aulas populações e quantitativa (2)
TRANSCRIPT

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
Faculdade de Engenharia de Ilha Solteira
Genética de
Populações e
Quantitativa
Prof. Dr. João Antonio da Costa Andrade
Departamento de Biologia e Zootecnia

Caracteres qualitativos
Descontinuidade, facilmente classificados em categorias
fenotípicas distintas.
Caracteres quantitativos
Continuidade (entre fenótipos extremos aparecem
inúmeros fenótipos intermediários), diferença entre
determinados fenótipos é mínima e os fenótipos, via de
regra, são mensuráveis.

Qualitativos Quantitativos
caracteres de tipo caracteres de grau
variação descontínua variação contínua
poucos locos gênicos muitos locos gênicos
genes com grande efeito genes com pequeno efeito
menor efeito ambiental maior efeito ambiental

CARÁTER
QUALITATIVO CARÁTER
QUANTITATIVO
Espécie com 2n = 16 cromossomos

Numero de genótipos diferentes com m locos e n alelos
por loco
NGD = [n(n+1)/2]m
Números fenótipos diferentes vai depender de:
• Ação gênica;
• Tipo de efeitos (aditivos iguais, aditivos
desiguais, oposicionais);
• Efeito do ambiente.

Base genética dos caracteres quantitativos são os Genes ou fatores múltiplos: -Bateria de locos com efeito sobre o mesmo caráter.
“A segregação dos genes envolvidos não pode ser seguida
individualmente, embora cada loco comporte-se
exatamente como Mendel postulou”

Sistemas de locos seriados – cada loco atuando em um passo do processo
loco A loco B loco C
enzima a enzima b enzima c
passo a passo b passo c
Substrato produto A produto B P.F.
Mutações drásticas, inativando enzimas, vão criar
fatores complementares (epistasia).
Mutações “leves” que causem variação na atividade das
enzimas, causando variação contínua na quantidade do
produto final, causarão variação quantitativa.

Efeito de dosagem – o caráter se expressa conforme o número de alelos favoráveis e/ou desfavoráveis presentes.
Exemplo: Vitamina A por g de endosperma (3n) de milho.
yyy – 0,05
Yyy – 2,25
YYy – 5,00
YYY – 7,50

Polimeria
Genes com efeitos aditivos e iguais.
Anisomeria
Genes com efeitos aditivos e desiguais.
Sistemas oposicionais
Genes com efeitos aditivos e subtrativos.

Método de Sewall Wright
Valores genotípicos (q2) (2pq) (p2) bb m Bb BB -a +a d BB = m + a (p2) Grau médio de dominância = gmd = d/a
Bb = m +d (2pq)
bb = m – a (q2) Média da pop. = g = m+a(p-q)+2pqd
P1 m F1 P2
-a +a d

Efeitos aditivos e iguais: Todos os alelos favoráveis
contribuem com o mesmo valor e os menos favoráveis
com outro valor.
Exemplo com três locos: A(a); B(b); C(c)
A=B=C=3; a=b=c=1
1 – Sem dominância: P1 – AABBCC 6 + 6 + 6 = 18 P2 – aabbcc 2 + 2 + 2 = 6 F1 – AaBbCc 4 + 4 + 4 = 12
F2

(1/8) ABC (1/8) ABc (1/8) AbC (1/8) Abc (1/8) aBC (1/8) aBc (1/8) abC (1/8) abc
(1/8) ABC
AABBCC (1/64)
AABBCc (1/64)
AABbCC (1/64)
AABbCc (1/64)
AaBBCC (1/64)
AaBBCc (1/64)
AaBbCC (1/64)
AaBbCc (1/64)
(1/8) ABc
AABBCc (1/64)
AABBcc (1/64)
AABbCc (1/64)
AABbcc (1/64)
AaBBCc (1/64)
AaBBcc (1/64)
AaBbCc (1/64)
AaBbcc (1/64)
(1/8) AbC
AABbCC (1/64)
AABbCc (1/64)
AAbbCC (1/64)
AAbbCc (1/64)
AaBbCC (1/64)
AaBbCc (1/64)
AabbCC (1/64)
AabbCc (1/64)
(1/8) Abc
AABbCc (1/64)
AABbcc (1/64)
AAbbCc (1/64)
Aabbcc (1/64)
AaBbCc (1/64)
AaBbcc (1/64)
AabbCc (1/64)
Aabbcc (1/64)
(1/8) aBC
AaBBCC (1/64)
AaBBCc (1/64)
AaBbCC (1/64)
AaBbCc (1/64)
aaBBCC (1/64)
aaBBCc (1/64)
aaBbCC (1/64)
aaBbCc (1/64)
(1/8) aBc
AaBBCc (1/64)
AaBBcc (1/64)
AaBbCc (1/64)
AaBbcc (1/64)
aaBBCc (1/64)
aaBBcc (1/64)
aaBbCc (1/64)
aaBbcc (1/64)
(1/8) abC
AaBbCC (1/64)
AaBbCc (1/64)
AabbCC (1/64)
AabbCc (1/64)
aaBbCC (1/64)
aaBbCc (1/64)
aabbCC (1/64)
aabbCc (1/64)
(1/8) abc
AaBbCc (1/64)
AaBbcc (1/64)
AabbCc (1/64)
Aabbcc (1/64)
aaBbCc (1/64)
aaBbcc (1/64)
aabbCc (1/64)
aabbcc (1/64)

Freq. Fenótipos Genótipos
1/64 18 AABBCC
6/64 16 AABBCc; AABbCC; AaBBCC
15/64 14 AABBcc; AAbbCC; aaBBCC; AaBbCC;
AaBBCc; AABbCc
20/64 12 AABbcc; AaBBcc; AAbbcC; AabbCC; aaBbCC; aaBBCc; AaBbCc
15/64 10 AAbbcc; aaBBcc; aabbCC; AaBbcc;
AabbCc; aaBbCc
6/64 8 Aabbcc; aaBbcc; aabbCc;
1/64 6 aabbcc
Frequência fenotípica em F2 sem dominância

Freq. Fenótipos Genótipos
3/64 18 AABBCC; AaBBCC
12/64 16 AABBCc; AaBBCc; AABbCC; AaBbCC
19/64 14 AABbCc; AaBBcc; AaBbCc; AAbbCC; AabbCC; aaBBCC; AABBcc
16/64 12 AABbcc; AaBbcc; AAbbCc; AabbCc; aaBBCc; aaBbCC
8/64 10 AAbbcc; Aabbcc; aaBBcc; aaBbCc;
aabbCC
5/64 8 aaBbcc; aabbCc
1/64 6 aabbcc
Freq. fenotípica em F2 com dominância no loco A(a)

