anÁlise taxonÔmica, molecular, ecolÓgica e/ou...
TRANSCRIPT

i
GUSTAVO HENRIQUE JERÔNIMO ALVES
ANÁLISE TAXONÔMICA, MOLECULAR, ECOLÓGICA E/OU
ULTRAESTRUTURAL DOS FILOS BLASTOCLADIOMYCOTA,
CHYTRIDIOMYCOTA E OOMYCOTA DE RESERVATÓRIOS DA BACIA
HIDROGRÁFICA DO SOROCABA/MÉDIO TIETÊ, ESTADO DE SÃO PAULO,
BRASIL
Tese apresentada ao Instituto de Botânica da
Secretaria do Meio Ambiente, como parte dos
requisitos exigidos para obtenção do título de
DOUTOR em BIODIVERSIDADE VEGETAL
E MEIO AMBIENTE, na Área de Concentração
de Plantas Avasculares e Fungos em Análises
Ambientais.
SÃO PAULO, 2018

ii
GUSTAVO HENRIQUE JERÔNIMO ALVES
ANÁLISE TAXONÔMICA, MOLECULAR, ECOLÓGICA E/OU ULTRAESTRUTURAL
DOS FILOS BLASTOCLADIOMYCOTA, CHYTRIDIOMYCOTA E OOMYCOTA DE
RESERVATÓRIOS DA BACIA HIDROGRÁFICA DO SOROCABA/MÉDIO TIETÊ,
ESTADO DE SÃO PAULO, BRASIL
Tese apresentada ao Instituto de Botânica da Secretaria do
Meio Ambiente, como parte dos requisitos exigidos para
obtenção do título de DOUTOR em BIODIVERSIDADE
VEGETAL E MEIO AMBIENTE, na Área de
Concentração de Plantas Avasculares e Fungos em
Análises Ambientais.
ORIENTADORA: DRA CARMEN LIDIA AMORIM PIRES ZOTTARELLI

Ficha Catalográfica elaborada pelo NÚCLEO DE BIBLIOTECA E MEMÓRIA
Alves, Gustavo Henrique Jerônimo
A474a Análise taxonômica, molecular, ecológica, e/ou ultraestrutural dos filos
Blastocladiomycota, Chytridiomycota e Oomycota de reservatórios da Bacia
Hidrográfica do Sorocaba/Médio Tietê, Estado de São Paulo, Brasil / Gustavo
Henrique Jerônimo Alves -- São Paulo, 2018.
150p. il.
Tese (Doutorado) -- Instituto de Botânica da Secretaria de Estado do Meio
Ambiente, 2018.
Bibliografia.
1. Ecologia. 2. Fungos zoospóricos. 3. Ultraestrutura dos zoóspros. I. Título.
CDU: 582.281

iii
À minha esposa e amiga, Carolina
Celestino Jerônimo Alves. Dedico

iv
AGRADECIMENTOS
Agradeço ...
Em especial à Dra. Carmen L.A. Pires Zottarelli, pela confiança em mim depositada
nesses seis anos de convivência, e principalmente pelo exemplo de profissionalismo,
dedicação e paixão pelo que faz.
Agradeço também, a minha esposa Carolina, por estar sempre ao meu lado em todos os
momentos, sempre me apoiando e incentivando em novos desafios e conquistas.
À Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP), pela concessão
das bolsas de Doutorado (n° processo FAPESP - 2014/16358-4) e Bolsa de Estágio de
Pesquisa no Exterior (BEPE n° processo FAPESP – 2016/25800-8), as quais me
permitiram crescer como profissional.
À Dra. Carla Ferragut, do Núcleo de Pesquisa em Ecologia do Instituto de Botânica,
pelo inestimável auxílio com as análises estatísticas e tratamento dos dados abióticos e
bióticos, sempre muito solícita e de bom humor.
À Dra. Denise Bicudo, do Núcleo de Pesquisa em Ecologia do Instituto de Botânica, por
todo incentivo inicial e suporte durante todo o período de estudo.
À Ana Lucia de Jesus, pela convivência e ajuda durante todo este período, sendo sempre
um exemplo de dedicação e trabalho duro.
À Vivis, Richi, Danilo, Luiz, Marcela, Sarah, Alex, Mauro, Nelson e Mari, pelo ótimo
ambiente de trabalho, conversas ao café e ajuda durante o desenvolvimento da tese.
Ao Stefano Zorzal (Jean), pelo apoio durante as coletas e análises laboratoriais, e
cervejas compartilhadas.

v
À Samantha, Elaine e Dorinha por toda ajuda durante as expedições de coleta, sempre
muito dispostas a me ajudar em todas as etapas desde a amostragem até o tratamento
das amostras coletadas.
À Pós-graduação do Instituto de Botânica (PG-IBt), por todo apoio institucional e
infraestrutura fornecida para desenvolvimento deste projeto de doutorado.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela bolsa
de doutorado fornecida durante o primeiro ano de projeto.
Ao Dr. Timothy Y. James (Tim) e University of Michigan por todo suporte, respeito e
confiança depositados durante o período de estágio, o qual foi muito importante para o
meu crescimento profissional e pessoal.
À Rabern Simmons, do laboratório de “Ecology and Evolutionary Biology” da
“University of Michigan”, pelos ensinamentos sobre as análises de ultraestrutura dos
zoósporos e pela amizade construída.
À Anat e Erik, por toda paciência e disposição em ajudar em todos os aspectos, sendo
primordiais na minha adaptação, principalmente nas semanas iniciais, sempre dispostos
a ajudar em qualquer situação.
À William Davis, Alisha, Jill, Lucas, Rob, Kevin and Buck pela agradável convivência e
respeito demonstrado durante os 6 meses de estágio na “University of Michigan”.
Ao Instituto Chico Mendes de preservação da Biodiversidade (ICM-Bio), pela permissão
de coleta de amostras de água e sedimento no reservatório Hedberg, localizado na
Floresta Nacional de Ipanema (FLONA de Ipanema), município de Iperó.
Ao grupo Votorantim, pelo suporte técnico e permissão de coleta de amostras de água e
sedimento nos reservatórios de Santa Helena e Itupararanga.
A todos que de certa forma tornaram possível a realização deste estudo.

vi
Lista de siglas
APA – Área de Proteção Ambiental
BEPE – Bolsa de Estágio de Pesquisa no Exterior
CBA – Companhia Brasileira de Alumínio
CBH-SMT – Conselho da Bacia Hidrográfica Sorocaba e Médio Tietê
CEPAGRI – Centro de Pesquisas Meteorológicas e Climáticas Aplicadas a Agricultura
CETESB – Companhia Ambiental do Estado de São Paulo
CONAMA – Conselho Nacional do Meio Ambiente
CCIBt – Coleção de Culturas de Algas, Fungos e Cianobácterias
DDSA – anidrido dodecenil-succínico
DMP 30 – Tris-Dimetilaminometil fenol
DMSO – Dimetilsulfato
EMS© – Electron Microscopy Sciences
FAPESP – Fundação de Ámparo à Pesquisa no Estado de São Paulo
FLONA de Ipanema – Floresta Nacional de Ipanema
IBAMA: Instituto Nacional do Meio Ambiente e Recursos Naturais Renováveis e da
Amazônia Legal
IET – Índice de Estado Trófico
NT – Nitrogênio Total
OD – Oxigênio dissolvido
PCA – Análise de Componentes Principais
PCoA – Análise de Coordenadas Principais
PEFI – Parque Estadual das Fontes do Ipiranga
PFLA – Análise de Ácidos Graxos Fosfolipídicos
PT – Fósforo Total
SigRH – Sistema de Gerenciamento de Recursos Hídricos
UGRHI – Unidade de Gerenciamento de Recursos Hídricos
UrAc – Acetato de Uranila

vii
SUMÁRIO
APRESENTAÇÃO .................................................................................................................................. viii
RESUMO .................................................................................................................................................. 09
ABSTRACT .............................................................................................................................................. 11
INTRODUÇÃO GERAL .......................................................................................................................... 13
OBJETIVOS ESPECÍFICOS ........................................................................................ .............................19
HIPÓTESES ................................................................................................................ .............................. 19
METODOLOGIA GERAL ....................................................................................................................... 21
REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................................................... 48
CAPÍTULO 1. Estrutura da comunidade de organismos zoospóricos heterotróficos (fungos
zoospóricos e oomicetos) da coluna d’água e sedimento superficial de reservatórios com distintos
estados tróficos
Resumo ...................................................................................................................................................... 56
Abstract ...................................................................................................................................................... 57
Introdução .................................................................................................................................................. 59
Material e Métodos ..................................................................................................... ............................... 62
Resultados & Discussão ............................................................................................................................. 65
Referências Bibliográficas ......................................................................................................................... 90
CAPÍTULO 2. Novelties for the Nowakowskiella genus: a new species and new combination
Abstract ……………………………………………………………………………….............................. 97
Introduction ……………………………………………………………………………............................ 97
Material and Methods ………………………………………………………………................................ 99
Results …………………………………………………………………………….................................. 103
Discussion ……………………………………………………………………………………………… 110
Literature cited ……………………………………………………………………………..................... 112
CAPÍTULO 3. Boothiomyces angulosum: a new combination into Terramycetaceae (Rhizophydiales,
Chytridiomycota)
Abstract ………………………………………………………………………………………………... 116
Introduction ……………………………………………………………………………………………. 117
Material and Methods …………………………………………………………………………………. 118
Results ……………………………………………………………………………................................. 121
Discussion …………………………………………………………………………………................... 126
References …………………………………………………………………………............................... 129
CONSIDERAÇÕES FINAIS ................................................................................................................ 131
Anexo I ............................................................................................................ ........................................ 132
Anexo II ......................................................................................... .......................................................... 134
Anexo III ................................................................................................................................. ................. 142
Apêndice I .................................................................................................................. .............................. 150

viii
APRESENTAÇÃO
A tese intitulada “Análise taxonômica, ecológica, molecular e/ou
ultraestrutural dos filos Blastocladiomycota, Chytridiomycota e Oomycota de
reservatórios da Bacia hidrográfica do Sorocaba/Médio Tietê, estado de São Paulo,
Brasil”, foi estruturada em três capítulos, os quais são precedidos por uma
INTRODUÇÃO GERAL, OBJETIVOS ESPECÍFICOS, HIPÓTESES E
METODOLOGIA GERAL.
O CAPÍTULO 1 aborda a diversidade de Blastocladiomycota, Chytridiomycota
e Oomycota na coluna d’água e sedimento superficial, de três reservatórios da Bacia
hidrográfica do Sorocaba/Médio Tietê, com distintos estados tróficos. Os padrões de
ocorrência e abundância desses grupos de organismos foram relacionados às flutuações
sazonais e aos diferentes fatores abióticos mensurados na coluna d’água e sedimento
superficial. As análises moleculares, neste contexto, foram utilizadas para corroborar as
identificações morfológicas.
O CAPÍTULO 2, elaborado de acordo com as normas da revista Mycologia, trata-
se de um estudo taxonômico e filogenético (análise combinada das regiões 18S, 5,8S e
28S do rDNA) de representantes da ordem Cladochytriales, com especial enfoque no
gênero Nowakowskiella J. Schröt., para o qual foi descrito uma nova espécie nomeada
como Nowakowskiella crenulata Jerônimo & Pires-Zottarelli, e proposto um novo
gênero, Karlingiella Jerônimo, Jesus & Pires-Zottarelli, considerando o posicionamento
filogenético de Nowakowskiella elongata Karling como grupo irmão do gênero
Cladochytrium Nowak.
O CAPÍTULO 3, elaborado de acordo com as normas da revista Nova Hedwigia,
inclui uma discussão morfológica, filogenética e ultraestrutural (baseado na ultraestrutura
dos zoósporos) da espécie Rhizophydium angulosum Karling, para a qual foi proposta
uma nova combinação (Boothiomyces angulosum (Karling) Jerônimo & Pires-Zottarelli).
Além disso, discutimos também a morfologia e posicionamento filogenético da espécie
Rhizophydium elyense Sparrow.

9
RESUMO GERAL
A bacia hidrográfica do Sorocaba/Médio Tietê (UGRHI 10) está situada na região
Centro-Sudeste do Estado de São Paulo, uma das mais desenvolvidas do interior do
estado. Nesta região, devido à crescente demanda por energia elétrica e abastecimento
público, diversos reservatórios foram criados, os quais estão sujeitos aos efeitos de ações
antrópicas locais. Dentre os impactos detectados estão a eutrofização e o uso desordenado
do solo, fatores que estão associados à perda da qualidade da água e da biodiversidade.
Representantes dos filos Blastocladiomycota e Chytridiomycota (fungos zoospóricos) e
do filo Oomycota, importantes agentes decompositores e parasitas, são pouco explorados
nestes ambientes aquáticos, existindo a necessidade de uma melhor compreensão do papel
destes organismos nos processos de transferência de energia nesses ecossistemas. Diante
disso, o objetivo deste estudo foi relacionar a riqueza e a abundância destes grupos de
organismos com a sazonalidade e alguns fatores abióticos mensurados na coluna d’água
e sedimento superficial de três reservatórios brasileiros (Santa Helena, Itupararanga e
Hedberg), inseridos na UGRHI 10, conhecidamente com distintos estados tróficos. Além
disso, disponibilizar sequências gênicas dos espécimes identificados em nível específico
e construir filogenias, confirmando ou rejeitando hipóteses filogenéticas dentre os
representantes de Chytridiomycota, por meio da análise da ultraestrutura dos zoósporos
destes fungos. Para isso, foram realizadas quatro coletas em cada reservatório, onde foram
mensuradas algumas variáveis físicas e químicas da água (temperatura, pH,
condutividade elétrica, oxigênio dissolvido, nitrogênio e fósforo total) e do sedimento
superficial (nitrogênio total, fósforo total e carbono orgânico total). A estrutura da
comunidade foi avaliada considerando-se a ocorrência, frequência e abundância dos
táxons, mediante cálculo da riqueza de espécies, do índice de diversidade de Shannon, do
índice de dominância de Simpson, da equitatividade e do índice de similaridade de
Sørensen. Durante o estudo, 52 táxons destes organismos zoospóricos heterotróficos
foram identificados, sendo 25 deles representantes dos fungos zoospóricos e 27 de
oomicetos. Dentre os fungos zoospóricos, uma nova espécie do gênero Nowakowskiella
J. Schröt foi descrita e o gênero Avachytrium Velez & Letcher relatado pela primeira vez
para o Brasil, assim como Plectospira gemmifera Drechsler a foi dentre os oomicetos. No
total, 102 sequências das regiões 18S, ITS, 28S, RPB1, RPB2 e COXI foram obtidas
durante o estudo, considerando os dois grupos de organismos, as quais representam 25
táxons, dentre os quais, Nowakowskiella ramosa Butler, uma nova espécie nomeada como

10
Nowakowskiella crenulata Jerônimo & Pires-Zottarelli, Rhizophydium angulosum
Karling (para o qual está sendo proposta uma nova combinação) e Plectospira gemmifera
Drechsler, todos inseridos pela primeira vez em análises filogenéticas. Foi observada uma
comunidade diversa com poucos táxons dominantes, sendo a maior parte deles observada
na estação chuvosa, com Cladochytrium replicatum Karling, Nowakowskiella
hemisphaerospora Shanor, Aphanomyces spp., Dictyuchus spp. e Pythium dos grupos P
e T, sendo os táxons mais frequentes e abundantes, considerando os reservatórios e
estações (seca e chuvosa). Os organismos zoospóricos estudados apresentaram uma
ampla distribuição ao longo de diferentes gradientes tróficos, existindo táxons
estritamente relacionados aos locais com as mais altas concentrações de nitrogênio e
fósforo total, e outros aos locais com as mais baixas concentrações destes nutrientes, e
que potencialmente poderiam ser indicativos da qualidade das águas. Concluímos,
portanto, que estes organismos zoospóricos heterotróficos são, geralmente, tolerantes às
amplas variações nas concentrações de nitrogênio e fósforo, mas alguns táxons são
limitados pela disponibilidade destes nutrientes, particularmente o fósforo.
Palavras-chave: diversidade, ecologia, filogenia, fungos zoospóricos, oomicetos,
ultraestrutura dos zoósporos.

11
ABSTRACT
The “Sorocaba/Médio Tietê” hydrographic basin (UGRHI 10) is located in the Center-
Southeast region of the São Paulo state, one of the most developed regions of the state.
In this region, due to the growing demand for electricity and public supply, several
reservoirs were built, which are subject to the effects of local anthropogenic actions.
Among the detected impacts the eutrophication and disorderly use of the land are factors
that are associated with the loss of water quality and biodiversity. Blastocladiomycota,
Chytridiomycota (zoosporic fungi) and Oomycota, important decomposing agents and
parasites, are poorly explored in these aquatic environments and additional studies are
necessary to clarify the role of these organisms in the energy transfer processes in these
ecosystems. Considering this, the aim of this study was to relate the species richness and
abundance of these groups with seasonality and some abiotic factors measured in the
water column and surface sediment of three Brazilian reservoirs (Santa Helena,
Itupararanga and Hedberg), located in UGRHI 10, with distinctive trophic states. In
addition, we also intend to provide gene sequences of the specimens identified at specific
level and build their phylogeny, confirming or rejecting phylogenetic hypotheses to the
Chytridiomycota representatives, through zoospore ultrastructural analysis. In order to do
this, four sampling were performed in each reservoir, in which physical and chemical
variables of the water (temperature, pH, electrical conductivity, dissolved oxygen,
nitrogen and total phosphorus) and surface sediment (total nitrogen, total phosphorus and
total organic carbon) were measured. The community structure was evaluated considering
the abundance, frequency, species richness, Shannon’s diversity index, Simpson’s index,
Evenness and similarity Sorensen index. During the study, 52 taxa of heterotrophic
zoosporic organisms were identified, 25 of which representative of Blastocladiomycota
and Chytridiomycota phyla (zoosporic fungi) and 27 of Oomycota phylum. On the
zoosporic fungi, a new species of the Nowakowskiella J. Schröt genus was described and
Avachytrium Velez & Letcher reported for the first time in Brazil, as well as Plectospira
gemmifera Drechsler to the Oomycota. In total, 102 sequences from the 18S, ITS, 28S,
RPB1, RPB2, and COXI regions were obtained during the study, considering the two
groups of organisms, which represent 25 taxa, being Nowakowskiella ramosa Butler, the
new species Nowakowskiella crenulata Jerônimo & Pires-Zottarelli, Rhizophydium
angulosum Karling (for which we are proposing a new combination) and Plectospira
gemmifera Drechsler included for first in phylogenetic analyzes. We observed a diverse

12
community with few dominant taxa, which occurred predominantly at rainy season, being
Cladochytrium replicatum Karling, Nowakowskiella hemisphaerospora Shanor,
Aphanomyces spp., Dictyuchus spp. and Pythium group P and T, the most frequent and
abundant, considering the reservoirs and seasons (dry and rainy). Zoosporic organisms
presented a wide distribution along different trophic gradients, with species strictly
related to locals with high concentrations of nitrogen and total phosphorus, and others
with low concentrations of these nutrients, which could potentially be taxa indicative of
quality of water. We conclude that heterotrophic zoosporic organisms are, in general,
tolerant to wide variations in nitrogen and phosphorus concentrations, but some taxa are
limited by the availability of these nutrients, particularly phosphorus.
Key words: diversity, ecology, phylogeny, oomycetes, zoosporic fungi, zoospore
ultrastructure.

13
INTRODUÇÃO GERAL
Os organismos zoospóricos heterotróficos dos filos Blastocladiomycota,
Chytridiomycota e Oomycota possuem como similaridade a produção de esporos
flagelados (zoósporos), os quais os permitem se dispersarem ativamente em ecossistemas
aquáticos e terrestres úmidos. Nestes ambientes, esses organismos atuam como
decompositores de matéria orgânica ou como parasitas de diferentes grupos de
hospedeiros, exercendo assim um importante papel na ciclagem do carbono (Shearer et
al. 2007, Sime-Ngando et al. 2011, Wurzbacher et al. 2016).
Blastocladiomycota e Chytridiomycota são filos incluídos no reino Fungi, cujos
representantes possuem esporos com um único flagelo liso, posteriormente inserido,
ocupando posicionamento basal na árvore filogenética dos fungos (James et al. 2006,
Hibbett et al. 2007). Chytridiomycota conta com aproximadamente 706 espécies
distribuídas atualmente em nove ordens (Chytridiales, Cladochytriales, Lobulomycetales,
Polychytriales, Rhizophlyctidales, Rhizophydiales, Mesochytriales, Gromochytriales e
Spizellomycetales), as quais estão inseridas na Classe Chytridiomycetes (Sparrow 1942,
Letcher et al. 2006, 2008, Kirk et al. 2008, Mozley-Standridge et al. 2009, Simmons et
al. 2009, Wakefield et al. 2010, Longcore & Simmons 2012). Já Blastocladiomycota, filo
criado a partir de Blastocladiales (ordem que anteriormente pertencia ao Filo
Chytridiomycota), possui uma única classe, Blastocladiomycetes, e é composto por 179
espécies (Kirk et al. 2008), cujos representantes divergiram do clado dos
Chytridiomycota e são considerados grupo irmão juntamente com Zygomycota,
Glomeromycota e Olpidium (James et al. 2006).
Representantes dos filos de fungos zoospóricos existem na Terra há pelo menos
400 milhões de anos como indicado por registros fósseis do baixo Devoniano (Taylor et
al. 1992, Remy et al. 1994) e são considerados de crucial importância nos processos de
decomposição e parasitismo nos ecossistemas aquáticos, onde são frequentemente
observados como parasitas de algas, cianobactérias, macrófitas, invertebrados, anfíbios,
fungos e oomicetos (Shearer et al. 2007, Sime-Ngando et al. 2012) e como sapróbios em
material orgânico particulado e dissolvido oriundos de fontes alóctones e autóctones
(Kagami et al. 2004). Embora a maioria das espécies conhecidas seja decompositora de
celulose, queratina e quitina (Powell 1993), algumas se destacam como importantes
parasitas de plantas de interesse econômico, como por exemplo, Physoderma maydis
(Miyabe) Miyabe que causa mancha marrom do milho e Synchytrium endobioticum
(Schilberszky) Percival responsável pela verruga da batata (Alexopoulos et al. 1996).

14
Espécies dos gêneros Catenaria Sorokin (Catenaria spinosa Martin, Catenaria
anguillulae Sorokin), Coelomomyces Keilin (Coelomomyces utahensis (Romney) Couch
& Nielsen) e Polycaryum Stempell (Polycaryum laeve Stempell) são parasitas comuns
em cladóceros, copépodos, rotíferos, nematóides e larvas de dípteros, atuando
decisivamente no controle dessas populações (Gleason et al. 2010). Gerphagnon et al.
(2013) relataram a presença de quitrídias parasitas de cianobactérias em eventos de
florações e evidenciaram o papel das mesmas na fragmentação mecânica dos filamentos
das cianobactérias, contribuindo assim, para o declínio destes eventos. Batrachochytrium
dendrobatidis Longcore, Pessier & D.K. Nichols é considerado o principal responsável
pelo declínio e extinção de mais de 200 espécies de anfíbios ao redor do mundo (Stuart
et al. 2004), tendo ainda sido descrita outra espécie para o gênero, Batrachochytrium
salamandrivoras Martel, a qual foi encontrada parasitando populações de Salamandra
salamandra Lineus (salamandras de fogo) na Holanda (Martel et al. 2013).
A classificação e as relações filogenéticas dos representantes dos fungos
zoospóricos (Blastocladiomycota e Chytridiomycota) vem passando por constantes
reestruturações tendo em vista as características ultraestruturais dos zoósporos e dados de
sequências genicas (James et al. 2000, 2006a, Letcher et al. 2008, Mozley-Standridge et
al. 2009, Longcore e Simmons 2012). O primeiro estudo de microscopia eletrônica de
fungos zoospóricos mostrou o incrível potencial da arquitetura ultraestrutural como
ferramenta para entendimento das relações evolutivas destes fungos (Koch 1961). Sua
aplicação direta na reclassificação de Chytridiomycota foi inicialmente proposta por Barr
(1980), o qual segregou Spizellomycetales de Chytridiales. Posteriormente Li et al.
(1993), também apoiados em dados ultraestruturais, transferiram os representantes
poliflagelados encontrados no rúmen de herbívoros de Spizellomycetales para
Neocallimastigales. Paquin et al. (1997), analisando genes mitocondriais, transferiram
Harpochytriaceae de Chytridiales (sensu Barr 1980) para Monoblepharidales. James et
al. (2000), ao proporem que as análises filogenéticas corroboravam as modificações
taxonômicas indicadas pelos dados ultraestruturais, aliaram essas duas ferramentas e
confirmaram a polifilia de Chytridiales e a divergência de Blastocladiales de Chytridiales
(James et al. 2006). Com isso, as ordens Blastocladiales e Neocallimastigales foram
elevadas a filo, Blastocladiomycota (James et al. 2006) e Neocallimastigomycota,
respectivamente, e os representantes de Monoblepharidales para
Monoblepharidomycetes, sendo então o filo Chytridiomycota redefinido (Hibbet et al.
2007). Embora as análises moleculares e ultraestruturais estejam constantemente

15
reconfigurando a classificação dentro de Blastocladiomycota e Chytridiomycota, estudos
com este enfoque na América do Sul podem ser resumidos ao de Velez et al. (2013) para
a Argentina, no qual foram descritos três novos gêneros, não havendo, no entanto,
nenhum estudo desta natureza no Brasil.
Já os representantes de Oomycota, apesar de serem também organismos
possuidores de esporos flagelados, estão inseridos no Reino Straminipila, pois apresentam
características morfológicas, fisiológicas, bioquímicas e moleculares que os distinguem
dos fungos (Alexopoulos et al. 1996, Beakes et al. 2014). Ainda hoje, estes organismos
têm sido estudados por micologistas, especialmente por ocuparem os mesmos nichos que
os fungos e por já terem sido classificados como tal (Sparrow 1960). Há dentre eles
representantes sapróbios, os quais são considerados importantes elementos na degradação
e ciclagem de nutrientes nos ecossistemas e também espécies parasitas de algas, peixes,
crustáceos, plantas, fungos, mamíferos (incluindo o homem). Alguns gêneros de
Oomycota, como por exemplo, Achlya, Aphanomyces e Saprolegnia, possuem espécies
que podem parasitar peixes e seus ovos, e em Pythium e Phytophthora se encontram
muitas espécies fitopatógenas (Alexopoulos et al. 1996). Pythium insidiosum tem se
destacado como importante parasita de vertebrados, causando pitiose em equinos,
bovinos, caprinos, cães, gatos e no homem (Alexopoulos et al. 1996), com relatos da
espécie no Brasil, especialmente na região sul (Santurio et al. 1998, Leal et al. 2001,
Bosco et al. 2005). Recente relato de Pythium aphanidermatum na espécie humana no
Afeganistão revela o quão oportunistas estes organismos podem ser (Calvano et al. 2011).
Mesmo diante da expressiva frequência e importância destes organismos
zoospóricos, que chamaremos ao longo do texto de fungos zoospóricos
(Blastocladiomycota e Chytridiomycota) e oomicetos (Oomycota), nos processos de
transferência de energia nos ecossistemas aquáticos, estudos abordando sua diversidade
e importância em reservatórios do Brasil ainda são escassos e restritos ao estado de São
Paulo, podendo ser resumidos aos trabalhos de Rocha & Merighi (1986) na Represa
Billings, Pires-Zottarelli (1990) na Represa do Lobo, Rocha & Pires-Zottarelli (2002) na
Represa do Guarapiranga e Rocha (2004) nos Lagos das Garças e das Ninféias do Parque
Estadual das Fontes do Ipiranga (PEFI). No entanto, somente o último discute de forma
mais aprofundada a influência dos fatores abióticos sobre os padrões de ocorrência destes
organismos. Dos 121 táxons de Chytridiomycota e 18 de Blastocladiomycota conhecidos
no Brasil (Lista de espécies da Flora do Brasil 2014), aproximadamente 79% foram
relatados para ecossistemas lóticos (rios, riachos e cachoeiras) e solos, principalmente em

16
áreas de Mata Atlântica (Beneke & Rogers 1962, 1970, Rogers et al. 1970, Lyra &
Milanez 1974, Milanez et al. 1994, Pires-Zottarelli et al. 1996, Pires-Zottarelli 1999,
Pires-Zottarelli & Rocha 2007, Pires-Zottarellli & Gomes 2007, Nascimento & Pires-
Zottarelli 2009, 2010, entre outros), sendo a diversidade em reservatórios representada
por somente 21% das espécies conhecidas no país. Este mesmo cenário pode também ser
atribuído para o filo Oomycota, que possui somente 19,5% da diversidade relatada para
ecossistemas lênticos (Rocha & Merighi 1986; Pires-Zottarelli, 1990; Rocha & Pires-
Zottarelli, 2002; Rocha, 2004), com maior representatividade dos gêneros Achlya,
Aphanomyces, Pythium e Saprolegnia.
Os reservatórios possuem grande importância devido aos diversos serviços que
prestam à população, principalmente no que se refere ao abastecimento público e geração
de energia elétrica, sendo a hidroeletricidade a principal fonte de energia do país e
responsável por 75% do total de energia produzida (Tolmasquim 2012). No entanto, sua
construção gera importantes, e muitas vezes, irreversíveis impactos, como perda da
biodiversidade e da qualidade da água e mudanças socioeconômicas regionais (Tundisi
et al. 2008). Dentre os diversos impactos antrópicos gerados, a eutrofização e o uso
desordenado do solo da bacia hidrográfica, são as variáveis que frequentemente têm sido
associadas com a perda da qualidade da água e da biodiversidade (Pusceddu et al. 2007,
Cunha et al. 2010). O processo de eutrofização tem sido bastante investigado por
representar uma das principais causas de alterações na qualidade da água e redução da
diversidade em águas continentais (Margalef 1983). Este processo geralmente associado
ao aumento da concentração de nutrientes, especialmente fósforo e nitrogênio, tem como
consequência o aumento de suas produtividades, podendo os ecossistemas aquáticos
passarem de oligotrófico ou mesotrófico para eutrófico ou mesmo hipereutrófico (Esteves
1998). Já os tipos de usos e a ocupação do solo na bacia hidrográfica possuem relações
significativas no que diz respeito às diversas substâncias químicas lançadas nos corpos
d’água, como efluentes domésticos, industriais e agrícolas, expondo diversos organismos
a uma grande variedade de compostos mutagênicos e citotóxicos (Amaral et al. 2007).
Nos reservatórios essas alterações ecológicas podem ser detectadas a partir do
monitoramento da água e do sedimento acumulado, seja este superficial ou depositado
em camadas (Birks & Birks 2006). A análise do sedimento fornece dados cumulativos e
constitui um arquivo de informações de natureza biogeoquímica, uma vez que as camadas
de deposição temporais se encontram sequencialmente acumuladas (Mozeto 2004).
Assim, o sedimento superficial representa uma amostra espacial e temporalmente

17
integrada dos eventos que se acumularam no passado recente (Smol 2008). O oxigênio,
se presente, está restrito as camadas superficiais do perfil sedimentar, e por isso, os fungos
presentes neste compartimento geralmente apresentam adaptações fisiológicas
(mecanismo fermentativo) e/ou morfológicas (esporos de resistência) para suportar tais
limitações (Wurzbacher et al. 2010). As leveduras e os representantes do filo
Chytridiomycota são considerados os grupos de fungos mais ativos no perfil sedimentar
de reservatórios (Slapeta et al. 2005) e técnicas de PFLA (Goedkoop et al. 2005,
Widenfalk et al. 2008) e clonagem (Dawson & Pace 2002, Luo et al. 2005), indicam estes
dois grupos como micota dominante nos sedimentos dos lagos e reservatórios, com
tendência de maior diversidade de espécies nos sedimentos localizados próximos as zonas
litorâneas (Collins & Willoughby 1962, Sparrow 1968). Além disso, assim como os
fungos zoospóricos, alguns poucos estudos têm apontado a importância dos oomicetos
nos processos de transferência de energia no sedimento, sendo alguns gêneros, tais como
Aphanomyces spp. e Pythium spp., considerados abundantes e ubíquos nas camadas
superficiais do perfil sedimentar (Sparrow 1968, Boopathi et al. 2015, Yang & Hong,
2015, Wurzbacher et al. 2016). No entanto, considerando a totalidade dos estudos
desenvolvidos em reservatórios, somente pouco deles (Goedkoop et al. 2005, Wildenfalk
et al. 2008, Zhang et al. 2015) analisaram a diversidade destes grupos de organismos no
sedimento superficial, sendo no Brasil estes relatos inexistentes.
Estudos abordando os efeitos dos fatores físicos, químicos e biológicos no
tamanho e composição das populações, natureza das interações entre parasitas e
hospedeiros e importância na dinâmica da ciclagem do carbono na cadeia alimentar, ainda
são escassos para estes organismos (Gleason et al. 2008). De acordo com Lefèvre et al.
(2007), os representantes do filo Chytridiomycota (quítridias) são muitas vezes
totalmente ignorados e erroneamente classificados em muitos estudos ecológicos,
especialmente os que abordam a biodiversidade em ecossistemas aquáticos. Em lagos e
reservatórios estes fungos têm sido principalmente reconhecidos como membros
proeminentes da comunidade planctônica, atuando decisivamente como parasitas,
afetando assim a sucessão do fitoplâncton e a produção primária. Além disso, tanto os
fungos zoospóricos como os oomicetos, atuam também como decompositores de
materiais recalcitrantes como quitina e celulose, e ainda disponibilizam energia dos
produtores primários e detritívoros para níveis tróficos superiores, ao terem seus
zoósporos predados pelo zooplâncton (Kagami et al. 2007, Gleason et al. 2008). Lefèvre
et al. (2012) baseando-se em análises moleculares evidenciaram que estes organismos

18
não somente compõem a comunidade planctônica dos reservatórios estudados, mas são
os organismos dominantes nestes locais. E ainda, segundo estes autores, o uso de técnicas
de clonagem mostrou que, a maioria das sequências recuperadas nestes ambientes não
combinou com qualquer táxon descrito, sugerindo que lagos e reservatórios abrigam uma
alta e inexplorada diversidade.
Embora as atuais técnicas de clonagem indiquem os fungos zoospóricos como
micota dominante de lagos e reservatórios, os fatores que afetam a variação temporal e a
estrutura da comunidade ainda permanecem pouco investigados (Wurzbacher et al.
2016). De maneira geral, quando consideramos os fungos zoospóricos e oomicetos, a
relação entre os padrões de ocorrência e distribuição em ambientes com diferentes trofias,
ainda é pouco explorada em escala global (Cooke 1976, Czeczuga et al. 1989, 1990a,b,
Czeczuga 1991a,b, Czeczuga & Woronowicz 1991, Lefranc et al. 2005, Chen et al. 2008,
Marano et al. 2011, Monchy et al. 2011), e principalmente no Brasil (Rocha 2004).
Muitos dos estudos conduzidos em ambientes com diferentes estados tróficos (Tan & Lim
1983, 1984, Czeczuga 1994, Rancovic 1998, Czeczuga & Muszynska 2001) listam
principalmente a diversidade em espécies, não relacionando os padrões de sua ocorrência
e distribuição com fatores abióticos. Czeczuga et al. (1990a,b) e Czeczuga (1991a,b)
apontaram estes organismos como tolerantes e adaptados a diferentes níveis de poluição
orgânica, mas relataram preferência dos mesmos por ambientes menos poluídos. Lefranc
et al. (2005), Chen et al. (2008), Lepère et al. (2008), Monchy et al. (2011) e Wurzbacher
et al. (2010, 2016) apontam, por meio de análises moleculares, que a diversidade tende a
ser menor em águas mais produtivas, sendo os reservatórios oligotróficos e mesotróficos
os que normalmente apresentam maior diversidade. Lefèvre et al. (2012) mostraram
maior diversidade de quitrídias em reservatórios meso-eutróficos do que em ambientes
oligotróficos, sugerindo que ambientes em estado trófico intermediário (mesotróficos)
sejam os mais propícios para ocorrência destes fungos. Para o Brasil, Rocha (2004)
sugeriu que a riqueza em espécies dos fungos zoospóricos e oomicetos dos lagos do
Parque Estadual das Fontes do Ipiranga (PEFI), com distintas trofias (mesotrófico e
eutrófico), apresentou correlação negativa com as concentrações de nitrogênio e fósforo,
tendo sido demonstrada e comprovada a natureza limáfoba ou limaxena destes
organismos. A autora pontua também que a ausência dos mesmos em locais amostrados
com altíssimas concentrações de nitrogênio e fósforo total indica a sua possível utilização
como bioindicadores de poluição, estando os mesmos completamente ausentes em tal
situação.

