análise proteômica em listeria...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
Análise proteômica em Listeria monocytogenes
Nadielly Xavier de Medeiros
Orientadora: Cíntia Silva Minafra e Rezende
GOIÂNIA 2011

ii
NADIELLY XAVIER DE MEDEIROS
Análise proteômica em Listeria monocytogenes
Seminário apresentado junto à Disciplina Seminários Aplicados do Programa de Pós-Graduação em Ciência Animal da Escola de Veterinária da Universidade Federal de Goiás. Nível: Mestrado
Área de Concentração:
Sanidade Animal, Higiene e Tecnologia de Alimentos
Linha de Pesquisa:
Controle de qualidade de alimentos
Orientador:
Profª. Drª. Cíntia Silva Minafra e Rezende – EVZ/UFG
Comitê de Orientação:
Prof. Dr. Albenones José de Mesquita – EVZ/UFG
Prof. Dr. Cristiano Sales Prado – EVZ/UFG
GOIÂNIA 2011

iii
SUMÁRIO
1 INTRODUÇÃO .............................................................................................. 1
2 REVISÃO DE LITERATURA ......................................................................... 2
2.1 Listeria monocytogenes ............................................................................. 2
2.1.1 Classificação ........................................................................................... 2
2.1.2 Habitat ..................................................................................................... 4
2.1.3 A doença ................................................................................................. 5
2.1.4 Fatores de virulência ............................................................................... 7
2.1.5 Mecanismo de invasão na célula hospedeira ......................................... 8
2.2 Análise proteômica................................................................................... 10
2.2.1 Proteômica ............................................................................................ 11
2.2.2 Métodos utilizados ................................................................................ 12
2.2.3 Preparação da amostra ......................................................................... 13
2.2.4 Eletrofore bidimensional (2-DE) ............................................................ 13
2.2.5 Espectrometria de massa ..................................................................... 14
2.2.6 Aplicações para Listeria monocytogenes .............................................. 15
3. CONSIDERAÇÕES FINAIS ....................................................................... 19
REFERÊNCIAS ............................................................................................. 20

iv
LISTA DE FIGURAS
FIGURA 1: Ocorrência de casos de listeriose por grupo de idade, Áustria
1997-2007................................................................................................ 7
FIGURA 2: Localização aproximada de genes de virulência L. monocytogenes
no cromossomo (3 Mb) e a localização de produtos de genes no meio
extracelular, da parede celular (tom escuro), membrana e citoplasma (tom
claro). ............................................................................................................... 9
FIGURA 3: Mecanismo ―ziper‖ usado por L. monocytogenes para entrada na
célula. ............................................................................................................. 10
FIGURA 4: Processo de infecção celular ....................................................... 11
FIGURA 5: Processo de formação da proteína. ............................................. 12
FIGURA 6: Fluxograma do processo da eletroforese bidimensional ............. 15

v
LISTA DE TABELA
TABELA 1. Classificação de Listeria monocytogenes com base em padrões
de impressões digitais genômicas e associação com surtos epidêmicos ........ 3

vi
LISTA DE QUADROS
QUADRO 1: Proteínas associadas à virulência em L. monocytogenes ........... 7
QUADRO2: Número de proteínas específicas e comuns de EGD e F2365....16

vii
LISTA DE ABREVIATURAS
DNA Ácido desoxirribonucleico
pH Potencial hidrogeniônico
pI Ponto isoelétrico
RNAm Ácido ribonucleico mensageiro

1
1 INTRODUÇÃO
A Listeria monocytogenes é um importante patógeno veiculado em
alimentos. Ocorrem descrições de que está amplamente distribuída no ambiente e
que pode ser encontrada em grande variedade de alimentos tanto frescos como
processados, principalmente de origem animal (MENA et al., 2004).
Listeriose é a doença de origem alimentar causada pela L. monocytogenes.
Surtos de listeriose são raros, mas sempre envolvem mortalidade, sobretudo em
grupos de riscos como pessoas imunodeprimidas, gestantes, neonatos e idosos
(BORGES, et al., 2009).
Uma forma de estudar as doenças e qualquer problema biológico complexo
é analisando as proteínas. Inicialmente é preciso compreender o que é a proteína
e como ela opera celularmente para entender a regulação dos mecanismos da
doença e posterior tratamento (CIERO et al., 2002).
O termo proteoma significa o conjunto de proteínas codificadas pelo
genoma de um organismo. Proteômica é definido como a compreensão da
estrutura, função e interações das proteínas. A análise proteômica é o estudo das
proteínas expressas por um genoma. O desenvolvimento desta técnica está
trazendo um melhor entendimento das vias bioquímicas e os papéis de interações
das proteínas ( MISHRA et. al., 2010).
Diversas proteínas estão relacionadas ao ciclo intracelular da L.
monocytogenes e estão associadas a importantes fatores de virulência como o
processo de adesão e invasão celular (CORSSANT, 2008).
A análise proteômica da Listeria monocytogenes pode identificar os
mecanismos de adaptação deste microrganismo quando está exposto a algum
tipo de estresse. Por este tipo de análise pode-se compreender as possíveis
diferenças entre as cepas virulentas e não virulentas. Diante do exposto,
objetivou-se com este trabalho, a abordagem sobre a análise proteômica de
Listeria monocytogenes.

2
2 REVISÃO DE LITERATURA
2.1 Listeria monocytogenes
O gênero Listeria compreende oito espécies, classificadas como
patogênicas e não patogênicas. De acordo com GERMANO & GERMANO (2008),
dentre as espécies patogênicas, pode-se apontar Listeria monocytogenes que é a
de maior importância em saúde pública e as espécies Listeria weshimeri, Listeria
seeligeri e Listeria ivanovii que raramente acometem o homem. No entanto,
considerando as espécies não patogênicas, menciona-se Listeria innocua e
Listeria grayi, Listeria marthii e Listeria rocourtiae, sendo que estas duas últimas
foram apontadas recentemente por GRAVES et al. (2010) e LECLERCQ no
mesmo ano.
Listeria monocytogenes é um bacilo Gram-positivo, não formador de
esporos, psicrotrófico, catalase positiva e anaéróbio facultativo, medindo entre 0,4
a 0,5 μm de diâmetro por 0,5 a 2,0 μm de comprimento. Apresenta mobilidade a
25°C devido a flagelos e movimento característico denominado tombamento
(GERMANO & GERMANO, 2008).
O agente etiológico da listeriose difere em muitos aspectos de outros
patógenos de origem alimentar por ser resistente a condições ambientais
adversas, como baixo pH e alta concentração de NaCl, sendo resistente a
temperaturas baixas (MARZOCCA, 2004).
2.1.1 Classificação
L. monocytogenes tem 13 sorovariedades denominados por 1/2a, 1/2b,
1/2c, 3a, 3b, 3c, 4a, 4b, 4c, 4d, 4e, 4ab, e 7. Destaca-se que os sorotipos
1/2a, 1/2b, e 4b são responsáveis por 98% dos surtos e o sorotipo 4b é
considerado o mais virulento. Este patógeno é agrupado em três linhagens com
base em sua ribotipagem e associação com surtos (Tabela 2). A linhagem I tem o
maior potencial patogênico e os sorotipos pertencentes a este grupo estão mais
envolvidos em surtos. A linhagem II tem potencial patogênico intermediário e,
possivelmente, responsável por surtos esporádicos, enquanto a linhagem III