Frequência sem dominância
Fenótipos Frequência com
dominância no loco A(a)
1/64 18 3/64
6/64 16 12/64
15/64 14 19/64
20/64 12 16/64
15/64 10 8/64
6/64 8 5/64
1/64 6 1/64
Comparação

Do
min
ân
cia
loco
A(a
)
a) Sem dominância
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=12
b) Dom. em A(a)
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=14
c) Dom. dois locos
P1 - AABBCC=18
P2 – aabbcc=6
F1 - AaBbCc=16
d) Dom. três locos
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=18
Efeitos aditivos e iguais: A=B=C=3; a=b=c=1

Efeitos aditivos e desiguais: A=4,5; B=C=2,25; a=1,5; b=c=0,75
a) Sem dominância
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=12
b) Dom. em A(a)
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=15
c) Dom. A(a) e B(b)
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=16,5
d) Dom. três locos
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=18

Efeitos oposicionais:
A=13; B=C= -2;
a=5; b=c= -1
a) Sem dominância
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=12
b) Dom. em A(a)
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=20
c) Dom. A(a) e B(b)
P1 - AABBCC=18
P2 - aabbcc=6
F1 - AaBbCc=19
d) Dom. em B(b) ou C(c)
P1 - AABBCC=18
P2 - aabbcc=6
F1 – AaBbCc=11

Genótipo Freqüência
em 64 indivíduos
Genótipo
para cada loco
Genótipo total Fenótipo para cada loco
AABBCC 1 2+2+2 6 (2 s2)+(2 s2)+(2 s2)
AABBCc 2 2+2+1 5 (2 s2)+(2 s2)+(1 s1)
AABBcc 1 2+2+0 4 (2 s2)+(2 s2)+(0 s0)
AABbCC 2 2+1+2 5 (2 s2)+(1 s1)+(2 s2)
AABbCc 4 2+1+1 4 (2 s2)+(1 s1)+(1 s1)
AABbcc 2 2+1+0 3 (2 s2)+(1 s1)+(0 s0)
AAbbCC 1 2+0+2 4 (2 s2)+(0 s0)+(2 s2)
AAbbCc 2 2+0+1 3 (2 s2)+(0 s0)+(1 s1)
AAbbcc 1 2+0+0 2 (2 s2)+(0 s0)+(0 s0)
AaBBCC 2 1+2+2 5 (1 s1)+(2 s2)+(2 s2)
AaBBCc 4 1+2+1 4 (1 s1)+(2 s2)+(1 s1)
AaBBcc 2 1+2+0 3 (1 s1)+(2 s2)+(0 s0)
AaBbCC 4 1+1+2 4 (1 s1)+(1 s1)+(2 s2)
AaBbCc 8 1+1+1 3 (1 s1)+(1 s1)+(1 s1)
AaBbcc 4 1+1+0 2 (1 s1)+(1 s1)+(0 s0)
AabbCC 2 1+0+2 3 (1 s1)+(0 s0)+(2 s2)
AabbCc 4 1+0+1 2 (1 s1)+(0 s0)+(1 s1)
Aabbcc 2 1+0+0 1 (1 s1)+(0 s0)+(0 s0)
aaBBCC 1 0+2+2 4 (0 s0)+(2 s2)+(2 s2)
aaBBCc 2 0+2+1 3 (0 s0)+(2 s2)+(1 s1)
aaBBcc 1 0+2+0 2 (0 s0)+(2 s2)+(0 s0)
aaBbCC 2 0+1+2 3 (0 s0)+(1 s1)+(2 s2)
aaBbCc 4 0+1+1 2 (0 s0)+(1 s1)+(1 s1)
aaBbcc 2 0+1+0 1 (0 s0)+(1 s1)+(0 s0)
aabbCC 1 0+0+2 2 (0 s0)+(0 s0)+(2 s2)
aabbCc 2 0+0+1 1 (0 s0)+(0 s0)+(1 s1)
aabbcc 1 0+0+0 0 (0 s0)+(0 s0)+(0 s0)

Generalizando
•Com grande número de locos com efeitos aditivos segregando, a F2 exibe baixa freqüência dos parentais, um grande número de fenótipos e, conseqüentemente, uma variação cada vez mais contínua;
•Genes com efeitos desiguais provocam ainda um maior número de classes fenotípicas, aumentando a “largura da curva de distribuição fenotípica”.
•O efeito do ambiente faz com que a curva de freqüência dos valores fenotípicos em F2 seja ainda mais contínua, mesmo sob alguma condição de dominância.

Grau de atividade
no inverno
Raça
Costeira
Raça
Alpina
F1
Segregação observada
em F2
Segregação esperada com 3
locos poliméricos
Completamente ativa + 43 15
Parcialmente ativa 139
945 Intermediária + 601 918
Parcialmente dormente 178
Completamente dormente + 14 15
Herança da dormência em Potentilla glandulosa
(Clausen e Hiesey, 1958)
Herança: três locos com efeitos aditivos e
aproximadamente iguais, sem dominância

Comprimento do estigma em mm
0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 1,2 1,3 1,4 1,5 N
P1 ca + +
P2 ch + +
F1 + +
F2 1 11 43 57 33 24 3 2 1 1 176
B1 (ca x ch) x ca 1 3 9 1 1 1 1 17
B2 (ca x ch) x ch 3 1 3 1 1 9
Distribuição de frequência de indivíduos com vários
comprimentos do lóbulo dos estigmas em Gilia capitata
capitata, Gilia capitata chamissonis e suas progênies
híbridas (Grant, 1950)
Herança: 3 ou 4 locos com pelo menos um com alelos
dominantes para estigmas curtos.

Distribuição de frequência do comprimento da roseta da
folha entre 996 F2 de um cruzamento interracial em
Potentilla glandulosa (Clausen e Hiesey, 1958)
Herança: Sistema de locos oposicionais, dominância
para folhas longas

Genes modificadores Sistema em que locos com efeitos muito pequenos (locos múltiplos) atuam a favor ou contra o efeito dos alelos de um gene maior (de grande efeito). Milho normal (alto) Milho braquítico (baixo) BB ++ - - ++ -- X bb - - ++ - - ++ F1 Bb +- +- +- +- ¼ BB (++ ++ ++ ++,..... +- +- +- +- ,....- - - - - - - -) F2 ½ Bb (++ ++ ++ ++,.... +- +- +- +-,. ....- - - - - - - -) ¼ bb (++ ++ ++ ++,... +- +- +- +-,......- - - - - - -)

Manchas nas pétalas do algodoeiro Gossypium barbadense Gossypium hirsutum
Manchado x Sem mancha Manchado x Sem mancha
3 manchados : 1 sem manchas em F2
Gossypium barbadense X Gossypium hirsutum Manchado Sem manchas
F1 intermediário
F2 com 22 graus diferentes de manchas devido a
diferenças alélicas entre as para modificadores.