19
Com relação aos padrões sazonais de distribuição desses organismos zoospóricos
em ecossistemas lênticos, verifica-se uma grande escassez de estudos. Em regiões
temperadas se estabelece normalmente o período chuvoso como o de maior diversidade
(Czeczuga 1986; 1995; Czeczuga et al. 2010). Em regiões tropicais, os únicos estudos a
abordarem a sazonalidade em ecossistemas lênticos brasileiros foram os de Pires-
Zottarelli (1990) e Rocha (2004), os quais também apontaram ser a maior diversidade
encontrada durante a estação chuvosa.
Diante do exposto, o objetivo geral deste estudo foi avaliar a ocorrência,
frequência e abundância dos fungos zoospóricos e oomicetos em reservatórios, com
distintos estados tróficos, da Bacia Hidrográfica do Sorocaba/Médio Rio Tietê.
Objetivos específicos
Analisar pela primeira vez a diversidade destes organismos no sedimento
superficial de reservatórios do Brasil;
Comparar a diversidade na coluna d´água e sedimento superficial;
Evidenciar, dentre os fatores abióticos mensurados, os mais atuantes na
ocorrência e distribuição dos táxons nos reservatórios amostrados;
Sequenciar as regiões SSU, ITS, LSU, COX1, RPB1 e RPB2 dos táxons
cultivados, bem como, incluí-los em análises filogenéticas;
Disponibilizar pela primeira vez dados de ultraestrutura dos zoósporos de
espécimes brasileiros de Chytridiomycota, com ênfase nas espécies de
classificação filogenética incerta e com ultraestrutura desconhecida.
Hipóteses
Se os fungos zoospóricos e oomicetos respondem a mudanças na disponilidade
de nutrientes (Gleason et al. 2008; Lefèvre et al. 2012; Wurzbacher et al. 2016),
então, espera-se encontrar diferenças na composição de espécies entre
reservatórios de diferentes trofias e, espécies associadas às diferentes
disponilidade de nutrientes;
Considerando que a maior parte dos estudos em reservatórios analisaram a
diversidade dos fungos zoospóricos e oomicetos na coluna d’água (Gleason et
al. 2008; Wurzbacher et al. 2016), espera-se que a maior riqueza destes
organismos seja observada neste compartimento;

20
Já que maior riqueza de espécies foi observada durante a estação chuvosa, em
estudos desenvolvidos em ecossistemas lênticos brasileiros (Pires-Zottarelli
1990; Rocha 2004), espera-se que a riqueza de espécies de fungos zoospóricos
e oomicetos também seja maior no período chuvoso nos reservatórios
estudados.

21
METODOLOGIA GERAL
Área de estudo
Caracterização geral da Bacia hidrográfica do Sorocaba/Médio Tietê
A Bacia Hidrográfica do Sorocaba/Médio Tietê foi definida como a “Unidade de
Gerenciamento de Recursos Hídricos – UGRHI 10 – Tietê/Sorocaba” pela Lei nº 9.034/94
de 27/12/1994, e o Comitê que gerencia esta bacia (CBH-SMT), foi formado em 1995
após a implantação do Sistema Estadual de Gerenciamento de Recursos Hídricos
(SigRH). As principais preocupações compartilhadas para a formação do comitê foram a
poluição das águas do rio Tietê e do reservatório de Itupararanga, principal manancial da
bacia do Sorocaba (SigRH, 2018).
A Bacia Hidrográfica do Sorocaba/Médio Tietê (UGRHI 10) está localizada na
região centro-sudeste do Estado de São Paulo e é constituída pela Bacia do rio Sorocaba
e pela bacia do Médio Tietê superior (Fig. 1). Compreende o trecho desde a barragem do
Rasgão, à montante, até a barragem de Barra Bonita, à jusante. A UGRHI 10 é delimitada
pelas bacias do Alto Tietê (UGRHI 06) a leste, Tietê/Jacaré (UGRHI 13) a noroeste e pela
UGRHI 05 a norte e nordeste, esta última formada pelas bacias dos rios Piracicaba,
Capivari e Jundiaí, que deságuam na margem direita do rio Tietê. Na porção sul-sudoeste-
noroeste, as bacias do Alto e Médio Paranapanema (UGRHIs 14 e 17, respectivamente)
são as porções limítrofes da UGRHI 10 (IPT 2008). A região do Médio Tietê é muito
influenciada pelas bacias a montante, tanto o Alto Tietê, que despeja grande parte do
esgoto não tratados da região metropolitana de São Paulo, quanto as Bacias do Piracicaba,
Capivari e Jundiaí que transportam os efluentes da Região Metropolitana de Campinas e
do Município de Jundiaí e deságuam no trecho médio do Rio Tietê dentro da UGRHI10
(CBH-SMT, 2012).

22
Figura 1. Localização da Bacia do Sorocaba/Médio Tietê (UGRHI 10) e seus limites com as demais
UGRHIs do Estado de São Paulo (Fonte: modificado de Silva & Matsumura-Tundisi 2010).
A área total da bacia do Sorocaba/Médio Tietê é de 12.099 km2, abrangendo uma
população de 1,650 milhões de habitantes em 54 municípios (SigRH, 2018). Está
subdividida em seis sub-bacias, três delas que compõem a Bacia do Rio Sorocaba e outras
três por drenagens de pequeno e médio porte, que drenam para o rio Tietê (Fig. 2). Neste
trecho, o rio Tietê tem como principais afluentes os rios Jundiaí, Capivari e Piracicaba,
enquanto o rio Sorocaba é formado pelos rios Una, Sorocabuçu e Sorocamirim, cujas
cabeceiras se localizam nos municípios de Ibiúna, Cotia, Vargem Grande Paulista e São
Roque. No início do Rio Sorocaba, suas águas foram represadas, formando o reservatório
de Itupararanga, importante manancial da região, o qual é especialmente prejudicado
devido à poluição que recebe da região metropolitana de São Paulo (SigRH 2018). O
reservatório de Itupararanga está ligado em um continuo ao de Santa Helena, o qual é
administrado pelo grupo Votorantim e utilizado para geração de energia às fábricas de
cimento. A bacia conta ainda, com unidades de conservação estaduais, municipais e com
a Floresta Nacional de Ipanema (FLONA de Ipanema), importante área de conservação
localizada no município de Iperó, na qual está localizado o reservatório Hedberg.

23
Figura 2. Localização das seis sub-bacias da UGRHI 10 (Fonte: Redes das Águas).
Caracterização dos reservatórios estudados
Três reservatórios inseridos na Bacia Hidrográfica do Sorocaba/Médio Tietê
foram inicialmente selecionados, considerando-se os seus diferentes estados tróficos.
Suas características morfométricas, profundidade máxima e média, área, ano de
construção e finalidade de utilização constam da tabela 1.
Tabela 1. Características morfométricas dos três reservatórios localizados na bacia
hidrográfica do Sorocaba/Médio Tietê (UGRHI 10).
Reservatórios Ano
construção
Estado
trófico
Prof.
máx. (m)
Prof.
méd.(m)
Vol.
máx.
(m3)
Área
(km2)
Tempo
residência da
água
Área de
proteção Utilização
Itupararanga 1912(b)
Mesotrófico(d)
19(d)
7.8(b)
286
milhões(e)
936.51
(e) 4-13 meses
(b)
APA de
Itupararanga(b)
Abastecimento público
(e)
Hedberg 1811(c)
Eutrófico(d)
- 4.0(c)
- 0.113(c)
- “FLONA de
Ipanema” (c)
Paisagismo, pesquisa e
educação ambiental (c)
St. Helena 1938 (f)
Oligotrófico(a)
10,5(a)
- - - - Não inclusa Geração de energia elétrica
a: Lucinda (2003); b: Secchin (2012); c: Plano de manejo - FLONA (2016); d: Projeto Acquased; e: Pedrazzi et al. (2014);
f: Eletromemoria-USP 2015; (-): informação indisponível.

24
Itupararanga (Figs. 3A, 4C e 4D) – O reservatório de Itupararanga foi construído em
1912 pela empresa São Paulo Eletric Company conhecida como “Light”, e teve seu
funcionamento iniciado em 1914. Inicialmente utilizado para geração de energia elétrica
à população, em 1976, a Companhia Brasileira de Alumínio (CBA) ficou responsável
pela sua operação e parte da energia gerada foi concentrada para abastecer a indústria de
alumínio (Pedrazzi et al. 2014). Localizado na sub-bacia do Alto Sorocaba, região onde
a economia está fortemente baseada em práticas agrícolas, este manancial tem
fundamental importância no abastecimento público da região, atendendo cerca de
1.000.000 de pessoas nos municípios de Ibiúna, Sorocaba, Mairinque e Votorantim
(Pereira 2008). O reservatório está incluído na Área de Proteção Ambiental (APA) de
Itupararanga, criada pela Lei Estadual nº 10.100, de 01 de dezembro de 1998 e alterada
pela Lei Estadual n° 11.579 de 02 de dezembro de 2003, cujo objetivo é o uso sustentável
e conservação ambiental da sub-bacia do Alto Sorocaba, a qual está fortemente ameaçada
pelo avanço dos loteamentos e uso intensivo de agrotóxicos (Plano Manejo, APA
Itupararanga). De acordo com a classificação das águas superficiais estabelecida pela
Resolução CONAMA n° 357, o reservatório se enquadra na Classe 2, podendo ser
utilizado para abastecimento e consumo humano (após tratamento convencional),
proteção das comunidades aquáticas, recreação de contato primário (natação, esportes
aquáticos e mergulho), irrigação e atividades de pesca.
Hedberg (Figs. 3B, 4A e 4B) – inserido na Floresta Nacional de Ipanema (FLONA), uma
unidade de conservação nacional criada em 1992 e atualmente administrada pelo Instituto
Nacional do Meio Ambiente e Recursos Naturais Renováveis e da Amazônia Legal
(IBAMA), o reservatório Hedberg foi formado pelo barramento do rio Ipanema e mesmo
estando inserido em uma Unidade de conservação, sofre com despejos de matéria
orgânica proveniente de áreas urbanizadas e agrícolas existentes à montante da FLONA.
Com barragem construída em 1811, durante a administração de Carl Gustav Hedberg, é
considerado o primeiro reservatório com fins de geração de energia hidráulica do país e
o segundo da América Latina. O reservatório foi importante na geração de energia para a
produção de ferro e atualmente é um elemento fundamental na composição da paisagem,
o qual teve sua margem recomposta por espécies de plantas nativas, área anteriormente
destinada à fins agrícolas (Plano de Manejo, 2016). Além disso, apresenta uma ictiofauna
típica de ambiente lêntico composta principalmente por saguirús, carás e lambaris e por

25
ser um reservatório antigo, a comunidade de peixes apresenta elevado grau de
estruturação (Plano de Manejo, 2016).
Em 2001, técnicos do IBAMA e CETESB, detectaram o vazamento de 40 mil
litros de óleo contendo ascarel na FLONA de Ipanema. Estudos toxicológicos
demonstraram que a mesma, tecnicamente denominada de alocloro 124, era altamente
tóxica e cancerígena, e potencialmente contaminante do solo e água (subterrânea ou
superficial). Na época do vazamento, os técnicos estimaram que a contaminação afetou
uma área de 5.000 m2 internos ao FLONA, e que poderia ter atingido o lençol freático, o
rio Ipanema, além do reservatório Hedberg.
Santa Helena (Figs. 3C, 4E e 4F) - localizado no entorno do município de Votorantim e
na divisa dos municípios de Mairinque e Alumínio, o reservatório é formado pelo
represamento do rio Sorocaba e está interligado ao reservatório de Itupararanga
(Eletromemoria-USP, 2015). As encostas do reservatório são ocupadas por atividades
pecuárias restritas, por remanescentes de mata nativa, reflorestamentos e áreas de lazer,
podendo ser observada a montante as instalações da fábrica de cimentos Santa Helena
(unidade industrial do Grupo Votorantim) (Eletromemória-USP, 2015).
Figura 3 A-C. Localização dos pontos de coleta nos reservatórios de Itupararanga (A), Hedberg (B) e Santa
Helena (C). Fonte: Projeto AcquaSed- FAPESP-2009/53898-9

26
Periodicidade de amostragem e seleção dos pontos de coleta
Durante os anos de 2014 e 2015 foram realizadas quatro amostragens (Agosto/2014;
Dezembro/2014; Março/2015 e Julho/2015), nas quais foram coletadas amostras de água
em diferentes profundidades da coluna d’água, bem como no sedimento superficial. Os
pontos de coleta (Tab. 2 e Fig. 3) foram definidos em estudo prévio (projeto temático
AcquaSed – n° processo FAPESP-2009/53898-9), considerando o corpo central dos
principais braços, a entrada dos principais tributários, além da proximidade com as
estações de captação de água (Figs. 3 e 4).
Tabela 2. Pontos de coleta dos três reservatórios da Bacia Hidrográfica do
Sorocaba/Médio Tietê (Fonte: Projeto AcquaSed- FAPESP-2009/53898-9)
Reservatório Ponto Coordenadas
Sul Oeste
Santa Helena
1 23°34'49,92'' 47°25'32,64''
2 23°34'58,56'' 47°25'50,52''
3 23°34'54,30'' 47°26'12,12''
Hedberg
1 23°25'55,86'' 47°35'33,06'
2 23°25'40,86'' 47°35'31,14''
3 23°25'33,72'' 47°35'42,66''
Itupararanga
1 23°37'11,58'' 47°13'59,34''
2 23°36'51,96'' 47°18'50,22''
3 23°37'12,96'' 47°19'37,20''
4 23°38'50,36'' 47°22'14,52''
5 23°36'53,10'' 47°23'34,70''
Em cada coleta foram amostrados 11 pontos, considerando-se os três reservatórios
(Fig. 3, 4 e Tab. 2), sendo em cada um deles coletadas amostras em diferentes
profundidades da coluna d’água (superfície, profundidade média e 1m acima do fundo),
bem como do sedimento superficial (2 cm superficiais).

27
Figura 4 A-F. Reservatórios amostrados durante o período de estudo. A-B. Hedberg. C-D. Itupararanga. E-
F. Santa Helena.
Amostragem do sedimento superficial e coluna d’água
As amostras de sedimento superficial de cada ponto foram coletadas com auxílio
do testemunhador de gravidade UWITEC™ (Fig. 5A-C), aproveitando-se os dois
primeiros centímetros superficiais, os quais geralmente integram de um a dois anos de
informação (Smol 2008), enquanto as diferentes profundidades da coluna d’água foram
coletadas em garrafa de Van Dorn (Fig. 5D e 5E).

28
Figura 5 A-E. Equipamentos utilizados para coleta de água e sedimento. A-C. Testemunhador de gravidade
UWITEC. D-E. Garrafa de Van Dorn
Fatores abióticos mensurados na coluna d’ água e sedimento superficial
Água: Em cada ponto de amostragem foram medidos in situ os perfis verticais de
temperatura, pH e condutividade elétrica a cada 0,5m até a estabilização do perfil térmico
e, a partir desse ponto, a cada 1,0m até o fundo, utilizando sonda U51-Horiba. A
transparência da água foi determinada a partir da profundidade de desaparecimento do
disco de Secchi e o cálculo da profundidade da zona eufótica feito multiplicando-se por
2,7 o valor da profundidade obtido pelo disco de Secchi (Cole 1994). O oxigênio
dissolvido foi analisado adotando-se o método de Winkler modificado (Golterman &
Clymo 1969), e o nitrogênio total e o fósforo total seguiram a metodologia de Valderrama
(1981).

29
Sedimento: Os marcadores geoquímicos avaliados no sedimento superficial foram o
fósforo total, o carbono orgânico total e o nitrogênio total (Valderrama 1981, Andersen
1976). Com o objetivo de compor a heterogeneidade das estações de amostragem, as
amostras de sedimento (n = 3) foram integradas antes da realização de cada análise,
utilizando-se sempre a mesma alíquota de cada amostra individual. Os dados foram
obtidos junto ao projeto Acquased (n° processo FAPESP-2009/53898-9), com o qual este
estudo está vinculado.
Cálculo do Índice de Estado Trófico (IET) e Clorofila-a
O Índice de Estado trófico (IET) dos reservatórios foi analisado mediante a
quantificação do fósforo total e clorofila-a. Esta última, pigmento clorofilado dominante
nas algas, além de ser utilizada para caracterizar o estado de trofia de um corpo de água,
pode ser usada para estimar a composição da biomassa algal (população fitoplanctônica).
Porém, deve-se observar que a concentração de pigmentos das algas pode variar
amplamente, dependendo do metabolismo, luz, temperatura, disponibilidade de
nutrientes, além de muitos outros fatores. Além disto, pigmentos clorofilados podem se
degradar para produtos relativamente estáveis, como as feofitinas, que interferem nos
métodos de determinação da clorofila-a (Wetzel 2001). Para quantificação deste
pigmento, as amostras foram filtradas a vácuo em filtros Whatman GF/F 47 mm, sob
baixa pressão (<0,5 atm), ainda no dia de coleta. Em seguida, foi empregado o método do
etanol 90% aquecido por 5 minutos, sem maceração (Sartory & Grobellar 1984). Os
cálculos foram baseados em Golterman et al. (1978) com correção da feofitina. Optou-se
por calcular o IET sem os dados do disco de Secchi, uma vez que os valores de
transparência muitas vezes não são representativos do estado de trofia, pois esta pode ser
afetada pela elevada turbidez decorrente de material mineral em suspensão e não apenas
pela densidade de organismos planctônicos (CETESB 2013). Para a classificação do grau
de trofia foi adotado o critério proposto por Carlson (1977) modificado por Lamparelli
(2004) (Tabela 3).

30
Tabela 3. Critério utilizado para classificação da trofia dos reservatórios segundo Carlson
(1977), modificado por Lamparelli (2004).
CLASSIFICAÇÃO IET
Ultraoligotrófico IET ≤ 47
Oligotrófico 47 < IET ≤ 52
Mesotrófico 52 < IET ≤ 59
Eutrófico 59 < IET ≤ 63
Supereutrófico 63 < IET ≤ 67
Hipereutrófico IET > 67
Plaqueamento e iscagem das amostras
Em laboratório, as amostras coletadas foram iscadas com substratos celulósicos
(epiderme de cebola, palha de milho, celofane e semente de Sorghum sp.), quitinoso
(exoesqueleto de camarão), queratinoso (ecdise de cobra), bem como grãos de pólen de
Pinus sp., seguindo metodologia descrita em Milanez et al. 1989. Todos os substratos
(iscas), quando possível, foram padronizados em discos de 0,6 cm de diâmetro, dos quais
foram adicionados dois pedaços por placa, no intuito de padronizar a metodologia e
cálculos comparativos. Para as amostras de diferentes profundidades da coluna d’água
foram utilizados 30 mL para cada amostra, enquanto para o sedimento superficial,
compartimento ainda pouco analisado com relação à diversidade de organismos
zoospóricos, foram padronizados 15 g para cada amostra, os quais foram diluídos em 30
mL de água destilada autoclavada. Esta quantidade foi adotada com base nos
procedimentos de iscagem de solo para os organismos zoospóricos (Jesus et al. 2013,
Jerônimo et al. 2015).
Análises estatísticas e índices de diversidade
A abundância (A) foi determinada com base no número de ocorrências,
considerando a presença ou ausência dos táxons em cada tipo de isca, nas diferentes
profundidades da coluna d’água (superfície, meio e fundo) e sedimento superficial
(Nascimento et al. 2011, adaptado). A frequência de ocorrência foi calculada em cada
estação (seca ou chuvosa) considerando-se: (número de amostras colonizadas por um
determinado táxon/número de amostras analisadas) × 100.
A estrutura da comunidade nas diferentes estações (seca e chuvosa) foi
caracterizada pela: (i) riqueza de espécies (S); (ii) Índice de diversidade de Shannon: (H’)
= -Σpi lnpi, onde pi é a abundância da espécie i na comunidade; (iii) Equitatividade (E) =

31
H’/Hmax’, em que Hmax’ é o valor máximo de diversidade para o número de táxons
presentes; (iv) Índice de dominância de Simpson: (D) = 1 - Σ (pi)2 baseado na abundância
dos táxons (Zak & Willig 2004); e (v) Índice de similaridade de Sorensen, utilizado para
comparar a composição dos táxons entre as estações (seca e chuvosa) e compartimentos
(coluna d’água e sedimento superficial), sendo SI = [2c/(a + b)] × 100, onde “c” é o
número de táxons comuns em ambas as estações ou compartimentos, “a” é o número de
táxons exclusivos da estação seca ou coluna d’água e “b” é o número de táxons exclusivos
da estação chuvosa ou sedimento superficial (Müeller-Dumbois & Ellemberd 1974).
A análise de componentes principais (PCA) foi utilizada para ordenar os dados
abióticos a partir de uma matriz de correlação, onde os dados foram transformados para
log (x +1), exceto pelo pH. As PCoAs dos dados bióticos da coluna d’água e sedimento
superficial foram calculadas com base na distância euclidiana com base na riqueza dos
táxons, para os quais foram desconsiderados os táxons com apenas uma ocorrência ao
longo do estudo. Ambas as análises, assim como a transformação, foram realizadas no
programa PC-ORD 6.0 (McCune & Mefford, 2011).
Variáveis climáticas
Os dados das variáveis climáticas foram obtidos nas estações meteorológicas mais
próximas dos locais de amostragem, sendo os dados de Itupararanga e Santa Helena
cedidos pelo Grupo Votorantim, enquanto os de Hedberg foram disponibilizados pela
sede da Marinha no município de Ipéro. As variáveis incluíram os valores médios de
precipitação pluviométrica e temperatura do ar, os quais foram representados de forma
gráfica.
Isolamento, cultivo e manutenção dos organismos zoospóricos
Os espécimes de organismos zoospóricos, isolados em iscas, foram inoculados em
meio de cultura a partir de fragmentos de micélio e rizomicélio ou por meio da captura
de zoósporos no momento da liberação dos mesmos (Sparrow 1960). Os espécimes foram
posteriormente purificados em meio de cultura sólido contendo 100 mg/L de penicilina
G e 100 mg/L de sulfato de estreptomicina (PmTG – 0,4 g leite peptonizado, 0,4 triptona,
2,5 g glicose, 16 g ágar, 1 L água destilada; CMA com glicose – 16 g corn meal ágar, 2,5
g glicose, 1 L água destilada; YpSs – 4 g extrato de levedura, 15 g amido solúvel, 1 g
K2HPO4, 0,5 g MgSO4 x 7H2O, 1 L água destilada e/ou MP5 – 4 g maltose, 1 g peptona,

32
16 g ágar, 1 L água destilada) e então depositados na Coleção de Cultura de Algas, Fungos
e Cianobactérias do Instituto de Botânica (CCIBt) (Fig. 6A-M). Os representantes de
Oomycota foram preservados em tubos com meio de cultura sólido (MP5, CMA), e em
frascos Wheaton contendo água destilada autoclavada, nos quais foram adicionados
fragmentos de meio de cultura contendo micélio em desenvolvimento, além de sementes
de Sorghum sp. Já os isolados de Chytridiomycota foram preservados em tubos contendo
meio de cultura sólido (CMA com glicose e PmTG), além de serem criopreservados
(método descrito abaixo). Os isolados de ambos os grupos foram mantidos em sala
climatizada (17°C), além de serem estocados em geladeira a cerca de 4°C.
Figura 6 A-M. Inclusão dos isolados de Chytridiomycota na Coleção de Cultura de Algas, Fungos e
Cianobactérias do Instituto de Botânica CCIBt. A. Culturas purificadas em placas. B-K. Crescimento dos
isolados em tubos contendo meio de cultura PmTG. L-M. Armazenamento na coleção

33
Criopreservação dos isolados de Chytridiomycota
A fim de preservar os isolados e mantê-los viáveis por um longo período, optamos
por criopreservá-los de acordo com o protocolo descrito em Boyle et al. (2004) e
resumido a seguir: cada isolado de Chytridiomycota foi cultivado em meio líquido PmTG
(35 mL) em tubos de centrífuga (tubos Falcon) por duas (espécies monocêntricas) ou
três semanas (quitridias policêntricas) (Fig. 7A). Após este período, os tubos foram
centrifugados a 4000 rpm durante 20 minutos e o sobrenadante removido para obtenção
da biomassa fúngica. Durante este processo, foi preparado o meio criopreservante (10 mL
de meio líquido PmTG, 1 mL de dimetilsulfato (DMSO) e 1 mL de soro fetal bovino) do
qual, foram adicionados 6 mL em cada tubo Falcon contendo o “pellet”. Esta mistura foi
pipetada para dissolver o “pellet” e depois 1 mL foi adicionado em cada criotubo (6
criotubos por isolado). Após o período de incubação apropriado (30 min em temperatura
ambiente), todos os criotubos foram colocados em um recipiente criopreservante e depois
estocados em ultra-freezer (Fig. 7B-F).

34
Figura 7 A-F. Protocolo de criopreservação. A. Adição da solução criopreservante nos tubos Falcon®. B.
Transferência da solução contendo o “pellet” de tecido para os criotubos. C-F. Estoque dos criotubos no
container criopreservante
Extração, amplificação, sequenciamento e análises filogenéticas dos organismos
zoospóricos
O DNA genômico dos espécimes cultivados foi extraído a partir de culturas puras,
com auxílio do kit de extração “PureLink Genomic DNA Kit” (Invitrogen™). Para os
fungos zoospóricos, as regiões SSU (18S), ITS e LSU (28S) do rDNA foram
amplificadas, respectivamente, com os “primers” NS4/SR1R, ITS4/ITS5 (White et al.
1990) e LR0R/LR5 (Vilgalys 2013), enquanto as regiões RPB1 e RPB2 (mDNA) foram
amplificadas com os “primers” RPB1Af/RPB1Dr (Stiller & Hall 1997) e
RPB26.5f/RPB211br (Liu et al. 1999), respectivamente. De acordo com Liu et al. (2006),
as regiões SSU (18S), RPB1 e RPB2, são altamente conservadas e fornecem informações

35
filogenéticas com alto grau de confiança para altos níveis hierárquicos (ordens, classes e
filos), enquanto as regiões ITS e 28S podem ser utilizadas para análises filogenéticas mais
precisas (espécies, gêneros e famílias) (James et al. 2006). Já para os representantes de
Oomycota, as regiões ITS, LSU e COX1 foram amplificadas com os “primers” UN-
up18S42/UN-lo28S22 (Levesque & de Cock 2004), LR0R/LR6-O (Riethmüller et al.
2002) e OomCoxI-Levup/OomCoxI-Levlo (Robideau et al. 2011), respectivamente.
Robideau et al. (2011), apontam que a região COX1 do mDNA juntamente com a ITS do
rDNA podem ser utilizadas para identificação dos táxons em nível especifico e que, em
alguns casos, o COX1 pode ser mais efetivo do que a região ITS. As reações de PCR
foram realizadas com o kit “PCR Supermix” (Invitrogen©) e os parâmetros inseridos no
termociclador foram os descritos em James et al. (2006): 94ºC por 1 minuto, seguido por
35 ciclos de desnaturação a 94ºC por 30 segundos, anelamento a 55ºC por 30 segundos,
extensão a 72ºC por 5 minutos e extensão final a 72ºC por 10 minutos. Posteriormente,
os amplicons foram purificados com o kit AxyPrep PCR Clean-up (Axygen©). As
sequências consenso foram geradas no programa “Sequencher™ 4.1.4” (Gene Codes),
enquanto o alinhamento foi realizado online pela ferramenta MAFFT 7.058 (Kazutaka &
Daron 2013). As diferentes regiões gênicas foram concatenadas no SequenceMatrix 1.8
(Vaidya et al. 2010), enquanto a escolha dos modelos de substituição de nucleotídeos foi
realizada no programa JModelTest 2.2.4. Os filogramas de Máxima Verossimilhança
foram construídos no Garli 2.01 (Bazinet & Cummings 2008) (para análises
concatenadas) e no PhyML 3.1 (Guindon and Gascuel 2003) para análises de uma única
região gênica.
Ultraestrutura dos zoósporos de representantes de Chytridiomycota
A ultraestrutura dos zoósporos é um tipo de análise onde se observa a presença ou
ausência de determinadas organelas, bem como sua posição relativa dentro da célula,
através de microscopia eletrônica de transmissão. Esta ferramenta é usualmente utilizada
no estudo dos fungos zoospóricos (Chytridiomycota, Blastocladiomycota e
Neocallimastigomycota), pois alguns autores como Li et al. (1993) e Letcher et al. (2005)
admitem que as características ultraestruturais são evolutivamente mais conservadas do
que a morfologia do talo e, portanto, fornecem informações importantes sobre a inter-
relação dos táxons.

36
O passo mais crítico para análise da ultraestrutura dos zoósporos é justamente a
produção de quantidade suficiente de zoósporos em meio de cultura sólido. Ao longo do
processo ocorrem inúmeras perdas, e caso a quantidade inicial não seja adequada,
dificilmente será possível observar todas as estruturas no final do processo. Com o intuito
de estimular a produção máxima de zoosporângios e zoósporos, foi preparado o meio de
cultura YpSs/8 utilizando água de solo, assim como mencionado por Lucarotti (1981).
De acordo com este autor, este meio foi capaz de estimular ou aumentar a produção de
zoosporângios e zoósporos nas espécies Cladochytrium replicatum Karling e
Nowakowskiella elegans Nowak. O meio foi preparado aquecendo-se 7 g de solo de
jardim (~65°C) com 0,05 g de carbonato de cálcio em 250 mL de água destilada durante
2 horas em 2 dias consecutivos. Esta mistura foi então filtrada duas vezes com filtro de
papel Whatman® n°.1, diluída (uma parte de água de solo para duas partes de água
destilada) e por fim autoclavada (Fig. 8A-F).
Figura 8 A-F. Preparo do meio YPSS/8 utilizando água de solo. A-C. Aquecimento do solo. D-E. Filtragem
do solo. F. Plaqueamento do meio YPSS/8

37
As análises de ultraestrutura dos isolados de Chytridiomycota foram realizadas
em parceria com o laboratório do Dr. Timothy Y. James (Evolutionary Genetics of Fungi
Laboratory) em Michigan (EUA), durante o período de Bolsa de Estágio de Pesquisa no
Exterior (BEPE), financiada pela Fapesp (n° processo FAPESP 2016/25800-8). O
protocolo utilizado está descrito e ilustrado com detalhes abaixo, por ser o primeiro desta
natureza descrito totalmente em português.
Protocolo de Microscopia Eletrônica de Transmissão (Chytridiomycota)
Crescimento dos isolados e coleta de zoósporos (Fig. 9A-C): Vinte placas de Petri
contendo meio de cultura sólido (PmTG) foram inoculadas com culturas puras, e então
incubadas a aproximadamente 25°C. Entre 7 e 15 dias, ou assim que os zoosporângios
tiverem crescido por toda a extensão da placa, as mesmas foram submersas com água
deionizada ultrapura (1 mL por placa) (Fig. 9A) e então, o líquido contendo os zoósporos
foi coletado após 1 hora de submersão, o qual foi adicionado a um tubo Falcon® (Fig.
9B, C).

38
Figura 9 A-C. Crescimento e coleta de zoósporos de quitridias A. Submergindo as placas. B-C. Fixando os
zoósporos com gluteraldeido 3%
Preparo dos reagentes: para as etapas de fixação é necessária a preparação prévia dos
reagentes descritos abaixo (Fig. 10A-G)
1. Tampão 0,2 M sym-colidina: foi utilizado o kit s-colidina comercializado pela
EMS© (Electron Microscopy Sciences). O kit contém ampolas de 9 mL de ácido
clorídrico (HCL a 2N) e de 5,34 mL de s-colidina. A solução foi preparada pela
adição de uma ampola de 9 mL de HCL (2N) e 1 ampola de 5,34 mL de s-colidina
em uma Becker de 500 mL de capacidade. Foram então adicionados 186 mL de
água deionizada ultrapura, resultando em uma solução tampão final de 200 mL.