3
relaciona-se a risco de baixa patogenicidade e raramente causa infecção humana
(BHUNIA, 2008).
TABELA 1. Classificação de Listeria monocytogenes com base em padrões de impressões digitais genômicas e associação com surtos epidêmicos .
Grupos Surtos Potencial patogênico Sorotipos
predominantes
Linhagem I
Responsável pelo
maior número de
casos
Alto
1/2b, 3b, 4b, 4d, 4e
Linhagem II Casos esporádicos
de listeriose
Médio 1/2a, 1/2c, 3c, 3a
Linhagem III Raramente causam
doença em
humanos
Baixo 4a, 4c
IIIA (Rham +)
4a (não virulenta) e
4c (virulenta)
IIIB (Rham -)
4a , 4c; 7
IIIC (Rham -)
Virulenta 4c
Fonte: Adaptado de BHUNIA, 2008.
Pesquisadores como ROBERTS et al., (2006) e ORSI et al., (2008)
indicaram que a linhagem III pode ser dividida em três subgrupos (IIIA, IIIB, e
IIIC). O subgrupo IIIA é composto por cepas típicas ramnose positivo, com
predominância do sorovar 4a e cepas virulentas do sorovar 4c; o subgrupo IIIB é
composto por cepas não virulentas, ramnose negativa dos sorovares 4a e 4c,
alguns dos quais podem estar relacionados com sorotipo 7; e o subgrupo IIIC é
composto por cepas virulentas, ramnose negativa, do sorovar 4c (LIU, 2006).

4
2.1.2 Habitat
Os membros do gênero Listeria normalmente são encontrados em
mamíferos, peixes, anfíbios, aves e insetos, na maioria dos casos portadores
assintomáticos que liberam a bactéria nas fezes, sem desenvolver infecção. O
reservatório primário parece ser o solo e a vegetação mas estão amplamente
distribuídos no ambiente e têm sido isolados de solo, água, ração animal, plantas
em decomposição, resíduos de matadouros e nas fezes de animais saudáveis,
incluindo seres humanos (SILVA et al., 2007). Estes microrganismos podem se
tornar endêmicos em ambientes de processamento de alimentos (MCLAUCHLIN
et al., 2004).
A rápida capacidade de adesão em superfícies faz com que a L.
monocytogenes seja um risco para saúde pública e indústrias de alimentos. Uma
vez presente na matéria-prima este microrganismo pode aderir rapidamente à
superfície de equipamentos em aço inoxidável e utensílios, sendo capaz de se
multiplicar, formando biofilmes, favorecendo a contaminação do produto final
(OLIVEIRA et al., 2010).
LIMA et al. (2005) observaram que a disseminação da L. monocytogenes
na planta de indústria produtora de linguiça frescal, promove a contaminação do
produto final. ROSSI et al. (2011) isolaram L. monocytogenes em 3,75% das
amostras de linguiça do tipo frescal.
A presença de L. monocytogenes no trato intestinal de animais possibilita a
contaminação dos criatórios e dos abatedouros e assim, podendo atuar como
uma fonte de introdução deste patógeno na linha de abate o que reforça a
possibilidade de contaminação de alimentos de origem animal (NALÉRIO et. al,
2009).
RIVOAL et al. (2010) relataram a presença de L. monocytogenes em
17,36% dos ovos crus e 2,1% em ovos pasteurizados. Eles também observaram
que a ocorrência no verão foi maior do que no inverno e no outono.
Ao analisar amostras de pele e intestino de peixe, CHEN et al. (2010) não
detectaram Listeria monocytogenes, mas observaram a presença deste patógeno
em 76,7% dos peixes refrigerados.