Gossypium barbadense Gossypium hirsutum
Manchado x Sem mancha Manchado x Sem mancha (MM ++ ++ ++)x(mm ++ ++ ++) (MM _ _ _ _ _ _)x(mm _ _ _ _ _ _)
3 manchados : 1 sem manchas em F2
(M_ ++ ++ ++) : (mm ++ ++ ++) (barbadense)
(M_ _ _ _ _ _ _) : (mm _ _ _ _ _ _) (hirsutum)
Gossypium barbadense X Gossypium hirsutum Manchado (MM ++ ++ ++) Sem manchas (mm _ _ _ _ _ _)
F1 intermediário (Mm +_ +_ +_)
¼ (MM ++ ++ ++;.... MM+_ +_ +_; ....MM _ _ _ _ _ _)
F2 ½ (Mm ++ ++ ++;.... Mm+_ +_ +_; .... Mm _ _ _ _ _ _)
¼ (mm ++ ++ ++;.... mm+_ +_ +_; .....mm _ _ _ _ _ _)

Como descobriram que G. barbadense possui
modificadores para aumentar as manchas e G
hirsutum para diminuir????
Caracteres quantitativos e interação gênica
“Como os caracteres quantitativos são
controlados por muitos locos, o que se procura
determinar é o tipo de interação gênica
predominante, uma vez que na prática é
impossível conhecer o tipo de interação em
cada loco e entre locos específicos.”




Médias e variâncias
0
0,5
1
1,5
2
2,5
m
F11 = m + g1 + e1;
F22 = m + g2 + e2;
F33 = m + g3 + e3;
F44 = m + g4 + e4;
F55 = m + g5 + e5;
F66 = m + g6 + e6;
F77 = m + g7 + e7;
: : : :
Fnn = m + gn + en;
n
Fm
ij
1
)(
11
)(
2222
2
n
n
FF
n
d
n
mFij
ijijij
F

Linhagem pura ou
Clone
F11 = m + g1 + e1;
F22 = m + g1 + e2;
F33 = m + g1 + e3;
F44 = m + g1 + e4;
F55 = m + g1 + e5;
F66 = m + g1 + e6;
F77 = m + g1 + e7;
: : : :
Fnn = m + g1 + en;
----------------------- 222 0 EGF
F11 = m + g1 + e1;
F22 = m + g2 + e2;
F33 = m + g3 + e3;
F44 = m + g4 + e4;
F55 = m + g5 + e5;
F66 = m + g6 + e6;
F77 = m + g7 + e7;
: : : :
Fnn = m + gn + en;
----------------------
Variedade
22 00 EF

Lote A -28 sementes, cada uma Lote B - 18 sementes da de uma planta diferente mesma planta mãe
3,8 6,6 4,7 9,0 7,3 6,9 5,4 5,2 4,8 3,9 4,3 4,1 5,7 9,7 5,6 5,8 7,3 4,2 10,2 8,8 6,1 6,0 5,9 4,9 5,5 5,8 7,4 6,2 3,9 6,2 8,6 10,4 4,3 3,8 6,1 5,2 4,0 4,2 4,4 3,8 2,6 4,4 5,9 4,3 3,6 9,3
Lote B Lote A
5,2 = m + G1 + E1 3,8 = m + G1 + E1 4,8 = m + G1 + E2 6,6 = m + G2 + E1 : : : : : : : : : : : : : : : : 4,4 = m + G1+ E18 9,3 = m + G18 + E1
0,69 = 0 + 0 = 2ˆe 4,98 = 0 +
22 ˆˆ eG
Portanto 2222 )/(29,4ˆˆˆ plkgBAG

Questões
•Qual o grau de confiança em reconhecer o valor
genotípico pelo valor fenotípico?
•As sementes colhidas das plantas superiores realmente
são superiores?
•A nova população produzida por essas sementes será
melhor que a população original? Quanto?
No exemplo, como a variância genética é bem superior
à variância ambiental, as medidas fenotípicas são boas
indicadoras dos respectivos valores genotípicos.

Coeficiente de herdabilidade para plantas autógamas
homozigóticas ou de propagação vegetativa
•São plantas que transmitem o genótipo integralmente para a
geração seguinte;
•É necessário saber quanto das diferenças fenotípicas (variância
fenotípica) é devido às diferenças genotípicas (variância
genotípica) entre indivíduos.
22
2
2
22
ˆˆ
ˆ100
ˆ
ˆ100ˆ
EG
G
F
Gh
%1,86
69,029,4
29,4100ˆ2
h

m0 ms ds
m0 m1 Gesp
ds = ms - m0 = diferencial de
seleção;
m0 = média da população original
ms = média dos indivíduos selecio-
nados;
Gesp = Progresso ou ganho com se-
leção;
Gesp = ds h2
m1 = média da população me-
lhorada
Melhoramento de populações

Observações importantes
•A herdabilidade é uma propriedade de um caráter em
uma dada população e em um dado ambiente;
•Podemos aumentar a herdabilidade ao uniformizar o
ambiente;
•O ganho esperado com seleção possui expressão
própria para cada método de seleção, mas que é
derivado da expressão original Gesp = ds h2.

Gmd 0 0 0 1 1 1 0,5 0,5 0,5
h2 100 50 20 100 50 20 100 50 20
Gen X1 X2 X3 X4 X5 X6 X7 X8 X9
AA 10,1* 10,05* 10,05 10,1* 10,12* 10,17* 10,10* 10,25* 10,33*
AA 10,1* 10,02 10,17* 10,1* 10,11 10,11* 10,10* 10,09* 10,02
AA 10,1* 10,16* 10,28* 10,1* 10,01 9,76 10,10* 10,16* 10,28*
AA 10,1* 10,13* 10,48* 10,1* 10,14* 10,01 10,10* 10,20* 10,02
AA 10,1* 10,12* 9,98 10,1* 9,98 9,91 10,10* 10,06 10,13*
Aa 10,0* 9,95 10,09* 10,1* 10,09 10,08 10,05* 9,96 10,07*
Aa 10,0* 10,17* 9,88 10,1* 10,37* 10,52* 10,05* 10,04 9,89
Aa 10,0* 10,14* 9,87 10,1* 10,11 10,03 10,05* 10,09* 10,13*
Aa 10,0 10,03* 10,09* 10,1 10,21* 10,28* 10,05 9,98 9,95
Aa 10,0 9,95 9,76 10,1 10,27* 10,04 10,05 10,17* 10,03
Aa 10,0 9,89 10,22* 10,1 10,14* 10,14* 10,05 10,02 9,86
Aa 10,0 10,01 10,11* 10,1 10,01 10,16* 10,05 10,12* 9,93
Aa 10,0 10,02 9,82 10,1 10,13* 9,99 10,05 10,17* 10,14*
Aa 10,0 9,95 10,05 10,1 10,24* 10,07 10,05 10,03 10,11*
Aa 10,0 10,08* 10,02 10,1 10,11 9,90 10,05 9,97 10,06
aa 9,90 9,97 9,93 9,9 9,86 9,80 9,90 9,90 9,55
aa 9,90 9,88 9,83 9,9 9,87 9,74 9,90 9,95 9,80
aa 9,90 9,9 10,23* 9,9 9,91 10,41* 9,90 10,06 10,14*
aa 9,90 9,95 9,93 9,9 9,93 10,10* 9,90 9,85 9,95
aa 9,90 9,99 10,01 9,9 9,99 9,99 9,90 9,95 9,91
Valores simulados (9 caracteres), com variação na dominância e herdabilidade

