biometria e o melhoramento de plantas na era da...
TRANSCRIPT

BIOMETRIA E O MELHORAMENTO DE PLANTAS NA ERA DA GENÔMICA
MAGNO ANTÔNIO PATTO RAMALHO1; EDUARDO DE SOUZA LAMBERT2
1Professor Titular, Depto de Biologia da Universidade Federal de Lavras. Caixa Postal 37, CEP. 37200-000 Lavras.MG. E-mail: [email protected] (autor para correspondência)2Doutorando em Genética e Melhoramento de Plantas, Depto de Biologia da Universidade Federal de Lavras.
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
RESUMO - O melhoramento genético começou com a domesticação de algumas espé-cies de plantas há mais de 10.000 anos e se intensificou para atender a demanda poralimentos com o crescimento populacional. Há pelo menos 85 anos, a Genética, o Me-lhoramento de Plantas e a Biometria estão intimamente associados. A união dessas áreasdo conhecimento originou a Genética Quantitativa. Pode ser facilmente comprovadoque essas ciências tiveram uma participação expressiva na produção de alimentos, ma-deira, fibras, etc., para atender as necessidades do homem no último século. Com oadvento da técnica dos marcadores moleculares e do seqüenciamento dos genomas, queiniciou a partir de 1974, vislumbrou-se a possibilidade de se realizar a seleção direta-mente no genótipo (DNA). Várias técnicas de laboratório foram implementadas para seobterem os marcadores moleculares e os processos de sequênciamento do DNA tiveramum crescimento explosivo, criando o que se denominou de era da genômica. Desenvol-veu-se a tecnologia do DNA recombinante, permitindo a transferência de genes entre asespécies, o que não era imaginado anteriormente. Alguns cientistas começaram a questi-onar a contribuição futura da Genética Quantitativa. O que se pretende mostrar com essapublicação é que todas as evidências disponíveis apontam que a sua importância, prova-velmente, será ainda maior do que no passado. As pesquisas mostraram que, especial-mente para caracteres quantitativos (QTL – quantitative traits loci), os marcadores de-vem estar associados a medidas do fenótipo e, para serem úteis, precisam ser obtidasinformações em experimentos de campo com a maior precisão experimental possível.Até mesmo para os transgênicos, necessita-se de avaliações extensivas, com o objetivode avaliar o efeito de genes exógenos no maior número de cultivares possível e emvários ambientes.Palavras-chave: melhoramento, genética quantitativa, biotecnologia, marcadoresmoleculares.
BIOMETRICS AND PLANT BREEDING IN THE GENOMICS AGE
ABSTRACT - Genetic improvement started with the domestication of some plant speciesmore than 10,000 years ago and has been intensified to meet the demands for foods withpopulation growth. Genetics, Plant Breeding and Biometrics have been closely associatedfor at least 85 years. The union of these areas of knowledge originated QuantitativeGenetics. It can be easily proved that these sciences played an important role in theproduction of foods, wood, fiber and so forth to meet men needs in the last century. Theadvent of molecular markers and genome sequencing techniques in 1974, opened up thepossibility of a selection strategy directly on the genotype (DNA). Several laboratory

229Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
techniques were implemented to obtain molecular markers and the DNA sequencingprocesses boomed and created what is called the genome era. The recombinant DNAtechnology development, allowing the transfer of genes among species that wereunimaginable previously, made some scientists begin to question the future contributionof Quantitative Genetics. The objective of this publication is to show that all the availableevidence points to the fact that Quantitative Genetics will grow more and more important.Research has shown that, especially for quantitative traits (QTLs - quantitative traitloci), the markers should be better associated to measurements of the phenotype and, tobe useful, data need to be obtained in field experiments with the greatest experimentalaccuracy. Even the GMO´s require extensive assessments of the exogenous gene in thegreatest number of cultivars and in the greatest number of environments.Key words: breeding, quantitative genetics, biothecnology, molecular markers.
O Melhoramento Genético antes de Mendele Fisher
Fehr (1987) definiu o melhoramentocomo sendo a arte e a ciência de promover omelhoramento genético das plantas. Contudo,antes das pesquisas de Mendel e Fisher, entreoutras, o melhoramento foi realizado exclusiva-mente como arte, por meio da habilidade visualdos nossos ancestrais em escolher as plantas paraos próximos plantios.
Existem mais de 350.000 espécies deplantas descritas e, dentre elas, o homem utilizaou utilizou como alimento menos de 3000, sen-do cultivadas atualmente aproximadamente 300.Ressalta-se, ainda, o fato de que apenas 15 espé-cies contribuem com mais de 90% dos alimentosconsumidos no mundo, sendo elas o arroz, trigo,milho, soja, sorgo, cevada, cana-de-açúcar, be-terraba açucareira, feijão, amendoim, batata in-glesa, batata-doce, mandioca, coco e banana(Paterniani, 2000).
Um questionamento que poderia ser fei-to é qual a razão de terem sido domesticadas cer-tas espécies de plantas e outras não. Um bomexemplo é o caso do trigo. A domesticação dessaespécie, juntamente com outros cereais de inver-no, como a cevada, o centeio e a aveia, ocorreuna região denominada de Crescente Fértil, atual
região que compreende a Jordânia, Israel, Líbano,Síria, Turquia, Iraque e Irã. Nessa região, o in-verno é ligeiramente úmido e o verão, quente eseco. Essa condição climática deve ter favoreci-do a domesticação das plantas de trigo (Allard,1999). Isso porque os grãos grandes dos ances-trais selvagens do trigo, quando comparados àssementes de outras espécies, provavelmente pro-piciavam germinação e emergência mais vigoro-sa, superando em crescimento inicial as plantascompetidoras. Além disso, como nessa região aprimavera é seca e coincidia com o momento doflorescimento, plantas com flores cleistogâmicastambém foram selecionadas. Essas flores, perma-necendo fechadas durante grande parte doflorescimento, diminuíam a dessecação dasanteras e estigmas, possibilitando maiorvingamento floral, o que contribuíam para umamaior produtividade de grãos. Deve-se conside-rar, ainda, que a menor variabilidade devido àautofecundação era favorável aos primitivos agri-cultores que, como nos dias atuais, almejavammaior uniformidade nos seus plantios.
A domesticação do trigo ocorreu aproxi-madamente há 9500 anos (Evans, 1998). Já osprimeiros relatos da chegada do trigo na Françasó ocorreram durante o século XI. Inicialmente,os franceses utilizavam para fabricação de pão, o

230
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
centeio, que era uma espécie já bem adaptada ecom boa produtividade. Entretanto, a aristocraciafrancesa preferia o pão de trigo e exigia dos cam-poneses a produção dessa gramínea, mesmo nãoestando bem adaptada às condições da França.Como o interesse dos camponeses pelo pão detrigo também aumentou, houve uma enorme pres-são para se incrementar a área cultivada. Isso es-timulou o processo seletivo, que contribuiu parao aumento da adaptação. Fato semelhante foiobservado em outros países, inclusive em regi-ões tropicais. Dessa forma, o trigo se adaptou àsmais diferentes regiões ecológicas, por meio dahabilidade visual dos agricultores para a seleção.
O caso do milho é um outro bom exem-plo. Estudos indicam que ele se originou no Mé-xico e, embora haja algumas controvérsias sobresua origem evolutiva, sabe-se que ele tem comoparente silvestre mais próximo o teosinte (Zeamays ssp. parviglumis). O teosinte é uma plantacom muitas ramificações laterais que saem de suahaste central e terminam em uma inflorescênciamasculina. Cada ramificação também possui umainflorescência feminina, que dá origem a uma es-piga pequena. O milho, por sua vez, não perfilhae tem espigas maiores. Foi demonstrado que essadiferença no perfilhamento é devido apenas a umgene, o tb1, que contribui para o incremento acen-tuado na dominância apical e, com isso, ocorre aconcentração dos fotoassimilados no caule cen-tral da planta e a supressão dos ramos axilares.Doebley et al. (1997) isolaram o gene e mostra-ram que o alelo tb1, presente no milho, se ex-pressa duas vezes mais nesta espécie do que noteosinte. Esse fato sugere que alterações no sis-tema regulatório do gene foram responsáveis peladivergência tão acentuada entre as duas espéci-es.
O fato de apresentar menor número deperfilhos e espigas maiores foi provavelmente
uma das razões da escolha do milho para adomesticação. Evidências indicam que tribos in-dígenas existentes no México iniciaram adomesticação e obtiveram algumas variedadesque se disseminaram para outras regiões da Amé-rica. Tanto foi assim, que quando CristóvãoColombo chegou à América, em 1492, já existi-am centenas de variedades de milho e ele levoualgumas amostras de milho para a Europa na oca-sião do seu retorno, após o descobrimento.
Duvick (1996) faz um interessante relatodo que ocorreu desde aquela época até o retornodo milho, já melhorado, para os Estados Unidos.Após esse acontecimento, houve aumento emprodutividade no século XIX, por meio da sele-ção nas variedades de polinização aberta, visan-do maior adaptação às condições de cultivo. Pos-teriormente, até por volta de 1930, não houvemodificações expressivas na produtividade, quese situava em torno de 2700 kg ha-1. A partir dadécada de 30, o milho híbrido passou a ser culti-vado, o que proporcionou uma verdadeira revo-lução na agricultura, não só americana, bem comona de todo o mundo.
A utilização dos híbridos tornou-se pos-sível após ter sido descoberta a possibilidade deperpetuar a heterose por meio do cruzamento daslinhagens, genótipo altamente endogâmico. Em-bora a heterose tenha sido uma das principaiscontribuições da ciência, quase um século se pas-sou desde o seu descobrimento e ainda há con-trovérsias a respeito das bases genéticas de suaocorrência (Crow, 1999).
Assim como ocorrera com o trigo, o me-lhoramento como “arte” propiciou que o milhofosse cultivado em praticamente todas as condi-ções ambientais em que é realizada a agriculturano planeta, evidenciando novamente a eficiênciada seleção. Esse mesmo fato ocorreu com gran-de parte das plantas cultivadas.

231Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
A origem da Genética Quantitativa
Mendel, um monge agostiniano, ficou nahistória por ter sido o primeiro a fornecer as basesdo controle genético dos caracteres. A publica-ção do seu trabalho ocorreu em 1865 e ficou pra-ticamente no anonimato até 1900, quando trêscientistas, Correns, De Vries e Tschermak, inde-pendentemente, mostraram que a teoria deMendel era correta. Assim, 1900 é considerado oano do nascimento da Genética.
Como Mendel trabalhou com caracteresqualitativos, isto é, aqueles que se expressam emclasses distintas, no início do século passado ima-ginava-se que a teoria não podia ser extrapoladapara os caracteres quantitativos. Essa idéia errô-nea foi gradativamente eliminada a partir de al-guns trabalhos como o de Yule, Nilsson-Ehle eEast, dentre outros. Já em 1903, um pesquisadordinamarquês, Wilhelm Johanssen, mostrou queo fenótipo de um caráter quantitativo era devidoà ação conjunta do genótipo mais o efeito do am-biente. Foi esse pesquisador que atribuiu o ter-mo gene aos fatores Mendelianos.
Também no início do século XX, come-çaram os trabalhos que mudariam a história dapesquisa agrícola, sendo Sir Ronald Fisher o prin-cipal responsável. As pessoas que o conheceramafirmam que ele era um excelente matemático,com profundo senso prático. Fisher começou seustrabalhos na Estação Experimental deRothamsted, na Inglaterra, dando origem à Esta-tística Experimental. Ele conciliou a GenéticaMendeliana e a Biometria em 1918, dando ori-gem à Genética Quantitativa, em um artigo pu-blicado na Transactions of the Royal Society ofEdinburg. Ressalta-se que essa publicação, defundamental importância para a ciência, não ti-nha sido aceita antes pela Royal Society ofLondon.
A publicação de Fisher (1918) enfocavaa covariância e a correlação genética entre indi-víduos aparentados. Com isso, foi possível de-compor a variância genética em variância aditiva,devido aos efeitos médios dos genes, a variânciade dominância, devido aos efeitos das interaçõesintralélicas e a variância epistática, devido àsinterações interalélicas. Após esse trabalho, sur-giram muitos outros, como o de Wright (1927),Cockerham (1954, 1956), Kempthorne, (1954) eSchnell (1963).
Comentando sobre os genes envolvidosno controle dos caracteres quantitativos, Mather(1949), de maneira semelhante a Fisher, dizia quedeveria estar envolvido um grande número degenes de pequeno efeito, os genes menores oumodificadores, denominados de poligenes. Des-sa maneira, os poligenes, que não podem ter seusefeitos isolados, quando associados, produzemalterações quantitativas que podem sermensuradas.
A teoria da Genética Quantitativa possi-bilitou que fosse adicionado ao melhoramentocomo arte, a ciência, como preconizado por Fehr(1987). No caso das plantas autógamas, os prin-cipais métodos de melhoramento já eram conhe-cidos antes do advento da Genética Quantitativa(Jensen, 1988). Contudo, os conhecimentos des-sa ciência permitiram avaliar os aspectos positi-vos e as deficiências de cada método e, assim,propor alternativas para melhorar a sua eficiên-cia e criar novos métodos. No caso das plantasalógamas, ficou evidenciado porque a seleçãomassal não estava sendo mais eficiente. Demons-trou-se a importância da seleção recorrente, so-bretudo após ter sido verificada a predominânciada variância aditiva no controle dos caracteres.Ainda, tornou-se claro que só por meio da sele-ção recorrente poder-se-ia ter aumento da fre-qüência dos alelos favoráveis e, por conseguinte,

232
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
incremento na expressão média do caráter sobseleção. Muitas outras informações importantespuderam ser obtidas e são relatadas em algumaspublicações (Hallauer & Miranda Filho, 1982;Falconer & Mackay, 1996; Wricke & Weber,1996; Kearsey & Pooni 1998).
O Melhoramento Genético convencionalapós o advento da Genética Quantitativa
Durante o século XX, a população do pla-neta passou de 1,5 bilhão de habitantes para 6,2bilhões. Esse espetacular incremento exerceu umaforte pressão sobre a necessidade de aumentar aprodutividade das principais espécies cultivadas.Efetivamente, esse fato ocorreu e foi uma dasmaiores contribuições da ciência para a socieda-de. O aumento em produtividade deveu-se, emparte, à melhoria do manejo, envolvendo o in-tensivo emprego de fertilizantes, especialmentenitrogenados; melhor controle de pragas, doen-ças e plantas daninhas (Cardwell, 1982). Mas,sem dúvida, o emprego de cultivares melhoradasteve uma expressiva participação.
São inúmeros os trabalhos que mostramo progresso genético obtido em várias espécies,dentre eles nos EUA (Wilcox, 2001), na Europa(Berzonsky & Lafever, 1993) e no Brasil(Vencovsky & Ramalho, 2000). Esse progressofoi devido à utilização da heterose, no caso dasalógamas, e também aos contínuos ciclos seleti-vos a que essas espécies cultivadas foram sub-metidas. Embora alguns pesquisadores insistamem afirmar que, atualmente, a produtividade dasprincipais espécies cultivadas atingiu um limitee não está ocorrendo ganho adicional, os dadosdisponíveis apontam exatamente em direção con-trária (Wilcox, 2001). Inclusive, tem sido mos-trado que existem alternativas para gerar varia-bilidade que antes eram desconhecidas(Rasmusson & Phillips, 1997). É certo que, devido
ao nível de produtividade atingido, as diferençasa serem detectadas exigem que os experimentossejam cada vez mais precisos, ou seja, é necessá-rio realizar o melhoramento com mais ciência doque arte.
O Melhoramento na Era da Genômica
A natureza química do material genéticocomeçou a ser elucidada por meio do trabalho deR. Griffith, em 1928, quando ele mencionou aexistência do que foi denominado de princípiotransformante. Mais tarde, em 1944, três pesqui-sadores, Avery, MacLeod e McCarty demonstra-ram que a substância responsável pela transmis-são da informação entre indivíduos e células erao DNA. A estrutura da molécula foi estabelecidaem 1953, por Watson e Crick, em um trabalhoque foi a base de todo o desenvolvimento da Bi-ologia Molecular.
A descoberta das enzimas de restrição,que cortam o DNA em pontos específicos, foifeita por Arber, em 1968. Esse foi um marco im-portante na transformação genética, sendo quelogo depois, em 1973, Stanley, Cohen e Browncombinaram uma enzima de restrição com oplasmídeo de Agrobacterium, para isolar um genee introduzí-lo em outro organismo. A manipula-ção do material genético começou efetivamenteem 1972, na Universidade Stanford, nos EstadosUnidos, quando o Dr. Paul Berg conseguiu unir aseqüência do DNA de Escherichia coli à do ví-rus Simian papiloma. Esse feito pode ser consi-derado o marco zero da denominada “era dagenômica”. Todas essas técnicas e muitas outrasnão comentadas têm levado a um melhor conhe-cimento da ação gênica. Atualmente, já são co-nhecidos mais detalhes da atuação dos genes emrelação ao que se dispunha há cinqüenta anos,quando a estrutura do DNA foi estabelecida.Sabe-se, hoje, por exemplo, que o gene depende

233Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
de regiões reguladoras situadas em várias posi-ções nos cromossomos. Além disso, possui umsítio promotor, que é uma região do DNA próxi-ma ao gene propriamente dito, onde se prende aRNA polimerase, enzima responsável pela trans-crição da informação. O gene estrutural, por suavez, pode ser dividido em duas partes, os íntronse éxons. A função dos íntrons ainda não está bemelucidada, porém acredita-se que os mesmos te-nham papel na regulação gênica. Já os éxons pos-suem a informação genética que dará origem aoRNA mensageiro. Esses comentários são indis-pensáveis no contexto dos marcadoresmoleculares, porque os diferentes alelos de umgene podem surgir por alterações em qualqueruma de suas partes constituintes e não apenas noseu exon. Glazier et al. (2002) discutem essesaspectos para vários genes em plantas e animaise apresentam uma relação de mutantes em várioscasos. Comentam, por exemplo, que em um dosgenes para o tamanho dos frutos em tomate, oOFRX, ocorre uma alteração no sítio promotor.Já na mesma espécie, a variação no conteúdo deaçúcar de frutos, resulta de uma alteração nosintrons do gene Lin 5. Em arroz, a sensibilidadeao fotoperíodo é devido à substituição de umadas bases na posição 91 do gene CK2α, no códonque codifica a lisina, dando origem a um pontofinal. Resta comentar, também, que apenas umaínfima percentagem do total de bases que com-põem o DNA de uma determinada espécie é efe-tivamente transcrita.
Entre as várias ferramentas de manipula-ção do DNA que foram desenvolvidas, a dosmarcadores moleculares foi uma das que maischamou atenção no melhoramento de plantas,sobretudo em função da possibilidade de identi-ficar-se os indivíduos superiores diretamente peloseu DNA. Nesta publicação não serão relatadasas diferentes técnicas moleculares existentes, queestão disponíveis em várias literaturas (Lander
& Botstein, 1989; Ferreira & Grattapaglia, 1998;Kumar, 1999; Lanza et al., 2000). Também nãoserão comentadas todas as possibilidades do em-prego de marcadores, que são encontradas emvárias publicações (Paterson et al. 1991; Stuber,1992; Lee, 1995). O que se deseja comentar é acontribuição da Biologia Molecular nos últimostrinta anos no melhoramento genético decaracteres quantitativos.
Marcadores Moleculares
1- Escolha de GenitoresEm qualquer espécie cultivada, o
germoplasma disponível possui milhares de li-nhagens e/ou cultivares. Em plantas autógamas,por exemplo, as possibilidades de combinaçãodesse germoplasma por hibridação são infinitas.Por isso, a escolha dos genitores a serem cruza-dos para obtenção da população segregante, paraser submetida à seleção, tem despertado a aten-ção dos geneticistas há muito tempo. SegundoBaezinger & Peterson (1991), as metodologiasdisponíveis podem ser agrupadas em duascategorais: aquelas que consideram o comporta-mento dos genitores per se e aquelas que envol-vem o desempenho de suas progênies. Entre asque utilizam o comportamento per se dosgenitores, estão as que envolvem a média e me-didas de divergência genética, tais como, coefi-ciente de parentesco, análise multivariada e osmarcadores moleculares. Quanto ao comporta-mento das progênies, os métodos que têm sidomais utilizados são os cruzamentos dialélicos, asestimativas das médias das n linhagens nageração (m + a) e a probabilidade de se obterlinhagens que superem um determinado padrãona geração (Jinks & Pooni, 1976). Na litera-tura, há inúmeros relatos do emprego de todasessas metodologias (Abreu, 1997; Souza Sobrinho,2001; Santos, 2000; Mendonça, 2001; Carneiro,2002).

234
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
Uma boa população para o melhoramen-to é aquela que associa média alta e a maiorvariabilidade possível para o caráter em ques-tão. Os que advogam o emprego de medidas dedivergência, especialmente a partir de marcadoresmoleculares, têm como princípio que, quantomaior a variabilidade, maior a probabilidade deencontrar um genótipo com bom desempenho.Entretanto, os melhoristas convencionais nãodesejam muita variabilidade. O seguinte exem-plo ilustra o que se pretende evidenciar. Supon-do que se deseje combinar em uma linhagem, 26locos com alelos favoráveis, sendo estes repre-sentados pelas letras maiúsculas. A finalidade é,portanto, obter-se uma linhagem que possua aconstituição AABB ... ZZ. Em uma primeira si-tuação, pode-se considerar dois genitores que secomplementam em todos os locos da maneiramostrada abaixo:
L1 L2
AA bb CC DD ... zz aa BB cc dd ... ZZEm todos os locos em que a L1 tem alelos
favoráveis, a L2 possui alelos desfavoráveis evice-versa. Desse modo, na geração F1 tem-se:Aa Bb Cc Dd ... Zz. Na geração F2, a segregaçãoé máxima e o indivíduo com todos alelos favorá-veis ocorre na freqüência de (¼)26. Na, que éa geração de interesse, a possibilidade de se ob-ter a linhagem AA BB CC DD ... ZZ, será de (½)26, isto é, uma em 67.108.864 linhagens.
Seja agora as linhagens L3 e L4, que este-jam segregando para apenas 13 locos, uma vezque elas são linhagens elites e têm 13 dos 26 locosjá fixados com alelos favoráveis:
L3 L4
AA bb CC DD ... zz AA BB CC dd ZZA F1 terá a constituição AA Bb CC Dd...
ZZ, segregando em 13 locos. A probabilidade dese obter na a linhagem desejada, nesse caso, será
de (½ )13 =. Uma probabilidade muito superior àanterior. É por essa razão que os melhoristas commaior experiência têm como prática sempre cruzar“bom x bom” (Rasmusson & Phillips, 1997;Dudley, 1997), sem se preocupar com variabili-dade em excesso. O princípio para isso é que omelhoramento deve ser uma acumulação de van-tagens.
Pelo exposto, é difícil decidir qual genitorcruzar, utilizando apenas a medida de diversida-de. Quando se deseja a produtividade de grãos,mas se escolhem os pais por meio de marcadoresmoleculares, é utilizada toda a divergência exis-tente. Os pais podem divergir para várioscaracteres e não apenas para a produtividade. Éprincipalmente por essa razão que, nas várias si-tuações em que se utilizaram marcadoresmoleculares para a escolha dos pais, os resulta-dos não foram tão promissores (Machado 2000;Mendonça, 2001). No futuro, quando estiveremdisponíveis marcadores moleculares específicospara o caráter sob seleção, a medida de divergên-cia poderá ser útil, se associada a dados de de-sempenho médio dos pais.
2- Teste de identidade genética para a prote-ção de cultivares
Com a lei de proteção de cultivares, fi-cou estabelecido que essas devem ser distintas,homogêneas e estáveis. Por isso, é exigida umadescrição detalhada, provando que a nova culti-var é distinta das pré-existentes. É fácil entenderque, para a maioria das espécies em que se dis-põe de alguns programas de melhoramento, como tempo, os descritores serão insuficientes paradiscriminar as novas cultivares. Uma alternativaé o emprego de marcadores moleculares com essafinalidade, o que se denomina de impressão di-gital do DNA (“DNA fingerprint”). Entre algu-mas das vantagens dessas metodologias, Ferreira

235Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
(2003) cita: a) independência da interaçãogenótipos x ambientes, o que não ocorre com osmarcadores morfológicos; b) o alto polimorfismono DNA, especialmente em regiões demicrossatélites; c) o baixo custo da genotipagem;d) o menor tempo requerido na análise do DNAem relação à análise fenotípica, para um grandenúmero de espécies vegetais.
Contudo, Troyer & Rocheford (2002),utilizando como referência a cultura do milho nosEUA, mostraram exemplos de que um granderelacionamento entre linhagens ou híbridos, ba-seados em marcadores moleculares, não exclu-em diferenças expressivas no desempenho agro-nômico. Em um dos exemplos, foram avaliados,em 3059 experimentos, dois híbridos simples,ambos obtidos a partir de uma linhagem comuma eles, a LH51, cruzada com outras duas linha-gens, a B73, para obtenção de um dos híbridos ea LH119, para o outro. Além disso, LH119 é de-rivada de um híbrido em que uma das linhagensé a B73, sendo, então, as duas linhagens muitorelacionadas. Foi verificado que elas possuem88,2% de similaridade genética, utilizandomarcadores RFLP. Observa-se, na Tabela 1, queos dois híbridos apresentaram a mesma produti-vidade de grãos, porém o híbrido B73-LH51 apre-sentou florescimento mais precoce, altura da plan-ta 3% menor e de espiga, 9% menor. O outro hí-brido foi 5% mais vigoroso na primavera, 15%superior em ‘stay green’, com redução de 18%em plantas quebradas. Assim, o híbrido LH119 xLH51 foi recomendado para regiões com maiorquantidade de chuvas durante o cultivo. Demons-trou-se também que a linhagem LH119 foi maistolerante ao calor e à seca do que a B73, noflorescimento.
Os autores salientam que, dependendo decomo essa tecnologia for empregada, com re-gras muito rígidas na identificação de linhagens
essencialmente derivadas, pode haver sérios pre-juízos ao futuro do melhoramento genético.
No Brasil, ainda não foi regulamentadoo emprego de marcadores moleculares comodescritores de cultivares a serem protegidas. NosEUA, é permitido, mas há controvérsia comrelação ao nível de similaridade a ser adotado.Smith & Smith’s (1989) propuseram o nível de75% de similaridade do genoma, obtido por meiode marcadores moleculares, como indicador deuma cultivar essencialmente derivada de outra.Se adotada essa proporção, a linhagem LH119seria considerada similar à B73, e o obtentor de-veria pagar royalties ao detentor da linhagem B73.Segundo Troyer & Rocheford (2002), se adotadoesse nível de similaridade, possivelmente linha-gens, como a LH119, que trouxeram vantagensaos agricultores, seriam pouco ou não utilizadasna produção de híbridos comerciais.
Esse não é um problema exclusivo dosmarcadores moleculares, mas sim de todos osdescritores utilizados para a proteção de cultiva-res. Entretanto, é preciso que o assunto seja am-plamente debatido, para não ser mais um entraveaos programas de melhoramento no Brasil. Istoporque as melhores linhagens ou cultivares, ouseja, aquelas que apresentam maior adaptação emvárias condições, certamente terão muitos locoscom constituições genéticas semelhantes.
3- Seleção assistidaO princípio da seleção assistida por
marcadores é a correlação genética entre a marcae os diferentes genes envolvidos no controle docaráter. A correlação genética pode ocorrer devi-do à pleiotropia e à ligação. A pleiotropia ocorrequando um gene afeta mais de um caráter aomesmo tempo. No caso dos marcadores, seriapreciso que a marca estivesse dentro do gene, demodo que eles estariam sempre juntos. Mesmopara um caráter monogênico, essa possibilidadeé muito pequena, e, para um caráter quantitativo,