5
GEBRETSADIK et al. (2011) detectaram L. monocytogenes em 13% das
amostras de leite cru, 4,3% das amostras de ovo líquido, 2,6% em carne crua e
1 % em queijo tipo cottage.
2.1.3 A doença
BUENO et al. (2010) relataram a forte associação genética entre as
estirpes de Listeria monocytogenes isoladas de alimentos e de espécimes clínicos
do Brasil, o que indica uma possível ligação de transmissão de L. monocytogenes
de alimentos para seres humanos.
Manifestações de listeriose incluem septicemia, meningite (ou
meningoencefalite), encefalite e infecções intra-uterinas ou cervical em mulheres
grávidas, o que pode resultar em aborto espontâneo. O início dos distúrbios
mencionados é geralmente precedido por sintomas de gripe, incluindo febre
persistente. Foi relatado que os sintomas gastrointestinais, como náuseas,
vômitos e diarréia podem preceder as formas mais graves de listeriose ou podem
ser os únicos sintomas expressos. O início do tempo às formas graves de
listeriose é desconhecido, mas pode variar de poucos dias a três semanas. O
tempo de início de sintomas gastrointestinais é desconhecido, mas provavelmente
é maior do que 12 horas (FDA, 2009).
Em 1929 o ocorreu o caso índice de listeriose sendo identificado e desde
então ficou comprovado que esta enfermidade apresenta-se esporadicamente,
em todo o mundo. L. monocytogenes é o agente etiológico de aproximadamente
98% dos casos que ocorrem em pessoas e 85% dos casos que ocorrem nos
animais (MANTILLA, 2007).
No geral, a taxa de letalidade devido à infecção é de cerca de
20-30%, com uma expectativa de 500 óbitos anualmente nos Estados Unidos. A
dose infectante para este patógeno não é conhecida, no entanto, é estimada em
cerca de 102-106 células, dependendo do estado imunológico do hospedeiro
(BHUNIA, 2008).
De acordo com as descrições de KASPER (2009), outro fator que maximiza
a listeriose relaciona-se às doenças associadas. O risco é aumentado para
indivíduos imunocomprometidos, gestantes e idosos. Pode-se mencionar que de
136 casos de listeriose na Áustria, 99 apresentavam uma ou mais doenças
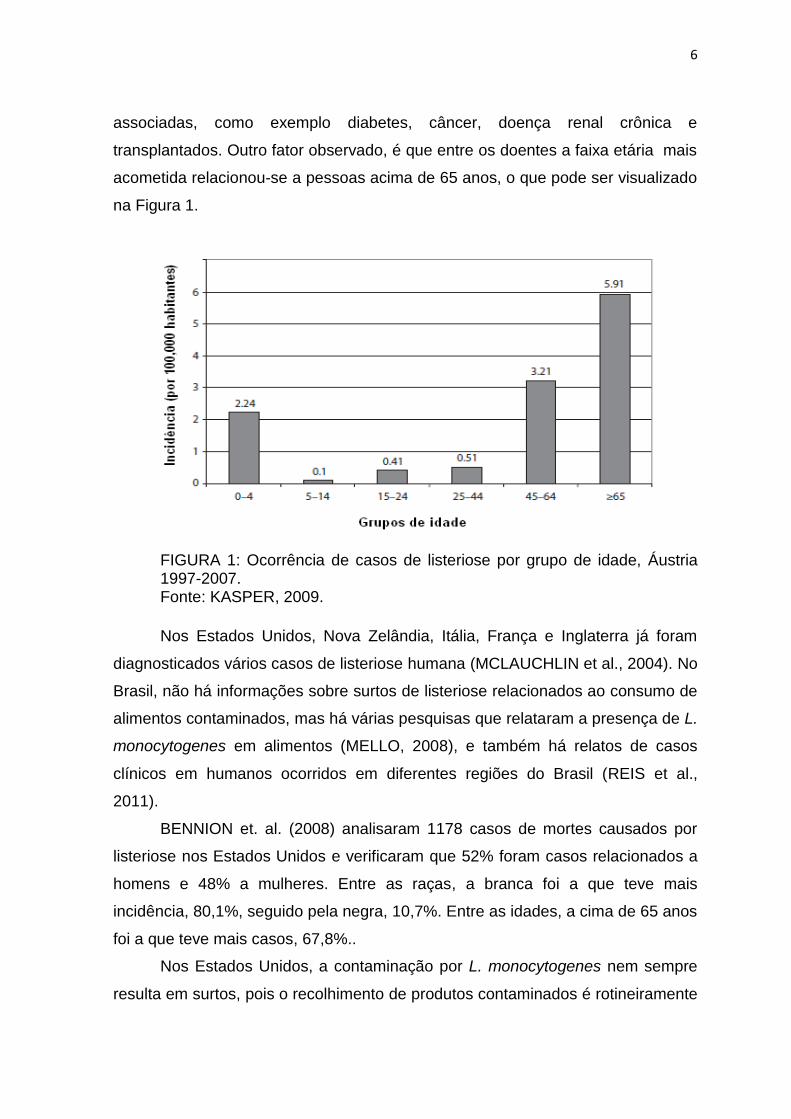
6
associadas, como exemplo diabetes, câncer, doença renal crônica e
transplantados. Outro fator observado, é que entre os doentes a faixa etária mais
acometida relacionou-se a pessoas acima de 65 anos, o que pode ser visualizado
na Figura 1.
FIGURA 1: Ocorrência de casos de listeriose por grupo de idade, Áustria 1997-2007. Fonte: KASPER, 2009.
Nos Estados Unidos, Nova Zelândia, Itália, França e Inglaterra já foram
diagnosticados vários casos de listeriose humana (MCLAUCHLIN et al., 2004). No
Brasil, não há informações sobre surtos de listeriose relacionados ao consumo de
alimentos contaminados, mas há várias pesquisas que relataram a presença de L.
monocytogenes em alimentos (MELLO, 2008), e também há relatos de casos
clínicos em humanos ocorridos em diferentes regiões do Brasil (REIS et al.,
2011).
BENNION et. al. (2008) analisaram 1178 casos de mortes causados por
listeriose nos Estados Unidos e verificaram que 52% foram casos relacionados a
homens e 48% a mulheres. Entre as raças, a branca foi a que teve mais
incidência, 80,1%, seguido pela negra, 10,7%. Entre as idades, a cima de 65 anos
foi a que teve mais casos, 67,8%..
Nos Estados Unidos, a contaminação por L. monocytogenes nem sempre
resulta em surtos, pois o recolhimento de produtos contaminados é rotineiramente

7
feitos. O custo estimado associados à contaminação por Listeria é
aproximadamente 2 bilhões de dólares por ano (BHUNIA, 2008).
2.1.4 Fatores de virulência
Cada etapa do ciclo de vida intracelular da L. monocytogenes depende da
produção e ação dos fatores de virulência. Diversas proteínas têm sido
reconhecidas como importantes fatores de virulência em L. monocytogenes por
estarem envolvidas no ciclo intracelular desta bactéria, incluindo a adesão,
entrada na célula hospedeira, movimento intracelular, lise celular, e para escapar
do sistema imunológico (Tabela 4) (BHUNIA, 2008).
QUADRO 1. Proteínas associadas à virulência em L. monocytogenes Proteínas Função
Fator proteína reguladora (Prfa) Regulador central de virulência
Internalina A (InlA) Entrada em células que expressam seu
receptor, a caderina-E
Internalina B (InlB) Entrada em células que expressam um dos
receptores, gC1qR HGF-SF, ou Met eo
glicoaminoglicanas (GAGs)
p60 (hidrólise da parede celular) Adesão e invasão celular
Proteína de adesão de Listeria (LAP) Adesão em células epiteliais intestinal
Fosfolipase (plcA -PI-PLC; plcB – PC-
PLC)
Lise da membrana do vacúolo
Listeriolisina O (LLO) Necessários para escapar do vacúolo por
lise da membrana do fagossomo
Proteína de virulência (Vip) Invasão em células epiteliais
Proteína polimerizadora de actina (ActA) Proteína de superfície necessária para
montagem da cauda de actina, que está
envolvida no movimento bacteriano no
interior do citoplasma
Fonte: adaptado de BHUNIA, 2008.

8
As principais proteínas para as etapas sucessivas desta bactéria no
processo infeccioso incluem a proteína de superfície Internalina A (InlA) e InlB,
para o processo de entrada, a toxina listeriolisina O (LLO) e a fosfolipases C (PI-
PLC), associadas ao processo de fuga do vacúolo, e a proteína de superfície
ACTA, responsável pela produção da cauda de actina (Figura 2) (CORSSANT,
2008).
FIGURA 2: Localização aproximada de genes de virulência L. monocytogenes no cromossomo (3 Mb) e a localização de produtos de genes no meio extracelular, da parede celular (tom escuro), membrana e citoplasma (tom claro). Fonte: BHUNIA, 2008.
2.1.5 Mecanismo de invasão na célula hospedeira
A L. monocytogenes é na maior parte, intracelular. Sua internalização
inicia-se com a adesão do microrganismo à superfície da célula, posteriormente
se tem a interação entre os ligantes da superfície bacteriana com os respectivos
receptores celulares, mecanismo conhecido como ―zíper‖ (figura3) (CORSSANT,
2008).