Caráter
Média
original
Selecio-
nados
p
Média
melhorada
Gs
AA Aa aa
X1 10,000 5 3 0 0,813 10,063 0,063
X2 10,000 4 4 0 0,750 10,050 0,050
X3 10,000 3 4 1 0,625 10,025 0,025
X4 10,050 5 3 0 0,813 10,093 0,043
X5 10,050 2 6 0 0,625 10,072 0,022
X6 10,050 2 4 2 0,500 10,050 0,000
X7 10,025 5 3 0 0,813 10,078 0,053
X8 10,025 4 4 0 0,750 10,069 0,044
X9 10,025 3 4 1 0,625 10,048 0,023
Genótipos selecionados, frequência alélica, média e ganho
com seleção

Estimativa do número de diferenças gênicas
No de locos Número de
alelos segregantes
Frequência de recuperação de um dos
fenótipos extremos em F2
1 2 (1/2)2 = 1/4 2 4 (1/2)4 = 1/16 3 6 (1/2)6 = 1/64 4 8 (1/2)8 = 1/256 . . . . . . n 2n (1/2)2n
Método da frequência dos fenótipos extremos em F2

B. Mexican x T. Thumb
(16,8 cm) (6,6 cm)
F1 (12,1 cm)
F2
Tamanho da espiga em duas raças de milho
Fenótipo (cm) Frequência Genótipos
16,8 1/16 AABB
14,2 4/16 2 AaBB;2 AABb
11,7 6/16 4 AaBb;1 aaBB; 1AAbb
9,1 4/16 2 aaBb; 2 Aabb
6,6 1/16 aabb
F2

Método de Sewall Wright
Valores genotípicos (q2) (2pq) (p2) bb m Bb BB -a +a d BB = m + a (p2) Grau médio de dominância = gmd = d/a
Bb = m +d (2pq)
bb = m – a (q2) Média da pop. = g = m+a(p-q)+2pqd
P1 m F1 P2
-a +a d
P2 P1

Para um loco: P P a1 2 2 ;
Para n locos: P P na1 2 2 a P P n ( )1 2 2 ;
Variância de F2 (interação aditiva): s naG
2 21 2 ( / ) ;
s n P P n P P nG
2
1 2
2
1 2
21 2 2 8 ( / ) [( ) ] ( )
nP P
sG
( )1 2
2
28
P1 m F1 P2
-a +a d
P2 P1

Requisitos necessários 1- Pais homozigóticos e contrastantes; 2 - Ausência de dominância e epistasia; 3 - Genes com efeito igual no fenótipo; 4 - Ausência de ligação.

Outro método
2
1
2
2
22 ]75,0[25,0
FF
DhhNG
)(21
12
11 PPPP
PFh
12PPD

Intensidade de seleção
•Seleção mais rigorosa ds maior menos
indivíduos selecionados;
•Seleção menos rigorosa ds menor mais
indivíduos selecionados;
Lote A -28 sementes, cada uma Lote B - 18 sementes da de uma planta diferente mesma planta mãe
3,8 6,6 4,7 9,0 7,3 6,9 5,4 5,2 4,8 3,9 4,3 4,1 5,7 9,7 5,6 5,8 7,3 4,2 10,2 8,8 6,1 6,0 5,9 4,9 5,5 5,8 7,4 6,2 3,9 6,2 8,6 10,4 4,3 3,8 6,1 5,2 4,0 4,2 4,4 3,8 2,6 4,4 5,9 4,3 3,6 9,3
No exemplo Seleção de cinco indivíduos de um total de 28
intensidade de seleção = 17,8%.

F
FF
idsdsds
i
2
Padronização: Índice de seleção
2
22
2
2
22)(
F
G
F
G
F
G
FFespiiihihdsG
•Esta fórmula é usada para se calcular o ganho
esperado na seleção truncada (quando se estabelece
apenas qual a intensidade de seleção que será utilizada);
•Neste caso o coeficiente da variância genética é um
devido ao fato de estarmos tratando de autógamas
homozigóticas ou plantas de propagação vegetativa.



Teste de progênie quando os indivíduos da
progênie têm o mesmo genótipo da planta mãe
•Não é o fenótipo do indivíduo que será usado para
avaliação, mas sim o comportamento médio da
progênie do indivíduo;
•Envolve retirada de várias sementes da mesma planta
ou a confecção de clones da mesma planta;

x x x x x x x x x x x x x x x x x x
x x x x x x x x x x x x x x x x x x
x x x x x x x x x x x x x x x x x x
x x x x x x x x x x x x x x x x x x

Exemplo: Produção de frutos em kg/planta
Plantas mãe (genótipos)
Plantas filhas (progênies)
Média das progênies
Variância ambiental
1 3,8 4,0 3,90 0,020 2 4,5 3,9 4,20 0,180 3 6,9 5,3 6,10 1,280 4 8,8 6,2 7,50 3,380 5 6,3 4,8 5,55 1,125 6 9,3 7,5 8,40 1,620 7 5,9 4,8 5,35 0,605 8 2,4 3,6 3,00 0,720 9 6,6 7,3 6,95 0,245
10 2,3 4,8 3,55 3,125 Média 5,45 1,230
2E

Plantas mãe (genótipos)
Plantas filhas (progênies)
Média das progênies
Variância ambiental
1 3,8 4,0 3,90 0,020 2 4,5 3,9 4,20 0,180
10 2,3 4,8 3,55 3,125 Média 5,745 1,230
020,0)0,48,3(2
10,48,3
12
12222
1
E

