antonio ferreira mendes de sousa estudo da inibição do sistema
TRANSCRIPT

AntonioFerreiraMendesdeSousa
Estudodainibiçãodosistemacomplementohumanopela
salivadeanofelinos(Diptera;Anophelinae)neotropicais
UniversidadeFederaldeMinasGerais
BeloHorizonte
Dezembro,2015

AntonioFerreiraMendesdeSousa
Estudodainibiçãodosistemacomplementohumanopela
salivadeanofelinos(Diptera;Anophelinae)neotropicais
Orientador:Prof.Dr.NelderdeFigueiredoGontijo
LaboratóriodeFisiologiadeInsetosHematófagos
DepartamentodeParasitologia-UFMG
BeloHorizonte–2015
TeseapresentadaaoProgramadePós-graduação em Parasitologia doInstituto de Ciências Biológicas daUniversidade Federal de Minas Geraiscomo requisito para obtenção do graudeDoutoremParasitologiaÁreadeconcentração:Entomologia

Aos meus pais, Antonio Ferreira de Sousa
SobrinhoeFranciscaMendesdeSousa,minha
maiorinspiração

Agradecimentos
Agradeçoaomeupaieàminhamãe,AntonioFerreiradeSousaSobrinho
eFranciscaMendesdeSousa,peloincentivoconstanteaalcançarmeusobjetivos,
peloapoioincondicionalepeloamorqueconfortaefortalece.Amovocês.
Ao meu orientador, Seu Nelder (Laboratório de Fisiologia de Insetos
Hematófagos–UFMG),pelaconfiança,ensinamentoseoportunidades.
Ao Dr. Jesus Valenzuela (Laboratory of Malaria and Vector Research –
NIH), por me receber em seu laboratório para realização do doutorado
sanduíche.
AoDr.JohnAndersen(LaboratoryofMalariaandVectorResearch–NIH),
pela ajuda direta na realização dos experimentos com as proteínas
recombinantes.
Ao Dr. Luciano Moreira e à Msc. Fernanda Rezende (Laboratório de
Malária–CPqRR/Fiocruz),pelagentilezaemnoscederosmosquitosdaespécie
Anophelesaquasalis.
AoDr.VladimirFazitodoVale(LaboratóriodeSimulídeoseOncocercose,
IOC/Fiocruz),pelaamizadeepelaajudadireta,críticasesugestõesrelacionadas
aosexperimentosrealizadosnestetrabalho,semprecommuitapaciênciaeboa
vontade.
Ao Daniel Costa Queiroz (Laboratório de Fisiologia de Insetos
Hematófagos–UFMG),oqualtiveoprazerdeorientarcomoalunodeiniciação
científica,pelaajudanarealizaçãodosexperimentosnoLFIHepelaamizade.
À Dra. Hélida Andrade e à Dra. Simone Pires (Laboratório de
Leishmanioses–UFMG),pelarealizaçãodaanálisedeespectrometriademassa
noInstitutodeCiênciasBiológicasdaUFMG.
AoDr.JoséMarcosRibeiro(LaboratoryofMalariaandVectorResearch–
NIH),pelaajudanaanálisedassequênciasdosgenesdeAnophelesalbimanus.
Ao Andre Laughinhouse e ao Kevin Lee, pela criação dos mosquitos
utilizadosnosexperimentosrealizadosnoNIH.
Aos amigos do Laboratório de Fisiologia de Insetos Hematófagos,
especialmenteaoCésarOliveira,portodaajudadentroeforadolaboratório.

AosamigosdoLaboratoryofMalariaandVectorResearch,especialmente
aoDr.FabianoOliveira,peloauxílioebomhumornolaboratório.
À Sumara e Sibele, secretárias da Pós-graduação em Parasitologia, pela
amabilidadeeporestaremsempredispostasaajudar.
À Luciana, Geni e Zuleika, secretárias da curso de Parasitologia para
graduação,pelaajudadurantemeuperíodocomobolsistaREUNI.
Ao Programade Pós-graduação emParasitologia daUFMG, nas pessoas
dos seus coordenadores, Prof.RicardoToshioFujiwara eProf.MarcosHorácio
Pereira, pelos ensinamentos de qualidade, essenciais à minha formação como
Parasitologista.
Às instituições de fomento que contribuíram para a realização deste
trabalho, CAPES, CNPq e FAPEMIG, e ao programa Ciências sem Fronteiras,
atravésdoqualrealizeioestágiodeDoutoradoSanduíchenoNationalInstitutes
ofHealth(NIH),EstadosUnidos.
A todosquediretaou indiretamente ajudaramaconcluireste trabalho,
deixoomeu“MuitoObrigado!”.

Sumário
Listadeabreviações i
Listadefiguras ii
Listadetabelas iii
Resumo iv
Abstract v
1.Introdução 15
1.1Maláriaeseusvetores 15
1.2AsalivadeAnophelessp. 20
1.3Osistemacomplemento 24
1.4Inibiçãodosistemacomplementoporartrópodeshematófagos26
2.Justificativa 29
3.Objetivos 31
3.1Objetivogeral 31
3.2Objetivosespecíficos 31
4.MaterialeMétodos 32
4.1Aprovaçãoemcomitêdeética 32
4.2Criaçãodosmosquitoseobtençãodeextratodeglândulas
salivares 32
4.3Ensaiohemolíticos 33
4.4Ensaiodeativaçãodaviadaslectinas 35
4.5Ensaiodedeposiçãodecomponentesdaviaalternativa 36
4.6WesternblotparadetecçãodeFatorBeC3a 38
4.7PurificaçãoeidentificaçãodoinibidorsalivardeA.albimanus 39
4.8RT-PCRparadetecçãodogenedaproteínagSG7emextratode
glândulassalivaresdeA.albimanus 41
4.9Clonagem,expressãoepurificaçãodasproteínasgSG7egSG7-2
deA.albimanusegSG7deA.darlingi 42
4.10IdentificaçãodoinibidorsalivardocomplementodeA.aquasalis44
4.11Westernbloteensaiohemolíticocomanticorpoanti-gSG7deA.
albimanus 45
4.12ELISAparadetectarligaçãodecomponentesdocomplementoaos
inibidoressalivares 47

4.13Ensaioderessonânciaplasmônicadesuperfície 48
4.14Análiseestatística 48
5.Resultados 49
5.1QuantificaçãodeproteínasdosEGSdasespéciesdeAnopheles
estudadas 49
5.2Ensaioshemolíticos 49
5.2.1Viaalternativa 49
5.2.2Viaclássica 51
5.3Ensaiodeativaçãodaviadaslectinas 52
5.4Ensaiodedeposiçãodecomponentesdaviaalternativa 52
5.5WesternblotparadetecçãodeFatorBeC3a 54
5.6WesternblotcomC3b,FatorBeFatorDpurificados 56
5.7PurificaçãodoinibidorsalivardeA.albimanus 56
5.8RT-PCRparadetecçãodogenedaproteínagSG7emextratode
glândulassalivaresdeA.albimanus 65
5.9ExpressãoepurificaçãodasproteínasgSG7egSG7-2deA.albimanus
egSG7deA.darlingi 66
5.10IdentificaçãodoinibidorsalivardeA.aquasalis 74
5.11WesternbloteensaiohemolíticocomIgGanti-gSG7de
A.albimanus 75
5.12ELISAparadetectarligaçãodecomponentesdocomplementoaos
inibidoressalivares 77
5.13Ensaioderessonânciaplasmônicadesuperfície 79
6.Discussão 80
7.Conclusões 90
8.Referênciasbibliográficas 91

Listadeabreviações
AAPP–proteínaanti-plaquetáriadeAnopheles(doinglês,Anophelesanti-plateletprotein)ADP–adenosinadifosfatoAMP–adenosinamonofosfatoATP–adenosinatrifosfatoBLAST–BasicLocalAlignmentSearchToolBSA–albuminadesorobovino(doinglês,bovineserumalbumine)CAM–complexodeataqueàmembranacDNA–DNAcomplementarDTT–ditiotreitolEDTA–ácidoetilenodiaminotetraacéticoEGS–extratodeglândulassalivaresEGTA–ácidoetilenoglicoltetraacéticoHPLC–cromatografialíquidadealtaeficiência(doinglês,highperformanceliquidchromatography)IL-10–interleucina10IPTG–isopropil-beta-D-tiogalactopiranosídeoISAC–Ixodesscapularisanti-complementka–constantedeassociaçãokd–constantededissociaçãokD-constantedeafinidadeMASP–serino-proteaseassociadaàMBL(doinglês,MBL-associatedserineprotease)MBL–lectinaligantedemanana(doinglês,mannanbindinglectin)MW–pesomolecular(doinglês,molecularweight)NCBI–CentroNacionalparaInformaçãoBiotecnológica(doingles,NationalCenterforBiotechnologyInformation)NIH–InstitutoNacionaldeSaúde(doinglês,NationalInstitutesofHealth)OPD–ortofenilenodiaminaPBS–soluçãosalinafosfatada(doinglês,phosphate-bufferedsaline)PCR–reaçãoemcadeiadepolimerase(doinglês,polymerasechainreaction)RNAi–RNAdeinterferênciaSDS-PAGE–geldepoliacrilamidacomdodecilsulfatodesódio(doinglês,sodiumdodecylsulfatepolyacrylamidegelelectrophoresis)SHN–SorohumanonormalTw–TweenUV–ultravioleta

ListadeFiguras
Figura1:Distribuiçãoatualdoscasosdemalárianomundo. 15Figura2:CiclobiológicodeAnophelessp.. 19Figura3:Distribuiçãoglobaldosprincipaisvetoresdemalária. 20Figura4:GlândulasalivardefêmeadeAnophelesaquasalis. 21Figura 5: Esquema representativo das três vias de ativação do sistemacomplemento. 25Figura6:SequênciasdosgenessintéticosutilizadosparaclonagemeexpressãodeproteínasrecombinantesdeA.albimanuseA.darlingi. 43Figura7:Açãodo extrato de glândulas salivares de seis espécies deAnophelessobreaviaalternativadosistemacomplementohumano. 50Figura8:EfeitodetratamentosdesnaturantessobreaatividadeinibidoradaviaalternativadoEGSdeA.albimanus. 51Figura 9: Ação do EGS de A. albimanus, A. aquasalis e A. gambiae sobre a viaclássicadosistemacomplemento. 52Figura10:Efeitodo extratode glândulas salivaresdeA.albimanus sobre a viadaslectinas. 52Figura 11: Ação do EGS de A. aquasalis e A. albimanus sobre a deposição decomponentesdaviaalternativadosistemacomplemento. 53Figura 12: Efeito doEGS deA.aquasalis sobre componentes da via alternativapreviamentedepositadosemsuperfícieativadora. 54Figura13:AçãodoEGSdeA.aquasaliseA.albimanusnaativaçãodoFatorB. 55Figura14:AçãodoEGSdeA.aquasaliseA.albimanusnaativaçãodocomponenteC3. 55Figura 15: Ação do EGS deA.albimanus na formação da C3-convertase da viaalternativa. 56Figura 16: Purificação do inibidor da via alternativa presente no EGS de A.albimanus. 57Figura17:SDS-PAGEcomfraçõesdacromatografiadefasereversadoEGSdeA.albimanus. 58

Figura 18: Quantificação das proteínas salivares de A. albimanus identificadasporespectrometriademassadafraçãopurificada1H12. 59Figura 19: Sequência do gene AALB001854 de A. albimanus disponível noVectorBase. 61Figura 20: Sequências dos dois genes anotados como gene AALB001854 deA.albimanus. 62Figura21:SequênciadaproteínasalivargSG7deA.albimanus. 63Figura22:SequênciadaproteínasalivargSG7-2deA.albimanus. 63Figura23:AlinhamentodassequênciasdospeptídeosdeproteínasalivardeA.albimanuscomasequênciadaproteínagSG7obtidanoVectorBase. 64Figura24:SequênciadaproteínasalivargSG7deA.darlingi. 64Figura25:ComparaçãodassequênciasdasproteínasgSG7dediferentesespéciesdeAnopheles. 65Figura 26: Eletroforese dos produtos da reação de PCR para amplificação dogenedaproteínagSG7deA.albimanus. 66Figura 27: Cromatografia de gel filtração para purificação das proteínasrecombinantes. 67Figura 28: SDS-PAGE com as proteínas recombinantes após o último passo depurificação. 67Figura29:Açãodasproteínas recombinantesgSG7egSG7-2deA.albimanus egSG7deA.darlingisobreaviaalternativadosistemacomplemento. 68Figura30:Açãodasproteínas recombinantesgSG7egSG7-2deA.albimanus egSG7deA.darlingisobreaviaclássicadosistemacomplemento. 68Figura31:Efeitodasproteínas gSG7e gSG7-2deA.albimanus sobre a viadaslectinas. 69Figura32:EfeitodasproteínasgSG7egSG7-2deA.albimanusnadeposiçãodoscomponentesC3,FatorB,properdinaeC9emplacascobertascomagarose.70Figura 33: Efeito da proteína gSG7 de A. darlingi sobre a deposição doscomponentesC3,FatorB,properdinaeC9emplacascobertascomagarose. 71Figura34:EfeitodagSG7deA.albimanus eFatorHsobrecomponentesdaviaalternativapreviamenteligadosàsuperfícieativadora. 72

Figura35:EfeitodasproteínasgSG7egSG7-2deA.albimanus naativaçãodoscomponentesFatorBeC3. 73Figura36:AçãodasproteínasrecombinantesgSG7egSG7-2deA.albimanusnaformaçãodaC3-convertasedaviaalternativa. 73Figura37:EletroforesedasproteínasgSG7recombinantesdeA.albimanus eA.darlingiedoEGSdeA.aquasalisemgeldepoliacrilamidacoradocomprata. 74Figura38:AlinhamentodasequênciadepeptídeogeradodeproteínasalivardeA.aquasaliscomasequênciadaproteínagSG7deA.darlingi. 75Figura39:Westernblotcomanticorpoanti-gSG7deA.albimanus. 75Figura40:WesternblotcomIgGanti-gSG7deA.albimanus. 76Figura41:EfeitodoIgGanti-gSG7sobreaatividadeanti-complementodoEGSedaproteínagSG7recombinante. 77Figura42:LigaçãodecomponentesdocomplementoàgSG7deA.albimanus.78Figura43:LigaçãodecomponentesdocomplementoaoEGSdeA.aquasalis.78Figura44:EnsaioderessonânciaplasmônicadesuperfíciecomgSG7egSG7-2deA.albimanus. 79

ListadeQuadros
Quadro1:CiclosdetemperaturautilizadosnasreaçõesdePCRparaamplificaçãodogenedaproteínagSG7deA.albimanus. 42Quadro 2: Proteínas salivares de A. albimanus identificadas na fração 1H12submetidaaespectrometriademassa. 59

Resumo
Mosquitos pertencentes ao gênero Anopheles são os transmissores
naturais de malária no mundo. A transmissão ocorre durante a hematofagia,
quando esses insetos inoculam na pele do hospedeiro formas infectantes do
Plasmodiumsp.juntamentecomoconteúdodesuasglândulassalivares.Asaliva
dessesmosquitoscontémmoléculasimportantesparaosucessodahematofagia,
como vasodilatadores e anticoagulantes. Entretanto, há pouca informação
existente sobre a interação de componentes salivares de anofelinos com o
sistema imune do hospedeiro vertebrado. Este trabalho teve como objetivo
demonstrar e caracterizar a atividade inibidora do sistema complemento na
saliva de anofelinos, principalmente A. albimanus e A. aquasalis, importantes
vetoresdemalárianaAméricaLatina.Paraisso,ensaioshemolíticos,ensaiosde
deposição e Western blots para detectar ativação de componentes do
complementoforamrealizadosnapresençadeextratodeglândulassalivaresdos
mosquitos.Osresultadosdessesexperimentosmostraraminibiçãosignificativae
dose-dependente da via alternativa do sistema complemento pelo extrato de
glândulassalivaresdasduasespéciescitadas.OinibidorsalivardeA.albimanus
foipurificadoemHPLCeidentificadoporespectrometriademassacomosendoa
proteínagSG7.Ogenedessaproteínafoientãoclonadoeexpressoembactérias.
Aproteínarecombinanteapresentouatividadetãopotentequantooextratode
glândulasalivardeA.albimanus.Anticorposproduzidoscontraarecombinante
foram capazes de reconhecer tanto a recombinante como a proteína nativa
presentenoextratodeglândulas,alémdebloquearaatividadedasduasformas
da proteína. Através de ensaio de ELISA e de ressonância plasmônica de
superfície, demonstramos que a gSG7 liga-se em properdina, um regulador
positivo da via alternativa do complemento, bloqueando a ação estabilizadora
dessavia.Estaéaprimeiradescriçãodeuminibidordocomplementonasaliva
demosquitos,eapresentapotencialimportâncianoprocessohematofágicoena
transmissãodoPlasmodiumsp..
Palavras-chave: Anopheles albimanus, Anopheles aquasalis, glândulas salivares,
sistemacomplemento,gSG7,inibiçãodocomplemento.

Abstract
Anophelesmosquitoesare thenaturalvectorsofmalaria throughout the
world. The transmission occurs during bloodmeal, when the insects inject
infectiveformsofPlasmodiumtogetherwiththecontentsoftheirsalivaryglands.
Their saliva presents active molecules important for hematophagy, such as
vasodilators, anticoagulant and platelet inhibitors. However, little is known
abouttheinteractionsofAnopheles’salivarycomponentswiththehost’simmune
system. The aim of this work was to demonstrate and to characterize anti-
complement activity in the saliva of neotropical anophelines, specially A.
albimanus and A. aquasalis, important malaria vectors in Latin America.
Hemolytic assays, ELISA assays and Western blots for detecting activation of
complement components were performed in the presence and absence of
salivaryglandsextractsofbothmosquitospecies.Theresultsshowedsignificant
and dose-dependent inhibition of the alternative pathway by the saliva of the
twospecies.ThesalivaryinhibitorofA.albimanuswasthenpurifiedusingHPLC
and identifiedbymassspectrometry,asbeingagSG7protein.Thegeneof that
proteinwas cloned and expressed inE.coli bacteria. The recombinant protein
presented anti-complement activity as potent as the salivary glands extracts.
Anti-recombinant gSG7 IgGantibodiesproduced in rabbitswere able todetect
both the recombinant and the native protein in the glands extract. The
antibodiesalsoblockedgSG7activityinthehemolyticassays.ThroughELISAand
surfaceplasmonresonance,weshowedthatgSG7bindsdirectlytoproperdin,a
positiveregulatorofthealternativepathwayofthecomplementsystem.Thisis
the first report of an anti-complementmolecule inmosquito saliva,which has
potentialroleinblood-feedingandparasitetransmissionbythevector.
Key-words: Anopheles albimanus, Anopheles aquasalis, salivary glands,
complement system, gSG7, complement inhibition.

15
1.Introdução1.1Maláriaeseusvetores A malária humana é uma das principais doenças parasitárias no mundo,
provocadaporprotozoáriosdogêneroPlasmodiumetransmitidapormosquitosdo
gêneroAnopheles.Adoençaéresponsávelporelevadamortalidadeemorbidadeem
países tropicais e subtropicais, sendo atualmente endêmica em 97 países,
acometendoanualmentecercade200milhõesdepessoasecommaisde45%da
populaçãomundial sob o risco de infecção (WHO, 2014). Estima-se que cerca de
580.000 pessoas morreram de malária em 2013, principalmente crianças com
menos de 5 anos de idade residentes no continente africano (Figura 1) (WHO,
2014).
Figura 1: Distribuição atual dos casos de malária no mundo (Adaptado deWHO,2014).
NasAméricas,amaláriaocorreem21países,comcercade420.000casos
confirmadose82mortesem2013.Brasil,ColômbiaeVenezuelaapresentaram
72%doscasosdestecontinente(WHO,2014).NoBrasil,adoençapredominana
região amazônica, onde ocorremmais de 99%dos casos notificados (Oliveira-
Ferreiraetal.2010).Amaláriaapresentadistribuiçãoheterogêneanessaregião,
sendoasáreasdemaiortransmissãoaquelasrecém-ocupadas,áreasdegarimpo

16
epopulaçõesribeirinhas.Nessasáreas,odesenvolvimentodovetoreexposição
da população aomesmo são favorecidos pelos hábitos dosmoradores e pelas
condiçõesdeficientesdemoradia, favorecendoassimamanutençãodamalária
endêmica(Alvesetal.2002,Sampaioetal.2015).
OsplasmódiossãoprotozoáriospertencentesaofiloApicomplexa,família
Plasmodiidae e ao gênero Plasmodium. Atualmente cinco espécies são
consideradas causadoras de malária humana: Plasmodium falciparum,
Plasmodium vivax, Plasmodium malariae, Plasmodium ovale e Plasmodium
knowlesi,sendoasduasprimeirasasmaisprevalentesnomundoeasprincipais
causadoras da doença em humanos (Cox-Singh & Singh 2010,WHO, 2014). P.
knowlesi possui macacos como hospedeiros vertebrados, mas recentemente
casosdemaláriahumanaprovocadaporessaespécietêmsidorelatadosnaÁsia
(Cox-Singh & Singh, 2010). De forma geral, a malária é caracterizada por
episódios de calafrios, febre com temperatura igual ou superior a 40ºC e
sudorese intensa.É comumopaciente apresentar juntamente cefaléia,mialgia,
náuseasevômitos(MS,2010).AmaláriagraveécausadaprincipalmenteporP.
falciparum, havendo obstrução de capilares sanguíneos em órgãos como rins,
pulmões, coração e cérebro, sendo comumente fatal se não tratada a tempo
(Miller et al. 2002, Weatherall et al. 2002). Emmulheres grávidas, a infecção
causaanemiaedisfunçãoplacentária, prejudicandoo crescimentodo feto, que
nasce com peso reduzido, favorecendo a mortalidade infantil nas áreas
endêmicas(Weatheralletal.,2002).Recentemente,casosdemaláriagraveporP.
vivax também têm sido relatados, caracterizados principalmente por anemia
severa,icterícia,trombocitopeniaedificuldaderespiratória(Lacerdaetal.2012).
Osplasmódiosapresentamciclobiológicoheteroxênico,divididoemciclo
assexuadonohospedeirovertebradoesexuadonomosquitovetor.Ainfecçãodo
hospedeiro vertebrado ocorre naturalmente quando fêmeas de anofelinos
infectadas, ao realizarem a hematofagia, inoculam juntamente com sua saliva
formas infectantesdoparasito,denominadasesporozoítos.Esta formaevolutiva
do Plasmodium acumula-se na glândula salivar do mosquito e, em um repasto
infectante,umafêmeainoculaemmédia120esporozoítosnapeledohospedeiro
(Medica&Innis2005).

17
Partedosparasitosinoculadosnapelepenetramemcapilarescutâneose
caem na corrente sanguínea. Os esporozoítos atravessam o citosol de várias
célulasparaativarasviasnecessáriasàinvasãoeaofuturodesenvolvimentodo
parasito no fígado (Mota & Rodriguez 2004). Em alguns minutos invadem
hepatócitos, diferenciam-se em formas arredondadas e iniciam o ciclo pré-
eritrocítico, no qual realizammerogonia e gerammilhares de células-filhas, os
merozoítos.Ohepatócitoparasitadoéentãorompido,liberandoosparasitosem
vesículas conhecidas como merossomos (Sturm et al. 2006). Muitos dos
merozoítos liberados são fagocitados pelas células de Kupffer, mas os que
conseguem sobreviver invadem hemácias e dão origem ao ciclo eritrocítico.
Nessascélulas,osplasmódiosmultiplicam-senovamentepormerogonia,gerando
dezenas de milhares de novos merozoítos. Com o rompimento da célula, os
merozoítos liberados invadem novas hemácias, repetindo novamente a
multiplicação assexuada. Após algum tempo de evolução da infecção (cerca de
seteadozediasparaP.falciparume24horasparaP.vivax),aparecemnointerior
das hemácias formas que não mais se reproduzem. São os gametócitos ou
gamontes(Weatheralletal.2002,Bousemaetal.2010).
Quando um anofelino se alimenta em hospedeiro com gametócitos
circulantes,eleingereestasformassexuadasdoparasito.DevidoaalteraçãodepH
e temperatura e produção de ácido xanturênico no trato digestivo do inseto, os
gametócitos se diferenciam em microgametas masculinos ou macrogameta
feminino (um gametócito pode dar origem a oito microgametas masculinos
flageladosouaumúnicomacrogametafemininoarredondado),quesefecundame
formamozigoto(Billkeretal.1997,Billkeretal.1998,Josling&Llinás2015).Cerca
de20horasapósafusãodosdoisnúcleos,ozigotocomeçaamovimentar-seepassa
aserchamadodeoocineto.Esteatravessaacélulaintestinaldoinseto,alojando-se
entreoepitélioeamembranabasaldointestino.Nesta localização, transforma-se
em oocisto e inicia o processo demultiplicação esporogônica, dando origem aos
esporozoítos.Ooocistomadurorompe-se,liberandoosesporozoítos,quecaemna
hemolinfa e migram para as glândulas salivares do inseto, invadindo-as. Em um
novo repasto sanguíneo, estas formas do parasito são inoculadas no hospedeiro

18
juntamente coma salivado vetor, completando seu ciclo biológico (Revisadopor
Pimentaetal.2015).
OsanofelinossãoinsetospertencentesàordemDiptera, famíliaCulicidaee
sub-famíliaAnophelinae.OgêneroAnopheles,compreendemaisde400espécies,
embora apenas aproximadamente 60 tenham importância epidemiológica para
malária (Harbach 2004). São vulgarmente conhecidos comomosquito-prego ou
carapanãnoBrasilediferenciam-sedeoutrosculicídeosporapresentarempouso
perpendicular em relação ao substrato, palpos do mesmo comprimento da
probóscide, asas manchadas, ovos com expansões cuticulares formando os
flutuadores, e larvas semsifão respiratório (Forattini, 2002).Taxonomicamente,
distribuem-se em seis subgêneros: Anopheles, Cellia, Kertezia, Lophopodomyia,
NyssorhynchuseStethomyia,dosquaisAnopheles,CelliaeNyssorhynchuspossuem
omaiornúmerodeespécies(Harbach2004).
Comotododíptero,osanofelinosapresentamcicloholometábolo,passando
pelas fases de ovo, quatro estádios larvais, pupa e adultos (Figura 2). As fases
imaturasdocicloocorrememcoleçõesdeáguacomcondiçõesvariáveisdeacordo
com a espécie de Anopheles, onde as larvas se alimentam de microrganismos
aquáticos. Os adultos se alimentam de sucos vegetais e apenas as fêmeas são
hematófagas, necessitando do sangue ingerido para maturação dos folículos
ovarianos.De formageral,acópuladosanofelinossedáemenxames, formando
‘nuvens’ nas quais os machos agarram as fêmeas com suas terminálias,
fecundando-as(Forattini,2002).
Entre as espécies do gênero Anopheles existem importantes diferenças
biológicas que permitem sua adaptação a diferentes ambientes, possibilitando a
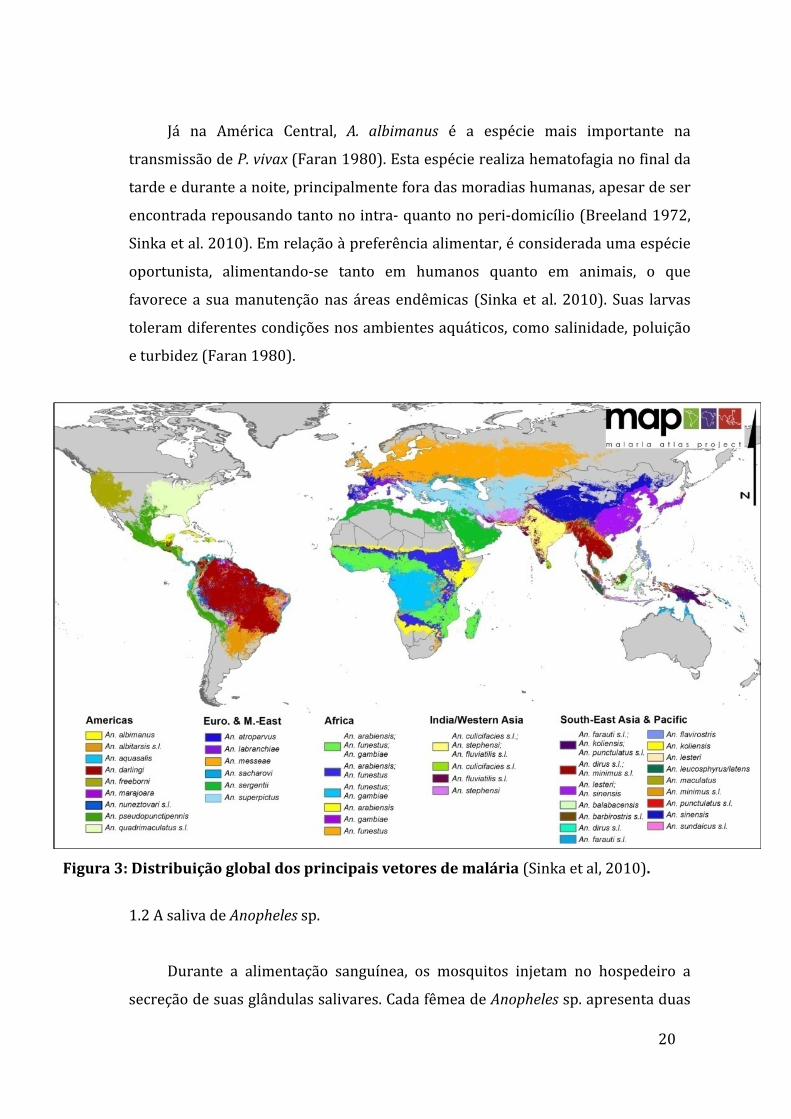
ampla distribuição desses mosquitos nos seis continentes (Figura 3), e a
transmissãoglobaldemalária (Sinkaetal.2010).Entreosprincipaisvetoresde
malária no Velho Mundo estão: A. gambiae (principal vetor na África sub-
sahariana), A. stephensi (principal vetor nas áreas urbanas da Índia) e A. dirus
(principalvetornosudesteasiático)(Sinkaetal.2010).NaAméricadoNorte,A.
pseudopunctipeniséconsideradooprincipaltransmissordemalárianoMéxico,e
A.freebornieA.quadrimaculatusforamimportantesvetoresdadoençanascostas

