caracterÍsticas biolÓgicas e antigÊnicas de...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
CARACTERÍSTICAS BIOLÓGICAS E ANTIGÊNICAS DE Escherichia coli COM ÊNFASE AOS GENES DE VIRULÊNCIA
Ana Maria de Souza Almeida Orientadora: Maria Auxiliadora Andrade
GOIÂNIA
2013

ii
ANA MARIA DE SOUZA ALMEIDA
CARACTERÍSTICAS BIOLÓGICAS E ANTIGÊNICAS DE Escherichia coli COM ÊNFASE AOS GENES DE VIRULÊNCIA
Seminário apresentado junto à Disciplina de Seminários Aplicados do Programa de Pós-Graduação em Ciência Animal da Escola de Veterinária e Zootecnia da Universidade Federal de Goiás Nível: Mestrado
Área de Concentração: Sanidade Animal Higiene e Tecnologia de Alimentos
Linha de Pesquisa:
Etiopatogenia, epidemiologia, diagnóstico e controle das doenças infecciosas e parasitárias dos animais.
Orientadora:
Maria Auxiliadora Andrade
Comitê de orientação: Valéria de Sá Jayme
Guido Fontgalland Coelho Linhares
GOIÂNIA
2013

iii
SUMÁRIO
1 INTRODUÇÃO .................................................................................................... 1
2 REVISÃO DE LITERATURA ............................................................................... 3
2.1 Escherichia coli ................................................................................................ 3
2.2 Patotipos .......................................................................................................... 3
2.3 Estruturas antigênicas ...................................................................................... 6
2.4 Patogênese ...................................................................................................... 7
2.5 Ilhas de patogenicidade.................................................................................. 10
2.6 Genes de virulência ........................................................................................ 11
2.6.1 Genes de aquisição de ferro ....................................................................... 11
2.6.2 Genes plasmidiais ....................................................................................... 12
2.6.3 Adesinas fimbriais ....................................................................................... 13
2.6.4 Adesinas afimbriais ..................................................................................... 15
2.7 Toxinas ........................................................................................................... 17
3. CONSIDERAÇÕES FINAIS ............................................................................. 20
REFERÊNCIAS .................................................................................................... 21

iv
LISTA DE ABREVIATURAS
A/E lesões de fixação e esfacelamento
Aerobactina sistema de aquisição de ferro
AmpC adenosina monofosfato
APEC Escherichia coli patogênica para aves
bfp feixe formador de pelos
CDT toxina de distinção citoletal
CNF fator citotóxico necrosante
CNF-1 citotoxina necrosante fator – 1
Crl gene de filamento curli
Csg gene de filamento curli regulada pela temperatura
eae intimina
EaggEC Escherichia coli enteroagregativa
EAST-1 toxinas termoestáveis enteroagregativas
EHEC Escherichia coli enterohemorrágica
EIEC Escherichia coli enteroinvasora
EPEC Escherichia coli enteropatogênica
ETEC Escherichia coli enterotoxigênica
Hly alfa hemolisina
IL interleucina
IP ilhas de patogenicidade
iss gene de alta sobrevivência ao soro
iuc hidroxamato sideróforo aerobactina
iut gene aerobactina de captação férrica
LEE Ilhas de patogenicidade locus para esfacelamento de
enterócitos
LPS lipopolissacarídeo

v
LT toxina termolábeis
MHC-1 complexo principal de histocompatibilidade classe 1
NMEC Escherichia coli de meningite neonatal
Pap fimbria P
REDEC Escherichia coli enteropatogênica para coelhos
Sab proteína autotransportadora para formação de biofilme.
Sfa Adesina S fimbria
ST toxina termo-estável
STEC Escherichia coli produtora de toxina Shiga
SSTT sistema de secreção tipo lll
STx Shiga-toxinas
Tir receptor de intimina
TNF-α fator de necrose tumoral
TraT gene estrutural para fator de sexo F de proteínas de
superfície de membrana externa
Tsh adesina regulada pela temperatura
UFC unidades formadoras de colônias
UPEC Escherichia coli uropatogênica

1 INTRODUÇÃO
Escherichia coli (E.coli) é um microrganismo comensal, presente no
intestino de mamíferos e aves (FERREIRA & KNÖBL, 2009), entretanto é
apontado como um dos agentes bacterianos mais frequentes em diarreias de
seres humanos e animais. Além de ter grande importância nas lesões
extraintestinais em aves.
Estudos recentes têm contribuido com descobertas a respeito de novas
estirpes, mutações, genes de virulência e sistemas de resistência indentificados
em diferentes bactérias. A alta capacidade adaptativa, mudanças fenotípicas e
genotípicas dos microrganismos tornam a pesquisa sobre o seu comportamento e
sua composição, essenciais para saúde pública e animal.
Colibacilose é uma das principais doenças de aves causadas por
E.coli. Apesar do grande conhecimento a respeito dessa doença, ela ainda
persiste entre as principais enfermidades endêmicas na avicultura mundial
(DZIVA & MARK, 2008). Isso ocorre devido a diversidade antigênica da E.coli,
que dificulta o controle e prevenção de todas enfermidades vinculadas a ela.
As manifestações de doenças por E.coli são associadas a diversos
fatores, destacando os genes de virulência codificados por plasmídeos,
bacteriófagos, ou ilhas de patogenicidade (IP) (GYLES & FAIRBROTHER, 2010).
Acredita-se que a maior parte dos sorogrupos de E.coli não
apresentem qualquer gene de virulência, porém, durante o processo evolutivo,
algumas cepas adquirem diferentes combinações de genes que lhes atribuem
capacidade de promover enfermidades (CHERNAKI-LEFFER et al., 2002).
Ressalta-se que as doenças vinculadas a esse patógeno trazem
grandes perdas econômicas como; queda na produção, custos com tratamento,
desvalorização dos animais, diminuição de índices reprodutivos, mortes e
reposição dos animais, além de sua relevante importância na saúde humana. Por
isso, a detecção e caracterização molecular de genes de virulência e de suas
combinações torna possível sua associação com a patogênese da doença e
possibilita o desenvolvimento de mecanismos eficazes no combate às infeções.
A elaboração de vacinas eficazes para a prevenção dessas doenças
vinculadas a E.coli é um dos principais objetivos das pesquisas atuais. Entretanto,

2
para eficácia dessas vacinas é necessário profundo conhecimento a respeito,
sobretudo, de seus genes de virulência.
O objetivo dessa revisão é descrever as principais características
biológicas e antigênicas da E.coli, com ênfase na patogênica para aves (APEC),
bem como seus principais genes de virulência e mecanismos ligados a resistência
no hospedeiro.

