mar no Japão. As espécies pertencentes a este gênero se caracterizam como bacilos Gram-
negativos, halofílicas, anaeróbias facultativas, móveis, catalase e oxidase positivas e de
morfologia colonial circular, mucóide e de cor marrom. Análises filogenéticas baseadas no
seqüenciamento do gene que codifica para o 16S rRNA demonstraram que este gênero
pertence ao filo Proteobacteria (Alphaproteobacteria) e está intimamente relacionado com
os gêneros Pannonibacter, Roseibium e Stappia (SHIEH et al., 2004; FUKUNAGA et al.,
2006).
Segundo Stackebrandt e Goebel (1994), duas estirpes definitivamente pertencem a
espécies diferentes se compartilham menos de 97,5% de similaridade de seqüência parcial
do gene que codifica para o 16S rRNA, e são membros de gêneros diferentes se
compartilham similaridade inferior a 93%. Desta forma, as estirpes Pm31 e Ca31
identificadas através do seqüenciamento parcial do gene que codifica para o 16S rRNA
como P. ascidiaceicola ou P. japonicus com 96% de homologia, e a estirpe Mm37
identificada como P. denitrificans também com 96% de homologia, podem representar uma
nova espécie deste gênero. As características fenotípicas principais do gênero foram
mantidas nas estirpes caracterizadas no presente trabalho, onde se apresentaram como
bacilos Gram-negativos, halofílicas, móveis, catalase e oxidase positivas e de morfologia
colonial circular, mucóide e de cor marrom. Testes adicionais serão necessários,
principalmente aqueles relacionados à capacidade de crescimento anaeróbio e de produção

84
de ácido a partir de determinadas fontes de carbono, no intuito de comparar com os
resultados observados para as espécies já descritas deste gênero.
Membros do gênero Pseudovibrio são freqüentemente encontrados dominando a
fração cultivável da comunidade bacteriana associada às esponjas (TAYLOR et al., 2007).
Além disso, membros deste gênero também foram detectados a partir de técnicas como a
eletroforese em gel com gradiente desnaturante (DGGE) e a hibridização in situ
(WEBSTER & HILL, 2001; THOMS et al., 2003; ENTICKNAP et al., 2006). Em um
trabalho realizado por Enticknap e colaboradores (2006), foi demonstrado a transferência
vertical destas estirpes, via gametas, para novas gerações da esponja Mycale laxissima.
Diante do exposto, há fortes evidências de que os membros deste gênero sejam simbiontes
verdadeiros.
As estirpes Pm31, Ca31 e Mm37 descritas no presente trabalho pertencem ao
mesmo gênero (Pseudovibrio sp.) que as estirpes marinhas produtoras de substâncias
antimicrobianas descritas por Muscholl-Silberhorn, Thiel e Imnhoff (2008), Thakur e
colaboradores (2005) e Hentschel e colaboradores (2001). No trabalho realizado por
Hentschel e colaboradores (2001) foi demonstrado que uma estirpe chamada de SB89,
isolada da esponja Aplysina aerophoba, apresentou atividade antimicrobiana contra as
estirpes-padrão E. coli e S. aureus com halos de inibição de crescimento de 13mm. Além
disso, esta estirpe marinha também foi testada contra estirpes clínicas de S. aureus e S.
epidermidis, onde observou-se halos de inibição de crescimento variando entre 13 e 15mm.
Em contrapartida, no presente trabalho, as estirpes Pm31, Ca31 e Mm37 foram ativas
contra cerca de 60% das estirpes indicadoras testadas, em sua maioria Gram-positivas. Em
relação a estirpe-padrão E. coli, somente a estirpe Mm37 conseguiu inibir o seu
crescimento apresentando halo de inibição de 12mm. Enquanto que, em relação a estirpe-

85
padrão S. aureus e as estirpes clínicas de S. aureus e S. epidermidis, as três estirpes
marinhas foram bastante ativas, apresentando halos de inibição de crescimento variando
entre 15 e 28mm.
Ainda entre os bacilos Gram-negativos isolados das esponjas, as estirpes H40 e H41
identificadas como P. fluorescens e a estirpe H51 identificada como P. putida foram
aquelas que apresentaram um maior espectro de ação, inibindo o crescimento de cerca de
90% das estirpes-padrão e clínicas utilizadas neste estudo. Recentemente, Ahmed e
colaboradores (2008) descreveram o isolamento de duas estirpes de Pseudomonas sp. (P.
putida e P. aeruginosa) a partir de esponjas coletadas no Paquistão. Os perfis de inibição de
crescimento das estirpes multirresistentes de S. aureus, S. epidermidis e E. coli foram
bastante semelhantes para as duas estirpes, com halos variando entre 14 e 18mm. As
estirpes H40, H41 e H51 também foram ativas contra estas indicadoras. Em relação às
estirpes-padrão de S. aureus, S. epidermidis e E. coli, os halos de inibição de crescimento
variaram entre 15 e 30mm. Interessantemente, para as estirpes clínicas destas espécies, os
halos de inibição de crescimento variaram entre 17 e 43mm. Em um outro estudo, uma
estirpe de P. fluorescens produtora de substância antimicrobiana foi isolada de uma espécie
de tunicata (um invertebrado marinho) coletada no Alaska (NEEDHAM et al., 1994). Esta
estirpe foi ativa contra estirpes clínicas multirresistentes de S. aureus e E. faecium e menos
ativa contra uma estirpe-padrão de P. aeruginosa. Até o presente momento, não foram
descritos trabalhos demonstrando o isolamento de estirpes de P. fluorescens produtoras de
substâncias antimicrobianas isoladas de esponjas marinhas. Porém, em estudos sobre
diversidade bacteriana, esta espécie já foi detectada como residente nas esponjas (KIM &
FUERST, 2006).

86
As estirpes Gram-positivas isoladas no presente trabalho e identificadas como
Bacillus pumilus apresentaram perfis de inibição bastante distintos. As estirpes Pc31 e Pc32
foram ativas contra cerca de 53% das estirpes indicadoras testadas. Por outro lado, a estirpe
Dr31 inibiu somente 13% destas. Além disso, a estirpe Dr31 só foi ativa contra estirpes
Gram-positivas, enquanto que, as estirpes Pc31 e Pc32 foram ativas tanto para Gram-
positivos quanto para Gram-negativos (em menor proporção). Em um trabalho realizado
por Pabel e colaboradores (2003), foi observado que dentre as 182 estirpes isoladas da
esponja Aplysina aerophoba, quatro produziram substâncias antimicrobianas e foram
identificadas como membros do gênero Bacillus. Três estirpes foram identificadas como B.
subtilis e uma como B. pumilus. As estirpes de B. subtilis foram ativas contra estirpes-
padrão de S. aureus e E. coli, enquanto que, a estirpe de B. pumilus foi ativa somente contra
S. aureus. Em um outro trabalho, realizado por Romanenko e colaboradores (2006), duas
estirpes de B. pumilus isoladas a partir de um molusco marinho também apresentaram
perfis de inibição distintos. A estirpe An114 foi ativa contra as estirpes-padrão de B.
subtilis (com halo de inibição de 10mm), E. faecium (com halo de inibição de 14mm) e S.
aureus (com halo de inibição de 20mm). Enquanto que, a estirpe An118 foi ativa somente
contra S. aureus, com halo de inibição de crescimento de 13mm. Além disso, estas estirpes
não foram ativas contra estirpes-padrão de Enterobacter cloacae e P. aeruginosa. Desta
forma, estes resultados sugerem que a atividade inibitória pode estar mais relacionada com
a estirpe produtora e com o tipo de substância antimicrobiana produzida, do que com a
espécie ao qual ela pertence (LO GIUDICE et al., 2007).
Neste trabalho, as características fenotípicas apresentadas por cada estirpe estavam
de acordo com aquelas descritas para a espécie ou para o gênero aos quais pertenciam. Uma
notável exceção foi a estirpe H51 (P. putida), que demonstrou a capacidade de crescer e de

87
produzir a substância também a 42°C. Em geral, as estirpes desta espécie não apresentam
tal característica (ROMANENKO et al., 2005). Este fato não nos surpreendeu, já que uma
estirpe de P. putida previamente isolada em nosso laboratório a partir da esponja Mycale
microsigmatosa também apresentou a capacidade de crescer e de produzir a substância
antimicrobiana à 42°C. Além disso, uma estirpe de P. putida com esta mesma característica
foi isolada a partir de águas subterrâneas na Pensilvânia. Esta estirpe foi patentiada (US
Patent 5,821,112) em 1998 e tem como aplicação principal a sua utilização na redução de
odores e degradação parcial de resíduos orgânicos de origens humana e animal, acelerando
a decomposição de resíduos biológicos (BOTTO & GEISLER, 1998).
As condições de temperatura para o crescimento foram analisadas e as estirpes
isoladas no presente trabalho foram consideradas mesófilas, pois todas cresceram em
temperaturas entre 15-42°C. As condições de salinidade para o crescimento foram
analisadas para as estirpes isoladas em meio BHI, que foram consideradas halofílicas
discretas (toleram concentrações de 1-6%), pois cresceram em concentrações de 1-2% de
NaCl. As estirpes Pm31, Ca31 e Mm37 (Pseudovibrio sp.) também foram consideradas
halofílicas discretas, pois cresceram em uma concentração de 2% de NaCl. Atualmente,
está sendo testada a capacidade de crescimento das nove estirpes frente a concentrações de
3, 5, 10, 15 e 20% de NaCl.
As estirpes Pm31 e Ca31 (Pseudovibrio sp.) cresceram aerobicamente no meio de
cultura em uma faixa de temperatura entre 15-30°C, sendo o crescimento ótimo ocorrendo
à temperatura ambiente. A estirpe Mm37 (Pseudovibrio sp.) cresceu em uma faixa de
temperatura entre 15-37°C, sendo o crescimento ótimo obtido entre 25-37°C. Não foi
observado crescimento a 42°C. Estes resultados foram semelhantes aos apresentados pelas
estirpes-padrão DN33 de P. denitrificans (SHIEH, LIN & JEAN, 2004), F423 de P.