39
2. Tampão 0,1 M sym-colidina: 50 mL da solução tampão a 0,2 M de s-colidina
(passo anterior, 1) foi diluída em 50 mL de água deionizada ultrapura, resultando
em uma solução tampão final de 100 mL de s-colidina a 0,1M.
3. Tampão 0,05 M sym-colidina: 25 mL da solução tampão de 0,1 M (passo 2) de s-
colidina foi diluída em 75 mL de água deionizada ultrapura, resultando em uma
solução tampão final de 100 mL de s-colidina a 0,05M.
4. Tampão de gluteraldeído 3% em 0,1 M de s-colidina: uma ampola de 10 mL de
gluteraldeído a 50% foi diluída em 156 mL de tampão s-colidina a 0,1 M,
resultando em 166 mL de gluteraldeído a 3%.
5. Acetona 100% com esferas de cerâmica (3A): a acetona 100% foi preparada com
adição de esferas de cerâmica (tamanho 3A) para remoção da água residual. Este
reagente deve ser preparado na semana que antecede o início do protocolo de
fixação.
6. Solução de tetróxido de ósmio 1%: foi utilizado o tetróxido de ósmio da EMS©.
Uma ampola de 2 mL (4% de concentração inicial) foi diluída em 2 mL de água
deionizada ultrapura, resultando em uma solução de 4 mL (2% de concentração).
Essa solução foi diluída novamente em mais 1 mL de água deionizada ultrapura,
resultando então na solução de trabalho de 1% de tetróxido de ósmio.
7. Acetato de uranila a 2%: foi utilizado o acetato de uranila (EMS©, 25 g UrAc), o
qual foi diluído em 10 mL de etanol 70% até a saturação. Essa solução adicionada
em um frasco âmbar, para evitar contanto com a luz, e então mantida em
temperatura ambiente.

40
Figura 10 A-G. Preparo dos reagentes. A-D. Preparo do gluteraldeído a 3%. E. Diluições de s-colidina. F.
Acetona a 100%. G. Acetato de uranila a 2%
Fixação dos zoósporos: igual volume de gluteraldeido 3% (20 mL) foi adicionado aos
tubos Falcon contendo a suspensão de zoósporos removidos das placas. Essa solução
penetra e fixa os zoósporos em cerca de hora de contato, mas para otimização dos efeitos,
os frascos foram mantidos na geladeira (~5°C) durante a noite (Fig. 9B, C).
Lavagem: o material fixado foi subsequentemente lavado por três vezes com solução
tampão de 0,1 M de s-colidina para remoção do gluteraldeído residual. Após cada
procedimento de lavagem, os zoósporos foram centrifugados a ~3G para formação de
“pellet” e o sobrenadante descartado, com posterior resuspensão do “pellet” em tampão
de s-colidina a 0,1 M.

41
Osmificação (Fig. 11A-D): O material anteriormente fixado e lavado foi então pós-fixado
em 1% de tetróxido de ósmio e mantido na geladeira durante a noite. Assim que a solução
de tetróxido de ósmio entra em contato com o “pellet’ de zoósporos, o mesmo adquire
uma coloração escurecida (Fig. 11D). Após o período de osmificação, o material foi
lavado uma vez com s-colidina a 0,1 M, seguido por três lavagens com água deionizada
ultrapura, para remoção de qualquer resíduo de tetróxido de ósmio.
Figura 11 A-D. Osmificação. A-C. Pós-fixação com 1% de tetróxido de ósmio. D. Reação do tetróxido de
ósmio com o “pellet” de zoósporos
Infusão em ágar (Fig. 12 A-P): Após as lavagens da etapa de pós-fixação, o “pellet” foi
infundido com 2% de ágar nobre. Para preparação do ágar nobre, 25 mL de água
deionizada ultrapura foi misturado com 0,5 g de ágar nobre e aquecido até o ágar se

42
dissolver por completo (Fig. 12A-E). Ainda quente, o ágar nobre foi adicionado em um
tubo de vidro de fundo cônico contendo o “pellet” de zoósporos, o qual foi deslocado para
o meio da coluna de ágar (esse talvez seja o passo mais delicado do processo, pois o
“pellet” pode se desfazer facilmente, diluindo tanto a suspensão de zoósporos, que os
encontrar no final seja muito complicado). Uma vez que “pellet” tenha sido infundido em
ágar aquecido, ele foi colocado na geladeira por 15 minutos para solidificar. Após a
solidificação do ágar, o bloco de ágar contendo o “pellet” no centro, foi removido do tubo
de vidro e colocado em uma placa de Petri (Fig. 12G-I). O material infundido foi
dissecado em pequenos blocos de ágar (Fig. 12J-N), os quais foram colocados em um
recipiente de plástico contendo 16 mL de água deionizada ultrapura (Fig. 12O-P).
Figura 12 A-P. Infusão do “pellet” com 2% de ágar nobre. A-E. Preparação do ágar nobre. F. Adição de
ágar nobre líquido. G-I. Remoção do ágar solidificado do tubo de vidro. J-N. Dissecção do bloco de ágar
em blocos menores. O-P. Adição de água deionizada ultrapura

43
Desidratação: para remoção de toda a água do passo anterior, um agente desidratante
(acetona) foi adicionado em uma serie gradual de 10%, 30%, 50%, 70%, 85%, 95% de
concentração durante 20 minutos em cada diluição, além de duas adições de acetona
100%, por 20 minutos, após as etapas anteriores.
Preparo da resina Epon-Araldite (Fig. 13A-P): essa resina foi preparada usando cinco
reagentes, os quais foram adicionados sequencialmente e misturados por 5 minutos, entre
cada nova adição (Fig. 13A-P). Inicialmente foram adicionados 11 mL de anidrido
dodecenil-succínico (DDSA) (Fig. 13B-F), seguido por 5,2 mL de resina Epon (812) (Fig.
13G-I), 8,1 mL de araldite (Fig. 13J-L), 0,75 mL de dibutilftalato (Fig. 13M) e 27 gotas
de Tris-dimetilaminometil fenol (DMP 30).
Figura 13A-P. Preparo da resina. A-F. Adição de DDSA. G-I. Adição de resina Epon 182. J-L. Adição de
Araldite. M. Adição de dibutilftalato. O-P. Resina finalizada após adição de DMP 30

44
Infiltração dos blocos com series diluídas de resina (Fig. 14A-M): a resina EPOX é o
material mais adequado para a preparação dos moldes finais, pois ela é solúvel em acetona
e, além disso, é um material fácil de seccionar, além de ser relativamente estável em feixe
de elétrons. O material desidratado (em acetona 100%) foi infiltrado em série graduada
de resina EPOX de 25% (25% de resina, 75% de acetona) por 2 horas, 50% (50% de
resina e 50% de acetona) e 75% (75% de resina, 25% de acetona) por 2 horas cada e 100%
de resina durante a noite, para eliminar todos os vestígios de acetona.
Figura 14 A-M. Infiltração dos blocos de ágar com diferentes diluições de resina
Preparo dos moldes (Fig. 15A-U): O material biológico totalmente infiltrado, bem como
as legendas correspondentes e o molde de inclusão (Fig. 15A-U), foram colocados na
máquina de vácuo durante a noite para remover quaisquer vestígios de bolhas de ar (Fig.
15D, E, F). Depois disso, o material biológico foi adicionado no molde, juntamente com

45
as legendas (Fig. 15G-Q) e então preenchido com resina Epon-Araldite recém-preparada
(Fig. 15F), a qual foi posteriormente polimerizada durante 72 horas a 65ºC. Os melhores
blocos foram selecionados para serem seccionados (Fig. 15R-U).
Figura 15 A-U. Última etapa da preparação da resina e adição dos blocos de ágar nos moldes
Secção dos blocos moldados (Fig. 16 A-H): A secção dos blocos moldados foi realizada
em um ultra micrótomo (Fig. 16A-H). O material foi inicialmente aparado (Fig. 16C-E)
à mão, até formação de um trapézio de aproximadamente 0,1 mm. O bloco foi então
aparado no ultra micrótomo (Fig. 16F) e depois cortado em secções de 80-120 nm, por
uma faca de diamante. Estas secções em série foram recolhidas em anéis de níquel (malha
100-300) (Fig. 16G-H) para então serem coradas com citrato de chumbo a 2%.

46
Figura 16 A-H. Secção dos blocos moldados em um ultra micrótomo e adição das secções em anéis de
níquel
Corando as secções (Fig. 17A-J): As secções foram então coradas durante 5 min em 2%
de citrato de chumbo para obtenção de contraste. Depois disso, as secções foram lavadas
duas vezes com água deionizada ultrapura para remoção de todos os vestígios de citrato
de chumbo.

47
Figura 17 A-J. Corando as secções com 2% de citrato de chumbo
Análise por microscopia eletrônica de transmissão: Depois das secções totalmente secas,
as mesmas foram examinadas em microscópio eletrônico de transmissão (modelo JEL-
1400) a 80 kV. Secções aleatórias foram observadas para obtenção de imagens
ultraestruturais.

48
REFERÊNCIAS BIBLIOGRÁFICAS
Amal, E. A., Aïcha, E. A. & Bernard, P. 2005. Achlya abortispora, a new Oomycete
isolated from water samples taken from a water reservoir in Morroco. Current
Microbiology 53: 60-67.
Amaral, A.M., Barbério, A., Voltolini, J.C. & Barros, L. 2007. Avaliação preliminar
da citotoxicidade e genotoxicidade da água da bacia do rio Tapanhon (SP-Brasil)
através do teste Allium (Allium cepa). Revista Brasileira de Toxicologia 20: 65‑72.
Birks, H.H. & Birks, H.J. 2006. Multi-proxy studies in palaeolimnology. Vegetation
History Archaeobotany 15: 235-251.
Carlson, R.E. 1977. A trophic state index for lakes. Limnology and Oceanography, 22:
361-369.
CEPAGRI – Centro de Pesquisas Meteorológicas e Climáticas Aplicadas a
Agricultura (2018). Disponível online em: http://www.cpa.unicamp.br/outras-
informacoes/clima-dos-municipios-paulistas.html. Acessado em 06 Junho de 2018.
CETESB – Companhia Ambiental do Estado de São Paulo. 2013. Relatório de
qualidade das águas superficiais no Estado de São Paulo. Parte 1. São Paulo, 301 p.
Chen, M., Chen, F., Yu, Y., et al. 2008. Genetic diversity of eukaryotic microorganisms
in Lake Taihu, a large shallow subtropical lake in China. Microbiology Ecology 56:
572−583
CONAMA, Conselho Nacional do Meio Ambiente. 2005. Resolução n° 357, de 17 de
março de 2005. Ministério do Meio Ambiente. p. 23.
Colins, C. & Willoughby, L.G. 1962. The distribution of bacteria and fungal spores in
Blelham Tarn with particular reference to an experimental overturn. Archiv für
Mikrobiologie 43: 294-307.
Cooke, W.B. 1976. Fungi in sewage. In. Gareth Jones, E.B (Ed.). recent advances in
Aquatic Mycology. London: Elek Science p. 389-434.
Czeczuga, B. Woronowicz, L., Brzozowska, K. & Chomutowska, H. 1989. Studies of
aquatic fungi IX. Mycoflora of different types springs. Acta Hydrobiologica 31: 273-
283.
Czeczuga, B. Woronowicz, L., Brzozowska, K. 1990a. Studies of aquatic fungi XII.
Aquatic fungi of the lowland river Biebrza. Acta Hydrobiologica 26: 77-83.

49
Czeczuga, B. Woronowicz, L., Brzozowska, K. 1990b. Studies of aquatic fungi XIII.
Mycoflora of river Czarna Hancza and its tributary, the river Marycha. Internationale
Revue der gesamten Hydrobiologie 75: 245-255.
Czeczuga, B. 1991a. Studies of aquatic fungi. Part XVI. Aquatic fungi of the river Pisa
and its tributary, the river Skroda. Acta hydrochimica et hydrobilogica 19: 57-65.
Czeczuga, B. 1991b. Studies of aquatic fungi. Part XXII. Mycoflora of the river
Wegorapa and its tributary, the river Goldapa-Jarka. Acta hydrochimica et
hydrobiologica 19: 517-528.
Czeczuga, B. & Woronowicz, L. 1991. The lake Mamry complex. Acta Mycologica 27:
93-103.
Czeczuga, B. 1994. Studies of aquatic fungi. Part XVII. Aquatic fungi of Lake Hancza
in the Suwalki Scenic Park and some adjacents lakes (northeastern Poland). Acta
Hydrobiologica 36: 371-385.
Czeczuga, B. & Muszynska, E. 2001. Zoosporic fungi growing on Gymnosperm pollen
in water of variety trophic state. Polish Journal of Environmental Studies 10: 89-94.
Dawson, S.C. & Pace, N.R. 2002. Novel Kingdom-level eukaryotic diversity in anoxic
enviroments. Proceedings Natural Academic Sciences of USA 99: 8324-8329.
Esteves, F.A. 2011. Fundamentos de limnologia. Rio de Janeiro. Interciencia. 3 ed. 826p.
FLONA – Floresta Nacional de Ipanema, Plano de Manejo. 2016. Instituto Chico
Mendes de Preservação da Biodiversidade.
Gerphagnon, M., Latour, D., Colombet, J. & Sime-Ngando, T. 2013. A double
staining method using SYTOX green and calcofluor white for studing fungal parasites
of phytoplankton. Applied and Environmental Microbiology
dx.doi.org/10.1128/AEM.00696-13.
Gleason, F. H., Karpov, S.A., Lilje, O., Macarthur, D.J. et al. 2014. In: Gareth Jones,
E. B., Hyde, K. D. e Pang, K. (Eds.). Zoosporic parasites of phytoplankton. Walter
de Gruyter GmbH, Berlin/Boston, pp.279-304.
Goedkoop, W., Widenfalk, A., Haglund, A.L., et al. 2005. Microbial characterization
of artificial sediment and comparisons with natural sediments – implications for
toxicity test. Environmental Taxicological Chemistry 24: 2725-2733.
Golterman, H.L. & Clymo, R.S. 1969. Methods for chemical analysis of freshwaters.
Oxford and Edinburg: Blackwell Scientific Publications. International Biological
Programme. 171p.

50
Grossart, H.P. & Rojas-Jimenez, K. 2016. Aquatic fungi: targeting a forgotten in
microbial ecology. Current opinion in Microbiology 31: 140-145.
James, T.Y., Letcher, P.M., Longcore, J.E., Mozley-Standridge, S.E. et al. 2006. A
molecular phylogeny of the flagellated fungi (Chytridiomycota) description of a new
phylum (Blastocladiomycota). Mycologia 98: 860-871.
Jerônimo, G.H., Jesus, A.L., Marano, A.V., et al. 2015. Diversidade de
Blastocladiomycota e Chytridiomycota do Parque Estadual da Ilha do Cardoso,
Cananéia, Estado de São Paulo, Brasil. Hoehnea 42: 135-163.
Jerônimo, G.H., Jesus, A.L., Rocha, S.C.O. Gonçalves, D.R. et al. 2017. New insights
into Plectospira genus (Oomycetes, Straminipila): morphological and molecular
analysis. Phytotaxa 307: 191-198.
Kagami, M., E. Van Donk, A. de Bruin, M. Rijkeboer & B. W. Ibelings, 2004.
Daphnia can protect diatoms from fungal parasitism. Limnology and Oceanography
49: 680-685.
Kagami, M., Bruin, A., Ibelings, B.A. & Donk, E.V. 2007. Parasitic chytrids: their
effects on phytoplankton communities and food-web dynamics. Hydrobiologia: 578:
113-129.
Kagami, M.; Helmsing, N. R.; Van Donk, E. 2011. Parasitic chytrids could promote
copepod survival by mediating material transfer from inedible diatoms.
Hydrobiologia 659: 49-54.
Kagami, M., Miki, T. & Takimoto, G. 2014. Mycoloop: chytrids in aquatic food webs.
Frontiers in Microbiology doi: 10.3389/fmicb.2014.00166.
Karling, J. S. 1977. Chytridiomycetarum Iconographia. Lubrecht & Cramer, Vaduz.
Lamparelli, M.C. 2004. Grau de trofia em corpos d’água do estado de São Paulo:
avaliação dos métodos de monitoramento.Tese de doutorado, Universidade de São
Paulo, 235 p.
Lefèvre, E., Bardot, C., Noël, C., Carrias, J.F., Viscogliosi, E. et al. 2007. Unveiling
fungal zooflagellates as members of freshwater picoeucakaryotes: evidence from a
molecular diversity study in a deep meromictic lake. Environmental Microbiology: 9:
61-71.
Lefèvre, E., Letcher, P. & Powell, M. 2012. Temporal variation of small eukaryotic
community in two freshwater lakes: emphasis on zoosporic fungi. Aquatic microbial
Ecology67: 91-105.

51
Lefranc, M., Thénot, A., Lepère, C. & Debroas, D. 2005. Genetic diversity of small
eukaryotes in lakes differing by their trofic status. Application Environmental
Microbiology 71: 5935-5942.
Lepère, C., Domaizon I. & Debroas, D. 2008. Unexpected importance of potencial
parasites in the composition of the freshwater small-eukaryote community. Applied
and Environmental Microbiology 74: 1029-1041.
Letcher, P.M., Powell, M.J., Churchill, P.F., Chambers, J.G. 2006. Ultrastructural and
molecular phylogenetic delineation of a new order, the Rhizophydiales
(Chytridiomycota). Mycological Research 110: 898-915.
Letcher, P.M., & Powell, M.J. 2014. Hypothesized evolutionary trends in zoospore
ultrastructural characters in Chytridiales (Chytridiomycota). Mycologia, 106: 379-
396.
Lucarotti, C.J. 1981. Zoospore ultrastructure of Nowakowskiella elegans and
Cladochytrium replicatum (Chytridiales). Canadian Journal of Botany, 59: 137-148.
Lucinda, I. 2003. Composição de Rotifera em corpos d’água da bacia do rio Tietê – São
Paulo, Brasil. Dissertação de mestrado. Universidade Federal de São Carlos: São
Carlos. 199p.
Marano, A.V., Zottarelli, C.L.A., Barrera, M.D. et al. 2011. Diversity, role in
decomposition, and sucession of zoosporic fungi and straminipiles on submerged
decaying leaves in a woodland stream. Hydrobiologia 659: 93-109.
Margalef, R. 1983. Limnologia. Omega, Barcelona.
McCune, B. & Mefford, M.J. 2011. PC-ORD. Multivariate analysis of ecological data.
Version 6. MjM Software, Gleneden Beach, Oregon, USA.
Milanez, A.I. 1989. Fungos de águas continentais. In: O. Fidalgo, V.L. Bononi (coords.).
Técnicas de coleta, preservação e herborização de material botânico. Série
Documentos. São Paulo, Instituto de Botânica. pp. 17-20.
Monchy, S., Sanciu, G., Jobard, M., Rasconi. et al. 2011. Exploring and quantifying
fungal in freshwater lake ecosystems using rDNA cloning/sequencing and SSU tag
pyrosequencing. Environmental Biology 13: 1433-1453.
Müeller-Dombois, D. & Ellemberd, H. 1974. Aims and methods of vegetation ecology.
New York, Jonh Wiley & Sons. pp. 547.

52
Nascimento, C.A., Gomes, E.P.C. & Pires-Zottarelli, C.L.A. 2011. Occurrence and
distribution of zoosporic organisms in water bodies from Brazilian Cerrado.
Mycologia 103: 261-272.
Pedrazzi, F.J.M., Conceição, F.T., Sardinha, D.S., Moschini-Carlos, V. et al. 2014.
Avaliação da qualidade da água do reservatório de Itupararanga, Bacia do Alto
Sorocaba. Geociências 33: 26-38.
Pires-Zottarelli, C.L.A. 1990. Levantamento dos fungos zoospóricos da Represa do
Lobo (“Broa”), São Carlos, SP, Brasil [Master dissertation]. São Paulo: Universidade
Estadual Paulista. 175 p.
Plaats-Niterink, V.J. 1981. Monograph of the genus Pythium. Studies in Micology 21:
1-244.
Pusceddu, A., Gambi, C., Manini, E. & Danovaro, R. 2007. Trophic state, ecosystem
efficiency and biodiversity of transitional aquatic ecosystems: analysis of
environmental quality based on different benthic indicators. Journal of Chemical
Ecology 23: 505‑515.
Rasconi, S. Jobard, M., Jouve, L. & Sime-Ngando, T. 2009. Use of calcofluor white
for detection, identification, and quantification of phytoplanktonic fungal parasites.
Applied and Environmental Microbiology 75: 2545–2553.
Rasconi, S.; Niquil, N.; Sime-Ngando, T. 2012. Phytoplankton chytridiomycosis:
community structure and infectivity of fungal parasites in aquatic ecosystems.
Environmental Microbiology, Bedford, v. 14, p. 2151–2170.
Rasconi, S., Grami, B., Niquil, N., Jobard, M. et al. 2014. Parasitic chytrids sustain
zooplankton growth during inedible algal bloom. Frontiers in Microbiology 5: 1-20.
Rocha, M. & Pires-Zottarelli, C.L.A. 2002. Chytridiomycota o Oomycota da represa
do Guarapiranga. Acta Botanica Brasilica 16: 287-309.
Rocha, M. 2004. Micota zoospórica de lagos com diferentes trofias do Parque Estadual
das Fontes do Ipiranga (PEFI), São Paulo, SP. Dissertação para obtenção do título de
Mestre em Microbiologia para o Instituto de Ciências Biomédicas da Universidade de
São Paulo, p. 1-85.
Sartory & Groberlaar. 1984. Extraction of chlorophyll-a from freshwater
phytoplankton for spectrophotometric analysis. Hidrobiology 114: 177-187.

53
Secchin, L.F. 2012. Caracterização ambiental e estimativa da produção de cargas difusas
na área de drenagem da represa de Itupararanga. Dissertação de mestrado.
Universidade de São Paulo: São Paulo. 122p.
Shearer, C.A., Descals, E., Kohlmeyer, B., Kohlmeyer, J. et al. 2007. Fungal
biodiversity in aquatic habitats. Biodiversity Conservation 16: 49-67.
Sime-Ngando, T., Lèfevre, E. & Gleason, F. 2011. Hidden diversity among aquatic
heterotrophic flagellates: ecological potentials of zoosporic fungi. Hydrobiologia
659: 5-22.
Slapeta, J., Moreira, D., Purificación, L.G. 2005. The extend of protist diversity:
insights from molecular ecology and freshwater. Proceedings of the Royal Society B
272: 2073-2081.
Smol, J.P. 2008. Pollution of lakes and rivers: a paleoenvironmental perspective.
Blackewell, 2 ed., Oxford University Press, New York.
Sparrow. F. K. 1968. The Fungi: An Advanced Treatise. In: Ainsworth, G.C. Sussman,
A.S. (eds.). The Fungal Population III, Academic Press, New York.
Tolmasquim, M.T. 2012. Perspectivas e planejamento do setor energético no Brasil.
Estados Avançados 26: 247-260.
Tundisi, J.G., Matsumura-Tundisi, T. & Tundisi, J.E.M. 2008. Reservoirs and human
well being: new challenges for evaluating impacts and benefits in the neotropics.
Brazilian Journal of Biology 68: 1133-1135.
Valderrama, G.C. 1981. The simultaneous analysis of total nitrogen and total
phosphorus in natural waters. Marine Chemistry 10: 109-122.
Wurzbacher, C.M., Barlocher, F. & Grossart, H.P. 2010. Fungi in lake ecosystems.
Aquatic microbial Ecology 59: 125-149.
Wurzbacher, C.M., Whartmann, N., Bourne, E., Attermeyer, K. et al. 2016. High
habitat-specific in fungal communities of oligo-mesotrophic temperate Lake Stechlin
(North-West Germany). Mycokeys 16: 17-44.
Wetzel, R.G. 2001. Structure and productivity of aquatic ecossystems. In: Limnology.
Academic Press, San Diego, pp. 206.
Widenfalk, A., Bertilsson, S., Sundh, I. & Goedkoop, W. 2008. Efects of pesticides on
community composition and activity of sediment microbes – response at various
levels of microbial community organization. Enviromental Polution 152: 576-584.
Wong, M.K.M., Goh, T.K., Hodgkiss, I.J. et al. 1998. Role of fungi in freshwater
ecossystems. Biodiversity and Conservation 7: 1187-1206.

54
Yang, X., & Hong, C.X. 2015. Diversity and populations of Phytophthora, Phytopythium
and Pythium species recovered from sediments in an agricultural run‐off
sedimentation reservoir. Plant Pathology 65: 1118-1125.
Zak, J.C. & Willig, M.R. 2004. Fungal biodiversity patterns. In: G.M. Mueller, G.F.
Bills, M.S. Foster (eds.). Biodiversity of fungi: Inventory and monitoring methods.
Elsevier Academic Press, San Diego, pp. 59–75.
Zappia, R.E., Hüberli, D., Hardy, G.S.J. & Bayliss, K.L. 2014. Fungi and oomycetes
in open irrigation systems: knowledge gaps and biosecurity implications. Plant
Pathology 63: 961-972.

55
CAPÍTULO 1
ESTRUTURA DA COMUNIDADE DE ORGANISMOS ZOOSPÓRICOS
HETEROTRÓFICOS (FUNGOS ZOOSPÓRICOS E OOMICETOS) DA
COLUNA D’ÁGUA E SEDIMENTO SUPERFICIAL DE RESERVATÓRIOS
COM DISTINTOS ESTADOS TRÓFICOS

56
Estrutura da comunidade de organismos zoospóricos heterotróficos (fungos
zoospóricos e oomicetos) da coluna d’água e sedimento superficial de reservatórios
com distintos estados tróficos
Autores: Jerônimo, G.H1. Ferragut, C2. & Pires-Zottarelli, C.L.A1.
1 Núcleo de Pesquisa em Micologia, Instituto de Botânica, São Paulo, Brasil, 04301-902
2 Núcleo de Pesquisa em Ecologia, Instituto de Botânica, São Paulo, Brasil, 04301-902
RESUMO
Historicamente, a maior parte do conhecimento sobre a produtividade dos ecossistemas
lênticos tem sido focada nos organismos autotróficos e produtores primários. Os
decompositores e parasitas, embora apresentem um papel importante na dinâmica da
cadeia alimentar, tem sido ao longo dos tempos negligenciados. Sequenciamentos de
amostras ambientais tem revelado que os fungos basais (Blastocladiomycota e
Chytridiomycota) representam a maior parte da micota presente nestes ecossistemas,
sendo juntamente com os oomicetos, cruciais para a ciclagem de nutrientes. No entanto,
pouco se sabe sobre a influência dos fatores ambientais sobre os padrões de ocorrência,
frequência e abundância destes organismos. Diante disto, o objetivo deste estudo foi
relacionar a riqueza e a abundância destes grupos de organismos com a sazonalidade e
alguns fatores abióticos mensurados na coluna d’água e sedimento superficial de três
reservatórios brasileiros (Santa Helena, Itupararanga e Hedberg), conhecidamente com
distintos estados tróficos. Para isso, foram realizadas quatro coletas em cada reservatório,
onde foram mensuradas algumas variáveis físicas e químicas da água (temperatura, pH,
condutividade elétrica, oxigênio dissolvido, nitrogênio e fósforo total) e do sedimento
superficial (nitrogênio total, fósforo total e carbono orgânico total). A estrutura da
comunidade foi avaliada considerando-se a ocorrência, frequência e abundância dos
táxons, mediante cálculo da riqueza de espécies, do índice de diversidade de Shannon, do
índice de dominância de Simpson, da equitatividade e do índice de similaridade de
Sørensen. Durante o estudo, 52 táxons de organismos zoospóricos heterotróficos foram
identificados, sendo 25 deles representantes dos filos Blastocladiomycota e
Chytridiomycota (fungos zoospóricos) e 27 do filo Oomycota. Dentre os fungos
zoospóricos, uma nova espécie do gênero Nowakowskiella J. Schröt foi descrita e o
gênero Avachytrium Velez & Letcher relatado pela primeira vez para o Brasil, assim
como Plectospira gemmifera Dreschler o foi dentre os oomicetos. Foi observada uma

57
comunidade diversa com poucos táxons dominantes, sendo a maior parte deles observada
na estação chuvosa, dentre os quais, Cladochytrium replicatum Karling, Nowakowskiella
hemisphaerospora Shanor, Aphanomyces spp., Dictyuchus spp. e Pythium dos grupos P
e T, foram os táxons mais frequentes e abundantes, considerando os reservatórios e
estações (seca e chuvosa). Os organismos zoospóricos apresentaram uma ampla
distribuição ao longo de diferentes gradientes tróficos, existindo espécies estritamente
relacionadas aos locais com as mais altas concentrações de nitrogênio e fósforo total, e
outras aos locais com as mais baixas concentrações destes nutrientes, e que
potencialmente poderiam ser táxons indicativos da qualidade das águas. Concluímos,
portanto, que os organismos zoospóricos heterotróficos são, no geral, tolerantes às amplas
variações nas concentrações de nitrogênio e fósforo, mas alguns táxons são limitados pela
disponibilidade destes nutrientes, particularmente o fósforo.
Palavras-chave: Blastocladiomycota, Chytridiomycota, ecologia, Oomycota
ABSTRACT
Historically, the most part of the knowledge about lentic ecosystems productivity has
been based on autotrophic organisms and primary producers. The decomposers and
parasites, although important in food chain dynamics, have been negligence in diversity
studies. Environmental sequencing data have been revealed that the basal fungi
(Blastocladiomycota e Chytridiomycota) compound the majority mycota in these
ecosystems, and together with oomycetes, are crucial on nutrient cycling. However, a few
is knowledge about the influence of the abiotic factors and seasonality on occurrence,
frequency and abundance patterns of these organisms. Considering this, the aim of this
study was to relate the species richness and abundance of these groups with seasonality
and some abiotic factors measured in the water column and surface sediment of three
Brazilian reservoirs (Santa Helena, Itupararanga and Hedberg), with distinctive trophic
states. In order to do this, four sampling were performed in each reservoir, in which
physical and chemical variables of the water (temperature, pH, electrical conductivity,
dissolved oxygen, nitrogen and total phosphorus) and surface sediment (total nitrogen,
total phosphorus and total organic carbon) were measured. The community structure was
evaluated considering the abundance, frequency, species richness, Shannon’s diversity
index, Simpson’s index, Evenness and similarity Sorensen index. During the study, 52
taxa of heterotrophic zoosporic organisms were identified, 25 of which representative of

58
Blastocladiomycota and Chytridiomycota phyla (zoosporic fungi) and 27 of Oomycota
phylum. On the zoosporic fungi, a new species of the Nowakowskiella J. Schröt genus
was described and Avachytrium Velez & Letcher reported for the first time in Brazil, as
well as Plectospira gemmifera Drechsler to the Oomycota. We observed a diverse
community with few dominant taxa, which occurred predominantly at rainy season, being
Cladochytrium replicatum Karling, Nowakowskiella hemisphaerospora Shanor,
Aphanomyces spp., Dictyuchus spp. and Pythium group P and T, the most frequent and
abundant, considering the reservoirs and seasons (dry and rainy). Zoosporic organisms
presented a wide distribution along different trophic gradients, with species strictly
related to locals with high concentrations of nitrogen and total phosphorus, and others
with low concentrations of these nutrients, which could potentially be taxa indicative of
quality of water. We conclude that heterotrophic zoosporic organisms are, in general,
tolerant to wide variations in nitrogen and phosphorus concentrations, but some taxa are
limited by the availability of these nutrients, particularly phosphorus.
Key words: Blastocladiomycota, Chytridiomycota, ecology, Oomycota

59
INTRODUÇÃO
Os organismos zoospóricos heterotróficos dos filos Blastocladiomycota,
Chytridiomycota e Oomycota possuem como similaridade a produção de esporos
flagelados (zoósporos), os quais os permitem se dispersarem ativamente em ecossistemas
aquáticos e terrestres úmidos. Nestes ambientes atuam como decompositores de matéria
orgânica ou como parasitas de diferentes grupos de hospedeiros, exercendo importante
papel na ciclagem do carbono nos ecossistemas (Shearer et al. 2007, Sime-Ngando et al.
2010, Wurzbacher et al. 2016).
Os ambientes aquáticos lênticos, em especial os reservatórios, possuem grande
importância devido aos diversos serviços que prestam à população, principalmente no
que se refere ao abastecimento público e geração de energia elétrica, sendo a
hidroeletricidade a principal fonte de energia do país e responsável por 75% do total de
energia produzida (Tolmasquim 2012). No entanto, sua construção gera importantes, e
muitas vezes, irreversíveis impactos, como a perda da biodiversidade e da qualidade da
água e mudanças socioeconômicas regionais (Tundisi et al. 2008). Dentre os diversos
impactos antrópicos gerados, a eutrofização e o uso desordenado do solo da bacia
hidrográfica, são as variáveis que frequentemente têm sido associadas com a perda da
qualidade da água e da biodiversidade (Pusceddu et al. 2007). O processo de eutrofização
tem sido bastante investigado por representar uma das principais causas de alterações na
qualidade da água e redução da diversidade em águas continentais (Margalef 1983). Este
processo geralmente associado ao aumento da concentração de nutrientes, especialmente
fósforo e nitrogênio, tem como consequência o aumento de suas produtividades (Esteves
1998). Já os tipos de usos e a ocupação do solo na bacia hidrográfica possuem relações
significativas no que diz respeito às diversas substâncias químicas lançadas nos corpos
d’água, como efluentes domésticos, industriais e agrícolas, expondo diversos organismos
a uma grande variedade de compostos mutagênicos e citotóxicos (Amaral et al. 2007).
Nos reservatórios essas alterações ecológicas podem ser detectadas a partir do
monitoramento da água e do sedimento acumulado, seja este superficial ou depositado
em camadas (Birks & Birks 2006). A análise do sedimento fornece dados cumulativos e
constitui um arquivo de informações de natureza biogeoquímica, uma vez que as camadas
de deposição temporais se encontram sequencialmente acumuladas (Mozeto 2004).
Assim, o sedimento superficial representa uma amostra espacial e temporalmente
integrada dos eventos que se acumularam no passado recente (Smol 2008). O oxigênio,