3
2 REVISÃO DE LITERATURA
2.1 Características da Escherichia coli
E.coli é uma bactéria Gram negativa da família Enterobacteriaceae,
não esporulada, anaeróbica facultativa, fermentativa, em sua maioria móveis
(flagelo peretríqueos) e pertence a microbiota entérica de mamíferos e aves.
Crescem em temperaturas de 18 a 44°C sendo 37°C é a temperatura ideal
(FERREIRA & KNÖBL, 2009).
Essa bactéria também é caracterizada por suas propriedades
bioquímicas; positiva para reação para indol, lisina, motilidade e reação de
vermelho metila; negativa para testes para urease e hidrogênio e utilização de
citrato. Além disso, algumas cepas podem produzir H2S (OLIVEIRA et al. 2004;
QUINN et al. 2005).
A produção de gás e ácido ocorre posteriormente à fermentação de
manitol, glicose, sorbitol, ramanose, maltose, manose, xilose, arabiose e glicerol.
A grande maioria das estirpes é capaz de fermentar lactose, embora algumas a
fermente tardiamente (ANDREATTI FILHO, 2007).
Em meios de nutrientes sólidos as unidades formadoras de colônias
(UFC) apresentam cerca de 1 a 3mm de diâmetro tanto com aspecto rugoso
quanto liso, no entanto podem existir colônias intermediárias e mucóides.
Colônias rugosas têm aspecto grosseiro e contornos irregulares, já as colônias
lisas são convexas, brilhantes e com bordos regulares (FERREIRA & KNÖBL,
2009).
Grande parte das E.coli são comensais, não apresentam qualquer
gene de virulência (CHERNAKI-LEFFER et al., 2002). Alguns estudos descrevem
que mesmo cepas de E.coli comensais podem conter um ou mais genes de
virulência com potencial de causar doenças em animais imunossuprimidos
(KARIYAWASAM et al., 2006).
2.2 Patotipos
Com base nos mecanismos de virulência específicos das cepas
patogênicas, E.coli pode ser classificada em patotipos. São eles;
enteropatogênica (EPEC), enterotoxigênica (ETEC), enteroinvasora (EIEC),
4
enterohemorrágica (EHEC), enteroagregativa (EaggEC), uropatogênica (UPEC),
de meningite neonatal (NMEC); enteropatogênica para coelhos (REDEC) e
patogênica para aves (APEC) (FERREIRA & KNÖBL, 2009).
E.coli produtora de toxina Shiga (STEC) é um dos patotipos mais
importantes para saúde pública. Embora sua maior característica seja a produção
da shiga toxina, ela é atribuída a algumas doenças veiculadas aos alimentos,
principalmente produtos cárneos, sendo capaz de colonizar o epitélio intestinal
(CADONA et al., 2013).
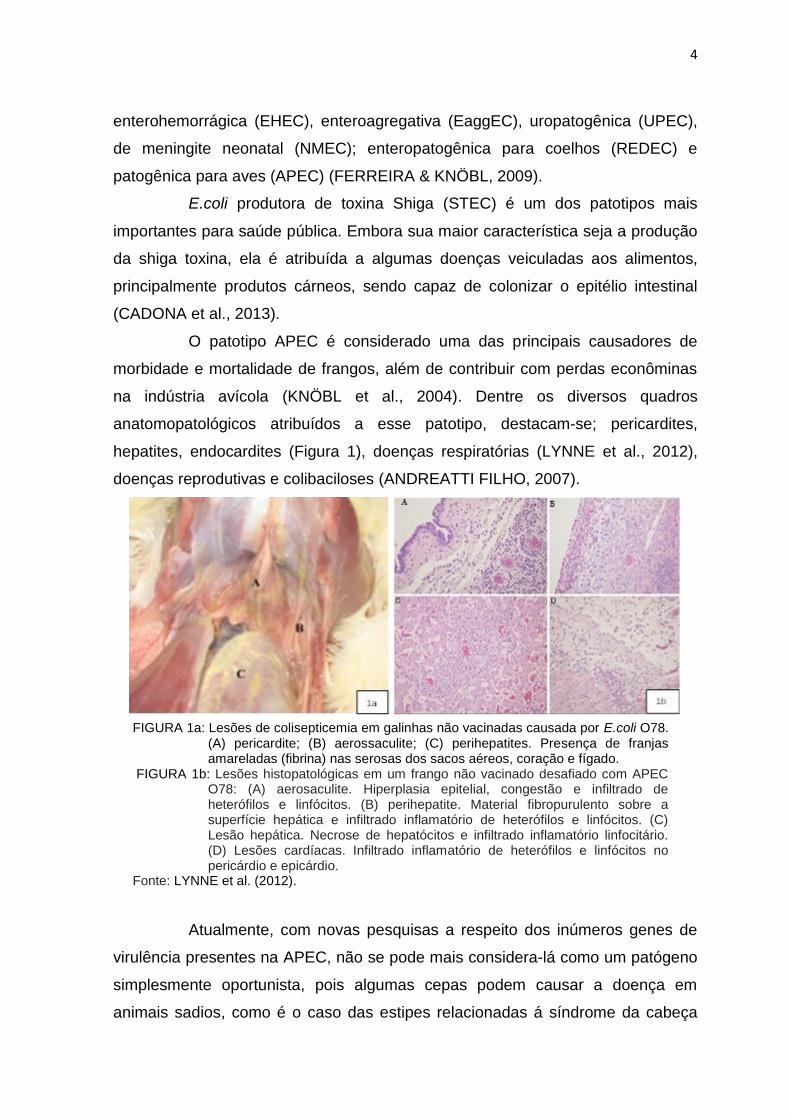
O patotipo APEC é considerado uma das principais causadores de
morbidade e mortalidade de frangos, além de contribuir com perdas econôminas
na indústria avícola (KNÖBL et al., 2004). Dentre os diversos quadros
anatomopatológicos atribuídos a esse patotipo, destacam-se; pericardites,
hepatites, endocardites (Figura 1), doenças respiratórias (LYNNE et al., 2012),
doenças reprodutivas e colibaciloses (ANDREATTI FILHO, 2007).
Atualmente, com novas pesquisas a respeito dos inúmeros genes de
virulência presentes na APEC, não se pode mais considera-lá como um patógeno
simplesmente oportunista, pois algumas cepas podem causar a doença em
animais sadios, como é o caso das estipes relacionadas á síndrome da cabeça
FIGURA 1a: Lesões de colisepticemia em galinhas não vacinadas causada por E.coli O78. (A) pericardite; (B) aerossaculite; (C) perihepatites. Presença de franjas amareladas (fibrina) nas serosas dos sacos aéreos, coração e fígado.
FIGURA 1b: Lesões histopatológicas em um frango não vacinado desafiado com APEC O78: (A) aerosaculite. Hiperplasia epitelial, congestão e infiltrado de heterófilos e linfócitos. (B) perihepatite. Material fibropurulento sobre a superfície hepática e infiltrado inflamatório de heterófilos e linfócitos. (C) Lesão hepática. Necrose de hepatócitos e infiltrado inflamatório linfocitário. (D) Lesões cardíacas. Infiltrado inflamatório de heterófilos e linfócitos no pericárdio e epicárdio.
Fonte: LYNNE et al. (2012).

5
inchada que são mais patogênicas e possuem características mais agressivas,
incluindo elevada letalidade, presença de ampla combinação de genes de
virulência e capacidade de se aderir aos diferentes tipos celulares (MATURANA et
al., 2011).
Além da diferenciação de patogenicidade e agressividade MATURANA
et al. (2011) distinguem as cepas de acordo com a doença causada por elas. Os
autores citam a existência de pelo menos dois subgrupos dentro do patotipo
APEC, classificados de acordo com seus genes de virulência e as doenças
desenvolvidas por elas. Uma delas é composta, principalmente, por estirpes que
determinam a síndrome da cabeça inchada e outra composta especialmente por
estirpes que causam onfalite. Os autores acrescentam ainda que a diversidade
existente entre estirpes que ocasionam diferentes doenças estão relacionadas ao
número de genes de virulência e suas caracteristicas fenotípicas como aderência
e letalidade. Estirpes que causam septicemia estão distribuídas entre essas duas
enfermidades. A aparente diversidade de estirpes que provocam septicemia pode
repletir na falta de identificação de um conjunto específico de genes de virulência
entre essas linhagens.
Cepas enteropatogênicas são comumente isoladas de aves com
diarreia e não isoladas de aves sadias. Porém, quando em ambiente adequado e
alimentação saudável, as aves podem albergar bactérias patogênicas sem
desenvolver a doença. Nesse caso, também podem ser isoladas de frangos
aparentemente hígidos cepas de EPEC e ETEC portadores de pili, gene que
auxilia na aderência da bactéria ao epitélio traqueal da ave, facilitando a
penetração e invasão da mucosa (KARIUKI et al., 2002).
Recentemente a preocupação com doenças ocasionadas por estirpes
de EHEC tem aumentado, pois surtos de diarreia hemorrágica estão sendo
relatados. Os bovinos são descritos como uma das principais fontes de infecção
de cepas de EHEC, que geralmente colonizam o terço final do intestino e são
excretadas pelas fezes, sem causar prejuizos ao animal (JORIS et al., 2013).