Planta mãe 1 (Genótipo - G1)
3,8 = m + G1 + E1 4,0 = m + g1 + E1’
Médias de progênies
2)(
2
1!11
1'11111
EEGmEGmEGmP
2)(
2
1!22
2'21222
EEGmEGmEGmP
2)(
2
1!
'
nn
nnnnnn
EEGmEGmEGmP
Generalizando

Plantas mãe Média de progênies 01 3,90 = m +G1+ ½(E1 + E1’) 02 4,20 = m +G2+ ½(E2 + E2’) 03 6,10 = m +G3+ ½(E3 + E3’) 04 7,50 = m +G4+ ½(E4 + E4’) 05 5,50 = m +G5+ ½(E5 + E5’) 06 8,40 = m +G6+ ½(E6 + E6’) 07 5,35 = m +G7+ ½(E7 + E7’) 08 3,00 = m +G8+ ½(E8 + E8’) 09 6,95 = m +G9+ ½(E9 + E9’) 10 3,55 = m +G10+ ½(E10 + E10’)
= 0 + + (½) = 3,241 2ˆF 2ˆG2ˆE
Generalizando para k indivíduos por progênie
)................(1
321 kiiEEEE
kGmP
2221
EGF
k

681,0ˆ
ˆ
ˆˆ
ˆˆ2
2
22
2
F
G
EG
Gh
81,0ˆ
ˆ
ˆ1
ˆ
ˆˆ2
2
22
2
F
G
EG
G
m
k
h
71,1ˆ
ˆ
ˆ
ˆ2
2
2
22
2
dshdsdsF
G
EG
G
g
03,2ˆ
ˆ
1ˆ
ˆ2
2
2
22
2
m
F
G
EG
G
g mdshds
k
ds
Herdabilidade em nível de
indivíduo.
Herdabilidade em nível de
médias de progênies.
Ganho com seleção em nível de indivíduos
Ganho com seleção em nível de médias de progênies
1,58
1,89

Componentes da variância genética Valores genotípicos (q2) (2pq) (p2) bb m Bb BB -a +a d BB = m + a (p2) Grau médio de dominância = gmd = d/a
Bb = m +d (2pq)
bb = m – a (q2) Média da pop. = g = m+a(p-q)+2pqd
Variância genética aditiva – variância devida aos efeitos
aditivos dos genes;
Variância genética dominante - variância devida aos
efeitos de dominância dos genes;
Variância genética epistática - variância devida aos
efeitos epistáticos dos genes;
2222 ˆˆˆˆIDAG
22222 ˆˆˆˆˆEIDAF

Definição matemática das variâncias genéticas aditiva e
dominante
Partindo-se da frequência alélica p=q=0,5, para o loco
B(b), no equilíbrio teremos:
Genótipo Frequência Valor genotípico
BB 1/4 m + a
Bb 1/2 m + d
bb 1/4 m - a
Valores genotípicos (q2) (2pq) (p2) bb m Bb BB -a +a d BB = m + a (p2) Grau médio de dominância = gmd = d/a
Bb = m +d (2pq)
bb = m – a (q2) Média da pop. = g = m+a(p-q)+2pqd
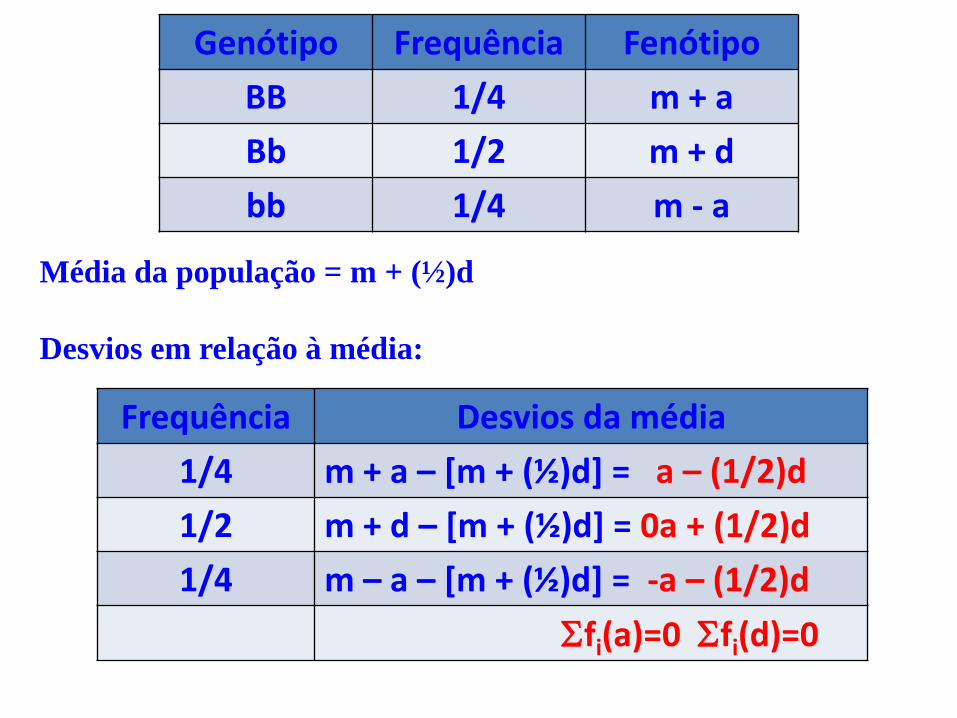
Genótipo Frequência Fenótipo
BB 1/4 m + a
Bb 1/2 m + d
bb 1/4 m - a
Média da população = m + (½)d
Desvios em relação à média:
Frequência Desvios da média
1/4 m + a – [m + (½)d] = a – (1/2)d
1/2 m + d – [m + (½)d] = 0a + (1/2)d
1/4 m – a – [m + (½)d] = -a – (1/2)d
fi(a)=0 fi(d)=0

Frequência Desvios da média
1/4 m + a – [m + (½)d] = a – (1/2)d
1/2 m + d – [m + (½)d] = 0a + (1/2)d
1/4 m – a – [m + (½)d] = -a – (1/2)d
fi(a)=0 fi(d)=0
Variância dos efeitos aditivos:
= (1/4)a² + (1/2)(0a²) + (1/4)(-a)² = (1/2)a²
Variância dos efeitos de dominância:
= (1/4)[(-1/2)d]² + (1/2)[(1/2)d]² + (1/4)[-(1/2)d]² = (1/4)d²
Para n locos teremos:
2
A
2
D
22
2
1iA
a 22
4
1iD d

Componentes da variâncias genética em uma geração F2
Genótipos Frequência Valores genotípicos
BB 1/4 m + a
Bb 1/2 m + d
bb 1/4 m – a
dmamdmamF2
1)(
4
1)(
2
1)(
4
12
2222 )]2
1([
4
1)]
2
1([
2
1)]
2
1([
4
12
dmamdmdmdmamGF
222
4
1
2
12
daGF
222
2 DAGF