60
se presente, está restrito as camadas superficiais do perfil sedimentar, e por isso, os fungos
presentes neste compartimento geralmente apresentam adaptações fisiológicas
(mecanismo fermentativo) e/ou morfológicas (esporos de resistência) para suportar tais
limitações (Wurzbacher et al. 2010). As leveduras e os representantes do filo
Chytridiomycota são considerados os grupos de fungos mais ativos no perfil sedimentar
de reservatórios (Slapeta et al. 2005), e técnicas de PFLA (Análise automática de ácidos
graxos e fosfolípedos) (Goedkoop et al. 2005, Widenfalk et al. 2008) e clonagem
(Dawson & Pace 2002, Luo et al. 2005), indicam estes dois grupos como micota
dominante nos sedimentos de lagos e reservatórios, com tendência de maior diversidade
de espécies nos sedimentos localizados próximos as zonas litorâneas (Collins &
Willoughby 1962, Sparrow 1968). Além disso, assim como os fungos zoospóricos, alguns
poucos estudos têm apontado a importância dos oomicetos nos processos de transferência
de energia no sedimento, sendo alguns gêneros, tais como Aphanomyces spp. e Pythium
spp., considerados abundantes e ubíquos nas camadas superficiais do perfil sedimentar
(Sparrow 1968, Boopathi et al. 2015, Yang & Hong, 2015, Wurzbacher et al. 2016). No
entanto, considerando a totalidade dos estudos desenvolvidos em reservatórios, somente
pouco deles analisaram a diversidade destes grupos de organismos no sedimento
superficial, sendo no Brasil estes relatos inexistentes.
Embora as atuais técnicas de clonagem indiquem os fungos zoospóricos como
micota dominante de lagos e reservatórios, os fatores que afetam a variação temporal e a
estrutura da comunidade ainda permanecem pouco investigados (Wurzbacher et al.
2016). De maneira geral, quando consideramos os fungos zoospóricos e oomicetos, a
relação entre os padrões de ocorrência e distribuição em ambientes com diferentes trofias,
ainda é pouco explorada em escala global (Cooke 1976, Czeczuga et al. 1989, 1990a,b,
Czeczuga 1991a,b, Czeczuga & Woronowicz 1991, Lefranc et al. 2005, Chen et al. 2008,
Marano et al. 2011, Monchy et al. 2011), e principalmente no Brasil (Rocha 2004).
Muitos dos estudos conduzidos em ambientes com diferentes estados tróficos (Tan & Lim
1983, 1984, Czeczuga 1994, Rancovic 1998, Czeczuga & Muszynska 2001) listam
principalmente a diversidade em espécies, não relacionando os padrões de sua ocorrência
e distribuição com fatores abióticos. Czeczuga et al. (1990a,b) e Czeczuga (1991a,b)
apontam estes organismos como tolerantes e adaptados a diferentes níveis de poluição
orgânica, mas relata preferência dos mesmos por ambientes menos poluídos e ricos em
matéria orgânica. Lefranc et al. (2005), Chen et al. (2008), Lepère et al. (2008), Monchy
et al. (2011) e Wurzbacher et al. (2010, 2016) apontam, por meio de análises moleculares,

61
que a diversidade tende a ser menor em águas mais produtivas, sendo os reservatórios
oligotróficos e mesotróficos os que normalmente apresentam maior diversidade. Lefèvre
et al. (2012) mostraram maior diversidade de quitrídias em reservatórios meso-eutróficos
do que em ambientes oligotróficos, sugerindo que ambientes em estado trófico
intermediário (mesotróficos) sejam os mais propícios para ocorrência destes fungos. Para
o Brasil, Rocha (2004) sugeriu que a riqueza em espécies dos fungos zoospóricos e
oomicetos dos lagos do Parque Estadual das Fontes do Ipiranga (PEFI), com distintas
trofias (mesotrófico e eutrófico), apresentou correlação negativa com as concentrações de
nitrogênio e fósforo, tendo sido demonstrada e comprovada a natureza limáfoba ou
limaxena destes organismos. A autora pontua também que a ausência dos mesmos em
locais amostrados com altíssimas concentrações de nitrogênio e fósforo total indica a sua
possível utilização como bioindicadores de poluição, estando os mesmos completamente
ausentes em tal situação.
Com relação aos padrões sazonais de distribuição desses organismos zoospóricos
em ecossistemas lênticos, verifica-se uma grande escassez de estudos. Em regiões
temperadas se estabelece normalmente o período chuvoso como o de maior diversidade
(Czeczuga 1986, 1995, Czeczuga et al. 2010). Em regiões tropicais, os únicos estudos a
abordarem a sazonalidade em ecossistemas lênticos brasileiros foram os de Pires-
Zottarelli (1990) e Rocha (2004), os quais também apontaram ser a maior diversidade
encontrada durante a estação chuvosa.
Mesmo diante da expressiva frequência e importância destes organismos
zoospóricos nos processos de transferência de energia nos ecossistemas aquáticos,
estudos abordando sua diversidade e importância em reservatórios do Brasil ainda são
escassos, podendo ser resumidos aos trabalhos de Rocha & Merighi (1986) na Represa
Billings, Pires-Zottarelli (1990) na Represa do Lobo, Rocha & Pires-Zottarelli (2002) na
Represa do Guarapiranga e Rocha (2004) nos Lagos das Garças e das Ninfeias do Parque
Estadual das Fontes do Ipiranga (PEFI), no entanto, somente o último discute de forma
mais aprofundada a influência dos fatores abióticos sobre os padrões de ocorrência destes
organismos. Dos 121 táxons de Chytridiomycota e 18 de Blastocladiomycota conhecidos
no Brasil (Lista de espécies da Flora do Brasil 2014), aproximadamente 79% foram
relatados para ecossistemas lóticos (rios, riachos e cachoeiras) e solos, principalmente em
áreas de Mata Atlântica (Beneke & Rogers 1962, 1970, Rogers et al. 1970, Lyra &
Milanez 1974, Milanez et al. 1994, Pires-Zottarelli et al. 1996, Pires-Zottarelli 1999,
Pires-Zottarelli & Rocha 2007, Pires-Zottarellli & Gomes 2007, Nascimento & Pires-

62
Zottarelli 2009, 2010, entre outros), sendo a diversidade em reservatórios representada
por somente 21% das espécies conhecidas no país. Este mesmo cenário pode também ser
atribuído para o filo Oomycota, que possui somente 19,5% da diversidade relatada para
ecossistemas lênticos (Rocha & Merighi 1986; Pires-Zottarelli, 1990; Rocha & Pires-
Zottarelli, 2002; Rocha, 2004), com maior representatividade dos gêneros Achlya,
Aphanomyces, Pythium e Saprolegnia.
Com isso, o objetivo deste estudo foi investigar a influência dos fatores abióticos
sobre os padrões de ocorrência, frequência e abundância dos organismos zoospóricos
heterotróficos (Blastocladiomycota, Chytridiomycota e Oomycota) em três reservatórios
tropicais com distintos estados tróficos. Além disso, baseado nos escassos relatos destes
grupos nas camadas sedimentares, esperamos encontrar uma maior diversidade de
organismos zoospóricos heterotróficos na coluna d’água, comparado ao sedimento
superficial, assim como durante a estação chuvosa.
MATERIAL E MÉTODOS
Amostragem e mensuração dos fatores abióticos
Três reservatórios, localizados na bacia hidrográfica do Sorocaba/Médio Tietê
(UGRHI10), foram selecionados considerando-se seus distintos estados tróficos e
características morfométricas (Tabela 1).
Tabela 1. Características morfométricas dos três reservatórios localizados na bacia
hidrográfica do Sorocaba/Médio Tietê (UGRHI 10).
Reservatórios Ano
construção
Estado
trófico
Prof.
máx. (m)
Prof.
méd.(m)
Vol.
máx.
(m3)
Área
(km2)
Tempo
residência
da água
Área de
proteção Utilização
St. Helena 1938 (f)
Oligotrófico(a)
10,5(a)
- - - - Não inclusa Geração de energia elétrica
Hedberg 1811(c)
Eutrófico(d)
- 4.0(c)
- 0.113(c)
- “FLONA de
Ipanema” (c)
Paisagismo, pesquisa e
educação ambiental (c)
Itupararanga 1912(b)
Mesotrófico(d)
19(d)
7.8(b)
286
milhões(e)
936.51
(e)
4-13
meses(b)
APA de
Itupararanga(b)
Abastecimento público
(e)
a: Lucinda (2003); b: Secchin (2012); c: Plano de manejo - FLONA (2016); d: Projeto Acquased; e: Pedrazzi et al. (2014); f:
Eletromemoria-USP 2015; (-): informação indisponível.
Durante o estudo, quatro coletas foram realizadas, duas na estação seca
(agosto/2014 e julho/2015) e duas na chuvosa (dezembro/2014 e março/2015), para as
quais foram obtidos os dados mensais de precipitação e temperatura. Onze pontos de

63
coleta foram selecionados nas zonas pelágicas destes reservatórios, considerando-se a
entrada dos principais tributários, as estações de captação de água, bem como a posição
da barragem (Tabela 2).
Tabela 2. Coordenadas geográficas dos pontos de coleta nos três reservatórios da bacia
do Sorocaba/Médio Tietê.
Reservatórios Pontos Coordenadas
Sul Oeste
Santa Helena
1 23°34'49,92'' 47°25'32,64''
2 23°34'58,56'' 47°25'50,52''
3 23°34'54,30'' 47°26'12,12''
Hedberg
1 23°25'55,86'' 47°35'33,06'
2 23°25'40,86'' 47°35'31,14''
3 23°25'33,72'' 47°35'42,66''
Itupararanga
1 23°37'11,58'' 47°13'59,34''
2 23°36'51,96'' 47°18'50,22''
3 23°37'12,96'' 47°19'37,20''
4 23°38'50,36'' 47°22'14,52''
5 23°36'53,10'' 47°23'34,70''
Em cada ponto de coleta foram amostradas diferentes profundidades da coluna
d’água (superfície, meio e fundo), para as quais, foram mensurados os seguintes fatores
abióticos: temperatura, pH, condutividade (sonda Horiba U-51), oxigênio dissolvido pelo
método de Winkler modificado (Golterman & Clymo 1969), nitrogênio total e fósforo
total (Valderrama 1981), clorofila-a (Sartory & Grobelaar 1984) e profundidade de
desaparecimento do disco de Secchi. Além disso, amostras de sedimento superficial (2
cm superficiais), foram coletadas e o carbono orgânico total, nitrogênio total e fósforo
total estimados (Valderrama 1981, Andersen 1976).

64
Iscagem, isolamento e identificação morfológica e molecular dos espécimes
Em laboratório as amostras de diferentes profundidades da coluna d’água, bem
como as de sedimento superficial, foram iscadas com substratos celulósicos, queratinosos
e quitinosos (grãos de pólen de Pinus sp., epiderme de cebola, palha de milho, celofane,
sementes de Sorghum sp., ecdise de cobra e exoesqueleto de camarão), conforme
metodologia descrita em Milanez et al. (1989).
Estruturas assexuadas (zoosporângios, zoósporos e esporos de resistência) e
sexuadas (anterídios, oogônios e oósporos) foram analisadas, considerando-se o tipo de
desenvolvimento, formato, tamanho, ornamentação, e utilizadas para a identificação
morfológica dos espécimes.
Análises de Máxima Verossimilhança de diferentes regiões do DNA (18S, ITS,
28S e COX1), foram utilizadas para corroborar as identificações morfológicas ou então
identificar os isolados nos quais não foram observadas estruturas sexuadas,
imprescindíveis para a identificação em nível de espécie. Os táxons isolados em meio
cultura e incluídos em análises filogenéticas, para os quais não foi possível a identificação
em nível especifico, foram considerados como táxons distintos (especialmente as do
gênero Pythium) com base nas diferenças moleculares e morfológicas.
Análises estatísticas e índices de diversidade
A abundância (A) foi determinada com base no número de ocorrências,
considerando a presença ou ausência dos táxons em cada tipo de isca, nas diferentes
profundidades da coluna d’água (superfície, meio e fundo) e sedimento superficial
(Nascimento et al. 2011, adaptado). A frequência de ocorrência foi calculada em cada
estação (seca ou chuvosa) considerando-se: (número de amostras colonizadas por um
determinado táxon/número de amostras analisadas) × 100.
A estrutura da comunidade nas diferentes estações (seca e chuvosa) foi
caracterizada pela: (i) riqueza de espécies (S); (ii) Índice de diversidade de Shannon: (H’)
= -Σpi lnpi, onde pi é a abundância da espécie i na comunidade; (iii) Equitatividade (E) =
H’/Hmax’, em que Hmax’ é o valor máximo de diversidade para o número de táxons
presentes; (iv) Índice de dominância de Simpson: (D) = 1 - Σ (pi)2 baseado na abundância
dos táxons (Zak & Willig 2004); e (v) Índice de similaridade de Sorensen, utilizado para
comparar a composição dos táxons entre as estações (seca e chuvosa) e compartimentos
(coluna d’água e sedimento superficial), sendo SI = [2c/(a + b)] × 100, onde “c” é o
número de táxons comuns em ambas as estações ou compartimentos, “a” é o número de

65
táxons exclusivos da estação seca ou coluna d’água e “b” é o número de táxons exclusivos
da estação chuvosa ou sedimento superficial (Müeller-Dumbois & Ellemberd 1974).
A análise de componentes principais (PCA) foi utilizada para ordenar os dados
abióticos a partir de uma matriz de correlação, onde os dados foram transformados para
log (x +1), exceto pelo pH. As PCoAs dos dados bióticos da coluna d’água e sedimento
superficial foram calculadas com base na distância euclidiana com base na riqueza dos
táxons, para os quais foram desconsiderados os táxons com apenas uma ocorrência ao
longo do estudo. Ambas as análises, assim como a transformação, foram realizadas no
programa PC-ORD 6.0 (McCune & Mefford 2011).
O teste t de "student” foi aplicado para verificar a existência de diferenças
significativas dentre os fatores abióticos mensurados nas diferentes estações e
reservatórios, bem como para os índices de diversidade, para o qual os dados foram
normalizados quando os mesmos não apresentavam uma distribuição normal. Foram
considerados resultados estatisticamente significativos valores de p≤0,05.
Índice de estado trófico (IET)
O cálculo do índice de estado trófico (IET) dos reservatórios foi baseado nas
concentrações de fósforo total e clorofila-a (Lamparelli 2004). Os valores da
profundidade do disco de Secchi não foram considerados para o cálculo do IET, já que
este pode não refletir a biomassa algal, mas o material inorgânico dissolvido na coluna
d’água (Cetesb 2013).
RESULTADOS & DISCUSSÃO
Dados climatológicos
O período de estudo compreendeu os anos de 2014 e 2015, os quais foram
atipicamente de baixa pluviosidade. De acordo com os dados do Cepagri (2018), as
médias mensais de precipitação durante os meses amostrados, para o município de
Votorantim (onde está inserido o reservatório de Santa Helena - Fig. 1A), foram muito
abaixo das médias históricas mensais para março (133,7 mm), julho (35,6 mm), agosto
(39,1 mm) e dezembro (191,7 mm), o mesmo sendo observado para os de Itupararanga
(141,2 mm em março, 44,7 mm em julho, 40 mm em agosto e 175,6 mm em dezembro),
localizado em Ibiúna, e Hedberg (141,0 mm em março, 56,1 mm em julho, 36 mm em

66
agosto e 190,3 mm em dezembro) em Iperó (Fig. 1B-C). Já as médias de temperatura
estão de acordo com as médias históricas observadas para os meses amostrados.
Figura 1. Médias mensais de precipitação (mm) e temperatura (°C) nos reservatórios de Santa Helena (A), Itupararanga (B) e
Hedberg (C) durante as amostragens.
Fatores abióticos da coluna d’água
Na análise de componentes principais, os dois primeiros eixos explicaram 63,4%
da variabilidade total dos dados (Fig. 2). O eixo 1, explicou a maior parte da variabilidade
dos dados (38,8%), sendo o fósforo total (PT) (r = - 0,47) e o nitrogênio total (NT) (r = -
0,45) as variáveis que mais influenciaram a variabilidade neste eixo. Independentemente
da estação, maior parte das unidades amostrais de Santa Helena e Itupararanga se
agruparam no lado positivo do eixo 1, relacionadas às mais baixas concentrações de PT
e NT, enquanto as de Hedberg se agruparam no lado negativo deste eixo. O eixo 2
explicou 24,6% da variabilidade dos dados, sendo a temperatura (r=- 0,60) no lado
negativo e o oxigênio dissolvido (r = 0,53) no lado positivo, as variáveis que mais

67
influenciaram a variabilidade dos dados. Todas as unidades amostrais da estação seca,
exceto pelo ponto 1 do reservatório Hedberg (Hsec1), se agruparam no lado positivo deste
eixo, enquanto as da estação chuvosa, se agruparam no lado negativo deste eixo. Portanto,
o eixo 1 representa um gradiente trófico, enquanto o eixo 2 um gradiente sazonal.
Figura 2. Ordenação das variáveis abióticas na análise de componentes principais (PCA) mensurados na
coluna d’água. O eixo 1 representa um gradiente trófico enquanto o eixo 2 um gradiente sazonal. Legendas:
Cloro: clorofila-a; Cond: Condutividade; NT: Nitrogênio total; OD: Oxigênio dissolvido; PT: fósforo
total; Secchi: transparência do disco de Secchi; Temp: temperatura. Stsec1: Santa Helena seca ponto 1;
Stsec2: Santa Helena seca ponto 2; Stsec3: Santa Helena seca ponto 3; Stchu1: Santa Helena chuvosa ponto
1; Stchu2: Santa Helena chuvosa ponto 2; Stchu3: Santa Helena chuvosa ponto 3; Hsec1: Hedberg seca
ponto 1; Hsec2: Hedberg seca ponto 2; Hsec3: Hedberg seca ponto 3; Hchu1: Hedberg chuvosa ponto 1;
Hchu2: Hedberg chuvosa ponto 2; Hchu3: Hedberg chuvosa ponto 3; Itsec1: Itupararanga seca ponto 1;
Itsec2: Itupararanga seca ponto 2; Itsec3: Itupararanga seca ponto 3; Itsec4: Itupararanga seca ponto 4;
Itsec5: Itupararanga seca ponto 5; Itchu1: Itupararanga chuvosa ponto 1; Itchu2: Itupararanga chuvosa
ponto 2; Itchu3: Itupararanga chuvosa ponto 3; Itchu4: Itupararanga chuvosa ponto 4; Itchu5: Itupararanga
chuvosa ponto 5.

68
Durante as amostragens foram observadas mudanças pontuais no estado trófico
dos reservatórios, quando comparados aos dados de literatura (Tab. 1). As concentrações
de fósforo total e a clorofila-a apresentaram uma ampla variação nas diferentes estações
e reservatórios, no entanto, estes últimos foram predominantemente mesotróficos durante
o período de estudo, exceto por Hedberg na estação seca (ago/14), o qual foi eutrófico e
Santa Helena no mesmo período (ago/14), onde foi observada condição oligotrófia (Fig.
3).
Figura 3. Concentração de clorofila-a e fósforo total e atribuição do índice de estado trófico nas estações
seca e chuvosa. E: Eutrófico; M: Mesotrófico; O: Oligotrófico. Classificação de trofia baseado em
Lamparelli (2004).
A variação dos fatores abióticos mensurados na coluna d’água está representada
na tabela 3 e figura 4 (A-H). De acordo com o teste t (p≤0,05), foram observadas
diferenças significativas entre estações ou reservatórios nas concentrações de fósforo total
(PT), nitrogênio total (NT), transparência da água (disco de Secchi), condutividade,
temperatura e oxigênio dissolvido.

69
Tabela 3. Valores máximos e mínimos (média ± desvio padrão) das variáveis abióticas mensuradas durante as estações
seca e chuvosa nos três reservatórios estudados.
Variáveis
St. Helena Hedberg Itupararanga
Seca Chuvosa Seca Chuvosa Seca Chuvosa
min-máx min-máx min-máx min-máx min-máx min-máx
(méd. ± DP) (méd. ± DP) (méd. ± DP) (méd. ± DP) (méd. ± DP) (méd. ± DP)
Temperatura (°C) 16,3-21,0 20,8-24,0 13,1-20,2 20,7-21,9 15,2-19,4 21,2-26,8
(18,0 ± 1,2) (22,9 ± 0,9) (17,1 ± 2,1) (22,6 ± 2,4) (17,0 ± 0,9) (24,1 ± 1,7)
Condutividade (µS cm-1) 67,0-106,0 97,0-119,0 111-175 26,0-244,0 73,0-126,0 33,0-200,0
(89,0 ± 12,4) (107,9 ± 7,9) (139,6 ± 22,6) (114,1 ± 51,4) (96,3 ± 14,2) (94,7 ± 29,7)
NT (µg L-1) 330,2-632,7 384,9-1035,0 1157,0-1764,6 212,9-1154,9 666,3-1107,4 307,2-1207,8
(523,0 ± 89,9) (660,3 ± 156,4) (1346,7 ± 182,0) (831,2 ± 254,5) (890,1 ± 110,4) (543,7 ± 202,0)
PT (µg L-1) 13,3-20,4 15,4-25,9 49,2-123,8 30,3-95,1 19,5-64,9 11,4-69,9
(17,0 ± 2,4) (19,8 ± 3,6) (87,9 ± 17,6) (67,2 ± 21,3) (38,2 ±12,5) (24,0 ± 15,8)
pH 4,9-7,4 5,7-8,3 6,2-9,4 5,4-7,2 4,4-7,6 6,2-8,7
(7,0 ± 0,7) (7,2 ± 0,7) (7,6 ±1,1) (6,4 ± 0,6) (6,1 ± 0,9) (7,1 ± 0,7)
OD (mg L-1) 5,5-8,2 1,8-9,1 2,9-10,0 0,9-9,9 4,0-9,1 0,5-9,2
(7,0 ± 0,7) (6,0 ± 0,8) (6,5 ± 1,8) (4,4 ± 2,5) (6,4 ±1,7) (4,3 ± 2,7)
Secchi (m) 1,2-1,9 1,1-1,6 0,6-1,0 0,2-0,8 0,8-1,7 0,7-1,4
(1,5 ± 0,3) (1,2 ± 0,2) (0,8 ± 0,2) (0,5 ± 0,2) (1,1 ± 0,3) (1,0 ± 0,2)
Clorofila-a (µg L-1) 0,9-18,6 3,8-17,9 1,4-153,8 1,1-15,4 0,5-62,0 1,3-69,0
(10,0 ± 7,0) (11,7 ± 4,5) (20,2 ± 36,8) (5,5 ± 4,0) (18,7 ± 15,8) (14,8 ±13,2)

70
Figura 4. Box-Plot das variáveis abióticas, clorofila-a e profundidade do disco de Secchi nos três
reservatórios durante as estações seca e chuvosa. Os círculos vazios (F e H) representam “outliers”.

71
Considerando as diferentes estações (seca e chuvosa), no reservatório de Santa
Helena, foram observadas as menores concentrações de fósforo total (PT), nitrogênio
total (NT) e clorofila-a, as quais estão próximas aos limites estabelecidos pela resolução
CONAMA (357/2005) que dispõe sobre a classificação dos corpos de água e padrões de
lançamento de efluentes e define valor máximo de 20 µg.L-1 para o fósforo total, 10 µg.L-
1 para a clorofila-a e valores entre 500 µg.L-1 a 3700 µg.L-1 para o nitrogênio total. No
entanto, em Itupararanga, durante as duas estações, foram observadas concentrações de
PT e clorofila-a acima dos valores estabelecidos pela resolução acima citada, o mesmo
sendo observado para o PT no Hedberg, o qual apresentou as maiores concentrações deste
nutriente dentre os reservatórios analisados (Fig. 4 e Tab. 3).
Os efeitos da variação da temperatura entre as diferentes camadas da coluna
d’água possui uma enorme importância na dinâmica dos ecossistemas lênticos, devido ao
fenômeno da estratificação térmica. Em reservatórios tropicais, diferenças de 1°C entre o
epilímnio e o hipolímnio, são suficientes para promover a estratificação térmica da coluna
d’água (Esteves 2011, Tundisi et al. 2008), alterando a densidade da água e promovendo
a formação de barreiras físicas para diversos grupos de organismos, especialmente os
microscópicos. No geral, os três reservatórios apresentaram perfil estratificado durante as
amostragens (Fig. 5), no entanto, durante a estação seca (jul/15), os três reservatórios
apresentaram perfil de mistura, o mesmo sendo observado para Santa Helena durante a
estação chuvosa de 2015 (mar/15). A mistura da coluna d’água refletiu diretamente na
composição de espécies nos diferentes compartimentos (coluna d’água e sedimento
superficial), os quais apresentaram uma alta similaridade (76% - 81%) se comparado aos
de perfil estratificado (54% - 62%).

72
Figura 5. Variação do perfil térmico na coluna d’água nos três reservatórios durante as estações seca e chuvosa. O eixo x representa
a temperatura (°C) e o eixo y a profundidade (m).

73
Sedimento superficial
No sedimento superficial foram mensurados o carbono orgânico total (COT%),
nitrogênio total (NT%) e fósforo total (PT%), além do oxigênio dissolvido do fundo da
coluna d’água. De acordo com Smol (2008), os dados do sedimento superficial (2 cm
superficiais) representam geralmente de um a dois de informação acumulada. Por isso, os
dados de COT (%), NT(%) e PT(%) foram extrapolados para as demais coletas, já que
estes parâmetros foram estimados somente uma vez durante todo o estudo. As três
variáveis apresentaram diferenças significativas entre os reservatórios (teste t, p≤0,05),
sendo os maiores valores de COT(%) e NT (%) observadas em Itupararanga e de PT (%)
em Hedberg. Já os menores valores de COT (%), NT (%) e PT (%) foram observados em
Santa Helena (Fig. 6A-C).
Figura 6. Box-Plot mostrando a variação do carbono orgânico total, fósforo total e nitrogênio total no
sedimento superficial de cada reservatório estudado.

74
No geral, os reservatórios apresentaram uma ampla variação no oxigênio
dissolvido no fundo da coluna d’água (Fig. 7), durante as estações seca e chuvosa, para
as quais foram observadas diferenças significativas de acordo com o teste t (p≤0,05). As
menores concentrações foram observadas em Hedberg e Itupararanga, durante a estação
chuvosa, enquanto a maior foi em Santa Helena durante a mesma estação. No fundo da
coluna d’água de todos os pontos de coleta em que o oxigênio apresentou concentrações
menores que 1 mg/L, não foram observadas ocorrências de fungos zoospóricos ou
oomicetos, independentemente do reservatório ou estação. Por outro lado, Santa Helena
durante a estação chuvosa apresentou as maiores concentrações de OD no fundo da coluna
d’água, onde foi observada uma alta abundância de organismos zoospóricos
heterotróficos. Estes dados indicam uma limitação destes organismos a baixas
concentrações de oxigênio dissolvido, assim como mencionado por Sparrow (1968),
Marano et al. 2011 e Wurbacher et al. (2010, 2016).
Figura 7. Variação na concentração de oxigênio dissolvido no fundo da coluna d’água, nos três reservatórios
durante a estação seca e chuvosa.

75
Análises filogenéticas
No total, 102 sequências das regiões 18S, ITS, 28S, RPB1, RPB2 e COXI foram
obtidas durante o estudo (Tabela 1, apêndice I), considerando os dois grupos de
organismos, as quais representam 25 táxons. Por meio das análises de máxima
verossimilhança, 33 espécimes foram identificados ou tiveram as identificações
morfológicas corroboradas, dos quais nove são de fungos zoospóricos e 24 de oomicetos.
Com relação aos últimos, Plectospira gemmifera Drechsler foi inserida pela primeira vez
em análises filogenéticas (Jerônimo et al. 2017 - Anexo I - Phytotaxa 307: 191-198). Já
dentre os fungos zoospóricos, N. ramosa Butler e a nova espécie de Nowakowskiella
(Nowakowskiella crenulata – Capítulo 2) foram inseridas pela primeira vez em
reconstruções filogenéticas, bem como Rhizophydium angulosum Karling, para o qual
está sendo proposta uma nova combinação (Capítulo 3). As demais filogenias (utilizadas
para confirmar ou identificar os isolados), assim como as imagens dos táxons
identificados em nível de espécie, foram inseridas no material complementar I.
Diversidade
A maioria dos estudos desenvolvidos em ecossistemas lênticos utilizaram técnicas
avançadas de sequenciamento de última geração, detectando assim grande parte da
riqueza dos táxons, no entanto, com uma baixa resolução taxonômica (geralmente em
nível de ordem), nos quais, os resultados estão diretamente relacionados à confiabilidade
das sequências disponíveis nos bancos de dados gênicos. Durante este estudo utilizamos
principalmente as características morfológicas para identificar os espécimes e as
ferramentas moleculares foram empregadas, no geral, para corroborar as identificações
morfológicas. Com isso, os dados obtidos apresentam uma maior especificidade
taxonômica e uma discussão mais aprofundada (em nível de espécie ou gênero).
Durante o estudo, 52 táxons de organismos zoospóricos heterotróficos (fungos
zoospóricos e oomicetos) foram identificados, sendo 25 deles representantes dos filos
Blastocladiomycota e Chytridiomycota (fungos zoospóricos) e 27 do filo Oomycota (Tab.
4). Dentre os fungos zoospóricos, 64 espécimes foram observados com 12 táxons
identificados em nível específico, dentro os quais, uma nova espécie para o gênero
Nowakowskiella J. Schröt foi descrita (Nowakowskiella crenulata Jerônimo & Pires-
Zottarelli – ver capitulo 2) e o gênero Avachytrium Velez & Letcher relatado pela primeira
vez para o Brasil. Os gêneros Nowakowskiella J. Schröt, Rhizophydium Schenk,

76
Diplophlyctis J. Schröt e Cladochytrium Nowak foram, respectivamente, os mais
frequentes e abundantes dentre os fungos zoospóricos (Tab. 4).
As análises de Máxima Verossimilhança (Anexo I, capítulos 2 e 3) foram
importantes para corroborar e/ou identificar nove isolados, dentre eles, Avachytrium sp.,
Asterophlyctis sarcoptoides, Nowakowskiella ramosa, Nowakowskiella
hemisphaerospora, Nowakowskiella crenulata, Nowakowskiella sp.1, Rhizophydium
angulosum, Rhizophydium elyense e Siphonaria sp.
Dentre os representantes do filo Oomycota, 84 espécimes foram observados e 13
táxons identificados em nível específico, sendo a espécie Plectospira gemmifera
Drechsler, isolada em amostras de sedimento superficial de Itupararanga, relatada pela
primeira vez para o Brasil (Anexo II - Jerônimo et al. 2017). Pythium Pringsheim,
Dictyuchus Leitg. e Aphanomyces de Bary foram os gêneros mais frequentes e abundantes
durante o estudo.
As análises de Máxima Verossimilhança foram importantes para corroborar e/ou
identificar 24 isolados, dentre eles, Achlya caroliniana, Achlya klebsiana, Achlya
prolifera, Aphanomyces laevis, Dictyuchus monosporus, Dictyuchus pseudodictyon,
Plectospira gemmifera, Pythium inflatum, Pythium midletonii, Pythium torulosum,
Pythium sp.1, Pythium sp.2, Pythium sp.3, Pythium sp.4, Pythium sp.5, Pythium sp.6,
Pythium sp.7, Phytopythium vexans, Saprolegnia aenigmatica e Saprolegnia subterranea
(Anexo III).
Os isolados pertencentes ao gênero Pythium, não identificados em nível de
espécie, foram divididos em grupos com base no tipo de zoosporângio segundo Plaats-
Niterink (1981), sendo o grupo dos filamentosos não inflados denominados de grupo P,
os filamentosos inflados de grupo F e os esféricos de grupo T.