88
ascidiaceicola (FUKUNAGA et al., 2006) e WSF2 de P. japonicus (HOSOYA &
YOKOTA, 2007). As estirpes Pm31 e Ca31 apresentaram a faixa e a temperatura ótima de
crescimento idênticas às observadas para a estirpe WSF2 de P. japonicus. No entanto, a
estirpe Mm37 apresentou uma faixa de crescimento mais abrangente, semelhante àquela
apresentada pela estirpe DN33 de P. denitrificans, na qual cresceu em uma faixa de
temperatura entre 20-35°C, com o crescimento ótimo ocorrendo entre 30-35°C.
As estirpes H40 e H41 (P. fluorescens) cresceram aerobicamente no meio de cultura
em uma faixa de temperatura entre 15-37°C, sendo o crescimento ótimo ocorrendo em toda
essa faixa. Não foi observado crescimento a 42°C. A estirpe H51 (P. putida) cresceu em
uma faixa de temperatura entre 15-42°C, sendo o crescimento ótimo obtido entre 25-42°C.
As estirpes H40 e H41 apresentaram a mesma faixa de crescimento descrita para a estirpe-
padrão de P. fluorescens (ROMANENKO et al., 2005). Enquanto que, a estirpe H51
apresentou uma faixa mais ampla do que a observada para a estirpe-padrão de P. putida
(15-37°C) (ROMANENKO et al., 2005). Até o presente momento, não foram descritos
trabalhos que comparassem o crescimento em diferentes temperaturas das estirpes de
Pseudomonas sp. isoladas de esponjas. Como via de regra, a maioria das estirpes marinhas
deste gênero teve o seu crescimento analisado somente à temperatura ambiente, o que
variou de uma região para outra (ROMANENKO et al., 2005; LO GIUDICE, BRUNI &
MICHAUD, 2007).
Em relação às condições de salinidade do meio, as estirpes de Pseudomonas sp.
apresentaram padrões distintos de crescimento. As estirpes H40 e H41 (P. fluorescens)
cresceram melhor nas concentrações de 1, 1,5 e 2% de NaCl. Enquanto que, a estirpe H51
(P. putida) apresentou um melhor crescimento na concentração de NaCl do próprio meio de
isolamento (meio BHI – 0,5% de NaCl). Estes resultados correspondem aos descritos para

89
as estirpes-padrão de P. fluorescens e P. putida, que não toleraram concentrações de NaCl
maiores que 8% (ROMANENKO et al., 2005). Em um trabalho realizado por Lo Giudice,
Bruni & Michaud (2007), uma estirpe de Pseudomonas sp. isolada a partir da água do mar
tolerou uma concentração mínima de 0% e máxima de 5% de NaCl. Normalmente, a
maioria dos trabalhos realizados para as estirpes marinhas de Pseudomonas sp. utilizaram
uma concentração de 2% ou 3% de NaCl, correspondente àquelas encontradas no meio de
cultivo padrão (meio Marine) e na água do mar, respectivamente (LO GIUDICE, BRUNI &
MICHAUD , 2007; AHMED et al., 2008).
As estirpes Pc31, Pc32 e Dr31 (B. pumilus) apresentaram padrões de crescimento
muito semelhantes. Estas cresceram aerobicamente no meio de cultura em uma faixa de
temperatura entre 15-42°C, sendo o crescimento ótimo ocorrendo entre 25-42°C. Por outro
lado, estas estirpes apresentaram padrões distintos de tolerância ao NaCl. A estirpe Dr31
cresceu melhor nas concentrações de 1, 1,5 e 2% de NaCl. Enquanto que, as estirpes Pc31 e
Pc32 apresentaram um melhor crescimento nas concentrações de NaCl do próprio meio de
isolamento (BHI – 0,5%). Em um trabalho realizado por Ivanova e colaboradores (1999),
estirpes marinhas de Bacillus sp. produtoras de substâncias antimicrobianas foram isoladas
a partir de esponjas marinhas coletadas no Japão e caracterizadas fenotipicamente. O
crescimento destas estirpes foi analisado nas temperaturas de 4°, 10°, 25°, 30°, 37°, 40°,
50° e 55°C e nas concentrações de 0, 1, 3, 7, 10, 12 e 15% de NaCl. As estirpes de B.
pumilus cresceram aerobicamente no meio de cultura em uma faixa de temperatura entre
15-50°C, sendo o crescimento ótimo ocorrendo entre 28-37°C. Além disso, foi observado
que estas estirpes foram capazes de crescer nas concentrações de 0-7% e 10% de NaCl. Os
resultados obtidos para as estirpes de B. pumilus isoladas no presente trabalho foram muito
semelhantes aos descritos para as estirpes isoladas por Ivanova e colaboradores (1999).

90
Estes resultados dão suporte a teoria de que a comunidade bacteriana das esponjas é estável
e que as características fenotípicas que definem as espécies são mantidas
independentemente da região geográfica ou da espécie de esponja estudada (LO GIUDICE,
BRUNI & MICHAUD, 2007).
Atualmente, há uma crescente preocupação com a presença de resíduos
farmacêuticos em ambientes aquáticos e seus possíveis impactos ambientais. Após a
administração de medicamentos, como, por exemplo, antibióticos e hormônios, uma parte
significativa destes fármacos é excretada por humanos no esgoto doméstico. No entanto,
estudos demonstraram que muitos resíduos de fármacos resistem a vários processos de
tratamento convencional da água (OLIVEIRA & VON SPERLING, 2007). Além disso,
muitos países em desenvolvimento, como o Brasil, despejam os esgotos não tratados
diretamente nos mares (COOPER, SIEWICKI & PHILLIPS, 2007). Este comportamento
pode gerar duas conseqüências principais: a exposição crônica dos organismos marinhos à
concentrações destes fármacos e o carreamento de diversos microrganismos nocivos, como
bactérias, vírus e larvas de parasitas (COOPER, SIEWICKI & PHILLIPS, 2007). No caso
dos antibióticos, a exposição crônica de bactérias marinhas a baixas concentrações destes
fármacos pode induzir a expressão de genes que codifiquem mecanismos de resistência às
drogas (HIRSCH et al., 1999). Kolpin e colaboradores (2002) detectaram concentrações
dos antibióticos (na ordem de ng/l e µg/l) fluoroquinolonas (ciprofloxacina e norfloxacina),
lincomicina, macrolídeos (claritromicina e roxitromicina), sulfonamidas (sulfadimetoxina,
sulfametazina, e sulfametoxazol), tetraciclinas (clorotetraciclina, oxitetraciclina e
tetraciclina), tilosina e trimetoprim, em amostras de águas superficiais nos Estados Unidos.
Resultados similares foram obtidos por Mudryk & Skórczewski (1998) a partir de amostras
coletadas no mar Báltico.

91
Além da resistência intrínseca, estudos indicaram que níveis variáveis de resistência
em bactérias marinhas também podem ser mediados pela transferência de plasmídeos de
resistência carreados por bactérias terrestres trazidas ao ambiente marinho por ação
antropogênica (CHANDRASEKARN, VENKATESH & LAITHAKUMARI, 1998;
TENDENCIA & DE LA PENA, 2001). Um exemplo desta assertiva foi demonstrado por
Thavasi e colaboradores (2007), em um estudo realizado na Índia. Estirpes bacterianas
foram isoladas a partir de amostras de sedimentos e da água do mar. Vinte e duas estirpes
apresentaram resistência a grande parte (entre 5 e 13) dos 20 antibióticos testados. Foi
demonstrado que estas estirpes pertenciam, basicamente, a sete gêneros e dez espécies.
Dentre as Gram-negativas, foram identificadas estirpes das espécies Citrobacter
intermedius, Enterobacter aerogenes, Klebsiella ozaenae, Moraxella catarrhalis, Neisseria
mucosa, Neisseria sicca e P. fluorescens. Dentre as Gram-positivas, foram identificadas
estirpes das espécies Bacillus megaterium, Bacillus subtilis e Micrococcus luteus. Quando
foi realizada a cura dos plasmídeos com laranja de acridina, a resistência contra
amoxicilina, ampicilina, canamicina, cloranfenicol, penicilina G e tetraciclina foi
totalmente perdida por todas as estirpes. Estes resultados confirmam o papel dos
plasmídeos em mediar a resistência aos antimicrobianos no ambiente marinho. Em nosso
laboratório, foi analisado o perfil plasmidial das estirpes isoladas no presente trabalho.
Análises preliminares demonstraram que as três estirpes de Bacillus sp. e uma de
Pseudomonas sp. apresentaram em eletroforese em gel de agarose pelo menos uma forma
plasmidial com tamanhos entre 8,0 e 10,4 kb e maior que 27kb, respectivamente.
Atualmente, estão sendo realizadas análises sobre a função destes plasmídeos nestas
estirpes marinhas.

92
As estirpes marinhas isoladas no presente trabalho tiveram os seus perfis de
susceptibilidade aos antimicrobianos analisados. As estirpes Pm31, Ca31 e Mm37 de
Pseudovibrio sp. foram resistentes aos antibióticos ciprofloxacina e gentamicina. Além
destes antibióticos, as estirpes Pm31 e Mm37 foram resistentes ao sulfazotrim, sendo esta
última também resistente à ceftazidima. Em um trabalho realizado por Shieh, Lin & Jean
(2004), foi analisado o perfil de susceptibilidade da estirpe-padrão DN33 de P.
denitrificans. Esta estirpe foi resistente à clindamicina, colistina, estreptomicina,
gentamicina, lincomicina, penicilina G e polimixina B. Além disso, demonstrou
sensibilidade intermediária ao ácido nalidíxico e à tetraciclina e foi sensível à ampicilina,
carbenicilina, cefalotina, cloranfenicol, eritromicina, neomicina, novobiocina e
vancomicina. A gentamicina foi o único antibiótico no qual as estirpes Pm31, Ca31 e
Mm37 também foram resistentes. Não foram descritos por Shieh, Lin & Jean (2004) os
tamanhos dos halos de inibição de crescimento da estirpe DN33 nos testes de
susceptibilidade aos antimicrobianos. Portanto, não foi possível comparar com os dados
observados e descrever os perfis de sensibilidade das estirpes de Pseudovibrio sp. isoladas
neste trabalho. Até o presente momento, não foram encontrados trabalhos que relatassem o
perfil de susceptibilidade das outras espécies do gênero Pseudovibrio.
As estirpes H40 e H41 (Pseudomonas fluorescens) e a estirpe H51 (Pseudomonas
putida) foram consideradas resistentes aos antibióticos cefalexina e sulfazotrim. Além
destes antibióticos, as estirpes H40 e H41 foram consideradas resistentes ao aztreonam e a
estirpe H51 foi considerada resistente ao cloranfenicol. Provavelmente, as estirpes H40 e
H41 também seriam resistentes ao cloranfenicol, pois apresentaram halos de inibição muito
pequenos (9 e 10mm, respectivamente). Até o presente momento, não foram encontrados
trabalhos que relatassem o perfil de sensibilidade das espécies do gênero Pseudomonas

