atributos funcionais morfológicos e anatômicos foliares ...‡Ão... · paliçádico, espessura...
TRANSCRIPT

Universidade Federal de Ouro Preto – UFOP
Instituto de Ciências Exatas e Biológicas – ICEB
Programa de Pós-Graduação em Ecologia de Biomas Tropicais
______________________________________________________________________
Dissertação de Mestrado
Denny Fernandes Eduardo
Atributos funcionais morfológicos e anatômicos foliares de
plantas de campo rupestre ferruginoso respondem
abundancia?
Ouro Preto 2017

Denny Fernandes Eduardo
Atributos funcionais morfológicos e anatômicos foliares de
plantas de campo rupestre ferruginoso respondem
abundancia?
Dissertação apresentada ao Departamento de
Evolução, Biodiversidade e Meio Ambiente do
Instituto de Ciências Exatas e Biológicas da
Universidade Federal de Ouro Preto como
requisito parcial para obtenção do título de Mestre
em Ecologia de Biomas Tropicais.
Orientador: Dr. Hildeberto Caldas de Sousa
Co-orientadora: Dra. Alessandra Rodrigues Kozovits
Ouro Preto, Minas Gerais, Brasil
Novembro de 2017



Epígrafe_______________________________________________Eduardo,D,F.2017
When your intelligence has passed out of the dense forest of delusion, you shall become
indiferente to all that has been heard and all that is to be heard
Bragavad Gita As It Is. His Divine Grace A. C. Bhaktivedanta Swami Prabhupada

Dedicatória
“Ao meu querido avô Sebastião Eduardo”

Agradecimentos
Agradeço esse trabalho primeiramente ao meu pai, minha mãe e minha irmã que me apoiaram
financeiramente durante as fases mais difíceis no começo do mestrado. E com muito carinho
ao meu avô pelos conselhos na hora certa. Com certeza sem vocês eu não teria conseguido
chegar aonde cheguei.
Quero agradecer a Alaine por me apresentar o lindo Morro São Sebastião onde morei o tempo
todo do meu mestrado.
Aos moradores do sítio, Leonardo, Ricardo, Igor, Alice pela convivência.
Aos ex-moradores da casa verde, Lucas, Arthur e Zito por me acolherem em tempo de
mudança na minha vida.
As Repúblicas Casa Blanca e Jardim zoológico pela recepção e acolhimento em horas
descontraídas.
Aos meus amigos Naaman e Tamires pelos ensinamentos yogis, que foram fundamentais para
eu conseguir-me concentrar para a realização deste trabalho.
Ao Bruno (polegada) pelos ensinamentos sobre taxonomia de plantas, etnobotânica e
medicina caseira. Nunca vou esquecer-me dos nossos rocks, que muitas vezes foram aleatórios
pela maravilhosa cidade de Ouro Preto.
A colega de mestrado Isabela Botelho por me ajudar no laboratório nas contagens dos
estômatos.
Quero agradecer a Marina Beirão, Fernanda, Grazi, Claúdio e professor Marcão pela ajuda
nas análises estatísticas.
Aos meus alunos de iniciação científica, Igor, Yuri, Francis, Julia Celane e em especial
Rafaela Lameira. Sem vocês com certeza este trabalho se tornaria inviável.
A Regislane pelos ensinamentos metodológicos em anatomia vegetal e pelos conselhos.
Ao Claúdio, técnico do laboratório, por me ajudar a me organizar para realizar os
experimentos.
A Deborah, técnica do Herbário, pela leitura da tese e sugestões.

A professora Maria Cristina Messias por me ajudar em um dos momentos de escolhas
importantes que passei na UFOP e me mostrar outros caminhos que eu pude seguir.
Ao meu caro orientador Hidelberto pelo seu carisma, acolhedor amigável. Na maioria dos dias
com um sorriso estampado no rosto e mesmo em dias turbulentos, cantarolando sambas
antigos pelos corredores da Universidade. Além do árduo trabalho, tivemos muitas conversas
descontraídas no laboratório durante os cafezinhos. Temos muitas histórias para contar! Muito
obrigado pelos ensinamentos sobre a vida e botânica. Fez-me mudar o jeito de ver e as plantas
e de gostar ainda mais destes organismos fantásticos.
A minha Co-orientadora Alessandra Kozovits pelo apoio e confiança no meu projeto.
A agência de fomento CAPES pela concessão da bolsa de Mestrado.
Ao NUPEB pela possibilidade de realizar a fotodocumentação.
Ao IEF pela disponibilização da licença para as coletas das plantas.
Ao Programa de Ecologia de Biomas Tropicais, em especial ao secretário Rubens Modesto,
que foi muito receptivo nas horas que mais precisei.

Sumário
Atributos funcionais morfológicos e anatômicos foliares de plantas de campo rupestre
ferruginoso respondem abundância? ........................................................................................ 1
Resumo ........................................................................................................................ 1
Introdução.................................................................................................................... 1
Material e métodos ...................................................................................................... 4
Análises estatísticas ..................................................................................................... 7
Atributos funcionais entre espécies mais e menos abundantes ................................... 7
Atributos funcionais morfológicos e fisiológicos por espécies ................................... 8
Características anatômicas foliares por espécies ......................................................... 9
Analise dos componentes principais ......................................................................... 11
Discussão ................................................................................................................... 12
Atributos funcionais fisiológicos e morfológicos ..................................................... 12
Atributos funcionais anatômicos foliares .................................................................. 20
Atributos funcionais que podem estar relacionados com a absorção e manutenção de
água e diminuição da temperatura foliar ................................................................... 21
Convergência evolutiva de atributos funcionais que podem estar relacionados com a
absorção e manutenção de água e diminuição da temperatura foliar ........................ 22
Densidade e tamanho dos estômatos ......................................................................... 22
Agrupamentos de características funcionais anatômicas e morfológicas das espécies
mais abundantes (PCA) ............................................................................................. 23
Agrupamentos de características funcionais anatômicas e morfológicas das espécies
menos abundantes (PCA) .......................................................................................... 24
Estrutura da comunidade de plantas .......................................................................... 24
Conclusão .................................................................................................................. 24
Referências bibliográficas ......................................................................................... 25

1
____________________________________________________________
Atributos funcionais morfológicos e anatômicos foliares de plantas de
campo rupestre ferruginoso respondem abundância?
Eduardo, D, F ¹, Sousa, H, C², Kozovits, A, R³
Instituto de ciências exatas e biológicas. Programa de Pós-Graduação em Ecologia de Biomas Tropicais. Universidade Federal de Ouro Preto, Minas
Gerais, Brasil. Correspondência ([email protected])
Resumo: Atributos funcionais foliares são utilizados para um melhor esclarecimento sobre as adaptações que cada
espécie tem em cada ambiente específico, e podem indicar quais características são mais importantes em espécies mais e
menos abundantes em um determinado espaço e tempo. Diferentes histórias de vida das plantas podem ter sucesso em
um mesmo espaço e tempo, podendo selecionar adaptações distintas com relação à economia de água, e sua aquisição e
manutenção pelas folhas. Os campos rupestres ferruginosos são ambientes montanos que ocorrem acima de 900m e
impõem severidade ambiental nas plantas por estresse hídrico devido à marcada estação seca, alta radiação, exposição a
ventos e a ocorrência de solos rasos. É comum a ocorrência de neblina durante o ano todo, e as plantas podem absorver
água pelas folhas quando ocorrem este evento o qual pode influenciar nos níveis de esclerofilia das folhas. Para analisar
as adaptações que as plantas possuem neste tipo de ambiente, foram selecionadas cinco espécies mais abundantes,
Tibouchina heteromalla, Baccharis reticularia, Diplusodon buxifolius, Periandra mediterranea e Erytroxylum
mirophyllum, e cinco menos abundantes Myrsine squarrosa, Miconia coralina, Eremanthus incanus, Ouratea semiserrata
e Trembleya laniflora, a partir do trabalho de (Arias 2016). E foram analisados a área foliar, área foliar específica, massa
foliar específica, espessura do limbo foliar, espessura da cutícula, espessura da epiderme adaxial, espessura do parênquima
paliçádico, espessura do parênquima lacunoso, espessura da epiderme abaxial, espessura da cutícula abaxial, densidade e
área estomática. As espécies mais abundantes apresentaram áreas foliares reduzidas, maiores valores de área foliar
específica, menores valores de massa foliar específica, menor espessura do limbo foliar, menor espessura da cutícula
adaxial, maiores células epidérmicas na face adaxial, menor espessura do parênquima lacunoso, maior espessura da
epiderme abaxial e menor espessura da cutícula abaxial, maior densidade estomática e também maiores valores de área
estomática. Além disso, T. heteromalla, B. reticularia e D. buxifolius são anfiestomáticas tais características podem
melhorar o desempenho fotossintético, refletindo na maior produtividade e crescimento das plantas mais abundantes. Em
outro contexto, as espécies apresentaram convergência evolutiva em atributos que favorecem a absorção de água e
manutenção nas folhas quando ocorrem eventos de neblina, como pubescência pesada e células mucilaginosas, além de
atributos que podem estar relacionados à diminuição da temperatura da folha, como drusas de oxalato de cálcio. Indicando
que a neblina é um fator muito importante na seleção das espécies que ocorrem nos campos rupestres ferruginosos.
Palavras chave: Atributos funcionais foliares, área foliar específica, massa foliar especifica, anatomia foliar, atributos que
podem estar relacionados com a absorção de água nas folhas.
Introdução
Entender como fatores ambientais
influenciam a distribuição e o funcionamento
das plantas tem sido um dos maiores objetivos
em estudos de ecologia (Grime 1997; McGill
et al. 2006, Craine 2007; Castro 2015). Para
esta finalidade, uma abordagem bastante
utilizada é o estudo das características
ecológicas funcionais das plantas (McGill et

2
al. 2006), muitas vezes representadas por
atributos anatômicos e morfológicos foliares
(Reich et al. 1992, Rozendaal 2006, Viole
2007, Rossatto e Kolb 2013, Hulshof 2013).
Estas características morfológicas
externas e internas, juntamente com atributos
fisiológicos, são fundamentais para o
estabelecimento, crescimento, reprodução e
colonização das espécies em um determinado
ambiente (Diaz 1998; Viole 2007; Rossatto e
Kolb 2013) e podem interferir nos serviços
ecossistêmicos como ciclagem de nutrientes e
sequestro de carbono (Reich et al. 1997,
Cornwell 2008).
Os atributos funcionais presentes
atualmente nas plantas foram selecionados ao
longo do tempo evolutivo em resposta à
variação de inúmeras pressões ambientais
(Gratani 2014), como disponibilidade de
água, luz e nutrientes, temperatura,
herbivoria, competição e outras interações
bióticas e abióticas (Violle 2007). Na mesma
comunidade vegetal, sob a ação dos
condicionantes edafo-climáticos similares,
espécies co-ocorrentes podem diferir bastante
em termos de atributos morfológicos e
anatômicos devido a trade-offs específicos
entre demandas relacionadas à economia de
nutrientes e às características hidráulicas (Li
et al. 2015), e possivelmente à especialização
no uso de recursos (Tilman 2007), mas
também podem convergir em características
funcionais em determinadas situações
ambientais (Ackerly & Cornwell 2007).
Plantas mais abundantes em locais
com forte pressão seletiva possuem traços
funcionais que são capazes de tolerar, resistir
ou evitar a severidade ambiental, utilizar
nichos ecológicos vazios, o que pode evitar a
exclusão competitiva por sobreposição de
nichos (MacArthur 1972). Por outro lado,
plantas mais raras estão mais propensas à
extinção quando ocorrem eventos estocásticos
(Rabinowitz et al. 1986).
Buscando entender como as plantas de
diferentes táxons convergem ou divergem em
suas características funcionais em um
determinado espaço e tempo e como estes
atributos influenciam a distribuição das
espécies, desenvolveu-se este estudo em um
Campo Rupestre Ferruginoso. Estes locais
ocorrem acima de 900 m de altitude, em
ambientes expostos a ventos e à alta irradiação
solar total, incluindo as bandas de ultra-
violeta, com alta taxa de evapotraspiração,
marcante sazonalidade de chuvas,
apresentando manchas de solos distróficos e
rasos, entremeados por afloramentos rochosos
(Giulietti 1987, Rapini et al. 2008). A
vegetação é dominada por espécies herbáceas
e arbustivas, cuja distribuição parece ser
fortemente influenciada pela microtopografia
do relevo, que se reflete em um mosaico de
ambientes edafo-climáticos (Jacobi et al.
2008, Ataíde 2011, Messias et al., 2012;
Messias et al. 2013; Carmo & Jacobi 2013;
Carmo et al. 2016). Tais condições ambientais
e a própria estrutura da vegetação indicam que