Componentes da variâncias genética em retrocruzamentos
222
1 1))](
2
1([
2
1))](
2
1([
2
1damamdamdm
GRC
addaGRC
2
1
4
1
4
1222
1 1
Genótipos Frequência Valores genotípicos
Bb 1/2 m + d
bb 1/2 m – a
)(2
1)(
2
1)(
2
11 1
damdmamCR
RC11 Bb x bb

222
2 1))](
2
1([
2
1))](
2
1([
2
1damdmdamam
GRC
addaGRC
2
1
4
1
4
1222
2 1
Genótipos Frequência Valores genotípicos
BB 1/2 m + a
Bb 1/2 m + d
)(2
1)(
2
1)(
2
12 1
damdmamCR
RC21 Bb x BB

addaGRC
2
1
4
1
4
1222
1 1
addaGRC
2
1
4
1
4
1222
2 1
222
2 1
2
1 1
2
1
2
1da
GRCGRC
222
2 1
2
1 12
DAGRCGRC

Freq. Progênies Valor genotípico
1/4 (1/2) BB +(1/2)Bb m + (1/2)(a + d)
1/2 (1/4) BB + (1/2) Bb + (1/4) bb m + (1/2)d
1/4 (1/2) Bb +(1/2)bb m + (1/2)(d – a)
Componentes de variância entre progênies de meios
irmãos (PMI)
População em equilíbrio: (1/4) BB; (1/2) Bb; (1/4) bb
Plantas mães Pólen
(1/4) BB
(1/2) Bb (1/2) B; (1/2) b
(1/4) bb

Freq. Progênies Valor genotípico
1/4 (1/2) BB +(1/2)Bb m + (1/2)(a + d)
1/2 (1/4) BB + (1/2) Bb + (1/4) bb m + (1/2)d
1/4 (1/2) Bb +(1/2)bb m + (1/2)(d – a)
)](4
1)(
2
1)(
4
1[
2
1)](
2
1)(
2
1[
4
1amdmamdmamIMP
)](2
1)(
2
1[
4
1amdm
dmIMP2
1

dmIMP2
1
22 )]2
1()(
2
1)(
2
1[
4
1dmdmam
PM I
2)]2
1()(
4
1)(
2
1)(
4
1[
2
1dmamdmam
2)]2
1()(
2
1)(
2
1[
4
1dmamdm
²8
12 aPM I 22
4
1APM I

Componentes de variância entre progênies de irmãos
germanos (PIG)
(1/4) BB (1/2) Bb (1/4) bb
(1/4) BB (1/16) BBxBB (1/8) BBxBb (1/16) BBxbb
(1/2) Bb (1/8) BbxBB (1/4) BbxBb (1/8) Bbxbb
(1/4) bb (1/16) bbxBB (1/8) bbxBb (1/16) bbxbb
População em equilíbrio: (1/4) BB; (1/2) Bb; (1/4) bb
Serão possíveis as seguintes progênies de IG:

Freq.
Cruz. Progênie Valores genotípicos
1/16 BBxBB BB m + a
4/16 BBxBb (1/2)BB + (1/2)Bb m + (1/2)a +(1/2)d
2/16 BBxbb Bb m + d
4/16 BbxBb (1/4)BB + (1/2)Bb + (1/4)bb m + (1/2)d
4/16 Bbxbb (1/2)Bb + (1/2)bb m - (1/2)a +(1/2)d
1/16 bbxbb bb m - a
)(16
2)]
2
1
2
1(
16
4)(
16
1dmdamamGIP
)(16
1)
2
1
2
1(
16
4)]
2
1(
16
4amdamdm
dmGIP2
1

dmGIP2
1
222 )]2
1(
2
1
2
1[
16
4)]
2
1()[(
16
1dmdamdmam
PIG
22 )]2
1(
2
1[
16
4)]
2
1([
16
2dmdmdmdm
22 )]2
1(
2
1
2
1[
16
4)]
2
1([
16
1dmdamdmam
22
16
1²
4
1da
PIG 222
4
1
2
1DAPIG

Componentes de variância entre progênies S1
Genó- tipos
Freq. Progênie S1 Valores
genotípicos
BB 1/4 BB m + a
Bb 1/2 (1/4)BB + (1/2)Bb + (1/4)bb m + (1/2)d
bb 1/4 bb m – a
)(4
1)
2
1(
2
1)(
4
11
amdmamS
dmS4
11

dmS4
11
222
1)]
4
1(
2
1[
2
1)]
4
1([
4
1dmdmdmam
S
2)]4
1([
4
1dmam
22
1
16
1²
2
1da
S
222
1
4
1DAS

22
4
1AEM I
222
4
1
2
1DAEIG
222
1
4
1DAES
222
2
16
3
2
3DAS
22 2 AS
222
4
3DADM I
222
4
3
2
1DADIG
222
1
2
1
2
1DADS
222
2
4
1
4
1DAd S
02 S

Autógamas ou seguidas autofecundações em alógamas
L1 x L2
F1
F2 (equivale a S0)
F3 (equivale a S1)
F4 (equivale a S1)
F∞ (equivale a S∞)
222
2 DAGF
222
1
4
1DAS
22 2AS
222
2
16
3
2
3DAS

Interpretação do QM residual
I II III TT
1 1 5 3 9
2 4 8 6 18
3 2 6 4 12
4 3 7 5 15
TB 10 26 18 54
Fontes de Variação G.L SQ QM F
Blocos 2 32 16 ∞
Tratamentos 3 15 5 ∞
Resíduo 6 0 0
Total 11 47

Análise em nível de indivíduos
Bloco I Bloco II TP
1 2 4 4 (10) 4 5 6 (15) 25
2 7 5 4 (16) 7 9 9 (25) 41
3 8 4 4 (16) 7 5 5 (17) 33
4 3 5 7 (15) 6 5 4 (15) 30
5 6 7 8 (21) 8 6 5 (19) 40
TB 78 91 169

F. V. G.L. SQ QM E(QM)
Blocos 1 5,6333 5,6333 ----
Progênies 4 30,4667 7,6167
Resíduo 4 12,8667 3,2167
Dentro 20 42,0000 2,1000
TOTAL 29 90,9667
222
pedkrk
22
edk
2
d
100,22 d
3722,02 e
7333,02 p
2055,32 F
2694,12 F
9332,22 A
9150,0ˆ h
5777,0ˆ m
h

Análise em nível de totais de parcelas
Bloco I Bloco II TP
1 10 15 25
2 16 25 41
3 16 17 33
4 15 15 30
5 21 19 40
TB 78 91 169