236
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
é impossível que isso ocorra. Assim, espera-seque os diferentes marcadores estejam ligados comos locos que contenham o alelo de interesse.
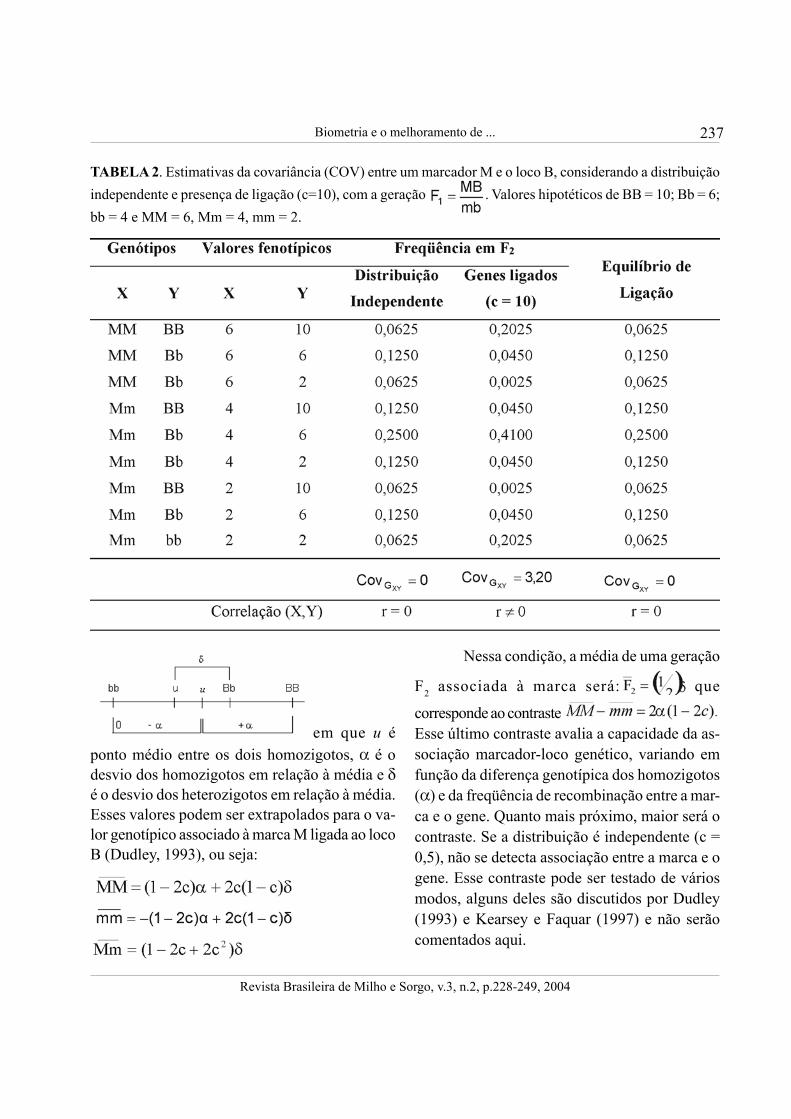
O princípio da ligação, que foi estabele-cido por um brilhante geneticista, Thomas HuntMorgan, em 1922, é o fundamento da seleçãoassistida por marcadores. É preciso salientar quea correlação devido à ligação não é permanente,ocorrendo somente quando há desequilíbrio deligação. Para exemplificar, considere doiscaracteres, o caráter X representado por M e ocaráter Y, representado pelo gene B. Na tabela 2,é mostrado o que ocorre na geração F2, quandoM e B não estão ligados e quando estão ligadoscom uma freqüência de recombinação (c) de 10%.É mostrado também o que irá ocorrer na presen-ça de ligação quando o equilíbrio for atingido.Veja que, quando a distribuição é independente,a covariância entre X e Y é zero, e, evidentemen-te, a correlação entre eles também será nula. Por-tanto, numa situação como essa, se M fosse uma
marca, ela não teria utilidade. Ocorrendo liga-ção, a geração F2 não está em equilíbrio, pois asfreqüências alélicas e genotípicas serão alteradascom o cruzamento ao acaso. Verifica-se que, nes-sa situação, como c = 10, a covariância entre MBé diferente de zero, e, portanto, a marca (M) podeser utilizada na identificação do gene B. Com odecorrer do intercruzamento, a população irá atin-gir o equilíbrio, mesmo com a seleção. Nota-se,ainda, na Tabela 2, que a covariância passa a sernula e a mesma observação feita para a distribui-ção independente é válida com a ligação. Esse éum ponto fundamental que, muitas vezes, passadespercebido pelos geneticistas.A teoria da seleção assistida por marcadores érelatada em algumas literaturas (Lande &Thompson, 1990; Stuber, 1992; Dudley, 1993;Bearzoti, 1997) e um sumário do que se encon-trou será comentado a seguir. O modelo mostra-do abaixo é usado para representar o valorgenotípico do loco B:
TABELA 1. Comparação entre as linhagens B73 e LH119 em combinação híbrida com a linhagem LH51.Resultados médios de avaliações realizadas em 3059 experimentos.
Fonte: adaptado de Troyer & Rocheford (2002).
237Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
em que u éponto médio entre os dois homozigotos, α é odesvio dos homozigotos em relação à média e δé o desvio dos heterozigotos em relação à média.Esses valores podem ser extrapolados para o va-lor genotípico associado à marca M ligada ao locoB (Dudley, 1993), ou seja:
TABELA 2. Estimativas da covariância (COV) entre um marcador M e o loco B, considerando a distribuiçãoindependente e presença de ligação (c=10), com a geração . Valores hipotéticos de BB = 10; Bb = 6;bb = 4 e MM = 6, Mm = 4, mm = 2.
Nessa condição, a média de uma geração
F2 associada à marca será: que
corresponde ao contraste Esse último contraste avalia a capacidade da as-sociação marcador-loco genético, variando emfunção da diferença genotípica dos homozigotos(α) e da freqüência de recombinação entre a mar-ca e o gene. Quanto mais próximo, maior será ocontraste. Se a distribuição é independente (c =0,5), não se detecta associação entre a marca e ogene. Esse contraste pode ser testado de váriosmodos, alguns deles são discutidos por Dudley(1993) e Kearsey e Faquar (1997) e não serãocomentados aqui.

238
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
fenotípica será: . E
como , pode-se escrever:
, e associado
ao marcador tem-se: Considerando iM
= iF e cM = cF, tem-se: . Fica evidente que o
ganho, utilizando o marcador, será superior à se-leção fenotípica quando p > h2.
Como foi visto, a seleção assistida pormarcadores será mais eficiente quando a propor-ção da variância genética aditiva explicada pelomarcador for maior que a herdabilidade. Então,ela só seria apropriada para caracteres de baixaherdabilidade, normalmente h2 inferior a 0,3.Contudo, para que o p seja confiável, ele tem queser obtido a partir de experimentos bem condu-zidos, em que a h2 necessariamente seja alta.Depreende-se que o benefício máximo da sele-ção assistida por marcadores de QTLs(quantitative traits loci) seria aquela praticada sobcondições de alta h2 e quando a característica deinteresse é difícil de ser medida (Dudley, 1993).
Tem ocorrido um acúmulo de informa-ções a respeito dos QTLs. Mackay (2001) discu-te alguns resultados com Drosophila, verifican-do que o tamanho médio do intervalo, contendoum QTL foi de 8,9 cM. Isso corresponde a umaregião do DNA, contendo 4459 Kb, sendo que avariação de tamanho foi de 0,1 a 44,7 cM. Comoo genoma da Drosophila contém 120Mb, esti-ma-se que ela possua 13600 genes, com tama-nho médio de 8,8 Kb. Assim, em um QTL médio
A variância genética associada aosmarcadores ( ) é fornecida por
e a variância genética não explicada pelo marcadoré:
em que: é a variância genética aditiva e éa variância de dominância (Souza Júnior, 1992).Se c = 0, toda variância é explicada pelomarcador, isto é, .Por outro lado, se c = ½,
. Pode-se, então, tera proporção da variância aditiva explicada pelo
marcador pela expressão . Ela é li-
nearmente associada ao coeficiente derecombinação c, ou seja, 1 ou 100% com c = 0 e0 quando c = 0,5.
A partir da expressão do ganho pela sele-ção fenotípica (GSF), utilizando indivíduos oufamílias, pode-se inferir sobre a eficiência da se-leção assistida por marcadores. A expressão doganho é , em que iF é a intensi-dade da seleção, cF é o controle parental, isto é,½ se a seleção ocorre após a polinização, 1,0 seantes e 2 se a recombinação é realizada comclones ou S1, é o desvio padrão fenotípicoda unidade seletiva e h2 a herdabilidade do cará-ter, considerando que, quando c = 0 ou muito pró-ximo de zero, o marcador explicará a maior par-te da variação e a herdabilidade associada aomarcador será 1. Assim, a equação do ganho coma seleção, utilizando o marcador, passa a ser
. Desse modo, a eficiência daseleção, utilizando o marcador em relação à

239Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
estão incluídos 507 genes, que podem variar de11 a 2191 genes. Como se observa, mesmo paraessa espécie, ainda está distante a possibilidadede utilizar o QTL efetivamente na seleção. Res-salta-se que o QTL não é um loco ou conjunto delocos propriamente dito, mas, sim, segmentos emdiferentes regiões dos cromossomos, associadosa outros genes que afetam o caráter quantitativo.Alguns trabalhos em que se utilizou somente aseleção fenotípica reforçam esse fato. Um dostrabalhos de maior duração de que se tem infor-mação é o que está sendo realizado para seleçãoem milho para teor de óleo e proteína (Dudley &Lambert, 1992). Já foram realizados mais de 100ciclos seletivos com resposta contínua à seleção.Dudley (1977) fez uma análise detalhada dos re-sultados com 76 ciclos seletivos. A populaçãooriginal possuía 1,09 mg/g de proteína no grão,passando para 2,89 mg/g por meio da seleção nosentido positivo (aumento do teor de proteína) ea 0,4 mg/g, com a seleção no sentido negativo.Houve um incremento de 165% no teor de prote-ína, que representou um ganho de 2,17% por ci-clo seletivo (Dudley & Lambert, 1992). Os auto-res comentam que a resposta contínua à seleçãosó seria possível se o caráter fosse controlado porum grande número de genes de pequeno efeito(poligenes ou modificadores). Utilizando algu-mas informações de genética quantitativa, eleestimou que o número de genes para esse caráterestá próximo de 200.
Posteriormente, Lynch & Walsh (1998)relataram resultados de marcadores molecularesassociados a esse caráter. Para isto, foram cruza-das as duas populações com alto e baixo teor deproteína, após 76 gerações. Desse cruzamento,foram avaliadas 100 famílias F2:3, procurando co-brir todo o genoma, utilizando-se marcadoresmoleculares espaçados de 20 cM. Foram encon-tradas 22 marcadores, distribuídos em todos os
10 cromossomos para o caráter. Contudo, ape-nas 6 marcadores explicaram 65% da variação.Segundo os autores, esse resultado pode ter ocorri-do devido ao pequeno poder dos testes, resultandoem uma superestimativa dos efeitos dosmarcadores. Apesar disso, esses resultados sãocoerentes com a proposição de Mather (1949),que sugeriu estarem os poligenes organizados noscromossomos em blocos gênicos. Nesses blocos,os vários locos envolvidos na expressão do cará-ter estariam próximos um dos outros, como porexemplo, + + - - - + - ++ -, em que + representaos locos com alelos favoráveis e - os desfavorá-veis. A liberação de variabilidade seria gradual,por meio da recombinação. Desse modo, seriaexplicada a resposta à seleção em longo prazo,observada para esses caracteres. Assim, ummarcador de QTL estaria ligado ao bloco gênico,sendo essa a razão de se encontrar um pequenonúmero de marcadores que explica grande parteda variação. Numa situação como essa, osmarcadores associados ao QTL seriam de peque-na utilidade.
Apesar de todos os avanços, ainda per-siste a incerteza sobre a natureza do controle doscaracteres quantitativos, a mesma dúvida que ti-nha Fisher, Mather e Comstock há mais de 60anos. O primeiro seqüenciamento completo deum genoma vegetal foi o da Arabidopsis. Nessaespécie, foi detectada a presença de 25500 genes,sendo que 45% deles estão em 4 ou mais cópias(Walsh, 2001). No caso do milho, tem sido cons-tatado também que mais de 1/3 do genoma re-presenta várias cópias repetidas de genes. Prova-velmente, essas cópias podem ser os genes can-didatos aos caracteres quantitativos, os poligenes.Tudo indica que o tamanho dos QTLs disponí-veis seja insuficiente para se promoverem os gan-hos esperados com a seleção assistida. Contudo,o acúmulo de informações a esse respeito deverá

240
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
propiciar melhor entendimento do controle ge-nético dos caracteres quantitativos.
4- Seleção assistida por marcadores noretrocruzamento
O retrocruzamento é um método ampla-mente utilizado quando se tem uma boa linha-gem, mas que é deficiente em uma ou poucas ca-racterísticas. Ele foi utilizado por Briggs por voltade 1922, visando a obtenção de linhagens de tri-go resistente a patógenos. Já, naquele momento,ele enfatizou um método perfeito, pois omelhorista pode prever o resultado a cadaretrocruzamento efetuado (Allard, 1999). Seja,por exemplo, uma linhagem boa (P1), que foi cru-zada com uma linhagem (P2) que possui um aleloque pode solucionar um problema no P1. Essegenitor (P1) é denominado de recorrente. A gera-ção F1 é novamente cruzada com o P1, obtendo-se o primeiro retrocruzamento (RC1). Nessa con-dição, em média, 75% dos alelos já serão do pairecorrente. No segundo retrocruzamento (RC2),esse número passa para 87,5%. Com xretrocruzamentos, a proporção de recuperação do
genitor recorrente é fornecida por . Veja
que a recuperação do pai recorrente ocorre demodo previsível e rapidamente. Na prática, sãoutilizados de 5 a 6 retrocruzamentos.
Uma das possibilidades de se utilizar aseleção assistida foi exatamente no método doretrocruzamento (Tanksley et al., 1989). Osmarcadores podem ser utilizados de dois modos(Hospital & Charcosset, 1997): a) para acelerar arecuperação dos alelos do genitor recorrente queestão distribuídos em todos os cromossomos eb) para auxiliar na eliminação dos alelos dogenitor não recorrente que estão ligados com o(s)loco(s) de interesse para os melhoristas.
Inicialmente, será enfatizado o primeirocaso, isto é, para acelerar a recuperação dos alelos
do genitor recorrente. Como já foi mencionado,o que se tem, a cada geração de retrocruzamento,é a freqüência média de recuperação. Como hávariação, por meio de emprego de marcadorespode-se identificar os indivíduos que mais seaproximam do genitor recorrente. Já foram reali-zados vários estudos teóricos que comprovam asua eficiência (Hospital et al., 1992; Frisch et al.,1999; Frisch & Melchinger, 2001). É importanteque se diga que as comparações foram efetuadasconsiderando que os melhoristas não realizamnenhuma seleção. Quando os dois genitores sãobem diferentes, a seleção que o melhorista reali-za também acelera o processo. Essa seleção serátanto mais eficiente quanto maior for o tamanhoda população. Nesse contexto, a seleção assisti-da proporciona alguma vantagem, mas, depen-dendo do custo do emprego dos marcadores, podenão compensar o investimento efetuado, princi-palmente se o melhorista realiza a seleção.
Infelizmente, não existem muitos resul-tados, na literatura, de situações em que osmarcadores foram efetivamente utilizados. Emuma das poucas referências encontradas, foi coma cultura do milho para transferência de genes dealto teor proteína (QPM) entre linhagens de mi-lho (Duarte, 2003). Ele considerou duas situa-ções, sendo que em uma delas a recuperação doparental recorrente foi de 86,5% e na segunda,de 85,5% após 2 retrocruzamentos, valores quesão muito próximos do esperado no RC2, que éde 87,5%. O autor concluiu que o uso domarcador não proporcionou a vantagem almejada.
O grande problema do retrocruzamentoé quando o loco de interesse é ligado próximo aoutro cujo fenótipo do genitor não recorrente éindesejável, que é a segunda situação menciona-da anteriormente. Com isso, a transferência dogene de interesse leva consigo outro(s)indesejável(veis). Esse fenômeno é conhecido por

241Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
‘linkage drag’. Segundo Souza (2001), a seleçãoassistida pode reduzir ou eliminar esse problemapor meio da identificação de recombinantes,utilizando os marcadores moleculares.
McClean (2003) sugere, após o primeiroretrocruzamento, utilizar um marcador a 1 cMdo gene de interesse. Aquelas linhas, em que ocor-re a recombinação, poderiam ser selecionadas.Em seguida, seriam novamente retrocruzadas e adescendência submetido a um novo marcador de1 cM no outro lado do gene. Assim, em somenteduas gerações a quantidade de DNA indesejávelseria reduzida a 2 cM. Como o próprio autorenfatiza, a dificuldade está em identificar essesdois marcadores tão próximos do gene de inte-resse. Além do mais, para se ter o recombinante,como já mencionado, a descendência deve ter umgrande número de indivíduos. Isso é factível paraalgumas espécies, mas para outras não, como ofeijão e a soja, em que os cruzamentos são difí-ceis e com poucos descendentes por cruzamen-to. Nesse contexto, é preciso salientar que, se hádisponibilidade de um grande número de descen-dentes, o melhorista pode realizar a seleçãofenotípica, porque, no caso de genes ligados comefeitos indesejáveis, a diferenciação fenotípica énormalmente fácil.
Plantas transgênicas no melhoramento
Os trabalhos de biologia molecular pro-piciaram o desenvolvimento de uma nova e im-portante ferramenta, que possibilita a introduçãode genes por meio não sexual. Com isso, tornou-se possível a transferência de genes entre espéci-es não relacionadas, como, por exemplo, de umabactéria para uma planta, ou do homem para bac-térias e muitas outras combinações. Surgiu, as-sim, a possibilidade de se criar variabilidade pormeio antes não imaginado. Essa nova tecnologiatem apresentado uma repercussão muito grande,com amplo envolvimento da mídia, gerando, até
mesmo, grupos bem distintos: aqueles que a de-fendem e os tradicionalmente contra a tecnologia.
Deixando de lado os aspectos mais ideo-lógicos do que científicos, o que se pretendemostrar é que, mesmo utilizando os transgênicos,ainda há um enorme envolvimento do melhora-mento tradicional e, conseqüentemente, tambémda biometria. Para ilustrar, vamos utilizar o exem-plo de maior sucesso até o momento, que foi aobtenção da soja resistente ao herbicida glifosato.O modo de ação do herbicida e como foi realiza-da a construção gênica são apresentados em de-talhe por Padgette et al. (1995). O importante émencionar que a incorporação realizou-se pormeio de aceleração de partículas – a biobalística,e, portanto, aleatoriamente inserido no DNA dasoja. Para a transferência, utilizou-se a linhagemA5403 e uma marca de seleção para identificaras células transformadas, as quais deram origemàs plantas com o provável gene de interesse. Nototal, foram obtidas 316 plantas e, na primeirageração, após a transformação (R0), foramidentificadas 40 linhas promissoras, avaliadas emcasa de vegetação (Figura 1). Uma das linhas, a40-3, não mostrou nenhum dano quando pulve-rizada com o glifosato e foi selecionada. Essaplanta foi representada na geração R2 por apenas4 famílias e as sementes das famílias foram ava-liadas em um ou dois locais, dependendo da dis-ponibilidade de sementes. Dessas linhas, desta-cou-se a 40-3-2, que foi retrocruzada para intro-duzir o gene em outras linhagens. Além disso,ela ainda foi avaliada em vários ambientes poralgumas gerações.
Como se observa, o processo se asseme-lha ao método genealógico, usualmente utiliza-do no melhoramento convencional para identifi-car-se uma linhagem desejável. Além do mais,foi necessária intensiva avaliação em vários lo-cais. Mesmo quando o gene foi transferido paraoutras linhagens, por meio de retrocruzamentos,

242
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
FIGURA 1. Esquema utilizado para obtenção do primeiro transgênico de soja resistente ao glifosato, a partirda identificação de uma linhagem promissora. Adaptado de Padgette et al. (1995).
houve necessidade de inúmeros testes para veri-ficar se outros caracteres da planta não foram afe-tados.
Algumas publicações têm procurado es-tabelecer paralelos entre o melhoramento conven-cional e por meio de transgênicos (Prakash, 2001;
Gepts, 2002). No melhoramento tradicional, nor-malmente são manuseados milhares de genessimultaneamente, e nos transgênicos, um ou pou-cos genes. Uma outra diferença apresentada porGepts (2002) é que os transgênicos atuais envol-vem um ganho de função com o gene inserido,

243Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
ao passo que no convencional, quase sempre háperda de função. O referido autor justificou suaproposição baseada no fato de que muitos alelosimportantes são recessivos, ou seja, não produ-zem a enzima ou alteram a estrutura da proteína,de modo que ela não funcione. Por outro lado,nos transgênicos é produzida uma nova proteínaque irá conferir resistência aos insetos ou tole-rância aos herbicidas, por exemplo. Contudo, háinúmeras exceções como alelos importantes nomelhoramento convencional que são dominan-tes e casos de transgênicos em que se procuraimpedir a síntese de alguma proteína da plantatransformada.
Não resta dúvida de que os transgênicospoderão dar uma importante contribuição para asociedade. Um questionamento que surge é que,até o momento, foram introduzidos nas plantasgenes que, pelo menos aparentemente, são sele-tivamente neutros. Ou seja, para a planta, o fatode ser resistente ou não aos herbicidas, por exem-plo, deve ter pequena importância na sua adapta-ção. Já quando estão envolvidas alterações emcaracteres controlados por mais de um gene, emenzimas de várias rotas metabólicas e que repre-sentam o resultado de milhares de anos de evo-lução, é provável que as dificuldades sejam bemmaiores. Há várias tentativas a esse respeito,como plantas com redução de lignina, maior to-lerância à seca e ao frio, etc. Só o tempo poderámostrar se os transgênicos serão eficientes tam-bém nesse caso. No entanto, até os resultadosaparecerem deve-se continuar utilizando os mé-todos tradicionais, que embora não produzammudanças com grande repercussão em curto pra-zo, mostraram-se capazes, em médio ou longoprazo, de moldarem as plantas com a seleção paraatender aos anseios dos homens. No Brasil, porexemplo, enquanto a discussão sobre o comér-cio ou não de transgênicos da soja se prolonga
por mais de 5 anos, os melhoristas prosseguiramcom a seleção. Nesses, trabalhos foram obtidasoutras linhagens resistentes ao cancro da haste eao nematóide do cisto, apenas para citar algumas,com importância superior à dos transgênicos parao agronegócio da soja.
Seqüenciamento do Genoma
O seqüenciamento do Genoma tem porobjetivo conhecer toda a seqüência de bases doDNA de uma determinada espécie. O que pare-cia ser um horizonte distante há pouco mais de10 anos, tornou-se uma realidade. Osseqüenciadores automáticos disponíveis atual-mente são capazes de seqüenciar alguns milhõesde bases por dia. Como já mencionado, a primei-ra espécie vegetal que teve o genoma completa-mente decifrado foi a Arabidopsis thaliana, quepossui 120 Mpb. A ênfase atual está sendodirecionada ao conhecimento de seqüência com-pleta do genoma estrutural (EST: “ExpressedSequence Tag”) de algumas espécies cultivadascomo o arroz, cana-de-açúcar e café, dentre ou-tras.
Muito se espera dos resultados a seremobtidos com o conhecimento detalhado dogenoma. A quantidade de recursos humanos e fi-nanceiros aplicados a esses estudos é enorme emtodos os países. Os mais otimistas, como Ferreira(2003), comentam que não será surpresa se esseconhecimento proporcionar uma verdadeira re-volução no melhoramento genético. Há que seressaltar que, embora os equipamentos deseqüenciamento sejam cada vez mais rápidos, ograu de complexidade envolvido é enorme. Émencionado, por exemplo, que o genoma do ho-mem e do macaco possui 99% de homologia. Emprincípio, isso mostra que evolutivamente elessão muito semelhantes como as teorias já previam.Contudo, considerando que o genoma humano