3
a disponibilidade de água deve ser um dos
principais fatores limitantes para o
estabelecimento de espécies arbóreas
(Scarano 2002, Jacobi et al. 2007, Sarthou et
al. 2009) e sugerem a presença de atributos
funcionais relacionados à eficiência na
aquisição e na manutenção da água nos
tecidos vegetais (Li et al. 2015).
Certos atributos morfológicos e
anatômicos foliares favorecem a tolerância e
resistência das plantas em ambientes com
déficit hídrico, tais como: paredes celulares
reforçadas, redução dos espaços intercelulares
(De Micco e Aronne 2012, Ackerly e
Cornwell 2007), epiderme mais espessa na
face adaxial (uni- ou pluri-estratificada)
(Turner 1994, Fahn e Cutler 1992), maior
espessura do mesofilo (Rossato e Kolb 2013),
parênquima paliçádico com mais de uma
camada de células, podendo ser encontrado
em ambos os lados da folha (simetria
isobilateral) (Marques 2000 e Raven 2001),
presença de células mucilaginosas, paredes
celulares lignificadas, folhas anfiestomáticas
(Fahn e Cutler 1992, Parkhurst 1978, Moot
1982) estômatos protegidos por tricomas
(Morretes 1969), maior densidade dos
estômatos e menor tamanho das células-
guarda (Beaulieu 2008, Franks et al. 2009,
Drake 2013), folhas com tamanho reduzido
(Gratani 2014), camada de cutícula espessa
(Larcher 2004), menor área foliar específica
(Wrigth et al, 2004). Além dos
atributos foliares relacionados à minimização
da perda de água que caracterizam níveis de
esclerofilia, estudos recentes chamam a
atenção para a importância de atributos
anatômicos que podem facilitar a captação de
água pelas folhas e sua manutenção nos
tecidos, como tricomas, células
mucilaginosas, cristais de oxalato de cálcio,
fibras lignificadas ou gelatinosas, cavidades
secretoras e maior extensão da bainha do feixe
vascular (Souza 2014).
Locais periodicamente sujeitos a
eventos de neblina, como tipicamente
ocorrem nos campos rupestres (Baeta 2012),
podem ter uma fonte adicional de água para a
vegetação, especialmente durante a estação
seca, minimizando o déficit hídrico (Simonin
2009; Eller 2013). Nos campos rupestres
ferruginosos, o déficit hídrico pode ter
selecionado atributos ao longo do processo
evolutivo das espécies para evitar a perda de
água. Concomitantemente, a presença de
neblina deve ter favorecido a seleção de
atributos que favoreçam a captação e
armazenamento de água nos tecidos foliares,
minimizando o déficit hídrico. Seria possível
separar atributos mais importantes para a
realização de cada uma dessas funções? Qual
o conjunto de atributos anatômicos e
morfológicos (para evitar a perda ou aumentar
a captação e manutenção de água na folha) é
dominante na comunidade de plantas dos
campos rupestres ferruginosos? No presente
trabalho, atributos anatômicos e morfológicos
foram mensurados em conjuntos de espécies

4
mais abundantes e mais raras em um
fragmento de campo rupestre ferruginoso,
localizado no município de Ouro Preto, Minas
Gerais. O objetivo deste trabalho foi verificar
a existência de padrões de agrupamentos de
atributos funcionais morfológicos e
anatômicos foliares que possam se relacionar
com a abundância de espécies em uma área de
campo rupestre ferruginoso. Tem-se como
primeira hipótese que as espécies mais
abundantes no Campo Ferruginoso na Serra
da Brígida não divergem muito em
agrupamentos de atributos funcionais
morfológicos e anatômicos das plantas menos
abundantes. A segunda hipótese é que
espécies de diferentes famílias possuem uma
convergência evolutiva adaptativa em
características anatômicas que podem estar
relacionadas com a captação e manutenção da
água pelas folhas como mucilagens, tricomas,
emergências e com a diminuição da
temperatura foliar, como cristais de oxalato de
cálcio e cavidades secretoras.
Material e métodos
A área de campo rupestre ferruginoso estudada
está inserida na Área de Proteção Ambiental
Estadual da Cachoeira das Andorinhas, na serra da
Brígida (20°21’35’’S, 43°30’11’’W), município
de Ouro Preto em Minas Gerais, Quadrilátero
Ferrífero, Brasil. A serra da Brígida possuí uma
área de 138,3ha (UFOP 2001), com amplitude de
altitude entre 1470 a 1500m, e o clima é Cwb,
temperado quente (úmido mesotérmico), segundo
a classificação de Köppen, apresentando estação
chuvosa de outubro a março e seca de abril a
setembro (Alvares et al. 2013). É comum a
ocorrência neblina inclusive na estação seca
(Baeta 2012, Souza 2014). A vegetação local é
composta principalmente de plantas herbáceas e
arbustivas, com alta taxa de endemismo, comum
em afloramentos rochosos nos campos rupestres
ferruginosos (Valim 2012, Messias 2013, Arias
2016).
Coleta dos dados Amostrais
No estudo realizado por Arias (2016) em 115
parcelas de 10x10m², pode ser averiguado que
Tibouchina heteromalla (D. Don) Cogn.
(Melastomataceae) tem 100% de abundância,
seguida por Baccharis reticularia (Vell.) Taub.
(Asteraceae) (98%), Diplusodon buxifolius
(Cham. & Schtdl) DC. (Lythraceae) (96%),
Periandra mediterranea (Vell. Taub.
(Leguminoase) (96%), Erythroxylum
microphyllum A.St.-Hil. (Erytroxylacea) (85%),
compondo o grupo de plantas mais abundantes,
enquanto, Myrsine squarrosa (Mez) M.F.Freitas &
Kin.-Gouv (Primulaceae) (41%), Miconia
corallina Spring (Melastomataceae) (35%),
Eremanthus incanus (Less.) Less. (Asteraceae)
(33%), Ouratea semiserrata (Mart. & Nees) Engl.
(Ochnaceae) (28%) e Trembleya laniflora (D.
Don) Cogn. (Melastomataceae) (23%), compõem
o grupo de espécies menos abundantes. Ao longo
de um transecto de 200m todos indivíduos das
espécies citadas acima foram marcados e sete
indivíduos das espécies mais abundantes e sete das

5
menos abundantes foram sorteadas. Dois ramos
contendo estruturas reprodutivas de cada espécie
foram coletados para a preparação exsicatas e
foram depositadas no Herbário Prof. José Baldini
na Universidade Federal de Ouro Preto.
Área foliar específica e massa foliar específica
Dez folhas de cinco indivíduos de cada espécie
foram coletadas e escaneadas em laboratório em
uma impressora multifuncional HP PSC 15-10,
para mesurar a área foliar, utilizando-se o
programa ImageJ Pro 4.0 (fig. 1). Posteriormente
as folhas foram secadas em uma estufa a 70ºC
durante três dias e pesadas em uma balança de
precisão. A área foliar específica foi determinada
pela razão entre a área e a massa foliar. A massa
foliar específica foi determinada pela razão entre a
massa e a área foliar (Reich 1997).
Anatomia foliar
Lâminas histológicas foram preparadas no
Laboratório de Anatomia Vegetal do
Departamento de Biodiversidade Evolução e
Ambiente (DEBIO/UFOP) para a caracterização
anatômica das plantas selecionadas. Sete folhas de
sete indivíduos das cinco espécies mais
abundantes e das cinco das mais raras foram
coletadas e fixadas em FAA 50% (Formaldeído a
37- 40%, etanol 50% e ácido acético PA, nas
proporções 5%: 5%: 90% respectivamente),
posteriormente foram submetidas a vácuo durante
48h e preservadas em álcool 70% (Johansen 1940).
Depois foram submetidas a vácuo durante 48h e
preservadas em álcool 70%. Foram feitos cortes
manuais transversais do limbo foliar na porção
mediana, na região da nervura central, e entre a
parte mediana e a margem, utilizando lâmina de
barbear. Os cortes foram corados com azul de astra
e fucsina básica e montados em glicerina a 50%.
Para obter cortes seriados foi utilizado a mesma
metodologia de fixação do material, posterior
desidratação pela série etanolica, e inclusão em
parafina (Kraus & Arduin, 1997) da parte mediana
e entre a parte mediana e a margem foliar. Foram
feitos cortes de 12 µm de espessura em um
micrótono rotativo, os quais foram reidratados e
corados com azul de astra e fucsina básica,
desidratados e montados em balsamo do Canadá.
A fotodocumentação foi realizada
utilizando o programa Leica application suite 4V.3
no Núcleo de Pesquisas em Ciências Biológicas na
UFOP (NUPEB). Posteriormente, utilizando o
Software Axio Vision, Zeiss imaging Sistems
(Zeiss 2008), foram feitas 10 medidas, em sete
cortes de sete folhas de cada espécie da espessura
da cutícula e das células da face adaxial da
epiderme, espessura do parênquima paliçádico,
espessura do parênquima lacunoso, espessura das
células da epiderme da face abaxial e da cutícula
da face abaxial. Também foi registrada se o
parênquima lacunoso é organizado de forma
conspícua ou inconspícua, e de características
funcionais que podem estar relacionadas com a
captação de água pelas folhas e diminuição da
temperatura foliar como, presença de tricomas,
emergências, cavidades secretoras, células
mucilaginosas e cristais de oxalato de cálcio.

6
Figura 1: Folhas das espécies mais abundantes: (A) T. heteromalla, (B) B. reticularia, (C) D. buxifolius (D)
P. mediterrânea (E) E. microphyllum e das espécies menos abundantes: (F) M. squarrosa (G) O.
semiserrata (H), E. incanus, (I) M. corallina, (J) T. laniflora ( 1 cm). Visão da face adaxial.
G
0
0
H
0
0
J
0
0
F
0
0
I
B
A
C
0
0
D
0
0
E
Espécies mais abundantes
Espécies menos abundantes