19
oesteeleste,respectivamente,dosEstadosUnidosatéadécadadequarenta(CDC,
2015).
Figura 2: Ciclo biológico de Anopheles sp.. Os anofelinos possuem cicloholometábolo,passandopelas fasesdeovo, larva,pupaeadulto [Fotos:StephenDogget(larvaepupa),SamCotton(adultos),RoxanneConnely(ovos)].
NaAméricadoSul,A.darlingiéaprincipalespécietransmissorademalária.
NoBrasil,elaéencontradaemquasetodoopaís,comexceçãodoextremosulede
certasáreasmuitosecasnaregiãoNordeste(Figura3).Éaltamentesusceptívelà
infecção por Plasmodium vivax e P. falciparum e apresenta alto grau de
antropofilia, alimentando-se predominantemente no peri-domicilio (Forattini,
2002,Giletal.2007).Entretanto,nasregiõescosteirasondeamaláriaéendêmica,
A. aquasalis é incriminado como o principal vetor. Essa espécie possui como
criadouroscoleçõesdeáguasalobrae,portanto,suaslarvassuportamaltosteores
desalinidade(Forattini,2002;Silvaetal.2006).Devidoaestacaracterística,sua
distribuiçãoéquaseexclusivamentelitorânea,ocorrendonasregiõescosteirasdas
AméricasCentraledoSul.EmBelém(PA),Povoaetal(2003)relataramíndicesde
infecçãonaturaldeA.aquasalisde1,18%(edeA.darlingide1,61%),contribuindo
deformasignificativanatransmissãodadoençanessacidade.
20
Já na América Central, A. albimanus é a espécie mais importante na
transmissãodeP.vivax(Faran1980).Estaespécierealizahematofagianofinalda
tardeeduranteanoite,principalmenteforadasmoradiashumanas,apesardeser
encontradarepousandotantonointra-quantonoperi-domicílio(Breeland1972,
Sinkaetal.2010).Emrelaçãoàpreferênciaalimentar,éconsideradaumaespécie
oportunista, alimentando-se tanto em humanos quanto em animais, o que
favorecea suamanutençãonasáreasendêmicas (Sinkaet al. 2010). Suas larvas
toleramdiferentescondiçõesnosambientesaquáticos,comosalinidade,poluição
eturbidez(Faran1980).
Figura3:Distribuiçãoglobaldosprincipaisvetoresdemalária(Sinkaetal,2010).
1.2AsalivadeAnophelessp.
Durante a alimentação sanguínea, os mosquitos injetam no hospedeiro a
secreçãodesuasglândulassalivares.CadafêmeadeAnophelessp.apresentaduas

21
glândulas salivares, cada uma formada por três lobos (Figura 4), localizadas no
interiordoseutórax(Jariyapan&Choochote2007).
Figura 4: Glândula salivar de fêmea de Anopheles aquasalis. As glândulassalivares de anofelinos são compostas por dois lobos laterais (LL) e um lobomedial(LM)(Foto:KelMartins).
A saliva dos anofelinos, assim como a de outros artrópodes hematófagos,
apresenta moléculas importantes para o processo de hematofagia, como
inibidores de agregação plaquetária, anti-coagulantes e vasodilatadores.
Recentemente, com estudos de sialomas (transcriptomas e proteomas de
glândulassalivares)dessesinsetos,algumasdessasmoléculasforamidentificadas
ecaracterizadas(Titusetal.2006).
Anopheles albimanus apresenta em sua saliva um peptídeo de 6,5 kDa
denominado Anophelina. Esta molécula apresenta atividade inibidora de α-
trombina, proteína responsável pela clivagem de fibrinogênio e consequente
geração de fibrina para formação do coágulo sanguíneo. Assim, a Anophelina
constituiumimportantecomponentesalivardificultandoahemostasianolocalda
picada,favorecendoorepastosanguíneopelovetor(Valenzuelaetal.1999).
A proteína anti-plaquetária de Anopheles (AAPP), descrita na saliva de A.
stephensi, inibe fortemente a agregação plaquetária ligando-se ao colágeno
subendotelialdovasosanguíneolesadopelaspeçasbucaisdovetor.Destaforma,
bloqueia a adesão e ativação de plaquetas ao colágeno. Seus altos níveis de
expressão sugerem que a AAPP possui um importante papel na localização de

22
vasossanguíneosenaprevençãodahemostasia(Yoshidaetal.2008).Tambémna
salivadeA.stephensi, foi encontradaumaproteínada famíliaD7que liga-se em
tromboxanoA2eleucotrienos,atuandocomoinibidordeagregaçãoplaquetáriae
possivelmentecomoanti-inflamatório(Alvarengaetal.2010).
Outra importante molécula inibidora de agregação plaquetária, a enzima
apirase,jáfoidescritanasalivadeA.darlingi,A.gambiaeeA.dirus(Moreiraetal.
2001,Jariyapan&Choochote2007).AapiraseéumanucleotidasequeclivaATPe
ADP em AMP e fosfato inorgânico e desse modo inibe a agregação plaquetária
dependente de ADP (Ribeiro 1995). Mosquitos que tiveram o gene codificador
destaproteínasilenciadoporRNAinecessitaramdodobrodotempodemosquitos
normais para localizar vasos sanguíneos, retardando assim a hematofagia
(Boissonetal.2006).
O sialomadeA.gambiaemostrou que esta espécie demosquito apresenta
várias proteínas salivares relacionadas à hematofagia (como mucinas, que
lubrificamaspeçasbucaisdoinseto)eàdigestãodeproteínasedecarboidratos,
(comopeptidaseseglicosidases,respectivamente).Comensaiosdesilenciamento
gênico por RNAi, ficou demonstrada a importância de determinadas proteínas
salivaresnalocalizaçãodovasosanguíneoenosucessodahematofagia(Dasetal.
2010).
Recentemente, Francischetti et al (2014) relataram atividade de
hemaglutininanoextratodeglândulassalivaresdecincoespéciesdeanofelinos.O
extratodeglândulassalivaresdeA.gambiaeapresentouatividadesobrehemácias
de diversas espécies de vertebrados, como humanos, equinos e bovinos. Entre
outras funções, essas lectinas podem ser ingeridas durante a hematofagia,
aglutinando hemácias no intestino do mosquito e favorecendo o processo
digestivo(Francischettietal.2014).
Além da importância no processo hematofágico, alguns trabalhos têm
demonstradoa importânciadasalivadovetorna infecçãopelosesporozoítosde
Plasmodiumsp.Utilizandomodelosmurinos,Vaughanetal(1999)demonstraram
queesporozoítosdeP.bergueiinoculadosporpicadasdeA.stephensiforammais
infectantes que esporozoítos inoculados intravenosamente por injeção com
seringa. Rocha et al (2004) encontraram resultados semelhantes, observando

23
níveisaumentadosdeparasitemiaporP.gallinaceumemgalinhasinoculadascom
esporozoítosjuntamentecomsalivadeAedesfluviatilis.
Assimcomodemonstradoparaoutrasdoençastransmitidasporartrópodes,
comoleishmaniose(Oliveiraetal.2015)edoençadeLyme(Schuijtetal.2011a),a
pré-imunizaçãodecamundongoscomsalivadovetorA.stephensi,protegeuesses
animais contra a infecção por P. yoelii (Donovan et al. 2007). Estes resultados
fortalecem a ideia da utilização de antígenos salivares como componentes de
vacinasparadoençastransmitidasporartrópodes.
Osmecanismospelosquaisasalivadeflebotomíneosecarrapatosdogênero
Ixodes favorecem a infecção por Leishmania sp. e Borrelia burgdorferi,
respectivamente, vêm sendo elucidados há mais de duas décadas e várias
informações já foram adquiridas sobre a interação da saliva desses artrópodes
comoparasitoporelestransmitidoeosistemaimunedohospedeiro(Titusetal.
2006). Entretanto, há relativamente poucos trabalhos na literatura sobre a
interação da saliva de anofelinos com o sistema imune de vertebrados e os
processospelosquaiselapodeauxiliar nainfecçãopelosplasmódios.Ospoucos
trabalhos descritos até o momento foram realizados apenas com extrato de
glândulasouproteínasalivarrecombinantedeumaespécie,A.stephensi.
Owhashietal(2001,2008)demonstraramapresençadeumaglicoproteína
salivar de 200 kDa com atividade quimiotática para neutrófilos e eosinófilos,
contribuindo para a resposta inflamatória no local da picada. Demeure et al.
(2005) demonstraram que saliva de A. stephensi induz a degranulação de
mastócitos na pele de camundongos, provocando extravasamento de líquido e
rápida infiltração de neutrófilos. Em linhagens de camundongos deficientes de
mastócitos,essainfiltraçãonãofoiobservada,reforçandoopapeldosmastócitos
na resposta inflamatória provocada pela picada de anofelinos. O mesmo grupo
relatou a diminuição de respostas por linfócitos T na presença de extrato de
glândulasalivar invitroe invivo.Osautoresdemonstraramqueadiminuiçãoda
resposta de hipersensibilidade do tipo tardia foi dependente da ativação de
mastócitos e mediada por IL-10 (Depinay et al. 2006). Entretanto, não há
informação sobre outras possíveis atividades imunomoduladoras em outras

24
espécies de anofelinos, permanecendo as interações na infecção malárica um
campodepesquisapromissor.
1.4Osistemacomplemento
O sistema complemento (SC) é um dos principaismediadores da resposta
imuneinatadosvertebrados,formadoporcercade30proteínassolúveisnoplasma
ou associadas à superfície celular. Este sistema atua em esquema de cascata
proteolítica, sendo sua principal função o reconhecimento, opsonização e lise de
microrganismos invasores e células alteradas do hospedeiro (Sim& Laich 2000).
Possui também importante papel na resposta imune adaptativa, auxiliando a
apresentaçãodeantígenosetornandoarespostahumoralmaiseficiente(Morganet
al.2005).
Sua ativação se dá através de três vias: a clássica, a das lectinas e a
alternativa (Figura 5). A via clássica é ativada quando o componente C1q,
juntamentecomasserino-proteasesC1reC1s(complexoC1),seligamaanticorpos
IgM e IgG aderidos à superfície de patógenos invasores. Uma vez ativado, o
complexoC1écapazdeclivaroscomponentesC4eC2,eseusfragmentosmaiores,
C4beC2a,formamocomplexoC4b2asobreasuperfícieativadora.Estecomplexo
possui a capacidade de clivar o componente C3, omais importante e abundante
componentedosistemacomplemento,emC3aeC3b(Dunkelberger&Song2010).
O fragmento C3a é uma anafilotoxina, responsável pela quimiotaxia de
leucócitos, epelo recrutamentoedegranulaçãodemastócitos, causandoaumento
da permeabilidade vascular e facilitando a infiltração celular em pontos de
inflamação (DiScipio & Schraufstatter 2007). Já C3b é uma opsonina que liga-se
covalentemente à superfície ativadora. Esta opsonizaçãomarca o patógeno como
“estranho”,sinalizandoparaquecélulasdedefesaofagocitem.Alémdisso,C3bse
ligapróximoaocomplexoC4b2a,formandoaC5convertase(C4b2a3b),queclivao
componenteC5emC5aeC5b.Estedáinícioàmontagemdocomplexodeataqueà
membrana (CAM). C6 se liga ao C5b recém ativado e logo forma um grande
complexocompostoporC5b,C6,C7,C8eC9.DuranteamontagemdoCAM,várias

25
moléculas do componente C9 inserem-se na bicamada lipídica do patógeno,
formandoporosquelevamàlisecelular(Dunkelberger&Song2010).
Figura 5: Esquema representativo das três vias de ativação do sistemacomplemento.AviaclássicaéativadaquandoC1qseligaaanticorposaderidosao antígeno; a via das lectinas quando a lectina ligante de manana (MBL)encontra carboidratos específicos associados a patógenos; e a via alternativa éativada quando C3 sofre hidrólise espontânea e forma a C3-convertase inicialdestavianapresençados fatoresBeD.As trêsviasconvergemparaumpontocomum,aativaçãodocomponenteC3,eculminamcomaformaçãodoComplexode Ataque a Membrana, responsável pela formação de poros na membranacelular(AdaptadodeLewis&Ram2014).
Aviadas lectinas foi amais recentementedescrita.Elaéativadade forma
similar à via clássica, entretanto é independente de anticorpos. As moléculas de
ativação desta via são a lectina ligante de manana (MBL) ou ficolinas, que
reconhecem carboidratos específicos presentes na superfície de patógenos, como
manana, N- acetilglicosamina ou fucose. AMBL é semelhante estruturalmente ao

26
componenteC1qdavia clássica, e como tal, umavez ligadaaopatógeno, ativaas
proteasesMASP-1eMASP-2.Umavezativadas,asMASPsclivamC4eC2,formando
aC3convertaseC4b2a,queativaocomponenteC3,dandocontinuidadeàcascata
proteolíticaatéaformaçãodoCAM.MBLeMASPssãomuitomenosabundantesno
soroqueC1q,C1reC1s,tendo,portanto,umaimportânciamenordentrodosistema
complemento(Dunkelberger&Song2010).
Aviaalternativaé iniciadapelahidróliseespontâneadeC3, com formação
deC3bsolúvel(C3b-H2O),queseligaaocomponenteFatorB.Umavezassociados,o
complexo C3b-H2O-B é clivado pela serino-protease Fator D, que cliva o Fator B,
formandoC3b-H2O-Bbsolúvel.Esteprodutoécapazdeativaroutrasmoléculasde
C3.SealgumamoléculadeC3bseligaaumasuperfícieestranha,comoasuperfície
deumpatógeno,elavaicontinuarativaeécapazderecebermoléculasdoFatorB
que é imediatamente ativado a Bb, formando C3bBb (a C3-convertase da via
alternativa) sobre o organismo estranho (Dunkelberger e Song, 2010). Com a
ligaçãodemaisumamoléculadeC3b,forma-seaC5convertase(C3bBb3b)davia
alternativa, que ativa o componente C5. Este processo também culmina na
formaçãodoCAM.Estaviapodeseramplificadaatravésdaligaçãodeproperdinaà
C3-convertase. A properdina (ou Fator P) estabiliza o instável complexo C3bBb
funcionandocomoumaproteínaativadora, ligando-seaomesmo tempoemsítios
dos componentes C3b e Bb que impossibilitam a ação de reguladores negativos
destavia(Torreiraetal.2009).Porestaremconstanteativação,estaviaapresenta
reguladores negativos específicos, como Fator H e Fator I. O Fator H atua na C3
convertase removendo Bb do complexo C3bBb e servindo como cofator para a
proteólisemediadaporFatorI(Ricklinetal.2010).
1.5Inibiçãodosistemacomplementoporartrópodeshematófagos
O sistema complemento do hospedeiro exerce forte pressão evolutiva nos
parasitos, que por sua vez desenvolvem mecanismos de proteção contra a ação
deletériadosefeitosdessesistema.Exemplodissosãoosartrópodeshematófagos,
quedesenvolveraminibidoresdocomplementopresentesnasalivae/ouconteúdo
intestinal(Barrosetal.2009,Schroederetal.2009,Mendes-Sousaetal.2013).

27
A inibição do sistema complemento por um artrópode hematófago foi
descrita pela primeira vez em estudos com saliva de Ixodes dammini, onde
demonstrou-se que a saliva desse carrapato possui a capacidade de inibir a via
alternativa do complemento humano (Ribeiro 1987), além de inativar as
anafilotoxinas produzidas durante a ativação da cascata (Ribeiro & Spielman
1986). Valenzuela et al. (2000) purificaram, clonaram e expressaram a proteína
salivar anti-complemento de Ixodes scapularis (ISAC) e demonstraram que essa
molécula de 18,5 kDa é capaz de inibir apenas a via alternativa, inibindo a
deposiçãodeC3b e fatorB em superfícies ativadoras, alémde remover o fatorB
ligado anteriormente. A saliva de Ixodes ricinus também possui a capacidade de
inibiraviaalternativaeprevinea clivagemdoFatorBeC3 (Lawrieet al.2005).
Moléculassemelhantesà ISACforamencontradasemsuasalivaeatuamdemodo
parecido,inibindoaativaçãodosmesmoscomponentes(Couvreuretal.2008).
Estudandoainibiçãodocomplementoportriatomíneos,Barrosetal(2009)
observaramquetantoasalivaquantooconteúdointestinaldasespéciesTriatoma
infestans,T. brasiliensis eRhodniusprolixus foram capazes de atuar sobre as vias
clássicae alternativado complementohumano, causandodiminuição significativa
daatividadedessasvias.
JáasalivadeLutzomyialongipalpis,principalvetordeLeishmaniainfantum
nas Américas, possui atividade inibidora da via clássica e da via alternativa do
complementohumano(Cavalcanteetal.2003).Vale(2011)descreveuomecanismo
de ação do inibidor salivar da via clássica desse vetor. Através de cromatografia
líquida (HPLC) e testes imunoenzimáticos,mostrou tratar-se de uma proteína de
aproximadamente 11 kDaque atua ligando-se ao componenteC1q, bloqueando a
ativaçãodeC1reC1se,consequentemente,ofluxonormaldacascata.Alémdisso,
demonstroutambémqueaproteínarecombinanteLJM19,cuja formanaturalestá
presentenaglândulasalivardeL.longipalpis,apresentaamesmaaçãosobreavia
clássica do sistema complemento de seres humanos (Vale, 2011). Essa atividade
também foi observada sobre o SC de cães, ratos e cobaias (Mendes-Sousa et al.,
2013).
Recentemente, Khattab et al (2015) demonstraram que uma das formas
pelas quais fêmeas deAnophelesgambiae eA. stephensi se evadem do ataque do

28
sistema complemento ao nível de intestino, é através da ligação de fator H do
sanguedohospedeiroàscélulasintestinais.Osautoresmostrarampormicroscopia
confocal a ligação do regulador às células epiteliais do intestino do mosquito e
verificaram que mosquitos que se alimentaram com sangue contendo anticorpo
anti-Fator H apresentaram maior índice de mortalidade e menor índice de
fecundidade(Khattabetal,2015).Paranossoconhecimento,porém,nãohárelato
naliteraturadeinibidoresdosistemacomplementonasalivadeculicídeos.
Para os vetores hematófagos, a inibição do SC tem funções muito
importantes. Durante a hematofagia, essa ação levaria à diminuição parcial da
resposta inflamatória no local da picada devido à produção reduzida de
anafilotoxinas, diminuindo assim a percepção pelo hospedeiro e permitindo o
repastosanguíneocompleto(Schroederetal,2009).Adicionalmente,épossívelque
nolocaldapicadaosinibidoressalivaresreduzamconsideravelmenteaquantidade
de antígenos salivares opsonizados com C3b, diminuindo assim a produção de
anticorpos contraproteínas salivares,preservandosuaspropriedadesnecessárias
paraosucessodahematofagia(Barrosetal,2009).
Alémdisso,afunçãoaparentementemaisóbviadessesinibidoressalivaresé
aproteçãodoepitélio intestinalcontra injúriasmediadaspelocomplemento.Uma
vezqueotratodigestivodosinsetoséformadoporapenasumacamadadecélulas
(Billingsley 1990), o dano provocado pela ativação do complexo de ataque à
membranapoderialevaràsuarupturaemortedoinseto(Barrosetal,2009).

29
2.Justificativa Apesardosprogramasdecontroledamalária teremconseguidoreduzir
em torno de 45% a incidência da doença nomundo nos últimos quinze anos,
devidoàausênciadeumavacinaeficiente,daresistênciacrescentedosparasitos
aos anti-maláricos existentes e da pouca eficácia dos métodos de controle do
vetor em áreas endêmicas, a malária ainda constitui um sério e ameaçador
problemadesaúdepúblicanessasáreas(WHO,2014).
AnophelesalbimanuseA.aquasalissãoimportantesvetoresdemaláriana
América Latina e, entre as espécies de anofelinos neotropicais, são as mais
comumente mantidas em colônias de laboratório. Isso proporciona acesso a
essesinsetos,permitindosuautilizaçãoemtrabalhosdepesquisaemdiferentes
campos,comooestudodasuarespostaimuneàinfecçãoporPlasmodium(Bahia
et al. 2011), estudos de resistência a inseticidas (Molina & Figueroa 2009),
observaçõessobreseudesenvolvimentoembrionário(deCarvalhoetal.2002)e
investigações demoléculas salivares com função anti-homeostática (Ribeiro &
Nussenzveig1993,Valenzuelaetal.1999).
Entretanto, para nosso conhecimento, na literatura não consta trabalho
sobreainteraçãodasalivadessasespéciesdeAnophelescomosistemaimunedo
hospedeiro vertebrado, nem a identificação e caracterização de moléculas
salivares específicas envolvidas na interação vetor-sistema complemento,
constituindo assim um importante e promissor campo de pesquisa. A
caracterização de como os artrópodes hematófagos conseguem se evadir do
sistema do complemento tem sido tema de vários estudos (Schroeder et al,
2009) e tem constituído uma das linhas de pesquisa do nosso laboratório
(Cavalcante et al, 2003; Barros et al, 2009; Mendes-Sousa et al, 2013), na
tentativadeelucidaraslacunasnoconhecimentodasinteraçõesvetor-parasito-
hospedeirovertebrado.
Estudosvoltadosparaainteraçãodasalivadovetorcomosistemaimune
dehospedeirosfazem-seaindamaisnecessáriosdepoisdademonstraçãoinvivo
deproteçãocontrainfecçãoporPlamosdiumsp.conferidaatravésdaimunização
porpicadasdeanofelinosnãoinfectados(Donovanetal,2007).Observaçõesde
comoantígenossalivarescompotencialvacinalinteragemcomosistemaimune

30
de seres humanos são importantes para a determinação de sua utilização em
possíveisvacinas.
Considerando este contexto, o estudo da inibição do sistema
complemento pela saliva de mosquitos do gênero Anopheles é de grande
importância para um melhor entendimento da interação destes importantes
vetorescomohospedeirohumano.

31
3.Objetivos3.1ObjetivogeralDemonstrar e caracterizar a atividade inibidora do sistema complemento
humano presente na saliva de Anopheles albimanus, A. aquasalis, A. dirus, A.
freeborni,A.gambiaeeA.stephensi.
3.2Objetivosespecíficos
3.2.1 Avaliar se o extrato de glândulas salivares (EGS) deAnopheles spp. é
capazdeinibirasviasclássica,das lectinasealternativadosistemacomplemento
humano;
3.2.2AvaliaroefeitodoEGSsobrecomponentesespecíficosdaviaalternativa;
3.2.3 Identificar, clonar e expressar possíveis moléculas salivares com
atividadeinibidoradosistemacomplemento;
3.2.4Produziranticorpocontrapossíveisinibidoressalivareseavaliarsua
capacidadedebloquearaatividadeanti-complementodosmesmos;
3.2.5Investigaromecanismodeaçãodepossíveisinibidoressalivares.

32
4.MaterialeMétodos
4.1Aprovaçãoemcomitêdeética
Os experimentos realizados neste estudo foram conduzidos de acordo
comosprincípioséticosdeexperimentaçãoanimaldaUniversidadeFederalde
MinasGeraisedoInstitutoNacionaldeSaúdedosEstadosUnidos.Estetrabalho
foi aprovado pelo Comitê de Ética emExperimentaçãoAnimal (CETEA/UFMG)
sob número de protocolo número 087/11, e pelo Comitê de Uso e Cuidado
AnimaldoNIH,comIDdeaprovaçãoASP-LMVR3.
4.2Criaçãodosmosquitoseobtençãodeextratodeglândulassalivares(EGS)
OsmosquitospertencentesàsespéciesAnophelesalbimanus,A.gambiae,
A. dirus, A. stephensi e A. freeborni foram mantidos em colônias fechadas em
insetáriosnoLaboratóriodePesquisadeMaláriaeVetores(LMVR)doInstituto
Nacional de Saúde (NIH), Estados Unidos, sob a coordenação do Sr. Andre
Laughinhouse.Osinsetosadultoserammantidosemgaiolascilíndricascomtela
napartesuperior,ondechumaçosdealgodãoembebidosemsoluçãodexarope
Karo® 10% eram oferecidos aos mosquitos. Galinhas imobilizadas eram
colocadas sobre a tela das gaiolas para que as fêmeas realizassem o repasto
sanguíneo.As formas imaturas(ovos, larvasepupas)erammantidasemcubas
de plástico, contendo água da torneira. As condições climáticas dos insetários
erampreservadasa27oCe75%deumidade.
Os mosquitos das espécies Anopheles aquasalis e Aedes aegypti foram
criadosnoinsetáriodoLaboratóriodeMalária(LAMAL),noCentrodePesquisa
RenéRachou–CPqRR/FIOCRUZ,sobacoordenaçãodoDr.LucianoMoreira.Para
a criaçãodeA.aquasalis, gaiolas cilíndricasdepapelão (Barripel®) coma face
superior telada eram utilizadas para manter os insetos adultos. Bolinhas de
algodão com solução de açúcar 10% e camundongos anestesiados eram
utilizadosparasuaalimentação.Asformasimaturaserammantidasemcubasde
plástico contendo água do mar diluída 1:10. Para a criação de A. aegypti, os

33
adultos eram mantidos em gaiolas quadriculares de plástico com as laterais
teladas(BugDorm®)contendobolinhasdealgodãocomsoluçãodeaçúcar10%.
Asformasimaturaseramcriadasemcubascontendoáguafiltradaedesclorada.
Os insetários de criação das duas espécies eram mantidos a 27oC e
aproximadamente80%deumidade.
ParaobtençãodoEGS,fêmeasde4a8diasnãoalimentadascomsangue
foramdissecadasemlâminadevidroescavadacontendoPBSousoluçãosalina
0,9%, com auxílio demicroscópio estereoscópico e estiletes entomológicos. As
glândulassalivaresforamcoletadasetransferidasparatubosde1,5mlcontendo
solução específica para o teste a ser realizado emantidos constantemente em
gelo.Ostubosforamentãosonicadospor60segundosecentrifugadosa10.000g
por10minutosa4oC.Osobrenadanteerautilizadonosensaios.Aconcentração
de proteínas nos extratos de glândulas salivares foi medida pelo método de
Bradford (Bradford 1976) ou pelo kit BCA (Thermo Scientific®), ambos
baseados em uma curva padrão de albumina de soro bovino, seguindo as
orientaçõesdomanualdeinstruções.
4.3Ensaiohemolítico(BaseadoemMendes-Sousaetal.,2013)
Nos testes de hemólise mediada pela via alternativa do SC, foram
utilizadas hemácias de coelho coletadas por punção da veia auricular de um
animalmantidonobiotériodoInstitutodeCiênciasBiológicasdaUFMG(paraos
ensaios com EGS de A. aquasalis) ou obtidas comercialmente da empresa
CompTech® (para os ensaios comEGSdas demais espécies). As células foram
lavadastrêsvezescomsoluçãoMg-EGTA(HEPES1mM,NaCl30mM,EGTA10
mM,MgCl27mM,glicose3%,gelatina0,02%,pH7,4)atravésdecentrifugaçãoa
600gpor5minutosa4ºCseguidadedescartedosobrenadanteeressuspensão
dashemáciasem1mldeMg-EGTA.Antesdosexperimentos,aconcentraçãodas
célulasfoiajustadapara1x108células/ml
Para os ensaios com EGS deA. aquasalis, foi utilizado um poolde soro
humanoprovenientedacoletadesanguededoadoresvoluntáriossaudáveisno
Laboratório de Fisiologia de Insetos Hematófagos (Departamento de

34
Parasitologia,ICB,UFMG).Nosensaioscomasoutrasespécies,foiutilizadosoro
humano normal adquirido da CompTech®. Em ambos os casos, o soro foi
aliquotadoemantidoa-80oCatéarealizaçãodosexperimentos,sendoquecada
alíquotafoiutilizadaapenasumavez.
Emtubosde1,5ml,foramadicionados25μldesorohumanodiluído1:20
emMg-EGTAjuntamentecom12,5μldePBScontendoaconcentraçãodesejada
deEGS.Emseguida,foramadicionados25µldesoluçãodehemáciaseostubos
foramentãoincubadosa37ºCpor30minutosparaaativaçãodocomplemento.
Apósa incubação,250µldePBSgelado foramacrescentadosa cada tubopara
parar a reação de hemólise. Após centrifugação a 1700 g por 30 segundos,
transferiu-se200µldosobrenadanteparaumaplacade96poços,paraserlida
em leitor de ELISA (Bio-Rad Benchmark®) a 415nm. Exclusivamente com A.
albimanus, foram realizados ensaios com EGS (quantidade referente a 4
glândulas)aquecidoemáguaferventepor30minutosouincubadopor3horasa
37oCcom0,2μgdeproteinaseK.
Emcadateste,foramutilizadostrêscontroles:controlehemólisetotal,no
qual eram acrescidos 250 µl de água destilada fria após a incubação a 37oC;
hemólise espontânea (controle negativo), no qual não havia adição de soro,
apenas25μldeMg-EGTA;ehemólisepelocomplemento(controlepositivo),no
qual era acrescentado soro, mas não havia adição de EGS. Os testes foram
realizadosemduplicata,comnomínimotrêsrepetições.Paraanálisedosdados,
a média dos controles negativos era subtraída dos demais resultados e os
mesmos transformadosemporcentagemdehemólise, considerandoo controle
positivocomo100%deatividadehemolítica.
Nos ensaios com a via clássica, foram utilizadas hemácias de carneiro.
Para os experimentos com EGS de A. aquasalis, foi coletado sangue da veia
jugular de um carneiro adulto mantido no ICB. O sangue era imediatamente
misturadocomsoluçãoconservadora(ácidocítrico0,05%,citratodesódio0,8%,
glicose2,05%,NaCl0,42%)naproporção1:1earmazenadoa4oC.Parapreparo
dashemácias,1mldo sangue foi centrifugadoa600gpor5minutosa4oCeo
sobrenadante descartado. As células precipitadas foram ressuspendidas e
homogeneizadas em1ml de soluçãoGHB-EDTA (HEPES5mM,NaCl 145mM,