9
FIGURA 3: Mecanismo ―ziper‖ usado por L. monocytogenes para entrada na célula. Fonte: Adaptado de ALONSO et al., 2004.
FIGURA 4: Processo de infecção celular
Fonte: COSSART, 2008.
Posteriormente ocorre o processo de espalhamento célula a célula, que é
ilustrado na Figura 4.
Após a entrada nas células, as bactérias são aprisionadas em um vacúolo,
a partir do qual elas escapam depois de cerca de 30 minutos. Uma vez dentro do
citosol elas se multiplicam com um tempo de geração de uma hora,
posteriormente começam a polimerizar actina, gerando uma rede de ramificações
de filamentos, a cauda de actina. Este processo de polimerização em um pólo da

10
bactéria produz energia para impulsionar as bactérias que se movem no
citoplasma a uma velocidade de cerca de 10 mm por minuto (CORSSANT, 2008).
Quando a bactéria alcança a membrana plasmática da célula infectada,
ocorre sua projeção e indução de formação de saliências, permitindo a invasão
nas células vizinhas, gerando um vacúolo com duas membranas, em seguida a
bactéria escapa deste vacúolo, iniciando-se um novo ciclo de replicação
(CORSSANT, 2008).
Acredita-se que esse fenômeno de invasão por célula direto permita
disseminação da bactéria em vários tecidos infectados e que esta bactéria fique
protegida dos mecanismos de defesas do hospedeiro (CORSSANT, 2008).
2.2 Análise proteômica
Antibióticos desempenham papel importante no combate às doenças
infecciosas. No entanto, a resistência à maioria dos antibióticos e o surgimento de
cepas altamente virulentas ameaça o tratamento de doenças que hoje são
curáveis. Os mecanismos determinantes da virulência bacteriana, do grau de
patogenicidade e resistência a antibióticos podem estar ligados à variabilidade
genética e à pressão evolutiva, que favorecem a adaptação dos microrganismos
sobreviventes, constituindo maior ameaça para a saúde humana uma vez que as
estratégias de defesa contra eles se tornam inúteis (MALMSTRO, 2011).
O genoma é o todo o conjunto de ácido desoxirribonucleico (DNA) de uma
célula. A sequência do genoma serve como um excelente começo para aumentar
a compreensão de como bactérias causam a doença e adaptam-se à pressão
evolutiva exercida pelo hospedeiro e pelo uso de antibióticos (MALMSTRO,
2011).
Atualmente onze genomas de Listeria monocytogenes estão disponíveis:
Listeria monocytogenes 08-5578, Listeria monocytogenes 08-5923 1/2a, Listeria
monocytogenes EGD-e 1/2a, Listeria monocytogenes HCC23 4a, Listeria
monocytogenes M7, Listeria monocytogenes str. 4a L99, Listeria monocytogenes
Clip81459 4b, Listeria monocytogenes strain 4b F2365, Listeria monocytogenes
J0161 1/2a, Listeria monocytogenes Finland 1988, Listeria monocytogenes FSL
R2-561 e Listeria monocytogenes 10403S (NCBI, 2011).

11
A sequência dos aminoácidos das proteínas pode ser deduzida a partir da
sequência do genoma. O ácido ribonucleico mensageiro (RNAm) é usado como
um modelo para montar a cadeia de aminoácidos que formam uma proteína
(Figura 5) CLANCY & BROWN, 2008).
FIGURA 5: Processo de formação da proteína. Fonte: SILVA et al., 2007
2.2.1 Proteômica
O termo proteoma se origina das palavras proteínas e genoma. O
proteoma representa todas proteínas codificadas pelo genoma de um organismo.
Proteômica, portanto, é definido como o teor de proteína total de uma célula ou de
um organismo. Proteômica é o entendimento da estrutura, função e interações
das proteínas do organismo. Proteínas controlam o fenótipo de uma célula
através da realização de todas as funções em uma célula (MISHRA et. al., 2010).
O estudo da proteômica tem implicações diretas em vários campos da
biologia e da biotecnologia. Dentre suas vertentes comprova-se a descoberta de
vias metabólicas nas diversas etapas celulares, favorecendo o conhecimento na
biologia celular e da bioquímica, viabilizando a identificação de moléculas
bioativas. Por conseqüência há o desenvolvimento de novos medicamentos, e
identificação e caracterização de moléculas exógenas ou endógenas específicas
a doenças (ROCHA et al., 2005).

12
2.2.2 Métodos utilizados
A análise proteômica é realizada por duas técnicas, a de separação das
proteínas e a de identificação. As tecnologias mais usadas para a análise de
proteoma são a eletroforese de duas dimensões (2-DE), que pode ser vista na
Figura 6, a qual realiza a separação das proteínas, e a espectrometria de massa,
usada para a identificação das proteínas (CIERO et. al, 2002).
FIGURA 6: Fluxograma do processo da eletroforese bidimensional Fonte: Adaptado de SILVA et al., 2007.

13
As principais etapas da eletroforese bidimensional são: preparação da
amostra; primeira dimensão; segunda dimensão e detecção das proteínas
(ROCHA et al., 2005).
2.2.3 Preparação da amostra
As proteínas devem ser parcialmente purificadas, completamente
solubilizadas, desagregadas, desnaturadas e reduzidas (ROCHA et al., 2005).
Primeiramente as amostras devem ser lavadas com tampões, depois
deverá ser feita a precipitação desta proteína. Após a lavagem das células, o
conteúdo protéico é extraído (SANTOS et al., 2004). Para a extração da proteína
é necessário que se faça a ruptura da parede celular e para isto vários métodos
podem ser utilizados, como a lise enzimática ou inibição da síntese da parede
celular, choque osmótico, aquecimento, congelamento e descongelamento,
secagem, em vórtex, com esferas de vidros, ultra-som, moinho de bolas, prensa
francesa e homogeneizador de alta pressão (PESSOA JÚNIOR et al., 2005).
O extrato bruto obtido no rompimento celular, geralmente contém outros
componentes celulares além da proteína tais como ácidos nucléicos, lipídios e
hidratos de carbono. Para eliminar ou reduzir a quantidade destes contaminantes
deve-se precipitar seletivamente as proteínas presentes nos extratos brutos,
usando, por exemplo, kits comerciais (SANTOS et al., 2004).
Para realizar a focagem isoelétrica, as amostras deverão ser solubilizadas
em uma solução desnaturante, não iônica, que permita a separação das proteínas
conforme o seu ponto isoelétrico (pI). Normalmente, estas soluções são
compostas por uréia, para desnaturar as proteínas, um agente redutor, para evitar
a oxidação das proteínas, um detergente não iônico para solubilizar as proteínas
e anfólitos para facilitar a focagem isoelétrica (SANTOS et al., 2004).
2.2.4 Eletrofore bidimensional (2-DE)
As proteínas são separadas de acordo com duas das suas propriedades: o
ponto isoelétrico, em uma primeira dimensão e massa molecular, numa segunda