7
Densidade e tamanho dos estômatos
As epidermes de quatro folhas de cada espécie
foram dissociadas pela solução de Jeffey (ácido
nítrico 10% e ácido crômico 10% 1/1 v/v)
(Johansen 1940), lavadas e posteriormente coradas
com Azul de Astra e Fucsina Básica (Kraus &
Arduin, 1997). Após este procedimento as
amostras foram montadas em gelatina glicerinada
de Kaiser 50% (Johansen 1940). Posteriormente
foram feitas microfotografias da face abaxial e
adaxial de cada espécie utilizando o software Leica
application suite 4V.3 no (NUPEB). A densidade
estomática (número de estômatos por unidade e
área) determinada pela média do número de
células estomáticas encontradas em 10 campos de
visualização (0,30 mm² em cada campo de
visualização). Para a contagem foi utilizado o
programa Adobe Photoshop versão 2015. A área
estomática foi medida em micrometros quadrados
utilizando o Software Axio Vision, Zeiss imagin
sistems (Zeiss 2008).
Análises estatísticas
Para comparar as médias dos parâmetros
funcionais morfológicos externos (área foliar, área
foliar específica, massa foliar específica) e
características funcionais anatômicas (espessura
dos tecidos anatômicos, densidade e tamanho dos
estômatos (variáveis y) entre as categorias
“espécies mais abundantes” e “espécies menos
abundantes” (variáveis x) foram realizados testes-
T para cada uma das variáveis y, separadamente.
Inicialmente, as suposições do teste –t estatístico
foram checadas através do teste de normalidade
(Shapiro-Wilk) das variáveis y e o teste F para
verificar a igualdade de variâncias entre as
amostras de acordo com Verzani (2014). Quando
as variáveis y não atenderam as suposições de
normalidade foi utilizado o teste não paramétrico
de Wilcoxon (Verzani 2014). Quando as mesmas
variáveis y foram comparadas entre cada uma das
espécies foi utilizado o teste de Kruskal-Wallis
(Sprent e Smeeton 2007), seguido de comparações
múltiplas pelo mínimo teste de diferenças
significativas (MDS), que se ajusta para múltiplos
testes, uma vez que as diferenças potenciais podem
não ser independentes (Siegel e Castellan, 1988).
Para analisar a separação dos agrupamentos de
características funcionais foi utilizado a análise de
componentes principais (PCA) usando a
metodologia de matriz de variância (Gotelli &
Ellison 2004) utilizando o programa R-studio.
Resultados
Atributos funcionais entre espécies mais e menos
abundantes
Quando as espécies foram comparadas entre os
agrupamentos categóricos das espécies mais e
menos abundantes, as mais abundantes
apresentaram menores áreas foliares médias (T=
10,10; P<0,001; fig. 2A). Apesar disso, a área
foliar específica desse grupo foi maior (T= 14,14;
P<0,001; fig 2B) e a massa foliar específica menor
(T=14,51; P<0,001; fig 2C) do que os valores
encontrados em folhas das espécies menos
abundantes.
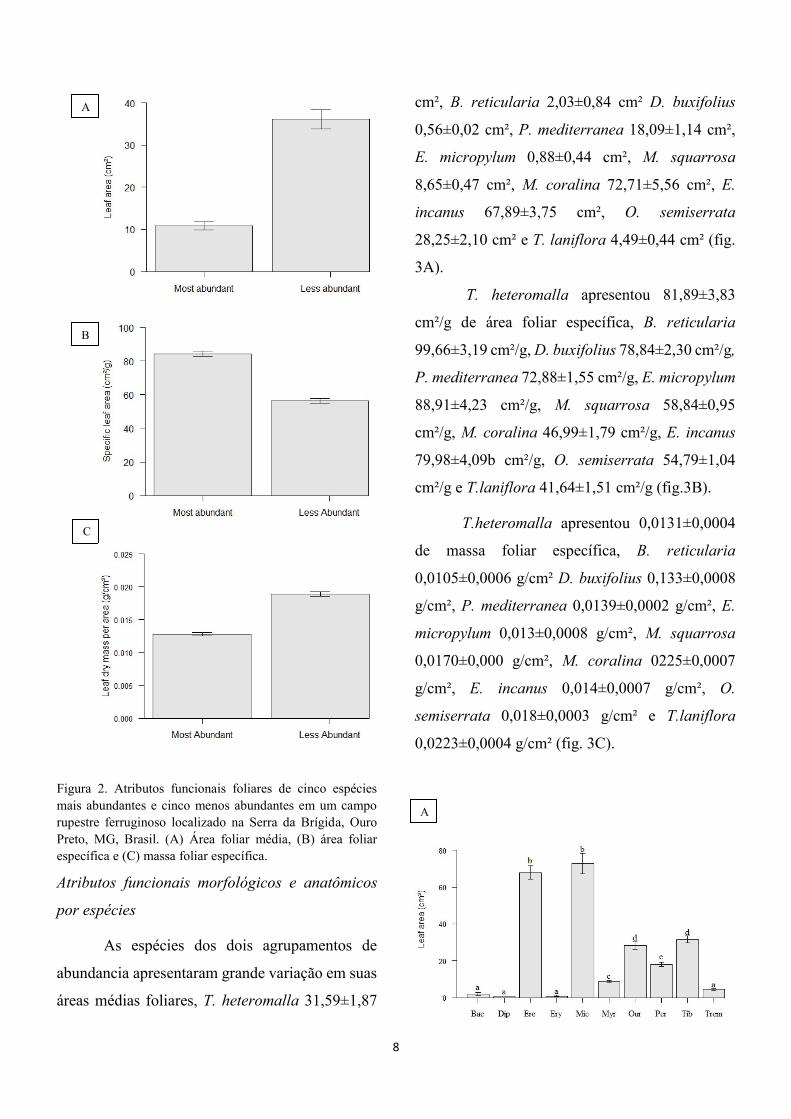
8
Figura 2. Atributos funcionais foliares de cinco espécies
mais abundantes e cinco menos abundantes em um campo
rupestre ferruginoso localizado na Serra da Brígida, Ouro
Preto, MG, Brasil. (A) Área foliar média, (B) área foliar
específica e (C) massa foliar específica.
Atributos funcionais morfológicos e anatômicos
por espécies
As espécies dos dois agrupamentos de
abundancia apresentaram grande variação em suas
áreas médias foliares, T. heteromalla 31,59±1,87
cm², B. reticularia 2,03±0,84 cm² D. buxifolius
0,56±0,02 cm², P. mediterranea 18,09±1,14 cm²,
E. micropylum 0,88±0,44 cm², M. squarrosa
8,65±0,47 cm², M. coralina 72,71±5,56 cm², E.
incanus 67,89±3,75 cm², O. semiserrata
28,25±2,10 cm² e T. laniflora 4,49±0,44 cm² (fig.
3A).
T. heteromalla apresentou 81,89±3,83
cm²/g de área foliar específica, B. reticularia
99,66±3,19 cm²/g, D. buxifolius 78,84±2,30 cm²/g,
P. mediterranea 72,88±1,55 cm²/g, E. micropylum
88,91±4,23 cm²/g, M. squarrosa 58,84±0,95
cm²/g, M. coralina 46,99±1,79 cm²/g, E. incanus
79,98±4,09b cm²/g, O. semiserrata 54,79±1,04
cm²/g e T.laniflora 41,64±1,51 cm²/g (fig.3B).
T.heteromalla apresentou 0,0131±0,0004
de massa foliar específica, B. reticularia
0,0105±0,0006 g/cm² D. buxifolius 0,133±0,0008
g/cm², P. mediterranea 0,0139±0,0002 g/cm², E.
micropylum 0,013±0,0008 g/cm², M. squarrosa
0,0170±0,000 g/cm², M. coralina 0225±0,0007
g/cm², E. incanus 0,014±0,0007 g/cm², O.
semiserrata 0,018±0,0003 g/cm² e T.laniflora
0,0223±0,0004 g/cm² (fig. 3C).
B
A
C
A
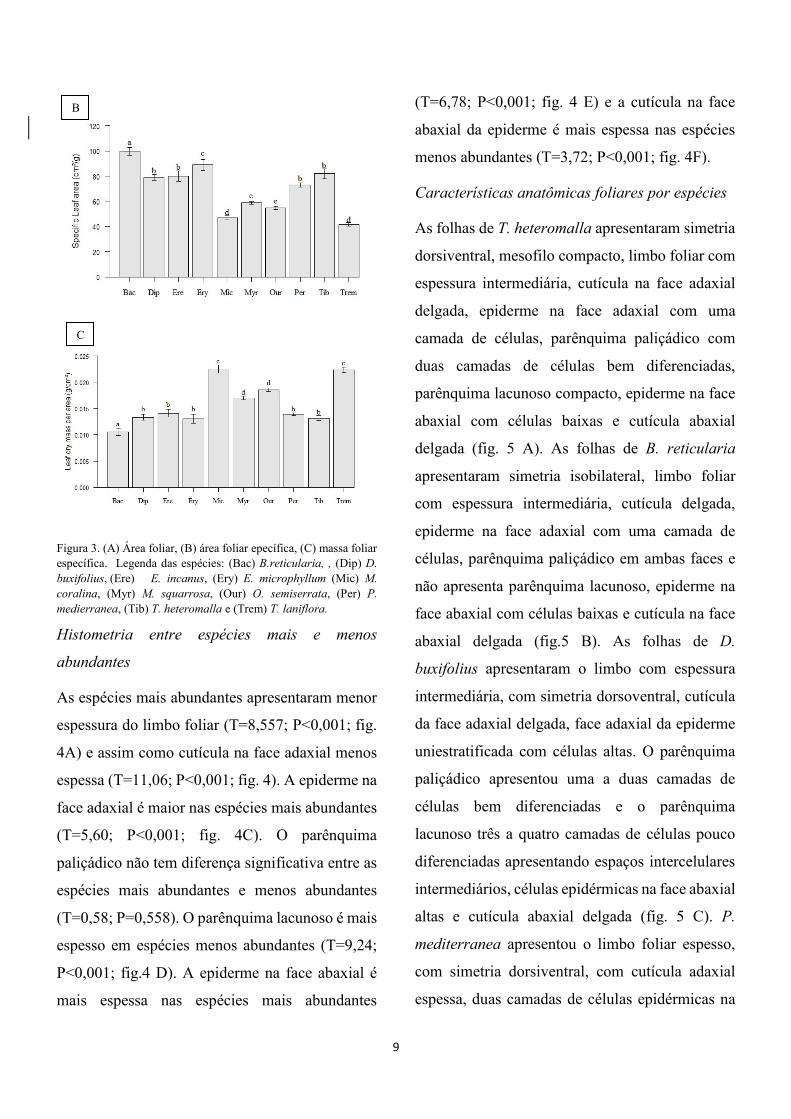
9
Figura 3. (A) Área foliar, (B) área foliar epecífica, (C) massa foliar
específica. Legenda das espécies: (Bac) B.reticularia, , (Dip) D.
buxifolius, (Ere) E. incanus, (Ery) E. microphyllum (Mic) M.
coralina, (Myr) M. squarrosa, (Our) O. semiserrata, (Per) P.
medierranea, (Tib) T. heteromalla e (Trem) T. laniflora.
Histometria entre espécies mais e menos
abundantes
As espécies mais abundantes apresentaram menor
espessura do limbo foliar (T=8,557; P<0,001; fig.
4A) e assim como cutícula na face adaxial menos
espessa (T=11,06; P<0,001; fig. 4). A epiderme na
face adaxial é maior nas espécies mais abundantes
(T=5,60; P<0,001; fig. 4C). O parênquima
paliçádico não tem diferença significativa entre as
espécies mais abundantes e menos abundantes
(T=0,58; P=0,558). O parênquima lacunoso é mais
espesso em espécies menos abundantes (T=9,24;
P<0,001; fig.4 D). A epiderme na face abaxial é
mais espessa nas espécies mais abundantes
(T=6,78; P<0,001; fig. 4 E) e a cutícula na face
abaxial da epiderme é mais espessa nas espécies
menos abundantes (T=3,72; P<0,001; fig. 4F).
Características anatômicas foliares por espécies
As folhas de T. heteromalla apresentaram simetria
dorsiventral, mesofilo compacto, limbo foliar com
espessura intermediária, cutícula na face adaxial
delgada, epiderme na face adaxial com uma
camada de células, parênquima paliçádico com
duas camadas de células bem diferenciadas,
parênquima lacunoso compacto, epiderme na face
abaxial com células baixas e cutícula abaxial
delgada (fig. 5 A). As folhas de B. reticularia
apresentaram simetria isobilateral, limbo foliar
com espessura intermediária, cutícula delgada,
epiderme na face adaxial com uma camada de
células, parênquima paliçádico em ambas faces e
não apresenta parênquima lacunoso, epiderme na
face abaxial com células baixas e cutícula na face
abaxial delgada (fig.5 B). As folhas de D.
buxifolius apresentaram o limbo com espessura
intermediária, com simetria dorsoventral, cutícula
da face adaxial delgada, face adaxial da epiderme
uniestratificada com células altas. O parênquima
paliçádico apresentou uma a duas camadas de
células bem diferenciadas e o parênquima
lacunoso três a quatro camadas de células pouco
diferenciadas apresentando espaços intercelulares
intermediários, células epidérmicas na face abaxial
altas e cutícula abaxial delgada (fig. 5 C). P.
mediterranea apresentou o limbo foliar espesso,
com simetria dorsiventral, com cutícula adaxial
espessa, duas camadas de células epidérmicas na
C
B
B

10
face adaxial, parênquima paliçádico com 3 a 4
camadas de células e o parênquima lacunoso
compacto, epiderme na face abaxial com células
altas, cutícula na face abaxial intermediária (fig.
5D) e fibras esclerênquimáticas. E. microphylum
apresentou o limbo foliar intermediário, com
organização dorsoventral, com cutícula na face
adaxial espessa, a face adaxial da epiderme
apresenta células altas, parênquima paliçádico
com duas camadas de células, parênquima
lacunoso conspícuo, epiderme na face abaxial com
células altas e cutícula na face abaxial delgada (fig.
5 E). O limbo foliar de M. squarrosa é
dorsoventral espesso com cutícula adaxial espessa,
epiderme adaxial uni-estratificada com
intermediária, parênquima paliçádico com duas
camadas de células, parênquima lacunoso com
espaços intercelulares conspícuos, epiderme
abaxial com células baixas e cutícula na face
abaxial espessa (fig. 6 A). O limbo de M. coralina
é espesso com simetria dorsoventral, a cutícula
adaxial espessa, epiderme adaxial com células
baixas, parênquima paliçádico com três a quatro
camadas de células e parênquima lacunoso
compacto, epiderme abaxial com células baixas e
a cutícula abaxial delgada e bainha
esclerenquimática que se estende em ambas as
faces da epiderme (fig. 6 B). O limbo foliar de O.
semiserrata é espesso com simetria dorsoventral,
cutícula adaxial espessa, epiderme adaxial com
células altas, parênquima paliçádico com duas
camadas de células, parênquima lacunoso
conspícuo, epiderme com células baixas, bainha
esclerenquimática que se estende entre a face
abaxial e adaxial da folha (fig. 6 C). O limbo foliar
de E. incanus é intermediário com simetria
dorsiventral, cutícula adaxial delgada, epiderme
adaxial com células baixas, parênquima paliçádico
com duas camadas de células bem diferenciadas,
parênquima lacunoso com três a quatro camadas
de células, epiderme abaxial uni-estratificada com
células epidérmicas baixas, cutícula abaxial
delgada e apresenta bainha parenquimática que se
estende em ambas as faces da epiderme (fig. 6 D).
O limbo foliar de T. laniflora é intermediário, com
cutícula da epiderme adaxial espessa, parênquima
paliçádico com três a quatro camadas, parênquima
lacunoso compacto, epiderme abaxial com células
baixas e cutícula abaxial delgada (fig. 6 E).
Atributos funcionais que podem estar
relacionados com a captação de água pela folha
T. heteromalla apresentou idioblastos com cristais
de oxalato de cálcio na epiderme adaxial (fig. 7.1)
e também na base de emergências, em ambos os
lados das folhas (fig. 7.2), onde também foi
possível encontrar células mucilaginosas (fig. 7.3).
B.reticularia apresenta cavidades secretoras (fig.
7.4) e tricomas (fig. 7.5). Em D. buxifolius pode
ser encontrado tricomas unicelulares tectores com
parede espessa e ápice agudo (fig.8.6), células
mucilaginosas (fig. 7.7) e cristais de oxalato de
cálcio (fig. 7.9). P. mediterranea apresenta bainha
esclerenquimática, células mucilaginosas (fig.
7.10) e tricomas (fig. 7.11), E. microphylum
apresenta células mucilaginosas (fig. 7.12). M.
squarrosa apresenta células mucilaginosas ao