35
EDTA 10mM e gelatina 0,1%, pH 7,4) e centrifugadas novamente. Apósmais
duas lavagens semelhantes, as células foram ressuspendidas em1ml deGHB-
EDTAeopsonizadascomanticorpoIgGanti-hemáciadecarneiroproduzidoem
coelho(Sigma®)diluído1:1000durante incubaçãoa37ºCpor30minutossob
leve agitação. Após a sensibilização, as células foram lavadas uma vez com
soluçãoGHB-EDTAe,emseguida,duasvezescomsoluçãoGHB2+(HEPES5mM,
NaCl 145mM, CaCl2 0,15mM,MgCl2 0,5mM e gelatina 0,1%, pH 7,4). Após a
últimalavagem,aconcentraçãodashemáciasfoiajustadapara2x108células/ml
em solução GHB2+. Para os experimentos com EGS das outras espécies de
mosquitos foram utilizadas hemácias de carneiro opsonizadas com anticorpo
adquiridas da CompTech®. Essas células foram lavadas três vezes em solução
GVB2+(gelatina0.1%,veronal5mM,NaCl145mM,NaN30.025%,CaCl20.15mM,
MgCl2 0, 5mM, pH 7,3) (CompTech®), como descritas anteriormente e a
concentraçãoajustadapara2x108células/ml.Orestantedosexperimentosse
deucomodescritoparaaviaalternativa,utilizandosorohumanodiluído1:60em
solução GHB2+ (nos ensaios com A. aquasalis) ou GVB2+ (nos ensaios com as
outrasespécies).
4.4Ensaiodeativaçãodaviadaslectinas(BaseadoemBergströmetal.2009)
PoçosdeumaplacadeELISA(Costar®)foramsensibilizadosovernighta
4ºCcom50μldetampãocarbonatodesódio35mMebicarbonatodesódio15
mMpH9,6 contendo100μg/mldemananaobtidadeSaccharomycescerevisiae
(Sigma®)ouBSA1%(controlenegativo).Ospoçosforamentãoincubadoscom
200μlde soluçãodebloqueio (PBS+BSA1%)porumahoraem temperatura
ambientesobleveagitação.Emseguida,foiadicionadosorohumanonormal1%
(diluídoemtampãoGVB2+)juntamentecomdiferentesconcentraçõesdeEGSde
A. albimanus (volume final de 100 μl por poço) e a placa incubada por 30
minutosa37oC.
Após a incubação, os poços foram lavados duas vezes com 200 μl de
soluçãodelavagem(PBS-Tw0,05%)sobagitaçãopordoisminutoseemseguida
incubados com 50 μl de solução de bloqueio contendo anticorpo anti-C3

36
(CompTech®) 1:1000 por 60 minutos em temperatura ambiente sob leve
agitação.
Depoisdemaisduaslavagens,ospoçosforamincubadossobagitaçãopor
60 minutos em temperatura ambiente com 50 μl de solução de bloqueio
contendo anticorpo anti-cabra conjugado com peroxidase (Sigma®) diluído
1:1500. Em seguida, os poços foram lavados novamente e 200 μl de tampão
citratodesódio50mMefosfatodesódio50mMpH5,0contendoOPD(Sigma®)
1mg/ml e peróxido de hidrogênio 0,075% foram adicionados aos poços para
revelaçãodaplaca.Aleiturafoifeitaa450nmnomodocinéticopor10minutos
a37oCemleitordeELISA(Bio-RadBenchmark®),utilizandoosoftwareSoftMax
Pro5.2.
Poços sensibilizados comBSA1%epoços sensibilizados commananae
incubados apenas com soro (sem EGS) foram utilizados como controles
negativos e positivos, respectivamente. Os ensaios foram realizados em
duplicata,compelomenostrêsrepetições.Paraocálculoestatístico,foicalculada
amédiadecadaduplicataeamédiadocontrolenegativosubtraídadasdemais.
OsresultadosforamexpressoscomoporcentagemdeativaçãodeC3daviadas
lectinas,considerandoocontrolepositivocomo100%deativação.
4.5Ensaiodedeposiçãodecomponentesdaviaalternativaemplacascobertas
comagarose(BaseadoemMendes-Sousaetal.,2013)
Placasde96poços (Costar®) foramcobertas com100μlde soluçãode
agarose0,1%edeixadassecara37ºCovernightparaaformaçãodeumapelícula
de agarose no fundo do poço. Então foram adicionados 20 µl de soro humano
normaldiluído1:5emsoluçãoHMEBN(HEPES5mM,MgCl27mM,EGTA10mM,
BSA5mg/ml,NaCl140mM,pH7,4), juntamentecomEGSdeA.aquasalisouA.
albimanusem80µldeHMEBN,totalizandoassim100µldevolumenopoço,ea
placaincubadaa37oCpor30minutos.
Emseguida,ospoçosforamlavadosduasvezescom200µldesoluçãode
lavagem(Tris10mM,NaCl140mMeBSA0,1%)pordoisminutossobagitaçãoe
foram adicionados 50 µl de anticorpo anti-C3 (CompTech®), anti-Fator B

37
(CompTech®) ou anti-Properdina (CompTech®) diluídos 1:1000, 1:500 e
1:1000, respectivamente, em solução contendoHEPES 10mM eNaCl 140mM
(pH 7,4) e a placa incubada por 30 minutos em temperatura ambiente sob
agitação.Apósduas lavagens, os poços foram tratados com50µl de anticorpo
anti-cabra conjugado com peroxidase (Sigma®) diluído 1:1500 na mesma
soluçãoe incubadosnovamentepor30minutosem temperaturaambientesob
agitação.Orestantedoensaiosedeucomodescritoparaoensaiodeativaçãoda
viadaslectinas,utilizandopoçosincubadossemsoroepoçosincubadosapenas
com soro (sem EGS ou proteína recombinante) como controles negativos e
positivos,respectivamente.Osensaiosforamrealizadosemduplicatae,emcada
ensaio, amédia do controle negativo foi subtraída dasmédias das velocidades
máximasdas reaçõesdocontrolepositivoedospoçoscom inibidor salivar.Os
resultados foram então transformados em porcentagem de deposição de cada
componente,considerandoocontrolepositivocomo100%dedeposição.
Para investigar se os inibidores salivares deA.aquasalis eA.albimanus
eram capazes de desligar componentes da via alternativa previamente
depositados sobre superfície ativadora, realizamos ensaios de deposição como
descritosacima,mascomalgumasmodificações.Inicialmente,foramadicionados
100μldeSHNdiluídoemHMEBN(SHN7%nosensaioscomA.aquasalise4%
nos ensaios comA.albimanus) e aplaca incubadaa37°Cpor30minutospara
ativaçãodaviaalternativaedeposiçãodosseuscomponentesnofundodopoço.
Apósduaslavagenscom200μldomesmotampão,100μldeHMEBNcontendo
EGS relativo a 15 glândulasdeA.aquasalis ou concentração final de20nMde
proteína recombinante deA.albimanus foram adicionados aos poços e a placa
incubada novamente por 30 minutos a 37°C. Poços incubados apenas com
tampãoforamutilizadoscomocontrolenegativo.Nosensaioscomoinibidorde
A. albimanus, 100 μg de Fator H purificado (CompTech) também foi utilizado
como controle para o desligamento de Fator B. Após mais duas lavagens,
anticorpoanti-FatorB,anti-C3ouanti-properdina foiadicionadoaospoçoseo
restantedoensaioocorreucomodescritoparaosensaiosdedeposição.

38
4.6WesternblotparadetecçãodeFatorBeC3a(BaseadoemLawrieetal.,2005)
Ensaiosdehemólisepelaviaalternativacomodescritosnoitem4.3foram
realizados utilizando apenas quantidade de EGS referente a 4 ou 15 glândulas
salivares(nosensaioscomA.albimanusouA.aquasalis,respectivamente)esoro
humanonormaldiluído1:35emMg-EGTA.Tubos comadiçãodePBS semEGS
foramutilizadoscomocontrolespositivos.Ostubosforamincubadosa37oCeem
diferentestemposdeincubação(tempos0’e30’,nosensaioscomA.aquasalis;e
tempos0’,30’e60’,nosensaioscomA.albimanus),foramcentrifugadosa1700g
por 30 segundos, e uma alíquota de 5 μl do sobrenadante foi coletado e
misturadocomtampãodaamostra4vezesconcentrado(LifeTechnologies®)e
agente redutor 10 vezes concentrado (Life Technologies®). Os tubos foram
então aquecidos a 90oC por 5 minutos e seu conteúdo aplicado em gel de
poliacrilamida 4-10%. Em cada gel, foi utilizado 10 ng de Bb ou 5 ng de C3a
purificados(CompTech®),umpoçoapenascomsorodiluído1:35nãoincubado
a 37°C e umpadrão de bandas pré-corado (SeeBlue Pre stained, Invitrogen®)
que,juntamentecomasamostras,correramporaproximadamente30minutosa
200V.
Após a corrida, as proteínas do gel foram transferidas para uma
membranadenitroceluloseduranteumahoraa30Ve,logoapósatransferência,
amembranafoibloqueadaovernightcomsoluçãobloqueadora(PBS-Tw0,05%+
leite em pó desnatado 10%) em constante agitação. Seguindo o bloqueio, a
membrana foi lavada três vezes por cinco minutos com PBS-Tw 0,05% e
incubada por uma hora em temperatura ambiente sob agitação com anticorpo
anti-FatorBouanti-C3a(CompTech®),ambosdiluídos1:1000emPBS-Tw+1%
BSA. Após mais três lavagens, a membrana foi incubada por uma hora com
anticorpoanti-cabra(nosensaiosparadetecçãodeFatorB)diluído1:2000em
PBS-Tw + 1% BSA ou anticorpo anti-coelho (ensaios para detecção de C3a)
diluído 1:3000 na mesma solução. Ambos anticorpos secundários eram
conjugados com peroxidase. Após a incubação, as membranas foram lavadas
maistrêsvezesearevelaçãodasbandasdeFatorBeC3asedeuutilizandookit

39
Peroxidase Substrate DAB (Vector Laboratories®), seguindo as instruções do
fabricante.
Para investigar a ação da saliva de A. albimanus na formação da C3
convertase da via alternativa, 200 ng de C3b purificado, 0,5 ng de Fator D
purificadoe0,5μgdeFatorBpurificado (CompTech®)em20μl deMg-EGTA
foram incubadosa37oC juntamente com20μldomesmo tampãocomou sem
EGSreferentea4glândulas.Nostempos0’,20’e40’deincubação,umaalíquota
dostubosfoicoletadaemisturadacomtampãodaamostraeagenteredutor.O
material foi então aquecido a 95oC por cinco minutos e corrido em gel de
poliacrilamida4-10%.
AativaçãodocomponenteFatorB (surgimentodasbandas referentesa
BbeBa)foiavaliadaatravésdeWesternblotrealizadocomodescritoacima.
4.7PurificaçãoeidentificaçãodoinibidorsalivardeA.albimanus
ParapurificaroinibidordocomplementodeA.albimanus,200glândulas
salivaresdefêmeasentre4e8diasdevidanãoalimentadascomsangueforam
dissecadasearmazenadasem100μldePBS.Omaterialfoisonicadopor30a60
segundos, centrifugado a 10.000 g por 10 minutos a 4oC e o sobrenadante
coletado. Uma alíquota do material coletado foi testada em ensaio hemolítico
paraconfirmarapresençadoinibidor.Orestantefoisubmetidoacromatografia
líquidadealtaeficiência(HPLC)usandoumacolunadefasereversa(Source15
RPC 3ml, GE Healthcare®) conectada em sistema ÄKTA Purifier (GE
Healthcare®).Acoluna foiequilibradacomtampãoTBS (50mMTris,150mM
NaCl) pH 7,4 (Tampão A) num fluxo de 1 ml/min e nossa amostra foi então
aplicada.Asproteínasforameluídasdeacordocomsuahidrofobicidade,usando
gradiente (0 a 100%) de acetonitrila contendo 0,1% de ácido trifluoroacético
(Tampão B) num fluxo de 1 ml/min e coletadas em frações de 250 μL. A
concentraçãodeproteínasfoiavaliadaa220nm,utilizandoosoftwareUnicorn
5.0(GEHealthcare®).
As frações coletadas foram centrifugadas em SpeedVac-SC 110 (Savant
Instruments®)a45oCpor3horasparacompletaevaporaçãodotampãoBe,em

40
seguida, resolubilizadas em 100 μl de PBS. Uma alíquota de 12,5 μl de cada
fração foi testada em ensaio hemolítico como descrito anteriormente para
identificar atividade inibidora da via alternativa. As frações que apresentaram
atividadeeassuasadjacentes, tiveramumaalíquotacoletadaemisturadacom
tampão da amostra e agente redutor, aquecidas a 95oC por cinco minutos,
corridas em gel de poliacrilamida (4-10%) e coradas com prata usando o kit
SilverQuest(Invitrogen®),seguindooseumanualdeinstruções.
Para identificação do inibidor, inicialmente a fração 1H12 inteira foi
enviada para a plataforma de espectrometria demassa noNIH. Omaterial foi
submetido a digestão por tripsina e os peptídeos gerados submetidos a
cromatografia líquida em nano-escala associada a espectrometria de massa
simultânea(nanoLC-MS/MS).Asproteínasencontradasnafraçãoenviadaforam
identificadas a partir de comparações com os bancos de dados disponíveis
(genoma e transcriptoma) no site do NCBI (http://www.ncbi.nlm.nih.gov) ou
VectorBase (www.vectorbase.org). Em um segundo momento, apenas uma
bandaentre6e14kDaqueapareciaexclusivamentenasfraçõescomatividade
foisubmetidaàanálisedeespectrometriademassa.
Aproteínaidentificadanosresultadosdaespectrometriacomooinibidor
salivar de A. albimanus foi encontrada no banco de dados do VectorBase e a
similaridade com proteínas de outras espécies de anofelinos foi investigada
atravésdeBLASTnomesmositeounoNCBI.Paraaverificaçãodapresençade
peptídeosinaledepossíveisdomíniosconhecidosdaproteína,foramutilizados
os softwares SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP) e
Pfam (http://pfam.xfam.org/), respectivamente. Para o alinhamento das
sequências de proteínas e desenho da árvore filogenética foram utilizados o
UniProt (http://www.uniprot.org/align/) e o PhyML (Seaview®),
respectivamente. Para investigar demais características dos genes e das
proteínas,foiutilizadoosoftwareDNAstarLasergene®.

41
4.8 RT-PCR para detecção do gene da proteína gSG7 em extrato de glândulas
salivaresdeA.albimanus
ParaconfirmarapresençadogenereferenteàproteínagSG7noextrato
de glândulas salivares de A. albimanus, foram sintetizados primers para a
sequência deste gene: albigsg7FOR1 (5’ GACAGTCATATTGCCGTTGG) e
albigsg7REV3 (5’ AACATGCGCTTTGCATACAG). Esses primers originam um
ampliconde477pb.
Inicialmente, foram dissecadas 50 glândulas salivares de fêmeas de A.
albimanus com 5 dias de vida não alimentadas com sangue. O material foi
dissecado e transferido imediatamente para 50 μl de Trizol® Reagent (Life
Technologies®). Após homogeneização do material com pistilo estéril, foi
realizadaextraçãodeRNAtotalseguindooprotocolodoTrizol®.Otubocomo
RNAextraído foi secado totalmente emSpeedVac e, em seguida, omaterial foi
dissolvidoem20μldeáguaultrapura(UltraPureDNAse/RNAseFreeDistilled
Water,LifeTechnologies®).
Utilizando o kit QuantiTect® Reverse Transcription kit (QIAGEN®),
produziu-se o DNA complementar (cDNA) a partir de 5 μl de RNA extraído,
seguindoomanualdeinstruçõesdokit,comumaalteração:aoinvésdeutilizaro
RTPrimerMixdokit,utilizamosoprimeralbigsg7REV3paraqueapenasoDNA
complementardanossasequênciafossegerado.
SeguindoomanualdeinstruçõesdoKitFastStartPCRMaster(Roche®),
comosprimersdescritosacimaeocDNAproduzidocomomolde, foi realizada
PCRparaverificar apresençadogenede interesse.Tubos comáguaultrapura
comomolde, sem cDNA, foramutilizados como controle negativo. Os ciclos da
reaçãosãoapresentadosnoQuadro1.
Nove microlitros dos produtos da PCR foram misturados com 1 μl de
tampãodaamostra10x (Invitrogen®),aplicadosemgeldeagarose1,2%com
SYBRSafestain(Invitrogen®)ecorridospor30minutosa100V.Comopadrão
depesomolecular,foiutilizadooGeneRuler1kb(ThermaScientific®).Apósa
corrida,ogelfoivisualizadoefotografadoemaparelhotransiluminadorcomluz
UV.

42
Quadro1:CiclosdetemperaturautilizadosnasreaçõesdePCRparaamplificaçãodogenedagSG7deA.albimanus
Ciclos Temperatura(ºC) Tempo
Desnaturação 1 95 4min
DesnaturaçãoAnelamentoAlongamento
30 955972
30seg30seg2min
Extensãofinal 1 72 7min
4.9 Clonagem, expressão e purificação das proteínas gSG7 e gSG7-2 de A.
albimanusegSG7deA.darlingi(BaseadoemAlvarengaetal.,2010)
GenessintéticosotimizadosdasproteínasgSG7egSG7-2deA.albimanus
e gSG7deA.darlingi sem a sequência do peptídeo sinal (Figura 6) ligados em
vetor Pet17b foramproduzidos (BioBasic®) e expressos emE. coli BL21 pLys
(NewEnglandBioLabs®).
SequênciadogenesintéticodagSG7deA.albimanus:
SequênciadogenesintéticodagSG7-2deA.albimanus:

43
SequênciadogenesintéticodagSG7deA.darlingi:
Figura 6: Sequências dos genes sintéticos utilizados para clonagem eexpressãodeproteínasrecombinantesdeA.albimanuseA.darlingi.
Asbactériasforamtransformadasporchoquetérmico(mantidasemgelo
por15minutos, transferidasparabanho-mariaa42ºCpor30segundose, logo
em seguida,mantidas emgelo novamente), transferidas para placa de ágar LB
contendoampicilina100μg/ml(KDMedical®)eincubadasovernighta37oC.As
colônias crescidas foram transferidas para um frasco com 50 ml de meio LB
Broth (KD Medical®) contendo 50 μl de ampicilina 100 mg/ml e 50 μl de
cloranfenicol35mg/mleincubadasa37oCovernight.
Quatrogarrafascom1litrodemeioLBcontendoampicilina100μg/mle
cloranfenicol 35 μg/ml foram semeadas com 10 ml da cultura crescida e
incubadasatéaconcentraçãodebactériasatingirA600de0,6a0,8.Emseguidaas
culturas foram induzidas com 1ml de IPTG 1M (Invitrogen®) e incubadas a
37oCpor3horas.Apósaincubação,asculturasforamcentrifugadasa7000rpm
por 15 minutos a 10oC e o sobrenadante descartado. O pellet formado foi
resolubilizado em 150 ml de TBS e centrifugado da mesma forma. O
sobrenadantefoidescartado,eopelletformadocongeladoa-20oCovernight.
Apósdescongelar,opelletfoiresolubilizadoem150mldeTBS,sonicado
em sonicador de ponta por aproximadamente 1minuto e centrifugado por 15
minutos a 7000 rpm a 10oC. O sobrenadante foi descartado e o pellet
resolubilizado, sonicado e centrifugado novamente. O pellet formado foi então
resolubilizadoem100mldeTBScontendo1gdeTritonX-100(Sigma-Aldrich®)
e agitado combarramagnética por 30minutos em temperatura ambiente. Em
seguida, o material foi centrifugado como anteriormente, o sobrenadante
descartadoeopelletresolubilizadoem100mldeTBS.Estalavagemfoirepetida

44
maisduasvezeseapósaultimacentrifugação,opelletfoiredissolvidoem50ml
detampãoTris20mMpH8,0contendohidrocloretodeguanidina(Invitrogen®)
6Meincubadopor1horaemtemperaturaambientesobagitaçãoconstante.Em
seguida, foi adicionado ditiotreitol (DTT) (Sigma-Aldrich®) na concentração
finalde10mMeo frasco incubadopor30minutossobagitaçãoconstanteem
temperatura ambiente. Após esse período, o material desnaturado pela
guanidina e pelo DTT foi adicionado gota a gota a 4 litros de tampão de
renovelamento(Trizma20mM,arginina300mM,dihidrocloretodecistamina2
mM,pH9,2)sempreemagitaçãocombarramagnéticae,emseguida, incubado
overnighta4oC.Asproteínasrenoveladasforamconcentradasusandoaparatode
concentração Millipore® pressionado por nitrogênio e filtro de 10 kDa
(MilliporeUltracell®)atéatingirovolumefinaldeaproximadamente10ml.
Parapurificaçãodasproteínas, omaterial foi aplicadoemcolunade gel
filtração HiPrep 16/60 Sephacryl S-100 HR (GE Healthcare®) conectada em
aparelhodeHPLC(ÄKTAPurifier,GEHealthcare®)easproteínascoletadasem
fraçõesde4mldeTBSpH7,4.UmaalíquotadecadafraçãofoicorridaemSDS-
PAGE, corado comComassie blue. As frações contendo a proteína de interesse
forammisturadaseconcentradasparaaproximadamente10mlusandofiltrode
10 kDa (Amicon®). As proteínas concentradas foram novamente purificadas
usando coluna de gel filtração Superdex 75 10/300 GL (GE Healthcare®) e a
purezadaproteínaavaliadaporSDS-PAGEcoradocomComassieblue.Asfrações
contendo a proteína de interesse foram misturadas, concentradas e a
concentraçãofinalcalculadapelocoeficientedeextinçãomolardecadaproteína.
Asproteínaspurificadasforamtestadasnosensaioshemolíticosparavia
alternativaeviaclássica,ensaiosdedeposiçãodecomponentesdocomplemento
(C3, Fator B, properdina e C9) e Western blots, como descritos previamente,
substituindooEGSpelasproteínasrecombinantes.
4.10IdentificaçãodoinibidordocomplementodasalivadeA.aquasalis
Em gel de poliacrilamida 12,5% foram corridos 0,5 μg de gSG7
recombinantedeA.albimanus,0,5μgdegSG7recombinantedeA.darlingie5μg

45
deEGSdeA.aquasaliseogelcoradocomprataparavisualizaçãodasproteínas
presentes.
Paraaidentificaçãodopossível inibidorsalivardeA.aquasalis,umnovo
gelfoicorridocom20μgdeEGSdessaespéciee10μgdegSG7recombinantede
A.darlingiecoradocomComassieblue.Aproteínarecombinanteeumaproteína
do EGS deA. aquasalis com peso similar foram cortadas do gel com ajuda de
bisturi,transferidasparatubosde1,5mletratadascombicarbonatodeamônio,
acetonitrila e iodoacetamina para descorar a banda e desidratar o gel. Em
seguida,asproteínasforamsubmetidasadigestãoportripsinaa37°Covernight.
Os peptídeos gerados pela tripsinização foram então sequenciados em
aparelho MALDI-TOF-TOF (Bruker®) mantido no Instituto de Ciências
BiológicasdaUFMG,utilizandoosoftwareMASCOT.Assequênciasdospeptídeos
gerados das duas amostras foram comparados com a sequência da gSG7 deA.
darlingi,utilizandoosoftwareUniProt(http://www.uniprot.org/align/).
4.11Westernbloteensaiohemolíticocomanticorpoanti-gSG7deA.albimanus
Anticorpos anti-gSG7 foram produzidos pela Spring Valley Laboratories
(Woodbine,Maryland,EUA),emcoelhoimunizadocomaproteínarecombinante
juntamente com adjuvante de Freunds. Após três imunizações, o animal foi
sangradoeseusorofoicoletadoeenviadoaoLaboratóriodePesquisadeMalária
eVetores.
Para purificação de IgG do soro, foi utilizado coluna de proteína A
(HiTrapTM 5 ml Protein A HP, GE Healthcare®), seguindo o seu manual de
instruções. O IgG purificado foi então testado por Western blot para ver o
reconhecimentodaproteínarecombinanteedaproteínapresentenoEGS.Para
isso, quantidade de EGS relativo a 20 glândulas salivares de fêmeas de A.
albimanus e 0,2 μg da proteína recombinante gSG7 foram corridos em gel de
poliacrilamida(4-12%)a200Vpor30minutos.Antesdeaplicadosnogel,oEGS
ouarecombinanteforammisturadoscomtampãodaamostraeagenteredutore
aquecidaspor5minutosa95ºC.

46
Apósacorrida,asproteínasdogelforamtransferidasparamembranade
nitrocelulose,quefoibloqueadaporduashorascomsoluçãobloqueadora(PBS-
Tw 0,05% + leite em pó desnatado 10%) em agitação constante. Seguindo o
bloqueio, a membrana foi lavada três vezes por cinco minutos com PBS-Tw
0,05% e incubada por uma hora em temperatura ambiente com IgG anti-gSG7
diluído 1:10000 em PBS + 1% BSA. Apósmais três lavagens, amembrana foi
incubada por uma hora com anticorpo anti-coelho conjugado com peroxidase
diluído 1:5000 na mesma solução. Após a incubação, as membranas foram
lavadasmaistrêsvezesearevelaçãosedeuutilizandookitPeroxidaseSubstrate
DAB(VectorLaboratories®).
Para verificar se o IgG anti-gSG7 de A. albimanus reconheceria outras
proteínassalivaresdeoutrasespéciesdemosquitos,foramaplicadosecorridos
emgeldepoliacrilamida12,5%,0,5μgdegSG7deA.albimanus,0,5μgdegSG7-2
de A. albimanus, 0,5 μg de gSG7 de A. darlingi, 5 μg de EGS de fêmeas de A.
aquasalise5μgdeEGSdefêmeasdeAedesaegypti.OWesternblotfoirealizado
como descrito acima, utilizando anticorpos primário e secundário diluídos
1:2000.
ParaavaliaracapacidadedoanticorpodebloquearaatividadedagSG7de
A.albimanus,ensaioshemolíticosforamrealizadosnapresençadoIgGanti-gSG7.
Emumtubode1,5ml,foramadicionados12,5μldePBScontendoEGSreferente
a1glândulasalivarougSG7naconcentraçãode30nM(concentraçãofinalde5
nM no tubo) e, em seguida, misturados com 12,5 μl de Mg-EGTA contendo
diferentesdiluiçõesdoIgGanti-gSG7.Apósadiçãode25μldesorodiluído1:20e
25μldehemáciasdecoelhosnaconcentraçãode1x108céls/ml,oconteúdodos
tubos foi homogeneizado e incubado a 37ºC por 30 minutos. Um controle foi
adicionado aos experimentos: um tubo com apenas hemácias e IgG anti-gSG7
diluído1:10(concentraçãomaisaltatestada),paraaveriguarseoanticorpopor
si só afetaria as hemácias. O restante do ensaio se deu como descrito
previamentenoItem4.3.

47
4.12 ELISA para detectar ligação de componentes do complemento aos
inibidoressalivaresdeA.aquasaliseA.albimanus
Placas de ELISA (Costar®) foram sensibilizadas com0,5 μg de gSG7 ou
gSG7-2 de A. albimanus ou 5 μg de EGS de A. aquasalis em 50 μl de tampão
carbonato de sódio 35 mM/bicarbonato de sódio 15 mM pH 9,6 durante
incubaçãoovernighta4oC.PoçosincubadoscomBSA1%foramutilizadoscomo
controlesnegativos.
Os poços foram bloqueados por uma hora com 200 μl de solução de
bloqueio(PBS+BSA1%)emtemperaturaambientesobagitaçãoe,emseguida,
incubadoscom0,2μgdecadacomponentedocomplementoasertestado(Fator
B,Bb,C3,C3b,FatorDeproperdina)em50μldePBSemtemperaturaambiente
por30minutos.Ospoçosforamentãolavadosduasvezescom200μldesolução
delavagem(PBS-Tw0,05%)por2minutossobagitaçãoeincubadoscom50μl
de solução de bloqueio contendo anticorpo específico para cada componente
testado(anti-C3foiutilizadoparadetectarC3eC3b;anti-FatorB,paradetectar
Fator B e Bb) diluído 1:5000 por 30 minutos em temperatura ambiente sob
agitação. Após mais duas lavagens, os poços foram tratados com anticorpo
secundário conjugado com peroxidase diluído 1:3000 em solução de bloqueio
por 30 minutos em temperatura ambiente e agitação constante. Seguindo a
incubação,maisduaslavagensforamrealizadaseospoçosforamreveladoscom
200μldesubstratoparaperoxidase(OPDeperóxidodehidrogênioemtampão
citratopH5,0,comodescritonosensaiosdeELISAnoItem4.5)eapós5minutos
a37oC,asplacasforamlidasa450nmemleitordeELISA.
Os ensaios foram realizados em duplicata, com 3 repetições. Asmédias
das absorvâncias das duplicatas de cada componente foram calculadas e
comparadas estatisticamente comamédiado seu respectivo controlenegativo
(poçosensibilizadocomBSA).

48
4.13Ensaioderessonânciaplasmônicadesuperfície
Ensaiosderessonânciaplasmônicadesuperfícieforamrealizadosusando
aparelho Biacore T100 (GE Healthcare®). As proteínas recombinantes gSG7 e
gSG7-2deA.albimanusdiluídasemtampãoacetatodesódio10mMpH5,0(20
μg/ml) foram imobilizadas em chips Sensor CM5 (GE Healthcare®) usando
grupamentoamina,comalvode imobilizaçãode1200unidadesderessonância
(RU). Células do chip sem proteína imobilizada foram utilizadas como branco
para subtração do efeito do tampão no sensorgrama. Para avaliar a ligação de
properdina às proteínas, experimentos de cinética foram realizados com
diferentesconcentraçõesdeproperdinaemtampãoHBS-P(HEPES0,01M,NaCl
0,15 M, Surfactante P20 0.005% v/v, pH 7,4, GE Healthcare®). O tempo de
contato das moléculas foi de 120 segundos em fluxo de 30 μl/min a 25ºC. A
dissociaçãodocomplexoproperdina-gSG7foimonitoradapor400segundosea
superfície do chip foi regenerada com um pulso de 10 segundos de tampão
glicina10mM.
Os sensorgramas foram montados usando o modelo 1:1 de ligação de
Langmuir das fases de associação e dissociação simultaneamente, usando o
softwareBiacoreEvaluationv2.0.3 (GEHealthcare®).Aconstantedeafinidade
(KD) foi calculada a partir da razão das constantes das taxas de dissociação e
associação(kd/ka).
4.14Análiseestatística
Os cálculos estatísticos foram realizados com o auxílio do software
GraphPad Prism 5.0. Em todos os experimentos, pelo menos três repetições
independentes foram feitas. A normalidade dos dados foi avaliada pelo teste
Kolmogorov-Smirnov. As variáveis com distribuição normal foram analisadas
atravésdotesteTpareado(nocasode2grupos)oupelostestesANOVAeTeste
de Tukey (no caso de mais de 2 grupos). Valor de p < 0,05 foi considerado
estatisticamentesignificativo(*).