77
Tabela 4. Ocorrência, frequência e abundância dos organismos zoospóricos heterotróficos nos três reservatórios da Bacia Hidrográfica do Sorocaba/Médio Tietê, durante as
estações seca e chuvosa. Legendas: FT: Frequência total; AT: Abundância total. Stsec: Santa Helena estação seca; Stchu: Santa Helena estação chuvosa; Hsec: Hedberg estação
seca; Hchu: Hedberg estação chuvosa; Itsec: Itupararanga estação seca; Itchu: Itupararanga estação chuvosa.
Fungos zoospóricos Stsec Stchu Frequência (%) Abundância Hsec Hchu Frequência (%) Abundância Itsec Itchu Frequência (%) Abundância FT (%) AT
Avachytrium sp.1 0 0 0 0 1 0 50 0,004 0 0 0 0 16,6 0,004
Catenochytrium sp.1 0 1 50 0,004 1 0 50 0,004 0 0 0 0 33,3 0,008
Catenomyces sp.1 0 0 0 0 1 0 50 0,004 0 0 0 0 16,6 0,004
Catenophlyctis variabilis (Karling)
Karling 0 0 0 0 0 5 50 0,021 0 0 0 0 16,6 0,021
Chytriomyces appendiculatus Karling 1 0 50 0,004 0 0 0 0 0 1 50 0,003 33,3 0,007
Cladochytrium replicatum Karling 1 0 50 0,004 1 2 100 0,013 1 0 50 0,003 66,6 0,019
Cladochytrium setigerum Karling 0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Cylindrochytridium sp.1 0 2 50 0,008 0 0 0 0 0 0 0 0 16,6 0,008
Diplophlyctis intestina (Schenk) J. Schröt
0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Diplophlyctis sarcoptoides (H.E.
Petersen) Dogma 3 0 50 0,013 0 1 50 0,004 1 0 50 0,003 50 0,019
Endochytrium sp. 1 0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Entophlyctis sp. 1 0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Karlingiomyces spp. 0 0 0 0 1 0 50 0,004 1 1 100 0,005 50 0,009
Nowakowskiella crenulata Jerônimo
& Pires-Zottarelli 0 2 50 0,008 1 2 100 0,013 0 0 0 0 50 0,021
Nowakowskiella hemisphaerospora Shanor
2 2 100 0,017 1 1 100 0,008 0 1 50 0,003 83,3 0,028
Nowakowskiella ramosa E.J. Butler 1 2 100 0,013 0 0 0 0 0 1 50 0,003 50 0,015
Nowakowskiella sp. 1 0 2 50 0,008 0 0 0 0 0 0 0 0 16,6 0,008
Nowakowskiella spp. 0 1 50 0,004 0 1 50 0,004 2 1 100 0,008 66,6 0,016
Phlyctochytrium sp. 1 0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Rhizophydium angulosum Karling 0 0 0 0 0 0 0 0 1 0 50 0,003 16,6 0,003
Rhizophydium elyense Sparrow 0 0 0 0 0 0 0 0 1 1 100 0,005 33,3 0,005
Rhizophydium sp. 1 0 0 0 0 0 0 0 0 0 1 50 0,003 16,6 0,003

78
Fungos zoospóricos Stsec Stchu Frequência (%) Abundância Hsec Hchu Frequência (%) Abundância Itsec Itchu Frequência (%) Abundância FT (%) AT
Rhizophydium spp. 0 0 0 0 0 0 0 0 0 1 50 0,003 16,6 0,003
Rhizophydium stipitatum Sparrow 0 0 0 0 1 0 50 0,004 0 0 0 0 16,6 0,004
Siphonaria sp.1 0 0 0 0 1 0 50 0,004 0 0 0 0 16,6 0,004
Riqueza de espécies 5 7 9 6 11 8
Oomycota Stsec Stchu Frequência (%) Abundância Hsec Hchu Frequência (%) Abundância Itsec Itchu Frequência (%) Abundância FT (%) AT
Achlya caroliniana Coker 1 0 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Achlya klebsiana Pieters 0 1 50 0,003 0 1 50 0,003 0 0 0 0 33,3 0,007
Achlya prolifera Nees 0 1 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Achlya spp. 3 0 50 0,01 3 1 100 0,014 1 0 50 0,002 66,6 0,026
Aphanomyces laevis de Bary 0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Aphanomyces spp. 3 1 100 0,014 2 7 100 0,031 0 2 50 0,004 83,3 0,049
Dictyuchus monosporus Leitg. 0 1 50 0,003 0 0 0 0 0 2 50 0,004 33,3 0,008
Dictyuchus pseudodictyon Coker &
Braxton 0 0 0 0 0 0 0 0 0 1 50 0,002 16,6 0,002
Dictyuchus spp. 2 3 100 0,017 4 2 100 0,021 1 1 100 0,004 100 0,042
Pythiogeton sp. 1 0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Phytopythium vexans Abad, de Cock,
Bala, Robideau, Lodhi & Levesque 0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Plectospira gemmifera Drechsler 0 0 0 0 0 0 0 0 1 0 50 0,002 16,6 0,002
Pythium inflatum V.D. Matthews 0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Pythium middletonii Sparrow 1 0 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Pythium torulosum Coker & P. Patt. 0 0 0 0 0 0 0 0 0 1 50 0,002 16,6 0,002
Pythium sp. 1 1 0 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Pythium sp. 2 1 0 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Pythium sp. 3 0 0 0 0 1 0 50 0,003 0 0 0 0 16,6 0,003
Pythium sp. 4 0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Pythium sp. 5 0 1 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Pythium sp. 6 0 1 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003

79
Fungos zoospóricos Stsec Stchu Frequência (%) Abundância Hsec Hchu Frequência (%) Abundância Itsec Itchu Frequência (%) Abundância FT (%) AT
Pythium sp. 7 1 0 50 0,003 0 0 0 0 0 0 0 0 16,6 0,003
Pythium spp. F 2 0 50 0,007 2 5 100 0,024 0 0 0 0 50 0,031
Pythium spp. P 0 4 50 0,014 2 3 100 0,017 0 3 50 0,006 66,6 0,038
Pythium spp. T 3 4 100 0,024 0 1 50 0,003 1 0 50 0,002 66,6 0,03
Saprolegnia aenigmatica Sand.-Sierra & Diég. Urib.
0 0 0 0 0 1 50 0,003 0 0 0 0 16,6 0,003
Saprolegnia subterranea Dissmann 0 0 0 0 0 0 0 0 1 0 50 0,002 16,6 0,002
Riqueza de espécies 10 9 6 13 5 6

80
Dentre os fungos zoospóricos, Diplophlyctis sarcoptoides (H.E. Petersen)
Dogma, Nowakowskiella hemisphaerospora Shanor e Cladochytrium replicatum Karling
foram comuns aos três reservatórios, durante o período de estudo. Por outro lado,
Cylindrochytridium sp.1 e Nowakowskiella sp.1 ocorrem exclusivamente em Santa
Helena, Avachytrium sp.1, Catenomyces sp., Catenophlyctis variabilis (Karling) Karling
e Cladochytrium setigerum Karling em Hedberg, enquanto Diplophlyctis intestina
(Schenk) J. Schröt, Endochytrium sp.1, Entophlyctis sp.1, Phlyctochytrium sp.1,
Rhizophydium angulosum Karling e R. elyense Sparrow o foram em Itupararanga
(Fig.8A). Já com relação aos representantes do filo Oomycota, Achlya spp. Aphanomyces
spp., Dictyuchus spp., Pythium spp. grupo T (zoosporângio esférico) e Pythium spp. grupo
P (zoosporângio filamentoso não inflado) ocorrem nos três reservatórios durante o
período de estudo. No entanto, Achlya caroliniana Coker, Achlya prolifera Ness,
Pythium middletonii Sparrow, Pythium sp.1, Pythium sp.2, Pythium sp.5, Pythium sp.6 e
Pythium sp.7, ocorreram exclusivamente em Santa Helena, Achlya klebsiana Pieters,
Aphanomyces laevis de Bary, Pythium sp.4, Phytopythium vexans Abad, de Cock, Bala,
Robideau, Lodhi & Levesque, Saprolegnia aenigmatica Sandoval-Sierra & Diéguez-
Uribeondo, Pythiogeton sp.1 e Pythium inflatum V.D. Matthews em Hedberg, enquanto
Dictyuchus monosporus Leitg., D. pseudodictyon Coker & Braxton, Pythium torulosum
Coker & P. Patt., Saprolegnia subterranea (Dissmann) Seym. e Plectospira gemmifera
Drechsler o foram em Itupararanga (Fig. 8B).

81
Figura 8. Número de ocorrência de fungos zoospóricos (A) e oomicetos (B) nos reservatórios analisados durante o estudo. Legendas (A): Siphsp1: Siphonaria sp.1; Rhizspp: Rhizophydium
spp.; Rhizsp1: Rhizophydium sp.1; Rhizstip: Rhizophydium stiptatum; Rhizely: Rhizophydium elyense; Rhizang: Rhizophydium angulosum; Phlycsp1: Phlyctochytrium sp.1; Nowspp:
Nowakowskiella spp.; Nowsp1: Nowakowskiella sp.1; Nowram: Nowakowskiella ramosa; Nowhem: Nowakowskiella hemisphaerospora; Nowcre: Nowakowskiella crenulata; continua ...

82
Karlspp: Karlingiomyces spp; Entosp1: Entophlyctis sp.1; Endosp1: Endochytrium sp.1. Diplsar: Diplophlyctis sarcoptoides; Diplinst: Diplophlyctis intestina; Cylin: Cylindrochytrium sp.1;
Cladset: Cladohytrium setigerum; Clarepl: Cladochytrium replicatum; Chyapp: Chytriomyces appendiculatus; Catvar: Catenophlyctis variabilis; Catenom: Catenomyces sp.1; Caten:
Catenochytridium sp.1; Avasp: Avachytrium sp.1. Legendas (B): Sapsub: Saprolegnia subterranea; Sapaen: Saprolegnia aenigmatica; Pytor: Pythium torulosum; Pyspp.T: Pythium group T;
Pyspp.P: Pythium group P; Pyspp.F: Pythium group F; Pysp7: Pythium sp.7; Pysp6: Pythium sp.6; Pysp5: Pythium sp.5; Pysp4: Pythium sp.4; Pysp2: Pythium sp.2; Pysp1: Pythium sp.1;
Pymidd: Pythium middletonii; Pyinf: Pythium inflatum; Plecgem: Plectospira gemmifera; Phytovex: Phytopythium vexans; Pytiosp1: Pythiogeton sp.1; Dicspp: Dictyuchus spp; Dicpseu:
Dictyuchus pseudodyction; Dicmon: Dictyuchus monosporus; Aphspp: Aphanomyces spp. Aphlae: Aphanomyces laevis; Achspp: Achlya spp; Achpro: Achlya prolifera; Achkle: Achlya
klebsiana; Achcar: Achlya caroliniana

83
De forma geral foi observada uma maior riqueza de espécies na estação
chuvosa, embora não tenham sido observadas diferenças significativas, de acordo com
teste t (p≤0,05), o que pode ser reflexo da baixa pluviosidade durante o período
amostrado, já que Pires-Zottarelli (1990) e Rocha (2004) encontraram maior diversidade
durante a estação chuvosa em lagos e reservatórios do estado de São Paulo. Ainda assim,
os reservatórios de Santa Helena e Hedberg apresentaram uma maior riqueza de espécies
de organismos zoospóricos heterotróficos durante a estação chuvosa, enquanto
Itupararanga o foi na estação seca. Já dentre os oomicetos, a maior riqueza foi observada
durante a estação chuvosa em Hedberg, enquanto os fungos zoospóricos, o foram durante
a estação seca em Itupararanga (Fig. 9).
Figura 9. Riqueza de espécies de fungos zoospóricos e oomicetos nos três reservatórios, durante as
estações seca e chuvosa, no período de estudo.
Considerando os diferentes compartimentos (coluna d'água e sedimento), os
oomicetos apresentaram maior riqueza de espécies na coluna de água (Fig. 10A),
enquanto os fungos zoospóricos o foram no sedimento superficial (Fig. 10B). No entanto,
na coluna d'água de Itupararanga, os fungos zoospóricos apresentaram maior riqueza de
espécies na estação chuvosa, enquanto nas amostras de sedimento de Hedberg, os
oomicetos apresentaram maior riqueza de espécies durante a estação seca. Além disso, os
oomicetos foram mais freqüentes e abundantes na coluna d'água, enquanto os fungos
zoospóricos o foram no sedimento superficial.

84
Figura 10. Riqueza de espécies de fungos zoospóricos e oomicetos na coluna d’água (A) e sedimento superficial (B) nos três
reservatórios, durante as estações seca e chuvosa, no período de estudo.
Durante o estudo, foi observada uma comunidade diversa com poucos táxons
dominantes (Tab. 5), sendo Cladochytrium replicatum, Nowakowskiella
hemisphaerospora, Aphanomyces spp., Dictyuchus spp., Pythium grupo P e Pythium
grupo T, os táxons mais frequentes e abundantes, considerando os reservatórios e estações
(seca e chuvosa). Wurzbacher et al. (2016) observaram a predominância de
representantes da ordem Cladochytriales (Nowakowskiella e Cladochytrium) em
ecossistemas lênticos de regiões temperadas, enquanto no Brasil, Rocha (2004)
mencionou a expressiva frequência dos representantes dos gêneros Nowakowskiella,
Dictyuchus e Pythium spp. do grupo T, em lagos do Parque Estadual das Fontes do
Ipiranga (PEFI), São Paulo, SP.
Tabela 5. Índices de diversidade dos organismos zoospóricos heterotróficos nos
reservatórios durante as estações seca e chuvosa. Letras iguais indicam que os valores
não apresentaram diferenças significativas entre as estações.
Índices Hedberg St. Helena Itupararanga
Seca Chuvosa Seca Chuvosa Seca Chuvosa
Riqueza de espécies (S) 15a 19a 15a 16a 16a 14a
Dominância de Simpson (D) 0,91a 0,90a 0,92a 0,92a 0,93a 0,92a
Índice de Shannon (H’) 2,60 a 2,60a 2,60a 2,60a 2,70a 2,60a
Equitatividade (E) 0,89a 0,75a 0,89a 0,87a 0,97a 0,94a

85
Uma similar composição de espécies foi observada entre as estações (SI = 88%),
nas quais, Chytriomyces appendiculatus, Cladochytrium replicatum, Diplophlyctis
sarcoptoides, Nowakowskiella crenulata, N. hemisphaerospora, N. ramosa, R. elyense,
Pythium spp. grupo P, Pythium spp. grupo T e Pythium spp. grupo F foram observados
tanto na estação seca como na chuvosa. Por outro lado, Catenophlyctis variabilis,
Cylindrochytridium sp.1, Nowakowskiella sp.1, Achlya klebsiana, A. prolifera,
Aphanomyces laevis, Dictyuchus monosporus, D. pseudodyction, Phytopythium vexans,
Pythium inflatum, P. torulosum e Saprolegnia aenigmatica foram restritos à estação
chuvosa, a qual apresentou maiores concentrações de nitrogênio total e fósforo total
(exceto por Hedberg), enquanto Cladochytrium setigerum, Diplophlyctis intestina,
Rhizophydium angulosum, R. stiptatum, Siphonaria sp.1, Achlya caroliniana, Plectospira
gemmifera, Pythium middletonii e Saprolegnia subterranea, o foram na estação seca,
associada à menores concentrações destes nutrientes (exceto por Hedberg). A alta
similaridade entre as estações pode ser reflexo da baixa pluviosidade observada durante
o período de estudo, especialmente na estação chuvosa, a qual apresentou volumes muito
abaixo das médias históricas para os meses amostrados, já que diversos estudos
(Czeczuga 1986, Pires-Zottarelli 1990, Czeczuga 1995, Rocha 2004, Marano et al. 2008,
Czeczuga et al. 2010, Marano et al. 2011) apontaram maior diversidade nos períodos de
maior pluviosidade.
Foi observada uma comunidade diversa, com poucos táxons dominantes entre os
compartimentos, os quais ocorreram de maneira geral na coluna d’água e sedimento
superficial (SI = 81%). No entanto, R. stipitatum, A. caroliniana, A. klebsiana, A.
prolifera, A. laevis, D. monosporus, P. vexans, P. middletonii, Pythium spp. grupo F, S.
aenigmatica ocorreram exclusivamente na coluna d’água, enquanto Chytriomyces
appendiculatus, C. setigerum, D. intestina, N. hemisphaerospora, R. angulosum, R.
elyense, Siphonaria sp.1, D. pseudodictyon, P. gemmifera, P. inflatum, P. torulosum e S.
subterranea o foram no sedimento superficial. Sparrow (1968), Czeczuga & Muszynska
(2001), Amal et al. (2005) e Zappia et al. (2014) também relataram a expressiva
frequência dos gêneros Achlya, Aphanomyces, Dictyuchus e Pythium em estudos de
distribuição no perfil da coluna d’água de ecossistemas lênticos. De acordo com estes
autores, Pythium spp. foram especialmente frequentes, assim como observado neste
estudo. No Brasil, Rocha & Pires-Zottarelli (2002) mencionaram, para o perfil da coluna
d’água do reservatório do Guarapiranga, os mesmos gêneros citados anteriormente,

86
embora Pythium não tenha se destacado como um dos mais frequentes. Já com relação ao
sedimento, Yang & Hong (2015) mencionaram a expressiva frequência de Pythium e
Phytophythium Abad, de Cock, Bala, Robideau, Lodhi & Lévesque, estando estes gêneros
restritos às camadas superficiais do perfil sedimentar, enquanto Wurzbacher et al. (2016)
demonstraram que Rhizophydiales e Cladochytriales apresentaram expressiva frequência
de ocorrência neste compartimento.
Considerando os gradientes trófico e sazonal observados na PCA (Fig. 2), estes
parâmetros foram relacionados à ocorrência e distribuição dos organismos zoospóricos
heterotróficos. A análise de coordenadas principais (PCoA) dos dados bióticos da coluna
d’água (Fig. 11) explicou 51,3% da variabilidade total dos dados, sendo o eixo 1 o que
explicou maior parte desta variação (31,8%). A PCoA indicou que os espécimes de
Dictyuchus estão relacionados com Santa Helena durante a estação chuvosa, enquanto os
espécimes de Achlya estão relacionados ao mesmo reservatório na estação seca, nas quais,
as mais baixas concentrações de fósforo total e nitrogênio total foram observadas. Por
outro lado, os espécimes de Aphanomyces, Pythium com zoosporângios filamentosos não
inflados (grupo P), estão relacionados com Hedberg na estação chuvosa, o qual
apresentou altas concentrações de PT e NT. Rocha (2004) também observou a relação
dos espécimes de Pythium do grupo P com as altas concentrações de NT e PT no lago das
Garças do Parque Estadual das Fontes do Ipiranga (PEFI), São Paulo, SP.

87
Figura 11. Análise de coordenadas principais (PCoA) dos dados bióticos da coluna d’água, durante as
estações seca e chuvosa. Os táxons com somente uma ocorrência ao longo do estudo não foram incluídos
na análise. Legendas: Achkl: Achlya klebsiana; Achspp: Achlya spp; Aphspp: Aphanomyces spp.; Clarep:
Cladochytrium replicatum; Dicmon: Dictyuchus monosporus; Dicspp: Dictyuchus spp., Karlspp:
Karlingiomyces spp.; Nowcre: Nowakowskiella crenulata; Nowspp: Nowakowskiella spp., PysppF:
Pythium spp. grupo F; PysppP: Pythium spp. grupo P; PysppT: Pythium spp. grupo T; Stsec1: Santa
Helena seca ponto 1; Stsec2: Santa Helena seca ponto 2; Stsec3: Santa Helena seca ponto 3; Stchu1: Santa
Helena chuvosa ponto 1; Stchu2: Santa Helena chuvosa ponto 2; Stchu3: Santa Helena chuvosa ponto 3;
Hsec1: Hedberg seca ponto 1; Hsec2: Hedberg seca ponto 2; Hsec3: Hedberg seca ponto 3; Hchu1:
Hedberg chuvosa ponto 1; Hchu2: Hedberg chuvosa ponto 2; Hchu3: Hedberg chuvosa ponto 3; Itsec1:
Itupararanga seca ponto 1; Itsec2: Itupararanga seca ponto 2; Itsec3: Itupararanga seca ponto 3; Itsec4:
Itupararanga seca ponto 4; Itsec5: Itupararanga seca ponto 5; Itchu1: Itupararanga chuvosa ponto 1;
Itchu2: Itupararanga chuvosa ponto 2; Itchu3: Itupararanga chuvosa ponto 3; Itchu4: Itupararanga chuvosa
ponto 4; Itchu5: Itupararanga chuvosa ponto 5.
A PCoA dos dados bióticos do sedimento superficial (Fig. 12) explicou 46,3%
da variabilidade total dos dados, com o eixo 1 explicando maior parte desta variação
(27,5%). A PCoA mostrou que Diplophlyctis sarcoptoides, Cladochytrium replicatum e
os espécimes de Pythium com zoosporângios esféricos (grupo T) estão relacionados com
com Santa Helena durante a estação seca, a qual apresentou baixas concentrações de PT
e NT, enquanto Catenophlyctis variabilis está relacionado com Hedberg durante a estação
chuvosa, onde foi observado altas concentrações de PT, NT e carbono orgânico total.
Rocha (2004) também relacionou as ocorrências de D. sarcoptoides aos locais com baixas

88
concentrações de PT e NT no Lago das Ninféias (São Paulo, SP) e de C. variabilis aos
locais de alta concentração destes nutrientes no Lago das Garças, embora estes relatos
estejam relacionados a distribuição ao longo da coluna d’água, uma vez que este é o
primeiro estudo a investigar a ocorrência destes organismos em sedimentos de
ecossistemas lênticos brasileiros.
Figura 12. Análise de coordenadas principais (PCoA) dos dados bióticos do sedimento superficial, durante
as estações seca e chuvosa. Os táxons com somente uma ocorrência ao longo do estudo não foram incluídos
na análise. Legendas: Achspp: Achlya spp; Aphspp: Aphanomyces spp.; Catvar: Catenophlyctis
variabilis; Chyapp: Chytriomyces appendiculatus; Clarep: Cladochytrium replicatum; Dipsar:
Diplophyctis sarcoptoides; Nowcre: Nowakowskiella crenulata; Nowhem: Nowakowskiella
hemisphaerospora; Nowram: Nowakowskiella ramosa; Nowsp1: Nowakowskiella sp.1; PysppP: Pythium
spp. grupo P; PysppT: Pythium spp. grupo T; Rhizely: Rhizophydium elyense; Stsec1: Santa Helena
seca ponto 1; Stsec2: Santa Helena seca ponto 2; Stsec3: Santa Helena seca ponto 3; Stchu1:
Santa Helena chuvosa ponto 1; Stchu2: Santa Helena chuvosa ponto 2; Stchu3: Santa Helena
chuvosa ponto 3; Hsec1: Hedberg seca ponto 1; Hsec2: Hedberg seca ponto 2; Hsec3: Hedberg

89
seca ponto 3; Hchu1: Hedberg chuvosa ponto 1; Hchu2: Hedberg chuvosa ponto 2; Hchu3:
Hedberg chuvosa ponto 3; Itsec1: Itupararanga seca ponto 1; Itsec2: Itupararanga seca ponto 2;
Itsec3: Itupararanga seca ponto 3; Itsec4: Itupararanga seca ponto 4; Itsec5: Itupararanga seca
ponto 5; Itchu1: Itupararanga chuvosa ponto 1; Itchu2: Itupararanga chuvosa ponto 2; Itchu3:
Itupararanga chuvosa ponto 3; Itchu4: Itupararanga chuvosa ponto 4; Itchu5: Itupararanga
chuvosa ponto 5.
Já que durante as amostragens foi observada maior frequência, abundância e
riqueza de táxons de fungos zoospóricos no sedimento superficial e dos oomicetos na
coluna d’água, podemos rejeitar a hipótese de que estes dois grupos ocorreriam
preferencialmente na coluna d’água. Os poucos relatos de fungos zoospóricos em
amostras de sedimento podem estar mais relacionadas a limitações de amostragens do
que necessariamente aos padrões ecológicos destes fungos. Além disso, como não foram
observadas diferenças significativas na riqueza de espécies entre as estações, não
confirmamos a hipótese de que em épocas mais chuvosas apresentariam uma maior
riqueza de organismos zoospóricos heterotróficos, comparado à estação seca, muito
provavelmente reflexo da baixa pluviosidade observada, principalmente, durante a
estação chuvosa.
Os organismos zoospóricos, apresentaram uma ampla distribuição ao longo de
diferentes gradientes tróficos, assim como mencionado por Rocha (2004), estando
algumas espécies relacionadas aos locais com as mais altas concentrações de nitrogênio
e fósforo total, como Catenophlyctis variabilis, enquanto outras estão à locais com as
mais baixas concentrações destes nutrientes (Diplophlyctis sarcoptoides e Cladochytrium
replicatum) e que poderiam potencialmente ser táxons indicativos da qualidade das
águas. Concluímos, portanto, que os organismos zoospóricos heterotróficos são, no geral,
tolerantes às amplas variações nas concentrações de nitrogênio e fósforo, mas alguns
táxons são limitados pela disponibilidade destes nutrientes, particularmente o fósforo.

90
Referências bibliográficas
Amal, E. A., Aïcha, E. A. & Bernard, P. 2005. Achlya abortispora, a new Oomycete
isolated from water samples taken from a water reservoir in Morroco. Current
Microbiology 53: 60-67.
Amaral, A.M., Barbério, A., Voltolini, J.C. & Barros, L. 2007. Avaliação preliminar
da citotoxicidade e genotoxicidade da água da bacia do rio Tapanhon (SP-Brasil)
através do teste Allium (Allium cepa). Revista Brasileira de Toxicologia 20: 65‑72.
Birks, H.H. & Birks, H.J. 2006. Multi-proxy studies in palaeolimnology. Vegetation
History Archaeobotany 15: 235-251.
Carlson, R.E. 1977. A trophic state index for lakes. Limnology and Oceanography, 22:
361-369.
CEPAGRI – Centro de Pesquisas Meteorológicas e Climáticas Aplicadas a
Agricultura (2018). Disponível online em: http://www.cpa.unicamp.br/outras-
informacoes/clima-dos-municipios-paulistas.html. Acessado em 06 Junho de 2018.
CETESB – Companhia Ambiental do Estado de São Paulo. 2013. Relatório de
qualidade das águas superficiais no Estado de São Paulo. Parte 1. São Paulo, 301 p.
Chen, M., Chen, F., Yu, Y., et al. 2008. Genetic diversity of eukaryotic microorganisms
in Lake Taihu, a large shallow subtropical lake in China. Microbiology Ecology 56:
572−583
CONAMA, Conselho Nacional do Meio Ambiente. 2005. Resolução n° 357, de 17 de
março de 2005. Ministério do Meio Ambiente. p. 23.
Colins, C. & Willoughby, L.G. 1962. The distribution of bacteria and fungal spores in
Blelham Tarn with particular reference to an experimental overturn. Archiv für
Mikrobiologie 43: 294-307.
Cooke, W.B. 1976. Fungi in sewage. In. Gareth Jones, E.B (Ed.). recent advances in
Aquatic Mycology. London: Elek Science p. 389-434.
Czeczuga, B. Woronowicz, L., Brzozowska, K. & Chomutowska, H. 1989. Studies of
aquatic fungi IX. Mycoflora of different types springs. Acta Hydrobiologica 31: 273-
283.
Czeczuga, B. Woronowicz, L., Brzozowska, K. 1990a. Studies of aquatic fungi XII.
Aquatic fungi of the lowland river Biebrza. Acta Hydrobiologica 26: 77-83.

91
Czeczuga, B. Woronowicz, L., Brzozowska, K. 1990b. Studies of aquatic fungi XIII.
Mycoflora of river Czarna Hancza and its tributary, the river Marycha. Internationale
Revue der gesamten Hydrobiologie 75: 245-255.
Czeczuga, B. 1991a. Studies of aquatic fungi. Part XVI. Aquatic fungi of the river Pisa
and its tributary, the river Skroda. Acta hydrochimica et hydrobilogica 19: 57-65.
Czeczuga, B. 1991b. Studies of aquatic fungi. Part XXII. Mycoflora of the river
Wegorapa and its tributary, the river Goldapa-Jarka. Acta hydrochimica et
hydrobilogica 19: 517-528.
Czeczuga, B. & Woronowicz, L. 1991. The lake Mamry complex. Acta Mycologica 27:
93-103.
Czeczuga, B. 1994. Studies of aquatic fungi. Part XVII. Aquatic fungi of Lake Hancza
in the Suwalki Scenic Park and some adjacents lakes (northeastern Poland). Acta
Hydrobiologica 36: 371-385.
Czeczuga, B. & Muszynska, E. 2001. Zoosporic fungi growing on Gymnosperm pollen
in water of variety trophic state. Polish Journal of Environmental Studies 10: 89-94.
Dawson, S.C. & Pace, N.R. 2002. Novel Kingdom-level eukaryotic diversity in anoxic
environents. Proceedings Natural Academic Sciences of USA 99: 8324-8329.
Esteves, F.A. 2011. Fundamentos de limnologia. Rio de Janeiro. Interciencia. 3 ed. 826p.
FLONA – Floresta Nacional de Ipanema, Plano de Manejo. 2016. Instituto Chico
Mendes de Preservação da Biodiversidade.
Gerphagnon, M., Latour, D., Colombet, J. & Sime-Ngando, T. 2013. A double
staining method using SYTOX green and calcofluor white for studing fungal parasites
of phytoplankton. Applied and Environmental Microbiology
dx.doi.org/10.1128/AEM.00696-13.
Gleason, F. H., Karpov, S.A., Lilje, O., Macarthur, D.J. et al. 2014. In: Gareth Jones,
E. B., Hyde, K. D. e Pang, K. (Eds.). Zoosporic parasites of phytoplankton. Walter
de Gruyter GmbH, Berlin/Boston, pp.279-304.
Golterman, H.L. & Clymo, R.S. 1969. Methods for chemical analysis of freshwaters.
Oxford and Edinburg: Blackwell Scientific Publications. International Biological
Programme. 171p.
Grossart, H.P. & Rojas-Jimenez, K. 2016. Aquatic fungi: targeting a forgotten in
microbial ecology. Current opinion in Microbiology 31: 140-145.

92
Jerônimo, G.H., Jesus, A.L., Rocha, S.C.O. Gonçalves, D.R. et al. 2017. New insights
into Plectospira genus (Oomycetes, Straminipila): morphological and molecular
analysis. Phytotaxa 307: 191-198.
Kagami, M., E. Van Donk, A. de Bruin, M. Rijkeboer & B. W. Ibelings, 2004.
Daphnia can protect diatoms from fungal parasitism. Limnology and Oceanography
49: 680-685.
Kagami, M., Bruin, A., Ibelings, B.A. & Donk, E.V. 2007. Parasitic chytrids: their
effects on phytoplankton communities and food-web dynamics. Hydrobiologia: 578:
113-129.
Kagami, M.; Helmsing, N. R.; Van Donk, E. 2011. Parasitic chytrids could promote
copepod survival by mediating material transfer from inedible diatoms.
Hydrobiologia 659: 49-54.
Kagami, M., Miki, T. & Takimoto, G. 2014. Mycoloop: chytrids in aquatic food webs.
Frontiers in Microbiology doi: 10.3389/fmicb.2014.00166.
Lamparelli, M.C. 2004. Grau de trofia em corpos d’água do estado de São Paulo:
avaliação dos métodos de monitoramento.Tese de doutorado, Universidade de São
Paulo, 235 p.
Lefèvre, E., Bardot, C., Noël, C., Carrias, J.F., Viscogliosi, E. et al. 2007. Unveiling
fungal zooflagellates as members of freshwater picoeucakaryotes: evidence from a
molecular diversity study in a deep meromictic lake. Environmental Microbiology: 9:
61-71.
Lefèvre, E., Letcher, P. & Powell, M. 2012. Temporal variation of small eukaryotic
community in two freshwater lakes: emphasis on zoosporic fungi. Aquatic microbial
Ecology67: 91-105.
Lefranc, M., Thénot, A., Lepère, C. & Debroas, D. 2005. Genetic diversity of small
eukaryotes in lakes differing by their trofic status. Application Environmental
Microbiology 71: 5935-5942.
Lepère, C., Domaizon I. & Debroas, D. 2008. Unexpected importance of potencial
parasites in the composition of the freshwater small-eukaryote community. Applied
and Environmental Microbiology 74: 1029-1041.
Lucinda, I. 2003. Composição de Rotifera em corpos d’água da bacia do rio Tietê – São
Paulo, Brasil. Dissertação de mestrado. Universidade Federal de São Carlos: São
Carlos. 199p.

93
Marano, A.V., Zottarelli, C.L.A., Barrera, M.D. et al. 2011. Diversity, role in
decomposition, and sucession of zoosporic fungi and straminipiles on submerged
decaying leaves in a woodland stream. Hydrobiologia 659: 93-109.
Margalef, R. 1983. Limnologia. Omega, Barcelona.
McCune, B. & Mefford, M.J. 2011. PC-ORD. Multivariate analysis of ecological data.
Version 6. MjM Software, Gleneden Beach, Oregon, USA.
Milanez, A.I. 1989. Fungos de águas continentais. In: O. Fidalgo, V.L. Bononi (coords.).
Técnicas de coleta, preservação e herborização de material botânico. Série
Documentos. São Paulo, Instituto de Botânica. pp. 17-20.
Monchy, S., Sanciu, G., Jobard, M., Rasconi. et al. 2011. Exploring and quantifying
fungal in freshwater lake ecosystems using rDNA cloning/sequencing and SSU tag
pyrosequencing. Environmental Biology 13: 1433-1453.
Müeller-Dombois, D. & Ellemberd, H. 1974. Aims and methods of vegetation ecology.
New York, Jonh Wiley & Sons. pp. 547.
Nascimento, C.A., Gomes, E.P.C. & Pires-Zottarelli, C.L.A. 2011. Occurrence and
distribution of zoosporic organisms in water bodies from Brazilian Cerrado.
Mycologia 103: 261-272.
Pedrazzi, F.J.M., Conceição, F.T., Sardinha, D.S., Moschini-Carlos, V. et al. 2014.
Avaliação da qualidade da água do reservatório de Itupararanga, Bacia do Alto
Sorocaba. Geociências 33: 26-38.
Pires-Zottarelli, C.L.A. 1990. Levantamento dos fungos zoospóricos da Represa do
Lobo (“Broa”), São Carlos, SP, Brasil [Master dissertation]. São Paulo: Universidade
Estadual Paulista. 175 p.
Plaats-Niterink, V.J. 1981. Monograph of the genus Pythium. Studies in Micology 21:
1-244.
Pusceddu, A., Gambi, C., Manini, E. & Danovaro, R. 2007. Trophic state, ecosystem
efficiency and biodiversity of transitional aquatic ecosystems: analysis of
environmental quality based on different benthic indicators. Journal of Chemical
Ecology 23: 505‑515.
Rasconi, S. Jobard, M., Jouve, L. & Sime-Ngando, T. 2009. Use of calcofluor white
for detection, identification, and quantification of phytoplanktonic fungal parasites.
Applied and Environmental Microbiology 75: 2545–2553.