11
longo da epiderme na face adaxial, cavidades
secretoras (fig.8.1) e tricomas glandulares sésseis
(fig. 8.2). M. coralina apresenta tricomas tectores
estrelados com paredes espessadas (fig. 8.3-4) e
idioblastos com drusas de oxalato de cálcio (fig.
8.5). E. incanus apresenta tricomas glandulares na
face adaxial com o pedículo curto (fig. 8.6)
agrupamentos de esclerênquima (8.7) e
emergências na face abaxial (8.8). Em O.
semiserrata podem ser encontrados tricomas
glandulares (fig. 8.9) e drusas de oxalato de cálcio
(fig. 8.10) e células mucilaginosas (fig. 9.11).
T.laniflora apresenta tricomas em ambas faces da
epiderme (fig. 8.12).
Convergência de atributos funcionais que podem
estar relacionados com a captação de água pela
folha
90% apresentaram tricomas, 80% células
mucilaginosas, 60% cristais de oxalato de cálcio e
20% apresentaram cavidades secretoras.
Disposição dos estômatos
As folhas de T. heteromalla são
anfiestomáticas apresentando estômatos no
mesmo alinhamento das demais células da
epiderme (fig.5A). B. reticularia apresenta folhas
anfiestomáticas no mesmo alinhamento das
demais células da epiderme (fig. 5B). As folhas de
D. buxifolius são anfiestomáticas e são nivelados
com as demais células epidérmicas (fig. 5C). As
folhas de P. mediterrânea são hipoestomáticas
apresentando estômatos em criptas (fig. 5D). As
folhas de E. microphylum são hipoestomáticas e os
estômatos são alinhados às demais células da
epiderme apresentando câmaras subestomáticas
conspícuas (fig. 5E). As folhas de M. squarrosa
são hipoestomáticas e os estômatos são dispostos
na mesma altura das demais células (fig. 6A). As
folhas de M. coralina são hipoestomáticas e os
estômatos são dispostos levemente nivelados em
relação com as demais células epidérmicas,
apresentando câmaras inconspícuas (fig. 6B). As
folhas de E. incanus são hipoestomáticas e os
estômatos são dispostos no mesmo nível das
demais células epidérmicas e apresentam câmaras
sub-estomáticas conspícuas (fig. 6C). As folhas de
O. semiserrata são hipoestomáticas e os estômatos
são alinhados ás demais células da epiderme (fig.
6D). As folhas de T. laniflora são hipoestomáticas
e os estômatos são encontrados em criptas (fig.
6E).
Densidade e tamanho dos estômatos
As espécies mais abundantes apresentaram
maiores densidades estomáticas (T= -3.287;
P=0.00106; fig. 9A), valores muito influenciados
(fig. 9C e 10) e apresentaram menores áreas
estomáticas (T= -3,258; P=0.00122; fig. 9B).
Analise dos componentes principais
A análise multivariada mostrou que o eixo PC1
explica 27% do dos dados de espessura foliar,
cutícula adaxial, parênquima lacunoso e massa
foliar específica e o eixo PC2 explica 24% dos
dados de área foliar, área estomática e cutícula
adaxial. As espécies mais abundantes
apresentaram os agrupamentos de atributos
funcionais como, área foliar específica, área
estomática, parênquima paliçádico, epiderme

12
adaxial, epiderme abaxial. Enquanto as espécies
menos abundantes apresentaram agrupamentos de
cutícula abaxial, espessura foliar, parênquima
lacunoso, cutícula adaxial, massa foliar específica,
densidade estomática e área foliar. Apesar das
plantas mais e menos abundantes apresentarem
dois agrupamentos de atributos funcionais as
espécies T. heteromalla, E, microphyllum, E .
incanus, P. mediterranea e M. squarrosa, foram
plotadas em uma área de intersecção entre os
agrupamentos (figura 11).
Discussão
Atributos funcionais morfológicos
As folhas são muito variáveis entre
espécies diferentes, podendo variar mesmo na
mesma espécie e até no mesmo indivíduo, como
resposta a diferentes intensidades luminosas
incidentes sobre a superfície foliar (Rossato 2013).
Quando as plantas são submetidas à alta
intensidade luminosa, uma estratégia adaptativa
morfológica para a diminuição da perda de água
por convecção entre a superfície foliar e o meio,
por transpiração, é a presença de folhas com áreas
reduzidas (Salisboury 2013, De Micco e Aronne
2012), o que é considerado um atributo funcional
xeromórfico. Como os campos rupestres
ferruginosos impõem uma alta demanda
evaporativa nas folhas pela exposição a ventos
fortes e alta radiação solar (Giulietti 1987, Rapini
et al, 2008), esperava-se que as plantas mais
abundantes, no presente estudo, apresentassem
áreas foliares menores quando comparadas com
espécies menos abundantes. Apesar das plantas
mais abundantes apresentaram folhas com áreas
significativamente menores, não necessariamente
todas apresentaram folhas reduzidas, T.
heteromalla (31,59±1,8cm²), e P. mediterranea
(18,09±1,14cm²), indicando que outros atributos
adaptativos para esse tipo de ambiente são
importantes para estas espécies.
A área foliar específica em geral apresenta
relação positiva com a taxa fotossintética,
produtividade primária e crescimento das plantas
(Reich 1997, Cornelissen et. al 2003) e portanto,
altos valores são em geral encontrados em plantas
pioneiras, com altas capacidades iniciais de
ocupação de espaço. Por exemplo, no estudo de
Boeger e Wisniewski (2003), Brachiaria
decumbens apresentou valores de AFE entre 241,3
cm²/g e 294,7 cm²/g e Arachis pintoi entre 218,9
cm²/g e 319,9 cm²/g. Os valores de AFE também
podem mudar na mesma espécie quando as folhas
são submetidas a diferentes intensidades solares,
por exemplo, em locais úmidos com moderado
défice hídrico Tabebuia heptaphyilla apresentou
234,51 cm²/g em locais sombreados e quando
submetidas a alta radiação solar 80,55 cm²/g
(Siebeneichler et. al 2008). Em florestas montanas
foi registrado 109,10 cm²/g de AFE (Boeger
2009). Já em ambientes esclerofilos, podem ser
encontrados valores médios próximos de 80 cm²/g
(Tuner e Tan 1995). Em florestas decíduas e
brevidecíduas, e em sempre verdes podem ser
encontrados valores próximos a 60,3 cm²/g,
indicando que nestes ambientes as espécies
apresentam crescimento lento (Araújo 2007).
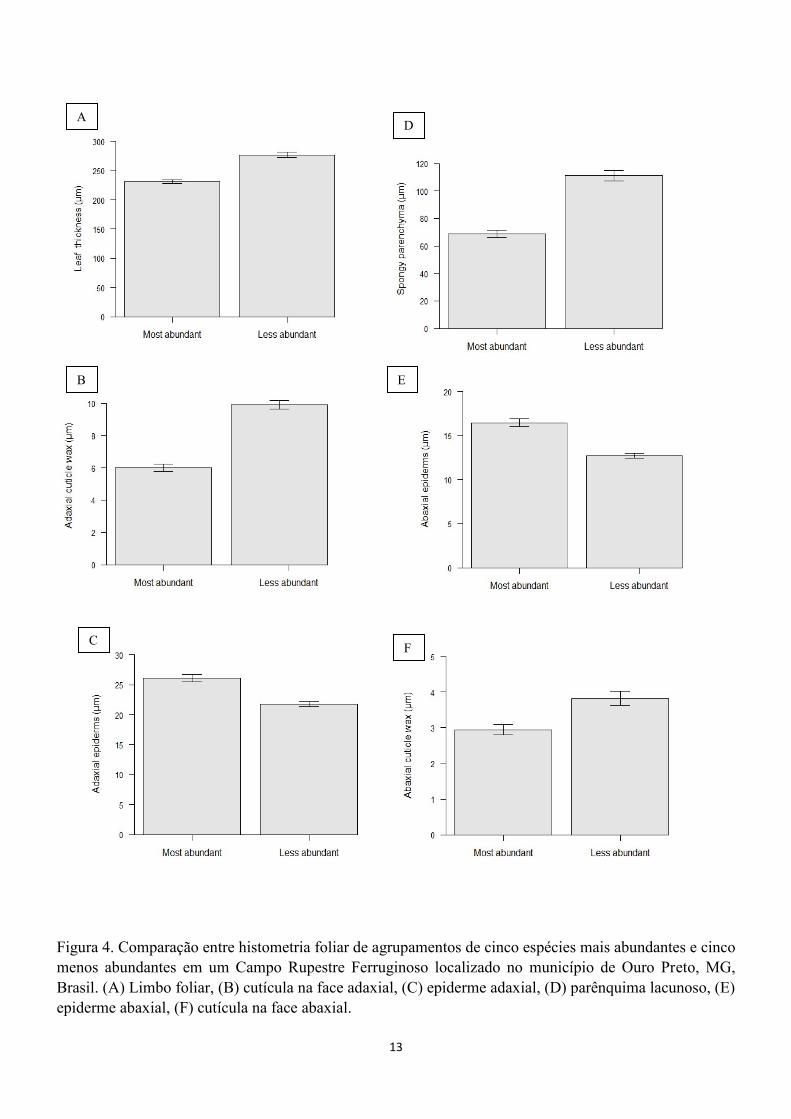
13
Figura 4. Comparação entre histometria foliar de agrupamentos de cinco espécies mais abundantes e cinco
menos abundantes em um Campo Rupestre Ferruginoso localizado no município de Ouro Preto, MG,
Brasil. (A) Limbo foliar, (B) cutícula na face adaxial, (C) epiderme adaxial, (D) parênquima lacunoso, (E)
epiderme abaxial, (F) cutícula na face abaxial.
A
S
h
a
p
i
r
o
-
W
i
l
k
D
S
h
a
p
i
r
o
-
W
i
l
k
B
S
h
a
p
i
r
o
-
W
i
l
k
C
S
h
a
p
i
r
o
-
W
i
l
k
E
S
h
a
p
i
r
o
-
W
i
l
k
F
S
h
a
p
i
r
o
-
W
i
l
k
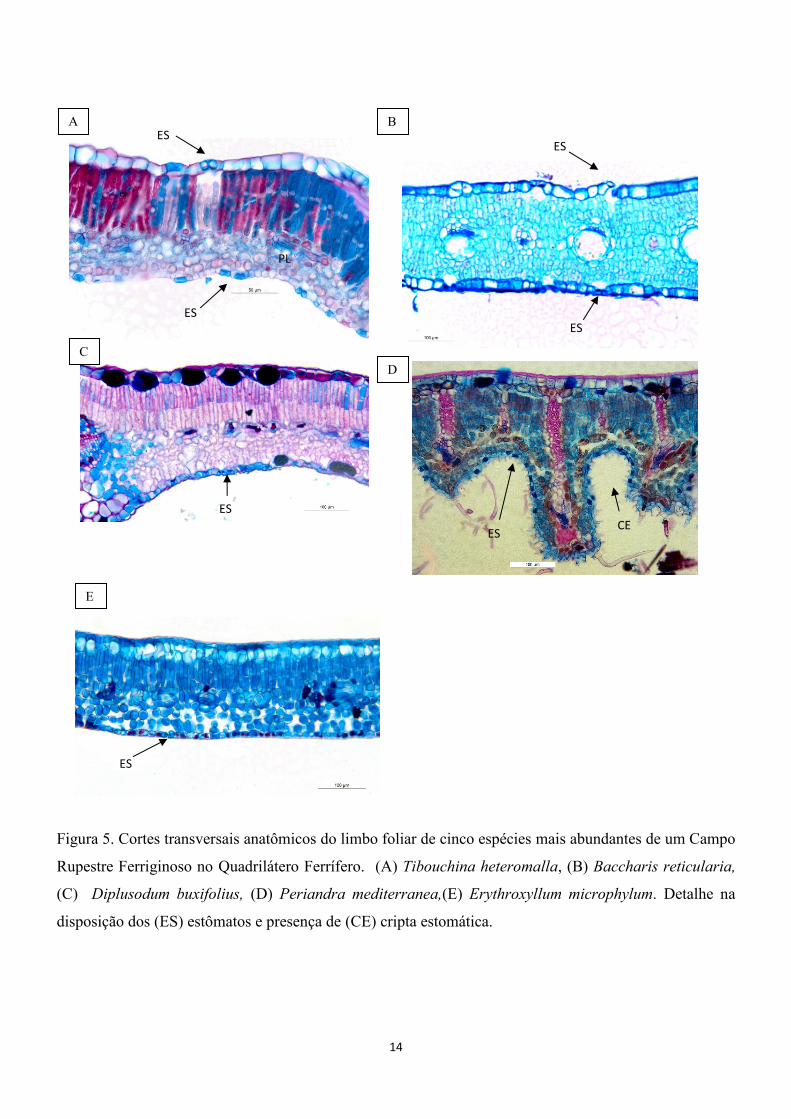
14
Figura 5. Cortes transversais anatômicos do limbo foliar de cinco espécies mais abundantes de um Campo
Rupestre Ferriginoso no Quadrilátero Ferrífero. (A) Tibouchina heteromalla, (B) Baccharis reticularia,
(C) Diplusodum buxifolius, (D) Periandra mediterranea,(E) Erythroxyllum microphylum. Detalhe na
disposição dos (ES) estômatos e presença de (CE) cripta estomática.
A
S
h
a
p
i
r
o
-
W
i
l
k
E
S
h
a
p
i
r
o
-
W
i
l
k
C
S
h
a
p
i
r
o
-
W
i
l
k
ES
D
S
h
a
p
i
r
o
-
W
i
l
k
B
S
h
a
p
i
r
o
-
W
i
l
k
PL
ES
ES
CE
ES
ES
ES
ES
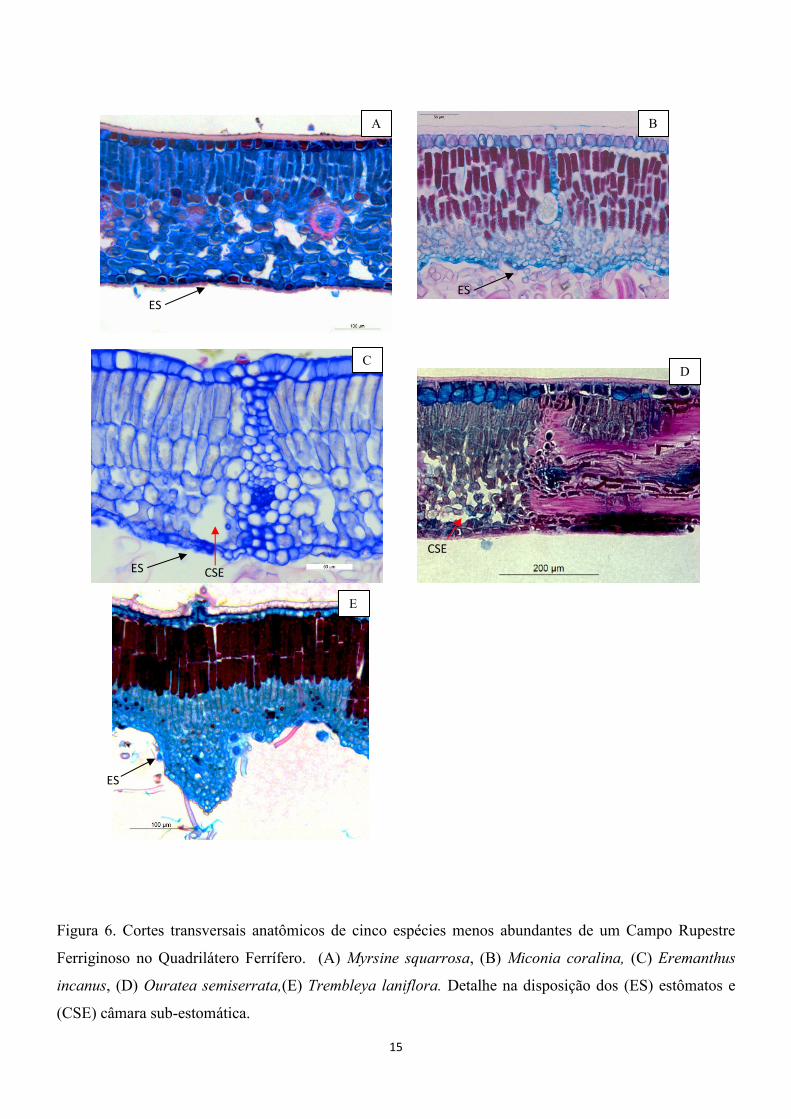
15
Figura 6. Cortes transversais anatômicos de cinco espécies menos abundantes de um Campo Rupestre
Ferriginoso no Quadrilátero Ferrífero. (A) Myrsine squarrosa, (B) Miconia coralina, (C) Eremanthus
incanus, (D) Ouratea semiserrata,(E) Trembleya laniflora. Detalhe na disposição dos (ES) estômatos e
(CSE) câmara sub-estomática.
A
S
h
a
p
i
r
o
-
W
i
l
k
B
S
h
a
p
i
r
o
-
W
i
l
k
C
S
h
a
p
i
r
o
-
W
i
l
k
D
S
h
a
p
i
r
o
-
W
i
l
k
E
S
h
a
p
i
r
o
-
W
i
l
k
ES
ES
ES ES
CSE
CSE
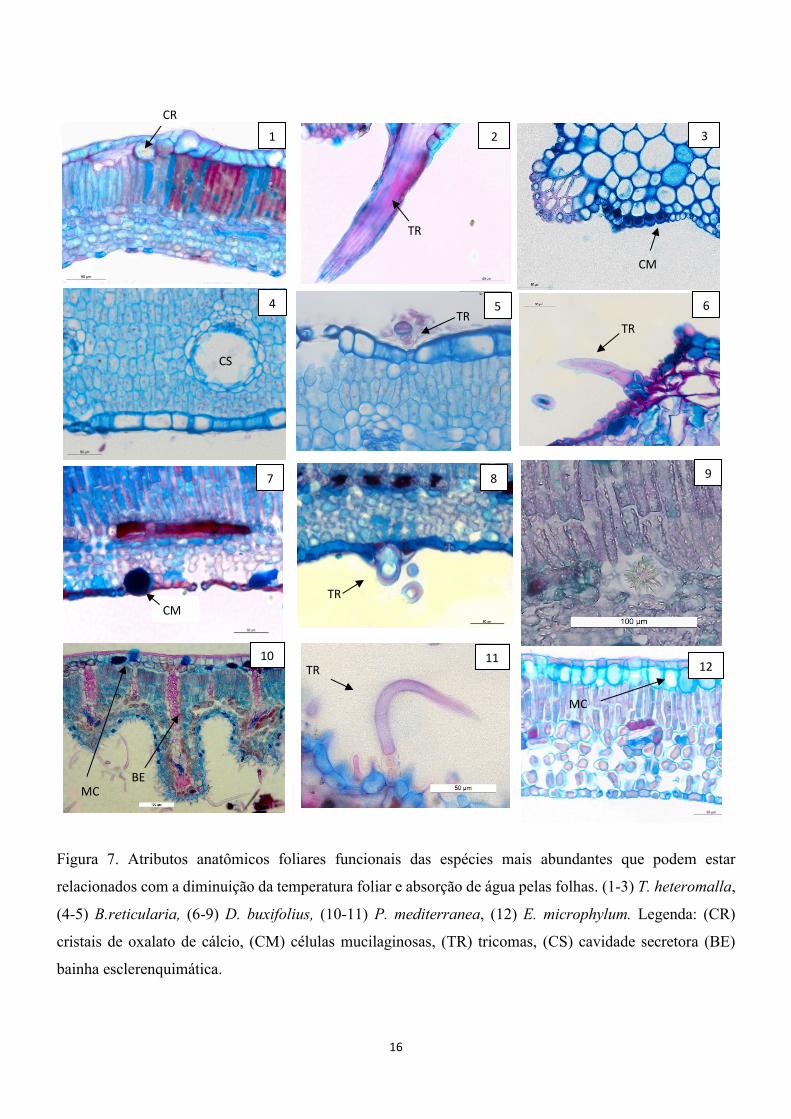
16
Figura 7. Atributos anatômicos foliares funcionais das espécies mais abundantes que podem estar
relacionados com a diminuição da temperatura foliar e absorção de água pelas folhas. (1-3) T. heteromalla,
(4-5) B.reticularia, (6-9) D. buxifolius, (10-11) P. mediterranea, (12) E. microphylum. Legenda: (CR)
cristais de oxalato de cálcio, (CM) células mucilaginosas, (TR) tricomas, (CS) cavidade secretora (BE)
bainha esclerenquimática.
CR
E
CR
1
c
2
CR
TR
CS
TR TR
CM TR
CM
MC
MC
TR
3
4
c
7 8
10 11 12
9
6
BE
5
5
c
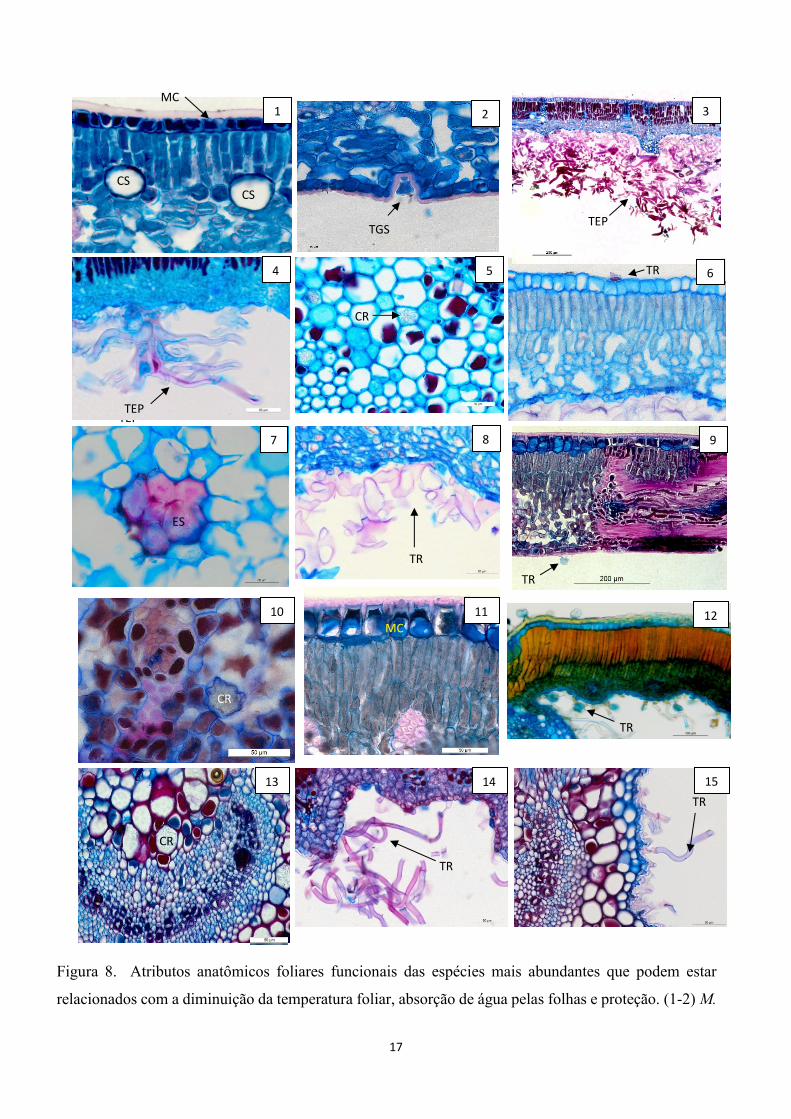
17
Figura 8. Atributos anatômicos foliares funcionais das espécies mais abundantes que podem estar
relacionados com a diminuição da temperatura foliar, absorção de água pelas folhas e proteção. (1-2) M.
ESC
TEP
TR
TR
CS CS
TGS
G
CR
TEP
TEP
ES
TR
TR
CR
MC
TR
CR
TR
1 2 3
4 5 6 TR
7 8 9
10 11 12
13 14 15
MC