93
isoladas das esponjas marinhas. Em contrapartida, foram relatados vários estudos sobre os
perfis de susceptibilidade de estirpes de Pseudomonas sp. isoladas da água do mar. Em um
trabalho realizado por De Souza e colaboradores (2006), estirpes do gênero Pseudomonas
foram isoladas a partir da água do mar coletada na costa indiana e testadas quanto ao perfil
de susceptibilidade aos antimicrobianos. Estas estirpes demonstraram perfis de resistência
variáveis aos antibióticos ampicilina, canamicina, cloranfenicol, estreptomicina e
tetraciclina. Em um outro trabalho, 33 estirpes de P. aeruginosa foram isoladas a partir da
água do mar coletada no Japão (KHAN et al., 2007). Dos 22 antibióticos testados, todas as
estirpes foram resistentes à pelo menos seis deles: amoxicilina-ácido clavulânico,
ampicilina, ampicilina-sulbactam, cefazolina, cefpodoxima e cefuroxima. Além disso, todas
foram sensíveis aos antibióticos amicacina, cefepima, ceftazidima, gentamicina, imipenem,
levofloxacina, meropenem, ofloxacina, piperacilina e piperaciclina-tazobactam. Por outro
lado, uma estirpe de Pseudomonas sp. isolada a partir da água do mar coletada na Antártida
demonstrou resistência à penicilia G e tetraciclina e foi sensível ao ácido nalidíxico,
ampicilina, cloranfenicol, polimixina B e tobramicina (LO GIUDICE, BRUNI &
MICHAUD, 2007). Estes resultados sugerem que os perfis de susceptibilidade aos
antimicrobianos são variáveis e dependentes tanto da diversidade metabólica de cada
espécie quanto do local de isolamento das estirpes deste gênero (MOORE et al., 1996;
ROMANENKO et al., 2005). Em nosso trabalho, as estirpes marinhas foram isoladas a
partir de esponjas coletadas na costa da cidade do Rio de Janeiro. Pode ser que este fato
tenha influenciado no perfil de resistência aos antimicrobianos apresentados por estas
estirpes.
As estirpes Pc31, Pc32 e Dr31 (B. pumilus) tiveram o crescimento inibido por todos
os antibióticos testados no presente trabalho. Resultados semelhantes foram observados

94
para as estirpes marinhas de Bacillus sp. produtoras de substâncias antimicrobianas isoladas
a partir de esponjas marinhas coletadas no Japão (IVANOVA et al., 1999). Estas estirpes
foram consideradas sensíveis à ampicilina, carbenicilina, oxacilina e penicilina G, com
halos de inibição de crescimento de 28-35mm, e ao cloranfenicol e à oleandomicina com
halos de 25mm. As estirpes Pc31, Pc32 e Dr31 apresentaram halos de inibição de
crescimento de 40-45mm para penicilina G, 22-25mm para o cloranfenicol e 35-39 para
eritromicina (análogo à oleandomicina). Desta forma, ao comparar os resultados pôde-se
concluir que estirpes Pc31, Pc32 e Dr31 foram sensíveis aos antibióticos cloranfenicol,
eritromicina e penicilina G. De um modo geral, as estirpes marinhas do gênero Bacillus
apresentam grande sensibilidade aos antibióticos, principalmente os penicilínicos. Um
exemplo desta assertiva foi descrito por Caccamo e colaboradores (2000), que descreveram
o isolamento de estirpes de uma nova espécie deste gênero, a partir de amostras de
sedimentos coletadas na Itália. Foi observado que, dos dez antibióticos testados, a
polimixina B foi o único antibiótico no qual estas estirpes apresentaram resistência. Desta
forma, estas foram sensíveis ao ácido nalidíxico, bacitracina, canamicina, cloranfenicol,
eritromicina, estreptomicina, novobiocina, penicilina G e tetraciclina.
Atualmente, sabe-se que as substâncias antimicrobianas de natureza protéica,
principalmente as bacteriocinas, podem ser sensíveis a várias enzimas proteolíticas ou
resistentes a uma ou outra enzima (NASCIMENTO et al., 2004). Em relação às enzimas
pronase, protease, proteinase K e tripsina, utilizadas no presente trabalho, a pronase é a
mais eficiente na clivagem proteolítica das substâncias. Esta propriedade é atribuída à sua
composição, que compreende uma mistura de endopeptidases (atuam preferencialmente nas
regiões internas da cadeia polipeptídica) e exopeptidases (atuam somente nas extremidades
das cadeias polipeptídicas). As enzimas protease, proteinase K e tripsina atuam somente

95
como endopeptidases (BARRETT, RAWLINGS & WOESSNER, 2003). Sabe-se também,
que a maior parte dos produtos naturais marinhos, principalmente os policetídeos, possuem
em sua estrutura grupamentos ácidos que são sensíveis à ação de hidróxidos cáusticos,
como o NaOH (PAUL et al., 2007).
Neste contexto, as estirpes Pm31, Ca31 e Mm37 (Pseudovibrio sp.) apresentaram
diferenças em relação à natureza química e às condições de temperatura para a produção de
suas substâncias antimicrobianas. A SAM produzida pela estirpe Ca31 foi mais ativa à
temperatura ambiente e foi sensível à ação da enzima proteolítica pronase e do NaOH,
indicando ter como características, natureza protéica e ácida. Enquanto que, a SAM
produzida pela estirpe Mm37 foi mais ativa em uma faixa entre 25-37°C e foi sensível à
fosfolipase C e ao NaOH, indicando ter como características, natureza lipídica e ácida.
Desta forma, esta SAM poderia pertencer à classe dos policetídeos, que são formados por
cadeias de ácidos graxos ligados a grupamentos carboxila e são ativos contra estirpes
Gram-positivas e Gram-negativas (STAUNTON & WEISSMAN, 2001). Por outro lado, a
SAM produzida pela estirpe Pm31 foi mais ativa à temperatura ambiente e foi resistente à
ação de todas as enzimas proteolíticas testadas, assim como, resistente à ação da fosfolipase
C e do NaOH. Provavelmente, a SAM produzida por esta estirpe não deve possuir uma
natureza protéica, fosfolipídica e nem ácida, ou a sua estrutura química a torna resistente à
ação enzimática e do NaOH. Um estudo descreveu o isolamento de uma estirpe de P.
denitrificans produtora de um pigmento vermelho a partir de uma ascídia (GUZMAN et al.,
2007). Este pigmento era derivado da prodigiosina, um composto heterocíclico, e
apresentou atividade contra S. aureus. Compostos heterocíclicos são compostos cíclicos
contendo, no anel, um ou mais átomos diferentes do carbono. Sabe-se que estes anéis
conferem grande estabilidade química ao composto, tornando-os extremamente resistentes

96
à ação enzimática (PAUL et al., 2007). Até o presente momento, o trabalho supracitado foi
o único que identificou e caracterizou uma substância antimicrobiana produzida por
estirpes do gênero Pseudovibrio.
Os metabólitos antibióticos de Pseudomonas sp. incluem alcalóides (moléculas
cíclicas), peptídeos cíclicos e seus derivados, piranos (moléculas cíclicas), policetídeos
(cadeias de ácidos graxos de tamanhos variados) e quinolonas (moléculas cíclicas)
(JAYATILAKE et al., 1996; BULTEL-PONCÉ et al., 1999; SINGH et al., 2003; ). A
substância antimicrobiana produzida pela estirpe H40 (P. fluorescens) foi mais ativa em
uma faixa entre 15-30°C e na concentração de 0,5% de NaCl. Além disso, foi sensível à
ação da fosfolipase C, indicando ter como característica uma natureza lipídica. Desta
forma, esta SAM poderia pertencer à classe dos policetídeos, semelhante a SAM produzida
pela estirpe Mm37 (Pseudovibrio sp.). As SAM das estirpes H41 (P. fluorescens) e H51 (P.
putida) foram mais ativas em uma faixa entre 15-30°C e 25-42°C, respectivamente, e na
concentração de 0,5% de NaCl. Além disso, foram resistentes à ação de enzimas
proteolíticas, fosfolipase C e NaOH. Desta forma, as SAM produzidas por estas estirpes
não devem possuir uma natureza protéica, fosfolipídica e nem ácida, ou as suas estruturas
químicas a tornam resistentes à ação enzimática e do NaOH. Neste contexto, estas SAM
poderiam pertencer às classes de alcalóides, peptídeos cíclicos e seus derivados, piranos,
polissacarídeos sulfatados ou quinolonas. Os polissacarídeos sulfatados possuem
características estruturais distintas e importantes atividades biológicas têm sido descritas
em uma diversidade de invertebrados e microrganismos marinhos. Atualmente, sabe-se
que, algumas bactérias apresentam a capacidade de produzir estes compostos em maior
quantidade quando cultivadas em meios de cultura ricos em nutrientes (BHAKUNI &
RAWAT, 2005). Em nosso trabalho, as estirpes H41 e H51 foram isoladas a partir do BHI,

97
que por ser um meio rico, poderia ser aquele que melhor ofereceria as condições
nutricionais como as encontradas no seu ambiente de origem.
Os metabólitos antibióticos de Bacillus sp. incluem lantibióticos, lipopeptídeos,
peptídeos cíclicos e seus derivados e policetídeos (DE ROSA, MITOVA &
TOMMONARO, 2003; NAGAI et al., 2003; PABEL et al., 2003). As substâncias
antimicrobianas produzidas pelas estirpes Pc31 e Pc32 (Bacillus pumilus) apresentaram
características muito semelhantes. Ambas foram resistentes à ação de enzimas proteolíticas,
fosfolipase C e NaOH, indicando não ter como características uma natureza protéica,
fosfolipídica e ácida. Assim como, foram mais ativas à temperatura ambiente. A única
diferença foi acerca das condições de salinidade para a produção das SAM. A SAM da
estirpe Pc31 apresentou a mesma atividade em todas as concentrações (0,5, 1, 1,5, 2%) de
NaCl testadas. Enquanto que, a SAM da estirpe Pc32 foi mais ativa na concentração de
0,5% de NaCl. Por outro lado, a SAM produzida pela estirpe Dr31 (B. pumilus) foi mais
ativa em uma faixa entre 25-42°C e na concentração de 0,5% de NaCl. Além disso, foi
sensível às enzimas pronase e proteinase K e ao NaOH, indicando ter como características,
natureza protéica e ácida. Como citado anteriormente, as SAM produzidas pelas estirpes
Pc31 e Pc32 foram as mais ativas contra as estirpes indicadoras Gram-positivas, enquanto
que, a SAM produzida pela estirpe Dr31 apresentou um espectro de ação bastante reduzido,
inibindo somente o crescimento de algumas estirpes Gram-positivas. Devido a sua natureza
protéica e ácida, e sua capacidade de inibir estirpes Gram-positivas, a SAM produzida pela
estirpe Dr31 poderia pertencer à classe das bacteriocinas. A maioria das bacteriocinas
demonstra um espectro de ação limitado, agindo somente contra estirpes filogeneticamente
relacionadas (BRUNO & MONTVILLE, 1993). No entanto, já foram descritas
bacteriocinas produzidas por estirpes Gram-positivas capazes de inibir diversos grupos de

98
estirpes Gram-positivas e alguns grupos de estirpes Gram-negativas (KLAENHAMMER,
1988; DE VUYST & VANDAMME, 1994; ZHENG et al., 1999). Além disso, as
bacteriocinas podem formar grandes complexos formados por proteínas, fosfolipídeos e/ou
carboidratos (BRUNO & MONTVILLE, 1993; EIJSINK et al., 1996). Desta forma, as
substâncias antimicrobianas produzidas pelas estirpes Pc31 e Pc32 também poderiam
pertencer à classe das bacteriocinas.
Diante do exposto, o isolamento e caracterização de estirpes produtoras de
substâncias antimicrobianas contra estirpes de importância médica, realizados no presente
trabalho, evidencia a possibilidade de se desenvolver novos agentes antimicrobianos a
partir de bactérias associadas às esponjas marinhas.