14
dimensão. A primeira dimensão é denominada focalização isoelétrica (IEF),
processo no qual as proteínas são separadas de acordo com a carga, pois elas
migram até quando o potencial hidrogeniônico (pH) for igual ao seu ponto
isoelétrico (pI). Na segunda dimensão, as proteínas são separadas de acordo
com suas massas moleculares (SANTOS, 2004).
Os parâmetros usados na primeira dimensão e na segunda dimensão são
independentes, então a separação tem um alto nível de resolução e isto permite a
visualização de centenas de proteínas diferentes ao mesmo tempo (SILVA et al.,
2007).
Para visualizar as proteínas, após a eletrofore 2-DE faz-se a coloração das
proteínas com corantes ou reagentes fluorescentes. A coloração mais usada é a
coloração por Azul de Coomassie e coloração por prata (CIERO et al., 2002).
Após a coloração obtém-se um perfil bidimensional de pontos, denominados de
spots, os quais contêm múltiplas cópias de uma proteína (SILVA et al., 2007).
As principais limitações da técnica 2-DE estão relacionadas com a baixa
detecção de classes específicas de proteínas, incluindo aquelas com pouca
abundância, baixa solubilidade, alta ou baixa massa molecular e com pI muitos
alcalinos (GYGI, et al., 2000).
2.2.5 Espectrometria de massa
A espectrometria de massa consiste em identificar as proteínas de
interesse. Esta técnica determina a massa molecular de fragmentos peptídicos
obtidos da digestão das proteínas com uma enzima proteolítica, como a tripsina.
O padrão de massas obtidos denomina-se ―peptide mass fingerprinting‖
(impressão digital da proteína). Para identificar a proteína deve-se fazer o
cruzamento dessa informação com informações disponíveis nas bases de dados
genômicas (SANTOS et al., 2004).
Nas últimas décadas grandes avanços foram obtidos na identificação das
proteínas por meio do uso da espectrometria de massa. Uma das principais
técnicas espectrométricas para a análise de proteínas é a espectrometria de
massa com base na dessorção e ionização das proteínas com laser, auxiliado por

15
uma matriz (MALDI, matrix assisted laser desorption/ionization), que analisa a
massa através do tempo de vôo dos íons no tubo de análise (ToF – Time of
Flying) (ROCHA, 2005).
MALDI é uma alternativa valiosa para identificação de proteínas e é
frequentemente utilizada para complementar resultados de análise de proteínas
obtidos por outras técnicas, por ser mais sensível e mais tolerante à presença de
contaminantes, como sais ou pequena quantidade de detergente (DOMON, et al.,
2006).
A técnica MALDI baseia-se na absorção da radiação ao comprimento de
onda do laser por uma solução saturada de uma matriz, que é misturada em
solução com quantidades muito pequenas de proteínas (o analito). Esta matriz
tem a função de absorver a radiação do laser para possibilitar uma transferência
eficiente da energia do pulso de radiação para o analito e isolar as moléculas de
analito umas das outras, para diminuir as ligações intermoleculares e permitir a
dessorção intacta das moléculas (FERNANDEZ-LIMA, et al., 2005).
A energia absorvida faz com que a matriz evapore e o analito passe para a
fase gasosa. A formação dos íons ocorre pela transferência de carga das
moléculas da matriz para o composto, assim sendo, os íons são separados no
analisador de massas Time-of-flinght (TOF) de acordo com o tempo de vôo de
cada um deles até atingir o detector, calculando-se então a massa molecular do
composto (MARQUEZ, 2006).
2.2.6 Aplicações para Listeria monocytogenes
A análise proteômica em L. monocytogenes permite identificar as proteínas
expressas ou reprimidas com diversos estímulos externos e diversas condições,
como a presença de NaCl, ácidos e baixas temperaturas (ESTEVES, 2009);
também é possível fazer comparação de variedades da mesma espécie (GIOTIS
al., 2008) e de espécies diferentes (CALVO et al. 2005).
Para compreender as diferenças entre as linhagem I e II de L.
monocytogenes, DONALDSON et al. (2009) analisaram as proteínas de
expressão do sorotipo 1/2a, cepa EGD, linhagem II e a cepa F2365, sorotipo 4b
pertencente à linhagem I. Os autores observaram um número surpreendente de

16
diferentes proteínas expressas entre as duas linhagens (Tabela 5) e relataram
que houve diferença entre elas na expressão de proteínas no envelope celular e
proteínas flagelares. De acordo com estes autores, estas diferenças entre estes
dois sorotipos podem ajudar a explicar por quê o sorotipo 4b está associado com
surtos de listeriose humana, enquanto o sorotipo 1/2a causa casos esporádicos
de listeriose humana.
No envelope celular e processos celulares foram identificadas proteínas
envolvidas na divisão celular, membrana bioenergética, mobilidade e quimiotaxia,
secreção de proteínas, transporte e ligação de proteínas. No metabolismo
intermediário foram identificadas proteínas relacionadas ao metabolismo de
carboidratos e moléculas relacionadas, metabolismo dos aminoácidos,
metabolismo de lipídeos, metabolismo de coenzimas metabolismo de fosfato, etc.
Nas vias de informações foram identificadas proteínas envolvidas na replicação
de DNA, recombinação de DNA, síntese de RNA, síntese de proteínas,
enovelamento de proteínas; proteínas ribossomais; restrição, modificação e
reparação do DNA (DONALDSON et al., 2009).
QUADRO 2: Número de proteínas específicas e comuns de EGD e F2365.
Função EGD F2365 Comum
Envelope celular e processos celulares 141 73 240
Metabolismo intermediário 132 73 286 Vias de informações 107 49 224
Fonte: DONALDSON et al., 2009
GIOTIS et al. (2008) analisaram o estudo das proteínas em Listeria
monocytogenes 10403S, pertencente ao sorotipo 1/2a. Neste estudo, os autores
analisaram cepas controle (exposição em pH 7,4 por uma hora) e cepas
adaptadas (exposição a pH 9,5 por uma hora). Em comparação com a cepa
controle, eles verificaram que 113 proteínas não foram expressas em bactérias
em estágio de estresse e oito proteínas foram recém sintetizadas. Entre as
proteínas identificadas como reguladoras, os autores encontraram duas proteínas
de choque térmico GroEL e DnaK, as quais também estão relacionadas à
tolerância ao estresse. Eles relataram uma forte evidência de que a proteína