18
squarrosa, (3-5) M. coralina, (6-8) E. incanus,(9-11) O. semiserrata, (12-15) T. laniflora. Legendas:
(CS) cavidade secretora, (TEP) tricoma estrelado pluricelular, (TGS) tricoma glândular séssil (ES)
esclerênquima (CR) cristais de oxalato de cálcio, (TR) tricomas.
Tabela 1: Atributos funcionais anatômicos que podem estar relacionados com a absorção de água e
diminuição da temperatura foliar de 10 espécies em um Campo Rupestre Ferruginoso na Serra da Brígida,
Ouro Preto, Brasil. (+) Presença (-) ausência de atributo funcional.
Figura 9. (A) Densidade estomática e (B) área estomática entre espécies mais e menos abundantes. Em um
Campo Rupestre Ferruginoso na Serra da Brígida, Ouro Preto, Brasil.
Espécies Drusas de oxalato de cálcio Cavidade secretora
Células
mucilaginosas Tricomas Emergências
T.heteromalla + - + + +
B. reticularia - + + + -
D. buxifolius + - + + -
P. medietarranea - - + + +
E. microphyllum - - + - -
M. squarrosa - + + + -
M. coralina + - + + +
E. incanus + - - + +
O. semiserrata + - + + -
T. laniflora + - - + +
A
B
c
B
19
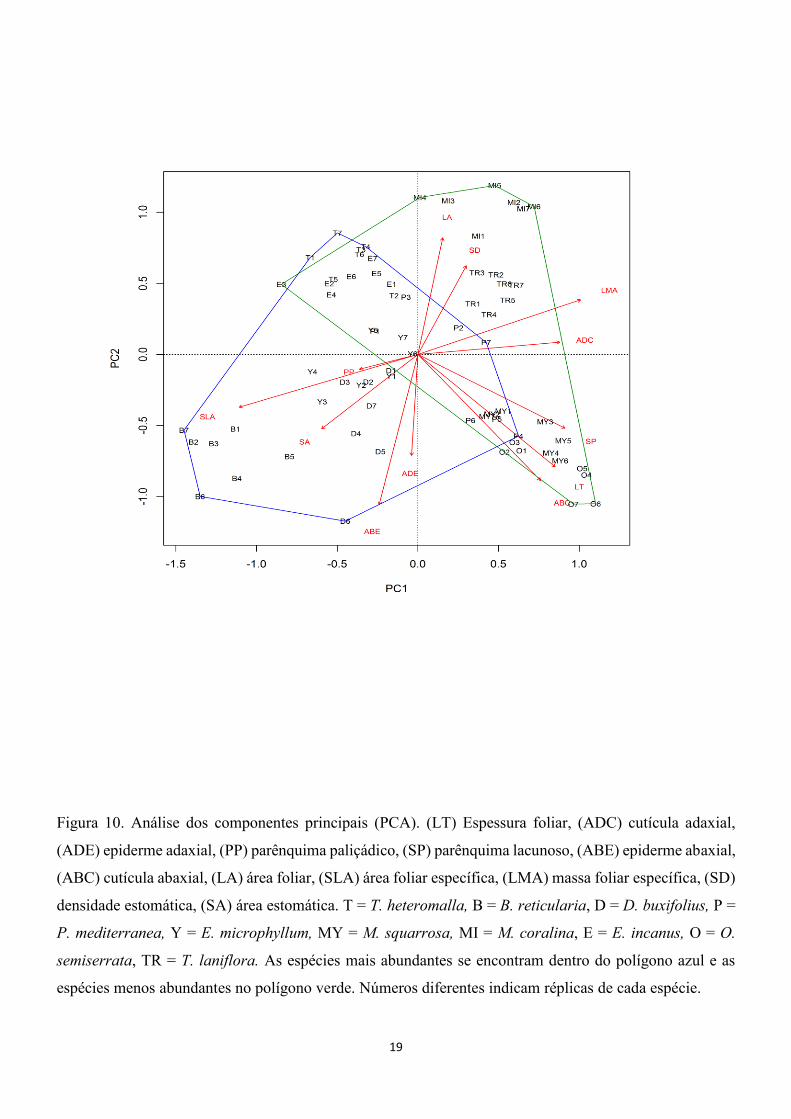
Figura 10. Análise dos componentes principais (PCA). (LT) Espessura foliar, (ADC) cutícula adaxial,
(ADE) epiderme adaxial, (PP) parênquima paliçádico, (SP) parênquima lacunoso, (ABE) epiderme abaxial,
(ABC) cutícula abaxial, (LA) área foliar, (SLA) área foliar específica, (LMA) massa foliar específica, (SD)
densidade estomática, (SA) área estomática. T = T. heteromalla, B = B. reticularia, D = D. buxifolius, P =
P. mediterranea, Y = E. microphyllum, MY = M. squarrosa, MI = M. coralina, E = E. incanus, O = O.
semiserrata, TR = T. laniflora. As espécies mais abundantes se encontram dentro do polígono azul e as
espécies menos abundantes no polígono verde. Números diferentes indicam réplicas de cada espécie.