99
6 BIBLIOGRAFIA
AHMED, N.; UZAIR, B.; AYAZ, S. & AHMED, V.U. Antibacterial activity of marine bacteria from Arab ian Sea of Pakistan. Int. J. Microbiol., 4(2), 104-114, 2008. ALONSO, C.; WARNECKE, F.; AMANN, R. & PERNTHALER, J. High local and global diversity of Flavobacteria in marine plankton. Environment. Microbiol., 9(5), 1253-1266, 2007. AMANN, R.I.; LUDWIG, W. & SCHLEIFER, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation . Microbiol. Rev., 59, 143-169, 1995. ANAND, T.P.; BHAT, A.W.; SHOUCHE, Y.S.; ROY, U.; SIDDHARTH, J. & SARMA, S.P. Antimicrobial activity of marine bacteria associated with sponges from the waters off the coast of South East India. Microbiol. Res., 161, 252-262, 2006. ANGELL, S.; BENCH, B.J.; WILLIAMS, H. & WATANABE, C.M.H. Pyocyanin isolated from a marine microbial population: synergistic production between two distinct bacterial species and mode of action. Chem. Biol., 13, 1349-1359, 2006. BABA, T. & SCHNEEWIND, O. Instruments of microbial warfare: bacteriocin synthesis, toxicity and immunity . Trends Microbiol., 6(2), 66-71, 1998. BAGLEY, M.C.; DALE, J.W.; MERRITT, E.A. & XIONG, X. Thiopeptide Antibiotics. Chem. Rev., 105(2), 685-714, 2005. BANNERMAN, T.L. & PEACOCK, S.J. Staphylococcus, Micrococcus, and other catalase-positive cocci. In: Manual of Clinical Microbiology, 390-411. Edited by Murray, P.R., Barron, E.J., Jorgensen, J., Pfaller, M., Landry, M.L., Washington, ASM press, USA, 2007. BARRETT, A.J.; RAWLINGS, N.D. & WOESSNER, J.F. The Handbook of Proteolytic Enzymes, 2nd ed. Academic Press, 2003, 2368pp. BAVESTRELLO, G.; ARILLO, A.; CALCINAI, B.; CATTANEO-VIETTI, R.; CERRANO, C.; GAINO, E.; PENNA, A. & SARA, M. Parasitic diatoms inside Antartic sponges. Biol. Bull., 198, 29-33, 2000. BHAKUNI, D.S. & RAWAT, D.S. Bioactive marine natural products, 1th ed., Anamaya Publishers, New Delhi, 396p, 2005. BIL, K.; TITLYANOV, E.; BERNER, T.; FOMINA, I. & MUSCATINE, L. Some aspects of the physiology and biochemistry of Lubomirska baikalensis, a sponge from Lake Baikal containing symbiotic algae. Symbiosis, 26, 179-191, 1999.

100
BLUNT, J.W.; COPP, B.R.; MUNRO, M.H.; NORTHCOTE, P.T. & PRINSEP, M.R. Marine natural products . Nat. Prod. Rep., 22, 15-61, 2005. BLUNT, J.W.; COPP, B.R.; MUNRO, M.H.; NORTHCOTE, P.T. & PRINSEP, M.R. Marine natural products . Nat. Prod. Rep., 23, 26-78, 2006. BONMATIN, J.M.; LAPREVOTE, O. & PEYPOUX, F. Diversity among microbial cyclic lipopeptides: iturins and surfactins. Activity-structure relationships to design new bioactive agents. J. Comb. Chem., 6, 541-556, 2003. BOTTO, W.S. & GEISLER, R.L. Biological odor metabolizing compositions and methods of use. US Patent 5,821,112 issued on October 1998. BOWMAN, J.P. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar. Drugs, 5, 220-241, 2007. BRANDILEONE, M.C.; CASAGRANDE, S.T.; GUERRA, M.L.; ZANELLA, R.C.; ANDRADE, A.L. & DI FABIO, J.L. Increase in numbers of beta-lactam-resistant invasive Streptococcus pneumoniae in Brazil and the impact of conjugate vaccine coverage. J. Med. Microbiol., 55, 567-574, 2006. BRINKHOFF, T.; BACH, G.; HEIDORN, T.; LIANG, L.; SCHLINGLOFF, A. & SIMON, M. Antibiotic production by a Roseobacter clade-affiliated species from the German Wadden Sea and its antagonistic effect on indigenous isolates. Appl. Environ. Microbiol., 70, 2560-2565, 2004. BRISTIANOU, M.; PANAGOU, C.; ADAMIS, T.; RAFTOGIANNIS, M.; ANTONOPOULOU, A.; CHRISOFOS, M.; GALANI, I.; KANELLAKOPOULOU, K.; TSAGANOS, T. & GIAMARELLOS-BOURBOULIS, J. The impact of multidrug resistance on the pathogenicity of Escherichia coli: an experimental study. Int. J. Antimicrob. Agents, 31(3), 216-223, 2008. BRUNO, M.E.C. & MONTVILLE, T.J. Common mechanistic action of bacteriocins from lactic acid bacteria. Appl. Environ. Microbiol., 59(9), 3003-3010, 1993. BULTEL-PONCÉ, V.; BERGE, J.P.; DEBITUS, C.; NICOLAS, J.L. & GUYOT, M. Metabolites from the Sponge-Associated Bacterium Pseudomonas Species. Mar. Biotechnol., 1, 384-390, 1999. BULTEL-PONCÉ, V.; DEBITUS, C.; BERGE, J.P.; CERCEAU, C. & GUYOT, M. Metabolites from the sponge-associated bacterium Micrococcus luteus. J. Mar. Biotechnol., 6, 233-236, 1998. BURJA, A.M. & HILL, R.T. Microbial symbionts of the Australian Great Barrier Reef sponge, Candidaspongia flabellata. Hydrobiologia, 461, 41-47, 2001. BUTLER, M.S. & BUSS, A.D. Natural products, the future scaffolds for novel antibiotics? Biochem. Pharmacol., 71(7), 919-929, 2006.

101
CACCAMO, D.; GUGLIANDOLO, C.; STACKEBRANDT, E. & MAUGERI, T.L. Bacillus vulcano sp. nov., a novel thermophilic species isolated from a shallow marine hidrotermal vent. Int. J. Syst. Evol. Microbiol., 50, 2009-2012, 2000. CASTILLO, I.; LODEIROS, C.; NÚÑEZ, M. & CAMPOS, I. Evaluación in vitro de sustancias antibacterianas producidas por bacterias aisladas de diferentes organismos marinos. Ver. Biol. Trop., 49, 1213-1221, 2001. CEGELSKI, L.; MARSHALL, G.R.; ELDRIDGE, G.R. & HULTGREN, S.J. The biology and future prospects of antivirulence therapies. Nat. Rev. Microbiol., 6(1), 17-27, 2008. CENTERS FOR DISEASE CONTROL AND PREVENTION (CDC). Reduced susceptibility of Staphylococcus aureus to vancomycin – Japan, 1996. Morb. Mortal. Wkly. Rep., 46(27), 624-626, 1997. CENTERS FOR DISEASE CONTROL AND PREVENTION (CDC). Staphylococcus aureus resistant to vancomycin – United States, 2002. Morb. Mortal. Wkly. Rep., 51(26), 565-567, 2002. CHANDRASEKARN, S.; VENKATESH, B. & LAITHAKUMARI, D. Transfer and expression of a multiple antibiotic resistance plasmid in marine bacteria. Curr. Microbiol., 37, 347-351, 1998. CHELOSSI, E.; MILANESE, M.; MILANO, A.; PRONZATO, R. & RICCARDI, G. Characterisation and antimicrobial activity of epibiotic bacteria from Petrosia ficiformis (Porifera, Demospongiae). J. Exp. Mar. Biol. Ecol., 309, 21-33, 2004. CLARDY, J. & WALSH, C. Lessons from natural molecules. Nature, 432(7019), 829-837, 2004. CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI). Performance standards for antimicrobial susceptibility testing. Eighteenth informational supplement M100-S18. CLSI, Waynae, PA, 2008. COOPER, E.R.; SIEWICKI, T.C. & PHILLIPS, K. Preliminary risk assessment database and risk ranking of pharmaceuticals in the environment. Sci. Total. Environ., 398(1-3), 26-33, 2007. COS, P.; VLIETINCK, A.J.; BERGHE, D.V. & MAES, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. J. Ethnopharmacol., 106, 290-302, 2006. DE MIRANDA, O.P.; SILVA-CARVALHO, M.C.; RIBEIRO, A.; PORTELA, F.; CORDEIRO, R.P.; CAETANO, N.; VIDAL, C.F. & FIGUEIREDO, A.M. Emergence in Brazil of methicillin-resistant Staphylococcus aureus isolates carrying SCCmecIV that are related genetically to USA800 clone. Clin. Microbiol. Infect., 13(12), 1165-1172, 2007. DE ROSA, S.; MITOVA, M. TOMMONARO, G. Marine bacteria associated with sponge as source of cyclic peptides. Biomol. Engineer., 20, 311-316, 2003.