17
encontrada, ALTR, minimiza a alcalinização excessiva e gastos de energia,
relacionados à mobilização de fontes de carbono disponível. Os autores
concluíram que, o pH alcalino proporciona múltiplo estresse de resistência em L.
monocytogenes, isto pode ser vital para a sobrevivência no trato gastrintestinal
humano, bem como no sistema de processamento de produtos alimentares, onde
as condições alcalinas prevalecem.
Analisando a parede celular da L. monocytogenes EGD-e e L. innocua
CLIP11262, CALVO et al. (2005) observaram que as proteínas P60, e a P45
estão presentes nas duas espécies. Já as proteínas PbpA e MurA/Nam estão
presentes somente em L. monocytogenes, resultados que podem explicar porquê
L. monocytogenes está mais envolvida em surtos.
ESTEVES (2009) analisou proteínas de quatro cepas de L. monocytogenes
pertencentes aos sorotipos 1/2a, 4b, 4c e 4d/4e, sob estresse nutricional (meio
mínimo – MWB, a baixa temperatura (11ºC), estresse ácido (pH 5,5) e estresse
salino (11% NaCl – w/v). No meio mínimo (MWB), a 37°C, foram identificadas as
proteínas p60, internalina C e listeriolisina O. Em MWB a 11°C a proteína p60 foi
identificada em todas as cepas, enquanto a listeriolisina O e a internalina C não
foi detectada em nenhuma cepa. A flagelina foi identificada nas cepas 4c e 4d/4e
e, com particular abundância, na cepa 1/2a, mas não foi encontrada na cepa 4b.
Na presença de 11% (m/v) de NaCl ou a pH 5,5 foram visualizadas, pelo menos,
duas proteínas sob expressas. A autora concluiu que a identificação de proteínas
diferencialmente expressas por L. monocytogenes poderá contribuir para o
controle efetivo deste patógeno.
Realizando a análise proteômica de cepas de L. monocytogenes cultivadas
a 37°C e 4°C, CACACE et al. (2010) observaram que houve um padrão
semelhante de proteínas expressas com uma média de 540 ± 10, das quais 134
teveram uma mudança significativa na intensidade. Os autores relataram que a
Listeria monocytogenes exerce uma regulação hierárquica para controlar as taxas
de reação glicolítica para compensar o efeito da temperatura baixa.
Utilizando a metodologia de análise proteômica, GUILBAUD et al. (2008)
observaram que a fumaça líquida inibiu o crescimento e a sobrevivência de L.
monocytogenes. Esta bactéria não desenvolveu uma resposta fisiológica
adaptativa e diferentes vias metabólicas foram afetadas. Fumaça líquida,

18
provavelmente, atuou sobre a membrana celular e, diretamente ou indiretamente,
sobre a estabilidade de LLO; isso poderia resultar em atenuação da virulência de
L. monocytogenes presentes em alimentos com fumaça líquida.
L. monocytogenes, em ambiente intracelular, sofre adaptações
metabólicas, tornando um meio mais favorável para sua sobrevivência do que
meio extracelular (VELDE et. al. 2009).
Ao avaliar a atividade da proteína σB em L. monocytogenes após a
exposição à vancomicina SHIN et al. (2010) observaram que esta proteína é
significativamente induzida pela adição deste princípio antimicrobiana, concluindo
que estes resultados sugerem que esta proteína pode contribuir para o
monitoramento da integridade da parede celular.
Pelo exposto, considerando a abrangência, versatilidade e dinamicidade
das sequências protéicas e sua interface com os microrganismos, a análise
proteômica permite uma série de avaliações tendo em vista os mecanismos de
patogenicidade destes agentes veiculados por alimentos.

19
3. CONSIDERAÇÕES FINAIS
As sequências de aminoácidos definem as proteínas, as quais são ricas em
informação e estão envolvidas em todos os processos biológicos. Propriedades
destas biomoléculas, como a expressão biológica e química, a atividade
específica, estado de modificação e associação a outras moléculas são
essenciais para a descrição da diversidade do sistema biológico
A proteômica veio para revolucionar os estudos do genoma. Com a análise
proteômica, pode-se obter dados sobre a organização e funções das proteínas e
então, compreender a função do gene.
Na busca de caracterizar o proteoma de uma determinada célula ou
organismo, deve-se lembrar que o proteoma é dinâmico, por refletir o ambiente
imediato em que é estudado. Em resposta a estímulos internos ou externos, as
proteínas podem ser modificadas por pós-traducionais. Assim, a análise
proteômica de determinada célula permite saber quais proteínas estão sendo
expressas em um certo momento.
A proteômica define informações importantes sobre L. monocytogenes,
destacando que com esta análise é possível compreender a alta resistência deste
patógenos a diversas condições ambientais, tais como baixas temperaturas, alta
concentração de NaCl, acidificação do meio e quais os mecanismos utilizados
para resistência a alguns antibióticos.
A comparação de proteínas em cepas patogênicas e não patogênicas de
microrganismos, permite identificar quais proteínas são expressas ou a ausência
de expressão e com isto pode-se entender o papel da proteína em associação ao
desencadeamento da doença e, por consequência, o seu controle. A identificação
das proteínas relacionadas a cepas virulentas auxilia no desenvolvimento de
métodos de diagnósticos da listeriose e assim, facilita o tratamento e cura dos
pacientes.
Por sua amplitude, estima-se que a aplicação deste tipo de análise
possibilitará enormes avanços ao diagnóstico laboratorial e conhecimento mais
específico e pontual de agentes patogênicos.

20
REFERÊNCIAS
1 Alonso, A., Portillo, F. G. Hijacking of eukaryotic functions by intracellular
bacterial pathogens. International Microbiology, Madri, v. 7, p. 181–191,
2004.
2 BENNION, J. R., SORVILLO, F., WISE, M. E., KRISHNA, S. MASCOLA L.
Decreasing Listeriosis Mortality in the United States, 1990–2005. Clinical
Infectious Diseases, Chicago, v. 47, p. 867–874, 2008.
3 BHUNIA, A. K. Foodborne Microbial Pathogens: Mechanisms and
Pathogenesis. Ed. Springer. 290p. 2008.
4 BORGES, M. F., ANDRADE A. P. C., ARCURI, E. F.KABUKI, D. Y.
KUAYE, A. Y. Listeria monocytogenes em leite e produtos lácteos.
Embrapa Agroindústria Tropical Fortaleza, CE. 2009. Empresa Brasileira
de Pesquisa Agropecuária Embrapa Agroindústria. Tropical Ministério da
Agricultura, Pecuária e Abastecimento, Brasília, 2009.
5 BUENO, V. F., BANERJEE, P., BANADA, P. P., MESQUITA, A. J.,
LEMES-MARQUES, E. G., BHUNIA, A. K. Characterization of Listeria
monocytogenes isolates of food and human origins from Brazil using
molecular typing procedures and in vitro cell culture assays. International
6 Journal of Environmental Health Research, Abingdon, v.20, n.1, p .43-
59, 2010.
7 CACACE, G., MAZZEO, M. F., SORRENTINO, A., SPADA, V., MALORNI,
A., SICILIANO, R. A. Proteomics for the elucidation of cold adaptation
mechanisms in Listeria monocytogenes. Journal of proteomics, Weinheim
V. 7 3, p. 2021 – 2030, 2010.
8 CALVO, E., PUCCIARELLI, M. G., BIERNE, H., COSSART, P., ALBAR, J.
P., PORTILLO, F. G. Analysis of the Listeria cell wall proteome by two-
dimensional nanoliquid chromatography coupled to mass spectrometry.
Proteomics, Weinheim, v. 5, p. 433–443, 2005.