20
Das dez espécies estudadas, as cinco mais
abundantes apresentaram área foliar específica
entre 72,88 cm²/g e 99,66 cm²/g, enquanto entre as
espécies menos abundantes o maior valor
encontrado foi em E. incanus (79,98 cm²/g) e as
outras entre 41,99 cm²/g e 58,84 cm²/g. Quando
foram comparadas entre os grupos mais e menos
abundantes, as espécies mais abundantes
apresentaram maiores valores de AFE
significativamente, o que deve estar relacionado
com uma maior produtividade, crescimento e
abundância dessas plantas (Reich 1997,
Cornelissen et. al 2003).
A massa foliar específica (MFE) é um
atributo que indica a dureza e níveis de esclerofilia
das folhas, apresentando altos valores em espécies
submetidas a stress hídrico consideradas como
economizadoras de água e nutrientes (Cornelissen
2003). Tem uma relação positiva com a radiação
solar (Ogaia e Peñuelas 2007) e inversamente
proporcional com a disponibilidade de água no
solo (De La Riva et. al 2016), precipitação (Ogaia
e Peñuelas 2007) e taxa fotossintética (Reich
1997), repercutindo na baixa produtividade e taxa
de crescimento lento (Rossatto 2013; Reich 2014).
Assim, seria esperado que nos campos rupestres
ferruginosos as plantas de uma forma geral
apresentassem altos níveis de MFE (Silveira et al.
2015), devido à baixa disponibilidade de água no
solo. De um modo geral as espécies arbóreas
apresentam amplitudes de MFE entre 30 e 330 g/
m−2 (Poorter et. al 2009, De La Riva et. al 2016).
Valores mais altos são encontrados em plantas
mais tolerantes a baixa disponibilidade de água no
ambiente. Por exemplo, plantas sempre verdes,
que possuem crescimento lento, apresentem
maiores valores de MFE do que espécies decíduas
(De La Riva et. al 2016).
Neste estudo, as cinco espécies mais
abundantes apresentaram valores de MFE entre
(105 e 139 g/ m−2), assim é possível indicar que as
espécies mais abundantes investem menos em
características funcionais relacionadas com
proteção, economia de água e nutrientes, quando
comparadas com as espécies menos abundantes
que apresentaram MFE entre (141 e 185 g/ m−2).
Portanto as espécies menos abundantes foram
consideradas como “economizadoras”, e as
espécies mais abundantes investem mais em
atributos funcionais relacionados com
características hidráulicas, como por exemplo, alta
densidade estomática (Salisboury 2013; Li et al,
2015).
Atributos anatômicos foliares funcionais
A presença de folhas mais espessas
também é considerada uma característica
adaptativa para ambientes sujeitos a períodos com
pouca água disponível, pela possibilidade de
aumentar a assimilação de CO² por unidade de área
e proteção contra a falta de água excessiva
(Rossato 2013). O aumento da espessura foliar
pode ser resultado do aumento de camadas de
parênquima paliçádico acarretando em uma maior
quantidade de cloroplastos por unidade de área,
repercutindo em uma maior taxa fotossintética
(Oguchi et al. 2005), o que poderia também

21
acarretar em vantagens competitivas nos campos
ferruginosos. No entanto, as variáveis categóricas
entre as espécies menos e mais abundantes não
apresentaram diferenças significativas na
espessura do parênquima paliçádico. Esse
resultado da maior espessura foliar em espécies
menos abundantes foi influenciado pela maior
espessura da cutícula e das células epidérmicas, na
face adaxial, além do parênquima lacunoso.
A maior espessura da cutícula, juntamente
com a camada de cera epicuticular, é descrita na
literatura como uma característica para evitar a
perda de água excessiva (Pyykko 1966, Fahn &
Cutler 1992, Larcher 2004), além de prevenir
contra a exposição excessiva a raios UV, quando
as folhas estão submetidas a altas intensidades
solares (Pyykko 1966). Também, a presença de
células epidérmicas espessas ou epiderme
pluriestratificada pode funcionar como
armazenadoras de água (Fahn & Cutler 1992). No
presente estudo a maioria das espécies apresentou
epiderme uni-estratificada, podendo ser
encontrado epiderme pluri-estratficada somente
em P. mediterranea e E. microphyllum, que fazem
parte das plantas mais abundantes. Além disso as
espécies mais abundantes também apresentaram
folhas com maiores espessuras das células
epidérmicas.
O parênquima lacunoso é menos espesso
nas espécies mais abundantes, característica que
pode estar relacionada com a regulação osmótica
da folha. Por outro lado, diminui a distância entre
os estômatos e o parênquima paliçádico em
espécies hipoestomáticas e com simetria
dorsiventral, o que também pode influenciar no
aumento da taxa fotossintética e da difusão de
gases (Lambers 1998), repercutindo na maior
produtividade dessas espécies.
Atributos funcionais que podem estar
relacionados com a absorção e manutenção de
água e diminuição da temperatura foliar
Tricomas e emergências influenciam na
difusão do vapor de água na superfície das folhas,
evitando a transpiração excessiva quando estas
estão submetidas a ventos e radiação UV (Woolley
1964), e também podem facilitar a absorção de
água pelas folhas, podendo aprisionar pequenas
gotículas de água da atmosfera, quando ocorrem
eventos de neblina. E consequentemente, por
diferença de potencial hídrico entre a atmosfera e
o interior das folhas, a água é absorvida (Johnson
1975).
Outros estudos mostraram que os Campos
Rupestres Ferruginosos apresentam solos
distróficos e elevadas concentrações de Fe e Al
(Machado 2011) o que pode ser prejudicial para
plantas que não possuem estratégias para evitar a
absorção desses elementos pelas raízes ou
acumulá-los em células especializadas nas folhas
(Hartwig et. al 2007, Duc et. al 2008). Algumas
plantas são acumuladoras de Al mas, não
necessariamente, a capacidade de acumulação de
Al está relacionada com a abundancia das plantas
do presente estudo (Arias 2016). Um dos atributos
funcionais que podem servir para destoxicação
destes metais é a presença de drusas de oxalato de
cálcio (Franceschi & Nakata 2005). Além disso,

22
esses cristais podem refratar a luz incidente sobre
a superfície foliar como um prisma, diminuindo a
temperatura da folha e, consequentemente, a taxa
de transpiração (Franceschi & Nakata 2005),
podendo trazer vantagens para espécies como
Tibouchina heteromalla e Diplusodon buxifolius,
que fazem parte do grupo de plantas mais
abundantes. Contudo, Miconia coralina, também
apresenta grande quantidade desses cristais e faz
parte das plantas menos abundantes.
Segundo Hafez (1952) ductos secretores,
também chamados de canais secretores, também
podem reduzir a difusão de vapor de água na
transpiração, mas esta função não é muito bem
descrita na literatura. Com outro viés, a alta
radiação solar pode influenciar na produção de
óleos essências secretados por cavidades e ductos
secretores (Budel 2012), o que pode resultar em
menores taxas de herbivoria (Rodriguez 2011). Os
indivíduos de Baccharis reticularia (Asteraceae)
possuem ductos secretores e Myrsine squarrosa
(Primulaceae) possui cavidades secretoras em suas
folhas. E a ocorrência dessas estruturas parecem
estar bem fixadas filogeneticamente tanto nas
Asteraceae (Pagni & Masini 1999, Trombin 2017)
quanto nas Primulaceae (Luna et al. 2014). Sendo
uma estratégia muito importante para estas plantas
no campo rupestre ferruginoso.
Células mucilaginosas são muito
importantes na economia de água nas folhas, pois
as mucilagens são polissacarídeos hidrofílicos
com capacidade de retenção de a água (Fahn
1992), podendo também favorecer a absorção e
manutenção da água na planta quando acontecem
eventos de neblina (Westyhoff 2009, Souza 2014).
Esse tipo de célula foi observado, em ambas as
faces da epiderme de todas as cinco espécies mais
abundantes. Como elas também foram encontradas
em três espécies do grupo das menos abundantes,
e em outras estudadas por Souza (2014) na mesma
área de estudo. E acredita-se que esse seja também
um caractere adaptativo importante em plantas de
campos ferruginosos.
Convergência evolutiva de atributos funcionais
que podem estar relacionados com a absorção e
manutenção de água e diminuição da temperatura
foliar
No campo rupestre ferruginoso onde foi
realizado este estudo, é comum a ocorrência de
eventos de neblina inclusive na estação seca
(Valim 2012, Souza 2014), funcionando como
uma segunda via de captação de água,
influenciando na seleção de plantas que possuem
características funcionais anatômicas que podem
facilitar a captação e manutenção da água nas
folhas. Assim, das 10 espécies estudadas, 90%
apresentaram tricomas, 80% células
mucilaginosas, 60% cristais de oxalato de cálcio,
50% emergências e 20% cavidades secretoras.
Densidade e tamanho dos estômatos
Alta densidade estomática por unidade de
área e com menores áreas estomáticas também são
consideradas como um atributo funcional
anatômico hidráulico xeromórfico (Beaulieu 2008,
Franks et al. 2009, Drake 2013), potencializar as
trocas gasosas e aumentar a velocidade da abertura

23
dos estômatos quando há disponibilidade de água
e velocidade de fechamento em situações de déficit
hídrico (Raven 2014). As espécies mais
abundantes deste estudo apresentaram maior
densidade estomática e menores áreas estomáticas
(fig. 9, A-B), indicando ser um dos atributos que
podem estar relacionado com a maior abundancia
deste grupo de plantas.
Outra caraterística que aumenta a taxa de
assimilação de CO² quando há disponibilidade de
água no ambiente é a presença de estômatos em
ambas as faces da epiderme (folhas
anfiestomáticas) (Parkhurst 1978, Moot 1982), e
também é considerada como xeromórfica (Bertsch
1969). As três espécies mais abundantes T.
heteromalla, B. reticularia e D. buxifolius
apresentaram folhas anfiestomáticas o que pode
ser relacionado com a maior produtividade destas
plantas. B. reticularia além de apresentar
anfiestomia suas folhas são isobilaterais o que
também aumenta o desempenho fotossintético
(Pykko 1966), também podendo repercutir na
maior abundancia dessa espécie.
Agrupamentos de características funcionais
anatômicas e morfológicas das espécies mais
abundantes (PCA)
As espécies mais abundantes apresentaram
maior espessura das células da epiderme tanto na
face adaxial quanto abaxial, maior área foliar
específica e maior densidade estomática e menores
áreas estomáticas D. buxifolius e E. microphylum
possuem cutícula delgada, o que parece ser
compensado pela espessura da epiderme e pela
redução da área foliar.
T. heteromalla, que é a espécie mais
abundante, possui folhas grandes e cutícula
delgada, o que pode ser compensado pela maior
espessura das células epidérmicas e densa
pubescência principalmente na face adaxial. Essas
características refratam e refletem o excesso de
radiação, evitam o superaquecimento da folha,
reduzem a taxa de transpiração (Pikko 1966,
Rozendaal 2006, Rossato 2013) e também podem
favorecer a absorção de água pelas folhas (Souza
2014). Outros atributos que podem estar
relacionados com a maior abundância desta
espécie é a alta densidade estomática (Beaulieu
2008, Franks et al. 2009, Drake 2013), o que
aumenta a assimilação de CO² e o reduzido
tamanho dos estômatos, o que permite rápida
abertura e fechamento dessas estruturas (Raven
2014). Estas características, juntamente com altos
valores de área foliar específica, possibilitam alta
taxa fotossintética e produtividade primária destas
plantas refletindo na taxa de crescimento rápido e
abundância destas plantas (Reich et. al 2014). Por
outro lado, B. reticularia, que é a segunda espécie
mais abundante, possui folhas pequenas
anfistomáticas com simetria dorsiventral
apresentando parênquima paliçádico espesso e
altos valores de área foliar específica, (Parkhust,
1978, Mott 1982, Reich et. al 1999, Reich et. al
2014) o que pode, também repercutir na
produtividade destas plantas, podendo compensar
a baixa densidade estomática e a presença de
estômatos grandes (1903±179,31µm²) (Raven
2014).