244
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
possui 5,81 x109 pares de bases na célula diplóide,um por cento do genoma ainda representa 58,1milhões de bases. Considerando que muitos alelosnovos surgem por alteração em uma ou poucasbases, pode-se inferir que ainda há diferençasgigantescas em vários genes que compõem essasespécies.
Alguns estudos com simulação têm sidodesenvolvidos para se verificar o potencial dagenômica em auxiliar o melhorista na seleção.Bernardo (2001), utilizando o BLUP (“Best Li-near Unbiased Predictors”), mostrou que a infor-mação do genoma apresentou limitado potencialpara auxiliar na seleção de caracteres quantitati-vos. Deve-se considerar, ainda, que a quantidadede informação a ser manuseada é enorme e, mes-mo com o desenvolvimento da Bioinformática,essa será a limitação mais séria. Por outro lado, éesperado um maior aprofundamento no conheci-mento dos genes, especialmente como eles sãoregulados. Essas informações deverão contribuirpara que os melhoristas possam conduzir os seustrabalhos com maior conhecimento científico doque atualmente o fazem.
Considerações Finais
A experiência do passado não deixa dú-vida de que a Biometria e o Melhoramento Ge-nético das Plantas tiveram importante papel emreduzir a fome no mundo. Não apenas isso, elascontribuíram também para aumento da produçãode madeira, fibras e muitos outros produtos es-senciais à vida do homem, sobretudo durante oséculo XX. Não há nenhuma evidência de que aimportância dessa ciência seja reduzida no futu-ro.
Normalmente, comenta-se que o trabalhode melhoramento genético é demorado, levandoem torno de 10 a 15 anos em média para obter-seuma nova linhagem. Esse tempo é relativo, pois,
se um programa de melhoramento já está implan-tado, a cada ano, é possível recomendar uma novacultivar. Essa constatação é válida para solucio-nar os problemas rotineiros nos programas demelhoramento. Se ocorrer um problema novo, éevidente que demandará mais tempo. Nesse últi-mo caso, a observação é válida mesmo quandose utilizaram técnicas biotecnológicas. Ostransgênicos, por exemplo, apresentam o mesmoproblema de uma cultivar obtida por métodosconvencionais, pois há necessidade de compro-vação de que ele é bom, somente possível apósintensiva avaliação.
Um outro aspecto refere-se ao custo deobtenção de uma nova cultivar. Ferreira (2003),utilizando a cultura de arroz como referência,escreveu que: “o desenvolvimento de uma novavariedade comercial leva de seis a oito anos, comcusto médio de milhares ou, tipicamente, milhõesde reais”. A questão do tempo já foi comentada,no entanto, se a obtenção de uma nova cultivarcustasse milhões de reais, é provável que, somen-te para espécies como soja e café, haveria condi-ções de se obterem novas cultivares. O recursodisponível, especialmente para culturas de sub-sistência, como arroz e feijão, tem sido bem in-ferior a esses relatados e, mesmo assim, novascultivares freqüentemente chegam ao mercado.Deve ser enfatizado que o custo de qualquer pro-grama de melhoramento, comparado com o damaioria das técnicas biotecnológicas, é bem in-ferior. Infelizmente, não há disponibilidade deinformaçôes de quanto foi investido pelos órgãosde fomento no Brasil, nos últimos vinte anos, emtécnicas biotecnológicas e no melhoramento con-vencional. Mas, certamente, os programas de me-lhoramento receberam quantia ínfima em rela-ção à biotecnologia. Esse, aliás, é o principal pro-blema enfrentado pelos melhoristas no Brasil. Osrecursos, sobretudo aqueles relativos a programas

245Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
especiais, nunca são destinados ao melhoramen-to convencional. Deve-se considerar que é noagronegócio que a economia do país tem-se sus-tentado e a obtenção de novas cultivares pelomelhoramento convencional é uma das principaisrazões desse sucesso. Paterniani (2002), comen-tando sobre as técnicas de manipulação genéti-ca, escreveu que “ainda é oportuno e desejável acondução de programas de melhoramento domilho empregando as técnicas convencionais jáconsagradas como eficientes e ainda não sufici-entemente exploradas”.
O treinamento de profissionais com ên-fase ao melhoramento genético têm-se reduzidoacentuadamente nos últimos anos. Nos cursos deGenética e Melhoramento de Plantas, por exem-plo, o número de matrículas na disciplina de Ge-nética Molecular,é pelo menos, três vezes supe-rior ao da Genética Quantitativa. Walsh (2001),comentando sobre as perspectivas da genéticaquantitativa na era da Genômica, enfatiza que éindispensável atualmente aos biologistasmoleculares o conhecimento profundo do traba-lho de Fisher (1918). A recíproca também é ver-dadeira, é importante que os melhoristas tenhamsólidos conhecimentos em biologia molecular,para avaliarem o real potencial e identificaremsituações em que as técnicas possam ser úteis.
Os conhecimentos básicos gerados pelaBiologia Molecular tornaram as ciências relaci-onadas ao Melhoramento de Plantas muito maisexcitantes. Os recursos computacionais disponí-veis possibilitam simulações de situações que ja-mais os pioneiros da Genética Quantitativa pu-deram imaginar. A cada dia que passa, novas in-formações são adicionadas ao conhecimento daatuação dos genes. Isso possibilitará que, numfuturo bem próximo, os continuadores do traba-lho de Fisher e Mather, tenham muito mais dadospara formularem os seus modelos e proporem
alternativas para tornar os programas de melho-ramento genético ainda mais eficazes. Os novosconhecimentos também têm servido para mos-trar a capacidade criativa e de imaginação de inú-meros cientistas que marcaram suas vidas por im-portantes contribuições à ciência. Um exemplo éo caso do geneticista russo Nikolai Vavilov, quededicou sua vida a elucidar o processo dedomesticação das espécies cultivadas, estabele-cendo o que foi conhecido por lei das sérieshomólogas da variação. Segundo essa lei, pode-se esperar que características encontradas em umaespécie também sejam encontradas em espéciesrelacionadas. Vavilov utilizou esse princípio paraprever características ainda não descobertas. To-dos os estudos com marcadores moleculares e,especialmente da genômica, comprovam a lei dasséries homólogas.
Literatura Citada
ABREU, A. F. B. Predição do potencial genéti-co de populações segregantes do feijpeiro uti-lizando genitores inter-raciais. 1997. 79 f. Tese(Doutorado em Genética e Melhoramento dePlantas) – Universidade Federal de Lavras, La-vras.
ALLARD, R. W. History of plant populationgenetics. Annual Review of Genetics, Palo Alto,v. 33, p. 1-27, 1999.
BAENZIGER, P. S.; PETERSON, E. J. Geneticvariation: its origin and use for breeding selfpollinated species. In: STALKER, H. T.;MULTRIPHY, J. P. Plant Breeding in the1990’s. Raleigh: North Caroline University, 1991.p. 69-100.
BEARZOTI, E. Simulação de seleção recor-rente assistida por marcadores molecularesem espécies autógamas. 1997. 230 f. Tese

246
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
(Doutorado) – Escola Superior de Agricultura“Luiz de Queiroz”, Universidade de São Paulo,Piracicaba.
BERNARDO, R. What if we knew all the genesfor a quantitative trait in hybrid crops? CropScience, Madison, v. 41, n.1, p.1-4, 2001.
BERZONSKY, W .A.; LAFEVER, H. N.Progress in Ohio soft red winter wheat breeding:grain yield and agronomic traits of cultivarsreleased from 1871 to 1987. Crop Science,Madison, v. 33, n. 6, p. 1382, 1993.
CARDWELL, V. B. 50 years of Minnesota cornproduction - sources of yield increase. AgronomyJournal, Madison, v. 74, n. 6, p. 984-990, 1982.
CARNEIRO, J. E. S. Alternativas para obten-ção e escolha de populações segregantes nofeijoeiro. 2002. 134 f. Tese (Doutorado em Ge-nética e Melhoramento de Plantas) - UniversdadeFederal de Lavras, Lavras.
CROW, J. F. Dominance and overdominance. In:CORRS, J. G.; PANDEY, S. (Ed.) The geneticsand exploitation of heterosis in crops. Madison:American Society of Agronomy, 1999. p. 49-58.
DUARTE, J. M Conversão de linhagens elitesem milho de alta qualidade protéica (QPM).2003. 129 f. Tese (Doutorado em Genética eMelhoramento de Plantas) – Universidade Fede-ral de Lavras, Lavras.
DOEBLEY, J.; STEC, A.; HUBBARD; L. Theevolution of apical dominance in maize. Nature,London, v. 386, p. 485-488, 1997.
DUDLEY, J. W. 76 genrations of selection foroil and protein percentage in maize. In:INTERNATIONAL CONFERENCE ONQUANTITATIVE GENETICS, 1976, Ames,
Iowa. Proceedings... Ames: Iowa StateUniversity Press, 1977. p. 459-474.
DUDLEY, J. W.; LAMBERT, R. J. Ninetygenerations of selection for oil and protein inmaize. Maydica, Bergamo, v. 37, p. 81-87, 1992.
DUDLEY, J. W. Molecular markers in plantimprovement: manipulation of genes affectingquantitative traits. Crop Science, Madison, v. 33,p. 660-667, 1993.
DUDLEY, J. W. Quantitative genetics and plantbreeding. Advances in Agronomy, New York,v. 59, p. 1-23, 1997.
DUVICK, D. N. Plant Breeding, an evolutionaryconcept. Crop Science, Madison, v. 36, p. 539-548, 1996.
EVANS, L. T. Feeding the ten billion: plants andpopulation growth. Cambridge: University Press,1998. 247 p.
FALCONER, D. S, MACKAY, T. E.Introduction to quantitative genetics. 4.ed.New York: Longman, 1996. 438 p.
FEHR, W. R. Principles of cultivardevelopment. New York: Mcmillian, 1987. 761p.
FERREIRA, M. E. Melhoramento genético doarroz: impactos da genômica. In: BORÉM, A.;GIÚDICE, M.; SEDIYAMA, T. (Ed.) Melhora-mento genômico. Viçosa: UFV, 2003. Cap. 4, p.73-127.
FERREIRA, M. E.; GRATTAPAGLIA, D. Intro-dução ao uso de marcadores moleculares emanálises genéticas. 3 ed. Brasília: EmbrapaCenargen, 1998. 220 p.
FISHER, R. A. The correlations between relativeson the supposition of Mendelian inheritance.

247Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Transactions of the Royal Society of Edinburg,Edinburg, v. 52, p. 399-433, 1918.
FRISCH, M.; BOHN, M.; MELCHINGER, A.E. Comparison of selection strategies for marker-assisted backrossing of a gene. Crop Science,Madison, v. 39, p. 1295-1301, 1999.
FRISCH, M.; MELCHINGER, A. E. Marker-Assisted backrossing for simultaneousintrogression of two genes. Crop Science,Madison, v .41, p. 1716-1725, 2001.
GLAZIER, A. M.; NADEAU, J. H.; ALTMAN,T. J. Finding genes that underlie complex traits.Science, Washington, v. 298, p. 2345-2349, Dec.2002.
HALLAUER, A. R.; MIRANDA FILHO, J. B.;Quantitative genetics in maize breeding. Ames:Iowa State University Press, 1981. 468 p.
HOSPITAL, F.; CHARCOSSET, A. Marker-assisted introgression of quantitative trait loci.Genetics, Baltimore, v. 147, p. 1469-1485, 1997.
GEPTS, P. A Comparison between cropdomestication, classical plant breeding, andgenetic engineering. Crop Science, Madison, v.42, p. 1780-1790, 2002.
HOSPITAL, F.; CHEVALET, C.; MULSANT, P.Using markers in gene introgression breedingprograms. Genetics, Baltimore, v. 132, p. 1199-1210, 1992.
JENSEN, N. F. Plant breeding methodology.New York: J. Wiley, New York, 1988. 676 p.
JINKS, J. L; POONI, H. S. Prediciting theproperties of recombinat inbred lines derived bysingle seed descent. Heredity, Essex, v. 36, n. 2,p. 253-266, 1976.
KEARSEY, M. J.; FARQUHAR, A. G. L. QTLanalysis in plants; where are we now? Heredity,Essex, v. 80, p. 137-142, 1998.
KEARSEY, M. J.; POONI, H. S. The geneticalanalysis of quantitative traits. Oxford:Chapman and Hall, 1996. 381 p.
KUMAR, L. S. DNA markers in plantimprovement: An overview. BiotechnologyAdvances, Oxford, v. 17, p. 143-182, 1999.
LANDER, E. S.; BOTSTEIN, D. Mappingmendelian factors underlying quantitative traitsusing RFLP linkage maps. Genetics, Maryland,v. 121, p. 185-199, 1989.
LANZA, M. A.; GUIMARÃES, C. T.;SCHUSTER, I.; Aplicação dos marcadoresmoleculares no melhoramento genético. Infor-me Agropecuário, Belo Horizonte, v. 21, n. 204,p. 97-108, 2000.
LEE, M. DNA markers and plant breedingprograms. Advances in Agronomy, New York,v. 55, p. 265-343, 1995.
LYNCH, M.; WALSH, B. Genetics and analysisof quantitative traits. Sunderland: SinauerAssociates, 1998. 980 p.
MACHADO, C. F.; SANTOS, J. B.; NUNES, G.H. S.; DUARTE, J. M. Efficiency of geneticdistance based on RAPD markers for choosingparents of common bean. Journal of Genetics& Breeding, Rome, v. 54, p. 252-258, 2000.
MACHADO, C. de F. Procedimentos para aescolha de genitores de feijão. 1999. 118 f. Dis-sertação (Mestrado em Genética e Melhoramen-to de Plantas) – Universidade Federal de Lavras,Lavras.

248
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
Ramalho & Lambert
MACKAY, T. F. C. The genetic architecture ofquantitative traits. Annual Review of Genetics,Palo Alto, v. 35, p. 303-339, 2001.
MATHER, K. Biometrical genetics: the studyof continuos variation. London: Methuen, 1949.162 p.
McCLEAN, P. Analysis of plant genomes withmolecular markers: linkage drag Disponível em:<http://www.cc.ndsu.nodak.edu/instruct/mcclean/plsc731/analysis/analysis6.htm >. Aces-so em: 20/10/2003.
MENDONÇA, H. A. de. Escolha de populaçõessegregantes de feijoeiro utilizando parâmetrosgenéticos, fenotípicos e marcadores RAPD.2001. 100 f. Tese (Doutorado em Genética eMelhoramento de Plantas) – Universidade Fede-ral de Lavras, Lavras.
PADGETTE, S. R.; KOLACZ, K. H.;DELANNAY, X.; RE, D. B.; LAVALLEE, B. J.;TINIUS, C. N.; RHODES, W. K.; OTERO, Y. I.;BARRY, G. F; EICHHOLTZ, D. A.; PESCHKE,V. M.; NIDA, D. L.; TAYLOR, N. B.; KISHORE,G. M. Development, identification, andcharacterization of a glyphosate-tolerant soybeanline. Crop Science, Madison, v. 35, p. 1451-1461,1995.
PATERNIANI, E. Breve História da Agriculturaaté a era da biotecnologia. In: PATERNIANI, E.(Ed.) Agricultura Brasileira e pesquisaagropecuária. Brasília: Embrapa Comunicacaopara Transferencia de Tecnologia,, 2000. cap.1,p.11-30.
PATERNIANI, E. Uma percepção crítica sobretécnicas de manipulação genética. Revista Bra-sileira de Milho e Sorgo, Sete Lagoas, v. 1, n. 1,p. 77-84, jan./abr. 2002.
PATERSON, A. H.; TANKSLEY, S. D.;SORRELLS, M. E. DNA markers in plantimprovement. Advances in Agronomy, NewYork, v. 46, p. 39-89, 1991.
PRAKASH, C. S. The genetically modified cropdebate in the context of agricultural evolution.Plant Physiology, Bethesda, v. 126, p. 8-15, 2001.
RASMUSSON, D. C.; PHILLIPS, R. L. Plantbreeding progress and genetic diversity from denovo variation and elevated epistasis. CropScience, Madison, v. 37, n. 2, p. 303-310, Mar./Apr. 1997.
SANTOS, P. G. Escolha de populaçõessegregantes para o programa de seleção dearroz em terras altas. 2000. 106 f. Tese (Dou-torado em Genética e Melhoramento de Plantas)– Universidade Federal de Lavras, Lavras.
SMITH, J. S. C.; SMITH, O. S. The descriptionand assessment of distances between inbreds ofmaize. II. The utilizty of morphological,biochemical, and genetic descriptors and ascheme for the testing of distinctiveness betweeninbreds. Maydica, Bergamo, v. 34, p. 151-156,1989.
SOUZA, A. P. Biologia molecular aplicada aomelhoramento. In: NASS, L. L.; VALOIS, A. C.C.; MELO, I. S.; VALADARES-INGLIS, M. C.(Ed.) Recursos genéticos e melhoramento deplantas. Rondonópolis: Fundacao MT, 2001. p.939-966.
SOUZA Jr., C. L de. Genetic variances ofmolecular markers of F2 and related populations.Revista Brasileira de Genética, Ribeirão Pre-to, v. 15, n. 2, p. 913-926, 1992.
SOUZA SOBRINHO, F. Divergência genéti-ca de híbridos simples e alternativas para

249Biometria e o melhoramento de ...
Revista Brasileira de Milho e Sorgo, v.3, n.2, p.228-249, 2004
obtenção de híbridos duplos de milho. 2001.96 f. Tese (Doutorado em Genética e Melhora-mento de Plantas) – Universidade Federal deLavras, Lavras
STUBER, C. W. Biochemical and molecularmarkers in plant breeding. Plant BreedingReviews, New York, v. 9, p. 37- 61, 1992.
TANKSLEY, S. D.; YOUNG, N. D.;PATTERSON, A. H.; BONIERBALE, M. W.RFLP mapping in plant breeding: new tools foran old science. Bio/Technology, New York, v. 7,n. 3, p. 257-263, 1989.
TROYER, A. F.; ROCHERFORD, T. R.Germplasm ownership: related corn inbreds.Crop Science, Madison, v. 42, n.1, p. 3-11, 2002.
VENCOVKY, R.; RAMALHO, M. A. P. Contri-buição do Melhoramento Genético de Plantas noBrasil. In: PATERNIANI, E. (Ed.) AgriculturaBrasileira e pesquisa agropecuária. Brasília:Embrapa Comunicação para Transferência deTecnologia, 2000. cap. 4, p. 57-89.
WALSH, B. Quantitative genetics in the age ofgenomics. Theoretical Population Biology, SanDiego, v. 59, p. 75-184, 2001.
WILCOX, J. R. Sixty years of improvement inpublicly developed elite soybean lines. CropScience, Madison, v. 49, p. 1711-1716, 2001.
WRICKE, G.; WEBER, E. Quantitative geneticsand selection in plant breeding. Berlin: Walterde Gruyter, 1986. p. 257-279.