F. V. G.L. SQ QM E(QM)
Blocos 1 16,9 16,9 ------- -------
Progênies 4 91,4 22,85
Resíduo 4 38,6 9,65
TOTAL 9 146,9
22
pr
2
75,92
55,62 p
2,262 A
3,162 F
425,112 F
5733,0ˆ
mh
)( 22222
pedrkkk
)( 222
edkk

Análise em nível de médias de parcelas
Bloco I Bloco II TP
1 3,3333 5,0000 8,3333
2 5.3333 8,3333 13,6666
3 5,3333 5,6666 11,0000
4 5,0000 5,0000 10,0000
5 7,0000 6,3333 13,3333
TB 26,0000 30,3333 56,3333

F. V. G.L. SQ QM E(QM)
Blocos 1 1,8778 1,8778 ------- -------
Progênies 4 10,1555 2,5389
Resíduo 4 4,2889 1,0722
TOTAL 9 16,3222
22
pr
2
0722,12
7333,02 p
9333,22 A
2693,12 F
5733,0ˆ m
h
)/( 222
pedrk
)/( 22
edk

CORRELAÇÃO ENTRE CARACTERES
Associação entre caracteres Covariância
•Medida da associação ou semelhança entre variáveis;
•Se for maior que zero associação positiva;
•Se for igual a zero ausência de associação;
•Se for menor que zero associação negativa.


•Percebe-se que a variância é um caso particular de
covariância de uma variável com ela mesma;
•A covariância tem um papel fundamental na seleção,
pois interessa a associação entre pais e descendentes;
•A semelhança entre genitores e descendentes garante
o progresso na seleção e pode ser medida pela
covariância;
•A covariância varia de - ∞ a + ∞;

•Por isso o parâmetro mais usado é o coeficiente de
correlação (r), que varia de -1 a +1.
•r expressa o efeito total dos genes em segregação. Alguns
podem aumentar ambos os caracteres, enquanto outros
aumentam um e reduzem outro;
•Exemplo: Genes que aumentam a taxa de crescimento
aumentam tanto a estatura como a massa, mas genes que
aumentam a gordura influenciam apenas a massa e não
são causa de correlação com a estatura.

Uso e importância no melhoramento
•Cuidado para evitar mudanças indesejáveis em um
caráter ao selecionar outro;
•Seleção indireta – quando a correlação for favorável e um
caráter possui herdabilidade muito baixa em relação ao
outro e/ou tenha problemas de medição e identificação;

Componentes da covariância
•Da mesma maneira que temos variâncias fenotípica,
genética, genética aditiva e ambiental, temos também as
covariâncias fenotípica, genética, genética aditiva e
ambiental;
•Apenas a covariância genética envolve uma associação de
natureza herdável, podendo ser utilizada na orientação de
programas de melhoramento;
•O ambiente torna-se causa de correlações quando dois
caracteres são influenciados pelas mesmas diferenças de
condições ambientais;

•Correlações ambientais negativas indicam que o ambiente
favorece um caráter e desfavorece outro;
•Correlações ambientais positivas indicam que os dois
caracteres são e beneficiados ou prejudicados pelas
mesmas causas de variação ambiental;
•Da mesma maneira que os demais parâmetros, os valores
de covariância e correlação são específicos para uma
população e um determinado ambiente.

Causas genéticas da covariância e correlação
•Pleiotropia – Situação em que locos atuam em dois ou
mais caracteres ao mesmo tempo (correlação permanente);
•Ligação gênica – Situação em que os locos atuam em
caracteres diferentes, mas estão próximos no mesmo
cromossomo (correlação não permanente, podendo ser
quebrada).

Decomposição das correlações fenotípica e genética
•Necessidade de esquema experimental como feito na
decomposição da variância fenotípica e genética;
•Os pacotes estatísticos já fazem a decomposição como
feito na variância.
Covariância (Produto Médio ) entre duas variáveis x e y
Método braçal
•Possível com a análise de variância de x, y e de uma
variável construída (x+y)

F. V. G. L. QMx QMy QM(x+y)
Blocos r-1 QMBx QMBy QMB(x+y)
Progênies t-1 QMPx QMPy QMP(x+y)
Resíduo (r-1)(t-1) QMRx QMRy QMR(x+y)
Dentro rt(k-1) QMDx QMDy QMD(x+y)
PMBxy = COVBxy = (1/2)(QMB(x+y) - QMBx - QMBy)
PMPxy = COVPxy = (1/2)(QMP(x+y) - QMPx - QMPy)
PMRxy = COVRxy = (1/2)(QMR(x+y) - QMRx - QMRy)
PMDxy = COVDxy = (1/2)(QMD(x+y) - QMDx - QMDy)

F. V. G. L. PMxy E(PM)
Blocos r-1 PMB -----------
Progênies t-1 PMP
Resíduo (r-1)(p-1) PMR
Dentro rp(k-1) PMD
pEdkrCOVkCOVCOV
edkCOVCOV
DCOV
PMDVOCdˆ
kPMDPMRVOCe
/ˆ
rPMRPMPVOCp
/)(ˆ
pAVOCVOC ˆ4ˆ
depFVOCVOCVOCVOC ˆˆˆ
rkVOCrVOCVOCVOCdepF/ˆ/ˆˆˆ

yx
xy
FF
F
F
QMPQMP
PMPVOCr
yx
xy
xy
..
ˆ
22
22 .
ˆ
yx
xy
xy
FF
F
F
VOCr
yx
xy
ee
e
e
QMRQMR
PMRVOCr
xyxy
xy
xy
..
ˆ
22
Coeficiente de correlação fenotípica
Coeficiente de correlação fenotípica média
Coeficiente de correlação ambiental

22 .
ˆ
yx
xy
xy
AA
A
A
VOCr
yxy
x
xy
x
xy
AAx
F
A
F
A
xyrhki
VOCki
VOCkdsRC
....
ˆ
.
ˆ
.2
2/
Coeficiente de correlação genética aditiva
Resposta correlacionada à seleção
•k=1, para seleção massal em ambos os sexos;
•k=1/2, para seleção massal em um sexo;
•k=1/4, para seleção entre MI em ambos os sexos;
•k=1/8, para seleção entre MI em um sexo.

ANÁLISES CONJUNTAS E
AGRUPADAS

Análise individual
Fontes de variação G.L. Q.M. F
Blocos r-1 QMB QMB/QMR
Tratamentos t-1 QMT QMT/QMR
Progênies p-1 QMP QMP/QMR
Testemunhas T-1 QMTe QMTe/QMR
Prog. vs Testem. (p-1)(T-1) QMPT QMPT/QMR
Erro (t-1)(r-1) QMR
Dentro de progênies pr(k-1) QMD
ANÁLISE AGRUPADA (modelo aleatório)

ANÁLISE AGRUPADA (modelo aleatório)
Fontes de Variação G.L. Q.M. E(QM)
Blocos/Experim. e(r-1)
Experimentos e-1 ---- ----
Progênies e(p-1) QMP
Erro médio e(p-1)(r-1) QMR
Dentro de progên. epr(k-1) QMD
22
5/ ed
2
d
222
5/ pedr

ANÁLISE CONJUNTA
Modelo aleatório

ANÁLISE CONJUNTA
Modelo fixo, exceto para blocos e erros

ANÁLISE CONJUNTA
Modelo aleatório para genótipos e fixos para locais

ANÁLISE CONJUNTA
Modelo aleatório com três fatores

ANÁLISE CONJUNTA
Modelo fixo com três fatores

PREDIÇÃO DA MÉDIA DE UM
CARÁTER EM POPULAÇÕES
OBTIDAS POR CRUZAMENTOS

PREDIÇÃO
Interessante pois, na maioria dos casos, o
número possíveis de híbridos, compostos e
sintéticos é extremamente elevado, sendo
impraticável obter todos e avaliá-los em
experimentos.

COMPOSTOS
• São materiais variáveis geneticamente,
formados pelo intercruzamento (recombinação)
de variedades, híbridos ou raças distintas, e
servem de base para o início de um processo de
seleção;
•Com n parentais é possível formar 2n – (n+1)
compostos diferentes.

SINTÉTICOS
• São materiais variáveis formados pelo
intercruzamento (recombinação) de linhagens e
servem como base para seleção, para uso
comercial direto ou para extração de novas
linhagens;
• Com n linhagens é possível formar 2n – (n+1)
sintéticos diferentes.

Com n linhagens diferentes poderemos obter:
híbridos simples (LA x LB);
híbridos duplos [(LA x LB) x (LC x LD);
híbridos triplos [(LA x LB) x LC].
2
nC
43n
C
33n
C

Com n linhagens de uma população A e p
linhagens de uma população B poderemos
obter:
np híbridos simples;
híbridos duplos;
híbridos triplos.
22
pnxCC
22
nppCnC

Processo de predição baseia-se em Mendel, Genética de
populações e Quantitativa. Considerando apenas um
loco, com presença de heterose, podemos prever a média
da geração F2:
A população F2 é composta por 25% de AA (P2), 50% de
Aa (F1) e 25 % de aa (P1).
AA = 10 unidades aa = 2 unidades; Aa = 12 unidades
2P P 1P 1F
h
A população F2 é composta por 25% de AA (P2), 50% de Aa (F1) e 25 % de aa (P1).
0,9)2(4
1)12(
2
1)10(
4
12 F hP
PFPF
2
1
4
2 2112
0,9)2(4
1)12(
2
1)10(
4
12
F
hPPFP
F2
1
4
2211
2

•Considerando que as médias dos pais e de F1 foram
obtidas de um grande número de repetições, o
ambiente terá pouca influência e a média de F2 será
bem estimada.
•A média F2 depende da frequência de cada material
parental na população final. Portanto a seguinte
expressão geral pode ser usada para qualquer tipo de
parental:
])([)]([]].[[11
i
ii
j
jjPfxPfYXM

Tanto os híbridos (triplos e duplos) como os compostos
e sintéticos podem ser preditos a partir das médias dos
parentais e dos seus cruzamentos simples.
Exemplo 01
Média de um híbrido duplo [(LA x LB) x (LC x LD)]
)](
2
1)(
2
1[)](
2
1)(
2
1[ DCxBADH
ABxCD
)(4
1DBCBDACADH
ABxCD

Exemplo 02
Média da F2 do híbrido duplo [(LA x LB) x (LC x LD)]
)](4
1)(
4
1)(
4
1)(
4
1[)](
4
1)(
4
1)(
4
1)(
4
1[
2DCBAxDCBAF
2
2)](
4
1)(
4
1)(
4
1)(
4
1[ DCBAF
)(16
2)](
16
12
DBDCCBDACABADCBAF

Exemplo 03
Composto com duas variedades (V1 e V2)
](2
1)(
2
1[)](
2
1)(
2
1[
21211 2VVxVVOC
4
22121
1 2
VVVVOC

Exemplo 04
Composto com três variedades (V1, V2 e V3)
)](3
1)(
3
1)(
3
1[)](
3
1)(
3
1)(
3
1[
3213211 2 3VVVxVVVOC
9
222323121321
1 2 3
VVVVVVVVVOC

Generalizando para n parentais teremos:
Como:
)](1
.......)(1
)(1
)(1
[)](1
.......)(1
)(1
)(1
[321321 nn
Vn
Vn
Vn
Vn
xVn
Vn
Vn
Vn
OC
).........................(2
123212122
321nnnn
n VVVVVVVVVVnn
VVVVOC
2nC F1s ou híbridos
2
1232121
1
).........................(
n
nnnn
C
VVVVVVVVVVF
Fnn
FxCVVVVVVVVVVnnnnn
2
)1(.........................
1
2
1232121

2
1232121
1
).........................(
n
nnnn
C
VVVVVVVVVVF
Fnn
FxCVVVVVVVVVVnnnnn
2
)1(.........................
1
2
1232121
1
11F
n
nP
nOC
hn
nPOC
1
)(1
11PF
nFOC

Predição de compostos ou sintéticos com participações
desiguais dos parentais
Utilizar a fórmula original
Exemplo
•Introgressão de 3 variedades em uma outra, de
maneira que o material resultante contenha 1/2 do
germoplasma da variedade original (V0) e a outra
metade seja composta por 1/3 de V1, 1/3 de V2 e 1/3 de
V3.
•Para isso há necessidade do cruzamento de V0 com as
outras três e a recombinação dos híbridos resultantes
(V0 x V1, V0 x V2 e V0 x V3).

V1 V2 V3
V0 10VV 20VV 30VV
)]3
1
3
1
3
1(
2
1
2
1[)]
3
1
3
1
3
1(
2
1
2
1[
32103210VVVVxVVVVOC
)]6
1
6
1
6
1
2
1[)]
6
1
6
1
6
1
2
1[
32103210VVVVxVVVVOC

Predição do cruzamentos de compostos
•Auxilia na identificação de pares de compostos que
mostram heterose, visando retirada de linhagens para
produção de híbridos ou para início de programa de
Seleção Recorrente Recíproca;
•Número de Pares de Compostos = 2
)]1(2[ nnC

Supondo um Composto A com n variedades e outro B
com p variedades teremos:
)].....(1
[)].....(1
['3'2'1321 pnBA
VVVVp
xVVVVn
OCxOC