49
5.Resultados
5.1QuantificaçãodeproteínasdosEGSdasespéciesdeAnophelesestudadas
A concentração de proteínas no extrato de glândulas salivares dos
anofelinos em estudo foi medida por teste de Bradford ou pelo kit BCA. As
médias e desvios padrões das concentrações de proteínas totais no EGS deA.
albimanus, A. aquasalis, A. dirus, A freeborni, A. gambiae e A. stephensi foram,
respectivamente, 0,75 ± 0,02 μg/glândula, 0,75 ± 0,05 μg/glândula, 0,59 ± 0,1
μg/glândula, 0,83 ± 0,04 μg/glândula, 0,56 ± 0,04 μg/glândula, e 0,71 ± 0,08
μg/glândula.
5.2Ensaioshemolíticos
5.2.1Viaalternativa
ConformedemonstradonaFigura7,oEGSdeA.albimanus,A.aquasalise
A. freeborni foi capaz de inibir a atividade hemolítica provocada pela via
alternativa de forma dose-dependente e estatisticamente significativa.
Curiosamente, o EGS de A. gambiae, A. dirus e A. stephensi não apresentou
nenhumaatividadesobreahemóliseprovocadapelocomplemento,mesmocom
quantidadedeEGSreferentea15glândulassalivares.
Quando tratado com aquecimento por fervura ou proteinase K, o EGS
referente a 4 glândulas de A. albimanus perdeu a atividade inibidora da via
alternativa(Figura8).
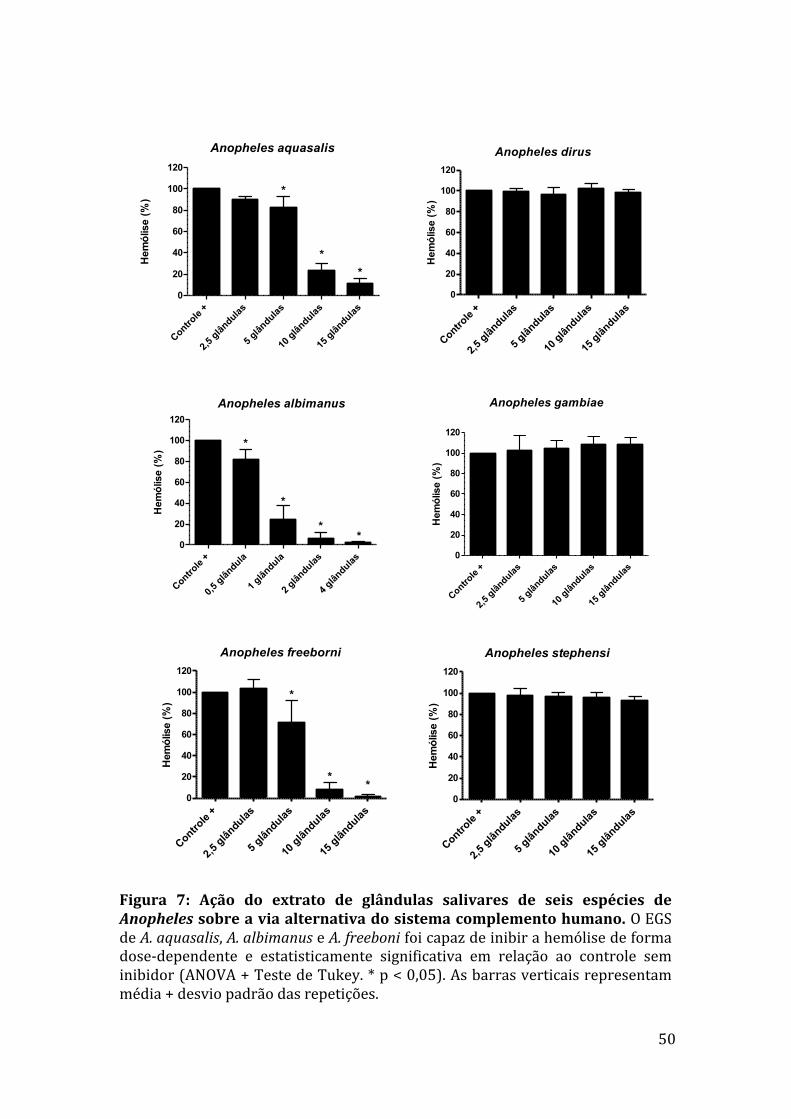
50
Anopheles aquasalis
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
**
*
**
Hem
ólis
e (%
)
Controle
+
0,5 glân
dula
1 glân
dula
2 glân
dulas
4 glân
dulas0
20
40
60
80
100
120
**
*
*
Anopheles albimanus
Hem
ólis
e (%
)
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
Anopheles freeborni
**
*
* *
Hem
ólis
e (%
)
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
Anopheles stephensi
Hem
ólis
e (%
)
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
Anopheles dirus
Hem
ólis
e (%
)
Anopheles gambiae
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120H
emól
ise
(%)
Figura 7: Ação do extrato de glândulas salivares de seis espécies deAnophelessobreaviaalternativadosistemacomplementohumano.OEGSdeA.aquasalis,A.albimanuseA.freebonifoicapazdeinibirahemólisedeformadose-dependente e estatisticamente significativa em relação ao controle seminibidor(ANOVA+TestedeTukey.*p<0,05).Asbarrasverticaisrepresentammédia+desviopadrãodasrepetições.

51
Figura8:EfeitodetratamentosdesnaturantessobreaatividadeinibidoradaviaalternativadoEGSdeA.albimanus.QuandotratadocomaquecimentoemáguaferventeoucomdigestãoporproteinaseK,oEGSperdeusuaatividade(ANOVA + Teste de Tukey). As barras verticais representam média + desviopadrãodasrepetições.
5.2.2Viaclássica
O EGS deA.gambiae eA.albimanus não foi capaz de inibir a atividade
hemolíticaprovocadapelaviaclássicadosistemacomplementohumano(Figura
9).QuandosalivadeA.aquasalisfoitestada,observamosumaleveinibiçãocom
diferençaestatísticasignificativaapenasquandoquantidadedesalivareferentea
10glândulasfoiutilizada(p=0.03402)(Figura9).
Controle
+
4 glân
dulas
4 glân
dulas aq
uecidas
4 glân
dulas +
protei
nase K
0
20
40
60
80
100
120
Hem
ólis
e (%
)
Anopheles albimanus
p < 0,001*
p < 0,001*
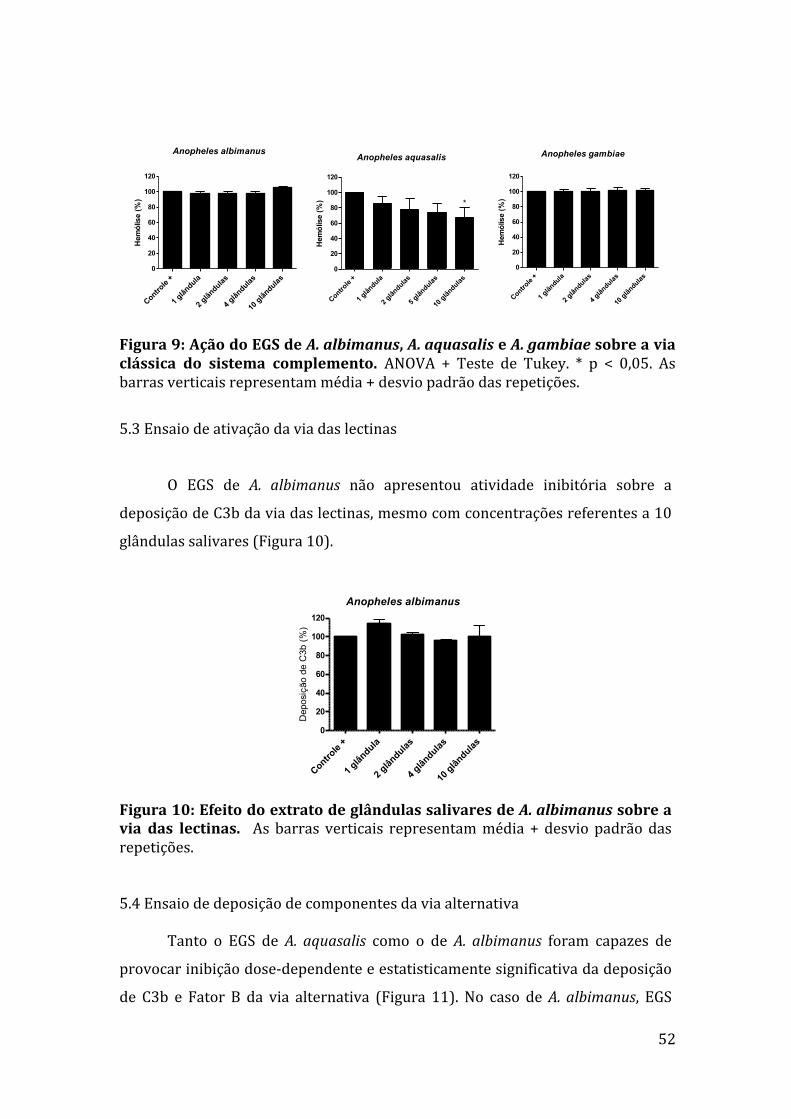
52
Anopheles albimanus
Controle
+
1 glân
dula
2 glân
dulas
4 glân
dulas
10 glân
dulas0
20
40
60
80
100
120
Hem
ólis
e (%
)
Anopheles aquasalis
Controle
+
1 glân
dula
2 glân
dulas
5 glân
dulas
10 glân
dulas0
20
40
60
80
100
120
*
Hem
ólis
e (%
)
Anopheles gambiae
Controle
+
1 glân
dula
2 glân
dulas
4 glân
dulas
10 glân
dulas0
20
40
60
80
100
120
Hem
ólis
e (%
)
Figura9:AçãodoEGSdeA.albimanus,A.aquasaliseA.gambiaesobreaviaclássica do sistema complemento. ANOVA + Teste de Tukey. * p < 0,05. Asbarrasverticaisrepresentammédia+desviopadrãodasrepetições.
5.3Ensaiodeativaçãodaviadaslectinas
O EGS de A. albimanus não apresentou atividade inibitória sobre a
deposiçãodeC3bdaviadaslectinas,mesmocomconcentraçõesreferentesa10
glândulassalivares(Figura10).
Controle
+
1 glân
dula
2 glân
dulas
4 glân
dulas
10 glân
dulas0
20
40
60
80
100
120
Anopheles albimanus
Dep
osiç
ão d
e C
3b (%
)
Figura10:EfeitodoextratodeglândulassalivaresdeA.albimanussobreavia das lectinas. Asbarras verticais representammédia+desviopadrãodasrepetições.5.4Ensaiodedeposiçãodecomponentesdaviaalternativa Tanto o EGS de A. aquasalis como o de A. albimanus foram capazes de
provocarinibiçãodose-dependenteeestatisticamentesignificativadadeposição
de C3b e Fator B da via alternativa (Figura 11). No caso deA.albimanus, EGS
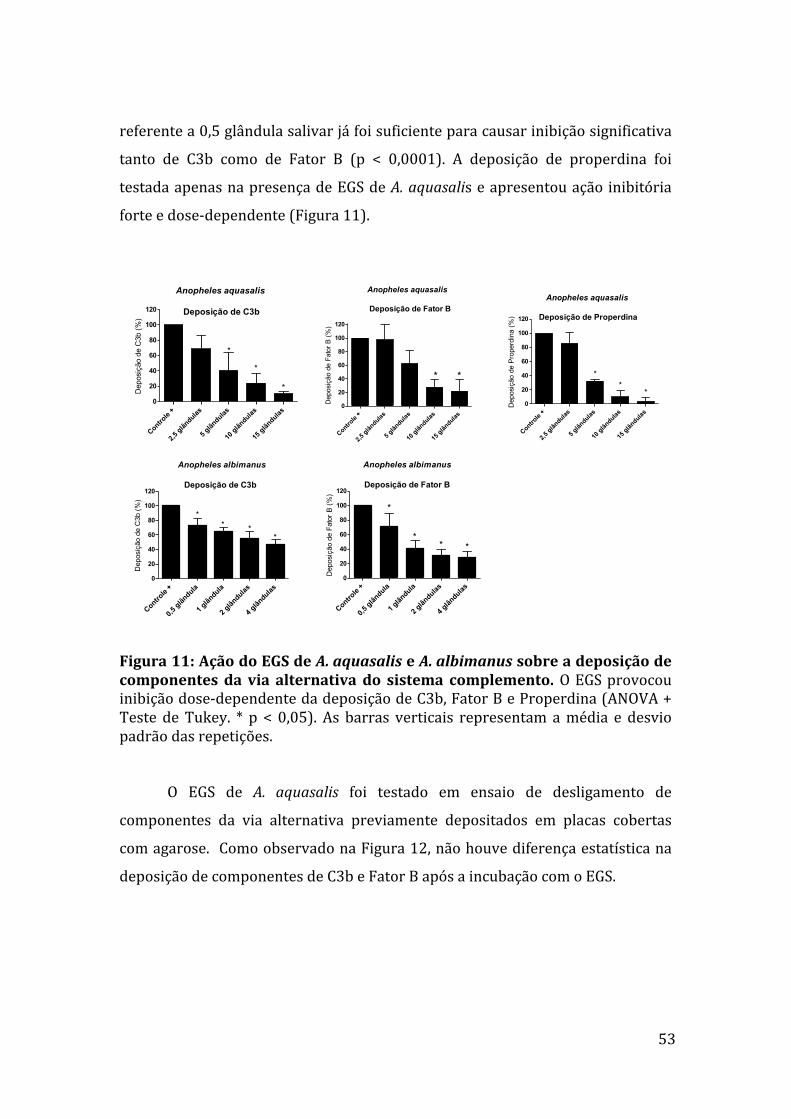
53
referentea0,5glândulasalivarjáfoisuficienteparacausarinibiçãosignificativa
tanto de C3b como de Fator B (p < 0,0001). A deposição de properdina foi
testadaapenasnapresençadeEGSdeA.aquasaliseapresentouaçãoinibitória
forteedose-dependente(Figura11).
Anopheles aquasalis
Deposição de C3b
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
*
*
*
Dep
osiç
ão d
e C
3b (%
)
Anopheles aquasalis
Deposição de Properdina
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
**
*
Dep
osiç
ão d
e Pr
oper
dina
(%)
Controle
+
2,5 glân
dulas
5 glân
dulas
10 glân
dulas
15 glân
dulas0
20
40
60
80
100
120
**
Anopheles aquasalis
Deposição de Fator B
Dep
osiç
ão d
e Fa
tor B
(%)
Controle
+
0,5 glân
dula
1 glân
dula
2 glân
dulas
4 glân
dulas0
20
40
60
80
100
120
**
*
*
Anopheles albimanus
Deposição de C3b
Dep
osiç
ão d
e C
3b (%
)
Controle
+
0,5 glân
dula
1 glân
dula
2 glân
dulas
4 glân
dulas0
20
40
60
80
100
120
Anopheles albimanus
Deposição de Fator B
*
*
**
Dep
osiç
ão d
e Fa
tor B
(%)
Figura11:AçãodoEGSdeA.aquasaliseA.albimanussobreadeposiçãodecomponentesda via alternativado sistema complemento.OEGSprovocouinibiçãodose-dependentedadeposiçãodeC3b,FatorBeProperdina(ANOVA+Teste deTukey. * p < 0,05). As barras verticais representamamédia e desviopadrãodasrepetições. O EGS de A. aquasalis foi testado em ensaio de desligamento de
componentes da via alternativa previamente depositados em placas cobertas
comagarose. ComoobservadonaFigura12,nãohouvediferençaestatísticana
deposiçãodecomponentesdeC3beFatorBapósaincubaçãocomoEGS.
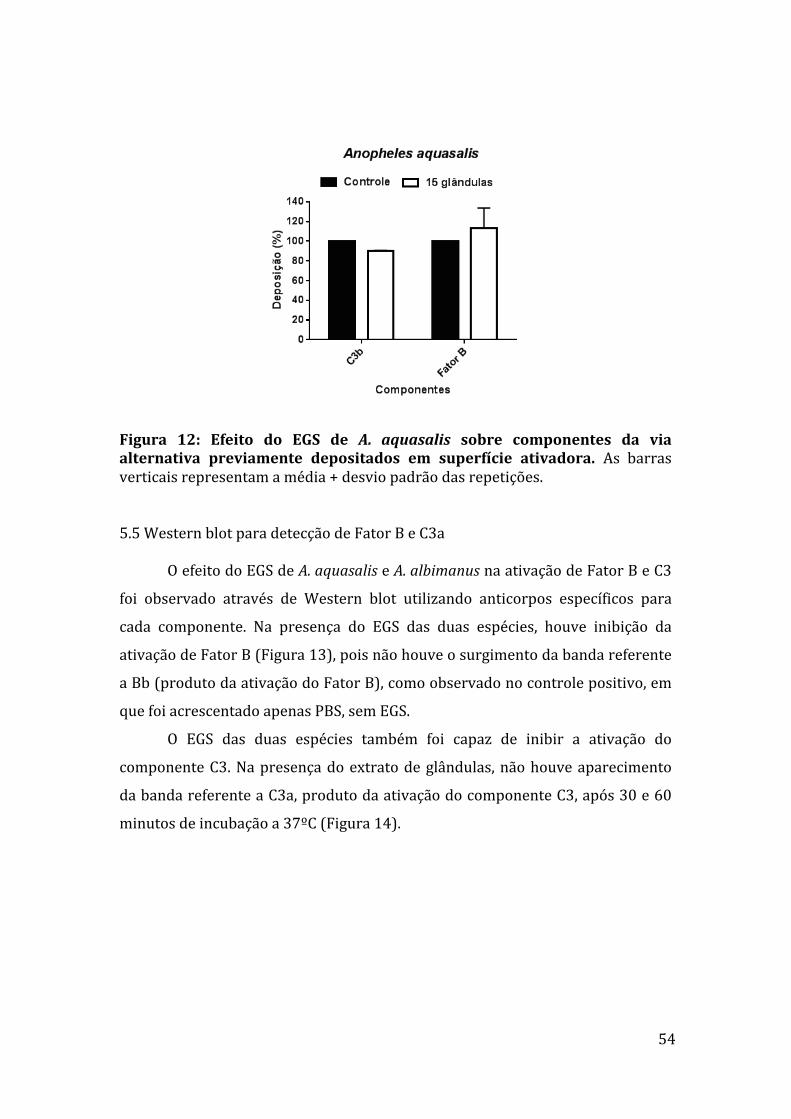
54
Figura 12: Efeito do EGS de A. aquasalis sobre componentes da viaalternativa previamente depositados em superfície ativadora. As barrasverticaisrepresentamamédia+desviopadrãodasrepetições.5.5WesternblotparadetecçãodeFatorBeC3a OefeitodoEGSdeA.aquasaliseA.albimanusnaativaçãodeFatorBeC3
foi observado através de Western blot utilizando anticorpos específicos para
cada componente. Na presença do EGS das duas espécies, houve inibição da
ativaçãodeFatorB(Figura13),poisnãohouveosurgimentodabandareferente
aBb(produtodaativaçãodoFatorB),comoobservadonocontrolepositivo,em
quefoiacrescentadoapenasPBS,semEGS.
O EGS das duas espécies também foi capaz de inibir a ativação do
componenteC3.Napresençadoextratodeglândulas,nãohouveaparecimento
dabandareferenteaC3a,produtodaativaçãodocomponenteC3,após30e60
minutosdeincubaçãoa37ºC(Figura14).
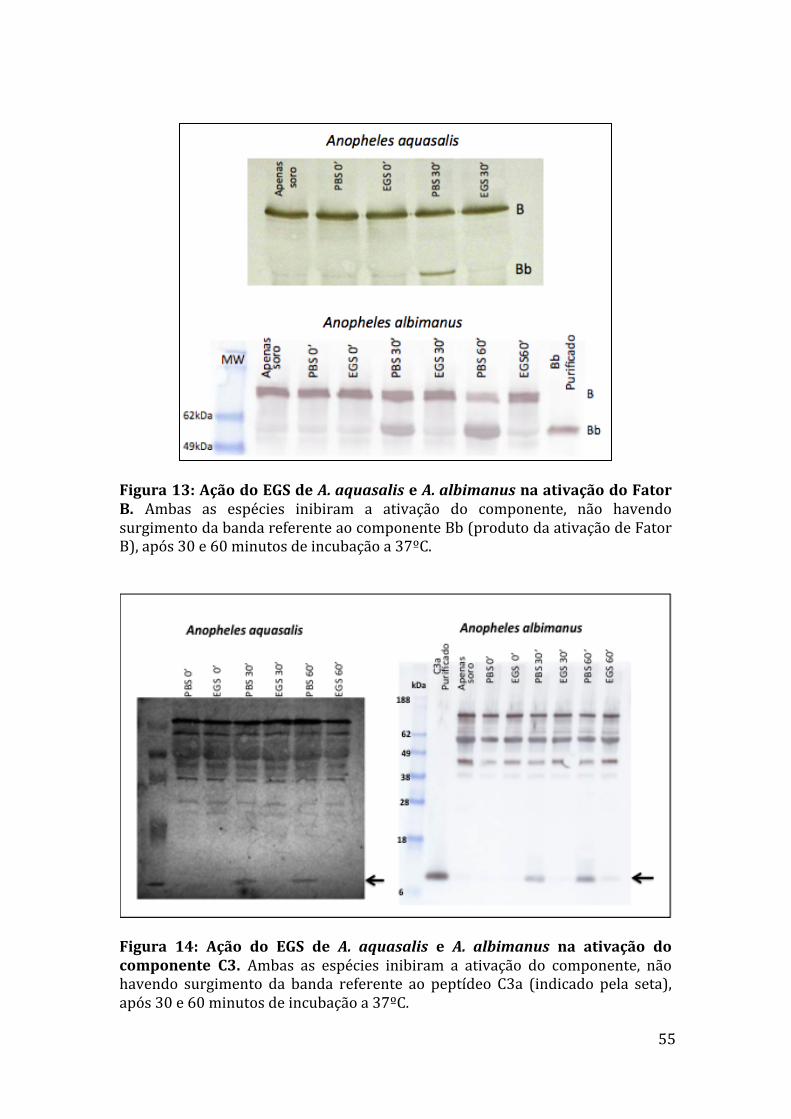
55
Figura13:AçãodoEGSdeA.aquasaliseA.albimanusnaativaçãodoFatorB. Ambas as espécies inibiram a ativação do componente, não havendosurgimentodabandareferenteaocomponenteBb(produtodaativaçãodeFatorB),após30e60minutosdeincubaçãoa37ºC.
Figura 14: Ação do EGS de A. aquasalis e A. albimanus na ativação docomponente C3. Ambas as espécies inibiram a ativação do componente, nãohavendo surgimento da banda referente ao peptídeo C3a (indicado pela seta),após30e60minutosdeincubaçãoa37ºC.