6
2.3 Estruturas antigênicas
A estrutura da APEC é composta de seguimentos antigênicos que
permitem a diferenciação sorológica e identificação de antígenos somáticos O,
flagelares H, fimbrial F e capsular K (Figura 2).
O antígeno somático O corresponde ao lipopolissacarídeo (LPS),
elemento termo-resistente que se projeta da membrana externa para o ambiente
extracelular. O lipídeo A (endotoxina), componente do LPS, é liberado durante a
multiplicação ou após a morte da bactéria, atua na ativação de macrófagos e de
mediadores da inflamação (FERREIRA & KNÖBL, 2009). O comprimento da
cadeia lateral na unidade de repetição do antígeno somático O impedem a ligação
do complexo de ataque a membrana do sistema complemento (HIRSH, 2003).
A colibacilose possui vários sorogrupos associados a sua patogenia,
porém os mais frequentes são O1, O2 e O78 (DZIVA et al., 2008). Já os
sorogrupos O2 e S8 de APEC, foram os mais encontrados por SILVEIRA et al.
(2002) e o O6 por KNÖBL el al. (2012), que caracteriza influência na prevalência
de diferentes estirpes de acordo com a região estudada.
Os antígenos capsulares K são polissacarídeos capsulares
relacionados à resistência bacteriana perante o sistema complemento. A cápsula
é um dos componentes bacterianos de menor patogenicidade, sendo removida
quando submetida a temperatura de 100°C por uma hora (FERREIRA & KNÖBL,
2009).
Antígenos flagelares H são compostos proteicos não utilizados com
frequência na identificação antigênica das cepas de E.coli, nem a sua
patogenicidade tem sido relacionada à presença do flagelo (ANDREATTI FILHO,
2007; FERREIRA & KNÖBL, 2009).
Os antígenos fimbriais F, também chamados de adesinas, pili ou
fímbrias, são moléculas proteicas que recobrem a superfície bacteriana, capazes
de reconhecer receptores específicos na superfície de células eucarióticas. A
expressão de adesinas é considerada um gene de virulência fundamental para
aderência e colonização dos tecidos do hospedeiro. Elas também conferem
especificidade de aderência da bactéria em relação a determinados tecidos e
órgãos do hospedeiro. Embora essas adesinas apresentem poucas diferenças

7
morfológicas existem características antigênicas e hemaglutinantes distintas
(FERREIRA & KNÖBL, 2009).
2.4 Patogênese
O conceito de patogenicidade é a capacidade de um microrganismo
causar a enfermidade. Os microrganismos da mesma espécie se diferenciam uns
dos outros por possuírem ou expressarem genes de virulência que proporcionam
colonização e expressão de inúmeros mecanismos que dificultam o combate do
hospedeiro, ocasionando assim a doença (VIEIRA, 2009).
De acordo com o patotipo, com o sorogrupo e com a presença de
genes de virulência as estirpes de E.coli podem causar desde quadro leves de
diarréia até doenças septicemicas graves. Adesinas, sistema de captação de
FIGURA 2 – Ilustração da estrutura antigênica molecular de E.coli. A= estrutura molecular do flagelo em relação ao corpo bacteriano; B= estrutura do polissacarídeo e a sua relação com lipídeo A; C= estrutura do pili em relação ou fímbria que se projeta da membrana externa da bactéria. PS= polissacarídeo; LA= localização do lipídeo A ou endotoxina; PG= peptideoglicano ou citoesqueleto das bactérias Gram negativas; MC= membrana citoplasmática em oposição à membrana externa que ancora o flagelo, pili e LPS.
Fonte: FERREIRA & KNÖBL (2009), com adaptações.

8
ferro, invasinas, toxinas e os fatores, inibitórios do sistema imune do hospedeiro e
genes de resistência a antimicrobianos são os genes mais importantes de
virulência (VIEIRA, 2009).
A infecção por ETEC se dá pela via oral e a fixação no epitélio do
intestino delgado ocorre pela ação de adesinas fimbriais (Figura 3). Após a
aderência entre a bactéria e a célula hospedeira há produção de enterotoxinas
que estimulam a secreção de água e eletrólitos no lúmen intestinal, ocasionando
diarreia líquida, desidratação, acidose metabólica e morte. Grandes quantidades
de LPS podem ser liberadas, aumentando o número de fatores de necrose
tumoral (TNF-α), interleucina L-1 e L-6 que contribuam para um quadro de choque
(WHITT & SALYERS, 2002; HISRH, 2003).
O estabelecimento da infecção por EPEC se dá por fraca fixação de
adesinas bfp (feixe formador de pelos) no cão e AF/R1, AF/R2 e Ral em coelhos
(REDEC). Um sinal transmitido da bactéria para a célula hospedeira, ativa
intiminas bacterianas (gene eae), provocando alterações no citoesqueleto celular
hospedeiro e posterior desenvolvimento de lesões de fixação e esfacelamento
(A/E) das vilosidades. O patotipo EHEC além das lesões de A/E produz também
toxinas shiga-like, shiga toxina ou verotoxinas. Stx-1 e Stx-2, codificadas por
bacteriófagos, que lesionam e provocam perda da integridade das células
endoteliais, causando diarréia hemorrágica (HIRSH, 2003).
FIGURA 3 – Ataque de ETEC as células epiteliais do íleo de um leitão desmamado, mostrando íntima aderência bacteriana (A) e o efeito nas células epiteliais. Barra = 1 μm.
Fonte: GYLES & FAIRBROTHER (2010), com adaptações.

9
A adesão ao trato respiratório pela APEC, também é mediado por
fimbrias, com destaque para fimbria tipo 1 (Figura 4), fimbria P (pap) e curli. A
adesina regulada pela temperatura (tsh) esta relacionada aos estágios iniciais da
infecção. O antígenos K e o gene de alta sobrevivência no soro (iss) tornam estas
estirpes mais resistentes ao efeito bactericina do soro, e o sistema de aquisição
de ferro (aerobactina) possibilitam a permanência e infecção do hospedeiro
(HIRSH, 2003; GYLES & FAIRBROTHER, 2010), ocasionando lesões sistêmicas
como; infecções respiratórias (aerossaculites), hepatites, perihepatites e
endocardites (LYNNE et al., 2012).
SZEMLAKO et al. (2013) afirmam que a ocorrência da combinação
fímbria P e genes que codificam alfa-hemolisina (hly) são mais frequentes entre
as E.coli que causam septicemias.
O início da colonização da UPEC ocorre na região periuretral e em
seguida move-se para uretra. A colonização da bexiga é mediada pelo pili tipo1 e
fimbria P. A presença de sideróforos e hemolisinas permitem a captação de ferro
e o antígeno K confere resistência à fagocitose, estimulando a produção de
citotoxina necrosante fator – 1 (CNF-1) e as alfa-hemolisinas lesionam as células
FIGURA 4 – Eletromicrografia de uma amostra de E.coli de origem aviária. Nota-se na superfície bacteriana a projeção de estruturas finas e longas denominadas de pili ou fímbrias, sendo caracterizado como pili Tipo 1. Coloração negativa – molibdato de amônio (30X).
Fonte: FERREIRA & KNÖBL (2009).