24
Agrupamentos de características funcionais
anatômicas e morfológicas das espécies menos
abundantes (PCA)
Espécies menos abundantes apresentaram
agrupamentos de atributos funcionais com maiores
valores de espessura foliar, cutícula adaxial,
cutícula abaxial, parênquima lacunoso, massa
foliar específica e área foliar (figura 10). T.
laniflora e M. coralina apresentaram maiores
valores de massa foliar específica indicando que
estas plantas investem em proteção e conservação
de nutrientes, assim este agrupamento de espécies
foi considerado como economizadoras de água (Li
et al. 2015).
Estrutura da comunidade de plantas
Características funcionais anatômicas e
fisiológicas adaptativas das plantas são
determinantes da abundância de cada espécie
(Cornwell 2010; Rossatto 2013). Apesar de ter a
possibilidade de separar alguns agrupamentos de
características funcionais entre espécies mais e
menos abundantes, cada espécie possuem
estratégias diferentes para tolerar ou evitar as
adversidades impostas pelo Campo Rupestre
Ferruginoso do presente estudo, podendo ser
similares ou divergentes entre as espécies. As
diferentes estratégias das plantas entre maior
produtividade e investir menos na proteção e
crescer rapidamente ou investir mais em proteção
e apresentar desenvolvimento lento, está
fortemente ligado às histórias de vida particulares
de cada espécie.
Espécies mais abundantes no presente
estudo estão mais ligadas a características
funcionais que favorecem maiores taxas
fotossintéticas (características hidráulicas) (Li et
al. 2015). Aumentando a produtividade, as plantas
aumentam a possibilidade de crescerem mais
rápido e propagarem mais do que outras, as
tornando mais abundantes e assim, mesmo que
sejam poucos indivíduos que consigam suportar a
pressão ambiental imposta pelo campo rupestre
ferruginoso, estes poucos conseguem crescer e
ocupar novamente estes locais rapidamente.
As plantas menos abundantes são
economizadoras de nutrientes e principalmente
água (Li et al. 2015). Podendo tolerar as pressões
ambientais nestes locais mesmo depois de eventos
estocásticos como fogo e chuvas de granizo.
Também é importante ressaltar que indivíduos de
uma mesma espécie podem apresentar atributos
funcionais diferentes em situações ambientais
diferentes, podendo se aclimatar e mudar sua taxa
fotossintética (Zhou 2016). Assim, as
características funcionais descritas no presente
trabalho podem mudar no futuro evolutivo devido
a outras pressões ambientais futuras como as
mudanças climáticas (Tian 2016).
Conclusão
Cada espécie possuí estratégias diferentes para a
mesma pressão imposta pelo campo rupestre
ferruginoso. As plantas mais e menos abundantes
no presente estudo apresentaram 51 % (resultados
da PCA) de variações em atributos relacionados à
tolerância para ambientes xeromórficos e taxa
fotossintética. Por outro lado, todas as plantas

25
apresentaram uma convergência evolutiva entre
atributos relacionados à absorção de água pelas
folhas. Assim, a neblina foi considerada muito
importante nos graus de xeromorfísmo das plantas.
A abundância das espécies no atual momento
evolutivo está relacionada com características
funcionais que proporcionam maior produtividade
das plantas mais abundantes, o que pode ser
indicado pelos valores de área foliar específica,
anfiestomia, alta densidade estomática. E as
plantas menos abundantes foram consideradas
com tolerantes e economizadoras, repercutindo na
menor taxa de crescimento e abundancia destas
plantas.
Referências bibliográficas
Ackerly, D. D., & Cornwell, W. K. (2007). A trait-based approach to
community assembly: Partitioning of species trait values into
within- and among-community components. Ecology Letters,
10(2), 135–145. http://doi.org/10.1111/j.1461-0248.2006.01006.x
Alvares, C. A., Stape, J. L., Sentelhas, P. C., de Moraes Gonçalves, J. L., &
Sparovek, G. (2013). Köppen’s climate classification map for
Brazil. Meteorologische Zeitschrift, 22(6), 711–728.
http://doi.org/10.1127/0941-2948/2013/0507
Araújo, J. F. De, & Haridasan, M. (2007). Relação entre deciduidade e
concentrações foliares de nutrientes em espécies lenhosas do
cerrado. Revista Brasileira de Botânica, 30(3), 533–542.
https://doi.org/10.1590/S0100-84042007000300017
Arias J,C,J. (2016). Do metal tolerance traits explain spatial distribution
patterns in metallicolous vascular plants species? Programa de Pós-
Graduação em Ecologia de Biomas Tropicais do Instituto de
Ciências Exatas e Biológicas da Universidade Federal de Ouro
Preto. Dissertação de Mestrado, 4-7
.http//www.repositorio.ufop.br/handle/123456789/6397
Ataíde, E. S., Castro, P. de T. A., & Fernandes, G. W. (2011). Florística e
caracterização de uma área de campo ferruginoso no Complexo
Minerário Alegria, Serra de Antônio Pereira, Ouro Preto, Minas
Gerais, Brasil. Revista Árvore, 1265–1275.
http://doi.org/10.1590/S0100-67622011000700013
Beaulieu, J. M., Leitch, I. J., Patel, S., Pendharkar, A., & Knight, C. A.
(2008). Genome size is a strong predictor of cell size and stomatal
density in angiosperms. New Phytologist, 179(4), 975–986.
http://doi.org/10.1111/j.1469-8137.2008.02528.x
Baêta, H, E 2012. Contribuição da deposição úmida (chuva e neblina) nas
relações hídricas e nutricionais de fisionomias de campos
ferruginosos na Serra da Brígida, Ouro Preto, MG. Programa de
Pós-graduação em Ecologia de Biomas Trópicais do Instituto de
Ciências Exatas e Biológicas da Universidade Federal de Ouro
Preto. Dissertação de mestrado.
Bertsch, A., & Domes, W. (1969). C02-Exchange in Amphistomatic
Leaves. 1. The influence of the distribution of stomata on both leaf
surfaces upon CO2 transport. Planta, 85(2), 183–193.
Boeger, M. R. T., & Wisniewski, C. (2003). Comparação da morfologia
foliar de espécies arbóreas de três estádios sucessionais distintos de
floresta ombrófila densa (Floresta Atlântica) no Sul do Brasil.
Revista Brasileira de Botânica, 26(1), 61–72.
https://doi.org/10.1590/S0100-84042003000100007
Boeger, M. R. T., Biu, C., & Goldenberg, R. (2009). Comparative leaf
architecture of Miconia sellowiana (DC.) Naudin
(Melastomataceae) from different plant physiognomies in Parana
State, Brazil. Acta Botanica Brasilica, 23(3), 657–665.
Budel, J. M., Duarte, M. R., Döll-Boscardin, P. M., Farago, P. V.,
Matzenbacher, N. I., Sartoratto, A., & Sales Maia, B. H. L. N.
(2012). Composition of essential oils and secretory structures of
Baccharis anomala , B. megapotamica and B. ochracea. Journal of
Essential Oil Research, 24(1), 19–24.
https://doi.org/10.1080/10412905.2012.645634
Diaz, S., Cabido, M., & Casanoves, F. (1998). Plant functional trait sand
environmental filters at a regional scale. Journal of Vegetation
Science, 9(1), 113–122. http://doi.org/10.2307/3237229
Duc, L., Carson, H., Da Conceição, S., Jane, M., De, C., Sereno, M., Neto,
B. (2008). Tolerância ao alumínio em plantas: toxicidade,
mecanismos e genes em espécies cultivadas aluminum tolerance in
plants: toxicity, mechanisms and genes in cultivated species.
Agrociência Pelotas, 14, 1–10.
https://doi.org/10.18539/cast.v14i3.1927

26
Carmo, F. F., & Jacobi, C. M. (2013). A vegetação de canga no Quadrilátero
Ferrífero, Minas Gerais: caracterização e contexto fitogeográfico.
Rodriguesia, 64(3), 527–541.
Carmo, F. F., de Campos, I. C., & Jacobi, C. M. (2016). Effects of fine-scale
surface heterogeneity on rock outcrop plant community structure.
Journal of Vegetation Science, 27(1), 50–59.
http://doi.org/10.1111/jvs.12342
Castro, S. A. B. (2015). Ecofisiologia de Melastomataceae dos Campos
Rupestres. Tese de Doutorado. Universidade Federal de Minas
Gerais, Instituto de Ciências Biológicas-ICB. Programa de Pós-
Graduação em Ecologia, Conservação de Manejo da Vida
Silvestre- ECMVS. Belo Horizonte.
Cornelissen, J. H. C., Lavorel, S., Garnier, E., Diaz, S., Buchmann, N.,
Gurvich, D. E., … Poorter, H. (2003). A handbook of protocols for
standardised and easy measurement of plant functional traits
worldwide. Australian Journal of Botany.
https://doi.org/10.1071/BT02124
Cornwell, W. K., Cornelissen, J. H. C., Amatangelo, K., Dorrepaal, E.,
Eviner, V. T., Godoy, O., … Westoby, M. (2008). Plant species
traits are the predominant control on litter decomposition rates
within biomes world wide. Ecology Letters, 11(10), 1065–1071.
http://doi.org/10.1111/j.1461-0248.2008.01219.x
Cornwell, W. K., & Ackerly, D. D. (2010). A link between plant traits and
abundance: evidence from coastal California woody plants. Journal
of Ecology, 98(4), 814–821. http://doi.org/10.1111/j.1365-
2745.2010.01662.x
Craine, J. M. (2007). Plant strategy theories: Replies to Grime and Tilman:
Forum. Journal of Ecology. http://doi.org/10.1111/j.1365-
2745.2007.01212.x
De Micco, V., & Aronne, G. (2012). Morpho-anatomical traits for plant
adaptation to drought. In Plant Responses to Drought Stress: From
Morphological to Molecular Features (Vol. 9783642326530, pp.
37–61). http://doi.org/10.1007/978-3-642-32653-0_2
De La Riva, E. G., Olmo, M., Poorter, H., Ubera, J. L., & Villar, R. (2016).
Leaf mass per area (LMA) and its relationship with leaf structure
and anatomy in 34 mediterranean woody species along a water
availability gradient. PLoS ONE, 11(2).
https://doi.org/10.1371/journal.pone.0148788
Drake, P. L., Froend, R. H., & Franks, P. J. (2013). Smaller, faster
stomata: Scaling of stomatal size, rate of response, and stomatal
conductance. Journal of Experimental Botany, 64(2), 495–505.
http://doi.org/10.1093/jxb/ers347
Eller, C.B., Lima, A.L., Oliveira R.S. (2013) Foliar uptake of fog water
and transport belowground alleviates drought effects in the cloud
forest tree species, Drimys brasiliensis (Winteraceae). New
Phytologist 199:151–162.
Fahn, B., Cutler. D.F. (1992). Xerophytes. Handbuch Der Pflanzenanatomie
Encycloperia of Plant Ananomy Traité D´Anatoie Vegetale.
Borntraeger: Berlin.
Franceschi, V. R., & Nakata, P. A. (2005). CALCIUM OXALATE IN
PLANTS: Formation and Function. Annual Review of Plant Biology,
56(1),41–71.
https://doi.org/10.1146/annurev.arplant.56.032604.144106
Franks, P. J., Drake, P. L., & Beerling, D. J. (2009). Plasticity in maximum
stomatal conductance constrained by negative correlation between
stomatal size and density: An analysis using Eucalyptus globulus.
Plant, Cell and Environment, 32(12), 1737–1748.
http://doi.org/10.1111/j.1365-3040.2009.002031.x
Giulietti, A.M.; Menezes, Nanuza Luiza; Pirani, José Rubens; Meguro, M.
& Wanderley, M. G. L. (1987). Flora da Serra do Cipó, MG:
caracterização e lista das espécies. Boletim de Botânica Da
Universidade de São Paulo. http://doi.org/10.11606/issn.2316-
9052.v9i0p1-151.
Gotelli, N.J., Ellison A, M. (2004). A primer of ecological statistics.
Sunderland (MA), Sinauer Associates.
Gratani, L. (2014). Plant Phenotypic Plasticity in Response to
Environmental Factors. Advances in Botany, 17.
doi:10.1155/2014/208747
Grime, J. P. P., Thompson, K., Hunt, R., Hodgson, J. G. G., Cornelissen, J.
H. C. H. C., Rorison, I. H. H., … Whitehouse, J. (1997). Integrated
Screening Validates Primary Axes of Specialisation in Plants.
Oikos, 79 (2), 259. http://doi.org/10.2307/3546011
Hafez, M Gamil. (1952) Effect of variable vapor concentrations of ethereal
oil upon transpiration and water abosrption Bull. Torrey Bot. Club
79,459,470.

27
Hartwig, I., Oliveira, A. C., Carvalho, F. I. F., Bertan, I., Silva, J. A. G.,
Schmidt, D. A. M., Reis, C. E. S. (2007). Associated mechanisms of
aluminum tolerance in plants. Semina: Ciências Agrárias, 28(2),
219–228.
Hulshof, C. M., Violle, C., Spasojevic, M. J., Mcgill, B., Damschen, E.,
Harrison, S., & Enquist, B. J. (2013). Intra-specific and inter-
specific variation in specific leaf area reveal the importance of
abiotic and biotic drivers of species diversity across elevation and
latitude. Journalof Vegetation Science, 24(5), 921–931.
http://doi.org/10.1111/jvs.12041
Jacobi, C. M., Do Carmo, F. F., Vincent, R. C., & Stehmann, J. R. (2007).
Plant communities on ironstone outcrops: A diverse and
endangered Brazilian ecosystem. Biodiversity and Conservation,
16(7), 2185–2200. http://doi.org/10.1007/s10531-007-9156-8
Jacobi, C. M., & Carmo, F. (2008). Diversidade dos campos rupestres
ferruginosos no Quadrilátero Ferrífero, MG. Megadiversidade,
4(1-2), 25–33.
Johansen, D. A., (1940). Plant Microtechnique. Mc. Graw-Hill Bood Co.,
New York.
Johnson, H. B. (1975). Plant pubescence: An ecological perspective. The
Botanical Review, 41(3), 233–258.
https://doi.org/10.1007/BF02860838
Kraus & Arduin, M (1997). Manual básico de métodos em morfologia
vegetal. Seropédica: Editora da Universidade Federal Rural do Rio
de Janeiro. v.1, 198.
Larcher W (2004). Ecofisiologia Vegetal. RiMa, São Carlos.
Lambers, H., Chapim , F.S., Pons, T.L. Plant physiological ecology. New
York: Springer, 1998. 540p.
Li, L., Mc Cormack, M. L., Ma, C., Kong, D., Zhang, Q., Chen, X., Guo, D.
(2015). Leaf economics and hydraulic traits are decoupled in five
species-rich tropical-subtropical forests. Ecology Letters, 18(9),
899–906. http://doi.org/10.1111/ele.12466
Luna, B. N. de, Defaveri, A. C. A. e, Sato, A., Bizzo, H. R., Freitas, M. de
F., & Barros, C. F. (2014). Leaf secretory tissues in Myrsine coriacea
and Myrsine venosa (Primulaceae): ontogeny, morphology, and
chemical composition of essential oils. Botany, 92, 757–766.
https://doi.org/10.1139/cjb-2014-0044.
Machado, N. A. M. (2011). Técnicas alternativas para restauração de áreas
de canga degradadas pela extração de bauxita. Dissertação de
mestrado. Universidade Federal de Ouro Preto, Brasil
MacArthur, R.H. (1972). Geographical Ecology. Harper & Row, New York.
McGill, B. J., Enquist, B. J., Weiher, E., &Westoby, M. (2006). Rebuilding
community ecology from functional traits. Trends in Ecology and
Evolution, 21(4), 178–185.
http://doi.org/10.1016/j.tree.2006.02.002
Marques, A. R., Garcia, Q. S., Passos Rezende, J. L., & Fernandes, G. W.
(2000). Variations in leaf characteristics of two species of Miconia
in the Brazilian cerrado under different light intensities. Tropical
Ecology, 41(1), 47–56. http://doi.org/10.1.1.318.6327
Messias, M. C. T. B., Leite, M. G. P., Meira-Neto, J. A. A., & Kozovits, A.
R. (2012). Fitossociologia de campos rupestres quartzíticos e
ferruginosos no Quadrilátero Ferrífero, Minas Gerais. Acta
BotanicaBrasilica, 26(1), 230–242. http://doi.org/10.1590/S0102-
33062012000100022
Messias, M. C. T. B., Leite, M. G. P., Meira Neto, J. A. A., Kozovits, A. R.,
& Tavares, R. (2013). Soil-Vegetation Relationship in
Quartziticand Ferruginous Brazilian Rocky Outcrops. Folia
Geobotanica, 48(4), 509–521. http://doi.org/10.1007/s12224-013-
9154-4
Morretes BL (1969) Contribuição estudo da anatomia das folhas de plantas
do cerrado III. Bol Bot USP 24:7–32
Mott, K. A., Gibson, A. C., & O’leary, J. W. (1982). The adaptive
significance of amphistomatic leaves. Plant, Cell & Environment.
https://doi.org/10.1111/1365-3040.ep11611750
Ogaya, R., & Peñuelas, J. (2007). Leaf mass per area ratio in Quercus ilex
leaves under a wide range of climatic conditions. The importance of
low temperatures. Acta Oecologica, 31(2), 168–173.
https://doi.org/10.1016/j.actao.2006.07.004
Oguchi, R., Hikosaka, K., & Hirose, T. (2005). Leaf anatomy as a constraint
for photosynthetic acclimation: Differential responses in leaf
anatomy to increasing growth irradiance among three deciduous
trees. Plant, Cell and Environment, 28(7), 916–927.
https://doi.org/10.1111/j.1365-3040.2005.01344.x
Pagni, A. M., & Masini, A. (1999). Morphology, distribution, and
histochemistry of secretory structures in vegetative organs of
santolina leucantha bertol. (asteraceae). Israel Journal of Plant

28
Sciences,47(4),257–263.
https://doi.org/10.1080/07929978.1999.10676782
Parkhurst, D. F. (1978). The adaptative significance of stomatal o on
ccurrenceone or both surfaces of leaves. Journal of Ecology, 66(2),
367–383. https://doi.org/10.2307/2259142
Poorter, H., Niinemets, Ü., Poorter, L., Wright, I. J., Villar, R., Niinemets,
U., … Villar, R. (2009). Causes and consequences of variation in
leaf mass per area (LMA):a meta-analysis. New Phytologist, 182(3),
565–588. https://doi.org/10.1111/j.1469-8137.2009.02830.x
Pyykko, Maire. (1966). Annales Botanici Fennici. Societas Zoologica
Botanica Fennnica Vanamo Helsinki, v.3, 607.
Rabinowitz, D., Cairns, S., & Dillon, T. (1986). Seven forms of rarity and
their frequency in the flora of the British Isles. In Conservation
Biology. The science of scarcity and diversity (pp. 182–204).
Rapini, A., Ribeiro, P. L., Lambert, S., & Pirani, J. R. (2008). A flora dos
campos rupestres da Cadeia do Espinhaço. Megadiversidade, 4(1-
2), 16–24.
Rapini, A., Ribeiro, P. L. U. Z., Lambert, S., , J. R., Jacobi, C. M., Fonseca,
F., … Taylor, N. (2008). Cadeia do Espinhaço: avaliação do
conhecimento científico e prioridade de conservação.
Megadiversidade, 4(1-2), 1–274.
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-
06832008000500032&lng=pt&nrm=iso&tlng=pt
Raven, P.H., Evert, R.F. & Eichhorn, S.E. 2001. Biologia Vegetal, 6a. ed.
Coord. Trad. J. E. Kraus. Editora Guanabara Koogan, Rio de
Janeiro. Raven, P.H. .607.
Raven, J. A. (2014). Speedy small stomata’. Journal of Experimental
Botany. https://doi.org/10.1093/jxb/eru032
Reich, P. B., Walters, M. B., & Ellsworth, D. S. (1992). Leaf Life-Span in
Relation to Leaf, Plant, and Stand Characteristics among Diverse
Ecosystems. EcologicalMonographs, 62 (3), 365–392.
http://doi.org/10.2307/2937116
Reich, P. B., Walters, M. B., & Ellsworth, D. S. (1997). From tropics to
tundra: Global convergence in plant functioning. Ecology, 94
(December), 13730–13734.
http://doi.org/10.1073/pnas.94.25.13730
Reich, P. B. (2014). The world-wide “fast-slow” plant economics spectrum:
A traits manifesto. Journal of Ecology, 102(2), 275–301.
https://doi.org/10.1111/1365-2745.12211
Rodrigues, T. M., Santos, D. C. Dos, & Machado, S. R. (2011). The role of
the parenchyma sheath and PCD during the development of oil
cavities in Pterodon pubescens (Leguminosae-Papilionoideae).
Comptes Rendus - Biologies, 334(7), 535–543.
https://doi.org/10.1016/j.crvi.2011.04.005
Rossatto, D., & Kolb, R. (2013). Leaf anatomical traits are correlated with
tree dominance in a Neotropical deciduous forest. New Zealand
Journal of Botany, 51(4), 242–251.
http://doi.org/10.1080/0028825X.2013.795904
Rozendaal, D. M. A., Hurtado, V. H., & Poorter, L. (2006). Plasticity in leaf
traits of 38 tropical tree species in response to light; relationships
with light demand and adult stature. Functional Ecology, 20(2),
207–216. http://doi.org/10.1111/j.1365-2435.2006.01105.x
Salisboury, F.B., Ross, C. W. (2013). Fisiologia das plantas. Tradução da 4
edição norte-americana. Cengage Learning. 623.
Sarthou, C., Kounda-Kiki, C., Vaçulik, A., Mora, P., & Ponge, J.F. (2009).
Successional patterns on tropical inselbergs: A case study on the
Nouragues inselberg (French Guiana). Flora - Morphology,
Distribution, Functional Ecology of Plants, 204(5), 396–407.
http://doi.org/10.1016/j.flora.2008.05.004
Siebeneichler, S, C; de Freitas, G,A., Silva, R,R.,Adorian, G,C,. Capellari,D
(2008).Morphophysiological characteristics in plants of Tabebuia
heptaphyilla (vell.) tol., in conditions of luminosity: Acta
Amaz. vol.38 no.3 Manaus.
http://dx.doi.org/10.1590/S0044-59672008000300011
Silveira, F. A. O., Negreiros, D., Barbosa, N. P. U., Buisson, E., Carmo, F.
F., Carstensen, D. W., … Lambers, H. (2016). Ecology and
evolution of plant diversity in the endangered campo rupestre: a
neglected conservation priority. Plant and Soil.
https://doi.org/10.1007/s11104-015-2637-8
Scarano, F. R. (2002). Structure, Function and Floristic Relationships of
Plant Communities in Stressful Habitats Marginal to the Brazilian
Atlantic Rain forest. Annals of Botany, 90(4), 517–524.
http://doi.org/10.1093/aob/mcf189
Siegel, S.; Castellan, N.J. 1988. Nonparametric Statistics for the Behavioral
Sciences. McGraw-Hill Inc., New York, USA, 2nd ed.

29
Simonin, K. A., Santiago, L. S., & Dawson, T. E. (2009). Fog interception
by Sequoia sempervirens (D. Don) crowns decouples physiology
from soil water deficit. Plant, Cell and Environment, 32(7), 882–
892. http://doi.org/10.1111/j.1365-3040.2009.01967.x
Souza, D. B. (2014). Absorção de Neblina Pelo Limbo Foliar de Espécies
Presentes em Complexo Rupestre. Departamento de Evolução,
Biodiversidade e Meio Ambiente do Instituto de Ciências Exatas e
Biológicas da Universidade Federal de Ouro Preto no programa de
Pós-Graduação em Ecologia de Biomas Tropicais. Dissertação de
Mestrado.
Sprent, P.; Smeeton, N.C. 2007. Applied Nonparametric Statistical
Methods. Chapman and Hall, Boca Raton, USA, 4th ed.
Tian, M., Yu, G., He, N., & Hou, J. (2016). Leaf morphological and
anatomical traits from tropical to temperate coniferous forests:
Mechanisms and influencing factors. Scientific Reports, 6(1), 19703.
https://doi.org/10.1038/srep19703
Tilman, D. (2007). Resource competition and plant traits: A response to
Craine et al. 2005: Forum. Journal of Ecology, 95(2), 231–234.
http://doi.org/10.1111/j.1365-2745.2007.01201.x
Trombin-Souza, M., Amaral, W., Pascoalino, J. A. L., Oliveira, R. A.,
Bizzo, H. R., & Deschamps, C. (2017). Chemical composition of the
essential oils of Baccharis species from southern Brazil: a
comparative study using multivariate statistical analysis. Journal of
Essential Oil ResearchOnline) Journal Journal of EssEntial Oil
rEsEarch, homepage (June), 1041–2905.
https://doi.org/10.1080/10412905.2017.1322007
Turner, I. M. (1994). Sclerophylly-primarily protective? Funcional Ecology
8:669-675.
Turner, I. M., Ong, B. L., & Tan, H. T. W. (1995). Vegetation Analysis,
Leaf Structure and Nutrient Status of a Malaysian Heath
Community. Biotropica, 27(1), 2–12.
Valim, E, A, R. (2012). Ciclagem de nutrientes no sistema atmosfera-solo-
planta em formação campestre e florestal sobre canga em Minas
Gerais. Departamento de Evolução, Biodiversidade e Meio
Ambiente do Instituto de Ciências Exatas e Biológicas da
Universidade Federal de Ouro Preto. Dissertação de mestrado 11-
12.
Verzani, J. 2014. Using R for Introductory statistics. Chapman and Hall,
Boca Raton, USA, 2nd ed.
Violle, C., Navas, M.-L., Vile, D., Kazakou, E., Fortunel, C., Hummel, I.,
& Garnier, E. (2007). Let the concept of trait be functional! Oikos,
116(5), 882–892. doi:10.1111/j.2007.0030-1299.15559.x
Westhoff, M., Zimmermann, D., Zimmermann, G., Gessner, P., Wegner, L.
H., Bentrup, F.-W., & Zimmermann, U. (2009). Distribution and
function of epistomatal mucilage plugs. Protoplasma, 235(1–4),
101–105. https://doi.org/10.1007/s00709-008-0029-0
Woolley, J. T. (1964). Water relations of soybean leaf hairs. Agronomy J.
56:569-571.
Zhou, S. X., Medlyn, B. E., & Prentice, I. C. (2016). Long-term water stress
leads to acclimation of drought sensitivity of photosynthetic capacity
in xeric but not riparian Eucalyptus species. Annals of Botany,
117(1), 133–144. https://doi.org/10.1093/aob/mcv161