56
5.6WesternblotcomC3b,FatorBeFatorDpurificados Para investigar se o EGS de A. albimanus interfere diretamente na
formação da C3-convertase da via alternativa, formou-se este complexo
enzimático artificialmente através da incubação a 37ºC dos componentes
purificados, na presença e ausência do EGS. Com o conteúdo do tubo em
diferentes tempos de incubação, foi realizado Western blot usando anticorpo
anti-Fator B. Conforme demonstrado na Figura 15, o EGS não interferiu na
ativaçãodoFatorB,sendopossíveladetecçãodosprodutosdoFatorB,BbeBa
após20e40minutosdeincubação,mesmonapresençadoextratodeglândulas.
Figura15:AçãodoEGSdeA.albimanusnaformaçãodaC3-convertasecomcomponentesC3b,FatorBeFatorDpurificados.OEGSnãoinibiuaativaçãodoFatorB, sendoobservadoasbandasreferentesaosprodutosdaativaçãodoFatorB(BbeBa)tantonascanaletascomesemEGS.5.7PurificaçãodoinibidorsalivardeA.albimanus
Acromatografiade fase reversaemaparelhodeHPLC foiutilizadapara
purificar o inibidor da via alternativa presente no extrato de 200 glândulas
salivaresdeA.albimanus.Ocromatogramageradoapresentoucercade10picos
deproteínas,separadasdeacordocomahidrofobicidade(Figura16-A).
Uma alíquota de cada fração a partir do primeiro pico foi testada no
ensaio hemolítico e quatro frações seguidas (frações 1H11, 1H12, 2A1 e 2A2)
apresentaram atividade anti-via alternativa (Figura 16-B). Sobrepondo o
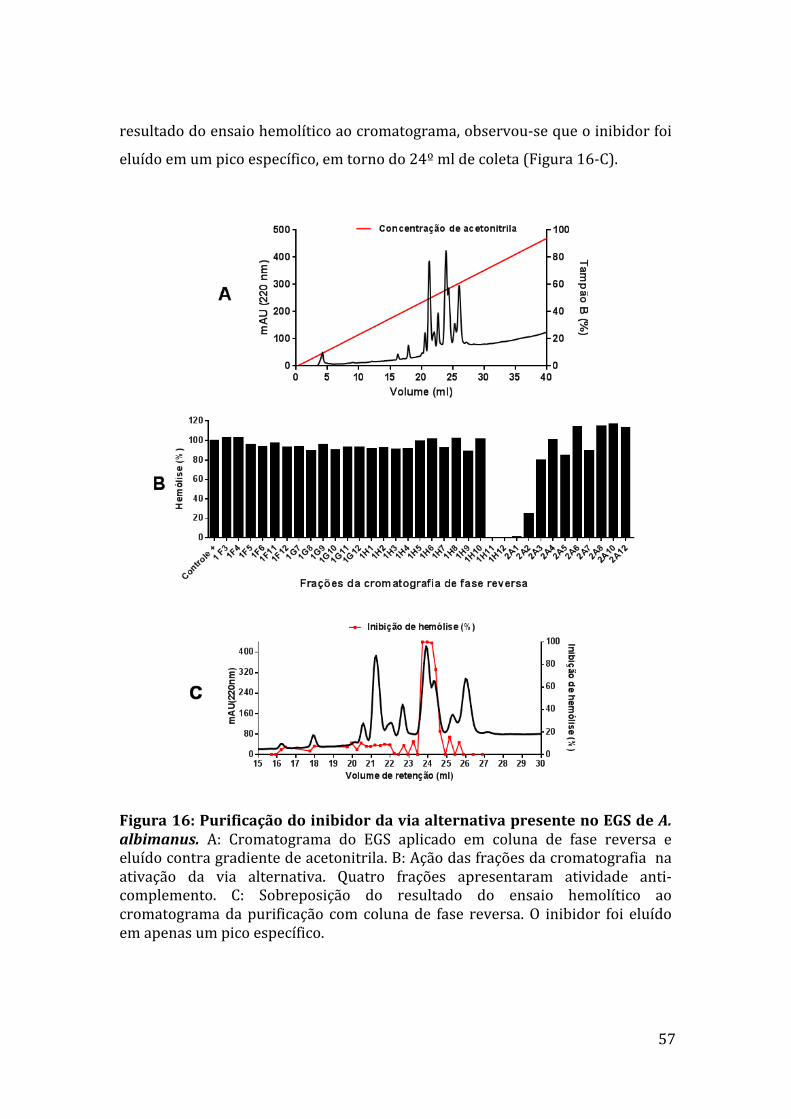
57
resultadodoensaiohemolíticoaocromatograma,observou-sequeoinibidorfoi
eluídoemumpicoespecífico,emtornodo24ºmldecoleta(Figura16-C).
Figura16:PurificaçãodoinibidordaviaalternativapresentenoEGSdeA.albimanus. A: Cromatograma do EGS aplicado em coluna de fase reversa eeluídocontragradientedeacetonitrila.B:Açãodasfraçõesdacromatografianaativação da via alternativa. Quatro frações apresentaram atividade anti-complemento. C: Sobreposição do resultado do ensaio hemolítico aocromatogramadapurificação comcolunade fase reversa.O inibidor foi eluídoemapenasumpicoespecífico.
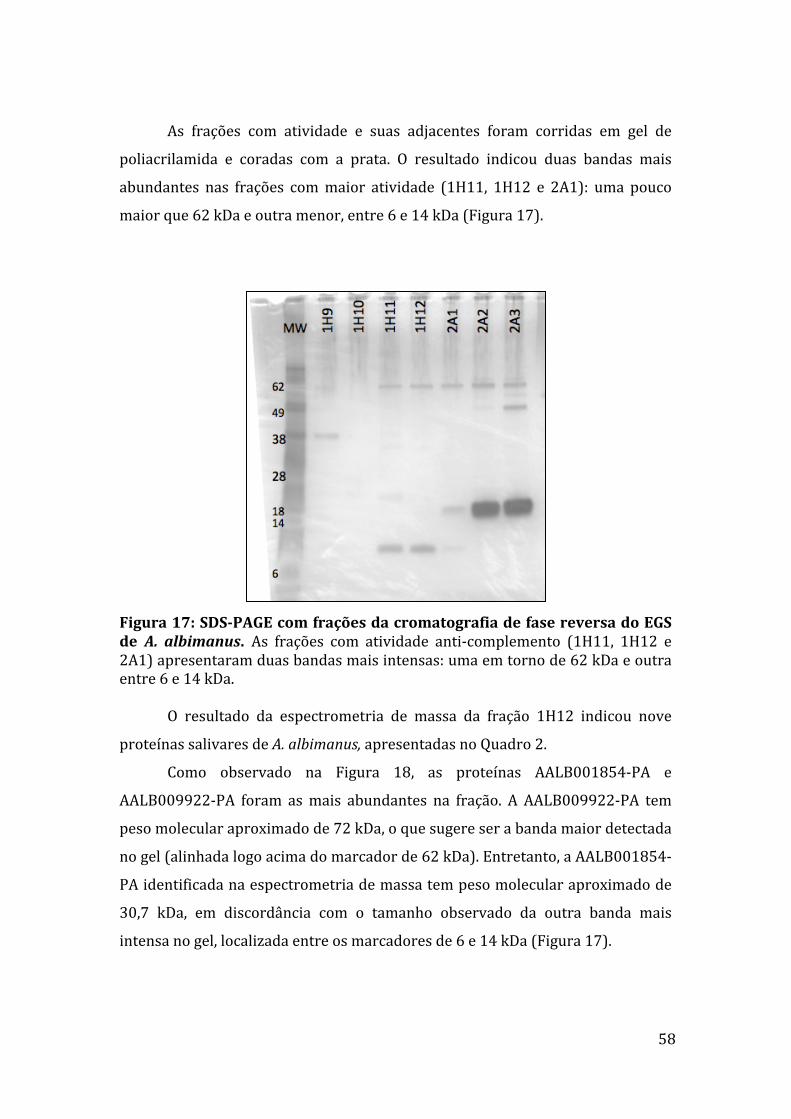
58
As frações com atividade e suas adjacentes foram corridas em gel de
poliacrilamida e coradas com a prata. O resultado indicou duas bandas mais
abundantes nas frações commaior atividade (1H11, 1H12 e 2A1): uma pouco
maiorque62kDaeoutramenor,entre6e14kDa(Figura17).
Figura17:SDS-PAGEcomfraçõesdacromatografiadefasereversadoEGSde A. albimanus. As frações com atividade anti-complemento (1H11, 1H12 e2A1)apresentaramduasbandasmaisintensas:umaemtornode62kDaeoutraentre6e14kDa. O resultado da espectrometria de massa da fração 1H12 indicou nove
proteínassalivaresdeA.albimanus,apresentadasnoQuadro2.
Como observado na Figura 18, as proteínas AALB001854-PA e
AALB009922-PA foram asmais abundantes na fração. A AALB009922-PA tem
pesomolecularaproximadode72kDa,oquesugereserabandamaiordetectada
nogel(alinhadalogoacimadomarcadorde62kDa).Entretanto,aAALB001854-
PAidentificadanaespectrometriademassatempesomolecularaproximadode
30,7 kDa, em discordância com o tamanho observado da outra banda mais
intensanogel,localizadaentreosmarcadoresde6e14kDa(Figura17).
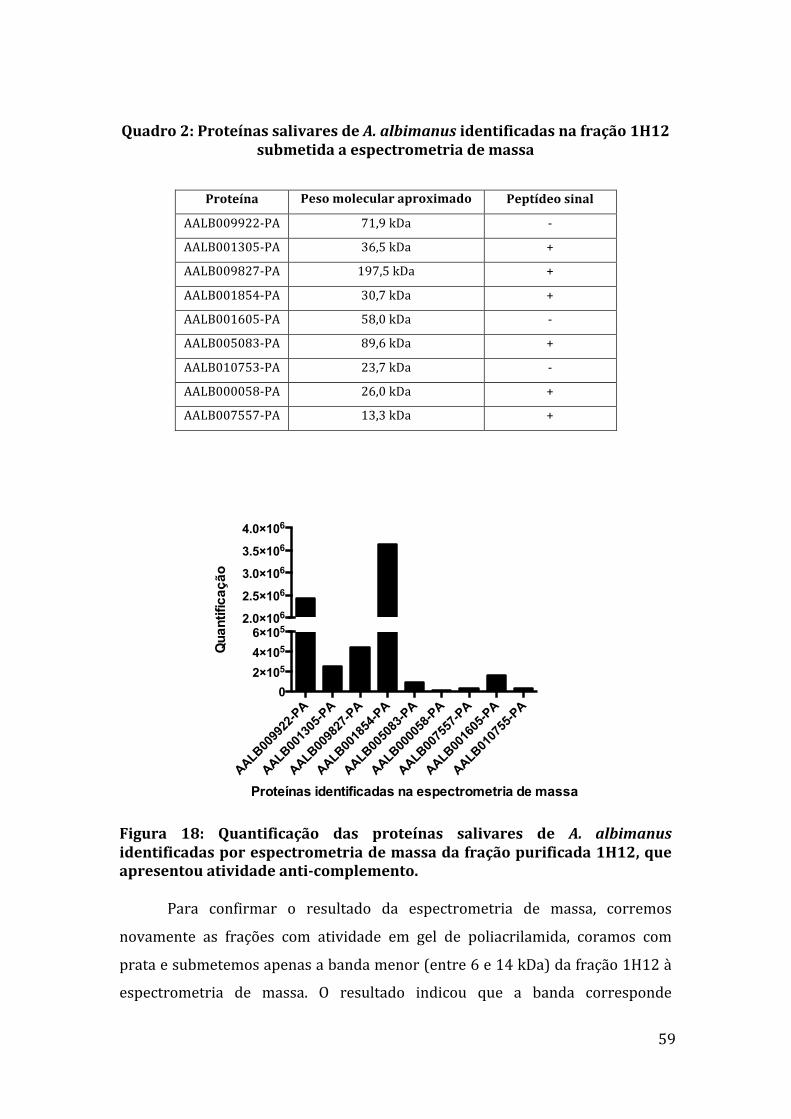
59
Quadro2:ProteínassalivaresdeA.albimanusidentificadasnafração1H12submetidaaespectrometriademassa
Figura 18: Quantificação das proteínas salivares de A. albimanusidentificadasporespectrometriademassadafraçãopurificada1H12,queapresentouatividadeanti-complemento.
Para confirmar o resultado da espectrometria de massa, corremos
novamente as frações com atividade em gel de poliacrilamida, coramos com
prataesubmetemosapenasabandamenor(entre6e14kDa)dafração1H12à
espectrometria de massa. O resultado indicou que a banda corresponde
AALB0099
22-P
A
AALB0013
05-P
A
AALB0098
27-P
A
AALB0018
54-P
A
AALB0050
83-P
A
AALB0000
58-P
A
AALB0075
57-P
A
AALB0016
05-P
A
AALB0107
55-P
A0
2×105
4×105
6×1052.0×106
2.5×106
3.0×106
3.5×106
4.0×106
Proteínas identificadas na espectrometria de massa
Qua
ntifi
caçã
o Proteína Pesomolecularaproximado Peptídeosinal
AALB009922-PA 71,9kDa -
AALB001305-PA 36,5kDa +
AALB009827-PA 197,5kDa +
AALB001854-PA 30,7kDa +
AALB001605-PA 58,0kDa -
AALB005083-PA 89,6kDa +
AALB010753-PA 23,7kDa -
AALB000058-PA 26,0kDa +
AALB007557-PA 13,3kDa +

60
realmente à proteína AALB001854-PA. Usando a sequência desta proteína
disponível no VectorBase, fizemos BLASTp no site do NCBI contra outros
anofelinosevimosqueestaproteína temalta similaridade (e-value de5e-32a
2e-41) com a proteína salivar gSG7 de A. darlingi, A. funestus, A. gambiae, A.
sinensiseA.stephensi.
No banco de dados do VectorBase, o gene AALB001854 desta proteína
apresenta seis éxons, com um longo íntron no meio da sequência, conforme
demonstradonaFigura19. Entretanto, ao analisarmosmais cuidadosamente a
sequência, notamos que, na verdade, se trata de dois genes distintos, que
provavelmenteforamanotadoserroneamentecomosendoumsónogenomade
A.albimanus.Oprimeirogenecomeçarealmentenostartcódondogeneanotado,
mas termina em stop códon (TAA) um pouco depois do final do terceiro éxon
(Figura20). Jáo segundogene, começano start códonumpoucoantesdoque
seriaoquartoéxondogeneanotado(Figura20)eterminanostopcódon(TAA)
comoanotadopreviamente.

61
TCCATGTCTACCAAAGACACCTTCCCCTTTCACACGACACCAGTGATTGCCAGAAATGTC CGATGTCACACAACAAACATCAACAATGGGGTCCAACTGATTATGGAAGTTATAATTTTA TTTACAACGCCAGCACGTACTACCATAACTTTGTGCACGCTCCTCCTGGCTGTGGCGTTG ACTGTGGCTTGCTGGTGCATGTGGAGGTACTGGCCGGAGGTTGGCTTGGTTTCCCCTTTC CCCAGTCTCCTCACTTGCAACCATTCATCAAAGTCACACAACATTTACTCAACCATGGTC GGTGTGAAACAGTTGGAGAGGAGTGCACCAGCAGTGATTGTACAAACAAGCAGTTTCCGT TTGTATTTAATTCAACTCTCTGTCTCCTAGCTTCCCTCTTTCTCTCTCTCTCTCTCTCTC TCTTTCTGTCTCTCTGGATATCTTCAACTCTCATCCCATCCGACCTTTGGCGAAACACTT GCGTAAACAGTTACTTCTTCACAGCGAATGCTCGTGAGGAAATAAAAAGGGAATCCATTA GCAATGGGAACCGGAGAAGTTGCTGGAAGTTAACCAATTATACCTGGAGTTCCTAATAGG ATGGCCGTTAGAATGACAGTCATATTGCCGTTGGCAATGGCCTTGATCTGCCTGATGCAG GTATGCAGCGCCTTTCGGCGCCCTACTACATCACATTCAATGCATAATTTCGCTTCCGAT TATTCACTTGCTAGGCTGAACCAGCTACGGCGGCAAATAACCACATCAGGACCGTTCTGA AGCTCTTCCGCACCATTGATCTGGACGATTCGAAAAAGAGCTTCTACCTAACAGCAGCCA AGTATGGCATCCAGACGCAGCTTCGCGAACCGATCATTCGAATCGTTGGAGGCTATCTTC CATCCACTAAGGTATGTCTATTCGATGATGCTCTGTGATCACATTAGTATCTGATCCAAC CGGGTCACTTGCAGCTATCAGAAGCATGCGTGAAGAATATGATCAGCGAGGTATACGAAA TAGAAGGCGACTTTTACTCCAAGTTCAGCTACGCGTGTGAAGACCATGCTCCCTACTCGG TCGAATGCCTGGAGGACGCTAGGGACGACTACCTCACCCAACTGGTGGAGCTGTTCAAGG AAACCAAGAAATGTTTGCGGGAATAAGATGAAGTATGTGACAAACTAAAATGCAGATAAA CCAATAAACTGTATGCAAAGCGCATGTTCCGGTACATGGAATTATGATTGAGTTGATTGC TTGAAACATGAGGTTTCTGTGAAGAACAGCAGGGCCTCCAAACAATATTCTATGGAAGCA TTGACTAATGGTGTGTCTTAACTGGAAACATTTATTACTTTGTCTGCATGTCAGCATTGA CCTCAGAATGTATTGTTCGTTCCATTCCTGCCAATTTTCAGAACACCAGCTTCCCAACGA GCTTCATTATTTCCCGAAGAGCGGATTCCCCCCAGAGACCGACAACAGTCTTGTTTATGA GGGATGGGATCAATATTGAGAAAGGTTCATCATCGTCCGATCCAGGTTTGCTCACCTGGC ATCCTATCCCATCAACCTGATACCCTATCCTGCAGAAAGTAATTGCATCAAGGCATGGTT TCTGTTTGCACTCACAGCTAGAGATAGGTGAGCAATCATTTGGTCTGGGACTGGGCTCCA TGCACCCTTTCTAACAAATCATTCGACGGGGAAAAACAGTATAAGTGTAGCAATTGAACG GAATTCACTTCAATCCAGTGTTGATTGTGCATCAGGATGGAACTTATTGGTACTAATGTT GTAATGTTAGCCGCATTTGCATTGATCAGCATAATGCAGGTACTGGAAATGCGACGTAGC AGTGCTTTCAGATACAGTACCTTCCTTGTAACTGTCGTGATTTATTGGGTTACTTCCAGT TCAATGCCATCGATGCGACACACGAGCACGCCAACAAAATGACAAACATCTTCAGGCGCG TCAAGCTAGATAAAACGAAAAACGCAGTTTATCAAGATTACGTCCAGAAGGGAATCAAGA CGCTGCTGAAGGATCCGCTCGTCTCGAAGGCAATGCTTCTGCCGGCCAGCAAATCGGTAA GCTGAATCGGTTAACTTCGACCTCCGATCCCAGTTAGTGTACTAGCTCCCTTTACTCCGT TATTTAAGCTTCCCGACGACTGCCTAAACGCCATGGTGGACGAAGCGAGAGAGCATGAGA ACAAGTTCTACGCGGTTTTTACGTACGACTGCCAAGATCACATCCCCACAGCTTTCCCAT GCTTGGAAAAGGGAGTAGCCACGTACTATGAGAACCTGAAAGCGCTGGAAAAATCCACTG AAAAGTGTTGCAACATGTAATAGCCGCTTTACCACTTTTATGATGACGATGATGATGATG TAAATGGTGATTGTGTTAGCATGAAATACAATGAAAATATCACAGGCAAATTGAATGATT GCTGTCCATTTTTATTCTCAAAACAATTTCCGATTTTCTCCCCAAAAAAAAAAATAGAAC CGGCCATTCGCTTGTGATGGTTGGAGCACGATTCCGAGTACACTGTCGACAGTTTTTGTC CAACCAACCGGGCGCAGATAGACTGGAAAAATTAAATAAACAAATCCTCACCCCGGAATA TCACTGAAAGCCTCCGATGAGCCGTTGGCACGTAAACGAATCCATGAAAAAAAGGGGTTG ATTTGTGGACCATCGCAAAAAGCCCACAGTCAACGGAACGTCCACCCAGGCCCAGCTCAC CCAGTGGATCATCGGGTCCACGGTCAGTTCAACCATTCTCGCTTATTCGTTCTCCAACAG AAAGGGCTGGAAAAAACGAGCAGAAGTTATTAAAATAATGCCACATCGAAGGGACTCTCC CAATGAAGGAAGCACGATCCTCACGGAAAAAAACAATAGCTCTTCACGTCCTCAGCTGGC AAAAAAAAAAAACAATGGCC
Figura19: Sequênciado geneAALB001854disponívelnoVectorBase.org.Emdestaque,asregiõesdasequênciaapresentadascomocodificadoras.
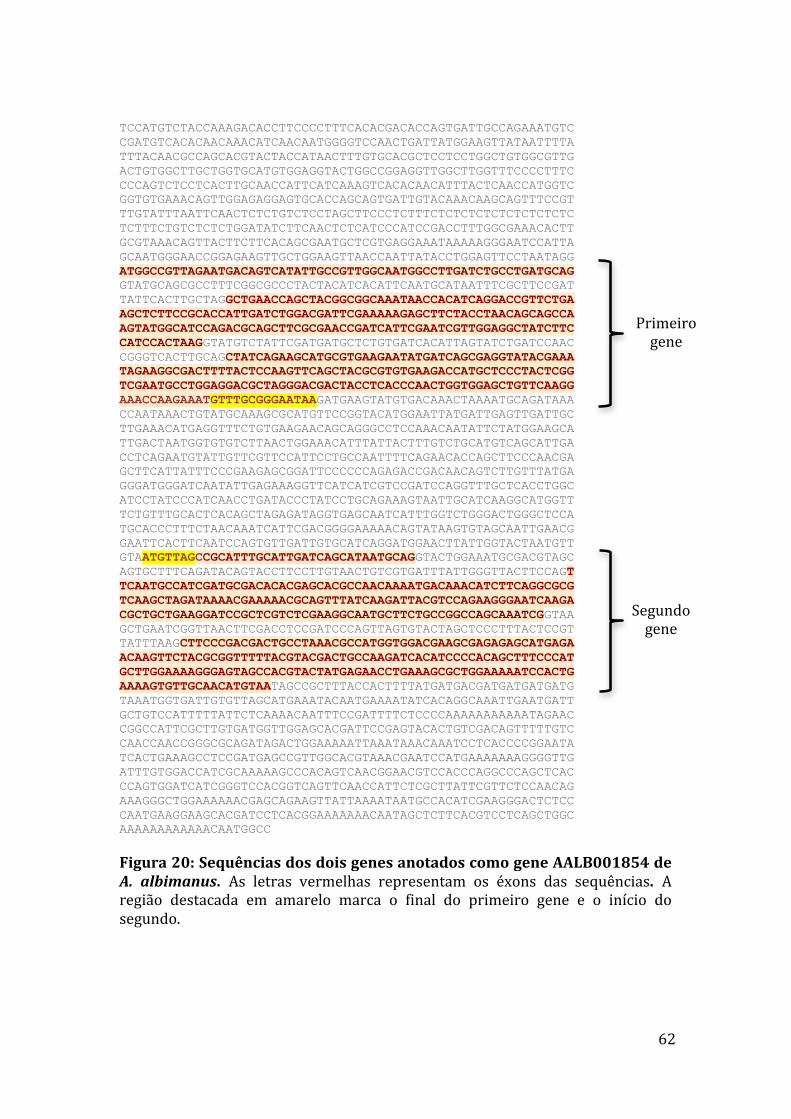
62
TCCATGTCTACCAAAGACACCTTCCCCTTTCACACGACACCAGTGATTGCCAGAAATGTC CGATGTCACACAACAAACATCAACAATGGGGTCCAACTGATTATGGAAGTTATAATTTTA TTTACAACGCCAGCACGTACTACCATAACTTTGTGCACGCTCCTCCTGGCTGTGGCGTTG ACTGTGGCTTGCTGGTGCATGTGGAGGTACTGGCCGGAGGTTGGCTTGGTTTCCCCTTTC CCCAGTCTCCTCACTTGCAACCATTCATCAAAGTCACACAACATTTACTCAACCATGGTC GGTGTGAAACAGTTGGAGAGGAGTGCACCAGCAGTGATTGTACAAACAAGCAGTTTCCGT TTGTATTTAATTCAACTCTCTGTCTCCTAGCTTCCCTCTTTCTCTCTCTCTCTCTCTCTC TCTTTCTGTCTCTCTGGATATCTTCAACTCTCATCCCATCCGACCTTTGGCGAAACACTT GCGTAAACAGTTACTTCTTCACAGCGAATGCTCGTGAGGAAATAAAAAGGGAATCCATTA GCAATGGGAACCGGAGAAGTTGCTGGAAGTTAACCAATTATACCTGGAGTTCCTAATAGG ATGGCCGTTAGAATGACAGTCATATTGCCGTTGGCAATGGCCTTGATCTGCCTGATGCAG GTATGCAGCGCCTTTCGGCGCCCTACTACATCACATTCAATGCATAATTTCGCTTCCGAT TATTCACTTGCTAGGCTGAACCAGCTACGGCGGCAAATAACCACATCAGGACCGTTCTGA AGCTCTTCCGCACCATTGATCTGGACGATTCGAAAAAGAGCTTCTACCTAACAGCAGCCA AGTATGGCATCCAGACGCAGCTTCGCGAACCGATCATTCGAATCGTTGGAGGCTATCTTC CATCCACTAAGGTATGTCTATTCGATGATGCTCTGTGATCACATTAGTATCTGATCCAAC CGGGTCACTTGCAGCTATCAGAAGCATGCGTGAAGAATATGATCAGCGAGGTATACGAAA TAGAAGGCGACTTTTACTCCAAGTTCAGCTACGCGTGTGAAGACCATGCTCCCTACTCGG TCGAATGCCTGGAGGACGCTAGGGACGACTACCTCACCCAACTGGTGGAGCTGTTCAAGG AAACCAAGAAATGTTTGCGGGAATAAGATGAAGTATGTGACAAACTAAAATGCAGATAAA CCAATAAACTGTATGCAAAGCGCATGTTCCGGTACATGGAATTATGATTGAGTTGATTGC TTGAAACATGAGGTTTCTGTGAAGAACAGCAGGGCCTCCAAACAATATTCTATGGAAGCA TTGACTAATGGTGTGTCTTAACTGGAAACATTTATTACTTTGTCTGCATGTCAGCATTGA CCTCAGAATGTATTGTTCGTTCCATTCCTGCCAATTTTCAGAACACCAGCTTCCCAACGA GCTTCATTATTTCCCGAAGAGCGGATTCCCCCCAGAGACCGACAACAGTCTTGTTTATGA GGGATGGGATCAATATTGAGAAAGGTTCATCATCGTCCGATCCAGGTTTGCTCACCTGGC ATCCTATCCCATCAACCTGATACCCTATCCTGCAGAAAGTAATTGCATCAAGGCATGGTT TCTGTTTGCACTCACAGCTAGAGATAGGTGAGCAATCATTTGGTCTGGGACTGGGCTCCA TGCACCCTTTCTAACAAATCATTCGACGGGGAAAAACAGTATAAGTGTAGCAATTGAACG GAATTCACTTCAATCCAGTGTTGATTGTGCATCAGGATGGAACTTATTGGTACTAATGTT GTAATGTTAGCCGCATTTGCATTGATCAGCATAATGCAGGTACTGGAAATGCGACGTAGC AGTGCTTTCAGATACAGTACCTTCCTTGTAACTGTCGTGATTTATTGGGTTACTTCCAGT TCAATGCCATCGATGCGACACACGAGCACGCCAACAAAATGACAAACATCTTCAGGCGCG TCAAGCTAGATAAAACGAAAAACGCAGTTTATCAAGATTACGTCCAGAAGGGAATCAAGA CGCTGCTGAAGGATCCGCTCGTCTCGAAGGCAATGCTTCTGCCGGCCAGCAAATCGGTAA GCTGAATCGGTTAACTTCGACCTCCGATCCCAGTTAGTGTACTAGCTCCCTTTACTCCGT TATTTAAGCTTCCCGACGACTGCCTAAACGCCATGGTGGACGAAGCGAGAGAGCATGAGA ACAAGTTCTACGCGGTTTTTACGTACGACTGCCAAGATCACATCCCCACAGCTTTCCCAT GCTTGGAAAAGGGAGTAGCCACGTACTATGAGAACCTGAAAGCGCTGGAAAAATCCACTG AAAAGTGTTGCAACATGTAATAGCCGCTTTACCACTTTTATGATGACGATGATGATGATG TAAATGGTGATTGTGTTAGCATGAAATACAATGAAAATATCACAGGCAAATTGAATGATT GCTGTCCATTTTTATTCTCAAAACAATTTCCGATTTTCTCCCCAAAAAAAAAAATAGAAC CGGCCATTCGCTTGTGATGGTTGGAGCACGATTCCGAGTACACTGTCGACAGTTTTTGTC CAACCAACCGGGCGCAGATAGACTGGAAAAATTAAATAAACAAATCCTCACCCCGGAATA TCACTGAAAGCCTCCGATGAGCCGTTGGCACGTAAACGAATCCATGAAAAAAAGGGGTTG ATTTGTGGACCATCGCAAAAAGCCCACAGTCAACGGAACGTCCACCCAGGCCCAGCTCAC CCAGTGGATCATCGGGTCCACGGTCAGTTCAACCATTCTCGCTTATTCGTTCTCCAACAG AAAGGGCTGGAAAAAACGAGCAGAAGTTATTAAAATAATGCCACATCGAAGGGACTCTCC CAATGAAGGAAGCACGATCCTCACGGAAAAAAACAATAGCTCTTCACGTCCTCAGCTGGC AAAAAAAAAAAACAATGGCC
Figura20:SequênciasdosdoisgenesanotadoscomogeneAALB001854deA. albimanus. As letras vermelhas representam os éxons das sequências. Aregião destacada em amarelo marca o final do primeiro gene e o início dosegundo.
Primeirogene
Segundogene

63
O primeiro gene codifica a proteína denominada de gSG7, cuja
sequênciaestádescritanaFigura21.Elaapresentapeptídeosinal,comsítiode
clivagem entre o 26º e 27º aminoácido. A proteína secretada (sem peptídeo
sinal),tempesomolecularde13,4kDa,écompostapor116aminoácidoseponto
isoelétrico5,39.
MAVRMTVILPLAMALICLMQAEPATAANNHIRTVLKLFRTIDLDDSKKSFYLTAAKYGIQTQLREPIIRIVGGYLPSTKLSEACVKNMISEVYEIEGDFYSKFSYACEDHAPYSVECLEDARDDYLTQLVELFKETKKCLRE
Figura21:SequênciadaproteínasalivargSG7deA.albimanus.Emvermelhoestá representado o sítio de clivagem do peptídeo sinal, entre o 26º e 27ºaminoácido.
JáosegundogenecodificaaproteínadenominadadegSG7-2,mostrada
na figura 22. Esta proteína também apresenta peptídeo sinal, sendo o sítio de
clivagem entre o 18º e o 19º aminoácido. Sem o peptídeo sinal, a gSG7-2 tem
peso molecular de 13,3 kDa, é composta por 116 aminoácidos e tem ponto
isoelétrico7,72.
MLAAFALISIMQFNAIDATHEHANKMTNIFRRVKLDKTKNAVYQDYVQKGIKTLLKDPLVSKAMLLPASKSLPDDCLNAMVDEAREHENKFYAVFTYDCQDHIPTAFPCLEKGVATYYENLKALEKSTEKCCNM
Figura 22: Sequência da proteína salivar gSG7-2 de A. albimanus. Emvermelhoestárepresentadoosítiodeclivagemdopeptídeosinal,entreo18ºe19ºaminoácido.
AsproteínasgSG7egSG7-2deA.albimanusapresentamapenas32,6%
de similaridade e, interessantemente, os peptídeos gerados na digestão por
tripsina para análise por espectrometria de massa da proteína salivar de A.
albimanusrecuperadadogelapresentaramsimilaridadeapenascomfragmentos
daprimeiraproteína(gSG7),conformedemonstradonaFigura23.

64
Figura 23: Alinhamento das sequências dos peptídeos gerados pelotratamento com tripsina da proteína salivar recuperada do gel com asequênciadagSG7deA.albimanusobtidanoVectorBase.
No NCBI, acessamos a sequência da gSG7 de A. darlingi (GenBank:
ACI30142.1)evimosqueelatambémtempeptídeosinal(entreosaminoácidos
26 e 27 da sequência), tem pesomolecular de 13,4 kDa, é composta por 116
aminoácidosepontoisoelétrico7,77(Figura24).
MAARMTIMLPLAVALICLLQTEPGMAAHSHIRKVLQLFRSIELDDSKKSFYLTAAKYGIQTQLREPLVRFAGGFAPSTRLSEACVKNAIARIYEIEGEFYAKFSYACENHDPYSVECLEEAQDDYPTKLGELFKKTKKCLRE
Figura24:SequênciadaproteínasalivargSG7deA.darlingi.Emvermelhoestá representado o sítio de clivagem do peptídeo sinal, entre o 26º e 27ºaminoácido.
Comparando as sequências das gSG7s de A. albimanus e A. darlingi,
pudemos ver que asmesmas apresentam 75% de similaridade (Figura 25). Já
quando comparada com as sequências de gSG7s de A. gambiae (GenBank:
CAC35523.1), A. funestus (GenBank: ABI83776.1), A. sinensis (GenBank:
KFB36874.1)eA.stephensi(GenBank:AAO06828.1),aproteínadeA.albimanus
apresentouapenascercade40%desimilaridade(Figura25).

65
Figura 25: Comparação das sequências das proteínas gSG7 de diferentesespéciesdeAnopheles.(A)AlinhamentodassequênciasdaproteínagSG7deA.albimanus, A. darlingi, A. funestus, A. gambiae, A. sinensis e A. stephensi. Osasteriscosindicamosaminoácidosidênticosemtodasasproteínas,doispontosindicamaminoácidoscomaltasimilaridadeeumpontoindicaaminoácidoscompoucasimilaridade.(B)ÁrvorefilogenéticabaseadanasequênciadasproteínasgSG7, confirmando a proximidade maior entre as gSG7s de A. albimanus e A.darlingi.
5.8 RT-PCR para detecção do gene da proteína gSG7 em extrato de glândulas
salivaresdeA.albimanus
Com o objetivo de detectar o gene da proteína gSG7 no extrato de
glândulas salivares de fêmeas de A. albimanus, 50 glândulas salivares foram
dissecadas e tiveram RNA total extraído pelo método de Trizol. A partir do
material extraído, foi produzido cDNA utilizando primer específico para
sequência do gene, e em seguida, foi realizada PCR convencional, utilizando o
cDNAcomomoldeeumpardeiniciadoresespecíficosparaasequênciadogene
citado.