94
Rasconi, S.; Niquil, N.; Sime-Ngando, T. 2012. Phytoplankton chytridiomycosis:
community structure and infectivity of fungal parasites in aquatic ecosystems.
Environmental Microbiology, Bedford, v. 14, p. 2151–2170.
Rasconi, S., Grami, B., Niquil, N., Jobard, M. et al. 2014. Parasitic chytrids sustain
zooplankton growth during inedible algal bloom. Frontiers in Microbiology 5: 1-20.
Rocha, M. & Pires-Zottarelli, C.L.A. 2002. Chytridiomycota o Oomycota da represa
do Guarapiranga. Acta Botanica Brasilica 16: 287-309.
Rocha, M. 2004. Micota zoospórica de lagos com diferentes trofias do Parque Estadual
das Fontes do Ipiranga (PEFI), São Paulo, SP. Dissertação para obtenção do título de
Mestre em Microbiologia para o Instituto de Ciências Biomédicas da Universidade de
São Paulo, p. 1-85.
Sartory & Groberlaar. 1984. Extraction of chlorophyll-a from freshwater
phytoplankton for spectrophotometric analysis. Hidrobiology 114: 177-187.
Shearer, C.A., Descals, E., Kohlmeyer, B., Kohlmeyer, J. et al. 2007. Fungal
biodiversity in aquatic habitats. Biodiversity Conservation 16: 49-67.
Secchin, L.F. 2012. Caracterização ambiental e estimativa da produção de cargas difusas
na área de drenagem da represa de Itupararanga. Dissertação de mestrado.
Universidade de São Paulo: São Paulo. 122p.
Slapeta, J., Moreira, D., Purificación, L.G. 2005. The extend of protist diversity:
insights from molecular ecology and freshwater. Proceedings of the Royal Society B
272: 2073-2081.
Smol, J.P. 2008. Pollution of lakes and rivers: a paleoenvironmental perspective.
Blackewell, 2 ed., Oxford University Press, New York.
Sparrow. F. K. 1968. The Fungi: An Advanced Treatise. In: Ainsworth, G.C. Sussman,
A.S. (eds.). The Fungal Population III, Academic Press, New York.
Tolmasquim, M.T. 2012. Perspectivas e planejamento do setor energético no Brasil.
Estados Avançados 26: 247-260.
Tundisi, J.G., Matsumura-Tundisi, T. & Tundisi, J.E.M. 2008. Reservoirs and human
well being: new challenges for evaluating impacts and benefits in the neotropics.
Brazilian Journal of Biology 68: 1133-1135.
Valderrama, G.C. 1981. The simultaneous analysis of total nitrogen and total
phosphorus in natural waters. Marine Chemistry 10: 109-122.
Wurzbacher, C.M., Barlocher, F. & Grossart, H.P. 2010. Fungi in lake ecosystems.
Aquatic microbial Ecology 59: 125-149.

95
Wurzbacher, C.M., Whartmann, N., Bourne, E., Attermeyer, K. et al. 2016. High
habitat-specific in fungal communities of oligo-mesotrophic temperate Lake Stechlin
(North-West Germany). Mycokeys 16: 17-44.
Wetzel, R.G. 2001. Structure and productivity of aquatic ecossystems. In: Limnology.
Academic Press, San Diego, pp. 206.
Widenfalk, A., Bertilsson, S., Sundh, I. & Goedkoop, W. 2008. Efects of pesticides on
community composition and activity of sediment microbes – response at various
levels of microbial community organization. Enviromental Polution 152: 576-584.
Wong, M.K.M., Goh, T.K., Hodgkiss, I.J. et al. 1998. Role of fungi in freshwater
ecossystems. Biodiversity and Conservation 7: 1187-1206.
Yang, X., & Hong, C.X. 2015. Diversity and populations of Phytophthora, Phytopythium
and Pythium species recovered from sediments in an agricultural run‐off
sedimentation reservoir. Plant Pathology 65: 1118-1125.
Zak, J.C. & Willig, M.R. 2004. Fungal biodiversity patterns. In: G.M. Mueller, G.F.
Bills, M.S. Foster (eds.). Biodiversity of fungi: Inventory and monitoring methods.
Elsevier Academic Press, San Diego, pp. 59–75.
Zappia, R.E., Hüberli, D., Hardy, G.S.J. & Bayliss, K.L. 2014. Fungi and oomycetes
in open irrigation systems: knowledge gaps and biosecurity implications. Plant
Pathology 63: 961-972.

96
CAPÍTULO 2
NOVELTIES FOR THE NOWAKOWSKIELLA GENUS (CLADOCHYTRIALES;
CHYTRIDIOMYCETES): NEW SPECIES AND NEW COMBINATION
Artigo elaborado de acordo com as normas da revista Mycologia

97
Novelties for the Nowakowskiella genus (Cladochytriales; Chytridiomycetes): new
species and new combination
Gustavo H. Jerônimo1*, Ana L. Jesus1, D. Rabern Simmons2, Timothy Y. James2, Carmen
L.A. Pires-Zottarelli1
1 “Núcleo de Pesquisa em Micologia”, Instituto de Botânica, Sao Paulo, Sao Paulo 04301-902
2 Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, Michigan 48109-
1085
* Corresponding author. Email: [email protected]
Abstract: Five Nowakowskiella species were identified and purified on nutrient agar and
included in phylogenetic analyses, this being the first such study for four of these species.
Our combined maximum likelihood analyses from SSU, 5.8S (ITS) and LSU regions of
rDNA showed that Nowakowskiella is not a monophyletic clade due the position of
Nowakowskiella elongata, which is more related to Cladochytrium. Therefore, we
reclassify N. elongata as the type of the new genus Karlingiella, in addition to describing
the new species N. crenulata within the monophyletic Nowakowskiella.
Key words: chytrid, Karlingiella, Nowakowskiella crenulata, phylogeny
INTRODUCTION
The advancement and popularization of molecular tools have deeply influenced
the systematics of several groups of fungi. In the basal Chytridiomycota (chytrid fungi or
chytrids), molecular data corroborating the relationships suggested by zoospore
ultrastructure have revealed that the classical taxonomic system, based on convergently-
derived thalli structures, is not phylogenetically accurate (Letcher et al. 2006; Letcher
and Powell 2014). Thus, numerous nomenclatural changes have been proposed, including
the description of new species, genera, families and orders (Letcher et al. 2005; James et
al. 2006; Letcher et al. 2006; Mozley-Standridge et al. 2009; Simmons et al. 2009;
Longcore and Simmons 2012; Vélez et al. 2013; Karpov et al. 2014) and major revisions
and reclassifications of polyphyletic groups (Vélez et al. 2011; Letcher and Powell 2014).

98
The description of the Cladochytriales, to include some genera classically
included in Chytridiales, was based on these species’ monophyly, independent of the
Chytridiales, in small (SSU) and large (LSU) subunit ribosomal DNA molecular
phylogenies (Mozley-Standridge et al. 2009). Additionally, of those examined by
transmission electron microscopy, they all produce zoospores with a unique constellation
of ultrastructural characters (Lucarroti 1981, Mozley-Standridge et al. 2009).
Representatives of this order have different types of development, reproductive
structures, and ecological preferences. The polycentric genera Nowakowskiella Schröt.
and Cladochytrium Nowak. are the largest of the order, yet only a few species have been
isolated, cultured, and included in molecular phylogenies, keeping their interspecific
relationships, and indeed their generic monophyly, uncertain.
Nowakowskiella was proposed by J. Schrot in 1893 to include the type species N.
elegans, previously named Cladochytrium elegans Nowak. Over the years, several
species have been described and currently this genus contains 18 legitimate species
(Mycobank 2018), which are morphologically recognized by production of polycentric
thalli, with operculated zoosporangia interconnected by rhizomycelia, which contain
occasional non-septate swellings (Sparrow 1960; Karling 1977). Originally included in
the Nowakowskiellaceae by Sparrow (1942), this genus was transferred shortly thereafter
to Megachytriaceae (Sparrow 1943). However, neither family was validly published, and
Mozley-Standridge et al. (2009) validly reinstated the original family name. Though taxa
have been recently reshuffled in higher taxonomic groups, the last new species, N.
keratinophila Hassan and Batko, was described around 20 years ago (Hassan & Batko
1988).
Therefore, the aim of this study is to introduce the new species Nowakowskiella
crenulata Jerônimo & Pires-Zottarelli, include N. multispora Karling and N. ramosa
Butler for the first time in phylogenetic reconstructions, besides to transfer N. elongata
Karling to a new genus, Karlingiella, and increase the knowledge about the phylogenetic
relationship within this order.

99
MATERIALS AND METHODS
Isolation and culture.—Sediment surface, soil and water samples were collected on lentic
and lotic freshwater habitats from several locations in the Brazilian Atlantic Rainforest
(TABLE I). These samples were baited with cellulosic substrates (onion skin, corn leaves,
and cellophane), infected substrates were aseptically removed and placed on nutrient agar
medium, on which chytrids were further isolated into pure culture through the collection
of aseptic zoospores and/or rhizomycelium onto fresh medium, including PmTG (1 g
peptonized milk, 1 g tryptone, 5 g dextrose, 8 g agar, 0.5 g streptomycin sulfate, 0.5 g
penicillin-G, 1 L deionized water), CMA plus glucose (16 g corn meal agar, 2.5 g glucose,
1 L deionized water) or YPSS/8 (4 g yeast extract, 15 g soluble starch, 1 g K2HPO4, 0.5
g MgSO4 × 7 H2O, 1 L deionized water). Pure cultures were deposited in the MMBF
culture collection (“Micoteca Mário Barreto Figueiredo”) belonging to WFCC (World
Federation for Culture Collections) and also to the CCIBt collection culture (“Coleção de
Cultura de Algas, Fungos e Cianobácterias do Instituto de Botânica”). Additionally, the
strains were cryopreserved according to the protocol of Boyle et al. (2003).
The collection culture or voucher numbers, the geographic coordinates and type
of samples and habitats collected are showed in the TABLE I.
TABLE I. Species, collection culture or voucher numbers, geographic coordinates and type of
samples and habitats collected for the 14 Cladochytriales strains isolated during this study.
Species
Collection
culture or
voucher
numbers
Geographic coordinates Municipality/State/
Country
Samples
(habitats)
Cladochytrium
replicatum CCIBt4014
25°03’05”/25°18’18”S-
47°53’48”/48°05’42”W
Cananéia/Sao
Paulo State/Brazil
water
(stream)
Cladochytrium
replicatum CCIBt3845 23°38’20,6”S/46°37’34,3”W
São Paulo/São
Paulo State/Brazil
water
(stream)
Cladochytrium
replicatum CCIBt4263
23°25'33,72''S/47°35'42,66''
W
Votorantim/São
Paulo State/Brazil
water
column
(reservoir)
Cladochytrium
replicatum ALJ05
24°30’25,09”S/47°15’42,08”
W
Juréia/São Paulo
State/Brazil
waterfall
(stream)
Nowakowskiella
crenulata CCIBt4258
23°34'49,92''S/47°25'32,64''
W
Votorantim/São
Paulo State/Brazil
sediment
surface
(reservoir)
Nowakowskiella
crenulata CCIBt4259
23°34'54,30''S/47°26'12,12''
W
Ibiúna/São Paulo
State/Brazil
sediment
surface
(reservoir)
Nowakowskiella
elongata CCIBt4016
25°03’05”/25°18’18”S-
47°53’48”/48°05’42”W
Cananéia/São
Paulo State/Brazil
water
(stream)
100
Species
Collection
culture or
voucher
numbers
Geographic coordinates Municipality/State/
Country
Samples
(habitats)
Nowakowskiella
elongata ALJ09
24°30’04,07”S/47°15’58,0”
W
Juréia/São Paulo
State/Brazil
submerged
leaves
(river)
Nowakowskiella
hemisphaerospora GHJ13
23°25'40,86''S/47°35'31,14''
W
Cananéia/São
Paulo State/Brazil
sediment
surface
(reservoir)
Nowakowskiella
multispora CCIBt4015
25°03’05”/25°18’18”S-
47°53’48”/48°05’42”W
Cananéia/São
Paulo State/Brazil
water
(stream)
Nowakowskiella
multispora CCIBt3864 23°38’18,8”S/46°37’31,7”W
São Paulo/São
Paulo State/Brazil
water
(stream)
Nowakowskiella
ramosa CCIBt4294
23°34'49,92''S/47°25'32,64''
W
Votorantim/São
Paulo State/Brazil
water
column
(reservoir)
Nowakowskiella sp.1 CCIBt4260 23°34'58,56''S/47°25'50,52''
W
Votorantim/São
Paulo State/Brazil
sediment
surface
(reservoir)
Nowakowskiella sp. ALJ23 24°22’51,04”S/47°20’39,0”
W
Juréia/São Paulo
State/Brazil soil
Polychytrium
aggregatum CCIBt4017
25°03’05”/25°18’18”S-
47°53’48”/48°05’42”W
Cananéia/São
Paulo State/Brazil soil
Polychytrium
aggregatum ALJ30
24°32’28,1”S/47°12’38,08”
W
Juréia/São Paulo
State/Brazil water (river)
Nowakowskiella species produce thick-walled resting spores that do not germinate
on standard nutrient agar media, and these structures are indicative of certain species. In
order to stimulate resting spore production, we cultivated our Nowakowskiella strains on
YPSS/8 media prepared with soil water as described by Lucarotti (1981). Lucarotti
argued that the micro-nutrients provided by the soil could stimulate the production of
zoospores, and we proposed to test if the same theory held true to resting spore
production. We applied 7 g of garden soil and 0.05 g calcium carbonate to 250 ml of
distillated water, steamed this mixture for 2 hours during two consecutive days, and then
filtered the solution twice with a Buchner funnel and Whatman n°1 filter paper to remove
all particulate material. After that, the solution was diluted in the ratio of two parts of
distillated water to 1 part of filtered solution, autoclaved, and used to prepare the YPSS/8
culture medium.
Morphology.—Pure cultures were observed growing in PmTG, CMA plus glucose and
YPSS/8 broth and on purified cellulosic substrates (onion skin, cellophane and corn
leaves). Examination of thallus morphology of all strains was performed on Leica
DMLB2. We used the Sparrow (1960) and Karling (1977) bibliographies along with the
original description of the species (Butler 1907, Shanor 1942, Karling 1944, 1963) to
identify our isolates.

101
DNA extraction, amplification and sequencing.—For DNA extraction, a small piece of
agar with zoosporangia or rhizomycelium was transferred to Erlenmeyer flasks
containing 50 ml of PmTG liquid medium prepared with autoclaved reserve-osmosis
water. After incubation for 5-10 days at 21C, the material was transferred to 2 mL
microcentrifuge tubes to obtain pellets for DNA extraction procedure. The extraction of
genomic DNA followed the protocol described in the PureLink Genomic DNA kit
(Invitrogen©). Genomic DNA was visualized by electrophoresis of the extracts on a 0.8%
agarose gel. Ribosomal regions were amplified with the PCR SuperMix kit (Invitrogen©)
for a final volume of 25 L, and the partial LSU, SSU and complete ITS regions of rDNA
were amplified using the primers LR0R/LR5, SR1R/NS4 (Vilgalys and Hester 1990) and
ITS4/ITS5 (White et al. 1990), respectively. Thermocycler parameters followed the cycle
described in James et al. (2006). Amplicons were purified with AxyPrep PCR Clean-up
kit (Axygen©) and sequenced at the “Centro de Pesquisa sobre o Genoma Humano e
células tronco, Universidade de São Paulo”. The contiguous sequences were assembled
into SEQUENCHER 5.3.
.Phylogenetic analyses.—For phylogenetic reconstruction, 24 isolates of Cladochytriales
were selected (TABLE II), which represent the major genera with available sequences in
GenBank while Polychytrium aggregatum strains (CCIBt4017, ALJ30 and JEL109) were
used as Polychytriales outgroup taxa. In addition, we are including 14 new
Cladochytriales strains isolated from different habitats at the Brazilian Atlantic Rainforest
(TABLE I). The contiguous sequences were assembled using Sequencher version 4.1.4
(Gene Codes) and alignment was performed online at Mafft version v. 7.058 (Kazutaka
and Daron 2013). The regions SSU, 5.8S (ITS) and LSU were concatenated through
SequenceMatrix v. 1.8 (Vaidya et al. 2010) resulting in a final length of 2427 base pairs.
The Maximum Likelihood (ML) analysis was conducted in Garli v. 2.01 (Bazinet and
Cummings 2008) adding the partition models generated by jModel Test v. 2.2.4 (Posada
2008). We performed the ML analysis with 1000 bootstrap replicates and the values
<70% were not shown in the final tree.

102
TABLE II. Species, strain/voucher and GenBank accession numbers. The isolates highlighted in
bold have never been cultured, sequenced, or included in phylogenetic analyses. NA: Not
available
Species Strain/voucher GenBank accession number
SSU ITS LSU
Allochytridium luteum ATCC 60989 JN940948 NA AY439066
Catenochytridium sp. JEL 145 EU828475 NA EU828503
Catenochytridium sp. JEL 044 EU828478 NA EU828506
Cladochytrium replicatum CCIBt4263 MH590084 MH590046 MH590071
Cladochytrium replicatum JEL 303 EU828461 NA EU828488
Cladochytrium replicatum CCIBt3845 MH590085 MH590047 MH590072
Cladochytrium replicatum CCIBt4390 MH590086 MH590048 MH590073
Cladochytrium replicatum JEL 180 NG017169 NA NG027614
Cladochytrium replicatum WJD 123 NA NA KC691378
Cladochytrium replicatum CCIBt 4014 KJ464414 NA KJ464415
Cladochytrium sp. SMS 013 EU828459 NA EU828486
Cladochytrium sp. JEL 153 EU828458 NA EU828485
Cladochytrium sp. BR 696 JN940946 JN943816 JN941001
Cylindrochytridium johnstonii JEL 596 JF796051 NA JF796052
Endochytriaceae JEL 072 EU828470 NA EU828497
Endochytrium ramosum JEL 402 EU828484 NA EU828513
Endochytrium sp. JEL 324 AY635844 AY997044 DQ273816
Nephrochytrium sp. JEL 125 AH009049 NA EU828511
Nowakowskiella elegans M 29 NA AY353257 AY349080
Nowakowskiella elegans JEL 046 EU828463 NA EU828490
Nowakowskiella elegans UCB 50 1 EU828464 NA EU828491
Nowakowskiella elegans JEL 157 EU828465 NA EU828492
Nowakowskiella elongata CCIBt4016 MH590087 MH590049 MH590074
Nowakowskiella elongata ALJ09 NA MH590050 MH590075
Nowakowskiella hemisphaerospora GHJ13 MH590088 MH590051 MH590076
Nowakowskiella multispora CCIBt3864 MH590089 MH590052 MH590077
Nowakowskiella multispora CCIBt4015 KJ539147 KJ539148 KJ539149
Nowakowskiella ramosa CCIBt4294 MH590090 NA MH590078
Nowakowskiella sp. ALJ 23 MH590091 MH590053 MH590079

103
Species Strain/voucher GenBank accession number
SSU ITS LSU
Nowakowskiella elegans JEL 127 EU828466 NA DQ273798
Nowakowskiella sp. JH HBR EU828469 NA EU828496
Nowakowskiella sp. JH CC2 EU828481 NA EU828509
Nowakowskiella sp. JH AS EU828482 NA EU828510
Nowakowskiella crenulata CCIBt4258 MH590092 MH590054 MH590080
Nowakowskiella crenulata CCIBt4259 MH590093 MH590055 MH590081
Nowakowskiella sp.1 CCIBt4260 MH590094 MH590056 MH590082
Polychytrium aggregatum JEL 109 NG017168 AY997074 AY349084
Polychytrium aggregatum ALJ30 MH590095 MH590057 MH590083
Polychytrium aggregatum CCIBt 4017 KJ464408 KJ464409 KJ464410
Septochytrium sp. JEL 177 EU828474 NA EU828502
Septochytrium variabile JEL 191 EU828483 NA EU828512
RESULTS
We were able to isolate 10 Nowakowskiella strains (TABLE I and II), six of which
could be identified as described species. The YPSS/8 soil media was effective at
stimulating the resting spore production of Nowakowskiella crenulata, but the same was
not observed with two unidentified strains, Nowakowskiella sp.1 CCIBt4260 and
Nowakowskiella sp. ALJ23, which only produced zoosporangia. Considering this, we
decided to maintain our identification of these both strains at generic level.
Brief morphological comments besides some pictures were included below
considering the poor availability information on these taxa.
Morphology.—Nowakowskiella ramosa Butler produces spherical to pyriform
zoosporangia and resistance structures with a parenchymal region (Pr), which support a
single or up to three resting spores with small incrustation in the wall (FIG. 1A–C). N.
hemisphaerospora produces ovoid to pyriform zoosporangia and septate resting spores,
that form two hemispheres, in which all contents migrate to one side while the other
remains empty (FIG. 1D,E). N. multispora produces subspherical to pyriform
zoosporangia and intercalary resistance structures, formed by consecutive and linked
resting spores with a homogenous content (FIG. 1F,G). Nowakowskiella sp.1 (FIG. 1H–

104
N) produces intercalary or terminal zoosporangia, which are commonly spherical,
subspherical or pyriform in shape (FIG. 1H–L), with a small discharge tube (FIG. 1K,N).
Additionally, prominent apophyses were observed (FIG. 1L) that are produced on both
sides of the zoosporangia when the development are intercalary. The zoospores have a
single hyaline lipid globule (FIG. 1M) and are released in a membrane-bound mass (FIG.
1K), in which the zoospores remain immobile for a short period before becoming mobile
and rupturing the binding membrane.
FIG. 1. Zoosporangia and resting spores morphology of Nowakowskiella ramosa, N. hemisphaerospora, N.
multispora and Nowakowskiella sp.1 CCIBt4260. A-C. Nowakowskiella ramosa CCIBt4294. A-B.
Ornamentate resting spores with a parenchymal region (Pr). C. Operculate (Op) zoosporangia. D-E.
Nowakowskiella hemisphaerospora GHJ13. D. Septate resting spores. E. Pyriform zoosporangia. F-G.
Nowakowskiella multispora CCIBt4015. F. Operculate (Op) zoosporangia. G. Resting spores. H-N.
Nowakowskiella sp. 1 CCIBt4260. H, I, J and L. Intercalary apophysate (Ap) zoosporangia. K, M.
Zoospores discharge. N. Mature and imature intercalary zoosporangia with a discharge tube (Tb). Ap:
Apophysis. Op: Operculum. Pr: Parenchymal region. Tb: discharge tube. Bars=10µm.

105
Phylogenetic analyses.—The combined data (SSU, 5.8S and LSU of the rDNA) had 2427
characters, with 1186 parsimony-informative sites. jModelTest indicated the most
appropriate models of DNA substitution were TrNef+I (SSU), TPM1uf+G (5.8S-ITS)
and TIM3ef+I+G (LSU), according to AIC criteria. Two strongly supported clades were
observed in our phylogenetic reconstruction, which represent the Nowakowskiellaceae
and Cladochytriaceae families proposed by Mozley-Standridge et al. (2009), though
internal configurations are altered and support values, specifically for the
Nowakowskiellaceae, are diminished (FIG. 2). In Nowakowskiellaceae, N. ramosa and
Nowakowskiella sp. (ALJ23) are sister group of the type species (N. elegans), the new
species N. crenulata is related to Nowakowskiella sp.1 (CCIBt4260), and N. multispora
in on a basal branch to other taxa in this genus. Clades of cultures identified as N. elegans
and Cladochytrium replicatum were monophyletic, respective to the species, but internal
branches suggest that these species could represent multiples lineages or possibly even
additional species, though more study would be required. In addition, the phylogenetic
placement of our Nowakowskiella elongata strains indicate that this species is not
associated to the other Nowakowskiella representatives and belongs within the new genus
named as Karlingiella, related to Cladochytriaceae and Nephrochytrium sp. JEL125.
FIG. 2. Phylogram inferred from Maximum Likelihood (ML) analyses of 37 ingroup isolates in
Cladochytriales order based on combined (SSU+5.8S+LSU) sequences data. Numbers at nodes represent

106
ML values. Strains highlighted in bold were isolated during this study and in grey represent the taxonomic
novelties.
TAXONOMY
We are proposing the new species, Nowakowskiella crenulata Jeronimo & Pires-
Zottarelli, based on the singular morphology of the zoosporangia and resting spores, along
with its phylogenetic placement. This species was isolated from sediment reservoir
samples and produces a characteristic resting spore (with crenulated ornamentations and
a septum delimiting the branch that supports this structure) and zoosporangia with
prominent and clavate apophyses. Also, we are transferring the species Nowakowskiella
elongata Karling to the new genus Karlingiella Jerônimo, Jesus & Pires-Zottarelli, based
on its disparate phylogenetic position as sister to the Cladochytriaceae.
Nowakowskiella crenulata G.H. Jerônimo and C.L.A. Pires-Zottarelli, sp. nov.
FIG. 3A-J
Typification: BRAZIL. SÃO PAULO. Votorantim, Santa Helena reservoir,
(23°34’54,30”S-47°26’12,12”W), from sediment samples baited with cellulosic
substrates (onion skin and corn leaves), 20 Mar 2014. Holotype preserved in lactophenol
and cottom blue and deposited in Herbarium SP. Ex-type, G.H. Jerônimo CCIBt4258 and
paratype G.H. Jerônimo CCIBt4259. GenBank: SSU: MH590092, MH590093; ITS:
MH590054, MH590055; LSU: MH590080, MH590081.
Fungus saprophytic. Polycentric rhizoidal system, branched, profuse, with numerous non-
septate swellings, broadly fusiform 4.5-5.0 × 3.0-4.5 µm or spherical 3.0-5.0 µm.
Zoosporangia terminal, occasionally intercalary, hyaline, smooth, spherical 20-23 µm,
subspherical 19-24 × 20-27 µm or ovoid 21-25 × 23-30 µm, usually apophysate, without
a discharge tube; wall thin, smooth, colorless; operculum smooth. Apophysis clavate 10-
25 × 5.0-12 µm or nearly spherical 10-14 m. Zoospores spherical, 2-3 µm in diameter,
with a single hyaline lipid globule. Resting spores spherical 7.0-7.5 µm or subspherical
7.0-7.5 × 7.0-7.5 µm in diameter, ornate with crenulated invaginations in the wall and
supported by a septate branch.

107
Etymology: The specific epithet refers to the shape of the resting spores, which produces
small crenulated invaginations in the wall.
Notes. The strains CCIBt4258 and CCIBt4259 grow indeterminantly on PmTG, CMA
plus glucose or even YPSS/8 media, but the resting spores production was only observed
in YPSS/8 broth. Both strains were isolated from sediment surface samples in a
oligotrophic reservoir (“Santa Helena” - 23°34’49,92”S-47°25’32,64”W ), but we were
also able to bait it from sediment samples in a eutrophic reservoir (“Hedberg” -
23°25’40,86”S-47°35’31,04”W).
FIG. 3. Morphology of Nowakowskiella crenulata on onion skin and YPSS/8 broth. A. Encysted zoospores in
YPSS/8. B. Early development stages in YPSS/8 broth. C-D. Zoosporangia and prominent clavate apophysis (Ap) on
onion skin. E-F. Operculate zoosporangia (Op) on onion skin. G. Rhizomycellium with non-septate swellings in
YPSS/8. H-J. Crenulate resting spores with a delimiting septum (Sp) in YPSS/8. Ap: Apophysis. Op: Operculum. Sp:
Septum. Bars=10µm.

108
Karlingiella Jerônimo, Jesus and Pires-Zottarelli, gen. et. nov.
Fungus saprophytic. Polycentric rhizoidal system, branched, profuse, with numerous non-
septate swellings. Operculate zoosporangia, terminal or intercalary, which could produce
a basal sterile portion, elongate, cylindrical or globose. Zoospores spherical, with a single
hyaline lipid globule. Resting spores intercalary, spherical or oval, hyaline, containing a
large refractive globule.
Etymology: The prefix of the genus name is proposed to honor the researcher John S.
Karling, who originally described several Nowakowskiella species during his visit to the
Brazilian Amazon Jungle, including N. elongata.
Type species: Karlingiella elongata (Karling) Jerônimo, Jesus and Pires-Zottarelli
Karlingiella elongata (Karling) Jerônimo, Jesus and Pires-Zottarelli, comb. nov.
FIG.4A-I
Basyonim: Nowakowskiella elongata Karling
Typification: As Karling did not designate a type in the original description (Karling
1944), we considered the illustrations (FIGS. 30-44 Karling 1944) as the lectotype.
Fungus saprophytic. Polycentric rhizoidal system, branched, profuse, with numerous non-
septate swellings, broadly fusiform 7-15 × 5-8 µm or spherical 6-15 µm. Zoosporangia
terminal or intercalary, frequently with a basal septate sterile portion, curved, elongate
25-85 × 10-33 µm, with an apical operculum. Zoospores spherical, 4-6 µm with a single
hyaline lipid globule, forming a temporary globular mass during the realizing process.
Intercalary resting spores, commonly spherical 16.5-33 µm, oval 20-25 × 18-23 µm,
hyaline, with a large refractive globule surrounded by several smaller droplets.
Diagnosis based on the strains CCIBt4016 and ALJ09.
Specimens examined: BRAZIL. SÃO PAULO. Cananéia, Ilha do Cardoso,
(25°03’05”/25°18’18”S-47°53’48”/48°05’42”W), soil sample from an Atlantic rainforest
area, baited with cellulosic substrates (onion skin and corn leaves), 6 Nov. 2012, G.H.
Jerônimo CCIBt 4016. GenBank: SSU: MH590087; ITS: MH590049; LSU: MH590074.

109
BRAZIL. SÃO PAULO. JURÉIA, “Mosaico de Unidades de Conservação Juréia-
Itatins”, (24°30’04,07”S/47°15’58,0”W), from submerged leaves samples in a Atlantic
Rainforest stream, baited with cellulosic substrates (onion skin and corn leaves), 24 Aug
2016, A. L. Jesus ALJ09. GenBank: ITS: MH590050; LSU: MH590075.
Notes: The morphologies of our isolates fit Karlings’s (1944) description of N. elongata,
with no additional or contradictory characteristics. These strains grow indeterminantly on
PmTG, CMA plus glucose or even YPSS/8 media. On agar media, the cultures are
predominantly rhizomycelial, but zoosporangia and resting spores are produced
occasionally. The elongated or cylindrical zoosporangia were commonly observed on
cellulosic baits (onion skin and corn leaves).
FIG 4. Morphology of Karlingiella elongata. A. Zoospores with a single hyaline lipid globule. B.
Zoosporangia and rhizomycelium in PmTG medium. C. Intercalary zoosporangia with elongate tube in
YPSS/8. D-F. Zoospores releasing and operculum (Op) in onion skin. G. Zoosporangium with a

110
parenchymal basal region (Pr) in onion skin. H. Rhizomycelium and non-septate swellings on PmTG. I.
Resting spores with a large lipid globule surrounded by small droplets in PmTG. Op: Operculum. Pr:
Parenchymal region. Bars=10µm.
DISCUSSION
Although Nowakowskiella is composed by numerous species, this genus is still
poorly studied phylogenetically with only Nowakowskiella elegans Nowak. and N.
hemisphaerospora Shanor representing the genus in phylogenetic studies (Mozley-
Standridge et al. 2009). In our combined phylogenetic analyses, Nowakowskiella does
not represent a monophyletic genus due to the placement of N. elongata Karling. In our
combined rDNA phylogeny, this species is sister to the Cladochytriaceae. Based on this
placement, we transferred N. elongata to the new genus Karlingiella as the new
combination Karlingiella elongata. Additionally, we described the new species
Nowakowskiella crenulata, which produces a crenulate resting spore and zoosporangia
supported by a prominent clavate apophysis. Our phylogeny represents the largest
analysis of the genus to date, and provides a phylogenetic backbone for the interspecific
relationships within Nowakowskiella. Branch lengths are long, and support values are
low, leaving a definitive phylogeny unknown until additional cultures can be examined.
As the morphological delineation of the Nowakowskiella species are based mainly
on resting spores development, we decided to maintain the identification of the strains
CCIBt4260 and ALJ27 at the generic level, although our phylogenetic analyses and
zoosporangia morphology have indicated that they could represent new species.
Despite Nowakowskiella ramosa being one of the first described species in this
genus, it has not been commonly reported in biodiversity studies, though it was recorded
twice in Brazil (Karling 1944; Booth 1979). On the other hand, the other Nowakowskiella
representatives isolated during this study (N. multispora and N. hemisphaerospora) have
been commonly reported in taxonomic studies in Brazil, especially in Atlantic rainforest
areas (Nascimento and Pires-Zottarelli 2009; Jesus et al. 2013; Jerônimo et al. 2015). The
records of N. crenulata in sediment samples of two reservoirs with opposite trophic states
(oligotrophic and eutrophic) is an indication that this species could survive in a wide range
of limnological conditions, probably due the production of resting spores.
This study brings important information about interrelationship of the
Nowakowskiella species, including four taxa in molecular phylogenetic reconstructions

111
for the first time, describing a new species, a new genus and a new combination, and
contribute to increase the knowledge about the morphology and phylogenetic diversity.
ACKNOWLEDGMENTS
This study was supported by Fapesp (“Fundação de Amparo a Pesquisa do Estado de São
Paulo) through scholarships awarded to G.H. Jerônimo (N° process 2014/16358-4 and
2016/25800-6 - BEPE), A.L. Jesus (N° process 2016/00697-0) and C.L.A. Pires-
Zottarelli (N° process 2012/50222-7 and 2016/11146-4). We are indebted to CAPES
(“Coordenação de Aperfoiçoamento de Pessoal de Nível Superior”), which supported
G.H. Jerônimo during his master degree and CNPq for the productive fellowship given
to C.L.A. Pires-Zottarelli (304493/2015-5). In addition, we are grateful to the University
of Michigan, Department of Ecology and Evolutionary Biology and all of the Michigan
Mycology lab team, for making me fell at home during the BEPE project period.
Additionally, we thank the thematic Acquased project (FAPESP Process N° 2009/53898-
9) for the productivity fellowship awarded to C.L.A. Pires Zottarelli (Process Nº
304493/2015-5). To conclude, Lucas Michelotti, Denise C. Bicudo, Samantha B.
Faustino, Elaine C.R. Bartozek, Stefano Zorzal de Almeida, Maria A.P.C. da Silva
(Dorinha) are recognized for their contribution collecting samples or laboratory support,
and we thank “Instituto Florestal” and “Grupo Votorantim” for the permission given to
collect samples.