102
DE SOUZA, M.J.; SHANTA, N.; BHARATHI, P.L. & CHANDRAMOHAN, D. Metal and antibiotic-resistance in psychrotrophic bacteria from Antarctic marine waters. Ecotoxicol., 15(4), 379-384, 2006. DE VUYST, L. & VANDAMME, E.J. Antimicrobial potential of lactic acid bacteria. In: Bacteriocins of Lactic Acid Bacteria, 91-142. Edited by De Vuyst, L., Vandamme, E.J., Glasgow, Blackie Academic & Professional, Scotland, 1994. DEBITUS, C.; GUELLA, G.; MANCINI, I.; WAIKEDRE, J.; GUEMAS, J.P.; NICOLAS, J.L. & PIETRA, F. Quinolones from a bacterium and tyrosine metabolites from its host sponge, Suberea creba from the Coral Sea. J. Mar. Biotechnol., 6, 136-141, 1998. DEWICK, P.M. Medicinal natural products: a biosynthetic approach, 2th ed., John Wiley, Chichester, 520p, 2002. DEY, J.; ROMAN, M.A.; MEE, B.J.; FROMONT, J. & SUTTON, D.C. Bacterial diversity and antibiotic activity in temperate Australian marine sponges. Boll. Mus. Ist. Biol. Univ. Genova, 68, 263-277, 2003. DONNER, C.D.; GILL, M. & TEWIERIK, L.M. Synthesis of pyran and pyranone natural products. Molecules, 9, 498-512, 2004. DRAGO, L. Epidemiology and mechanisms of resistance: clinical and environmental impact. Infez. Med., 2, 6-12, 2007. EIJSINK, V.G.H.; BRURBERG, M.B.; MIDDELLHOVEN, P.H. & NES, I.F. Induction of bacteriocin production in Lactobacillus sake by a secreted peptide. J. Bacteriol., 178(8), 2232-2237, 1996. ENTICKNAP, J.J.; KELLY, M.; PERAUD, O. & HILL, R.T. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Appl. Environment. Microbiol., 72(5), 3724-3732, 2006. FALAGAS, M.E. & KARAGEORGOPOULOS, D.E. Clinical microbiology and infectious diseases – 18th European Congress. Drug resistance among Gram-negative and Gram-positive bacteria. I. Drugs, 11(6), 409-411, 2008. FALAGAS, M.E. Pandrug-resistant Gram-negative bacteria: the dawn of the post-antibiotic era? Int. J. Antimicrob. Agents, 29(6), 630-636, 2007. FIESELER, L.; HORN, M.; WAGNER, M. & HENTSCHEL, U. Discovery of the novel candidate phylum Poribacteria in marine sponges. Appl. Environ. Microbiol., 70, 3724-3732, 2004. FRIEDRICH, A.B.; FISCHER, I.; PROKSCH, P.; HACKER, J. & HENTSCHEL, U. Temporal variation of the microbial community associated with the Mediterranean sponge Aplysina aerophoba. FEMS Microbiol. Ecol., 38, 105-113, 2001.

103
FRIEDRICH, A.B.; MERKERT, H.; FENDERT, T.; HACKER, J.; PROKSCH, P. & HENTSCHEL, U. Microbial diversity in the marine sponge Aplysina cavernicola (formerly Verongia cavernicola) analyzed by fluorescence in situ hybridization (FISH) . Mar. Biol., 134, 461-470, 1999. FUKUNAGA, Y.; KURAHASHI, M.; TANAKA, K.; YANAGI, K. ; YOKOTA, A. & HARAYAMA, S. Pseudovibrio ascidiaceicola sp. nov., isolated from ascidians (sea squirts). Int. J. Syst. Evol. Microbiol., 56, 343-347, 2006. FULLER, A.T.; MELLOWS, G.; WOOLFORD, M.; BANKS, G.T.; BARROW, K.D. & CHAIN, E.B. Pseudomonic acid: an antibiotic produced by Pseudomonas fluorescens. Nature, 234, 416-417, 1971. GAINO, E.; SCISCIOLI, M.; LEPORE, E.; REBORA, M. & CORRIERO, G. Association of the sponge Tethya orphei (Porifera, Demospongiae) with filamentous cyanobacteria. Invertebr. Biol., 125, 281-287, 2006. GARSON, M.J.; FLOWERS, A.E.; WEBB, R.I.; CHARAN, R.D. & MCCAFFREY, E.J. A sponge/dinoflagellate association in the haplosclerid sponge Haliclona sp.: cellular origin of cytotoxic alkaloids by Percoll density gradient centrifugation. Cell Tissue Res., 293, 365-373, 1998. GERNERT, C.; GLOCKNER, F.O.; KROHNE, G. & HENTSCHEL, U. Microbial diversity of the freshwater sponge Spongilla lacustris. Microb. Ecol., 50, 206-212, 2005. GIAMBIAGI-DEMARVAL, M.; MAFRA, M.A.; PENIDO, E.G.C. & BASTOS, M.C.F. Distinct groups of plasmids correlated with bacteriocin production in Staphylococcus aureus. J. Gen. Microbiol., 136, 1591-1599, 1990. GIDDENS, S.R. & BEAN, D.C. Investigations into the in vitro antimicrobial activity and mode of action of the phenazine antibiotic D-alanylgriseoluteic acid. Int. J. Antimicrob. Agents, 29, 93-97, 2007. GRAEBER, I.; KAESLER, I.; DIECKMANN, R.; VON DÖHREN, H. & SZEWZYK, U. Sponge associated bacteria from boreal sponges. Boll. Mus. Ist. Biol. Univ. Genova, 68, 357-363, 2003. GUZMAN, A.A.S.; PREDICALA, R.Z.; BERNARDO, E.B.; NEILAN, B.A.; ELARDO, S.P.; MANGALINDAN, G.C.; TASDEMIR, D.; IRELAND, C.M.; BARRAQUIO, W.L. & CONCEPCION, G.P. Pseudovibrio denitrificans strain Z143-1, a heptylprodigiosin-producing bacterium isolated from a Philippine tunicate. FEMS Microbiol. Lett., 277(2), 188-196, 2007. HADJU, E. & LOPES, D.A. Checklist of Brazilian deep-sea sponges. In: Porifera Research - Biodiversity, Innovation and Sustainability, 353-359. Edited by Custódio, M.R., Lôbo-Hajdu, G., Hajdu, E., Muricy, G., Rio de Janeiro, Museu Nacional - Série Livros, Brasil, 2007. HAJDU, E. & DESQUEYROUX-FAUNDEZ, R. A synopsis of South American Mycale (Mycale) (Poecilosclerida, Demospongiae), with the description of three new species and a cladistic analysis of Mycalidae. Revue Suis. Zool., 101(3), 563-600, 1994.

104
HAJDU, E. & VAN SOEST, R.W.M. A revision of Atlantic Asteropus Sollas, 1888 (Demospongiae), including a description of three new species, and with a review of the family Coppatiidae Topsent, 1898. Bijdragen tot de Dierkunde, 62, 3-19, 1992. HENTSCHEL, U.; FIESELER, L.; WEHRL, M.; GERNERT, C.; STEINERT, M.; HACKER, J. & HORN, M. Microbial diversity of marine sponges. Prog. Mol. Subcell. Biol., 37, 59-88, 2003. HENTSCHEL, U.; HOPKE, J.; HORN, M.; FRIEDRICH, A.B.; WAGNER, M.; HACKER, J. & MOORE, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol., 68, 4431-4440, 2002. HENTSCHEL, U.; SCHMID, M.; WAGNER, M.; FIESELER, L.; GERNERT, C. & HACKER, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol., 35, 305-312, 2001. HEWAVITHARANA, A.K.; SHAW, P.N.; KIM, T.K. & FUERST, J.A. Screening of rifamycin producing marine sponge bacteria by LC–MS–MS. J. Chromatogr. B, 852, 362-366, 2007. HILL, M.; HILL, A.; LOPEZ, N. & HARRIOTT, O. Sponge-specific bacterial symbionts in the Caribbean sponge, Chondrilla nucula (Demospongiae, Chondrosida). Mar. Biol., 148, 1221-1230, 2006. HILL, R.T. Microbes from marine sponges: a treasure trove of biodiversity for natural products discovery. In: Microbial diversity and bioprospecting, 177-190. Edited by Bull, A.T., Washington DC, ASM Press, EUA, 2004. HIRSCH, R.; TERNES, T.; HABERER, K. & KRATZ, K.L. Occurrence of antibiotics in the aquatic environment. Sci. Total Environ., 225, 109-118, 1999. HÖLLER, U.; WRIGHT, A.D.; MATTHEE, G.F.; KÖNIG, G.M.; DRAEGER, S.; AUST, H.-J. & SCHULZ, B. Fungi from marine sponges: diversity, biological activity and secondary metabolites. Mycol. Res., 104, 1354-1365, 2000. HOLMES, B. & BLANCH, H. Genus-specific associations of marine sponges with group I crenarchaeotes. Mar. Biol., 150, 759-772, 2006. HOOPER, J.N.A. & VAN SOEST, R.W.M. Systema Porifera: a guide to the classification of sponges, 1th ed., Plenum Publishers, New York, 1810p, 2002. HOPWOOD, D.A. Genetic contributions to understanding polyketide synthases. Chem. Rev., 97, 2485-2497, 1997. HOSOYA, S. & YOKOTA, A. Pseudovibrio japonicus sp. nov., isolated from coastal seawater in Japan. Int. J. Syst. Evol. Microbiol., 57, 1952-1955, 2007.