66
O produto da PCR corrido no gel apresentou apenas uma banda, em
tornode500paresdebase(Figura26),compatívelcomotamanhodoamplicon
gerado pelos primers utilizados (477pb). No controle negativo, não houve
amplificaçãodenenhummaterial,indicandoausênciadecontaminação.
Figura26:EletroforesedosprodutosdareaçãodePCRparaamplificaçãodogenedaproteínagSG7deA.albimanus.UsandoocDNAproduzidoapartirdeRNAextraídodeglândulassalivares,houveamplificaçãoapenasdeumgene,que coincide com o tamanho aproximado do amplicon gerado pelos primersutilizados na reação (477pb). No tubo controle, não houve amplificação denenhumfragmentodeDNA.
5.9ExpressãoepurificaçãodasproteínasgSG7egSG7-2deA.albimanusegSG7
deA.darlingi
AsproteínasgSG7egSG7-2deA.albimanus egSG7deA.darlingi foram
expressas emE.coli BL21 pLys e purificadas através de cromatografias de gel
filtração em HPLC. Na Figura 27 estão representados os cromatogramas do
último passo da purificação das recombinantes, com formação de apenas um
pico,indicandoapurezadasnossasamostras.
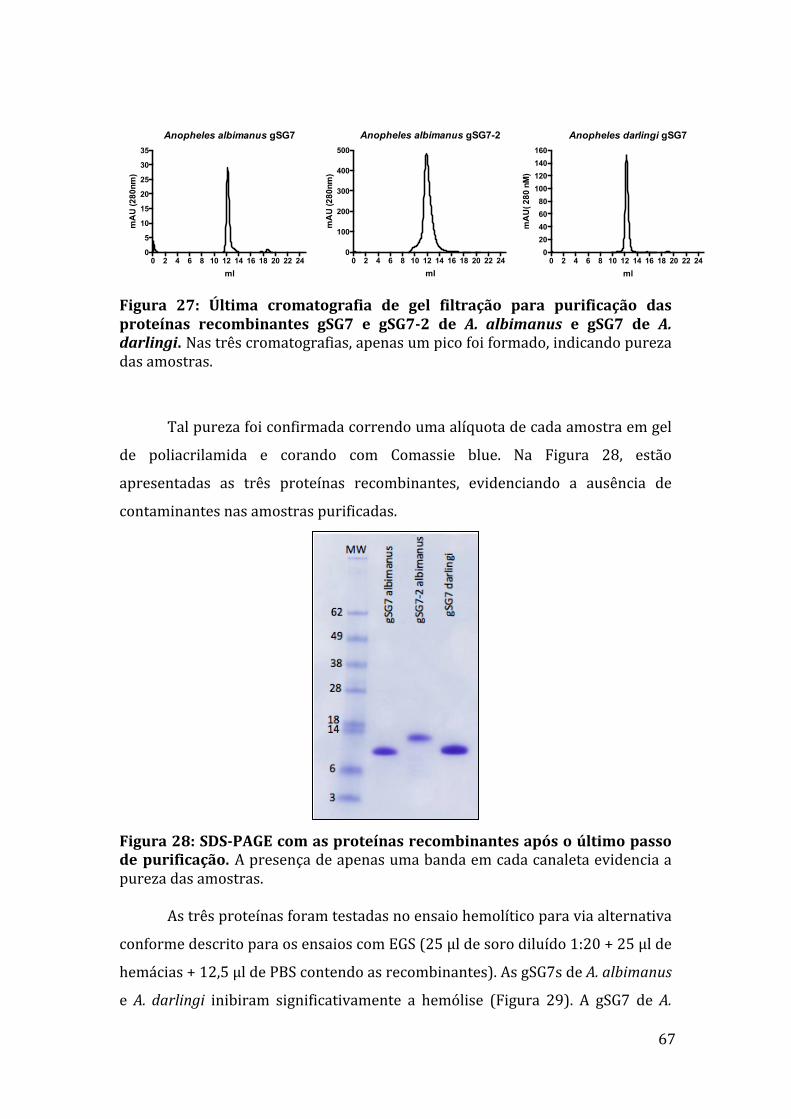
67
Figura 27: Última cromatografia de gel filtração para purificação dasproteínas recombinantes gSG7 e gSG7-2 de A. albimanus e gSG7 de A.darlingi.Nastrêscromatografias,apenasumpicofoiformado,indicandopurezadasamostras.
Talpurezafoiconfirmadacorrendoumaalíquotadecadaamostraemgel
de poliacrilamida e corando com Comassie blue. Na Figura 28, estão
apresentadas as três proteínas recombinantes, evidenciando a ausência de
contaminantesnasamostraspurificadas.
Figura28:SDS-PAGEcomasproteínasrecombinantesapósoúltimopassodepurificação.Apresençadeapenasumabandaemcadacanaletaevidenciaapurezadasamostras. Astrêsproteínasforamtestadasnoensaiohemolíticoparaviaalternativa
conformedescritoparaosensaioscomEGS(25μldesorodiluído1:20+25μlde
hemácias+12,5μldePBScontendoasrecombinantes).AsgSG7sdeA.albimanus
eA. darlingi inibiram significativamente a hemólise (Figura 29). A gSG7 deA.
0 2 4 6 8 10 12 14 16 18 20 22 240
5
10
15
20
25
30
35
ml
mA
U (2
80nm
)Anopheles albimanus gSG7
0 2 4 6 8 10 12 14 16 18 20 22 240
100
200
300
400
500
ml
mA
U (2
80nm
)
Anopheles albimanus gSG7-2
0 2 4 6 8 10 12 14 16 18 20 22 240
20406080
100120140160
ml
Anopheles darlingi gSG7
mA
U( 2
80 n
M)
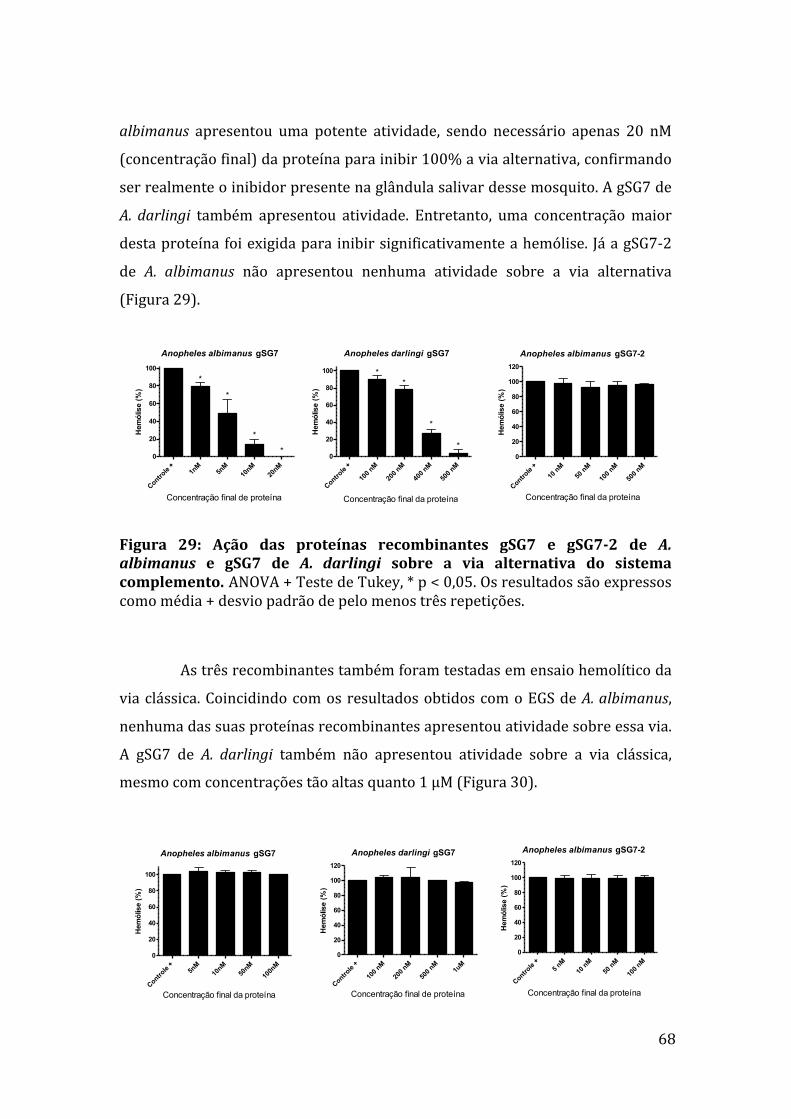
68
albimanus apresentou uma potente atividade, sendo necessário apenas 20 nM
(concentraçãofinal)daproteínaparainibir100%aviaalternativa,confirmando
serrealmenteoinibidorpresentenaglândulasalivardessemosquito.AgSG7de
A.darlingi também apresentou atividade. Entretanto, uma concentraçãomaior
destaproteínafoiexigidaparainibirsignificativamenteahemólise. JáagSG7-2
de A. albimanus não apresentou nenhuma atividade sobre a via alternativa
(Figura29).
Controle
+1n
M5n
M10
nM20
nM0
20
40
60
80
100
Anopheles albimanus gSG7
*
*
*
*
Concentração final de proteína
Hem
ólis
e (%
)
Controle
+10
nM50
nM
100 n
M
500 n
M0
20
40
60
80
100
120
Anopheles albimanus gSG7-2
Concentração final da proteínaH
emól
ise
(%)
Controle
+
100 n
M
200 n
M
400 n
M
500 n
M0
20
40
60
80
100
Anopheles darlingi gSG7
*
*
**
Concentração final da proteína
Hem
ólis
e (%
)
Figura 29: Ação das proteínas recombinantes gSG7 e gSG7-2 de A.albimanus e gSG7 de A. darlingi sobre a via alternativa do sistemacomplemento.ANOVA+TestedeTukey,*p<0,05.Osresultadossãoexpressoscomomédia+desviopadrãodepelomenostrêsrepetições.
Astrêsrecombinantestambémforamtestadasemensaiohemolíticoda
viaclássica.CoincidindocomosresultadosobtidoscomoEGSdeA.albimanus,
nenhumadassuasproteínasrecombinantesapresentouatividadesobreessavia.
A gSG7 de A. darlingi também não apresentou atividade sobre a via clássica,
mesmocomconcentraçõestãoaltasquanto1μM(Figura30).
Controle
+
100 n
M
200 n
M
500 n
M1u
M0
20
40
60
80
100
120
Anopheles darlingi gSG7
Concentração final de proteína
Hem
ólis
e (%
)
Controle
+5 n
M10
nM50
nM
100 n
M0
20
40
60
80
100
120
Anopheles albimanus gSG7-2
Concentração final da proteína
Hem
ólis
e (%
)
Controle
+5n
M10
nM50
nM10
0nM
0
20
40
60
80
100
Anopheles albimanus gSG7
Concentração final da proteína
Hem
ólis
e (%
)

69
Figura 30: Ação das proteínas recombinantes gSG7 e gSG7-2 de A.albimanus e gSG7 de A. darlingi sobre a via clássica do sistemacomplemento.Osresultadossãoexpressoscomomédia+desviopadrãodepelomenostrêsrepetições.
AsproteínasrecombinantesdeA.albimanusforamtestadastambémno
ensaiodeativaçãodaviadaslectinas.Emnenhumadasconcentraçõestestadas
as proteínas apresentaram inibição da deposição de C3b ativado por esta via
(Figura31).
Anopheles albimanus gSG7
Controle
+5 n
M10
nM20
nM
100 n
M0
20
40
60
80
100
120
Concentração final da proteína
Dep
osiç
ão d
e C
3b (%
)
Anopheles albimanus gSG7-2
Controle
+5 n
M10
nM20
nM
100 n
M0
20
40
60
80
100
120
Concentração final da proteína
Dep
osiç
ão d
e C
3b (%
)
Figura31:EfeitodasproteínasgSG7egSG7-2deA.albimanus sobreaviadas lectinas. As barras representam a média e desvios padrões dasporcentagens de deposição de C3b em relação ao controle positivo (seminibidor).
EnsaiosdedeposiçãodeC3b,FatorB,properdinaeC9napresençade
gSG7deA.albimanuseA.darlingiedagSG7-2deA.albimanusemplacacoberta
com agarose foram realizados, conforme descrito para os ensaios comEGS no
Material e Métodos (Item 4.5). Houve inibição dose dependente e
estatisticamente significativa da deposição de todos os componentes testados
apenas na presença da proteína gSG7 (Figuras 32 e 33). A proteína de A.
albimanussemostroumaispotente,precisandodeconcentraçõesfinaisemtorno
de20nMparainibirquase100%adeposiçãodetodososcomponentes(Figura
32).
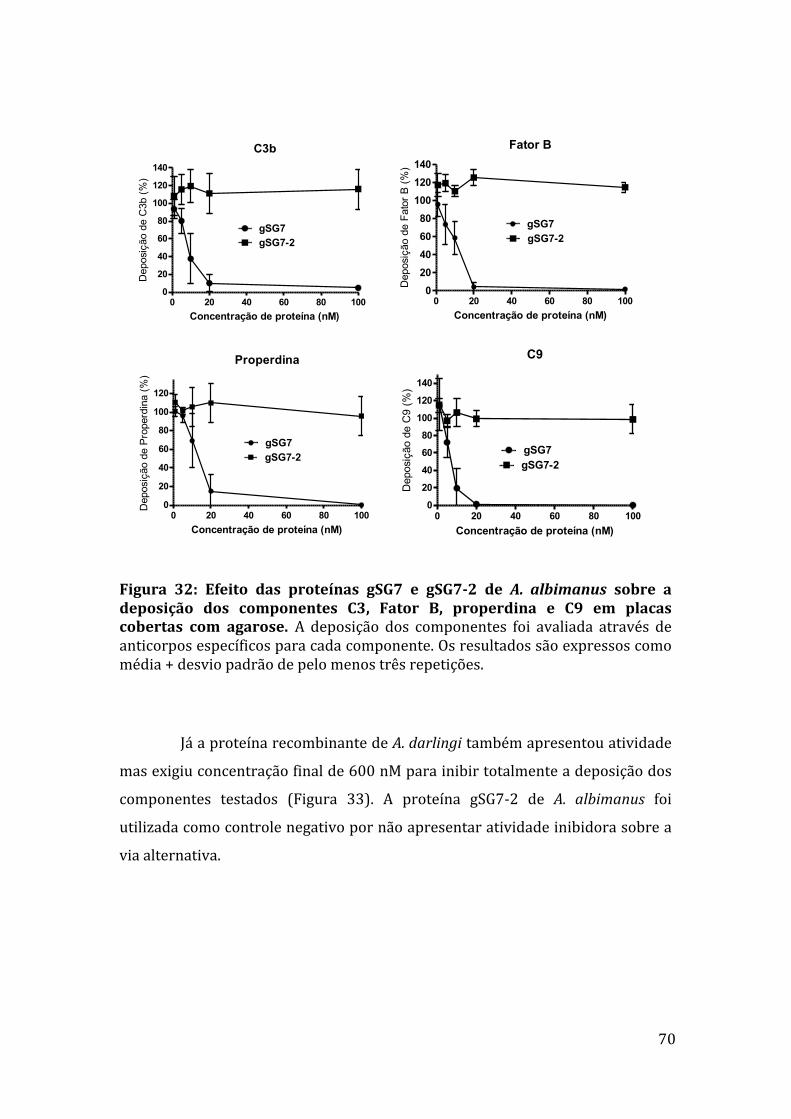
70
C3b
0 20 40 60 80 1000
20
40
60
80
100
120
140
gSG7gSG7-2
Concentração de proteína (nM)
Dep
osiç
ão d
e C
3b (%
)Fator B
0 20 40 60 80 1000
20406080
100120140
gSG7gSG7-2
Concentração de proteína (nM)
Dep
osiç
ão d
e Fa
tor B
(%)
Properdina
0 20 40 60 80 1000
20
40
60
80
100
120
gSG7gSG7-2
Concentração de proteína (nM)
Dep
osiç
ão d
e Pr
oper
dina
(%)
C9
0 20 40 60 80 1000
20
40
60
80
100
120
140
gSG7gSG7-2
Concentração de proteína (nM)
Dep
osiç
ão d
e C
9 (%
)
Figura 32: Efeito das proteínas gSG7 e gSG7-2 de A. albimanus sobre adeposição dos componentes C3, Fator B, properdina e C9 em placascobertas com agarose. A deposição dos componentes foi avaliada através deanticorposespecíficosparacadacomponente.Osresultadossãoexpressoscomomédia+desviopadrãodepelomenostrêsrepetições.
JáaproteínarecombinantedeA.darlingitambémapresentouatividade
masexigiuconcentraçãofinalde600nMparainibirtotalmenteadeposiçãodos
componentes testados (Figura 33). A proteína gSG7-2 de A. albimanus foi
utilizadacomocontrolenegativopornãoapresentaratividadeinibidorasobrea
viaalternativa.

71
Properdina
0.0 0.2 0.4 0.6 0.8 1.00
20
40
60
80
100
120
gSG7gSG7-2
Concentração de proteína (µM)
Dep
osiç
ão d
e Pr
oper
dina
(%)
C9
0.0 0.2 0.4 0.6 0.8 1.00
20
40
60
80
100
120
gSG7gSG7-2
Concentração de proteína ( µM)
Dep
osiç
ão d
e C
9 (%
)
Fator B
0.0 0.2 0.4 0.6 0.8 1.00
20
40
60
80
100
120
gSG7gSG7-2
Concentração de proteína (µM)
Dep
osiç
ão d
e Fa
tor B
(%)
C3b
0.0 0.2 0.4 0.6 0.8 1.00
20
40
60
80
100
120
gSG7gSG7-2
Concentração de proteína (µM)
Dep
osiç
ão d
e C
3b (%
)
Figura33:EfeitodasproteínasgSG7deA.darlingi sobreadeposiçãodoscomponentesC3,FatorB,properdinaeC9emplacascobertascomagarose.Adeposiçãodoscomponentesfoiavaliadaatravésdeanticorposespecíficosparacadacomponente.Osresultadossãoexpressoscomomédia+desviopadrãodepelomenostrêsrepetições.
A proteína gSG7 de A. albimanus foi também avaliada em ensaio de
desligamento de componentes da via alternativa previamente depositados em
superfície ativadora. Não houve diferença estatística entre os resultados de
deposição de C3b, FatorB ou properdina na presença ou ausência do inibidor
(Figura 34). Quando Fator H foi adicionado aos poços, houve diminuição
estatisticamentesignificativadadeposiçãodeFatorB(Figura34).
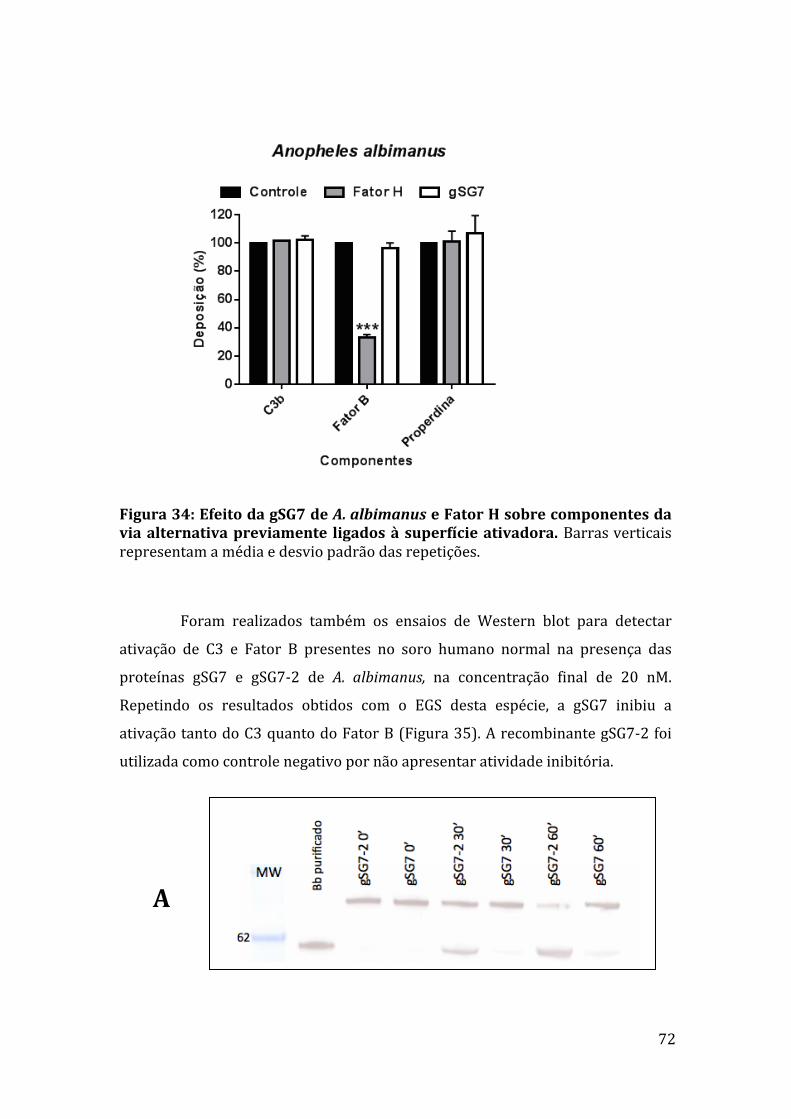
72
Figura34:EfeitodagSG7deA.albimanuseFatorHsobrecomponentesdaviaalternativapreviamente ligadosàsuperfícieativadora.Barrasverticaisrepresentamamédiaedesviopadrãodasrepetições.
Foram realizados também os ensaios de Western blot para detectar
ativação de C3 e Fator B presentes no soro humano normal na presença das
proteínas gSG7 e gSG7-2 de A. albimanus, na concentração final de 20 nM.
Repetindo os resultados obtidos com o EGS desta espécie, a gSG7 inibiu a
ativaçãotantodoC3quantodoFatorB(Figura35).ArecombinantegSG7-2foi
utilizadacomocontrolenegativopornãoapresentaratividadeinibitória.
A

73
Figura35:EfeitodasproteínasgSG7egSG7-2deA.albimanusnaativaçãodoscomponentesFatorB(A)eC3(B).NascanaletascomagSG7,nãohouveaparecimentodasbandasreferentesaBb(A)eC3a(B),produtosdaativaçãodeFatorBeC3,respectivamente.
OWesternblotcomoscomponentespurificadosC3b,FatorBeFatorD
também foi repetido, substituindo o EGS pelas proteínas recombinantes na
concentraçãofinalde20nM.NemagSG7nemagSG7-2 inibiramaativaçãode
FatorBnesseexperimento(Figura36).
Figura 36: Ação das proteínas recombinantes gSG7 e gSG7-2 de A.albimanusnaformaçãodaC3-convertasecomcomponentesC3b,FatorBeFator D purificados. As proteínas não inibiram a ativação do Fator B, sendoobservadoasbandasreferentesaosprodutosdaativaçãodoFatorB(BbeBa)nasrespectivascanaletas.
B

74
5.10IdentificaçãodoinibidorsalivardeA.aquasalis
AsproteínasgSG7recombinantesdeA.albimanusedeA.darlingiforam
corridasaoladodeEGSdefêmeasdeA.aquasalisemgeldepoliacrilamida12,5%
e coradas comprata.Nas canaletas emque foramaplicadas as recombinantes,
apenas uma banda com peso molecular de aproximadamente 13 kDa foi
observada.NacanaletaemqueoEGSfoiaplicado,aproximadamenteoitobandas
foram detectadas, incluindo uma com peso molecular muito próximo ao das
gSG7s(Figura37).
Figura 37: Eletroforese das proteínas gSG7 recombinantes de (1) A.albimanus e (2) A. darlingi e do (3) EGS de A. aquasalis em gel depoliacrilamida corado com prata. A seta indica uma banda no EGS com omesmopesomoleculardasproteínasrecombinantesdasoutrasespécies.
AsbandasreferentesàgSG7deA.darlingieasuacorrespondentenoEGS
deA.aquasalis foramextraídasdo gel e submetidas a tripsinização seguidade
espectrometria de massa. Dos peptídeos gerados a partir da amostra de A.
darlingi,umalinhoucom100%desimilaridadedeaminoácidoscomasequência
dagSG7dessaespécie.Interessantemente,umdospeptídeosgeradosapartirda
amostra de A. aquasalis também alinhou com 100% de similaridade com um
fragmentodiferentedasequênciadagSG7deA.darlingi(Figura38).

75
Figura 38: Alinhamento da sequência de peptídeo gerado da proteínasalivar de A. aquasalis (Pep1) com a sequência da proteína gSG7 de A.darlingi.
5.11WesternbloteensaiohemolíticocomIgGanti-gSG7deA.albimanus Para verificar se o anticorpo anti-gSG7 reconheceria a proteína
recombinanteeaproteínapresentenoEGS,foirealizadoWesternblotusandoo
anticorponadiluiçãode1:10000.ComomostradonaFigura39,oanticorpofoi
capaz de identificar as duas formas testadas da proteína, apresentando uma
únicabandaemcadacanaleta.
Figura 39: Western blot com anticorpo anti-gSG7 de A. albimanus. O IgGproduzidoemcoelhoreconheceutantoaproteínarecombinantecomoaproteínanativapresentenoextratodeglândulassalivares.