10
epiteliais hospedeira, provocando quimiotaxia de neutrófilos, causando cistites e
pielonefrites (WHITT & SALYERS, 2002).
Cepas EaggEC produzem toxinas termolábeis (LT), toxinas
termoestáveis enteroagregativas codificadas por um plasmídeo (EAST-1), toxinas
termo-estáveis (ST), toxina do tipo hemolisina e em alguns casos produzem
Shiga-toxinas. Essas toxinas auxiliam a instalação da infecção (CAMPOS &
TRABULSI, 2002).
2.5 Ilhas de patogenicidade
Ilhas de patogenicidade (IP) são ilhas genômicas, constituídas por
amplas regiões cromossômicas (10.000 a 200.000 pares de bases) de alta
instabilidade e com características distintas do restante do genoma bacteriano.
Codificam um ou mais genes de virulência, e parte ou todo arsenal molecular para
que esses genes possam alcançar sua célula alvo (VIEIRA, 2009).
IP são compostas por guanidina e citosina (G+C), diferentes do
restante do cromossomo e estão comumente associadas a genes que codificam
RNA transportador. Possuem hot spots em suas extremidades, que são
elementos envolvidos na mobilidade genética (local de inserção ou deleção
genômica simplificada), como; fagos temperados, sequências de inserção e
integrases (VIEIRA, 2009).
Estudos a respeito de IP são essenciais para detecção de genes de
virulência, que por sua vez permitem a identificação e caracterização de E.coli
patogênicas. Um exemplo são as lesões do fenótipo A/E codificadas na IP locus
para esfacelamento de enterócitos (LEE), presente principalmente no patotipo
EPEC, STEC (GYLES & FAIRBROTHER, 2010) e EHEC (JORIS et al., 2013).
LEE codifica também adesinas de membrana externa denominada
intimina (gene eae), sistema de secreção tipo lll - SSTT (Esc e proteína Sep),
chaperones (proteínas Ces), translocadores (EspA, EspB e EspD), proteínas
efetoras (EspF, EspG e Map) e receptores translocados intimina (Tir) (GIRARD,
2005), demostrando a importância de cada IP na patogênese bactéria.
Estudos recentes com finalidade de determinar se a presença de
anticorpos e/ou a cinética das respostas de anticorpos contra eae, Tir, EspA,

11
EspB pode ser usada para monitorar infecções por EHEC, foram feitos rebanhos
de gado de corte no Brasil. Os resultados demonstraram que apenas respostas
séricas à EspA e EspB podem ser utilizadas para monitorar o estado de infecção
de rebanhos bovinos com diarreia causada por EHEC. A correlação entre a
presença de anticorpos contra eae ou Tir e a baixa prevalência de EHEC não
pode ser presumida, pois, rebanhos negativos para essa estirpe podem
apresentar esses mesmos anticorpos (JORIS et al., 2013).
Fimbria P, fimbria S, hemolisinas, CNF-1 e sistema de sequestro de
ferro também são genes codificados em IP. No entanto, alguns podem ser
encontrados tanto em IP quanto em plasmídeos, como os genes hly e
aerobactina. Existem ainda, aqueles codificados do genoma nuclear, são eles;
fimbria tipo 1, antígenos especificos O (comumente O1, O2, O4, O6 e O25), e
aerobactina (GYLES & FAIRBROTHER, 2010).
2.6 Genes de virulência
2.6.1 Genes de aquisição de ferro
A aquisição de ferro é um importante fator de virulência para bactérias
(WHITT & SALYERS, 2002). E.coli possui recursos de aquisição e assimilação de
ferro proveniente do hospedeiro (GYLES & FAIRBROTHER, 2010). São eles; os
sideróforos (enterobactina, aerobactina e outros), que promovem esse sequestro
e aquisição através da remoção do ferro de proteínas carreadoras (HIRSH, 2003;
GYLES & FAIRBROTHER, 2010) e as hemolisinas, que capturam ferro obtido da
lise de eritrócitos (ROCHA et al., 2002).
O “operon” aerobactina contém o gene responsável pela produção de
hidroxamato sideróforo aerobactina (iuc), com suas respectivas variantes
(iucABCD), e o gene aerobactina de captação férrica (iutA). A capacidade de
obtenção de ferro pelo sistema aerobactina é devidamente estabelecida, porém,
em algumas situações como é o caso da estirpe APEC O2, esse sistema é
incompleto, pela ausência de iucA. No entanto, a presença de “operons”
cromossômicos yersinabacterina e enterobactina nessa estirpe, compensam a
perda de iucA, tornando a captação de ferro possível (LING et al., 2013).

12
2.6.2 Genes plasmidiais
A transferência de material genético entre as bactérias é outra forma de
aquisição de genes de virulência, com destaque para a conjugação, em que o
simples contato com outras bactérias vivas possibilita a transmissão de genes de
virulência presentes em uma molécula de DNA extra-cromossômico, denominada
plasmídeo (VIEIRA, 2009).
O rápido desenvolvimento de resistência das enterobactérias às
cefalosporinas de largo espectro está relacionada a cepas cujos plasmídeos
produzem beta-lactamases de espectro estendido – ESBLs e beta-lactamase tipo
AmpC, que hidrolisam o anel beta-lactâmico destes antimicrobianos. Em 2006, o
Programa de Vigilância a resistência antimicrobiana Holandês, isolou 153 E.coli
obtidas de amostras cecais de frangos saudáveis em diferentes abatedouros na
Holanda, em que 80% das amostras resistentes a cefotaxima carreavam o gene
ESBL e 17% carreavam o gene AmpC (DIERIKX et al., 2010).
Diferente de outros patotipos, a STEC é capaz de transportar grandes
plasmídeos, que codificam dentre outros genes de virulência, o gene sab que se
adere tanto à superfícies abióticas quanto a células epiteliais (CADONA et al.,
2013), produzindo biofilme (BUVENS & PIÉRARD, 2012). Já em pesquisas feitas
na Bélgica a partir STEC isoladas de humanos, e carnes cruas de ruminantes e
animais silvestres, o gene sab foi infrequente nesse patotipo quando LEE era
negativo (BUVENS & PIÉRARD, 2012). Os autores acrescentam que talvez sab
possa promover recursos alternativos nos mecanismos de adesão em STEC LEE
negativos, porém precisa-se de maiores estudos à respeito. Outro plasmídeo,
vinculado principalmente a certas estirpes de STEC como O157: H7 é o
plasmídeo-hemolisina, que condifica enterohemolisinas (GYLES &
FAIRBROTHER, 2010).
Outro gene plasmidial de grande importância é o brp ou gene colicina,
que produz e secreta colicinas. As colicinas são toxinas proteicas acionadas por
bactérias no combate à célula hospedeira (WHITT & SALYERS, 2002; LLOUBES
et al., 2013).

13
TraT e iss também são genes plasmidiais, codificados pelo plasmídeo
ColV. São genes determinantes para resistência sérica, aos efeitos bactericidas
do sistema complemento e à fagocitose, comumente relacionados a cepas
septicêmicas (GYLES & FAIRBROTHER, 2010). Essa resistência é atribuída a
produção de proteínas de membrana externa (OMPs), produzida por esses genes
(KNÖBL et al., 2001; MONROY et al., 2005). O “operon” aerobactina também é
codificado pelo plasmídeo ColV em estirpes APEC O2 (LING et al., 2013).
Além da codificação via plasmídeo, o gene iss também pode ser
codificado no cromossomo bacteriano. Ultimamente sabe-se que muitas cepas de
E.coli tem iss em seus cromossomos e sua seleção para os diferentes alelos iss
possibilita uma melhor detecção desse gene entre estirpes (JOHNSON et al.,
2008). O gene iss vem sendo isolado de papagaios com colibacilose (KNÖBL et
al., 2008), porém não há estudos a respeito do seu local de codificação.
Análises das variantes alélicas tornam possível o estudo sobre a
evolução iss e o conhecimento de suas variantes, afim de explorar a possibilidade
que esse gene e a proteína codificada por ele são ferramentas úteis para
preparação de vacinas (JOHNSON et al., 2008).
Um estudo piloto para produção de vacina contra a colibacilose em
aves baseado no gene iss mostrou bons resultados, em que galinhas imunizadas
com glutationa-S-transferase-Iss (GST-Iss) foram capazes de produzir anticorpos
contra GST-Iss e iss, além de induzirem resposta imunoprotetora em desafios
com duas estirpes de APEC diferentes. A forte associação entre iss e APEC,
juntamente com a localização de iss na membrana externa, possibilita estratégias
de uso desse gene em vacinas para controle de colibacilose (LYNNE et al., 2012).
2.6.3 Adesinas fimbriais
A maioria das estirpes de E.coli apresentam algum tipo de aderência
(Figura 5). Um exemplo é a intimina eae, responsável pela ligação íntima entre a
bactéria e o epitélio intestinal do hospedeiro (SILVEIRA et al., 2002). A sua
ausência promove diminuição significativa na aderência bacteriana, como em
algumas estirpes de STEP e EPEC, por exemplo (GIRARD, 2005).