112
LITERATURE CITED
Bazinet AL, Cummings MO. 2008. The lattice project: a grid research and production
environment combining multiple grid computing models. In: Distributed & Grid
Supporting Communities (Weber MHW, ed.) Marbug: Rechenkraft.net, pp 2-13.
Booth T. 1979. Strategies for study of fungi in marine and marine influenced ecosystems.
Revista de Microbiologia 10: 123-138.
Boyle D G, Hyatt AD, Daszak P, Berger L, Longcore J. E, Porter D, Hengstberger SG,
Olsen V. 2013. Cryo-archiving of Batrachochytrium dendrobatidis and other
chytridiomycetes. Diseases of Aquatic Organisms 56:59–64.
Butler EJ. 1907. An account of the genus Pythium ans some Chytridiaceae. Memorians
of the Department of Agriculture India, Botanical Series 1: 1–160.
Hassan, S.K.M. & Batko, A. 1988. Nowakowskiella keratinophila sp. nov. a
keratinophilic fungus from the brackish water. Acta Mycologica Warszawa 22:
193-198.
James TY, Letcher PM, Longcore JE, Mozley-Standridge SE, Porter D, Powell MJ,
Griffith GW, Vilgalys R. 2006. A molecular phylogeny of the flagellated fungi
(Chytridiomycota) and description of a new phylum (Blastocladiomycota).
Mycologia 98:860–871.
Jerônimo GH, Jesus AL, Marano AV, James TY, de Souza JI, Rocha SCO, Pires-
Zottarelli CLA. 2015. Diversidade de Blastocladiomycota e Chytridiomycota do
Parque Estadual da Ilha do Cardoso, Cananéia, Brasil. Hoehnea 42: 135-163
Jesus AL, Marano AV, Schoenlein-Crusius IH, Pires-Zottarelli CLA. Diversidade de
organismos zoospóricos heterotróficos do Parque Estadual das Fontes do Ipiranga,
São Paulo, Brasil: novas citações. Hoehnea 40: 167-180.
Karling JS. 1944. Brazilian chytrids. I. Species of Nowakowskiella. Bulletin of the Torrey
Botanical Club 71: 374–389.
———. 1963. Indian chytrids. IV. Nowakowskiella multispora sp. nov. and other
polycentric species. Sydowia 17: 314–319.
———. 1977. Chytridiomycetarum Iconographia. Vaduz, J. Cramer.

113
Karpov SA, Kobseva AA, Mamkaeva MA, Mamkaeva KA, Mikhailov KV, Mirzaeva
GS, Aleoshin VV. 2014. Gromochytrium mamkaevae gen & sp. nov. and two new
orders: Gromochytriales and Mesochytriales (Chytridiomycetes). Persoonia
32:115–126.
Kazutaka K, Daron M. 2013. MAFFT Multiple Sequence Alignment Software Version
7: Improvements in Performance and Usability. Available from:
http://mafft.cbrc.jp/alignment/server (accessed 27 March 2018).
Letcher PM, Powell MJ. 2005. Kappamyces, a new genus in the Chytridiales
(Chytridiomycota). Nova Hedwigia 80: 115–133.
———, ———. Churchill PF, Chamber JG. 2006. Ultrastructural and molecular
deliniation of a new order, the Rhizophydiales (Chytridiomycota). Mycological
Research 110: 898–915.
———, ———. 2014. Hypothesized evolutionary trends in zoospore ultrastructural
characters in Chytridiales (Chytridiomycota). Mycologia, 106: 379–396.
Longcore JE, Simmons DR. 2012. The Polychytriales ord. nov.contains chitinophilic
members of the rhizophlyctoid alliance. Mycologia 104:276–294.
Lucarotti CJ. Zoospore ultrastructure of Nowakowskiella elegans and Cladochytrium
replicatum (Chytridiales). Canadian Journal of Botany 59:137–148.
Mozley-Standridge SE, Letcher PM, Longcore JE, Porter D, Simmons DR. 2009.
Cladochytriales—a new order in Chytridiomycota. Mycological Research
113:498–507.
Nascimento CA, Pires-Zottarelli CLA. 2009. Chytriales (Chytridiomycota) do Parque
Estadual da Serra da Cantareira, SP, Brasil. Acta Botanica Brasilica 23: 459–473.
Posada D. 2008. jModelTest: phylogenetic model averaging. Molecular Biology and
Evolution 2: 1253–1256.
Shanor L. 1942. A new fungus belonging to the Cladochytriaceae. American Journal of
Botany 29: 174–179.
Simmons DR, James TY, Meyer AF, Longcore JE. 2009. Lobulomycetales, a new order
in the Chytridiomycota. Mycological Research 113:450–460.

114
Sparrow FK. 1942. A classification of aquatic phycomycetes. Mycoloia 34: 113–116.
———. 1943. The aquatic Phycomycetes, exclusive of the Saprolegniaceae and Pythium.
University of Michigan Press. Ann Arbor,
———.1960. Aquatic Phycomycetes. University of Michigan Press, Ann Arbor.
Vélez CG, Letcher PM, Schultz S, Mataloni G, Lefevre E, Powell MJ. 2013. Three new
genera from aquatic habitats in Argentina. Mycologia 105:1251–1265.
———, ———, ———, Powell MJ, Churchill PF. 2011. Molecular phylogenetic and
zoospore ultrastructural analyses of Chytridium olla establish the limits of a
monophyletic Chytridiales. Mycologia 103:118–130.
Vaidya G, Lohman DJ, Meier R. 2010. SequenceMatrix: concatenation software for the
fast assembly of multigene datasets with character set and codon information.
Cladistics 27: 171–180.
Vilgalys R, Hester M. 1990. Rapid genetic identification and mapping of enzymatically
amplified ribosomal DNA from several Cryptococus species. Journal of
Bacteriology 172: 4238-4246.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal
ribosomal RNA genes for phylogenetics. In Part Three: Genetics and Evolution.
Academic Press.

115
CAPíTULO 3
BOOTHIOMYCES ANGULOSUM: A NEW COMBINATION INTO
TERRAMYCETACEAE (RHIZOPHYDIALES, CHYTRIDIOMYCOTA)
Artigo elaborado de acordo com as normas da revista Nova Hedwigia

116
Boothiomyces angulosum: a new combination into Terramycetaceae
(Rhizophydiales, Chytridiomycota)
Authors: Gustavo Henrique Jerônimo1*, Carmen Lidia Amorim Pires-Zottarelli1, David
Rabern Simmons2 and Timothy Yong James2
1 Núcleo de Pesquisa em Micologia, Instituto de Botânica, São Paulo, SP, Brazil
2 Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor,
MI , USA
*corresponding author: [email protected]
With 25 figures and 1 table
Abstract: The genus Rhizophydium was proposed by A. Schenk in 1858 to accommodate
the inoperculate taxa previously inserted in Chytridium. This genus, which has already
included 235 species, have been segregated in different genera based on molecular and
zoospore ultrastructural analyses. However, some taxa have never been investigated
about its phylogenetic position or zoospore ultrastructural characters. The aim of this
study is to discuss, through morphology, zoospore ultrastructure and/or molecular
analyzes, the relation of our isolates of Rhizophydium angulosum and R. elyense in the
Rhizophydiales phylogeny. These isolates produced angular zoosporangia, characteristic
of the Terramycetaceae representatives, and grouped within Boothiomyces clade,
considering the concatenate analyses of complete ITS and partial LSU regions of rDNA.
The transmission electronic microscopy (TEM) analyses revealed that R. angulosum
produces zoospores with the same ultrastructural characters described to Boothiomyces
representatives, while R. elyense presented sufficient characteristics that supports its
morphological delineation from Boothiomyces macroporosum. Based on molecular and
ultrastructural analyses, we are transferring R. angulosum to Boothiomyces genus besides
to debate the morphology of its representatives.
Key words: chytrid, phylogeny, Rhizophydium, ultrastructure

117
Introduction
The genus Rhizophydium was proposed by A. Schenk in 1858 to accommodate the
inoperculate taxa previously inserted in Chytridium (Letcher et al. 2006). The
morphological delineation encompassed around 235 species, which were in the general
characterized by production of eucarpic, monocentric and inoperculate thallus, differing
by the shape and size of zoosporangia, number and position of discharge pores and
relation with substratum or host (Sparrow 1960). However, the circumscription of the
species within the genus was complex because the taxa present a few morphological
distinctions and according to Sparrow (1960), the number of discharge pores, the size and
shape of the zoosporangia and the nature of the substratum, are not always decisive. Thus,
different approaches as zoospores ultrastructure and molecular analyses have been used
to understand the evolutionary relation among the taxa classified in this genus.
Barr (1978, 1980) showed that Rhizophydium species, included in this time in
Chytridiales, produced zoospores with a unique ultrastructural configuration, differing
from other members of Chytridiales. In the following years, with the advance of the
molecular methods allied to the ultrastructural tools, the genus Rhizophydium was
segregated in 11 clades (Letcher et al. 2006, 2008a, 2015) and new genera and species
were proposed (Letcher & Powell 2005; Lepelletier et al. 2014; Letcher et al. 2008a,b,
2012, 2015; Powell et al. 2011), which were grouped in the Rhizophydiales. Currently,
this order comprises 16 families (Letcher et al. 2015), which differ by zoospore
ultrastructural configuration, molecular placement and morphological characteristics.
Among them, Terramycetaceae Letcher includes two genera, Terramyces Letcher and
Boothiomyces Letcher, which produce zoosporangia predominantly angular in shape and
zoospores with a similar ultrastructural configuration. However, subtle ultrastructural
differences were observed by the authors as the type of cisterna and shape of the structure
associated to kinetossome (KAS) (Letcher et al. 2006).
Considering all described species, some of them were not purified neither
investigated regarding the zoospore ultrastructural configuration and molecular
placement. Among these is Rhizophydium angulosum Karling, which is characterized by
production of zoosporangia that occur singly or in aggregates, predominantly angular in
shape but also with subspherical, irregularly clavate, stellate, ovoid, pyriform or citriform
forms. The pores are apical, subapical or lateral, and rhizoidal system is composed by
abundant and finely branched rhizoids (Karling 1968).

118
The aim of this study is to investigate, through morphology, zoospore
ultrastructure as well as molecular analyzes, the position of R. angulosum into the
Rhizophydiales phylogeny. In addition, we discussed the phylogenetic relationship of
Rhizophydium elyense Sparrow.
Materials and methods
Isolation and culture
Sediment surface samples were collected in the Itupararanga reservoir
(23°45’37”-23°35’02”S/47°21’00”-46°57’29”W), located in a protected Atlantic
Rainforest area at São Paulo state, Brazil. These samples were baited with shrimp
exoskeleton, snakeskin and onion skin and posteriorly isolated on nutritive medium
through zoospores inoculation. Thalli developed on nutrient medium (CMA plus glucose
- 16 g corn meal agar, 2.5 g glucose, 1 L deionized water) was purified and posteriorly
deposited in MMBF culture collection (“Micoteca Mário Barreto Figueiredo”) belonging
to WFCC (World Federation for Culture Collections) and CCIBt collection culture
(“Coleção de Cultura de Algas, Fungos e Cianobácterias do Instituto de Botânica”).
Besides that, the strains were cryopreserved according to the protocol of Boyle et al.
(2003).
Morphology
Cultures of the strains CCIBt4261 and GHJ30 were observed growing in CMA
plus glucose and on purified shrimp exoskeleton and snakeskin. Examination of thallus
morphology was performed on Leica DMLB2. We used Sparrow (1960) and Karling
(1977) monographs besides the original descriptions (Sparrow 1957, Karling 1968) to
identify morphologically these isolates.
DNA extraction, amplification and sequencing
For DNA extraction, a small piece of agar with zoosporangia was transferred to
Erlenmeyer`s containing 50 ml of PmTG liquid medium (peptonized milk, tryptone and
glucose) prepared with autoclaved reserve-osmosis water. After incubation for 5-10 days
at 21C, the material was transferred to 2 mL microfuge tubes in order to obtain pellets for
DNA extraction procedure. The extraction of genomic DNA followed the protocol
described in the PureLink Genomic DNA kit (Invitrogen©). Electrophoresis was
performed using 0.8% (p/v) agarose gel and the partial LSU and complete ITS regions of

119
rDNA were amplified using the primers LR0R/LR5 (Vilgalys & Hester 1990) and
ITS4/ITS5 (White et al. 1990), respectively. The thermocycler parameters followed the
cycle described by James et al. (2006). The ribosomal regions were amplified with the
PCR SuperMix kit (Invitrogen©) for a final volume of 25 L and the amplicons were
purified with AxyPrep PCR Clean-up kit (Axygen©), and sequenced at the “Centro de
Pesquisa sobre o Genoma Humano e células tronco, Universidade de São Paulo”. The
contiguous sequences were assembled into SEQUENCHER 5.3.
.
Phylogenetic analyses
For phylogenetic reconstruction, 36 isolates of Rhizophydiales were selected
(Table 1), which represents genera with available sequences in GenBank while
Spizellomyces representatives (SW001 and CBS66873) were used as outgroup taxa. Our
contiguous sequences were assembled using Sequencher version 4.1.4 (Gene Codes)
and the alignment performed online at Mafft version v. 7.058 (Kazutaka & Daron 2013).
The regions ITS and LSU of rDNA were concatenate through SequenceMatrix v. 1.8
(Vaidya et al. 2010) resulting in a final alignment of 1800 base pairs while the phylogeny
was built on PhyML v. 3.1 (Guindon & Gascuel, 2003) adding the model generated by
jModel Test v. 2.2.4 (Posada 2008). The Maximum Likelihood analysis was performed
with 1000 bootstrap and the values below to the 70% were not inserted in the final tree.
Table 1. Isolates, GenBank accession number of ITS and LSU region of rDNA used in the analyses. The
strains highlighted in bold represent the taxa isolated during this study.
Isolates
GenBank acession
number
ITS LSU
Alphamyces chaetifer ARG025 EF585646 EF585606
Alphamyces chaetifer ARG028 EF585649 EF585609
Alphamyces chaetifer ARG058 EF585663 EF585623
Angulomyces argentinensis ARG008 EF585635 EF585595
Angulomyces argentinensis ARG015 EF585640 EF585600
Angulomyces argentinensis ARG070 EF585667 EF585627
Aquamyces chlorogonii ARG018 EF585643 EF585603

120
Isolates
GenBank acession
number
ITS LSU
Aquamyces chlorogonii Barr102 DQ485600 DQ485536
Boothiomyces macroporosum PLAUS021 NR119591
Boothiomyces sp. ARG040 EF585656 EF585616
Globomyces pollinis-pini ARG068 EF585665 EF585625
Globomyces pollinis-pini ARG069 EF585666 EF585626
Gorgonomyces haynaldii ARG024 EF585645 EF585605
Gorgonomyces haynaldii ARG026 EF585647 EF585607
Gorgonomyces haynaldii Barr106 DQ485601 DQ485537
Kappamyces sp. PL120 DQ485672 DQ485581
Kappamyces sp. PL117 EF585670 EF585630
Pateramyces corrientinensis ARG046 EF585657 EF585617
Protrudomyces lateralis ARG047 EF585658 EF585618
Protrudomyces lateralis ARG071 EF585668 EF585628
Protrudomyces lateralis ARG072 EF585669 EF585629
Protrudomyces lateralis Barr004 DQ485597 DQ485533
Rhizophydium carpophilum JEL134 DQ485612 DQ485548
Rhizophydium angulosum CCIBt4261
Rhizophydium elyense GHJ30
Rhizophydium elyense PLAUS9 AY439049
Rhizophydium sphaerotheca BR89 JN943817 JN941000
Rhizophydium sp. PL139 DQ485677 DQ485586
Rhizophydium sp. PL010 DQ485638 DQ485566
Rhizophydium sp. PL073 DQ485660
Rhizophydium sp. PL042 DQ485658
Rhizophydium sp. Barr436 DQ485607 DQ485543
Rhizophydium sp. PLAUS002 DQ485632
Terramyces subangulosum PL001 DQ485652
Terramyces subangulosum PL003 DQ485653 AY439041
Terramyces subangulosum PL122 DQ485673 DQ485582
Urceomyces sphaerocarpus ARG038 EF585655 EF585615
Urceomyces sphaerocarpus ARG048 EF555659 EF555619

121
Isolates
GenBank acession
number
ITS LSU
Spizellomyces punctatus SW001 EU379241 EU379198
Spizellomyces plurigibbosus CBS66873 NR145374 JN941011
Transmission Electronic Microscopy (TEM) analyses
The fixation and transmission electronic microscopy (TEM) were performed
according to the protocol described in Letcher & Powell (2005) and it is summarized with
details at material and methods of this thesis (Pags. 34 to 46). We examined the presence
or absence of a vesicular region, fibrillar bridge (connecting the kinetossome and non-
flagellated centriole) and the type and extension of the cisterna. We also analyzed the
shape of the structure associated to kinetossome (KAS), organization of the microtubules
and relation of the microbody lipid complex (MLC) with the mitochondria.
Results
Our two strains (CCIBt4261 and GHJ30) produced angular zoosporangia with
multiple discharge pores, which are typical of Terramycetaceae representatives.
Morphology Figs 1-16
The strain CCIBt4261 was isolated from sediment surface sample at a depth of 19
meters in a mesotrophic reservoir (Itupararanga) (Figs. 1-11). It grows in chitin (shrimp
exoskeleton) as well as cellulosic (onion skin) substrates and produces a characteristic
aggregated development in baits (Fig. 11). In these substrates, the zoosporangia are
predominantly angular or stellate (Figs. 8-9), while on nutrient agar the clavate,
subspherical and reniform shapes were observed (Figs. 4-11). We are able to note on
nutrient agar, zoosporangia with nearly angular shapes in the early stages of development
(Figs.1-3) besides production of a dilated region at the base (Fig. 6). The zoospores,
usually released by two lateral pores, have a single hyaline lipid globule (Fig. 10).
Considering all these morphological aspects, we identified this strain as Rhizophydium
angulosum Karling.

122
The strain GHJ30 was isolated from sediment sample at the same mesotrophic
reservoir (Itupararanga) using keratinous substrate (snakeskin) (Figs. 12-16). On
snakeskin, it produces sessile and angular (polygonal) zoosporangia at maturity, which
are mostly citriform in shape (Fig. 12 and 13). The rhizoids generally disconnect from
the zoosporangia (sessile) and the zoospores with a single hyaline lipid globule are
released through 2 or 3 pores (Fig. 14). The resting spores are spherical, small, smooth
and produce a thick wall (Fig. 16). On nutrient agar, the zoosporangia are predominantly
spherical, while the rhizoidal system are extensive, delicate and branched (Fig. 15).
Considering all characteristics mentioned above we identified this strain as Rhizophydium
elyense Sparrow.
Figures 1-16. Different development stages of Rhizophydium angulosum and Rhizophydium elyense. 1-11.
Development stages of Rhizophydium angulosum. 1-3. Early development stages in nutrient agar medium.
4. Clavate zoosporangium in nutrient agar medium. 5. Reniform shape in nutrient agar medium. 6. Clavate
zoosporangium with a prominent basal region (br) in nutrient agar medium. 7. Subspherical zoosporangium

123
with prominent papillae in nutrient agar medium. 8. Stellate shape in shrimp exoskeleton. 9. Angular shape
in shrimp exoskeleton. 10. Zoospores releasing on shrimp exoskeleton. 11. Aggregated development on
shrimp exoskeleton. 12-16. Rhizophydium elyense. 12-13. Citriform zoosporangia on snakeskin. 14. Empty
zoosporangia showing pores on snakeskin. 15. Spherical shape in nutrient agar medium. 16. Resting spores
on snakeskin. Scale bars: 10 µm
Phylogenetic analyses Fig 17
The combined alignment from complete ITS and partial LSU regions of rDNA
had 1800 base pairs, from which 994 were parsimony-informative. The substitution
model generated by jModelTest to combined regions was Tamura-Nei plus gamma shape
distribution (TrN+G). The maximum likelihood analyses showed that our isolates
grouped together with Boothiomyces representatives with a high bootstrap value (75%)
(Fig. 17). Besides that, the strains GHJ30 and CCIBt4261 isolated from sediment samples
in the Itupararanga reservoir, are sister taxa within Boothiomyces clade.
Figure 17. Combined topology (complete ITS and partial LSU regions of rDNA) of the maximum
likelihood analysis. Our isolates are highlighted in bold and the numbers next to the branches represents
the boostrap values. Values below to 70% were not inserted in the three.
Transmission electronic microscopy analysis Figs 18-25

124
We were able to observe in R. angulosum (CCIBt4261), all ultrastructural
characters mentioned by Letcher et al. (2006) to Boothiomyces representatives (Figs. 18-
25). These characters include the presence of a fenestrate cisterna (Fc) partially covering
the lipid globule (L) (Figs. 18, 19 and 22), a microbody lipid complex (MLC) associated
to the mitochondria (M) (Fig. 20) and the shape of the structure associated to kinetossome
(KAS) similar to a spur (Sp), which contain a spherical knob in the edge (Fig. 21).
Besides that, we observed the presence of an evident vesicular region (VR) (Figs. 21 and
23) and microtubules, the later organized as a bundle (Mt) (Fig. 24). The presence of a
fibrillar bridge (Fb) were also observed connecting the parallel kinetossome and non-
flagellated centriole (Figs. 24 and 25).
Figures 18-25. Zoospore ultrastructure of Rhizophydium angulosum. 18-19. Transversal section of the lipid
globule and fenestrate cisterna. 20, 21 and 23. Longitudinal section showing the nucleus position (partially
inserted in ribosomal aggregation; mitochondria outside of ribosomal aggregation and associated to

125
microbody lipid globule; structure associated to kinetossome (KAS) like a spur containing a spherical knob
in the edge. 22. Longitudinal section. 24. Longitudinal section showing the kinetossome and non-flagellated
centriole linked by a fibrillary bridge and microtubules organized as a bundle. 25. Transversal section
showing showing the kinetossome and non-flagellated centriole. Fc: fenestrate cisterna. M: mitochondria.
N: nucleus. R: ribossomal aggregation. Sp: Spur. L: lipid globule. MLC: microbody lipid complex. VR:
vesicular region. Mt: microtubules. Fb: fibrillar bridge. NfC: non-flagellated centriole. K: kinetossome.
Scale bar: 200 nm

126
Taxonomy
Boothiomyces angulosum (Karling) Jerônimo & C.L.A. Pires-Zottarelli, comb. nov.
Figs.1-11
Basionym: Rhizophydium angulosum Karling
Type: As Karling (1968) did not designed any material as a type, we followed the
recommendation of Letcher & Powell (2012), which suggested considering the original
illustration (Fig. 1 M-X, Arch. Mikrobiol. 61: 115. 1968) as the lectotype of this species.
Zoosporangia angular and stellate in baits and clavate, subspherical or reniform
on nutrient agar. Zoospores ultrastructural characters as described by Letcher et al. (2006)
to Boothiomyces representatives, which include the presence of a vesicular region,
fenestrate cisterna, microbody lipidic complex (MLC) related to mitochondria, structure
associated to kinetossome (KAS) as a spur containing a spherical knob in the edge and
kinetossome and non-flagellated centriole connected by a fibrillar bridge.
Diagnosis based on the isolate CCIBt4261 obtained from shrimp exoskeleton at sediment
sample of the Itupararanga reservoir (23°36’53,10”S/47°23’34,70”W).
Specimen examined: Brazil, Sao Paulo, Ibiúna municipality, sediment sample from
Itupararanga reservoir, 23°36’53,10”S - 47°23’34,70”W, shrimp exoskeleton.
Discussion
Morphology
Despite Letcher et al. (2006) consider B. macroporosum and R. elyense
morphologically similar and potentially the same species, we were able to find
fundamental differences to support our morphological identification of the strain GHJ30
as R. elyense. In R. elyense GHJ30, we were able to observe the production of sessile
zoosporangia (Fig. 12 and 13) besides spherical resting spores with a granulated content
(Fig. 16), corroboring the original description (Sparrow 1957), which were not cited by
Karling (1968) or Letcher et al. (2006) to B. macroporosum isolates.
Differently from Boothiomyces macroporosum (Karling) Letcher, R. angulosum
CCIBt4261 produced zoosporangia with a dilated region on the base (basal region) (Fig.

127
6). Besides that, this strain produced clavate (on nutrient agar) and stellate (on shrimp
exoskeleton) zoosporangia, which were not cited to B. macroporosum or R. elyense. In
addition, in R. angulosum CCIBt4261, the rhizoidal system is short and branched, while
in B. macroporosum and R. elyense it is extensive.
Although these three taxa produce angular zoosporangia with prominent papillae,
which morphologically characterize the Terramycetaceae representatives, only in B.
macroporosum were observed the deliquescence of the dome-shaped papillae and
formation of a hyaline protuberance in the exit orifice as illustrated by Karling (1968) in
the figures 15, 16 and 17.
Phylogeny
Our concatenate analyses showed that R. angulosum CCIBt4261 belongs to
Boothiomyces genus as well as R. elyense GHJ30, which are sister taxa. Although Letcher
et al. (2006) consider Boothiomyces as a uni-specific genus and admit that B.
macroporosum and R. elyense (PLAUS9) are probably the same species, the inclusion of
one another species (R. angulosum CCIBt4261) within this clade, showed that it is a
complex of species with a few interspecific variation based on ITS and LSU regions.
Probably other regions of DNA besides other specimens are necessary to elucidate the
relationship of the species in this clade. In addition, our sequence of R. elyense (GHJ30)
presented enough phylogenetic distinctions if compared to B. macroporosum PLAUS21
and R. elyense PLAUS9, indicating that this species should also be transferred to
Boothiomyces clade. However, as we did not performed the ultrastructural analyses of R.
elyense (GHJ30), we decided not to transfer it herein.
Zoospore ultrastructure
The genera inserted in Terramycetaceae family, besides to produce angular
zoosporangia, produce zoospores with a similar constellation of ultrastructural characters
differing by the type of cisterna, which are fenestrate in Boothiomyces and simple in
Terramyces. The structure associated to kinetossome (KAS) is a spur in both groups, but
only in Terramyces, Letcher et al. (2006) observed the production of two spherical knobs
in the edge of this structure.
The strain CCIBt4261 presented all ultrastructural characteristics described by
Letcher et al. (2006) to Boothiomyces clade, including the fenestrate cisterna partially

128
covering the lipid globule and structure associated to kinetossome (KAS) as a spur, with
one spherical knob in the edge.
Based on our morphological identification, phylogenetic reconstruction and
zoospore ultrastructural analyses, we transferred R. angulosum to Boothiomyces genus as
the new combination, B. angulosum, and pointed that B. macroporosum and R. elyense
are morphologically distinctive, and the later should be transfer to Boothiomyces.
Acknowledgements
We would like to thank FAPESP (“Fundação de Amparo à Pesquisa do Estado de São
Paulo) and CAPES (“Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for
the PhD fellowships given to G.H. Jerônimo Alves, Fapesp Process N° 2014/16358-4 and
CAPES. We are also grateful to FAPESP to support, through BEPE fellowships (“Bolsa
de Estágio de Pesquisa no Exterior” Process N° 2016/25800-8), the internship of G.H.
Jerônimo Alves at Timothy James lab, in Michigan (USA), to perform the zoospores
ultrastructural analyses. In addition, we are grateful to the University of Michigan,
Department of Ecology and Evolutionary Biology and all Timothy James lab team, for
the making me fell at home during the BEPE project period. Besides that, the thematic
Acquased project (FAPESP Process N° 2009/53898-9) and productivity fellowship given
to C.L.A. Pires Zottarelli (Process Nº 304493/2015-5) by CNPq (“Conselho Nacional de
Desenvolvimento Científico e Tecnológico”) is acknowledged. To conclude, Lucas
Michelotti, L., Denise C. Bicudo, Samantha B. Faustino, Elaine C.R. Bartozek, Stefano
Zorzal de Almeida, Maria A.P.C. da Silva (Dorinha) are recognized for their contribution
collecting samples or laboratory support besides “Instituto Florestal” and “Grupo
Votorantim” for the permission given to collect samples.

129
References
BARR, D. J. S. & HADLAND-HATMANN, V.E. (1978). Zoospore ultrastructure in the
genus Rhizophydium (Chytrdiales). - Canad. J. Bot. 56: 2380-2404.
BARR, D. J. S. (1980). An outline for the reclassification of the Chytridiales, and for a
new order, the Spizellomycetales. - Canad. J. Bot. 58: 2380-2394.
BOYLE, D.G., HYATT, A.D., DASZAK, P., BERGER, L., LONGCORE, J.E.,
PORTER, D., HENGSTBERGER, S.G. & OLSEN, V. (2003). Cryo-archiving of
Batrochochytrium dendrobatidis and other Chytridiomycetes. - Dis. Aqua. Organ. 56: 59-
64.
GUINDON S. & GASCUEL, O. (2003). PhyML: A simple, fast and accurate algorithm
to estimate large phylogenies by maximum likelihood. - Syst. Biol. 52: 696-704.
JAMES, T.Y., LETCHER, P.M., LONGCORE, J.E., MOZLEY-STANDRIDGE, S.E. ET
AL. (2006). A molecular phylogeny of the flagellated fungi (Chytridiomycota)
description of a new phylum (Blastocladiomycota). Mycologia 98: 860-871.
KARLING, J. S. (1968). Zoosporic fungi of Oceania. - Arch. Mikrobiol. 61: 112-127.
KARLING, J. S. (1977). Chytridiomycetarum Iconographia. - J. Cramer, Vaduz
KAZUTAKA, K. & DARON, M. (2013). MAFFT Multiple Sequence Alignment
Software Version 7: Improvements in Performance and Usability. Available from:
http://mafft.cbrc.jp/alignment/server (accessed 27 April 2018).
LEPELLETIER, F., KARPOV, S.A., ALACID, E., LE PANSE, S., BIGEARD, E.,
GARCÉS, E., JEANTHON, C. & GUILLOU, L. (2014). Dinomyces arenyensis gen. et.
sp. nov. (Rhizophydiales, Dinomycetaceae fam. nov.) a chytrid infecting marine
dinoflagellates. - Protist 165: 230-244.
LETCHER, P. & POWELL, M.J. (2005). Kappamyces, a new genus in the Chytridiales
(Chytridiomycota). Nova Hedwigia 80: 115-133.
LETCHER, P., POWELL, M.J., CHURCHILL, P.F. & CHAMBER, J.G. (2006).
Ultrastructural and molecular deliniation of a new order, the Rhizophydiales
(Chytridiomycota). – Mycol. Res. 110: 898-915.

130
LETCHER, P.M. VELEZ, C.G., BARRANTES, M.E., POWELL, M.J., CHURCHILL,
P.A. & WAKEFIELD, W.S. (2008a). Ultrastructural and molecular analyses of
Rhizophydiales (Chytridiomycota) isolates from North America and Argentina. – Mycol.
Res. 112: 759-782.
LETCHER, P., POWELL, M.J. & VIUSENT, M.C. (2008b). Rediscovering of an unsual
chytridiaceous fungus new to the order Rhizophydiales. - Mycologia 100: 325-334.
LETCHER & POWELL, M. (2012). A taxonomic sumary and revision of Rhizophydium
(Rhizophydiales, Chytridiomycota). University Printing, University of Alabama,
Tuscaloosa, 222 p.
LETCHER, P., POWELL, M.J. & DAVIS, W.J. (2015). A new family and four new
genera in Rhizophydiales (Chytridiomycota). - Mycologia 107: 808-830.
POSADA, D. (2008). jModelTest: phylogenetic model averaging. - Mol. Biol. Evol. 2:
1253–1256.
POWELL, M.J. LETCHER, P.M. & LONGCORE, J.E. (2011). Operculomyces is a new
genus in the order Rhizophydiales. - Mycologia 102: 854-862.
SPARROW, F.K. (1957). A further contribution to the phycomycete flora of Great
Britain. – Trans. Br. Mycol. Soc. 40: 523-535.
SPARROW, F.K. (1960). Aquatic Phycomycetes. 2nd revised ed. - University of
Michigan Press, Ann Arbor.
VAIDYA, G., LOHMAN, D.J. & MEIER, R. (2010). SequenceMatrix: concatenation
software for the fast assembly of multigene datasets with character set and codon
information. - Cladistics 27: 171–180.
VILGALYS, R. & HESTER, M. (1990). Rapid genetic identification and mapping of
enzymatically amplified ribosomal DNA from several Cryptococus species. – J.
Bacteriol. 172: 4238-4246.
WHITE, T. J., BRUNS, T., LEE, S. & TAYLOR, J. (1990). Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In Part Three: Genetics
and Evolution. Academic Press (ed.). pp. 315-322.

131
CONSIDERAÇÕES FINAIS
O estudo desenvolvido evidência a importância de estudos taxonômicos e
ecológicos em ecossistemas pouco explorados, como é o caso dos reservatórios. Em
especial, o estudo do sedimento superficial, compartimento até então ignorado com
relação à diversidade destes organismos no Brasil, apresentou resultados muito
significativos. Neste compartimento, foi descrita uma nova espécie de Nowakowskiella
(Nowakowskiella crenulata), bem como observada pela primeira vez para no Brasil a
espécie Plectospira gemmifera. Além disso, maior parte da riqueza de espécies dos filos
Blastocladiomycota e Chytridiomycota foi observada neste compartimento. Como
durante as amostragens foi observada maior frequência, abundância e riqueza de táxons
de fungos zoospóricos no sedimento superficial e dos oomicetos na coluna d’água,
podemos rejeitar a hipótese de que estes dois grupos ocorreriam preferencialmente na
coluna d’água. Os poucos relatos de fungos zoospóricos em amostras de sedimento
podem estar mais relacionados às limitações de amostragem, do que necessariamente aos
padrões ecológicos destes fungos.
Os organismos zoospóricos apresentaram uma ampla distribuição ao longo de
diferentes gradientes tróficos, assim como mencionado por Rocha (2004), estando
algumas espécies relacionadas aos locais com as mais altas concentrações de nitrogênio
e fósforo total, como Catenophlyctis variabilis, enquanto outras à locais com as mais
baixas concentrações destes nutrientes (Diplophlyctis sarcoptoides e Cladochytrium
replicatum), e que poderiam potencialmente ser táxons indicativos da qualidade das
águas. Concluímos, portanto, que os organismos zoospóricos heterotróficos são, no geral,
tolerantes às amplas variações nas concentrações de nitrogênio e fósforo, mas alguns
táxons são limitados pela disponibilidade destes nutrientes, particularmente o fósforo.
Com isso, consideramos que os ecossistemas lênticos representam ambientes ricos
em diversidade de organismos zoospóricos e que merecem maior atenção, em especial o
estudo de suas camadas sedimentares e as relações de ocorrência, frequência e abundância
destes organismos frente aos padrões ambientais, os quais ainda são enormemente
negligenciados em termos mundiais.

132
Anexo I
Zoosporângios de Siphonaria sp.1 (A-B), Diplophlyctis sarcoptoides (C) e Avachytrium sp.1 (D) em meio
de cultura PmTG. Barras = 10 µm.

133
Análise concatenada de Máxima Verossimilhança das regiões SSU e LSU do rDNA. Os isolados obtidos durante este estudo estão destacados em negrito.
Os números próximos aos ramos representam os valores de “bootstratp”, para os quais foram incluídos na topologia somente os valores acima de 75%.