76
Diferentemente, o anticorpo anti-gSG7deA.albimanus não foi capazde
reconheceroutrasproteínassalivares,comoagSG7-2dessaespécieeagSG7de
A.darlingi,nemproteínasdoEGSdeA.aquasalisouA.aegypti(Figura40).
Figura 40: Western blot com IgG anti-gSG7 de A. albimanus. O anticorporeconheceu a gSG7 recombinante de A. albimanus (1) mas não reagiu contragSG7-2 da mesma espécie (2) nem com gSG7 de A. darlingi (3). O anticorpotambémnãoreconheceunenhumaproteínanoEGSdeA.aquasalis(4)ouEGSdeA.aegypti(5).
O IgG anti-gSG7 foi capaz de bloquear a atividade inibidora tanto da
proteínarecombinantequantodaproteínanoEGS.QuandotestadosobreoEGS,
oanticorpo foi capazdebloquearaatividadeestatisticamente significativanas
diluiçõesde1:10e1:100.Jáquandotestadocomaproteínarecombinante,oIgG
bloqueou de forma estatisticamente significativa até na diluição de 1:1000
(Figura 41). Quando apenas o anticorpo diluído 1:10 foi incubado com as
hemácias,nenhumahemólisefoiobservada,mostrandoqueoanticorpoporsisó
nãoatuacomohemolisina.
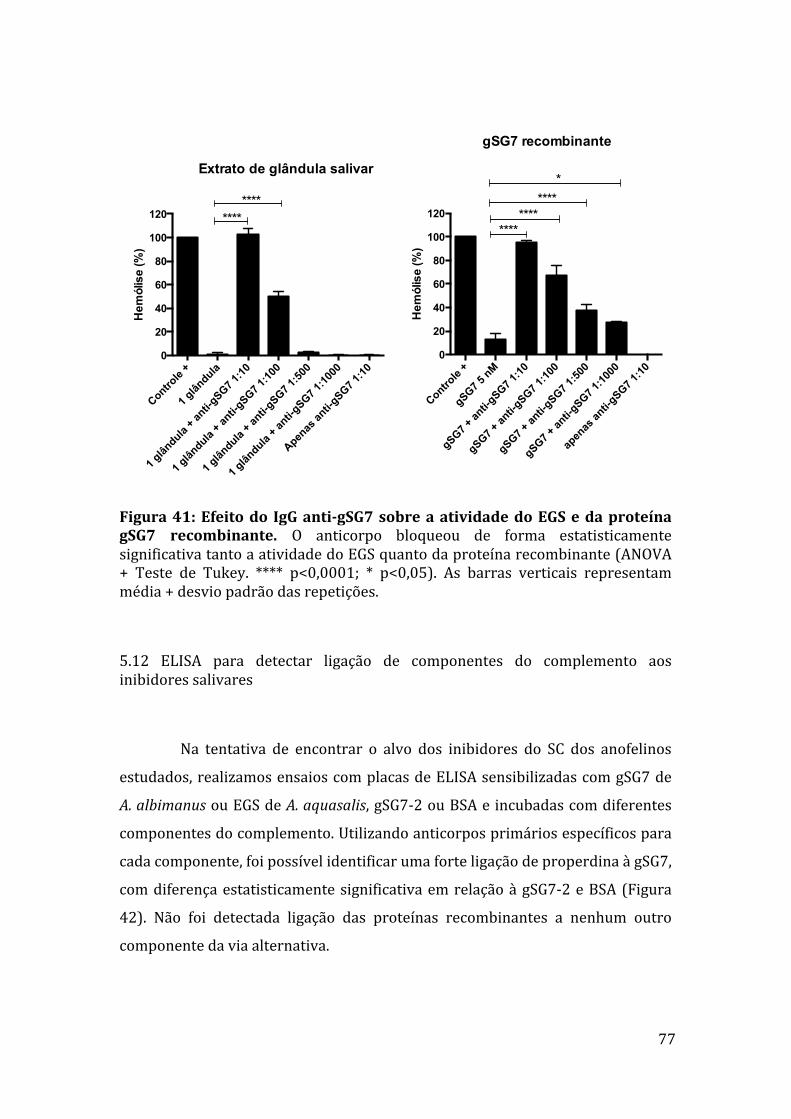
77
Figura41:Efeitodo IgGanti-gSG7sobreaatividadedoEGSedaproteínagSG7 recombinante. O anticorpo bloqueou de forma estatisticamentesignificativatantoaatividadedoEGSquantodaproteínarecombinante(ANOVA+ Teste de Tukey. **** p<0,0001; * p<0,05). As barras verticais representammédia+desviopadrãodasrepetições.
5.12 ELISA para detectar ligação de componentes do complemento aosinibidoressalivares
Na tentativa de encontrar o alvo dos inibidores do SC dos anofelinos
estudados,realizamosensaioscomplacasdeELISAsensibilizadascomgSG7de
A.albimanusouEGSdeA.aquasalis,gSG7-2ouBSAeincubadascomdiferentes
componentesdocomplemento.Utilizandoanticorposprimáriosespecíficospara
cadacomponente,foipossívelidentificarumaforteligaçãodeproperdinaàgSG7,
comdiferençaestatisticamentesignificativaemrelaçãoàgSG7-2eBSA(Figura
42). Não foi detectada ligação das proteínas recombinantes a nenhum outro
componentedaviaalternativa.
Controle
+
1 glân
dula
1 glân
dula + a
nti-gSG7 1
:10
1 glân
dula + a
nti-gSG7 1
:100
1 glân
dula + a
nti-gSG7 1
:500
1 glân
dula + a
nti-gSG7 1
:1000
Apenas
anti-
gSG7 1:10
0
20
40
60
80
100
120
Hem
ólis
e (%
)
********
Extrato de glândula salivar
Controle
+
gSG7 5 nM
gSG7 + an
ti-gSG7 1
:10
gSG7 + an
ti-gSG7 1
:100
gSG7 + an
ti-gSG7 1
:500
gSG7 + an
ti-gSG7 1
:1000
apen
as an
ti-gSG7 1
:100
20
40
60
80
100
120
Hem
ólis
e (%
)
gSG7 recombinante
*************
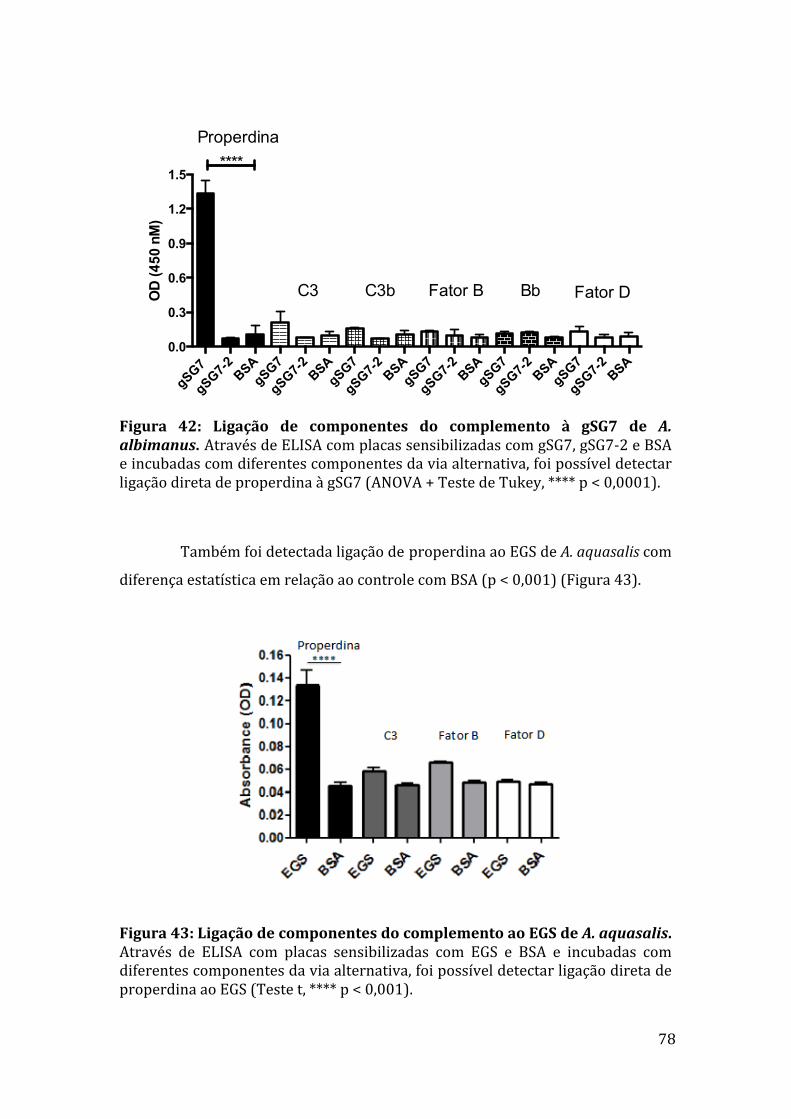
78
Figura 42: Ligação de componentes do complemento à gSG7 de A.albimanus.AtravésdeELISAcomplacassensibilizadascomgSG7,gSG7-2eBSAeincubadascomdiferentescomponentesdaviaalternativa,foipossíveldetectarligaçãodiretadeproperdinaàgSG7(ANOVA+TestedeTukey,****p<0,0001).
TambémfoidetectadaligaçãodeproperdinaaoEGSdeA.aquasaliscom
diferençaestatísticaemrelaçãoaocontrolecomBSA(p<0,001)(Figura43).
Figura43:LigaçãodecomponentesdocomplementoaoEGSdeA.aquasalis.Através de ELISA com placas sensibilizadas com EGS e BSA e incubadas comdiferentescomponentesdaviaalternativa,foipossíveldetectarligaçãodiretadeproperdinaaoEGS(Testet,****p<0,001).
gSG7
gSG7-2 BSAgSG7
gSG7-2 BSAgSG7
gSG7-2 BSAgSG7
gSG7-2 BSAgSG7
gSG7-2 BSAgSG7
gSG7-2 BSA0.0
0.3
0.6
0.9
1.2
1.5O
D (4
50 n
M)
Properdina
C3 C3b Fator B Bb Fator D
****
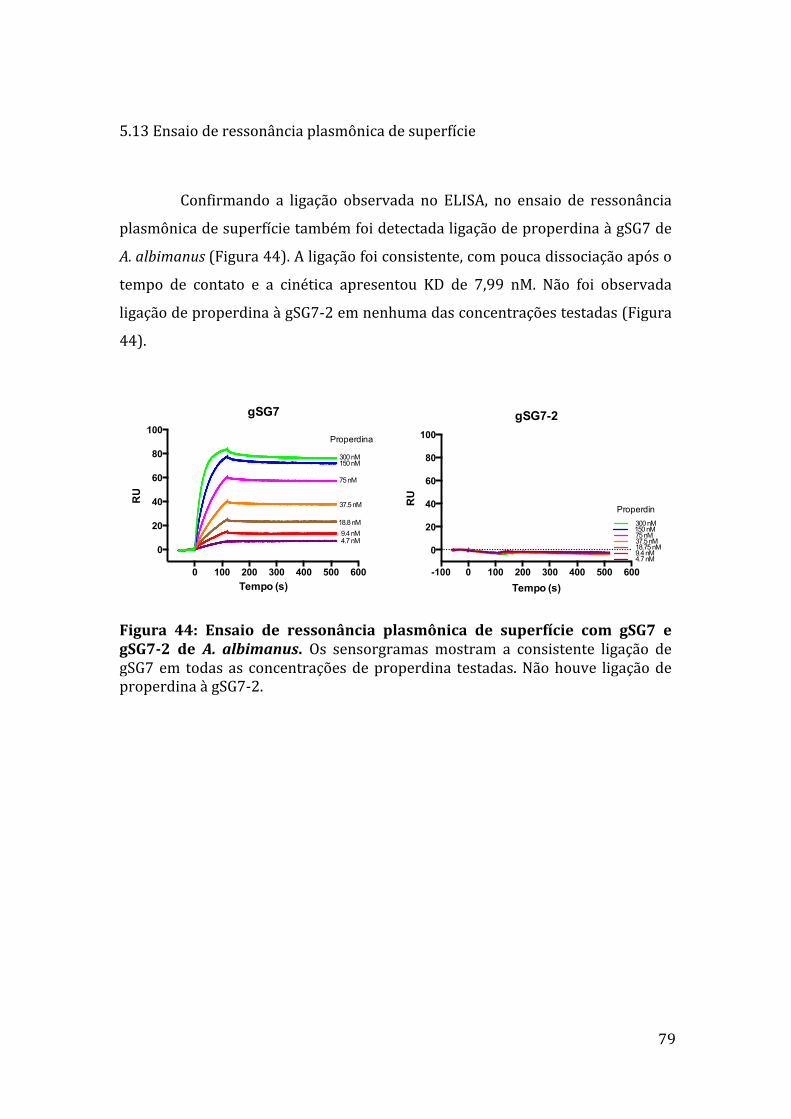
79
5.13Ensaioderessonânciaplasmônicadesuperfície
Confirmando a ligação observada no ELISA, no ensaio de ressonância
plasmônicadesuperfícietambémfoidetectadaligaçãodeproperdinaàgSG7de
A.albimanus(Figura44).Aligaçãofoiconsistente,compoucadissociaçãoapóso
tempo de contato e a cinética apresentou KD de 7,99 nM. Não foi observada
ligaçãodeproperdinaàgSG7-2emnenhumadasconcentraçõestestadas(Figura
44).
Figura 44: Ensaio de ressonância plasmônica de superfície com gSG7 egSG7-2 de A. albimanus. Os sensorgramasmostram a consistente ligação degSG7em todasas concentraçõesdeproperdina testadas.Nãohouve ligaçãodeproperdinaàgSG7-2.
0 100 200 300 400 500 600
0
20
40
60
80
100
Tempo (s)
RU
gSG7
300 nM150 nM
75 nM
37.5 nM
18.8 nM9.4 nM4.7 nM
Properdina
-100 0 100 200 300 400 500 600
0
20
40
60
80
100
Tempo (s)
RU
gSG7-2
4.7 nM9.4 nM18.75 nM37.5 nM75 nM150 nM300 nM
Properdin

80
6.Discussão
As proteínas do SC presente no sangue constituem uma importante
armadosistemaimuneinatocontrapatógenosinvasoresecélulasalteradasdo
hospedeiro.Umavezativado,oSCinduzorecrutamentoeativaçãodefagócitos,
alémdeformarporosnamembranacelular,levandoaodesequilíbrioosmóticoe
morte dos organismos estranhos. Para garantir sua sobrevivência, esses
organismosdesenvolverammecanismosdeescapedoSC,sendoasformasmais
comuns a secreção de inibidores de componentes específicos da cascata de
ativação e a adsorçãode reguladoresnegativosdoSCpresentesno sangueem
sua superfície. Mecanismos de evasão do SC já foram descritos em bactérias
(Potempa & Potempa 2012), fungos (Luo et al. 2013), vírus (Avirutnan et al.
2010),protozoários(Sacks&Sher2002),helmintos(Da’dara&Krautz-Peterson
2014)eatémesmoemcélulascancerígenashumanas(Okrojetal.2015).
Para inibição do SC, artrópodes hematófagos apresentam moléculas
específicas na saliva ouno intestino (Schroeder et al. 2009), pois esse sistema
permanece intacto por até uma hora no sangue ingerido e pode ser ativado
dentrodolúmenintestinaldos insetos(Simonetal.2013,Khattabetal.2015),
causandodanoaotratodigestivocasonãosejainativadoporinibidoressalivares
ouintestinais(Barrosetal.2009,Khattabetal.2015).Assim,sejaanívelsalivar
ouintestinal,a inibiçãodoSCpodeserconsideradaumprocessocrucialparaa
sobrevivênciadeartrópodesquesealimentamdesangue.
Neste estudo, a atividade anti-complemento foi pesquisada pela
primeiraveznoEGSdemosquitospertencentesaogêneroAnopheles.Aatividade
foiobservadanosEGSsdasespéciesneotropicaisA.albimanus,A.aquasaliseA.
freeborni.OEGSdeA.albimanusapresentoupotenteatividadeexclusivasobrea
viaalternativa,cominibiçãodeaproximadamente80%comquantidadedesaliva
referente a apenas uma glândula (Figura 7). Quando submetido a tratamentos
desnaturantes, comoaquecimentoedigestãoporproteinaseK,oEGSperdeua
atividade(Figura8),indicandoqueoinibidorédenaturezaproteica.OEGSdeA.
aquasalis, além de inibir a via alternativa de maneira dose-dependente,
apresentou leve inibição da via clássica, embora tenha sido necessário uma

81
grandequantidadedesaliva(referentea10glândulassalivares).OefeitodoEGS
de A. freeborni sobre outras vias do complemento não foi avaliado. As três
espéciesqueapresentaramatividadeanti-complementopossuemconcentrações
semelhantes de proteína por glândula salivar (aproximadamente 0,8 μg de
proteína/glândula) e, portanto, a diferença na intensidade da inibição da via
alternativapelosEGSsdastrêsespéciespodeserdevidoaoutrosmotivos,como
mecanismosdiferentesdeinibição,diferençanaexpressãodo(s)inibidor(es)ou
mesmodiferençasestruturaisdosmesmos.
AinibiçãodaviaalternativapeloEGSdeA.albimanuseA.aquasalisfoi
confirmada em ensaios de deposição em placas cobertas com agarose, onde
observamos a inibiçãodadeposiçãodeC3b, FatorB eproperdina (Figura11).
Por ensaiosdeWesternblot, demonstramosquenapresençadoEGSdasduas
espécies não há ativação dos componentes Fator B e C3 (Figura 13 e 14),
corroborandoos resultadosobservadosnos ensaiosdedeposição.Essesdados
indicam que os inibidores atuam logo no início da cascata de ativação da via
alternativa.QuandooWesternblotfoirealizadoutilizandoC3b,FatorBeFatorD
purificados,oEGSdeA.albimanusnãoinibiuaativaçãodocomponenteFatorB
(Figura15),indicandoqueoinibidornãoatuadiretamenteemnenhumdostrês
componentesutilizados.
A via alternativa possui grande importância na ativação do CS pois
independedemoléculasativadorase,porser iniciadaporhidróliseespontânea
do componente C3, está constantemente sendo ativada em baixos níveis na
circulação (Dunkelberger& Song 2010). Alémdisso, ela amplifica em cerca de
80%aatividadedaviaclássica(produçãodeC5aeCAM),poisoC3bgeradopor
estaviapode formarasC3- eC5-convertasesdavia alternativa, ativandomais
moléculas de C3 e C5, respectivamente, e consequentemente, formando mais
CAM (Harboe et al. 2004). Apesar de não observado em nossos resultados
(apenas com alta concentração de saliva de A. aquasalis), é possível que os
inibidoressalivaresdaviaalternativaatuem indiretamentesobreasatividades
das vias clássica e das lectinas no local da picada e dentro do intestino do
mosquito,diminuindoaatividadedessasvias.

82
Curiosamente, os EGSs de anofelinos do Velho Mundo (A. dirus, A.
gambiaeeA.stephensi) não apresentaramatividade anti-complemento,mesmo
com altas concentrações de proteínas salivares (Figura 7). Recentemente,
Khattabetal(2015)demonstraramqueA.gambiaeeA.stephensiligamFatorH,
um regulador negativo do SC, ao seu epitélio intestinal, inativando o SC. Isso
explicariaporqueessesmosquitosnãoapresentaminibidoressalivares,umavez
que os inibidores intestinais seriam suficientes para a proteção do trato
digestivo desses insetos.De fato, inibidores do sistema complemento têm sido
descritosnointestinodebarbeirosTriatomainfestans,T.brasiliensiseRhodnius
prolixus (Barros et al. 2009), do flebotomíneo Lutzomyia longipalpis (Mendes-
Sousa et al. 2013), damosca tsé-tséGlossinamorsitans (Ooi et al. 2015) e do
ácaroSarcoptesscabiei(Bergströmetal,2009).Embarbeiroseflebotomíneos,já
foramdescritosinibidoresdocomplementotantonasalivaquantonointestino,
evidenciando a importância desses inibidores para a hematofagia e
sobrevivência desses vetores. É possível que existam também inibidores do
complemento no intestino dos anofelinos do Novo Mundo, atuando
sinergicamentecomosinibidoressalivaresnaproteçãodotratodigestivodesses
mosquitos.
Nosensaioshemolíticosparaativaçãodaviaalternativa,usamos25µl
deSHNdiluído1:20.Portanto,emcadatuboutilizadohavia1,25µldeSHN.EGS
referente a uma glândula salivar deA. albimanus foi suficiente para inibir em
torno de 80% a atividade dessa via (Figura 7). Considerando que: a) um
anofelino ingereemtornode2,5µldesangue(Jeffery1956,Phasomkusolsilet
al. 2015) e que o hematócrito humano permanece em torno de 45%, logo o
volume de soro ingerido pelomosquito é de aproximadamente 1,4µl; e b) os
anofelinos injetam na pele do hospedeiro até metade do conteúdo das suas
glândulas salivares (portanto, o conteúdo de uma glândula) (Golenda et al.
1995), a quantidade de saliva de A. albimanus inoculada seria suficiente para
inibir o SC ingerido no respasto sanguíneo. Já A. aquasalis e A. freeborni
necessitaram de quantidades maiores de proteínas salivares para inibir
significativamente o SC (em torno de 10 glândulas). Esses dados reforçam a
hipótese da existência de inibidores intestinais nos anofelinos neotropicais,

83
sendonecessáriosparaagirjuntamentecomosinibidoressalivaresnaproteção
dointestinodessasespéciescontraaativaçãodoCAM.
O inibidor salivar de A. albimanus foi purificado e identificado por
espectrometria de massa como pertencente à família gSG7 (gambiae Salivary
Gene7),queéamplamentedistribuídanasglândulassalivaresdeanofelinos.O
gene codificador dessa proteína foi descrito inicialmente no sialoma de A.
gambiae, sendo expresso exclusivamente nas glândulas salivares de formas
adultas do mosquito (Lanfrancotti et al. 2002). Posteriormente, esse gene foi
tambémdescritoemA.stephensi(Valenzuelaetal.2003),A.darlingi(Calvoetal.
2009),A.funestus(Calvoetal.2007)eA.sinensis(Zhouetal.2014).Essegrupo
de proteínas possui pouca similaridade de sequência com outros grupos de
proteínas de outros organismos, apresentam peptídeo sinal, não possuem
domíniosconhecidoseaindanãopossuiestruturacaracterizada.
A gSG7 recombinante de A. albimanus apresentou atividade anti-
complemento de forma semelhante ao EGS dessa espécie em todos os
experimentostestados(ensaiohemolítico,ensaiosdedeposiçãoeWesternblot),
confirmandoserrealmenteoinibidordocomplementopresentenasalivadeA.
albimanus.AssequênciasdasgSG7sdeanofelinosdoVelhoMundoapresentam
aproximadamente 45% de identidade de aminoácidos com a gSG7 de A.
albimanus. As diferenças nas sequências dos aminoácidos destas proteínas
devemocorrernosítiodeaçãodagSG7dosinsetosdoNovoMundo,justificando
aausênciadeatividadeanti-complementonosanofelinosdoVelhoMundo. Jáa
mesmaproteínamaduradeA.darlingiapresenta77%deidentidadecomadeA.
albimanus e também apresenta atividade anti-complemento. Esses dados
sugerem que a atividade anti-complemento é característica das gSG7s de
mosquitosdoNovoMundoeevoluíramapósadivergênciadeformasdoNovoe
VelhoMundos.
No EGS deA. aquasalisobservamos uma banda namesma altura das
gSG7sdeA.albimanuseA.darlingi(Figura37).Oresultadodaespectrometriade
massa indicou um peptídeo com 100% de similaridade com um fragmento da
sequência de aminoácidos da gSG7 de A. darlingi. Considerando que as três
espécies pertencem ao mesmo subgênero (Nyssorhynchus) e são

84
filogeneticamentemaispróximas,essesresultadossugeremaexistênciadeuma
proteína da família gSG7 no EGS deA.aquasalis, possivelmente o seu inibidor
salivar do SC. O transcriptoma total de larvas L3, L4 e fêmeas dessa espécie
alimentadascomaçúcarousanguefoirecentementepublicado(Costa-da-Silvaet
al. 2014), com algumas sequências depositadas nos bancos de dados virtuais.
Entretanto, nenhuma sequência de A. aquasalis apresentou similaridade
significativa com o gene ou a proteína gSG7 de outros anofelinos. RNAm de
glândulas salivares dessa espécie foram extraídos e atualmente estão sendo
sequenciados e analisados para a anotação do sialotranscriptoma. Quando o
mesmoestiverfinalizado,poderemosinvestigarcommaiorprecisãoapresença
dealgumasequênciacomsimilaridadecomagSG7deA.albimanuseA.darlingi.
Isawa et al (2007) produziram a gSG7 recombinante de A. stephensi,
denominadaanophensina,edemonstraramqueamesmaéinibidoradosistema
cinina-calicreína, ligando-seemFatorXII e cininogêniodealtopesomolecular,
bloqueando a produção de bradicinina, importante mediador de resposta
inflamatória. Em nosso estudo, outras atividades da gSG7 de A. albimanus,
incluindoaatividadeacimadescrita,nãoforaminvestigadasmasépossívelque
ela também apresente outras finalidades além de inibir o complemento.
Considerando que esta espécie de mosquito apresenta dezesseis proteínas no
seuEGSequeapenasnovedelassãosecretadas(Fontaineetal.2012),nãoseria
de se admirar que suas moléculas salivares apresentemmais de uma função,
agindosinergicamentedemodoafavorecerahematofagia.
Por ensaios de ELISA, demonstramos a ligação de properdina à gSG7
recombinante de A. albimanus e ao EGS de A. aquasalis utilizados para
sensibilizar a placa. Essa ligação foi confirmada para o inibidor da primeira
espécieatravésdeensaiode ressonânciaplasmônicadesuperfície (Figura44),
ondeobservamos forte ligaçãodaproperdinaao inibidor.Este componentedo
SCéumaglicoproteínade53kDaformadapor7domíniosdetrombospondina,
encontrada comumente na forma de oligômeros, principalmente dímeros,
trimerosetetrâmeroscirculantesnoplasmacomconcentraçãode5a20µg/mL
(Pangburn 1989). A properdina liga-se em C3bB e C3bBb, estabilizando a C3-
convertase da via alternativa, aumentando em até 10 vezes sua vida média,

85
permitindoaativaçãodemuitomaismoléculasdeC3(Fearon&Austen1975).
Além disso, protege esse complexo enzimático de reguladores negativos como
FatorH,queinduzadissociaçãodeFatorBdeC3beatuacomoreceptordeFator
I,queenzimaticamenteinativaC3bemiC3b(C3binativado)(Medicusetal.1976,
Hourcade2006,Torreiraetal.2009).Defato,jáfoidemonstradoqueanticorpos
IgG específicos contra properdina são capazes de inibir a atividade lítica e a
deposição de componentes da via alternativa, ressaltando a importância desse
componente para a ativação dessa via (Gupta-Bansal et al. 2000, Pauly et al.
2014). Essa ligação explica porque o inibidor salivar de A. albimanus atua
somente sobre a via alternativa e porquenosWesternblots realizados apenas
comoscomponentesC3b,FatorBeFatorDpurificados(semproperdina)nãofoi
observadanenhuma inibiçãodaativaçãodocomponenteFatorB (Figuras15e
35).
Proteínas salivares de artrópodes hematófagos com atividade anti-
complemento foram identificadas na saliva dos carrapatos Ixodes scapularis
(Valenzuela et al., 2000) e I. ricinus (Tyson et al. 2008), vetores daDoença de
LymenaAméricadoNorteeEuropa,respectivamente.NasalivadeI.scapularis
foramdescritas a ISACe a Salp20, proteínasque inibemexclusivamente a via
alternativa. Duas famílias de proteínas homólogas anti-via alternativa foram
encontradas na saliva de I. ricinus: as IRACs (Daix et al. 2007) e as IxACs
(Couvreur et al. 2008). Salp20 e as IxACs inibem a via alternativa ligando em
properdina(Tysonetal.2008;Couvreuretal.2008).Ainibiçãodaviaalternativa
já foidescrita tambémnasalivadeL.longipalpis (Cavalcanteetal.,2003).Vale
(2011)caracterizouparcialmenteessaatividade,demonstrandoqueoEGSatua
noiníciodacascata inibindoaativaçãodoFatorB,masquenãoage inibindoa
atividadeenzimáticadocomponenteFatorD.Recentemente,descobrimosqueo
inibidor salivar desse flebotomíneo trata-se da proteína Lufaxin, inicialmente
descrita como o anti-coagulante salivar de L. longipalpis (Collin et al. 2012).
Resultados preliminares demonstraram que essa proteína recombinante
bloqueia exclusivamente a ativação da via alternativa através da ligação em
properdina(manuscritoempreparação).

86
Porligarememproperdina,essasproteínassalivaresinibemsualigação
na C3-convertase da via alternativa, desestabilizando o complexo enzimático,
acelerandosuadissociação,alémdepermitiraligaçãodereguladoresnegativos
da C3-convertase, como Fator I, inativando a cascata dessa via. Baseado em
nossos resultados,podemosassumirqueos inibidoressalivaresdosanofelinos
apresentamummecanismodeaçãosimilar.Entretanto,agSG7inibeadeposição
deFatorBeoutroscomponentesemplacascobertascomagarosemas,diferente
de ISAC e Salp20 (Valenzuela et al., 2000; Tyson et al., 2007), não remove
componentes previamente depositados (Figura 34). Assim, podemos concluir
queela é capazdebloquear a formaçãodaC3-convertase,masnãoatua sobre
complexos pré-formados, assim como tem sido descrito para o EGS de L.
longipalpis (Vale, 2011) e as SMIPPs, inibidores do complemento presentes no
intestinodeSarcoptesscabiei,que também inibemaviaalternativa ligandoem
properdina(Bergstrometal.,2009).
JáfoidemonstradoqueasalivadeL.longipalpisfavoreceainfecçãopor
Leishmania no hospedeiro vertebrado, exacerbando o tamanho e a carga
parasitárianalesãocutânea(Titus&Ribeiro1988)eque,apesardeapresentar
mecanismosprópriosdeevasãodocomplemento(Späthetal.2003),a inibição
do SC pela saliva do vetor ajuda amanter a viabilidade dos parasitos quando
expostos ao soro humano (Nascimento 2011). Também já se sabe que TSLPI,
umaproteínasalivardocarrapatoI.scapularisqueinibeoSC,éimportantepara
atransmissãodaespiroquetacausadoradeDoençadeLyme,Borreliaburgdorferi
(Schuijt et al. 2011b). O silenciamento do gene dessa proteína ou anticorpos
contra ela provocaram uma diminuição da sobrevivência da bactéria tanto no
hospedeiro vertebrado quanto no intestino do carrapato (Schuijt et al. 2011).
Recentemente, foi demonstrado que merozoítos e gametas de P. falciparum
captamFatorHpresentenosanguecirculanteenosangue ingeridopelovetor,
respectivamente,paraevadirdalisepelocomplemento(Simonetal.2013,Rosa
et al. 2015). Entretanto, não há relatos de como esporozoítos, as formas
infectantes inoculadas pelo vetor, evadem desse sistema. Essas formas são
depositadasnapeledohospedeirodurantea fasede sondagemdomosquitoe
apósalgunsminutosinvademcapilarescutâneos,atingindoacorrentesanguínea

87
(Aminoetal.2006),permanecendoexpostosaoSCdurantetodoesseprocesso.
Épossíveleprovávelqueesta formaevolutivadoparasitotenhaseuspróprios
mecanismosdeescapedoSC,masapresençadeinibidoresnasalivadosvetores
doNovoMundodeveajudarnosucessodainfecção,tornandomenosinóspitoo
ambientedeinoculaçãodosesporozoítos.
Neste trabalhodemonstramosqueanticorpos IgGespecíficoscontraa
gSG7recombinantedeA.albimanusreconhecemtantooinibidornativopresente
no EGS quanto o recombinante (Figura 39), confirmando serem a mesma
proteína. Entretanto, os anticorpos não reconheceram outras proteínas dessa
mesmaespécieoudeoutrasespéciesdemosquitos,comoaproteínagSG7deA.
darlingiouproteínasdoEGSdeA.aquasalisouA.aegypti(Figura40),sugerindo
queosítioimunogênicodaproteínaéespecíficoparacadaespécie.Dessaforma,
se proteínas dessa família forem imunogênicas e estimularem produção de
anticorpos em indivíduos expostos aos mosquitos em áreas endêmicas para
malária, elas poderiam ser bons candidatos para marcadores de exposição
espécie-específicos,permitindoa identificaçãodeespéciescommaiorpotencial
vetorial, especialmente em áreas onde diferentes espécies se sobrepõem.
Adicionalmente,adetecçãodeanticorposanti-salivadovetorpoderiaserusada
para indicar infecção por Plasmodium, além de atuar como marcador da
severidadedadoença(Andradeetal.2009).
Atualmente, muitos trabalhos têm focado na pesquisa para o
desenvolvimentodevacinasdebloqueiodetransmissão(VBT)contraparasitos
transmitidos por artrópodes, cujo objetivo é impedir o desenvolvimento do
patógenonovetoresesuasubsequentetransmissão(Carter2001).Antígenosde
estágios sexuados do Plasmodium (encontrados no intestino do vetor) e
moléculassalivareseintestinaisdomosquitotêmdemonstradopotencialvacinal
(Carter2001,Lavazec&Bourgouin2008).Nocasodasleishmanioses,estudosde
VBTcomantígenossalivaresde flebotomíneosdoVelhoedoNovoMundotêm
sidodescritos.Gomesetal.(2008)verificaramqueaimunidadeaumaproteína
salivarespecíficadeL.longipalpis,aLJM19(queéinibidordaviaclássicadoSC),
propiciouproteçãosignificativacontraaleishmaniosevisceralfatalemhamster.
Recentemente,Oliveiraetal(2015)demonstraramqueaimunizaçãodemacacos

88
do gênero Rhesus com a proteína salivar PdSP-15 de Phlebotomus duboscqi é
capaz de proteger esses animais contra o surgimento de leishmaniose cutânea
provocadaporL.major.EmestudoscomcarrapatosI.scapularis, tambémjáfoi
descrito que a imunização de camundongos com a proteína inibidora do
complemento TSLPI levou a diminuição da carga parasitária em diferentes
tecidos do hospedeiro vertebrado (Schuijt et al. 2011). Considerando a
importância da evasão do SC por esporozoítos de Plasmodium nos primeiros
momentos da infecção, se um anofelino infectado inocular o parasito em um
indivíduo imunizado com gSG7 e, portanto, com este inibidor inativado por
anticorpos,osesporozoítosinoculadosficarãomaissusceptíveisaoataquedoSC,
resultandoemdiminuiçãodaquantidadedeparasitos viáveisno sangue.Além
disso, os anticorpos poderiam bloquear o inibidor no intestino do mosquito,
deixando o complemento ativo neste órgão, podendo causar danos ao epitélio
intestinal do mesmo. Caso estas hipóteses sejam verdadeiras, a gSG7 teria
potencial como um componente de vacina de bloqueio de transmissão para
malária.
Atualmente, doenças auto-imunes mediadas pelo complemento têm
recebidomuita atenção devido à sua severidade, como a SíndromeHemolítica
Urêmicaatípica(SHUa)eaHemoglobinúriaParoxísticaNoturna(HPN),doenças
decarátercrônicoquelevamaoóbitoempoucosanossenãotratadas(Bottoet
al. 2009, Ricklin & Lambris 2013, DeZern & Brodsky 2015). A causa dessas
gravesdoençaséadesregulaçãodaviaalternativadoSCpordiferentesrazões,
como ausência ou produção insuficiente de reguladores de membrana, como
CD55eCD59, emutaçõesemgenes codificadoresde reguladoresdoSC (como
FatorHeFatorI),levandoaproduçãodeformasalteradasdessasproteínasque
nãoapresentamadevidafunção(Roumeninaetal.2011).Essasalteraçõeslevam
àativaçãoinapropriadadessavia,queatacacélulaspróprias,provocandodanos
em tecidos normais do hospedeiro, principalmente eritrócitos. Atualmente, a
drogadisponívelparao tratamentodaSHUaedaHPNéumanticorpoanti-C5
(Soliris®),quebloqueiaaativaçãodocomponenteC5,prevenindoaativaçãodo
CAMnasuperfíciedecélulasnormais(Brodskyetal.2008).Entretanto,comoC5
éumcomponentecomumdastrêsviasdeativaçãodocomplemento,suainibição

89
levaàsusceptibilidadedohospedeiroapatógenosinvasores,levandoainfecções
respiratóriasouurinárias(Brodskyetal.2008).Assim,agSG7deA.albimanus,
umaproteínadebaixopesomolecularqueligaemproperdinaeinibeapenasa
via alternativa, poderia ser um candidato promissor para testes de uma nova
droga para doenças auto-imunes mediadas pelo sistema complemento,
diminuindo o risco de infecções durante o tratamento, uma vez que as vias
clássicaedaslectinasaindaestariamativascontrapatógenosinvasores.
Tendo em vista os aspectos levantados nesta discussão, o
descobrimentodeumamoléculadasalivadeanofelinoscapazdeinibirosistema
complementoconstituiumgrandepassonoestudodainteraçãovetor-parasito-
hospedeiro vertebrado. Além disso, nosso trabalho abre portas para novos
estudosvoltadosparaafisiologiaeevoluçãodeinsetoshematófagos,vacinasde
bloqueio de transmissão, marcadores de exposição e o desenvolvimento de
drogas para síndromes graves e fatais causadas pela desregulação do sistema
complemento.