14
Os genes de virulência pap ou fímbria P (ROCHA et al., 2002; KNÖBL
et al., 2004) e sfa são fímbrias D+ manose resistentes na mediação de adesão da
E. coli para diferentes tecidos do hospedeiro. A presença dos genes pap e sfa
varia entre estirpes associadas a diferentes doenças. O gene pap é encontrado
com maior frequência em aves com síndrome na cabeça inchada e doença
respiratória crônica, já o gene sfa é mais comum em isolados de onfalites,
sugerindo assim, que esses genes possibilitam infecções extra-intestinais em
diferentes locais (KNÖBL et al., 2004).
A utilização do carboidrato D+ manose permite classificar as fímbrias
em: manose sensíveis, quando a hemaglutinação é inibida pela presença do
carboidrato, e manose resistentes quando a hemaglutinação ocorre na presença
de manose. Estudos mostram que as adesinas manose sensíveis (pili tipo 1 ou
tipo1 like) são responsáveis pela colonização de traquéia e sacos aéreos durante
a fase inicial da doença, enquanto a colonização de órgãos internos e o
desenvolvimento de septicemias dependa da expressão de adesinas manose
resistentes (fímbria P) (FERREIRA & KNÖBL, 2009).
Mudanças no padrão de adesão observadas na presença ou na
ausência de D+manose sugerem associação de mais de um tipo de fímbria. A
adesão exclusivamente na ausência de D+manose indica que o pili tipo 1 ou tipo
FIGURA 5 – Aderência de E.coli O78 patogênica para aves nas células ciliadas da traquéia, aderência ao muco secretado pelas células secretórias de muco da traquéia. Giemsa 140X.
Fonte: FERREIRA & KNÖBL (2009).

15
1 like, podem participar da patogênese de certos patotipos (SILVEIRA et al.,
2002).
Cada variante do gene pap (papA, papC, papE, papF, etc) é
responsável por codificar diferentes subunidades das fímbrias, como o papA que
codifica o eixo principal da fímbria. Dentre as variantes do gene pap destaca-se o
papC por sua maior frequência em APEC do que nos demais patotipos. Genes
pap não são essenciais para a patogênese da APEC, porém podem desempenhar
um papel significativo nesse processo. No entanto, papA e APEC são
antigenicamente correlacionadas e é provável que esse gene proporcione uma
vantagem evolutiva pela aquisição de DNA estranho ou supressão de
informações genéticas existentes, resultando no desenvolvimento de novas
estirpes e até mesmo patotipos (KARIYAWASAM & NOLAN, 2011).
PapA, diferente dos genes papC e papG, é isolado apenas de cepas
patogênicas, não sendo encontrado em cepas comensais avícolas
(KARIYAWASAM et al., 2006). Estudos relacionados ao sequenciamento genético
de papA em APEC, revelaram a possibilidade da presença de um transposon
Tn10 (resistência a tetraciclina), possivelmente selecionado devido à prática
comum de utilização desses antimicrobianos como promotores de crescimento e
na terapéutica das aves. A incorporação de genes de resistência antimicrobiana
ao genoma do “operon” pap pode trazer vantagens na sobrevivência dessas
bactérias (KARIYAWASAM & NOLAN, 2011).
Além das características decritas acima SZEMLAKO et al. (2013)
sugerem que pap com capacidade para causar resposta inflamatória grave podem
auxiliar na ultrapassagem de barreiras dos tecidos, possibilitando o acesso à
corrente sanguínea, causando bacteremia.
2.6.4 Adesinas afimbriais
O receptor de intimina Tir é uma proteína transportada através do
sistema de secreção tipo lll (SSTT) (Figura 6) para o citoplasma da célula
hospedeira, onde atua como receptor de eae, que é expressa na superfície
bacteriana, resultando em íntima ligação com a célula hospedeira. A
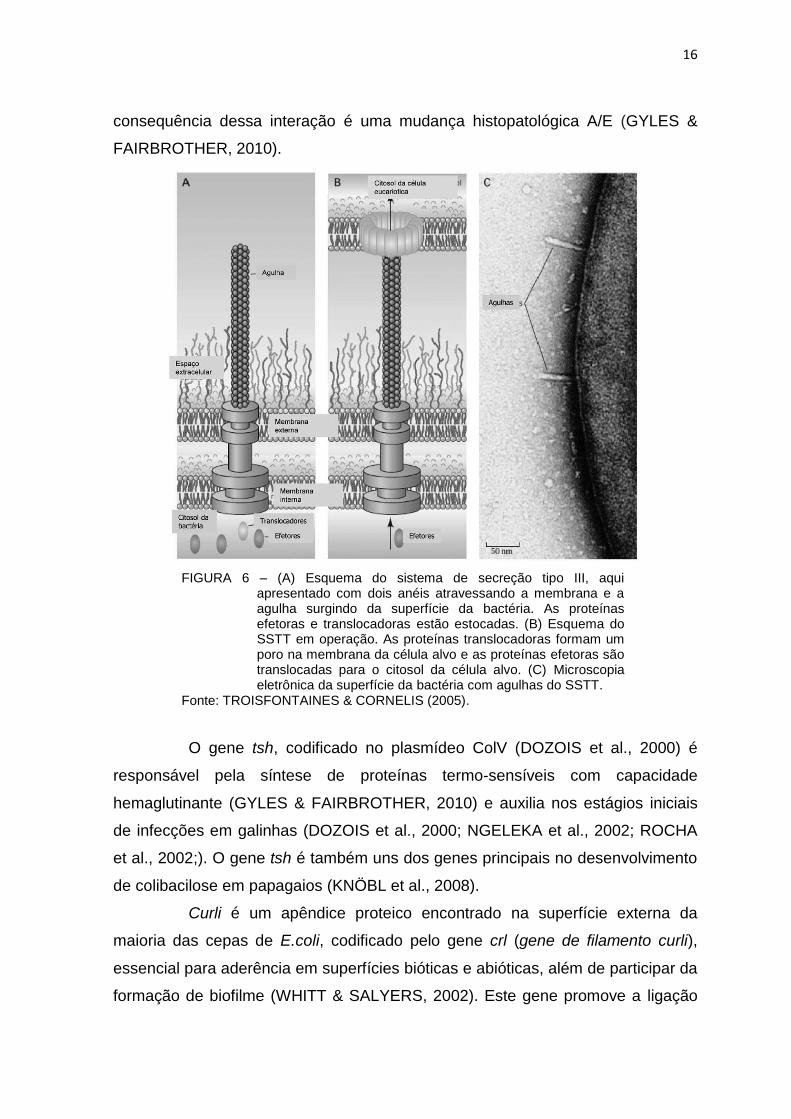
16
consequência dessa interação é uma mudança histopatológica A/E (GYLES &
FAIRBROTHER, 2010).
O gene tsh, codificado no plasmídeo ColV (DOZOIS et al., 2000) é
responsável pela síntese de proteínas termo-sensíveis com capacidade
hemaglutinante (GYLES & FAIRBROTHER, 2010) e auxilia nos estágios iniciais
de infecções em galinhas (DOZOIS et al., 2000; NGELEKA et al., 2002; ROCHA
et al., 2002;). O gene tsh é também uns dos genes principais no desenvolvimento
de colibacilose em papagaios (KNÖBL et al., 2008).
Curli é um apêndice proteico encontrado na superfície externa da
maioria das cepas de E.coli, codificado pelo gene crl (gene de filamento curli),
essencial para aderência em superfícies bióticas e abióticas, além de participar da
formação de biofilme (WHITT & SALYERS, 2002). Este gene promove a ligação
FIGURA 6 – (A) Esquema do sistema de secreção tipo III, aqui apresentado com dois anéis atravessando a membrana e a agulha surgindo da superfície da bactéria. As proteínas efetoras e translocadoras estão estocadas. (B) Esquema do SSTT em operação. As proteínas translocadoras formam um poro na membrana da célula alvo e as proteínas efetoras são translocadas para o citosol da célula alvo. (C) Microscopia eletrônica da superfície da bactéria com agulhas do SSTT.
Fonte: TROISFONTAINES & CORNELIS (2005).