21
9 CHEN, B.Y., PYLA, R., KIM, T.J., SILVA, J.L., JUNG, Y.S. Prevalence and
contamination patterns of Listeria monocytogenes in catfish processing
environment and fresh fillets. Food Microbiology, Londom, v. 27p. 645-
652, 2010.
10 CIERO, L., BELLATO, C. M. PROTEOMA: Avanços Recentes em Técnicas
de Eletroforese Bidimensional e Espectrometria de Massa. Biotecnologia
Ciência & Desenvolvimento, Brasília, v. 29 p. 158 – 164. 2002.
11 CLANCY, S., BROWN, W. Translation: DNA to mRNA to protein. Nature,
Hawthorne, v. 1. P. 1-3., 2008. Disponível em:
http://www.nature.com/scitable/topicpage/translation-dna-to-mrna-to-
protein-393. Acesso em: 23/08/2011.
12 COSSART P., TOLEDO-ARANA, A.. Listeria monocytogenes, a unique
model in infection biology: an overview. Microbes and Infection, Paris,
v.10, p.1041-1050, 2008.
13 DOMON B., AEBERSOLD, R. Mass Spectrometry and Protein Analysis.
Science, Washington, v. 312, n. 5771, p. 212-217, 2006.
14 . DONALDSON, J. R., NANDURI, B., BURGESS, S. C., LAWRENCE, M. L.
Comparative Proteomic Analysis of Listeria monocytogenes Strains F2365
and EGD. APPLIED AND ENVIRONMENTAL MICROBIOLOGY,
Washington. v. 75, p. 366–373, 2009.
15 ESTEVES, B. S. I. Análise comparativa de proteínas secretadas em
Listeria monocytogenes. Mestrado em Engenharia Alimentar - Qualidade e
Segurança Alimentar - Instituto Superior de Agronomia. 2009. 60p.
http://hdl.handle.net/10400.5/1991.
16 FERNANDEZ-LIMA, F. A., PONCIANO, C.R., PEDRERO E.,. SILVEIRA,
E.F. Acoplamento das espectrometrias de mobilidade iônica e de massa
maldi-TOF. Revista Brasileira de Aplicações de Vácuo, Rio de Janeiro ,
v. 24, n. 2, 74-80, 2005.

22
17 FDA. Food and Drug Administration. FDA Center for Food Safety and
Applied Nutrition, 2009. Foodborne pathogenic microorganisms and natural
toxins handbook (Bad Bug Book). Disponível em
http://www.fda.gov/Food/FoodSafety/FoodborneIllness/FoodborneIllnessFo
odbornePathogensNaturalToxins/BadBugBook/ucm070064.htm Acesso
em: 02/05/2011.
18 GEBRETSADIK, S., KASSA, T., ALEMAYEHU, H., HURUYA, K.,
KEBEDEB, N. Isolation and characterization of Listeria monocytogenes and
other Listeria species in foods of animal origin in Addis Ababa, Ethiopia.
Journal of Infection and Public Health, Riyadh, v. 4, p. 22—29, 2011.
19 GERMANO, P. M. L., GERMANO, M. I. S.Higiene e vigilância sanitária
de alimentos. 3 ed. Barueri, Manole, 2008. 629 p.
20 GIOTIS, E. S, MUTHAIYAN, A., BLAIR, I. S, WILKINSON B. J.,.
MCDOWELL, D. A. Genomic and proteomic analysis of the Alkali-Tolerance
Response (AlTR) in Listeria monocytogenes 10403S. BioMed Central
Microbiology, London, 2008, 8:102. P. 1-11.
21 GRAVES, L. M., HELSEL, L. O., STEIGERWALT, A. G., MOREY, R. E.,
DANESHVAR, M. I., ROOF, S. E., ORSI, R. H., FORTES, E. D., MILILLO,
S. R., DEN BAKKER, H. C., WIEDMANN, M., SWAMINATHAN, B.,
SAUNDERS. B. D. Listeria marthii sp. nov., isolated from the natural
environment, Finger Lakes National Forest. International journal of
systematic and evolutionary microbiology, Reading, v.60, n. 6 p.1280-
1288, 2010.
22 GUILBAUD, M., CHAFSEY I., PILET, M.-F., LEROI, F., PRÉVOST, H.,
HE´BRAUD, M., DOUSSET, X. Response of Listeria monocytogenes to
liquid smoke. Journal of Applied Microbiology, Oxford, n.104, p. 1744–
1753, 2008.

23
23 Gygi SP, Aebersold R. Mass spectrometry and proteomics. Current
Opinion in Chemical Biology, London, v. 4, p. 489–494, 2000.
24 KASPER, S., HUHULESCU, S., AUER, B., HELLER, I., KARNER, F.,
WÜRZNER, R., WAGNER, M., ALLERBERGER, F. Epidemiology of
listeriosis in Austria. Wien Klin Wochenschr, Wien, v. 121, p. 113–119.
2009.
25 LECLERCQ, A., CLERMONT, D., BIZET, C., GRIMONT, P. A., LE
FLÈCHE-MATÉOS, A., ROCHE, S. M., BUCHRIESER, C., CADET-
DANIEL, V., LE MONNIER, A., LECUIT, M., ALLERBERGER, F. Listeria
rocourtiae sp. nov. International journal of systematic and evolutionary
microbiology, Reading,. v. 60 n.9 p. 2210-2214, 2010.
26 LIMA, A. S., VON LAER, A. E., TRINDADE P. S., SILVA, W. P.
Disseminação de Listeria monocytogenes no processamento de lingüiça
mista frescal avaliada por sorologia e rapd. Alimentos e nutrição [online],
Araraquara v.16, n.3, p. 245-251, 2005. Disponível em: http://serv-
bib.fcfar.unesp.br/seer/index.php/alimentos/article/viewFile/475/442.
Acesso em: 03/08/2011.
27 LIU, D., M. L. LAWRENCE, M. WIEDMANN, L. GORSKI, R. E.
MANDRELL, A. J. AINSWORTH, AUSTIN, F. W. Listeria monocytogenes
subgroups IIIA, IIIB, and IIIC delineate genetically distinct populations with
varied pathogenic potential. Journal of Clinical Microbiology. Washington, v.
44, p. 4229–4233, 2006.
28 MALMSTRO, L. MALMSTRO, J., AEBERSOLD, R.. Quantitative
proteomics of microbes: Principles and applications to virulence.
Proteomics, Weinheim, v. 11, p. 2947–2956. 2011
29 MANTILLA, S. P. S., FRANCO, R. M., OLIVEIRA, L. A. T., SANTOS, E. B.,
GOUVÊA R. Ocorrência de Listeria spp. em amostras de carne bovina
moída comercializadas no município de niterói, RJ, Brasil. Ciências
Agrotecnologia, Lavras, v. 31, n. 4, p. 1225-1230, 2007.