134
ANEXO II– Jerônimo et al.
2017
Phytotaxa 307 (3): 191–198
http://www.mapress.com/j/pt/
Copyright © 2017 Magnolia
Press
Article
https://doi.org/10.11646/phytotaxa.307.3.3
New insights into Plectospira genus (Oomycetes, Straminipila): morphological and molecular analyses
GUSTAVO HENRIQUE JERÔNIMO1,2, ANA L. JESUS1, SARAH C.O. ROCHA1, DANILO R. GONÇALVES &
CARMEN L.A. PIRES-ZOTTARELLI1
1 Instituto de Botânica, Núcleo de Pesquisa em Micologia, 3687 Miguel Stéfano Ave., 04301-902, São Paulo, SP, Brazil
2 E-mail: [email protected]
Abstract
We analyzed the morphology and phylogenetic placement of three isolates of Plectospira belonging to Plectospira gem-
mifera and P. myriandra that were isolated from soil, water bodies and reservoir sediment. The molecular data are from the
partial large subunit and the complete internal transcribed regions of the ribosomal DNA. The genus is characterized by the
presence of lobulated zoosporangia and mode of zoospore discharge forming an evanescent cluster in the exit orifice. Our
isolates clustered together with the single isolate of the genus available in GenBank, however, our phylogenetic analysis and
morphological characterization showed that the isolate identified as Plectospira myriandra CBS 523.87 is possibly another
taxon. In addition, P. gemmifera is recorded for the first time in Brazil and its sequences were inserted for the first time in a
phylogenetic reconstruction.
Keywords: morphology, phylogeny, Plectospira
Introduction
The genus Plectospira was proposed by Drechsler (1927) to include Plectospira myriandra, a phytopathogenic species
originally isolated from sugarcane roots in USA. Initially placed in Saprolegniaceae due its morphological characteristics, the
genus was transferred to Leptolegniaceae by Dick et al. (1999) and more recently to Verrucalvaceae by Beakes et al. (2014)
considering its phylogenetic position together with the genera Aphanomyces, Pachymetra and Verrucalvus. The genus present
mycelial growth and a type of zoosporangium composed typically of a complex of inflated elements together with well-
developed evacuation hyphae (Drechsler 1927).
Currently, according to MycoBank, four phytopathogenic species of Plectospira are described and considered legitimate
(Plectospira myriandra Drechsler, P. gemmifera Drechsler, P. agama Drechsler and P. plagiocaula Drechsler) since Plectospira
dubia Atkins was transferred to Atkinsiella by Vishiniac (1958) considering the morphological characteristics.
The species of Plectospira are responsible for causing diseases in crops of economic importance such as tomato (Solanum
lycopersicum L.) (Drechsler 1927) and sugarcane (Saccharum officinarum L.), and also in higher-plants such as Prunella
vulgaris L. and Rumex acetosella L. (Watanabe 1987), although its infectious mechanism remains poorly investigated.
Considering the importance of Plectospira species as parasite of different crops, is surprising as mentioned by Johnson et al.
(2002), that only few studies have reported the occurrence of the genus worldwide. After their original descriptions, only P.
myriandra were reported again in literature, firstly in Japan (Watanabe 1987) and then in Brazil (Gomes et al. 2003, Rocha
2006, Miranda & Pires-Zottarelli 2012, Jesus et al. 2013), but always as saprotrophs in vegetal debris. On the other hand,
there is no record of P. gemmifera and the other species in literature after their publication. According to Johnson et al. (2002),
the few reports can be related to a frequent misidentification of the species as belonging to Aphanomyces, another genus from
Verrucalvaceae that presents morphological similarities and is composed by species considered parasites of plants (Scott 1961,
Gaulin et al. 2007).
The aim of this study was to characterize morphologically the isolates of P. myriandra and P. gemmifera and include both
species in a phylogenetic reconstruction.
ISSN 1179-3155 (print edition)
PHYTOTAXA

135
Material & Methods
Origin of isolates
We used three Brazilian isolates of Plectospira in our analysis, which were deposited in CCIBtculturecollection (“Coleção de
Culturas de Algas, Cianobactérias e Fungos do Instituto de Botânica”, São Paulo, SP, Brazil) and MMBF culture collection
(“Micoteca Mário Barreto Figueiredo”) belonging to WFCC (World Federation for Culture Collections). The isolates CCIBt
3992 and CCIBt 3372 of Plectospira myriandra was isolated from soil (23°03’05’’–25°18’18’’S and 47°53’48’’–
48°05’42’’W) and freshwater samples (23º38’08”–23º40’18”S and 46º36’48”–46º38’00”W) respectively, of the Atlantic
Rainforest areas, São Paulo State, Brazil. The isolate of Plectospira gemmifera (CCIBt 4250, MMBF 04/16) was isolated
from sediment sample of Itupararanga reservoir (23°37’11.58”S and 47°13’59.34”W).
Morphological studies
Sexual and asexual structures of the isolates were characterized and measured once the pure cultures were obtained.
Identification was made according to the main literatures of the genus including Drechsler (1927, 1929) and Johnson et al.
(2002). Due to the similarity of the species of the genus, a comparative table has been prepared to help in the morphological
identification of these two species. The morphological description of P. myriandra was made considering the isolates CCIBt
3992 and CCIBt 3372, the latter described in Jesus et al. (2013).
DNA extraction, PCR amplification and sequencing
For DNA extraction, the isolates were cultivated initially onto solid medium with 0.20 gL-1 of each streptomycin sulphate
and penicillin G, as antibiotics. A small piece of agar with mycelium from the pure culture was transferred to Erlenmeyer’s
containing 50 mL of MP5 liquid medium (maltose-peptone) prepared with autoclaved reverse-osmosis water. After incubation
for 5–10 days at 21°C, the mycelium was transferred to 2.0 mL microfuge tubes in order to obtain mycelial pellets with
enough biomass for DNA extraction. DNA genomic extraction followed the protocol described in the “PureLink Genomic
DNA Kit” (InvitrogenTM). Electrophoresis was performed using 1% (p/v) agarose gel. The partial LSU and complete ITS
regions were amplified using the primers LR0R/LR6-O (Riethmüller et al. 2002) and UN-up 18S42/UN-up 28S22 (Robideau
et al. 2011) respectively. DNA was amplified with the PCR SuperMix kit (Invitrogen®) for a final volume of 25 µl in a C1000
Touch™ Thermal Cycler Bio-Rad. The PCR amplification technique was performed following the conditions described by
Marano et al. (2014). Amplicons were purified with AxyPrep PCR Clean-up kit (Axygen®).
Phylogenetic analysis
For phylogenetic reconstruction were selected 23 isolates of Verrucalvaceae, which represents the genera with sequences
available in GenBank (Table 1). The isolates Achlya primoachlya (Coker & Couch) T.W. Johnson & R.L. Seymour (CCIBt
3982) and Achlya caroliniana Coker (CCIBt 3870) were used as the outgroup taxa. Initially, the contiguous sequences were
assembled using Sequencher™ version 4.1.4 (Gene Codes®) and the ambiguous bases were manually edited. The alignment
was performed in MAFFT version 7 (Kazutaka & Daron 2013) following standart default. JModelTest 0.1.1 (Posada 2008),
with Akaike Information Criterion, was used to determine the best model of base substitution. For sequences of ITS and LSU
of rDNA regions the Transversion model (TVM+G) plus gamma distribution (G) was indicated as the best substitution model.
The complete ITS (763 bp) and partial LSU regions (1261 bp) were concatenated in SequenceMatrix 1.8 (Vaidya et al. 2010)
resulting in a fragment of 2024 bp in length. The Maximum likelihood (ML) analysis were performed in Garli 2.01 (Bazinet
& Cummings 2008) using partition models and branch swapping by best TBR, while the support of nodes, was determined
with bootstrap analysis with 1000 replications. Tree was rooted with members of the Achlya Nees.
TABLE 1. Taxa, voucher, GenBank accession number of the isolates used in phylogenetic reconstruction. NA: not available.
* Isolates from this study.
Taxa Voucher GenBank accession number
ITS LSU
Aphanomyces repetans CBS 1268.86 NA HQ395662
Aphanomyces repetans CBS 1268.87 NA HQ395671
Aphanomyces laevis CBS 478.71 HQ643122 HQ665242
Aphanomyces laevis CCIBt 4070 KP006463 KP006453

136
Taxa Voucher GenBank accession number
ITS LSU
Aphanomyces sinensis ATCC 4825 JQ070116 JQ070142
Aphanomyces stellatus CCIBt 3994 KP006462 KP006454
Aphanomyces stellatus AR 51 NA AF119587
Aphanomyces salsuginosus ATCC 4775 JQ070107 JQ070132
Aphanomyces astaci CBS 1215.37 NA JX115216
Aphanomyces iridis SAP 356 FM999227 NA
Aphanomyces iridis CBS 524.87 HQ643121 HQ665248
Aphanomyces euteiches CBS 154.73 NA HQ665129
Aphanomyces euteiches 315 KM486065 NA
Aphanomyces euteiches 1309 KM486066 NA
Aphanomyces euteiches CBS 156.73 NA HQ665132
Aphanomyces cochlioides CBS 477.71 HQ643115 HQ665241
Pachymetra chaunorhiza CBS 960.87 NA AF119598
Plectospira myriandra* CCIBt 3992 KR063220 KR063219
Plectospira myriandra* CCIBt 3372 KT935288 KX353796
Plectospira myriandra CBS 523.87 HQ643402 HQ665247
Plectospira gemmifera* CCIBt 4250 KX084705 KX084707
Achlya primoachlya CCIBt 3982 KM058757 KM058754
Achlya caroliniana CCIBt 3870 KP006458 KP006451
Results
Plectospira gemmifera and P. myriandra had common characteristics of the genus such as the presence of lobulate
zoosporangia and mode of zoospore discharge forming an evanescent cluster in exit orifice (Fig. 1–2). Although these species
are morphologically similar, some structures can be used to help in distinguishing them (Table 2). Plectospira myriandra
presents an evident pellucid spot and scattered pits in the internal wall of the oospores (Fig. 2) while P. gemmifera produces
abundant gemmae in the whole life cycle (Fig. 1). Besides that, our isolates presented proportion of antheridia production
different that was described by Drechsler (1927, 1929). We observed that our isolates of P. myriandra frequently present
abundant antheridia while in P. gemmifera they are rare or not frequent. Therefore, we advise that antheridia proportion should
be carefully observed in future taxonomic studies. This is the second record of P. gemmifera in the world and the first record
in Brazil.
Morphological analysis
Plectospira gemmifera Drechsler (1929: 358)
Fig 1. A–M
Mycelium dense; hyphae slender, sparingly branched. Inflated elements of sporangia abundant, sometimes very extensive,
which produces a long tube to release the zoospores. Zoospores forming an evanescent cluster in exit orifice and encysting with
usually 10–12 µm diam. Gemmae abundant, spherical, 20–35 µm diam. and frequently pyriform or obpyriform, 40–35 × 25–
28 µm diam. Oogonia mostly terminal on short branches or on longer hyphae, smooth, subsphaerical, 18–22 × 19–24 µm
diam., spherical, 20–30 µm diam. provided with wall 2.5–3.0 µm in thickness. Antheridia absent or when present, diclinous.
Oospore single, colorless, subcentric, spherical, 15–20 µm diam. and subsphaerical, 18–20 × 20–22 µm diam.
Specimen examined—BRAZIL. São Paulo: Votorantim, Itupararanga reservoir, from sample of surface sediment located 2
meters under the water column, 29 august 2012, CCIBt 4250 and MMBF 04/16.
Notes: The isolate CCIBt 4250 produces abundant gemmae of different formats when cultivated in culture medium (CMA +
glucose) and in water cultures growing in Sorghum sp. seeds. In contrast with the original description of Drechsler (1929),
that observed antheridia always present, in the isolate studied the antheridia production were rare or not frequent and most
oogonia are abortive and/or parthenogenetic. Despite this, the abundant production of gemmae and the characteristics of
oogonium, oospores and zoosporangium are in agreement with Drechsler (1929) and confirm our identification as Plectospira
gemmifera.

137
TABLE 2. Comparison of morphological characteristics between isolates of Plectospira myriandra and P. gemmifera collected
during this study.
Structures Taxa (number of the culture collection)
Plectospira myriandra (CCIBt 3772/CCIBt 3992) Plectospira gemmifera (CCIBt 4250)
Antheridia diclinous, monoclinous, frequent diclinous, rare
Oogonium scattered pits in the wall unpitted wall
Oospores presence of pellucid spot absence of pellucid spot
Gemmae absent present
Notes: Johnson et al. (2002) mentioned that P. gemmifera presents zoosporangia larger and with more lobules
than P. myriandra, however, we did not include this characteristic in the table above once it varies depending
on the bait used and thus it is difficult to observe and compare.
FIGURE 1. A–M. Plectospira gemmifera CCIBt 4250. A–D. Different formats of gemmae. E. Lobulated zoosporangia. F–G. Zoosporangium with
a long tube and zoospores cluster. H. Catenulate gemmae. I. Spherical gemmae with germinative tube. J. Oogonium and antheridia. K. Subcentric
oospores. L. Empty gemmae. M. Piriform gemmae germinating. Bars: 10 µm.

138
FIGURE 2. A–J. Plectospira myriandra. A–E. Plectospira myriandra CCIBt 3992. A. Lobulated zoosporangium. B. Oogonium and subcentric
oospore. C. Oogonium showing pellucid spot (ps). D. Oogonium showing scattered pits in the wall (pw). E. Oogonium and abundant antheridia. F–J.
Plectospira myriandra CCIBt 3372. F. Lobulated zoosporagium. G. Oogonium and antheridia. H. Oogonium and pellucid spot (ps) showed in indirect
focus (arrow). I. Oogonium and subcentric oospore. J. Oogonia with pellucid spot indicated by the arrow. ps: pellucid spot. pw: scattered pits in the
wall. Bars: 10 µm.
Plectospira myriandra Drechsler (1927: 294)
Fig 2. A–J
Mycelium dense; hyphae slender, sparingly branched. Inflated elements of sporangia abundant, which produces a
long tube to release the zoospores. Zoospores forming an evanescent cluster in exit orifice and encysting with
usually 8–11 µm diam. Gemmae absent. Oogonia mostly laterally, rarely terminal on short branches, smooth,
spherical, 20–30 µm diam., provided with wall of 1.0–2.0 µm in thickness. Antheridia always present, diclinous,
rarely monoclinous, ranging from 2 to 15 or more. Oospore single, yellow, subcentric, spherical, 13.7–22 µm in
diam.

139
Specimen examined:—BRAZIL. São Paulo: Cananéia, “Parque Estadual da Ilha do Cardoso (PEIC)”, from soil
sample, 20 August 2012, CCIBt 3992.
Notes:—The isolates CCIBt 3992 and CCIBt 3372 (Jesus et al. 2013) of Plectospira myriandra presented all
characteristics mentioned in its original descriptions (Drechsler 1927), and are in agreement with other three studies
previously published in Brazil (Rocha 2006, Gomes et al. 2003, Miranda & Pires-Zottarelli 2012). In addition, the
features mentioned above, which are shown in Table 2 and Figure 2, were crucial for a correct identification of this
species.
Phylogenetic placement
Our concatenate analysis showed two major clades (Fig. 3) which presented the same configuration of the
functional groups mentioned by Diéguez-Uribeondo et al. (2009). The clade I is composed by saprophytic and/or
opportunistic taxa (subclade I-a) and by species considered parasites of animals (subclade I-b). The clade II is
composed by parasites of plants, and is divided in two subclades (II-a and II-b) strongly supported (100% and
100%, respectively). The subclade II-b is composed by four isolates of Plectospira genus (P. myriandra CCIBt
3372, CCIBt 3992, CBS 523.87 and P. gemmifera CCIBt 4250) and can be considered as sister group of the
Aphanomyces and Pachymetra species parasites of plants (clade II-a). Besides that, our phylogenetic reconstruction
indicated with high bootstrap value (100%) that the isolate of P. myriandra CBS 523.87 possibly do not belong to
this taxon.
FIGURE 3. Concatenate analysis of complete ITS and partial LSU rDNA regions of Verrucalvaceae family inferred
by Maximum Likelihood method. The numbers next the branches indicate the bootstrap values (%) and the scale bar
the numbers of substitution per site. In bold the three Brazilian isolates of Plectospira and in grey the whole clade.
Discussion
Our morphological and phylogenetic analyses added important contributions to a poorly investigated genus inside
the Verrucalvaceae sensu Beakes et al. (2014). Both species (P. gemmifera and P. myriandra) analyzed in this study
showed morphological features that contributed to their identification. In addition, their position in the
phylogenetic analysis of two DNA combined regions (complete ITS and partial LSU of rDNA) corroborated our
morphological characterization.
The Plectospira species formed a well-supported monophyletic subclade (100%) positioned as a sister group of
subclade II-a, composed by parasites of plants (Riethmüller et al. 2002, Diéguez-Uribeondo et al. 2009). Although
the phylogenetic analysis showed a closely relation between the isolates from this study with plant pathogenic

140
species (subclade II-a), it is interesting to mention that our isolates were isolated as saprophytes from soil,
freshwater and surface sediment of reservoirs, which shows the great adaptability of these species in different
environments.
Inside the genus Plectospira, the three isolates from this study clustered with the single isolate of the genus
identified as P. myriandra (CBS 523.87) available in GenBank, although our two isolates of P. myriandra are not
very closely related with the specimen from GenBank. Considering the position of P. myriandra from GenBank
together with P. gemmifera with high support value (100%), as well as the presence in our P. myriandra isolates
of singular characteristics mentioned in Table 2, we suggest that P. myriandra (CBS 523.87) described by
Watanabe (1987) is possibly another species of Plectospira.
This study brings important information about the occurrence and provides new molecular data of a poorly studied
genus. Although both species herein related have been previously described as parasites of plants, we found them
acting as saprophytes in soil, freshwater habitats and sediment reservoir. The ability of both species as facultative
parasites shows the great flexibility of these taxa to different environmental pressures.
Acknowledgments
We would like to thank FAPESP (“Fundação de Amparo à Pesquisa do Estado de São Paulo), for the fellowships
given to G.H. Jerônimo (Process N° 2014/16358-4) and for financial support given to C.L.A. Pires Zottarelli
(Process N° 2012/50222-7) and to thematic project Acquased (Process N° 2009/53898-9). Also CNPq (“Conselho
Nacional de Desenvolvimento Científico e Tecnológico”) is acknowledged for the fellowship to S.C.O. Rocha
(Process N° 117238/2012-0) and C.L.A. Pires Zottarelli (Process N° 304493/2015-5). In addition, D.C. Bicudo,
S.B. Faustino, E.C.R. Bartozek, S. Zorzal de Almeida, M.A.P.C. da Silva (Dorinha) are recognized for their
contribution in collecting samples and in laboratory analysis. To conclude I would like to thank “Instituto Florestal”
and “Grupo Votorantim” for the permission given to collect samples.
References
Bazinet, A.L., & Cummings, M.P. (2008) The Lattice Project: a grid research and production environment
combining multiple grid computing models. In: Weber, M.H.W. (Ed.) Distributed & Grid Computing - Science
Made Transparent for Everyone. Principles, Applications and Supporting Communities, pp. 2–13.
Beakes, G.W., Honda, D. & Thines, M. (2014) Systematics of the Straminipila: Labyrinthulomycota,
Hyphochytriomycota, and Oomycota. In: McLaughlin DJ, Spatafora JW (Eds.) The Mycota VIII Part A. Springer-
Verlag, Berlin, pp. 39–97. http://doi.org/10.1007/978-3-642-55318-93
Dick, M.W., Vick, M.C., Gibbings, J.G., Hedderson, T.A. & Lopez-Lastra, C.C. (1999) 18S rDNA for species of
Leptolegnia and other Peronosporomycetes: justification for the subclass taxa Saprolegniomycetidae and
Peronosporomycetidae and division of the Saprolegniaceae sensu lato into the families Leptolegniaceae and
Saprolegniaceae. Mycological Research 103: 1119–1125. http://doi.org/10.1017/S0953756299008643
Dick, M.W. (2001) Straminipilous fungi. Kluwer Academic Publishers, Dordrecht, 670 pp.
https://doi.org/10.1007/978-94-015-9733-3.
Diéguez-Uribeondo, J., García, M. A., Cerenius, L., Kozubíková, E., Ballesteros, I., Windels, C., Weiland, J.,
Kator, H., Söderhäll, K. & Martín, M.P. (2009) Phylogenetic relationships among plant and animal parasites, and
saprotrophs in Aphanomyces (Oomycetes). Fungal Genetics and Biology 46: 365–376.
http://doi.org/10.1016/j.fgb.2009.02.004
Drechsler, C. (1927) Two water molds causing tomato rootlet injury. Journal of Agricultural Research 34: 287–296.
Drechsler, C. (1929) The beet water mold and several related root parasites. Journal of Agricultural Research 38:
309–361.

141
Gaulin, E., Jacquet, C., Bottin, A. & Dumas, A. (2007) Root rot disease of legumes caused by Aphanomyves
euteiches. Molecular Plant Pathology 8: 538–548. http://doi.org/10.1111/j.1364-3703.2007.00413.x
Gomes, A.L., Pires-Zottarelli, C.L.A., Rocha, M., & Milanez, A.I. (2003) Saprolegniaceae de áreas de cerrado do
estado de São Paulo, Brasil. Hoehnea 30: 95–110.
Jesus, A.L., Marano, A.V., Schoenlein-Crusius, I.H. & Pires-Zottarelli, C.L.A. (2013) Diversidade de organismos
zoospóricos heterotróficos do Parque Estadual das Fontes do Ipiranga, São Paulo, SP, Brasil: novas citações.
Hoehnea 40: 167–180. http://doi.org/10.1590/S2236-89062013000100009
Johnson, T.W. Jr., Seymour, R.L. & Padgett, D.E. (2002) Biology and systematics of Saprolegniaceae. Available
from: http://www.ucnw. edu/people/padgett/book (accessed 30 May 2016)
Kazutaka, K. & Daron, M. (2013) MAFFT Multiple Sequence Alignment Software Version 7: Improvements in
Performance and Usability. Available from: http://mafft.cbrc.jp/alignment/server (accessed 27 February 2016)
https://doi.org/10.1093/molbev/mst010
Marano, A.V., Jesus, A.L., de Souza, J.I., Leãno, E.M., James, T.Y., Jerônimo, G.H., de Cock, A.W.A.W. & Pires-
Zottarelli, C.L.A. (2014) A new combination in Phytopythium: P. kandeliae (Oomycetes, Straminipila).
Mycosphere 5: 510–522. http://dx.doi.org/10.5943/mycosphere/5/4/3
Miranda, M.L. & Pires-Zottarelli, C.L.A. (2012) Oomicetos do Parque Estadual da Serra da Cantareira, São Paulo,
SP, Brasil. Hoehnea 39: 95–112. http://doi.org/10.1590/S2236-89062012000100005
Posada, D. (2008) jModelTest: phylogenetic model averaging. Molecular Biology and Evolution 2: 1253–1256.
https://doi.org/10.1093/molbev/msn083
Riethmüller, A., Voglmayr, H., Göker, M., Weiß, M. & Oberwinkler, F. (2002) Phylogenetic relationships of the
downy mildews (Peronosporales) and related groups based on nuclear large subunit ribosomal DNA sequences.
Mycologia 94: 834–849. http://doi.org/10.2307/3761698
Robideau, G.P., de Cock, A.W.A.M., Coffey, M.D., Voglmayr, H., Brouwer, H., Bala, K., Chitty, D.W., Désaulniers,
N., Eggertson, Q.A., Gachon, C.M.M., Hu, C.H., Küpper, F.C., Rintoul, T.L., Sarhan, E., Verstappen, E.C.P.,
Zhang, Y., Bonants, P.J.M., Ristaino, J.B. & Lévesque, A.C. (2011) DNA barcoding of oomycetes with cytochrome
c oxidase subunit I and internal transcribed spacer. Molecular and Ecological Resources 11: 1002–1011.
Rocha, J.R.S. (2006) Filos Chytridiomycota e Oomycota. In: Gusmão, L.F.P & Maia, L.C. (Eds.) Diversidade e
caracterização dos fungos do semi-árido brasileiro. pp. 75–95.
Vaidya, G., Lohman, D.J. & Meier, R. (2010) SequenceMatrix: concatenation software for the fast assembly of
multigene datasets with character set and codon information. Cladistics 27: 171–180.
Visniac, H.S. (1958) A new marine phycomycete. Mycologia 50: 66-79. http://doi.org/10.2307/3756037
Watanabe, T. (1987) Plectopira myriandra, a rediscovered water mold in Japanese soil. Mycologia 79: 77–81.

142
Anexo III
Saprolegnia aenigmatica CCIBt4347 (A) e Saprolegnia subterranea CCIBt4346 (B-E). A. Zoosporângio
de Saprolegnia aenigmatica com proliferação interna. B. Zoosporângio de Saprolegnia subterranea com
proliferação interna. C-E. Detalhes do anterídio, oogônio, oósporo e tudo germinativo de Saprolegnia
subterranea. Barras = 10 µm.

143
Análise concatenada de Máxima Verossimilhança da região ITS do rDNA. Os isolados obtidos durante este estudo estão destacados em negrito. Os números próximos aos
ramos representam os valores de “bootstratp”, para os quais foram incluídos na topologia somente os valores acima de 75%.

144
Achlya caroliniana (A-E), Achlya klebsiana (F-H), Dictyuchus pseudodictyon (H-L). A-B. Zoosporângios
de Achlya caroliniana mostrando liberação aclióide. C-E. Oogônios com oósporos. F. Zoosporângio de
Achlya klebsiana mostrando liberação aclióide. G. Oogônio e oósporos de Achlya klebsiana. H.
Zoosporângio de Dictyuchus pseudodictyon. I-L. Oogônio e oósporos de Dictyuchus pseudodictyon. Barras
= 10 µm.
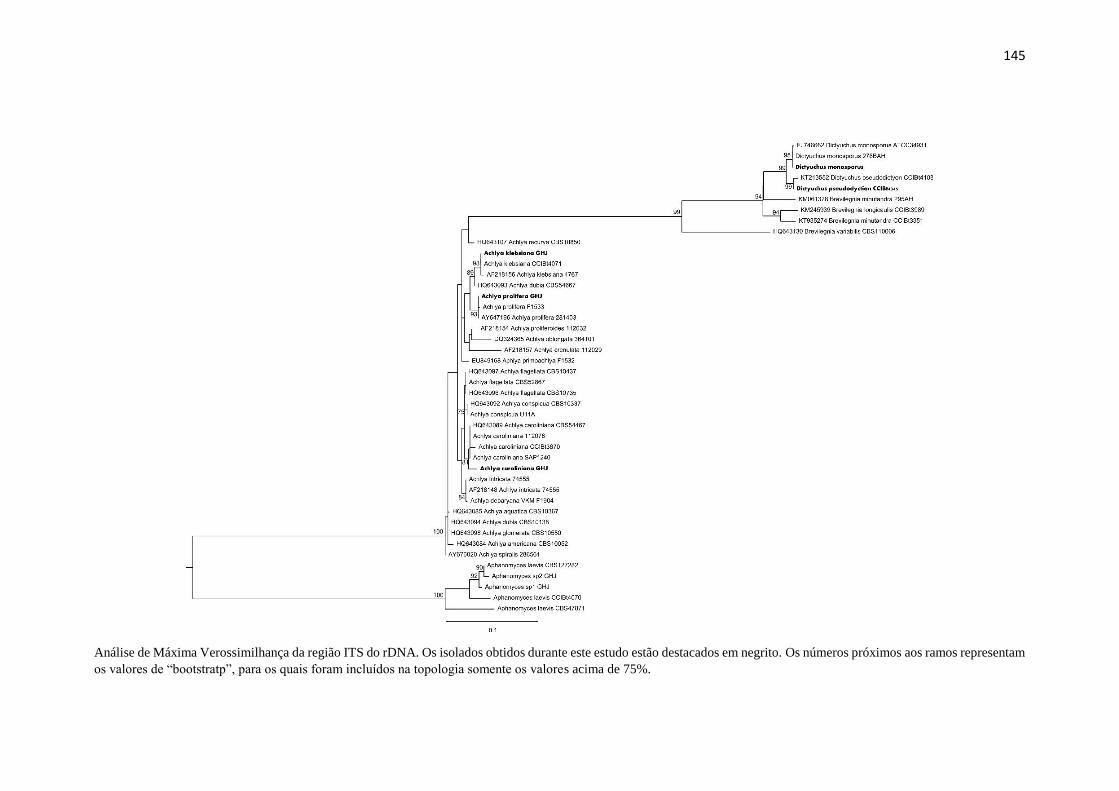
145
Análise de Máxima Verossimilhança da região ITS do rDNA. Os isolados obtidos durante este estudo estão destacados em negrito. Os números próximos aos ramos representam
os valores de “bootstratp”, para os quais foram incluídos na topologia somente os valores acima de 75%.
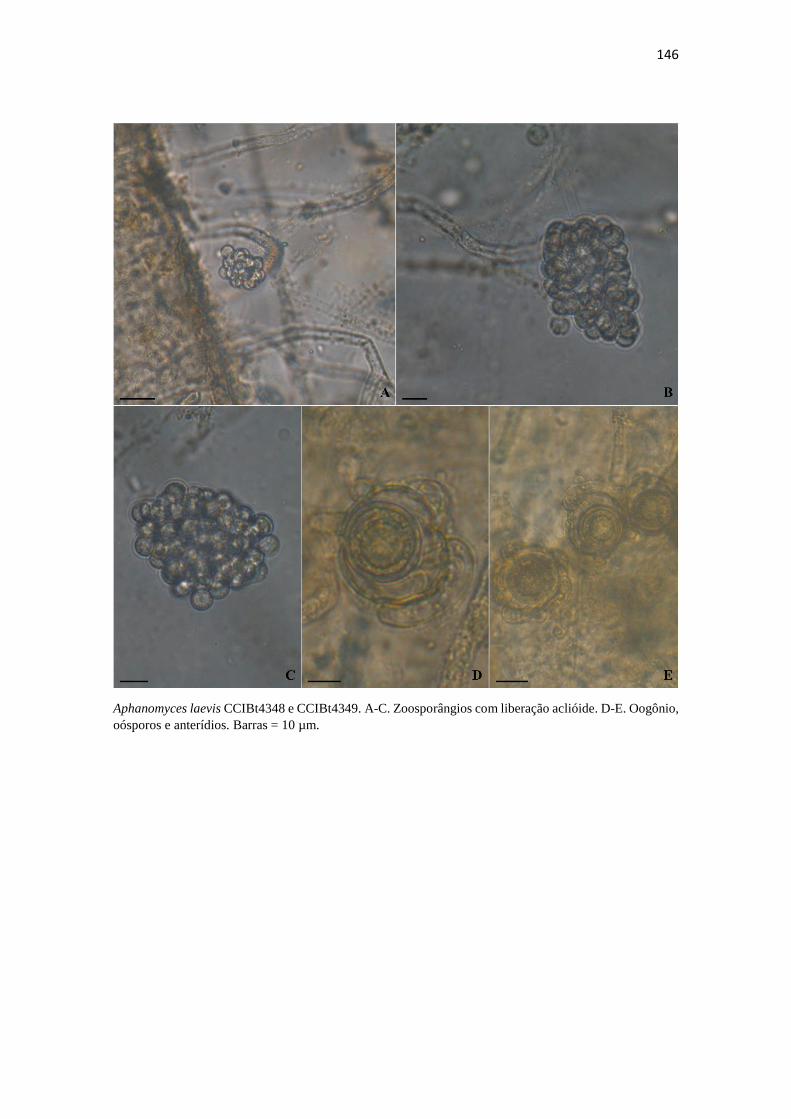
146
Aphanomyces laevis CCIBt4348 e CCIBt4349. A-C. Zoosporângios com liberação aclióide. D-E. Oogônio,
oósporos e anterídios. Barras = 10 µm.

147
Análise de Máxima Verossimilhança da região ITS do rDNA. Os isolados obtidos durante este estudo estão destacados em negrito. Os números próximos aos ramos representam
os valores de “bootstratp”, para os quais foram incluídos na topologia somente os valores acima de 75%.

148
Pythium torulosum (A-E) e Phytopythium vexans (F-I). A-B. Zoosporângios filamentosos inflados de
Pythium torulosum. C-E. Oogônio, oósporos e anterídios de Pythium torulosum. F-H. Zoosporângio
esférico de Phytopythium vexans. I. Oogônio e anterídio de Phytopythium vexans. Barras = 10 µm.

149
Análise concatenada de Máxima Verossimilhança das regiões ITS do rDNA e COX1 do mDNA. Os
isolados obtidos durante este estudo estão destacados em negrito. Os números próximos aos ramos
representam os valores de “bootstratp”, para os quais foram incluídos na topologia somente os valores
acima de 75%.

150
Apêndice I
Táxons
Regiões gênicas
18S ITS 28S RPB1 RPB2 COX1
Chytridiomycota
Asterophlyctis sarcoptoides * * *
Avachytrium sp. * * *
Cladochytrium replicatum * * * * *
Nowakowskiella hemisphaerospora * * *
Nowakowskiella ramosa * * *
*
Nowakowskiella crenulata * * *
Nowakowskiella sp.1 * * * * *
Rhizophydium angulosum * * * * *
Rhizophydium elyense * * *
Siphonaria sp. * * * *
Oomycota
Achlya caroliniana * *
Achlya klebsiana * *
Achlya prolifera * *
Aphanomyces laevis CCIBt4348 * *
Aphanomyces laevis CCIBt4349 * *
Dictyuchus monosporus * *
Dictyuchus pseudodictyon * *
Plectospira gemmifera * *
Pythium inflatum * *
*
Pythium midletonii * *
*
Pythium torulosum * *
*
Pythium sp.1 * *
*
Pythium sp.2 * *
*
Pythium sp.3 * *
*
Pythium sp.4 * *
*
Pythium sp.5 * *
*
Pythium sp.6 * *
*
Pythium sp.7 * *
*
Phytopythium vexans * *
*
Saprolegnia aenigmatica * *
Saprolegnia subterranea * *
Saprolegnia subterranea * *
Saprolegnia subterranea * *