17
entre o complexo principal de histocompatibilidade classe 1 (MHC-1), entre a
matriz extracelular, entre as proteínas séricas e entre as células intestinais
(NGELEKA et al., 2002; WHITT & SALYERS, 2002).
NENNINGER et al. (2011) citaram que as variantes do gene curli, csgG
e csgE, agem em conjunto para proporcionar estabilidade na secreção de outras
variantes como; csgA, csgB e csgF. O gene csgE pode inibir a polimerização de
csgA purificado nas fibras amilóides, sugerindo que certas estirpes de E.coli
podem impedir a formação de amilóide. Os autores acrescentaram, que cepas
que superexpressam csgG são sensíveis à Eritromicina, porém quando há
indução da expressão de csgE essas cepas se tornam novamente resistentes a
este antimicrobiano.
Em isolados de E. coli enteropatogênicas de humanos com diarréria
pode-se encontrar modificações na presença de variantes do gene curli, como
nos estudos feitos por HERNANDES et al. (2012), onde 97% dos isolados de
EPEC possuiam o gene csgD e apenas 19,7% apresentavam gene csgA, que
indica alta diversidade na sequencia de nucleotídeos deste ultimo gene.
SAIDENBERG et al. (2013) detectaram os genes iss, pap, iuc, hlyA e
tsh de E.coli isoladas de Mutum-do-Nordeste (Pauxi mitu), ave considerada em
extinção, aparentemente sadios. Esses achados foram motivo de preocupação,
pois fatores de estresse, como: início da época de reprodução; doenças
concomitantes e manejo impróprio do ambiente e da criação podem desencadear
desequilíbrios no organismo e assim conduzir a manifestação de colibacilose, que
pode ter resultados graves em uma espécie nativa ameaçada de extinção.
2.7 Toxinas
As toxinas produzidas pelas diferentes estirpes de E.coli também são
fatores importantes para patogênese das enfermidades atribuídas a essa bactéria.
GYLES & FAIRBROTHER (2010) pontuam as enterotoxinas termo-estável (STa,
STb), termo-estável enteroagregativa (EAST-1) e termo-lábil (LT) como
promotoras de distúrbios metabólicos dos fluidos intestinais.
A STa induz acúmulo de líquido na luz intestinal de camundongos
lactentes e leitões (HIRSH, 2003; QUINN et al. 2005; GYLES & FAIRBROTHER,

18
2010), provocando diminuição e atrofia das vilosidades. Já o STb é inativado pela
presença da tripsina (GYLES & FAIRBROTHER, 2010) e afeta leitões lactentes e
desmamados (HIRSH et al., 2003).
ARAGÃO et al. (2012), descrevem que a detecção de determinadas
toxinas, como STa isolada por eles, na maioria dos isolados de ETEC obtidos de
um surto de colibacilose em carneiros, possibilita a identificação do patotipo e
sugerem que esta toxina contribua para o desenvolvimento da enfermidade, até
mesmo em casos de ausência das demais toxinas.
Já a LT induz a secreção de fluidos através da estimulação de
prostaglandinas e citocinas no sistema nervoso entérico, possui efeitos
imunomoduladores potentes e ainda pode induzir a apoptose de leucócitos. O
subtipo LT – l torna o enterócito afetado, um hipersecretor de AMPc, provocando
secreção excessiva de eletrólitos e água, causando desidratação, acidose
metabólica e morte. Os mecanismos de LT – 2 ainda são obscuros e necessitam
de maiores estudos (GYLES & FAIRBROTHER, 2010).
Stx são proteínas citotóxicas inibidoras da síntese proteica nas células
hospedeiras, semelhantes à da toxina produzida por Shigela (HIRSH, 2003).
Elevados índices de produção dessa toxina estão relacionados à indução de
bacteriófagos, como durante a ação dos antimicrobianos, que causam danos ao
DNA bacteriano ou na parede celular da bactéria. Stx1 está envolvida na
regulação de ferro, presente em maior número nos ambientes de privação de
ferro. Stx2 geralmente esta associada à doença do edema (ataque à células
endoteliais) ou em outras enfermidades vinculadas a suínos (GYLES &
FAIRBROTHER, 2010).
Outras toxinas consideradas importantes são toxina de distinção
citoletal (CDT), fator citotóxico necrosante (CNF), hemolisinas e as
enterohemolisinas necrosantes (MEHDIPOUR et al., 2012). A CDT tem como
função desencadear a parada do ciclo celular, induzindo a apoptose celular. Já as
CNF são atribuídas a diarreia e septicemia em bezerros (WHITT & SALYERS,
2002)
Pesquisas feitas para avaliar a ocorrência e funcionalidade CNF e CDT
em E.coli isoladas de bezerros e cães apresentaram inúmeras combinações de
genes de virulência e foi possível associar a cistite demonstrada por alguns

19
animais, com a presença de CNF-1 correlacionada com papC, hlyA e sfa
(SALVARANI et al., 2012), sugerindo que combinações de genes e dessa toxina
podem ser necessárias para o desenvolvimento da cistite. CNF e CDT foram as
toxinas mais detectadas de E.coli isoladas de carcaças de carneiros iranianos em
pesquisas de MEHDIPOUR et al. (2012), definindo a importância dessas toxinas
em diferentes espécies.
O gene hly (alfa hemolisina), além de lesionar as células hospedeiras,
suprime a produção de citocinas e interleucinas pela célula eucariótica, permitindo
que a bactéria se estabeleça no organismo hospedeiro. Cepas que não possuem
genes que promovem hemólise consequentemente não apresentam citotoxicidade
elevada e não são capazes de suprimir citocinas, dificultando assim sua
colonização no hospedeiro (HILBERT et al., 2012). Já a pesquisa feita com a
UPEC sugerem que a hly é um gene de virulência que proporciona a migração do
trato urinário para a corrente sanguínea, causando septicemia (SZEMLAKO et al.,
2013).
CNF e hly são toxinas conhecidamente importantes no estabelecimento
de doenças em mamíferos, porém não há dados aprofundados em relação as
aves. Alguns estudos associados a APEC sugeriram que essas toxinas atuam
como facilitador na transposição da barreira interespécies (KNÖBL el al., 2012).
As colicinas são proteínas tóxicas produzidas e secretadas no meio
extracelular pelas E.coli e outras bactérias (WHITT & SALYERS, 2002). O ataque
às células alvo se dá por despolarização da membrana citoplasmática, por
atividade citotóxica contra ácidos nucléicos citoplasmáticos, ou por interferirem
com a biossíntese de peptiodeoglicanos (WHITT & SALYERS, 2002; LLOUBES et
al., 2013).
Em estudos realizados em isolados de E.coli obtidos de papagaios com
colibacilose no Brasil, não foram identificados genes que codifiquem toxinas,
tornando o conhecimento mais aprofundado sobre esses e outros genes de
virulência em psitacídeos relevante (KNÖBL et al., 2008).