105
HSIEH, F.-C.; LIN, T.-C.; MENG, M. & KAO, S.-S. Comparing methods for identifying Bacillus strains capable of producing the antifungal lipopeptide iturin A . Curr. Microbiol., 56(1), 1-5, 2008. HUGHES, J. & MELLOWS, G. Interaction of pseudomonic acid A with Escherichia coli B isoleucyl-tRNA synthetase. Biochem. J., 191(1), 209-219, 1980. HUGHES, J.; MELLOWS, G. & SOUGHTON, S. How does Pseudomonas fluorescens, the producing organism of the antibiotic pseudomonic acid A, avoid suicide? FEBS Lett., 122(2), 322-324, 1980. IMHOFF, J.F. & STÖHR, R. Sponge-associated bacteria: general overview and special aspects of the diversity of bacteria associated with Halichondria panicea. In: Marine Molecular Biotechnology, 35-57. Edited by Müller, W.E.G., New York, Springer, USA, 2003. ISBISTER, G.K. & HOOPER, J.N. Clinical effects of stings by sponges of the genus Tedania and a review of sponge stings worldwide. Toxicon, 46(7), 782-785, 2005. ISNANSETYO, A. & KAMEI, Y. Pseudoalteromonas phenolica sp. nov., a novel marine bacterium that produces phenolic anti-methicillin-resistant Staphylococcus aureus substances. Int. J. Syst. Evol. Microbiol. 53(2), 583-588, 2003. IVANOVA, E.P.; VYSOTSKII, M.V.; SVETASHEV, V.I.; NEDASHKOVSKAYA, O.I.; GORSHKOVA, N.M.; MIKHAILOV, V.V.; YUMOTO, N.; SHIGERI, Y.; TAGICHI, T. & YOSHIKAWA, S. Characterization of Bacillus strains of marine origin. Internatl. Microbiol., 2, 267-271, 1999. JAYATILAKE, G.S.; THORNTON, M.P.; LEONARD, A.C.; GRIMWADE, J.E. & BAKER, B.J. Metabolites from an Antarctic sponge-associated bacterium, Pseudomonas aeruginosa. J. Nat. Prod., 59, 293-296, 1996. JOHN, J.F. & HARVIN, A.M. History and evolution of antibiotic resistance in coagulase-negative staphylococci: susceptibility profiles of new anti-staphylococcal agents. Ther. Clin. Risk Manag., 3(6), 1143-1152, 2007. KHAN, N.H.; ISHII, Y.; KINO, N.K.; ESAKI, H.; NISHINO, T.; NISHIMURA, M. & KOGURE, K. Isolation of Pseudomonas aeruginosa from open ocean and comparison with freshwater, clinical and animal isolates. Microbial Ecol., 53, 173-186, 2007. KIM, T.K. & FUERST, J.A. Diversity of polyketide synthase genes from bacteria associated with the marine sponge Pseudoceratina clavata: culture-dependent and culture-independent approaches. Environ. Microbiol., 8(8), 1460-1470, 2006. KLAENHAMMER, T.R. Bacteriocins of lactic acid bacteria. Biochemie, 70, 337-349, 1988.

106
KOLPIN, D.W.; FURLONG, E.T.; MEYER, M.T.; THURMAN, E.M.; ZAUGG, S.D.; BARBER, L.B. & BUXTON, H.T. Pharmaceuticals, hormones, and other organic wastewater contaminants in U.S. streams, 1999-2000: a national reconnaissance. Environ. Sci. Technol., 36(6), 1202-1211, 2002. KUTI, J.L.; KIFFER, C.R.; MENDES, C.M. & NICOLAU, D.P. Pharmacodynamic comparison of linezolid, teicoplanin and vancomycin against clinical isolates of Staphylococcus aureus and coagulase-negative staphylococci collected from hospitals in Brazil. Clin. Microbiol. Infect., 14(2), 116-123, 2008. LAFI, F.F.; GARSON, M.J. & FUERST, J.A. Culturable bacterial symbionts isolated from two distinct sponge species (Pseudoceratina clavata and Rhabdastrella globostellata) from the Great Barrier Reef display similar phylogenetic diversity. Microb. Ecol., 50, 213-220, 2005. LANNA, E.; ROSSI, A.L.; CAVALCANTES, F.; HAJDU, E. & KLAUTAU, M. Calcareous sponges from São Paulo state, Brazil (Porifera: Calcarea: Calcinea) with the description of two new species. J. Mar. Biol. As. United Kingdom, 87, 1553-1561, 2007. LAPORT, M.S.; SANTOS, O.C.S. & MURICY, G. Marine sponges: potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol., in press. LIU, H.; HE, Y.; JIANG, H.; PENG, H.; HUANG, X.; ZHANG, X.; THOMASHOW, L.S. & XU, Y. Characterization of a phenazine-producing strain Pseudomonas chlororaphis GP72 with broad-spectrum antifungal activity from green pepper rhizosphere. Microbiol., 54, 302-306, 2007. LO GIUDICE, A.; BRILLI, M.; BRUNI, V.; DE DOMENICO, M.; FANI, N. & MICHAUD, L. Bacterium-bacterium inhibitory interactions among psychrotrophic bacteria isolated from Antarctic seawater (Terra Nova Bay, Ross Sea). FEMS Microbiol. Ecol., 60, 383-396, 2007. LO GIUDICE, A.; BRUNI, V. & MICHAUD, L. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. J. Basic. Microbiol., 47, 496-505, 2007. LOPEZ, J.V.; MCCARTHY, P.J.; JANDA, K.E.; WILLOUGHBY, R. & POMPONI, S.A. Molecular techniques reveal wide phyletic diversity of heterotrophic microbes associated with Discodermia spp. (Porifera: Demospongiae). Mem. Qld. Mus., 44, 329-341, 1999. LUZHETSKYY, A.; PELZER, S. & BECHTHOLD, A. The future of natural products as a source of new antibiotics. Curr. Opin. Investig. Drugs., 8(8), 608-613, 2007. MACFADDIN, J.F. Biochemical tests for identification of medical bacteria, 3th ed., Lippincott Williams & Wilkins, Philadelphia, 912p, 2000. MALDONADO, M.; CORTADELLAS, N.; TRILLAS, M.I. & RUTZLER, K. Endosymbiotic yeast maternally transmitted in a marine sponge. Biol. Bull., 209, 94-106, 2005.

107
MARAHIEL, M.A.; NAKANO, M.M. & ZUBER, P. Regulation of peptide antibiotic production in Bacillus. Mol. Microbiol., 7, 631-636, 1993. MARGOT, H.; ACEBAL, C.; TORIL, E.; AMILS, R. & FERNANDEZ PUENTES, J.L. Consistent association of crenarchaeal archaea with sponges of the genus Axinella. Mar. Biol., 140, 739-745, 2002. MARTIN, M.F. & LIRAS, P. Organization and expression of genes involved in the biosynthesis of antibiotics and other secondary metabolites. Annu. Rev. Microbiol., 43, 173-206, 1989. MARTINS, A. & CUNHA, M.L. Methicillin resistance in Staphylococcus aureus and coagulase-negative staphylococci: epidemiological and molecular aspects. Microbiol. Immunol., 51(9), 787-795, 2007. MCDANIEL, R.; WELCH, M. & HUTCHINSON, C.R. Genetic approaches to polyketide antibiotics. Chem. Ver., 105, 543-558, 2005. MEARNS-SPRAGG, M.; BREGU, M.; BOYD, K.G.; & BURGESS, J.G. Cross-species induction and enhancement of antimicrobial activity produced by epibiotic bacteria from marine algae and invertebrates, after exposure to terrestrial bacteria. Lett. Appl. Microbiol., 27, 142-146, 1998. MITOVA, M.; POPOV, S. & DE ROSA, S. Cyclic peptides from a Ruegeria strain of bacteria associated with the sponge Suberites domuncula. J. Nat. Prod., 67, 1178-1181, 2004. MOFFITT, M.C. & NEILAN, B.A. The expansion of mechanistic and organismic diversity associated with non-ribosomal peptides. FEMS Microbiol. Lett., 191, 159-167, 2000. MONTALVO, N.F.; MOHAMED, N.M.; ENTICKNAP, J.J. & HILL, R.T. Novel actinobacteria from marine sponges. Antonie Van Leeuwenhoek, 87(1), 29-36, 2005. MOORE, E.R.B.; MAU, M.; ARNSCHEIDT, A.; BÖTTGER, E.C.; HUTSON, R.A.; COLLINS, M.D.; VAN DE PEER, Y.; DE WACHTER, R. & TIMMIS, K.T. The determination and comparison of the 16S rDNA gene sequence of species of the genus Pseudomonas (sensu stricto) and estimation of the natural generic relationships. Syst. Appl. Microbiol., 19, 478-492, 1996. MOTHES, B.; HAJDU, E. & VAN SOEST, R.W.M. Tedania brasiliensis new species (Demospongiae, Poecilosclerida, Tedaniidae) from Brazil, with some remarks about the genus Tedania in the Tropical Southwestern Atlantic. Bull. Mar. Sci., 66(1), 1-11, 2000. MOULIN, G.; CAVALIÉ, P.; PELLANNE, I.; CHEVANCE, A.; LAVAL, A.; MILLEMANN, Y.; COLIN, P. & CHAUVIN, C. A comparison of antimicrobial usage in human and veterinary medicine in France from 1999 to 2005. J. Antimicrob. Chemother., Oxford, 2008. Disponível em: <http://jac.oxfordjournals.org/cgi/content/ abstract/dkn213>. Acesso em: 08 julho 2008. MUDRYK, Z. & SKÓRCZEWSKI, P. Antibiotic resistance in marine neustonic and planktonic bacteria isolated from the Gdansk Deep. Oceanol., 40, 125-136, 1998.

108
MÜLLER, W.E.G. & MÜLLER, I.M. Origin of the metazoan immune system: identification of the molecules and their functions in sponges. Integr. Comp. Biol., 43, 281-292, 2003. MURICY, G. & MORAES, F. Marine sponges of Pernambuco State, NE Brazil. Rev. Brasil. Oceanogr., 46(2), 213-217, 1999. MURICY, G. & SILVA, O.C. Esponjas marinhas do Estado do Rio de janeiro: Um recurso renovável inexplorado. In: Ecologia dos ambientes costeiros do Estado do Rio de Janeiro, 155-178. Edited by Silva, S.H.G., Lavrado, H.P., Rio de Janeiro, PPGE-UFRJ, Brasil, 1999. MUSCHOLL-SILBERHORN, A.; THIEL, V. & IMHOFF, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microbial. Ecol., 55(1), 94-106, 2008. NAGAI, K.; KAMIGIRI, K.; ARAO, N.; SUZUMURA, K.; KAWANO, Y.; YAMAOKA, M.; ZHANG, H.; WATANABE, M. & SUZUKI, K. YM-266183 and YM-266184, novel thiopeptide antibiotics produced by Bacillus cereus isolated from a marine sponge. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological properties. J. Antibiotics, 56, 123-128, 2003. NASCIMENTO, J.S.; ABRANTES, J.; GIAMBIAGI-DEMARVAL, M. & BASTOS, M.C.F. Growth conditions required for bacteriocin production by strains of Staphylococcus aureus. World J. Microbiol. Biotechnol., 20, 941-947, 2004. NEEDHAM, J.; KELLY, M.T.; ISHIGE, M. & ANDERSEN, R.J. Andrimid and moiramides A-C, metabolites produced in culture by a marine isolate of the bacterium Pseudomonas fluorescens: structure elucidation and biosynthesis of outstanding interest. J. Org. Chem., 59, 2058-2063, 1994. OCLARIT, J.M.; OKADA, H.; OHTA, S.; KAMINURA, K.; YAMAOKA, Y.; IIZUKA, T.; MIYASHIRO, S. & IKEGAMI, S. Anti- bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios, 78, 7-16, 1994. OKEKE, I.N.; LAXMANINARAYAN, R.; BHUTTA, Z.A.; DUSE, A.G.; JENKINS, P.; O’BRIEN, T.F.; PABLOS-MENDEZ, A. & KLUGMAN, K.P. Antimicrobial resistance in developing countries. Part 1: recent trends and current status. Lancet Infect. Dis., 5, 481-493, 2005. OLIVEIRA, C.J.; CARVALHO, L.F.; FERNANDES, S.A.; TAVECHIO, A.T.; MENEZES, C.C. & DOMINGUES, F.J. Antimicrobial resistance of Salmonella serotypes isolated from slaughter-age pigs and environmental samples. Microb. Drug Resist., 8(4), 407-411, 2002. OLIVEIRA, S.C. & VON SPERLING, M. Análise da confiabilidade de estações de tratamento de esgotos. Eng. Sanit. Ambient., 12(4), 389-398, 2007. PABEL, C.T.; VATER, J.; WILDE, C.; FRANKE, P.; HOFEMEISTER, J.; ADLER, B.; BRINGMANN, G.; HACKER, J. & HENTSCHEL, U. Antimicrobial activities and matrix-assisted laser desorption/ionization mass spectrometry of Bacillus isolates from the marine sponge Aplysina aerophoba. Mar. Biotechnol., 5, 424-434, 2003.