90
7.Conclusões
• EGSdeA.albimanus,A.aquasaliseA.freeborniinibemaviaalternativado
sistemacomplementohumano;
• O inibidor salivar de A. albimanus, A. darlingi e A. aquasalis pertence à
famíliadeproteínasgSG7;
• AnticorposIgGcontraagSG7recombinantedeA.albimanusreconhecem
ebloqueiamosinibidoresrecombinanteenativodesseanofelino;
• A gSG7 atua ligando-se em properdina, o regulador positivo da via
alternativaimpedindoqueelaestabilizeaC3convertasedessavia.

91
8.Referênciasbibliográficas
AlvarengaPH,FrancischettiIMB,CalvoE,Sá-NunesA,RibeiroJMC,AndersenJF2010.Thefunctionandthree-dimensionalstructureofathromboxaneA2/cysteinylleukotriene-bindingproteinfromthesalivaofamosquitovectorofthemalariaparasite.PLoSBiol.8:e1000547.
AlvesFB,DurlacherRR,MenéndezR,KreigerH,SilvaLPDa,CamargoEP2002.HighprevalenceofasymptomaticPlasmodiumvivaxandPlasmodiumfalciparuminfectionsinnativaAmazonianpopulations.AmJTropMedHyg66:641–648.
AminoR,ThibergeS,MartinB,CelliS,ShorteS,FrischknechtF,MénardR2006.QuantitativeimagingofPlasmodiumtransmissionfrommosquitotomammal.Nat.Med.12:220–224.
AndradeBB,RochaBC,Reis-FilhoA,CamargoLMA,TadeiWP,MoreiraLA,BarralA,Barral-NettoM2009.Anti-AnophelesdarlingisalivaantibodiesasmarkerofPlasmodiumvivaxinfectionandclinicalimmunityintheBrazilianAmazon.Malar.J.8:121.
AvirutnanP,FuchsA,HauhartRE,SomnukeP,YounS,DiamondMS,AtkinsonJP2010.AntagonismofthecomplementcomponentC4byflavivirusnonstructuralproteinNS1.J.Exp.Med.207:793–806.
BahiaAC,KubotaMS,TemponeAJ,AraújoHRC,GuedesBAM,OrfanóAS,TadeiWP,Ríos-VelásquezCM,HanYS,SecundinoNFC,Barillas-MuryC,PimentaPFP,Traub-CseköYM2011.TheJAK-STATpathwaycontrolsPlasmodiumvivaxloadinearlystagesofAnophelesaquasalisinfection.PLoSNegl.Trop.Dis.5:e1317.
BarrosVC,AssumpçãoJG,CadeteAM,SantosVC,CavalcanteRR,AraújoRN,PereiraMH,GontijoNF2009.Theroleofsalivaryandintestinalcomplementsysteminhibitorsinthemidgutprotectionoftriatominesandmosquitoes.PLoSOne4:e6047.
BergströmFC,ReynoldsS,JohnstoneM,PikeRN,BuckleAM,KempDJ,FischerK,BlomAM2009.Scabiesmiteinactivatedserineproteaseparalogsinhibitthehumancomplementsystem.J.Immunol.182:7809–7817.
BillingsleyPF1990.TheMidgutUltrastructureofHematophagousInsects.Annu.Rev.Entomol.35:219–248.
BillkerO,LindoV,PanicoM,EtienneaE,PaxtonT,Della,RogersM,SindenRE,MorrisHR1998.Identificationofxanthurenicacidastheputativeinducerofmalariadevelopmentinthemosquito.Nature392:289–292.
BILLKERO,SHAWMK,MARGOSG,SINDENRE1997.Therolesoftemperature,pHandmosquitofactorsastriggersofmaleandfemalegametogenesisofPlasmodiumbergheiinvitro.Parasitology115:1–7.
BoissonB,ClaudeJ,ChoumetV,MartinE,XuJ,VernickK,BourgouinC2006.

92
GenesilencinginmosquitosalivaryglandsbyRNAi.580:1988–1992.
BottoM,KirschfinkM,MacorP,PickeringMC,WürznerR,TedescoF2009.Complementinhumandiseases:Lessonsfromcomplementdeficiencies.Mol.Immunol.46:2774–2783.
BousemaT,OkellL,ShekalagheS,GriffinJT,OmarS,SawaP,SutherlandC,SauerweinR,GhaniAC,DrakeleyC2010.RevisitingthecirculationtimeofPlasmodiumfalciparumgametocytes:moleculardetectionmethodstoestimatethedurationofgametocytecarriageandtheeffectofgametocytocidaldrugs.Malar.J.9:136.
BradfordMM1976.Arapidandsensitivemethodforthequantitationofmicrogramquantitiesofproteinutilizingtheprincipleofprotein-dyebinding.Anal.Biochem.72:248–254.
BreelandSG1972.STUDIESONTHEECOLOGYOFANOPHELESALBIMANUS.21:751–754.
BrodskyRA,YoungNS,AntonioliE,RisitanoAM,SchrezenmeierH,SchubertJ,GayaA,CoyleL,CastroCde,FuC-L,MaciejewskiJP,BesslerM,KroonH-A,RotherRP,HillmenP2008.Multicenterphase3studyofthecomplementinhibitoreculizumabforthetreatmentofpatientswithparoxysmalnocturnalhemoglobinuria.Blood111:1840–1847.
CalvoE,DaoA,PhamVM,RibeiroJMC2007.AninsightintothesialomeofAnophelesfunestusrevealsanemergingpatterninanophelinesalivaryproteinfamilies.InsectBiochem.Mol.Biol.37:164–175.
CalvoE,PhamVM,MarinottiO,AndersenJF,RibeiroJMC2009.ThesalivaryglandtranscriptomeoftheneotropicalmalariavectorAnophelesdarlingirevealsacceleratedevolutionofgenesrelevanttohematophagy.BMCGenomics10:57.
CarterR2001.Transmissionblockingmalariavaccines.Vaccine19:2309–2314.
CarvalhoSCGde,MartinsJuniorAdeJ,LimaJBP,ValleD2002.TemperatureinfluenceonembryonicdevelopmentofAnophelesalbitarsisandAnophelesaquasalis.Mem.Inst.OswaldoCruz97:1117–1120.
CavalcanteRR,PereiraMH,GontijoNF2003.Anti-complementactivityinthesalivaofphlebotominesandfliesandotherhaematophagousinsects.Parasitology127:87–93.
CollinN,AssumpçãoTCF,MizuriniDM,GilmoreDC,Dutra-OliveiraA,KotsyfakisM,Sá-NunesA,TeixeiraC,RibeiroJMC,MonteiroRQ,ValenzuelaJG,FrancischettiIMB2012.Lufaxin,anovelfactorXainhibitorfromthesalivaryglandofthesandflylutzomyialongipalpisblocksprotease-activatedreceptor2activationandinhibitsinflammationandthrombosisinvivo.Arterioscler.Thromb.Vasc.Biol.32:2185–2196.
Costa-da-SilvaAL,MarinottiO,RibeiroJMC,SilvaMCP,LopesAR,BarrosMS,Sá-NunesA,KojinBB,CarvalhoE,SuesdekL,Silva-NetoMAC,JamesAa.,CapurroML2014.TranscriptomeSequencingandDevelopmental

93
RegulationofGeneExpressioninAnophelesaquasalis.PLoSNegl.Trop.Dis.8:1–11.
CouvreurB,BeaufaysJ,CharonC,LahayeK,GensaleF,DenisV,CharloteauxB,DecremY,PrévôtP-P,BrossardM,VanhammeL,GodfroidE2008.VariabilityandactionmechanismofafamilyofanticomplementproteinsinIxodesricinus.PLoSOne3:e1400.
Cox-singhJ,SinghB2010.Knowlesimalaria :newlyemergentandofpublichealthimportance ?TrendsParasitol.24:406–410.
Da’daraAa.,Krautz-PetersonG2014.NewinsightsintothereactionofSchistosomamansonicercariatothehumancomplementsystem.Parasitol.Res.113:3685–3696.
DaixV,SchroederH,PraetN,GeorginJ-P,ChiappinoI,GilletL,FaysKde,DecremY,LeboulleG,GodfroidE,Bollena,PastoretP-P,GernL,SharpPM,Vanderplasschena2007.IxodesticksbelongingtotheIxodesricinuscomplexencodeafamilyofanticomplementproteins.InsectMol.Biol.16:155–166.
DasS,RadtkeA,ChoiY-J,MendesAM,ValenzuelaJG,DimopoulosG2010.TranscriptomicandfunctionalanalysisoftheAnophelesgambiaesalivaryglandinrelationtobloodfeeding.BMCGenomics11:566.
DemeureCE,BrahimiK,HaciniF,MarchandF,PéronetR,HuerreM,NicolasJ,BreyP,StP2005.Anophelesmosquitobitesactivatecutaneousmastcellsleadingtoalocalinflammatoryresponseandlymphnodehyperplasia.J.Immunol.174:3932–3940.
DepinayN,HaciniF,BeghdadiW,PeronetR,MeS2006.MastCell-dependentdown-regulationofantigen-specificimmuneresponsesbymosquitobites.J.Immunol.:4141–4146.
DeZernAE,BrodskyRa.2015.ParoxysmalNocturnalHemoglobinuria.Hematol.Oncol.Clin.NorthAm.29:479–494.
DiScipioRG,SchraufstatterIU2007.Theroleofthecomplementanaphylatoxinsintherecruitmentofeosinophils.Int.Immunopharmacol.7:1909–1923.
DonovanMJ,MessmoreAS,ScraffordDa.,SacksDL,KamhawiS,McDowellMA2007.Uninfectedmosquitobitesconferprotectionagainstinfectionwithmalariaparasites.Infect.Immun.75:2523–2530.
DunkelbergerJR,SongW-C2010.Complementanditsroleininnateandadaptiveimmuneresponses.CellRes.20:34–50.
FaranME1980.ArevisionofthealbimanussectionofthesubgenusNyssorhyncysofAnopheles.15.
FearonDT,AustenKF1975.Properdin:bindingtoC3bandstabilizationoftheC3b-dependentC3convertase.J.Exp.Med.142:856–863.
FontaineA,FusaïT,BriolantS,BuffetS,VillardC,BaudeletE,PophillatM,GranjeaudS,RogierC,AlmerasL2012.Anophelessalivaryglandproteomes

94
frommajormalariavectors.BMCGenomics13:614.
FrancischettiIMB,MaD,AndersenJF,RibeiroJMC2014.Evidenceforalectinspecificforsulfatedglycansinthesalivaryglandofthemalariavector,Anophelesgambiae.PLoSOne9:e107295.
GilLHS,TadaMS,KatsuragawaTH,RibollaPEM,SilvaLHPda2007.UrbanandsuburbanmalariainRondônia(BrazilianWesternAmazon)II:perennialtransmissionswithhighanophelinedensitiesareassociatedwithhumanenvironmentalchanges.Mem.Inst.OswaldoCruz102:271–276.
GolendaCF,KleinT,ColemanR,BurgeR,WardRA,SeeleyDC1995.DepletionofTotalSalivaryGlandProteininBlood-FedAnophelesMosquitoes.J.Med.Entomol.32:300–305.
GomesR,TeixeiraC,TeixeiraMJ,OliveiraF,MenezesMJ,SilvaC,OliveiraCIde,MirandaJC,ElnaiemD-E,KamhawiS,ValenzuelaJG,BrodskynCI2008.Immunitytoasalivaryproteinofasandflyvectorprotectsagainstthefataloutcomeofvisceralleishmaniasisinahamstermodel.Proc.Natl.Acad.Sci.U.S.A.105:7845–7850.
Gupta-BansalR,ParentJB,BrundenKR2000.Inhibitionofcomplementalternativepathwayfunctionwithanti-properdinmonoclonalantibodies.Mol.Immunol.37:191–201.
HarbachRE2004.TheclassificationofgenusAnopheles(Diptera:Culicidae):aworkinghypothesisofphylogeneticrelationships.Bull.Entomol.Res.94:537–553.
HarboeM,UlvundG,VienL,FungM,MollnesTE2004.Thequantitativeroleofalternativepathwayamplificationinclassicalpathwayinducedterminalcomplementactivation.Clin.Exp.Immunol.138:439–446.
HourcadeDE2006.TheroleofproperdinintheassemblyofthealternativepathwayC3convertasesofcomplement.J.Biol.Chem.281:2128–2132.
IsawaH,OritoY,IwanagaS,JingushiN,MoritaA,ChinzeiY,YudaM2007.Identificationandcharacterizationofanewkallikrein-kininsysteminhibitorfromthesalivaryglandsofthemalariavectormosquitoAnophelesstephensi.InsectBiochem.Mol.Biol.37:466–477.
JariyapanN,ChoochoteW2007.Salivaryglandproteinsofthehumanmalariavector,AnophelesdirusB(Diptera:Culicidae).Rev.doInst.…49:5–10.
JefferyGM1956.BloodmealvolumeinAnophelesquadrimaculatus,A.albimanusandAedesaegypti.Exp.Parasitol.5:371–375.
JoslingGA,LlinásM2015.SexualdevelopmentinPlasmodiumparasites:knowingwhenit’stimetocommit.Nat.Rev.Microbiol.13:573–587.
KhattabA,BarrosoM,MiettinenT,MeriS2015.AnophelesMidgutEpitheliumEvadesHumanComplementActivitybyCapturingFactorHfromtheBloodMeal.PLoSNegl.Trop.Dis.9:e0003513.
LacerdaMV,MourãoMP,AlexandreMA,SiqueiraAM,MagalhãesBM,Martinez-

95
EspinosaFE,SantanaFilhoFS,BrasilP,VenturaAM,TadaMS,CoutoVS,SilvaAR,SilvaRS,AlecrimMG2012.UnderstandingtheclinicalspectrumofcomplicatedPlasmodiumvivaxmalaria:asystematicreviewonthecontributionsoftheBrazilianliterature.Malar.J.11:12.
LanfrancottiA,LombardoF,SantolamazzaF,VeneriM,CastrignanòT,ColuzziM,ArcàB2002.NovelcDNAsencodingsalivaryproteinsfromthemalariavectorAnophelesgambiae.FEBSLett.517:67–71.
LavazecC,BourgouinC2008.Mosquito-basedtransmissionblockingvaccinesforinterruptingPlasmodiumdevelopment.MicrobesInfect.10:845–849.
LawrieCH,SimRB,NuttallPa2005.Investigationofthemechanismsofanti-complementactivityinIxodesricinusticks.Mol.Immunol.42:31–38.
LewisLA,RamS2014.Meningococcaldiseaseandthecomplementsystem.Virulence5:98–126.
LuoS,SkerkaC,KurzaiO,ZipfelPF2013.ComplementandinnateimmuneevasionstrategiesofthehumanpathogenicfungusCandidaalbicans.Mol.Immunol.56:161–169.
MedicaDL,InnisP2005.QuantitativeDynamicsofPlasmodiumyoeliiSporozoiteTransmissionbyInfectedAnophelineMosquitoes.Infect.Immun.73:4363–4369.
MedicusRG,GötzeO,Müller-EberhardHJ1976.Alternativepathwayofcomplement:recruitmentofprecursorproperdinbythelabileC3/C5convertaseandthepotentiationofthepathway.J.Exp.Med.144:1076–1093.
Mendes-SousaAF,NascimentoAAS,QueirozDC,ValeVF,FujiwaraRT,AraújoRN,PereiraMH,GontijoNF2013.DifferenthostcomplementsystemsandtheirinteractionswithsalivafromLutzomyialongipalpis(Diptera,Psychodidae)andLeishmaniainfantumpromastigotes.PLoSOne8:e79787.
MillerLH,BaruchDI,MarshK,DoumboOK2002.Thepathogenicbasisofmalaria.Nature415:673–679.
MolinaD,FigueroaLE2009.[MetabolicresistancetoorganophosphateinsecticidesinAnophelesaquasalisCurry1932,Libertadormunicipality,SucreState,Venezuela].Biomedica29:604–615.
MoreiraCK,MarrelliMT,LimaSP,MarinottiO2001.AnalysisofSalivaryGlandProteinsoftheMosquitoAnophelesdarlingi(Diptera :Culicidae)AnalysisofSalivaryGlandProteinsoftheMosquitoAnophelesdarlingi(Diptera :Culicidae).38:763–767.
MorganBP,MarchbankKJ,LonghiMP,HarrisCL,GallimoreAM2005.Complement:centraltoinnateimmunityandbridgingtoadaptiveresponses.Immunol.Lett.97:171–179.
MotaMM,RodriguezA2004.Migrationthroughhostcells:thefirststepsofPlasmodiumsporozoitesinthemammalianhost.Cell.Microbiol.6:1113–

96
1118.
NascimentoA2011.AtividadelíticapelosistemacomplementoempromastigotasdeLeishmania(Leishmania)infantumfrenteacomponentessalivaresdeLutzomyia(Lutzomyia)longipalpis(Diptera :Psychodidae)Atividadelíticapelosistemacomplementoempromastigotasde.
OkrojM,HolmquistE,NilssonE,AnagnostakiL,JirströmK,BlomAM2015.LocalexpressionofcomplementfactorIinbreastcancercellscorrelateswithpoorsurvivalandrecurrence.CancerImmunol.Immunother.64:467–478.
OliveiraF,RowtonE,AslanH,GomesR,CastrovinciPA,AlvarengaPH,AbdeladhimM,TeixeiraC,MenesesC,KleemanLT,Guimarães-CostaAB,RowlandTE,GilmoreD,DoumbiaS,ReedSG,LawyerPG,AndersenJF,KamhawiS,ValenzuelaJG2015.Asandflysalivaryproteinvaccineshowsefficacyagainstvector-transmittedcutaneousleishmaniasisinnonhumanprimates.Sci.Transl.Med.7:290ra90.
Oliveira-FerreiraJ,LacerdaMVG,BrasilP,LadislauJLB,TauilPL,Daniel-RibeiroCT2010.MalariainBrazil:anoverview.Malar.J.9:115.
OoiC-P,HainesLR,SouthernDM,LehaneMJ,Acosta-SerranoA2015.TsetseGmmSRPN10HasAnti-complementActivityandIsImportantforSuccessfulEstablishmentofTrypanosomeInfectionsintheFlyMidgut.PLoSNegl.Trop.Dis.9:e3448.
OwhashiM,HaradaM,SuguriS,OhmaeH,IshiiA2001.TheroleofsalivaofAnophelesstephensiininflammatoryresponse:identificationofahighmolecularweightneutrophilchemotacticfactor.Parasitol.Res.87:376–382.
OwhashiM,HaradaM,SuguriS,OmaeH,IshiiA2008.Identificationofaneosinophilchemotacticfactorfromanophelinemosquitoesasachitinasefamilyprotein.Parasitol.Res.102:357–363.
PangburnMK1989.Analysisofthenaturalpolymericformsofhumanproperdinandtheirfunctionsincomplementactivation.J.Immunol.142:202–207.
PaulyD,NagelBM,ReindersJ,KillianT,WulfM,AckermannS,EhrensteinB,ZipfelPF,SkerkaC,WeberBHF2014.Anovelantibodyagainsthumanproperdininhibitsthealternativecomplementsystemandspecificallydetectsproperdinfrombloodsamples.PLoSOne9:e96371.
PhasomkusolsilS,PantuwattanaK,TawongJ,KhongtakW,KertmaneeY,MonkannaN,KleinTA,KimH,MccardlePW2015.ActaTropicaTherelationshipbetweenwinglength,bloodmealvolume,andfecundityforsevencoloniesofAnophelesspecieshousedattheArmedForcesResearchInstituteofMedicalSciences,Bangkok,Thailand.ActaTrop.i:220–227.
PimentaPF,OrfanoAS,BahiaAC,DuarteAP,Ríos-VelásquezCM,MeloFF,PessoaFA,OliveiraGA,CamposKM,VillegasLM,RodriguesNB,Nacif-PimentaR,SimõesRC,MonteiroWM,AminoR,Traub-CsekoYM,LimaJB,BarbosaMG,LacerdaMV,TadeiWP,SecundinoNF2015.AnoverviewofmalariatransmissionfromtheperspectiveofAmazonAnophelesvectors.Mem.Inst.

97
OswaldoCruz110:23–47.
PotempaM,PotempaJ2012.Protease-dependentmechanismsofcomplementevasionbybacterialpathogens.BiolChem393:873–888.
PóvoaMM,ConnJE,SchlichtingCD,AmaralJCOF,SeguraMNO,SilvaANMDa,SantosCCBDos,LacerdaRNL,SouzaRTLDe,GalizaD,SantaRosaEP,WirtzRa2003.Malariavectors,epidemiology,andthere-emergenceofAnophelesdarlingiinBelém,Pará,Brazil.J.Med.Entomol.40:379–386.
RibeiroJM1987.lxodesdammini :SalivaryAnti-complementActivity.Exp.Parasitol.353:347–353.
RibeiroJM,NussenzveigRH1993.Thesalivarycatecholoxidase/peroxidaseactivitiesofthemosquitoAnophelesalbimanus.J.Exp.Biol.179:273–287.
RibeiroJM,Spielmana1986.Ixodesdammini:salivaryanaphylatoxininactivatingactivity.Exp.Parasitol.62:292–297.
RicklinD,HajishengallisG,YangK,LambrisJD2010.Complement:akeysystemforimmunesurveillanceandhomeostasis.Nat.Immunol.11:785–797.
RicklinD,LambrisJD2013.ProgressandTrendsinComplementTherapeutics(JDLambris,VMHolers,andDRicklin,Eds.).AdvExpMedBiol734:1–22.
RochaACda,BragaÉM,AraújoMS,FranklinBS,PimentaPF2004.EffectoftheAedesfluviatilissalivaonthedevelopmentofPlasmodiumgallinaceuminfectioninGallus(gallus)domesticus.Mem.Inst.OswaldoCruz99:709–715.
RosaTFA,FlammersfeldA,NgwaCJ,KiesowM,FischerR,ZipfelPF,SkerkaC,PradelG2015.ThePlasmodiumfalciparumbloodstagesacquirefactorHfamilyproteinstoevadedestructionbyhumancomplement.Cell.Microbiol.
RoumeninaLT,LoiratC,Dragon-DureyM-A,Halbwachs-MecarelliL,Sautes-FridmanC,Fremeaux-BacchiV2011.AlternativecomplementpathwayassessmentinpatientswithatypicalHUS.J.Immunol.Methods365:8–26.
SacksD,SherA2002.Evasionofinnateimmunitybyparasiticprotozoa.Nat.Immunol.3:1041–1047.
SampaioVS,SiqueiraAM,AlecrimMdasGC,MourãoMPG,MarchesiniPB,AlbuquerqueBC,NascimentoJ,FigueiraÉAG,AlecrimWD,MonteiroWM,LacerdaMVG2015.MalariaintheStateofAmazonas:atypicalBraziliantropicaldiseaseinfluencedbywavesofeconomicdevelopment.Rev.Soc.Bras.Med.Trop.48Suppl1:4–11.
SaúdeMda2010.GuiapráticodetratamentodamalárianoBrasil.Bras.-MinistériodaSaúde:36.
SchroederH,SkellyPJ,ZipfelPF,LossonB,VanderplasschenA2009.Subversionofcomplementbyhematophagousparasites.Dev.Comp.Immunol.33:5–13.
SchuijtTJ,CoumouJ,NarasimhanS,DaiJ,DeponteK,WoutersD,BrouwerM,OeiA,RoelofsJJTH,DamAPvan,PollTvander,Van’tVeerC,HoviusJW,FikrigE

98
2011a.Atickmannose-bindinglectininhibitorinterfereswiththevertebratecomplementcascadetoenhancetransmissionofthelymediseaseagent.CellHostMicrobe10:136–146.
SchuijtTJJ,NarasimhanS,DaffreS,DePonteK,HoviusJWWR,Van’tVeerC,PollTvander,BakhtiariK,MeijersJCM,BoderET,DamAPvan,FikrigE,CoumouJ,NarasimhanS,DaiJ,DePonteK,WoutersD,BrouwerM,OeiA,RoelofsJJTH,vanDamAP,vanderPollT,van’tVeerC,HoviusJWWR,FikrigE2011b.IdentificationandcharacterizationofIxodesscapularisantigensthatelicittickimmunityusingyeastsurfacedisplay.CellHostMicrobe6:e15926.
SilvaANDa,SantosCCDos,LacerdaRN,SantaRosaEP,SouzaRTDe,GalizaD,SucupiraI,ConnJE,PóvoaMM2006.LaboratorycolonizationofAnophelesaquasalis(Diptera:Culicidae)inBelém,Pará,Brazil.J.Med.Entomol.43:107–109.
SimRB,LaichA2000.Serineproteasesofthecomplementsystem.Biochem.Soc.Trans.28:545–550.
SimonN,LasonderE,ScheuermayerM,KuehnA,TewsS,FischerR,ZipfelPF,SkerkaC,PradelG2013.Malariaparasitesco-opthumanfactorhtopreventcomplement-mediatedlysisinthemosquitomidgut.CellHostMicrobe13:29–41.
SinkaME,Rubio-PalisY,ManguinS,PatilAP,TemperleyWH,GethingPW,BoeckelTVan,KabariaCW,HarbachRE,HaySI2010.ThedominantAnophelesvectorsofhumanmalariaintheAmericas:occurrencedata,distributionmapsandbionomicprécis.Parasit.Vectors3:72.
SpäthGF,GarrawayLa,TurcoSJ,BeverleySM2003.Therole(s)oflipophosphoglycan(LPG)intheestablishmentofLeishmaniamajorinfectionsinmammalianhosts.Proc.Natl.Acad.Sci.U.S.A.100:9536–9541.
SturmA,AminoR,SandCvande,RegenT,RetzlaffS,RennenbergA,KruegerA,PollokJ-M,MenardR,HeusslerV2006.ManipulationofHostHepatocytesbytheMalariaParasiteforDeliveryintoLiverSinusoids.Science(80-.).313:1287–1290.
TitusRG,BishopJV,MejiaJS2006.Theimmunomodulatoryfactorsofarthropodsalivaandthepotentialforthesefactorstoserveasvaccinetargetstopreventpathogentransmission.ParasiteImmunol.28:131–141.
TitusRG,RibeiroJM1988.SalivaryglandlysatesfromthesandflyLutzomyialongipalpisenhanceLeishmaniainfectivity.Science239:1306–1308.
TorreiraE,TortajadaA,MontesT,CordobaS,LlorcaO2009.3DstructureoftheC3bBcomplexprovidesinsightsintotheactivationandregulationofthecomplementalternativepathwayconvertase.Proc.Natl.Acad.Sci.U.S.A.106:882–887.
TysonKR,ElkinsC,Silvaa.Mde2008.ANovelMechanismofComplementInhibitionUnmaskedbyaTickSalivaryProteinThatBindstoProperdin.J.Immunol.180:3964–3968.

99
ValenzuelaJG,CharlabR,MatherTN,RibeiroJM2000.Purification,cloning,andexpressionofanovelsalivaryanticomplementproteinfromthetick,Ixodesscapularis.J.Biol.Chem.275:18717–18723.
ValenzuelaJG,FrancischettiIMB,PhamVM,GarfieldMK,RibeiroJMC2003.ExploringthesalivaryglandtranscriptomeandproteomeoftheAnophelesstephensimosquito.InsectBiochem.Mol.Biol.33:717–732.
ValenzuelaJG,FrancischettiIM,RibeiroJM1999.Purification,cloning,andsynthesisofanovelsalivaryanti-thrombinfromthemosquitoAnophelesalbimanus.Biochemistry38:11209–11215.
VaughanJa.,SchellerLF,WirtzRa.,AzadAF1999.InfectivityofPlasmodiumbergheisporozoitesdeliveredbyintravenousinoculationversusmosquitobite:Implicationsforsporozoitevaccinetrials.Infect.Immun.67:4285–4289.
VLADIMIRFAZITODOVALE2011.EstudodaatividadeinibidoradosistemadocomplementohumanopresentenasalivaeconteúdointestinaldeLutzomyialongipalpis(Diptera:Psychodidae).
WeatherallDJ,MillerLH,BaruchDI,MarshK,DoumboOK,Casals-PascualC,RobertsDJ2002.Malariaandtheredcell.HematologyAm.Soc.Hematol.Educ.Program:35–57.
WorldHealthOrganization2014.Worldmalariareport2014.
YoshidaS,SudoT,NiimiM,TaoL,SunB,KambayashiJ,WatanabeH,LuoE,MatsuokaH2008.Inhibitionofcollagen-inducedplateletaggregationbyanophelineantiplateletprotein,asalivaproteinfromamalariavectormosquito.Blood111:2007–2014.
ZhouD,ZhangD,DingG,ShiL,HouQ,YeY,XuY,ZhouH,XiongC,LiS,YuJ,HongS,YuX,ZouP,ChenC,ChangX,WangW,LvY,SunY,MaL,ShenB,ZhuC2014.GenomesequenceofAnophelessinensisprovidesinsightintogeneticsbasisofmosquitocompetenceformalariaparasites.BMCGenomics15:42.