20
3. CONSIDERAÇÕES FINAIS
Escherichia coli possui diferentes classificações perante seus
sorogrupos e mecanismos de virulência. Essa diversidade antigênica e de
patotipos pode explicar a natureza da manifestação clinico-patológica da maioria
das doenças causadas por ela.
Os inúmeros genes de virulência, que podem estar presentes em
isolados de E.coli, desenvolvem mecanismos complexos para invasão e
colonização da célula hospedeira. Mais estudos são necessários para
identificação e caracterização destes genes, para proporcionar maior clareza do
desenvolvimento dos processos patogênicos utilizados por essas bactérias para
infectar seus hospedeiros.
A facilidade de transmissão dos genes plasmidiais dificulta o controle
de determinadas bactérias e é importante no aumento da patogenicidade de
estirpes. Genes de aquisição de ferro e genes de resistência sérrica são
essenciais para E. Coli, pois, facilitam a infecção do hospedeiro mesmo em
ambientes adversos. As adesinas são elementos essenciais nos estágios iniciais
da invasão ao organismo hospedeiro. A combinação de genes de virulência com
diferentes finalidades tornam estirpes cada vem mais difícil o seu combate pelo
organismo hospedeiro e dificultam a ação dos antimicrobianos.
Além disso, conhecimentos mais aprofundados a respeito da
diversidade antigênica da E.coli poderão, futuramente, contribuir para elaboração
de vacinas eficazes no controle de enfermidades atribuídas a essa bactéria.

21
REFERÊNCIAS
1. ANDREATTI FILHO, L. R. Saúde aviária e doenças. São Paulo: Roca, 2007. vol. 10, p. 112-117.
2. ARAGÃO, A. Z. B.; TEOCCHI, M. A.; FREGOLENTE, M. C.; GATTI, M. S. V.; PIRES, A. V.; YANO, T. Colibacillosis in lambs is associated to type 1 heat-stable enterotoxin in a farm São Paulo State, Brazil. Ciência Rural, Santa Catarina, v.42, n.5, p.854-857, 2012. [NOTE]
3. BUVENS, G.; PIÉRARD, D. Low prevalence of STEC autotransportador contibunting to biofilm (Sab) in verocytotoxin-producinf Escherichia coli isolates of humans and raw meats. European Journal of Clinical Microbiology e Infectious Diseases [online], Wiesbaden, v.31, n.7, p.1463-1465, 2012. Disponível em: http://link.springer.com/article/10.1007%2Fs10096-011-1464-y. Acesso em: 5 set 2013
4. CADONA, J. S.; BUSTAMANTE, A. V.; PARMA, A. E.; LUCCHESE, P. M. A.;
SANSO, A. M. Distribution of additional virulence factors related to adhesion and toxicity in Shiga toxin-producing Escherichia coli isolated from raw products in Argentina. Applied Microbiology, Washington, v.56, n.1 p.449-455, 2013.
5. CAMPOS, L.C.; TRABULSI, L.R. Escherichia. In.: TRABULSI, L.R. et al.
Microbiologia. 3 ed. São Paulo: Atheneu, 2002, p.215-228.
6. CHERNAKI-LEFFER, A.M.; BIESDORF, S.M.; ALMEIDA, L.M.; LEFFER, E.V.B; VIGNE, F. Isolamento de enterobactérias em Alphitobius diaperinus e na cama de aviários no oeste do estado do Paraná. Revista Brasileira de Ciência Avícola, Campinas, v.4, n.3, p. 243-247, 2002.
7. DIERIKX, C.; ESSEN-ZANDBERGEN, A. V.; VELDMAN, K.; SMITH, H.;
MEVIUS, D. Detection of extended spectrum beta-lactamase producing Salmonella enterica and Escherichia coli isolates from poultry. Veterinary Microbiology, Amsterdam, v.145, n.1, p.273-278, 2010.
8. DOZOIS, C. M.; DHO-MOULIN, M.; BRÉE, A.; FAIRBROTHER, J. M.;
DESAUTELS, C.; CURTISS, R. Infection and Immunity, Washington, v.68, n.7, p.4145-4154, 2000.
9. DZIVA, F.; MARK, S. Colibacillosis in poultry: Unravelling the molecular basis
of virulence of avian pathogenic Escherichia coli in their natural hosts. Avian Pathology, Huntingdon, v.37, n.1, p.355-366, 2008.

22
10. FERREIRA, A. J. P.; KNÖBL, T. Enfermidades bacterianas. In: JÚNIOR BERCHIERI, A.; SILVA, NEPOMUCENO, E.; DI FÁBIO, J.; SESTI, L.; ZUANAZE, M. A. F. Doenças das aves. Campinas: Facta, 2009. cap.4, p. 457-474.
11. GIRARD, F.; BATISSOON, I.; FRANKEL, G. M.; HAREL, J.; FAIRBROTHER,
J. M. Interaction of enteropathogenic and shiga toxin-producing Escherichia coli and porcine intestinal mucosa: Role of intimin and tir in adherence. Infection and Immunity, Washington, v.73, n.9, p.6005-6016, 2005.
12. GYLES, C. L.; FAIRBROTHER, J. M. Escherichia coli. In: GYLES, G. L.;
PRESCOTT, J. F.; SONGER, G.; THOEN, C. O. Pathogenesis of bacterial infections in animals. 4d. Iowa: Blackwell Publishing, 2010. cap. 15, p. 266-308.
13. HERNANDES, R. T.; VELSKO, I.; SAMPAIO, S. C. F.; ELIAS, W. P.;
ROBINS-BROWNE, R. M.; GOMES, T. A. T.; GIRÓN, J. A. Fimbrial adhesins produced by atypical enteropathogenic Escherichia coli strains. Applied and Environmental Microbiology, Washington, v.77, n.23, p.8391-8399, 2011.
14. HILBERT, D. W.; PAULISH-MILLER, T. E.; TAN, C. K.; CAREY, A. J.; ULETT,
G. C.; MORDECHAI, E.; ADELSON, M. E.; GYGAX, S. E.; TRAMA, J. P. Clinical Escherichia coli isolates utiliza alpha-hemolysin inhibit in vitro epitelial cytokine production. Microbes and Infection, Paris, v.14, n.7-8, p.628-638, 2012.
15. HIRSH, D. D. Escherichia coli. In: ____. Microbiologia Veterinária. 2 ed. Rio
de Janeiro: Granabara Koogan SA, 2003. cap. 9, p. 63-69. 16. JOHNSON, T. J.; WANNEMUEHLER, Y. M.; NOLAN, L. K. Evolution of the iss
gene in Escherichia coli. Applied and Environmental Microbiology, Washington, v.74, n.8, p.2360-2369, 2008.
17. JORIS, M. A.; VANROMPAY, D.; VERSTRAETE, K.; REU, K.; ZUTTER, L.;
COX, E. Use of antibody responses against locus of enterocyte effacement (LEE) – Encoded antigens to monitor enterohemorragic Escherichia coli infections on cattle farms. Applied and Environmental Microbiology, Washington, v.79, n.12, 3677-3683, 2013.
18. KARIUKI, C.; GILKS, J.; MUYODI, J.; GETTY, B.; HART, A. Carriage of
potentially pathogenic Escherichia coli in chickens. Avian Diseases, Kennet Square, v.46, n.3, p.721-724, 2002.
19. KARIYAWASAM, S.; JOHNSON, T. J.; DEBROY, C.; NOLAN, L. K.
Occurrence of pathogenicity island lAPEC-O1 genes among Escherichia coli implicated in avian colibacillosis. Avian diseases, Kennet Square, v.50, p.405-410, 2006.