109
PAUL, V.J.; ARTHUR, K.E.; RITSON-WILLIAMS, R.; ROSS, C. & SHARP, K. Chemical defenses: from compounds to communities. Biol. Bull., 213, 226-251, 2007. PIEL, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep., 21, 519-538, 2004. PILE, A.J.; PATTERSON, M.R. & WITMAN, J.D. In situ grazing on plankton <10µm by the boreal sponge Mycale lingua. Mar. Ecol. Prog. Ser., 141, 95-102, 1996. PINHEIRO, U.S. & HAJDU, E. Shallow-water Aplysina Nardo (Aplysinidae, Verongida, Demospongiae) from the São Sebastião Channel and its environs (Tropical southwestern Atlantic), with the description of new species and literature review of other Brazilian records of the genus. Revta. Bras. Zool., 18(1), 143-160, 2001. PITCHER, D.G.; SAUNDERS, N.A. & OWEN, R.J. Rapid extration of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol., 8, 151-156, 1989. POULAKOU, G. & GIAMARELLOU, H. Oritavancin: a new promising agent in the treatment of infections due to Gram-positive pathogens. Expert. Opin. Investig. Drugs, 17(2), 225-243, 2008. PRESTON, C.M.; WU, K.Y.; MOLINSKI, T.F. & DELONG, E.F. A psychrophilic crenarchaeon inhabits a marine sponge: Crenarchaeum symbiosum gen. nov., sp. nov. Proc. Natl. Acad. Sci. USA, 93, 6241-6246, 1996. REISWIG, H.M. Particle feeding in natural populations of three marine demosponges. Biol. Bull., 141, 568-591, 1971. RHEE, K.-H. Cyclic dipeptides exhibit synergistic, broad spectrum antimicrobial effects and have anti-mutagenic properties. Int. J. Antimicrob. Agents, 24, 423-427, 2004. RIBEIRO, A.; CORONADO, A.Z.; SILVA-CARVALHO, M.C.; FERREIRA-CARVALHO, B.T.; DIAS, C.; ROZENBAUM, R.; DEL PELOSO, P.F.; DA COSTA FERREIRA LEITE, C.; TEIXEIRA, L.A. & FIGUEIREDO, A.M. Detection and characterization of international community-acquired infections by methicillin-resistant Staphylococcus aureus clones in Rio de Janeiro and Porto Alegre cities causing both community- and hospital-associated diseases. Diagn. Microbiol. Infect. Dis., 59(3), 339-345, 2007. RICE, L.B. Emerging issues in the management of infections caused by multidrug-resistant gram-negative bacteria. Cleve Clin. J. Med., 4, 12-20, 2007. ROMANENKO, L.A.; UCHINO, M.; FALSEN, E.; FROLOVA, G.M.; ZHUKOVA, N.V. & MIKHAILOV, V.V. Pseudomonas pachastrellae sp. nov., isolated from a marine sponge. Int. J. Syst. Evol. Microbiol., 55, 919-924, 2005. ROMANENKO, L.A.; UCHINO, M.; KALINOVSKAYA, N.I. & M IKHAILOV, V.V. Isolation, phylogenetic analysis and screening of marine mollusc-associated bacteria for antimicrobial, hemolytic and surface activities. Microbiol. Res., doi:10.1016/j.micres.2006.10.001, 2006.

110
RUPPERT, E.E. & BARNES, R.D. Invertebrate zoology, 6th ed., Harcourt Brace College Publishers, Texas, 928p, 1994. SAMBROOK, J. & RUSSELL, D.W. Molecular cloning: a laboratory manual, 3th ed., Cold Spring Harbor Laboratory Press, New York, 999p, 2001. SANTAVY, D.L. & COLWELL, R.R. Comparison of bacterial communities associated with the Caribbean sclerosponge Ceratoporella nicholsoni and ambient seawater. Mar. Ecol. Prog. Ser., 67, 73-82, 1990. SARMENTO, F. & CORREIA, M.D. Description of ecological and external morphological parameters of Porifera at the ponta verde coral reef, Alagoas, Brazil. Rev. Bras. Zoociências, 4(2), 203-214, 2002. SCHIRMER, A.; GADKARI, R.; REEVES, C.D.; IBRAHIM, F.; DELONG, E.F. & HUTCHINSON, C.R. Metagenomic analysis reveals diverse polyketide synthase gene clusters in microorganisms associated with the marine sponge Discodermia dissoluta. Appl. Environ. Microbiol., 71, 4840-4849, 2005. SCHMITZ, F.J.; VANDERAH, D.J.; HOLLENBEAK, K.H.; ENWALL, C.E.L.; GOPICHAND, Y.; SENGUPTA, P.K.; HOSSAIN, M.B. & VANDERHELM, D. Metabolites from the marine sponge Tedania ignis - a new atisanediol and several known diketopiperazines. J. Org. Chem., 48, 3941, 1983. SCHÖNBERG, C.H.L. & LOH, W.K.W. Molecular identity of the unique symbiotic dinoflagellates found in the bioeroding demosponge Cliona orientalis. Mar. Ecol. Prog. Ser., 299, 157-166, 2005. SCHUPP, T.; TOUPET, C.; ENGEL, N. & GOFF, S. Cloning and sequence analysis of the putative rifamycin polyketide synthase gene cluster from Amycolatopsis mediterranei. FEMS Microbiol. Lett., 159, 201-207, 1998. SELVIN, J.S.; JOSEPH, S.; ASHA, K.R.T.; MANJUSHA, W.A.; SANGEETHA, V.S.; JAYASEEMA, D.M.; ANTONY, M.C. & VINITHA, A.J.D. Antibacterial potential of antagonistic Streptomyces sp. isolated from marine sponge Dendrilla nigra. FEMS Microbiol. Ecol., 50, 117-122, 2004. SHARP, K.H.; EAM, B.; FAULKNER, D.J. & HAYGOOD, M.G. Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl. Environm. Microbiol., 73(2), 622-629, 2007. SHIEH, W.Y.; LIN, Y.T. & JEAN, W.D. Pseudovibrio denitrificans gen. nov., sp. nov., a marine, facultatively anaerobic, fermentative bacterium capable of denitrification . Int. J. Syst. Evol. Microbiol., 54, 2307-2312, 2004. SIEBER, S.A. & MARAHIEL, M.A. Molecular mechanisms underlying nonribosomal peptide synthesis: approaches to new antibiotics. Chem. Rev., 105, 715-738, 2005. SIMPSON, T.L. The cell biology of sponges, 1th ed., Springer-Verlag, New York, 681p, 1984.

111
SINGH, M.P.; KONG, F.; JANSO, J.E.; ARIAS, D.A.; SUAREZ, P.A.; BERNAN, V.S.; PETERSEN, P.J.; WEISS, W.J.; CARTER, G. & GREENSTEIN, M. Novel α-pyrones produced by a marine Pseudomonas sp. F92S91: taxonomy and biological activities. J. Antibiotics, 56, 1033-1044, 2003. STACKEBRANDT, E. & GOEBEL, B.M. Taxonomic note: a place for DNA-DNA reassociation and rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol., 44, 846-849, 1994. STARLIPER, C. & MORRISON, P. Bacterial pathogen contagion studies among freshwater bivalves and salmonid fishes. J. Shell. Res., 19, 251-258, 2000. STAUNTON, J. & WEISSMAN, K.J. Polyketide biosynthesis: a millennium review. Nat. Prod. Rep., 18, 380-416, 2001. STIERLE, A.C.; CARDELLINA, J.H. & SINGLETON, F.L. A marine Micrococcus produces metabolites ascribed to the sponge Tedania ignis. Experientia, 44, 1021, 1988. SUZUMURA, K.; YOKOI, T.; FUNATSU, M.; NAGAI, K.; TANAKA, K.; ZHANG, H. & SUZUKI, K. YM-266183 and YM-266184, novel thiopeptide antibiotics produced by Bacillus cereus isolated from a marine sponge II. Structure elucidation. J. Antibiotics, 56, 129-134, 2003. TAYLOR, M.W.; RADAX, R.; STEGER, D. & WAGNER, M. Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev., 71(2), 295-347, 2007. TAYLOR, M.W.; SCHUPP, P.J.; DAHLLOF, I.; KJELLEBERG, S. & STEINBERG, P.D. Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity . Environ. Microbiol., 6, 121-130, 2004. TAYLOR, M.W.; SCHUPP, P.J.; DE NYS, R.; KJELLEBERG, S. & STEINBERG, P.D. Biogeography of bacteria associated with the marine sponge Cymbastela concentrica. Environ. Microbiol., 7, 419-433, 2005. TENDENCIA, E.A. & DE LA PENA, L.D. Antibiotic resistance of bacteria from shrimp ponds. Aquaculture, 195, 193-204, 2001. THAKUR, A.N.; THAKUR, N.L.; INDAP, M.M.; PANDIT, R.A.; DATAR, V.V. & MÜLLER, W.E.G. Antiangiogenic, antimicrobial, and cytotoxic potential of sponge-associated bacteria. Mar. Biotechnol., 7, 245-252, 2005. THAVASI, R.; APARNADEVI, K.; JAYALAKSHMI, S. & BALA SUBRAMANIAN, T. Plasmid mediated antibiotic resistance in marine bacteria. J. Environ. Biol., 28(3), 617-621, 2007. THIEL, V. & IMHOFF, J.F. Phylogenetic identification of bacteria with antimicrobial activities isolated from Mediterranean sponges. Biomol. Engineer., 20, 421-423, 2003.