24
30 MARZOCCA M.A., MARUCCI P.L., SICA M.G., ALVAREZ E.E. Detección
de Listeria monocytogenes en distintos productos alimenticios y en
muestras ambientales de una amplia cadena de supermercados de la
ciudad de Bahía Blanca (Argentina). Revista Argentina de Microbiologia.
Buenos Aires [online], v.36, n.4, 2004. Disponível em:
http://www.scielo.org.ar/scielo.php?pid=S0325-
75412004000400006&script=sci_arttext. Acesso em: 25/03/2009.
31 MCLAUCHLIN, J., MITCHELLB, R.T. SMERDONC, W.J JEWELLD K.
Listeria monocytogenes and listeriosis: a review of hazard characterisation
for use in microbiological risk assessment of foodsInternational. Journal of
Food Microbiology, Ithaca, v. 92 p. 15–33, 2004.
32 MELLO, J. F; EINSFELDT, K.; FRAZZON, A. P. G.; COSTA, M.;
FRAZZON, J. Molecular analysis of the iap gene of listeria monocytogenes
isolated from cheeses in Rio Grande do Sul, Brazil. Brazilian Journal of
Microbiology, São Paulo, v. 3, p. 169-172, 2008.
33 MENA, C., ALMEIDAA, G., CARNEIROA, L., TEIXEIRA, P., HOGGA, T.,
GIBBSA, P. A. Incidence of Listeria monocytogenes in different food
products commercialized in Portugal. Food Microbiology, London, v. 21,
p. 213–216, 2004.
34 MISHRA, N. C., BLOBEL, G. Introduction to Proteomics: Principles
and Applications. Ed. Wiley, 2010, p. 200.
35 NALÉRIO, E S., ARAÚJO, M. R., MENDONÇA, K. S., BASSANI, M. T.,
SILVA, W. P. Listeria monocytogenes: monitoramento desse perigo
biológico na cadeia produtiva de frangos do sul do Rio Grande do Sul.
Ciências Tecnologia de Alimentos, Campinas, v. 29, n. 3, p. 626-630,
2009.
37 NBCI. National Center for Biotechnology Information. Complete microbial
genome, 2011. Disponível em:
http://www.ncbi.nlm.nih.gov/genomes/lproks.cgi. Acesso em 17/08/2011.

25
38 OLIVEIRA, M. M. M.; BRUGNERA, D. F.; ALVES, E.; PICCOLI, R. H.
Biofilm formation by Listeria monocytogenes on stainless steel surface and
biotransfer potential. Brazilian Journal of Microbiology, São Paulo, v. 41,
p. 97-106, 2010.
39 PESSOA JÚNIOR, A.; KILIKIAN, B. V. Purificação de produtos
biotecnológicos. Barueri: Manole, 2005. 444 p.
40 ORSI R. H.,. SUN. Q. WIEDMANN M. Genome-wide analyses reveal
lineage specific contributions of positive selection and recombination to the
evolution of Listeria monocytogenes. BMC Evolutionary Biology 2008,
8:233-254
41 Reis, C. M. F., BARBOSA, A. V., RUSAK, L. A., VALLIM D. C., HOFER, E.
Antimicrobial susceptibilities of Listeria monocytogenes human strains
isolated from 1970 to 2008 in Brazil. Revista da Sociedade Brasileira de
Medicina Tropical, v.44, n.2, p.173-176, 2011.
42 RIVOAL K., QUÉGUINER S., BOSCHER E., BOUGEARD S., ERMEL G.,
SALVAT G., FEDERIGHI M., JUGIAU F., PROTAIS, J. Detection of Listeria
monocytogenes in raw and pasteurized liquid whole eggs and
characterization by PFGE. International Journal of Food Microbiology,
Amsterdam, v. 138, p. 56–62, 2010.
43 ROBERTS, A., NIGHTINGALE, K., JEFFERS, G., FORTES, E., KONGO, J.
M., WIEDMANN, M. Genetic and phenotypic characterization of Listeria
monocytogenes lineage III. Microbiology (2006), 152, 685–693
44 ROCHA. L., COSTA. P. H. A., MAGALHÃES, J. C. C., EVARISTO , R. G.
S., VASCONCELOS, E. A. R., COUTINHO, M. V., PAES N. S., SILVA, M.
C. M., GROSSI-DE-SÁ, M. F. Comunicado técnico. Eletroforese
bidimensional e análise de proteomas. Comunicado técnico, Embrapa,
Brasília, p. 1-12, 2005.
45 ROSSI, L. P.R., ALMEIDA, R. C.C., LOPES, L. S., FIGUEIREDO, A. C.L.,
RAMOS, M. P.P., ALMEIDA, P. F. Occurrence of Listeria spp. in Brazilian

26
fresh sausage and control of Listeria monocytogenes using bacteriophage
P100. Food Control, Guildford, v. 22, p. 954-958, 2011.
46 SANTOS, P. M., TEIXEIRA, M. C., SÁ-CORREIA, I. A Análise Proteómica
quantitativa na Revelação de Mecanismos de Resposta a stresse químico
em microrganismos: Métodos em Biotecnologia - Proteômica Quantitativa.
Boletim de biotecnologia, São Paulo, n. 77 p. 7-19, 2004.
47 SHIN JH, KIM J, KIM SM, KIM S, LEE JC, AHN JM, CHO JY. Sigma B-
dependent protein induction in Listeria monocytogenes during vancomycin
stress. FEMS microbiology letters, Amsterdam, v. 308, n. 1, p. 94-100,
2010.
48 SILVA, A. M. S., CORREA, G. C., REIS, E. M. PROTEOMICA - UMA
ABORDAGEM FUNCIONAL DO ESTUDO DO GENOMA. Saúde &
Ambiente em Revista, Duque de Caxias, v.2, n.2, p. 01-10, 2007.
49 SILVA, N., JUNQUEIRA, V. C. A., SILVEIRA, N. F. A., TANIWAKI, M. H.,
SANTOS, R. F. S., GOMES, R. A. R. Manual de métodos de análise
microbiológica de alimentos. 3. ed, São Paulo: Livraria Varela, p.536, 2007.
50 VELDE, V. S., DELAIVE, E., DIEU, M., CARRYN, S., VAN, B. F.,
DEVREESE, B., RAES, M., TULKENS, P.M. Isolation and 2-D-DIGE
proteomic analysis of intracellular and extracellular forms of Listeria
monocytogenes. Proteomics, Weinheim, v. 9, n. 24, p. 5484-5496, 2009.