23
20. KARIYAWASAM, S.; NOLAN, L. K. PapA gene of avian pathogenic Escherichia coli. Avian diseases , Kennet Square, v.55, n.1, p.532-538, 2011.
21. KNÖBL, T.; BACCARO, M. R.; MORENO, A. M.; VIEIRA, M. A.; FERREIRA, C. S.; FERREIRA, A. J. Virulence properties of Escherichia coli isolated from ostriches with respiratory disease. Veterinary Microbiology, Amsterdam, v.2283, n.1, p.71-80, 2001.
22. KNÖBL, T.; GODOY, S. N.; MATUSHIMA, E. R.; GUIMARÃES, M. B.; PIANTINO, A. J. Caracterização molecular dos fatores de virulência de estirpes de Escherichia coli isoladas de papagaios com colibacilose aviária. Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v.45, n.1, p.54-60, 2008.
23. KNÖBL, T.; GOMES, T. A. T.; VIEIRA, M. A. M.; BOTTINOA, J. A.;
FERREIRA, A. J. P. F. Detection of pap, sfa, afa and fim adhesin-encoding operons in avian pathogenic Escherichia coli. International Journal of Applied Research in Veterinary Medicine, Washington, v.2, n.2, p. 135-141, 2004.
24. KNÖBL, T.; MORENO, A. M.; PAIXÃO, R.; GOMES, T. A. T.; VIEIRA, M. A.
M.; LEITE, D. S.; BLANCO, J. E.; FERREIRA, A. J. P. Prevalence of avian pathogenic Escherichia coli (APEC) clone harboring sfa gene in Brazil. The Cientific World Journal [online],Nova York , v. 2012, p.1-7, 2012. Disponível em : http://www.hindawi.com/journals/tswj/2012/437342/. Acesso: 3 set. 13.
25. LING, J.; PAN, H.; GAO, Q.; XIONG, L.; ZHOU, Y.; ZHANG, D.; GAO, S.; LIU,
X. Aerobactin synthesis genes iucA and iucC contribute to the pathogenicity of avian pathogenic Escherichia coli O2 strain E058. Plos one [online], São Franscico, v.8, n.2, p.1 – 10, 2013. Disponível em: http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0057794. Acessado em: 4 set. 13.
26. LLOUBES, R.; BERNADAC, A.; HOUOT, L.; POMMIER, S. Non classical
secretion systems. Research in Microbiology, Paris, v.164, n.1, p.655-663, 2013.
27. LYNNE, A. M.; KARIYAWASAM, S.; WANNEMUEHLER, Y.; JOHNSON, T. J.;
JOHNSON, S. J.; SINHA, A. S.; LYNNE, D. K.; MOON, H. W.; JORDAN, D. M.; LOGUE, G. M.; FOLEY, S. L.; NOLAN, L. K. Recombinant Iss as a Potential Vaccine for Avian Colibacillosis. Avian Diseases, Kennet Square, v.56, n.1, p.192-199, 2012.
28. MATURANA, V. G.; PACE, F.; CARLOS, C.; PIRES, M. M.; CAMPOS, T. A.; NAKAZATO, G.; STHELING, E. G.; LOGUE, C. M.; NOLAN, L. K.; SILVEIRA, W. D. Subpathotypes of avian pathogenic Escherichia coli (APEC) exist as defined by their syndromes and virulence traits. Open Microbiology Journal, Sharjah, v.5, n.55, p. 55-64, 2011.

24
29. MEHDIPOUR, S.; DOOSTI, A.; DEHKORDI, P. G. Detection of citolethal distending toxin (CDT) and cytotoxic necrotizing factor (CNF) genes among Escherichia coli isolates from Iraniam sheep carcasses. Comparative clinical pathology [online], Heidelberg, v.21, p.1683-1688, 2012. Disponível em: http://link.springer.com/content/pdf/10.1007%2Fs00580-011-1349-6.pdf. Acesso em: 5 set 2013.
30. MONROY, M. A.; KNÖBL, T.; BOTTINO, J. A.; FERREIRA, C. S.; FERREIRA, A. J. Virulence characteristics of Escherichia coli isolates obtained from broilers breeders with salpingites. Comparative Immunology, Microbiology and Infectious Diseases, Oxford, v.28, n.1, p.1-15, 2005.
31. NENNINGER, A. A.; ROBINSON, L. S.; HAMMER, N. D.; EPSTEIN, E. A.;
BADTKE, M. P.; HULTGREN, S. J.; CHAPMAN, M. R. CsgE is a curli secretion specificity factor that preventes amyloid fibre aggregation. Molecular Microbiology [online], Malden, v.81, n.2, p.486-499, 2011.. Disponível em: http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2958.2011.07706.x/pdf. Acesso em: 4 set 13.
32. NGELEKA, M.; BRERETON, L.; BROWN, G.; FAIRBROTHER, J. M.
Pathotypes of avian Escherichia coli as related to tsh-, pap-, pil-, and iuc-DNA sequences, and antibiotic sensitivy of isolates from internal tissues and the cloacae of broilers. Avian diseases, Kennet Square, v.46, n.1, p.143-152, 2002.
33. OLIVEIRA, W. F.; CARDOSO, W. M.; MARQUES, L. R,; SALLES, R. P.; C.;
AGUIAR FILHO, J.B.; TEIXEIRA, R. S. C.; ROMÃO, R. S. C.; LIMA, A. C. P. Utilização de diferentes meios de cultura para isolamento de enterobactérias em amostras fecais de frango de corte procedentes de exportação industriais do Estado do Ceará, Brasil. Revista portuguesa de ciências veterinárias, Lisboa, v.99, n.552, p.211-214, 2004.
34. QUINN, P.J.; MARKEY, B.K.; CARTER, M.E.; DONNELLY, W.J.; LEONARD,
F.C. Microbiologia Veterinária e Doenças Infecciosas. 1ª ed. Porto Alegre: editora Artmed 512p, 2005.
35. ROCHA, A. C. G. P.; SILVA, A. B.; BRITO, B. G.; MORAES, H. L.; PONTES,
A. P.; CÉ, M. C.; NASCIMENTOS, V. P.; SALLE, C. T. P. Virulence factors of avian pathogenic Escherichia coli isolated from broilers from the South of Brazil. Avian diseases, Kennet Square, v. 46, n.3, p. 749-753, 2002.
36. SAIDENBERG, A. A. B.; ALLEGRETTI, L.; ASTOLFI-FERREIRA, A. A. S.;
FERREIRA, A. J. P.; ALMEIDA, M. A.; RASO, T. F. Some virulence genes of Escherichia coli isolated from cloacal swabs of healthy Alagoas Curassows (Pauxi mitu) in Brazil. Pesquisa Veterinária Brasileira, Rio de Janeiro, v.33, n.4, p.523-527, 2013.
37. SALVARANI, S.; TRAMUTA, P.; NEBBIA, P.; ROBINO, P. Occurence and
functionality of cycle inhibiting fator, cytotoxic necrotising factors and cytolethal

25
distending toxins in Escherichia coli isolated from calves and dogs in Italy. Research in Veterinary Science, Londres, v.92, n.1, p.372-377, 2012.
38. SILVEIRA, W. D.; FERREIRA, A.; BROCCHI, M.; HOLLANDA, L. M.;
CASTRO, A. F. P.; YAMADA, A. T.; LANCELLOTTI, M. Biological characteristics and pathogenicity of avian Escherichia coli strains. Veterinary Microbiology, Amsterdan, v.85, n.1, p.47-53, 2002.
39. SZEMLAKO, K.; KRAWCZYK, B.; SAMET, A.; SLEDZIÚSKA, A.; NOWICKI, B.; NOWICKI, S.; KUR, J. A subset of two adherence systems, acute pro-inflammatory pap genes and invasion coding dra, fim, or sfa, increases the risk of Escherichia coli translocation to the bloodstream. European journal of clinical microbiology & infection diseases [on line], Wiebaden, v.1, n.1, p. 1-4, 2013. Disponível em: http://link.springer.com/article/10.1007%2Fs10096-013-1913-x. Acesso 9 set. 2013.
40. TROISFONTAINES, P.; CORNELIS, G. R. Type III secretion: more systems than you think. Physiology, Switzerland, v.20, p.326-339, 2005.
41. VIEIRA, M. A. M. Ilhas de patogenicidade. O mundo da saúde, São Paulo, v.33, n.4, p.406-414, 2009.
42. WHITT, D. D.; SALYERS, A. A. Escherichia coli extraintestinal. In:____.
Bacterial pathogenesis, as molecular approach. 2 ed. Washington: ASMPress, 2002. cap.29, p. 422-436.