112
THOMPSON, F.L.; IIDA, T. & SWINGS, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev., 68(3), 403-431, 2004. THOMS, C.; HORN, M.; WAGNER, M.; HENTSCHEL, U. & PROKSCH, P. Monitoring microbial diversity and natural product profiles of the sponge Aplysina cavernicola following transplantation. Mar. Biol., 142(4), 685-692, 2003. THORNTON, M.P. Characterization of secondary metabolite bioactivity and production in bacteria associated with the Antarctic marine sponge Isodictya setifera, MS Thesis, Melbourne, 51p, 1995. VACELET, J. & DONADEY, C. Electron microscope study of the association between some sponges and bacteria. J. Exp. Mar. Biol. Ecol., 30, 301-314, 1977. VACELET, J. & GALLISSIAN, M.F. Virus-like particles in cells of the sponge Verongia cavernicola (Demospongiae, Dictyoceratida) and accompanying tissue changes. J. Invertebr. Pathol., 31, 246-254, 1978. VACELET, J. Etude en microscopie electronique de l’association entre bacteries et spongiaires du genre Verongia (Dictyoceratida). J. Microsc. Biol. Cell., 23, 271-288, 1975. VILELA, M.A.; SOUZA, S.L.; PALAZZO, I.C.; FERREIRA, J.C.; MORAIS, M.A.; DARINI, A.L. & MORAIS, M.M. Identification and molecular characterization of Van-A-type vancomycin-resistant Enterococcus faecalis in Northeast of Brazil. Mem. Inst. Oswaldo Cruz, 101, 715-719, 2006. VINING, L.C. Functions of secondary metabolites. Annu. Rev. Microbiol., 44, 395-427, 1990. VOGEL, S. Current-induced flow through living sponges in nature. Proc. Natl. Acad. Sci. USA, 74, 2069-2071, 1977. WAGNER, M. & HORN, M. The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol., 17, 241-249, 2006. WALSH, C.T. Polyketide and nonribosomal peptide antibiotics: modularity and versatility . Science, 303, 1805-1810, 2004. WEBSTER, N.S. & HILL, R.T. The culturable microbial community of the Great Barrier Reef sponge Rhopaloeides odorabile is dominated by an α-proteobacterium. Mar. Biol., 138, 843-851, 2001. WEHRL, M.; STEINERT, M. & HENTSCHEL, U. Bacterial uptake by the marine sponge Aplysina aerophoba. Microb. Ecol., 53, 355-365, 2007. WEISBURG, W.G.; BARNS, S.M.; PELLETIER, D.A. & LANE, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol., 173(2), 697-703, 1991.

113
WELKER, M. & VON DÖHREN, H. Cyanobacterial peptide – nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev., 30(4), 530-563, 2006. WILKINSON, C.R. Immunological evidence for the Precambrian origin of bacterial symbioses in marine sponges. Proc. R. Soc. Lond. B, 220, 509-517, 1984. WILKINSON, C.R. Interocean differences in size and nutrition of coral reef sponge populations. Science, 236, 1654-1657, 1987. WILKINSON, C.R. Microbial associations in sponges. I. Ecology, physiology and microbial populations of coral reef sponges. Mar. Biol., 49, 161-167, 1978a. WILKINSON, C.R. Microbial associations in sponges. II. Numerical analysis of sponge and water bacterial populations. Mar. Biol., 49, 169-176, 1978b. WILKINSON, C.R. Microbial associations in sponges. III. Ultrastructure of the in situ associations in coral reef sponges. Mar. Biol., 49, 177-185, 1978c. WILKINSON, C.R. Net primary productivity in coral reef sponges. Science, 219, 410-412, 1983. WILKINSON, C.R. Nutrient translocation from symbiotic cyanobacteria to coral reef sponges. In: Biologie des spongiaires, 373-380. Edited by Levi, C., and Boury-Esnault, N., Paris, Colloques Internationaux du Centre National de la Recherche Scientifique, France, 1979. YAHEL, G.; SHARP, J.H.; MARIE, D.; HASE, C. & GENIN, A. In situ feeding and element removal in the symbiont-bearing sponge Theonella swinhoei: bulk DOC is the major source for carbon. Limnol. Oceanogr., 48, 141-149, 2003. YANAGISAWA, T. & KAWAKAMI, M. How does Pseudomonas fluorescens avoid suicide from its antibiotic pseudomonic acid? J. Biol. Chem., 278(28), 25887-25894, 2003. ZHENG, G. & SLAVIK, M.F. Isolation, partial purification and characterization of a bacteriocin produced by a newly isolated Bacillus subtilis strain. Lett. Appl. Microbiol., 28, 263-367, 1999. ZHENG, Z.; ZENG, W.; HUANG, Y.; YANG, Z.; LI, J.; CAI, H. & SU, W. Detection of antitumor and antimicrobial activities in marine organism associated actinomycetes isolated from the Taiwan Strait, China. FEMS Microbiol. Lett., 188(1), 87-91, 2000.

114
SPECIAL ISSUE OF CURRENT PHARMACEUTICAL BIOTECHNOLO GY
EDITORIAL
Maria do Carmo de Freire Bastos Departamento de Microbiologia Geral, Instituto de Microbiologia Prof. Paulo de Góes, Universidade Federal
do Rio de Janeiro, 21941-590, Rio de Janeiro, Brasil
The spread of microbial resistance leads to a growing demand for novel antimicrobial drugs.
However, despite the significant efforts in academia and the pharmaceutical industry, no genuinely new class
of antimicrobial compounds has reached the market in the last years. However, a large variety of substances
with potential antimicrobial activity is widely distributed in nature being produced by both prokaryotes and
eukaryotes. Therefore, in this special issue of Current Pharmaceutical Biotechnology, we attempt to provide
the reader with an overview on antimicrobial substances produced by bacteria, marine sponges, and tropical
plants. Concerning prokaryotes, the reviews will focus on bacteriocins, antimicrobial peptides produced by
bacteria.
Bierbaum and co-workers focus their review on the current developments in the lantibiotic (class I
bacteriocins) field and on recent findings concerning mode of action, biosynthesis and engineering of
lantibiotics produced by Gram-positive bacteria, while the review written by Nissen-Meyer and co-workers
focuses on the structure and mode of action of class II bacteriocins. Structure-function analysis of bacteriocins
is particularly relevant for illuminating how these potent bactericidal agents function at a molecular level.
Such knowledge is fundamental when considering the rational design of new bacteriocin variants with
improved properties that make them especially useful for medical and biotechnological applications.
Bastos and co-workers describe the relevant features of staphylococcins, bacteriocins produced by
staphylococci, discussing examples of their potential biotechnological applications mainly as antibacterial
agents.
Diep and co-workers discuss some well-characterised quorum sensing networks involved in
regulation of bacteriocin production in lactobacilli, with special focus on the use of the regulatory components
in gene expression and on lactobacilli as potential delivery vehicle for therapeutic and vaccine purposes.

115
The review of Lagos and co-workers is focused on microcin E492 features, a bacteriocin that,
besides an antimicrobial activity, also displays a cytotoxic effect on tumor cell lines involving apoptosis
induction. This trait makes this bacteriocin also suitable for cancer treatment.
Laport and co-workers describe an overwhelming number of bioactive substances that have been
discovered in sponges and their associated microorganisms. Sponges are considered the most prolific marine
producers of novel substances exhibiting antimicrobial activity with a variety of biotechnologically relevant
properties.
Finally, the main focus of the review written by Alviano and Alviano are the antimicrobial and
antioxidant properties of bioactive compounds obtained from crude plant extracts and essential oils of
Brazilian plants used empirically in folk medicine to treat various infectious disorders.
From the data presented in these reviews, we can realize that considerable research to date has led to
the identification of a growing number of antimicrobial substances that may provide valuable addition to the
current range of antimicrobial compounds available for animal and human medicine. However, we should
keep in mind that, although substantial progress has been made in identifying novel drug leads from a variety
of living organisms, great efforts are still need to advance to their clinical applications. Therefore, I hope that
this special issue of Current Pharmaceutical Biotechnology can be useful to those readers already working on
antimicrobial substances, stimulating discussion and further studies, but principally serving to attract new
researchers from other fields interested in exploitation of the biotechnological applications of these
substances.

116
Marine sponges: potential sources of new antimicrobial drugs
M. S. Laport1*, O. C. S. Santosa, and G. Muricy1
1Instituto de Microbiologia Prof. Paulo de Góes, Universidade Federal do Rio de Janeiro, Bloco I, CCS,
Avenida Carlos Chagas Filho, 373, Cidade Universitária, 21491-590, Rio de Janeiro, RJ, Brazil.
2 Departamento de Invertebrados, Museu Nacional, Universidade Federal do Rio de Janeiro, Quinta da Boa
Vista s/n°, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brazil.
* Corresponding author:
Marinella Silva Laport
Departamento de Microbiologia Médica
Instituto de Microbiologia Prof. Paulo de Góes - IMPPG
Bloco I - CCS – UFRJ - Avenida Carlos Chagas Filho, 373
Cidade Universitária - 21941-590 - Rio de Janeiro - RJ - BRAZIL
e-mail: [email protected]
Key words: antibacterial, antifungal, antimicrobial, antiprotozoal, antiviral, resistance, sponges, sponge-
associated bacteria

117
Abstract: Sponges (phylum Porifera) are sessile marine filter feeders that have developed efficient defense
mechanisms against foreign attackers such as viruses, bacteria, or eukaryotic organisms. Marine sponges are
among the richest sources of pharmacologically-active chemicals from marine organisms. It is suggested that
(at least) some of the bioactive secondary metabolites isolated from sponges are produced by functional
enzyme clusters, which originated from the sponges and their associated microorganisms. More than 5,300
different products are known from sponges and their associated microorganisms, and more than 200 new
metabolites from sponges are reported each year. As infectious microorganisms evolve and develop resistance
to existing pharmaceuticals, the marine sponge provides novel leads against bacterial, viral, fungal and
parasitic diseases. Many marine natural products have successfully advanced to the late stages of clinical
trials, as for example ara-A (vidarabine), an anti-viral drug used against the herpes simplex encephalitis virus.
This substance is in clinical use for many years. Moreover, a growing number of candidates have been
selected as promising leads for extended preclinical assessment, including manzamine A (activity against
malaria, tuberculosis, HIV, and others), lasonolides (antifungal activity) and psammaplin A (antibacterial
activity). In this review we have surveyed the discoveries of products derived from marine sponges and
associated bacteria that have shown in vivo efficacy or potent in vitro activity against infectious and parasitic
diseases, including bacterial, viral, fungal and protozoan infections. Our objective was to highlight the
susbtances that have the greatest potential to lead to clinically useful treatments.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo