abundÂncia e sucessÃo de palythoa caribaeorum - repositório institucional da … ·...
TRANSCRIPT

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS APLICADAS E EDUCAÇÃO LITORAL NORTE – RIO TINTO/PB
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E MONITORAMENTO AMBIENTAL
ABUNDÂNCIA E SUCESSÃO DE Palythoa caribaeorum
(DUCHASSAING E MICHELOTTI, 1860): UM ESTUDO
EXPERIMENTAL NO BANCO DOS ABROLHOS, BA
PATRÍCIA SILVA BASÍLIO
RIO TINTO-PARAÍBA
2013

ii
PATRÍCIA SILVA BASÍLIO
ABUNDÂNCIA E SUCESSÃO DE Palythoa caribaeorum
(DUCHASSAING E MICHELOTTI, 1860): UM ESTUDO EXPERIMENTAL NO
BANCO DOS ABROLHOS, BA
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia e Monitoramento Ambiental da Universidade Federal da Paraíba, como requisito parcial para a obtenção do título de Mestre.
Orientador: Dr. Ronaldo Bastos Francini Filho
RIO TINTO-PARAÍBA
2013

iii
PATRÍCIA SILVA BASILIO
ABUNDÂNCIA E SUCESSÃO DE Palythoa caribaeorum
(DUCHASSAING E MICHELOTTI, 1860): UM ESTUDO EXPERIMENTAL NO
BANCO DOS ABROLHOS, BA
BANCA EXAMINADORA
___________________________________________________
Dr. Ronaldo Bastos Francini Filho (Orientador)
____________________________________________________
Dra. Adriane Pereira Wandeness (membro interno - titular)
____________________________________________________
Dra. Cristiane Francisca da Costa (membro externo – titular)
____________________________________________________
Dra. Fabiana Cézar Félix Hackradt (membro externo - suplente)
____________________________________________________
Dr. George Emmanuel Cavalcanti de Miranda (membro interno - suplente)

iv
AGRADECIMENTOS
A todos que contribuíram com este trabalho;
Aos meus familiares, em especial à minha Mãe (Edeilta Silva Basílio) e ao meu
irmão (Daniel Silva Basílio);
Ao meu noivo, que sempre esteve ao meu lado e a quem eu dedico meu amor,
Rafael dos Santos Dantas;
À todos os amigos, professores e coordenadores do Programa De Pós-Graduação
Em Ecologia E Monitoramento Ambiental, assim como aos do Programa De Pós-
Graduação Em Ciências Biológicas da Universidade Federal da Paraíba;
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pela
bolsa de mestrado concedida;
À equipe da Conservação Internacional do Brasil em Caravelas, pelo suporte
logístico e todos que participaram diretamente na coleta de dados;
Ao meu orientador, Dr. Ronaldo Bastos Francini-Filho pelas oportunidades e
conhecimentos repassados;
Meus sinceros agradecimentos.

v
SUMÁRIO
LISTA DE FIGURAS ............................................................................................... VII
LISTA DE TABELAS .............................................................................................. VIII
RESUMO .................................................................................................................. IX
ABSTRACT ............................................................................................................... X
1. INTRODUÇÃO .................................................................................................... 12
2. OBJETIVOS ....................................................................................................... 20
2.1. Objetivos gerais ............................................................................................. 20
2.2. Objetivos específicos..................................................................................... 20
3. MATERIAL E MÉTODOS ................................................................................... 22
3.1. Área de Estudo .............................................................................................. 22
3.2. Delineamento amostral.................................................................................. 27
3.2.1. Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos e a influência de variáveis bióticas e abióticas ........................................................................................................ 27
3.2.2. Experimento de exclusão de Palythoa caribaeorum ..................................... 32
a. Análise de recolonização e vitalidade de Palythoa caribaeorum ................... 35
b. Análise da sucessão bentônica nas áreas experimentais ............................. 37
3.3. Análises estatísticas ........................................................................................... 38
3.3.1. Variações espaciais e temporais na cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos e a influência de variáveis bióticas e abióticas. ....................................................................................................... 38
3.3.2. Experimento de exclusão de Palythoa caribaeorum ..................................... 38
a. Recolonização e vitalidade da Palythoa caribaeorum ................................... 38
b. Sucessão de organismos bentônicos ............................................................ 39
4. RESULTADOS ................................................................................................... 41
4.1. Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos e a influência de variáveis bióticas e abióticas ........................................................................................................ 41
4.2. Experimento de exclusão .............................................................................. 44
4.2.1. Recolonização da Palythoa caribaeorum ...................................................... 44
4.2.2. Variação na vitalidade de Palythoa caribaeorum .......................................... 51
4.3 Sucessão de organismos bentônicos ............................................................ 56
5. DISCUSSÃO ....................................................................................................... 61

vi
5.1 Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos e a influência de variáveis bióticas e abióticas ........ 61
5.2 Experimento de exclusão: Recolonização e vitalidade da Palythoa caribaeorum. ................................................................................................. 62
5.3 Sucessão de organismos bentônicos ............................................................ 69
6. CONCLUSÃO ..................................................................................................... 76
7. REFERÊNCIAS .................................................................................................. 77

vii
LISTA DE FIGURAS
FIGURA 1. MAPA DO BANCO DOS ABROLHOS MOSTRANDO OS SÍTIOS ESTUDADOS E ÁREAS MARINHAS PROTEGIDAS. A) RECIFES ITACOLOMIS, B) TIMBEBAS, C) PARCEL DAS PAREDES, D) SEBASTIÃO GOMES, E) PARCEL DOS ABROLHOS E F) ARQUIPÉLAGO DOS ABROLHOS. ................................................................... 25
FIGURA 2. MERGULHADOR COLETANDO DADOS SOBRE COBERTURA DO SUBSTRATO ATRAVÉS DO MÉTODO DE FOTO-QUADRADO (CF. FRANCINI-FILHO ET AL., 2013). FOTO: PABLO PITA ORTUNDA. .................................................................... 28
FIGURA 3. PAINEL DE ANÁLISES DO PROGRAMA CORAL POINT COUNT WITH EXCEL EXTENSIONS 4.1 (CPCE) (KOHLER E GILL, 2006). CADA LETRA REPRESENTA UM DOS 20 PONTOS ALEATÓRIOS UTILIZADOS NA IDENTIFICAÇÃO DOS ORGANISMOS (IMAGEM AMPLIFICADA). ........................... 29
FIGURA 4. METODOLOGIA DE CENSO VISUAL ESTACIONÁRIO (CF. MINTE-VERA ET AL., 2008). NO RAIO DE DOIS METROS SÃO CONTABILIZADOS OS PEIXES MENORES QUE 10 CM CT E NO RAIO DE QUATRO METROS SÃO CONTABILIZADOS INDIVÍDUOS >10 CM CT. .......................................................... 30
FIGURA 5. PEIXES PREDADORES DE PALYTHOA CARIBAEORUM REGISTRADOS NO BANCO DOS ABROLHOS, BA: A) ABUDEFDUF SAXATILIS, B) ALUTERUS SCRIPTUS, C) CANTHERHINES MACROCERUS, D) CHAETODON STRIATUS, E) CHAETODON SEDENTARIUS, F) HOLACANTHUS CILIARIS, G) POMACANTHUS ARCUATUS E H) POMACANTHUS PARU. FOTOS: R.B. FRANCINI-FILHO. ..................................................................................................... 31

viii
LISTA DE TABELAS
TABELA 1. CARACTERÍSTICAS AMBIENTAIS E BIOMASSA DE PREDADORES DE ZOANTÍDEOS NOS SÍTIOS ESTUDADOS. .............................................................. 26
TABELA 2. CARACTERIZAÇÃO TEMPORAL DAS COLETAS REALIZADAS DURANTE O EXPERIMENTO DE EXCLUSÃO DE PALYTHOA CARIBAEORUM. T0: DADOS DE COBERTURA OBTIDOS ANTES DA RETIRADA DA PALYTHOA CARIBAEORUM, T1: DADOS DE COBERTURA COLETADOS IMEDIATAMENTE APÓS A RETIRADA DA PALYTHOA CARIBAEORUM. OS DEMAIS TEMPOS SÃO REFERENTES AOS DIAS PASSADOS DESDE O INICIO DO EXPERIMENTO (I.E. RETIRADA DA P.CARIBAEORUM). .......................................................................... 33
TABELA 3. ANÁLISE DE VARIÂNCIA (ANOVA) FATORIAL TESTANDO O EFEITO DOS FATORES SÍTIO E ANO NA COBERTURA RELATIVA DE PALYTHOA CARIBAEORUM NO BANCO DOS ABROLHOS. ...................................................... 42
TABELA 4. RESULTADO DA REGRESSÃO MÚLTIPLA MOSTRANDO A INFLUÊNCIA RELATIVA DA DISTANCIA DA COSTA, PROFUNDIDADE, LATITUDE E BIOMASSA DE PEIXES PREDADORES DE ZOANTÍDEOS NA COBERTURA DE PALYTHOA CARIBAEORUM. ................................................................................... 44
TABELA 5. ANÁLISE DE VARIÂNCIA (ANOVA) FATORIAL TESTANDO OS EFEITOS DE SÍTIO, TEMPO, RASPAGEM (RASPADO OU NÃO RASPADO) E TRATAMENTO (COM GAIOLA, COM GAIOLA ABERTA E SEM GAIOLA) NA COBERTURA DE PALYTHOA CARIBAEORUM. ................................................................................... 45
TABELA 6. TAXAS DE CRESCIMENTO DA P.CARIBAEORUM (CM².DIA-1) PARA CADA TRATAMENTO NAS ÁREAS MANIPULADAS E CONTROLE,NOS SÍTIOS PAB4 E ARENG. .................................................................................................................... 47
TABELA 7. ANÁLISE DE VARIÂNCIA (ANOVA) FATORIAL TESTANDO O EFEITO DO TEMPO, RASPAGEM E TRATAMENTO NA COBERTURA DE PALYTHOA CARIBAEORUM EM ARENG. ............................................................................................................. 48

ix
RESUMO
Os recifes de corais são as maiores estruturas biológicas dos oceanos, formados, principalmente, por organismos construtores (ex. corais escleractíneos e algas calcárias). Nas ultimas cinco décadas, foram registrados declínios acentuados na cobertura de corais em todo o mundo. Sobrepesca, poluição e aumento da sedimentação costeira são apontados como os principais impactos responsáveis por esses declínios e pelo aumento na abundância de organismos não construtores (e.g. algas e zoantideos), processo conhecido como “mudança de fase”. O zoantídeo Palythoa caribaeorum é abundante no Atlântico Sul, particularmente no Brasil. Ele é capaz de tolerar alta variabilidade ambiental e apresenta a maior taxa de crescimento entre os antozoários, além de estratégias elaboradas de competição por espaço, as quais garantem sua superioridade competitiva sobre a maioria de outros invertebrados bentônicos recifais. O presente trabalho foi realizado no Banco dos Abrolhos, BA, e tem como objetivos principais: 1) descrever os padrões espaço-temporais de cobertura de P. caribaeorum ao longo do Banco dos Abrolhos, 2) avaliar a influência da distância da costa, profundidade, latitude e biomassa de peixes predadores de zoantídeo na abundância de P. caribaeorum, 3) descrever a sucessão bentônica em áreas nas quais P. caribaeorum foi retirada. O estudo dos padrões espaço-temporais foi conduzido em seis
recifes de 2006 a 2008: Parcel dos Abrolhos (cinco sítios), Arquipélago (cinco sítios), Timbebas (três sítios), Itacolomis (oito sítios), Parcel das Paredes (dois sítios) e Sebastião Gomes (um sítio). O experimento de exclusão foi realizado de 2006 a 2009 no topo dos sítios Arenguera (Parcel das Paredes) e PAB4 (Parcel dos Abrolhos). Em cada sitio foram selecionadas quatro áreas de 5 x 5 m, sendo duas manipuladas (cobertura de P. caribaeorum retirada). Três tratamentos foram aplicados dentro de cada área: gaiola
fechada, gaiola aberta e sem gaiola. Os métodos utilizados foram foto-quadrado (caracterização do bentos) e censo visual estacionário (quantificação de peixes). Os resultados mostraram que P. caribaeorum domina no topo dos recifes, com coberturas insignificantes na parede, demonstrando a preferência de P. caribaeorum por ambientes mais iluminados e horizontais. A cobertura relativa de P. caribaeorum foi maior nos sítios PAB4, PAB5, TIM2, TIM3, ARENG e SG, tendo sido influenciada positivamente pela profundidade e negativamente pela distância da costa. A biomassa de predadores variou entre os sítios mas não influenciou significativamente na distribuição da P. caribaeorum. As cinco espécies de predadores de P. caribaeorum mais abundantes em termos de biomassa foram Abudefduf saxatilis, Pomacanthus paru, Pomacanthus arcuatus, Holacanthus ciliares e Chaetodon striatus. No experimento de exclusão foi registrada recuperação lenta de P. caribaeorum na área manipulada, não sendo registrada diferença nos padrões de recolonização entre os dois sítios (próximo e afastado da costa). Um dos fatores que pode ter inibido o assentamento da P. caribaeorum na área manipulada foi a proliferação de algas
epilíticas, principalmente dentro das gaiolas fechadas, fato conhecido como uma das primeiras fases de degradação nos ambientes recifais. A taxa de recolonização (i.e. assentamento seguido de crescimento) foi maior em ARENG (0,23 cm-2.dia-1) do que em PAB4 (0,12 cm-2.dia-1) na área manipulada. Entretanto, na área controle, PAB4 apresentou taxa de recuperação de 3,18 cm-2.dia-1 enquanto que em ARENG a área de P. caribaeorum
declinou (-0,25 cm2.dia-1). Pode-se concluir: 1) variáveis ambientais, como profundidade e distância da costa, são mais importantes do que variáveis bióticas (i.e predação) na dinâmica de P. caribaeorum, 2) apesar da cobertura de P. caribaeorum apresentar relação negativa com a distância da costa, as taxas de recolonização não foram maiores no sítio mais próximo a costa, 3) a cobertura de P. caribaeorum aumenta em períodos próximos ao verão, em ambos os sítios e 3) a baixa densidade de Palythoa foi desfavorável para um processo mais rápido de recolonização. Palavras chave: Zoantídeos; sucessão bentônica; recifes de corais; sedimentação.

x
ABSTRACT
Coral reefs are recognized as the largest biological structures in the oceans. They formed by building organisms (e.g. scleractinian corals and calcareous algae). In the last five decades, there have been declines in coral cover worldwide. Overfishing, pollution and increased coastal sedimentation are pointed as the main impacts responsible for declines in abundance of reef-building organisms and increases in abundance of non-building ones (e.g. algae), a process known as "phase shift". The zoanthid Palythoa caribaeorum is abundant in the
Atlantic, particularly in Brazil. It is able to tolerate high environmental variability and displays the highest growth rate among all Anthozoans. Moreover, it has elaborated strategies of competition for space, which makes it competitively superior to most other benthic invertebrates. The present work was performed in the Abrolhos Bank, Bahia, with the following main objectives: 1) to describe the spatio-temporal patterns in cover of P. caribaeorum across the Abrolhos Bank, 2) evaluate the relative influence of distance offshore, depth, latitude and biomass of fish zoanthid predators in the abundance of P. caribaeorum; 3) describe the benthic succession in two areas in which P. caribaeorum was excluded (one next to the coast and subject to high sedimentation levels and another far from the coast and subject to low sedimentation). The study of spatio-temporal patterns was conducted in six reefs from 2006 to 2008: Parcel dos Abrolhos (five sites), Archipelago (five sites), Timbebas (three sites), Itacolomis (eight sites), Parcel das Paredes (two sites) and Sebastião Gomes (one site). The exclusion experiment was conducted from 2006 to 2009 in the top of the reefs of sites Arenguera (Parcel das Paredes) and PAB4 (Parcel dos Abrolhos). Four 5 x 5 m areas were selected at each site. Two of them were manipulataded (all P. caribaeorum removed) and the other two were not manipulated (control). Three
treatments were applied within each area: 1) closed cage, to exclude reef fish known to predate P. caribaeorum, 2) open cage (control for the effect of the cage), 3) without cage. The methods used were photo-quadrats (benthic assessments) and stationary visual census (fish assessments). The results showed that P. caribaeorum dominates the top of the reefs, with insignificant coverage in the wall. This suggests the preference of P. caribaeorum for horizontal inclination and high light levels. The relative cover of P. caribaeorum was higher in
the sites PAB4, PAB5, TIM2, TIM3, ARENG and SG, and was positively influenced by depth and negatively influenced by distance offshore. The five most abundant reef fish species recognized as predatos of zoanthids in terms of biomass were A. saxatilis, P. paru, P. arcuatus, H. ciliares, and C. striatus. In the exclusion experiment, a slow recovery of P. caribaeorum was recorded in the manipulated areas, with no differences in recolonization rates between the two sites (near or away from the coast). Settlement of P.caribaeorum at the manipulated areas may have been inhibited by the proliferation iof fast growing epilithic algae, particularly inside closed cages. The overall average growth rate was higher in ARENG (0.23 cm-2.day-1) than in PAB4 (0.12 cm-2.day-1) in the manipulated area. However, in the control area of PAB4 P. caribaeorum showed a growth rate of 3.18 cm-2.day-1, while in the control area of ARENG it declined (-0.25 cm-2.day-1). Total coverage of P. caribaeorum
declined in the control area of ARENG due to a bleaching event that occurred throughout the study. Conclude that: 1) environmental variables such as depth and distance from the coast, are more important than biotic (eg predation) in the dynamics of P. caribaeorum, 2) despite the coverage of P. caribaeorum present negative relationship with distance from the coast, recolonization rates were not higher at the site closest to shore, 3) coverage of P. caribaeorum increases in periods near the summer at both sites, and 3) low density of Palythoa was unfavorable for a faster process of recolonization.
Keywords: Zoanthids; benthic succession; coral reefs, sedimentation.

11
1. INTRODUÇÃO

BASILIO, P. S. 12
1. INTRODUÇÃO
Os recifes de corais são altamente complexos e diversificados,
apresentando fundamental importância ecológica e econômica e sendo
reconhecidos como as maiores estruturas biológicas dos oceanos (CONNELL,
1978). Eles funcionam como barreira de proteção física para as praias e como fonte
de recursos e renda para milhões de pessoas em todo o mundo (MOBERG E
FOLKE, 1999; SPALDING et al., 2001).
Ambientes recifais podem ser definidos tanto por sua estrutura física quanto
pelos organismos que neles se encontram (SPALDING et al., 2001; BELLWOOD et
al., 2004). Os recifes de corais são estruturas tridimensionais dominadas por corais
escleractíneos e outros organismos construtores como algas calcárias (QUINN,
1982; GHERARDI E BOSENCE, 2001; BELLWOOD et al., 2004). Os corais
escleractínios estão geralmente presentes em águas rasas e de baixa turbidez, isso
devido à dependência de microalgas fotossintetisantes simbiontes (zooxantelas) à
luz (TITLYANOV E TITLYANOVA, 2002).
A complexidade estrutural (topográfica) do substrato recifal, como o número
e tamanho das fendas, arquitetura e inclinação do mesmo (AULT E JOHNSON,
1998; GRATWICKE E SPEIGHT, 2005), representam fatores de fundamental
importância na estruturação, funcionamento, resiliência e dinâmicas de comunidades
recifais (HUGHES et al., 2003; ALVAREZ-FILIP et al., 2009).
A medida que os organismos construtores morrem, são disponibilizados
novos espaços para colonização, tanto para organismos construtores quanto para
não construtores (e.g. algas e zoantídeos; SEBENS, 1982; BELLWOOD et al.,
2004). O equilíbrio entre a abundância de organismos construtores e não-
construtores é tratado como parâmetro para determinação das fases de degradação
dos recifes (HUGHES, 1994; BELLWOOD et al., 2004, FRANCINI-FILHO et al.,
2013).
Nas ultimas cinco décadas foram registrados declínios acentuados na
cobertura de corais em todo o mundo, aumentando assim a preocupação com os
possíveis efeitos deletérios e permanentes a estes ecossistemas (GARDNER et al.,
2003; PANDOLFI et al., 2003; BELLWOOD et al., 2004; BRUNO E SELIG, 2007).

BASILIO, P. S. 13
Os principais distúrbios que contribuem com o declínio dos recifes são os
seguintes: 1) sobrepesca (JACKSON et al., 2001; SCHEFFER et al., 2005; SANDIN
et al., 2008), 2) aquecimento e acidificação da água superficial do mar (DONNER et
al., 2005; HOEGH-GULDBERG et al., 2007; DE’ATH et al., 2009), 3) poluição
costeira, particularmente a descarga de nutrientes (PASTOROK E BILYARD, 1985;
LOYA E KRAMARSKY-WINTER, 2003; BAUMAN et al., 2010), 4) degradação de
habitat (HUGHES, 1994; PANDOLFI et al., 2003) e sedimentação (LOYA, 1976;
SEGAL et al., 2008; RIEGL E BRANCH, 1995; PHILIPP E FABRICIUS, 2003;
FABRICIUS, 2005). Este último pode ser causado tanto por ressuspensão de
sedimentos do fundo marinho quanto pelo escoamento de águas continentais
(WEBER et al., 2012).
Além de fatores naturais, como chuvas intensas e tempestades, a ação
antrópica vem favorecendo o aumento da sedimentação em áreas recifais,
principalmente através da ocupação de regiões costeiras e dragagens de estuários
(VAN KATWIJK et al.,1993). Os impactos ocasionados pelo desenvolvimento
costeiro interferem diretamente na estrutura, distribuição (YOSHIOKA E YOSHIOKA,
1989; DUTRA, 2003) e vitalidade dos organismos recifais bentônicos (FABRICIUS et
al., 2003). Os efeitos do elevado aporte de sedimento em corais incluem a remoção
de colônias (WOOLFE E LARCOMBE, 1999), morte nos estágios iniciais de vida
(FABRICIUS et al., 2003), redução de taxas de crescimento (ABDEL-SALAM E
PORTER, 1988; ROGERS, 1990), lesões no tecido vivo (ROGERS, 1990), inibição
do recrutamento (TE, 1992), asfixia (ABDEL-SALAM E PORTER, 1988) e redução
da capacidade fotossintetizante das zooxantelas simbiontes (WEBER et al., 2012).
Taxas de sedimentação acima de 10 mg.cm-2.dia-1 são consideradas
limitantes para vitalidade e sobrevivência de corais (ROGERS, 1990; DUTRA et al.
2006). Portanto, para garantir a sobrevivência, muitos cnidários assumem
estratégias de zonação (e.g. preferência por habitats com menor acúmulo de
sedimentos) e adotam formas ramificadas que dificultam o acúmulo e facilitam a
remoção de sedimento da superfície da colônia (ROGERS, 1990).
Existe uma relação íntima entre a mortalidade dos corais e a presença de
microrganismos associados ao sedimento (HODGSON, 1990; WEBER et al.2012).
Em experimento realizado em laboratório, Weber et al. (2012) mostraram que os
componentes microbianos no sedimento, através de processos bioquímicos, liberam

BASILIO, P. S. 14
compostos que degradam os tecidos dos corais pela redução do oxigênio e pH. No
entanto, cada espécie reage de maneira diferente à sedimentação, a qual afeta
principalmente espécies ineficientes em rejeitar sedimento e com baixa capacidade
de sobreviver em ambientes sombreados (ROGERS, 1990; KLEYPAS, 1996). A
capacidade de remoção de sedimento da superfície corpórea dos corais está
relacionada ao tamanho dos pólipos, com espécies que possuem pólipos maiores
apresentando maior capacidade de remoção de sedimentos (HUBBARD E POCOCK
1972; LEÃO et al., 2003).
Os ecossistemas estão adaptados para responder a mudanças naturais em
condições ambientais (CONNEL et al., 1997; SCHEFFER, 2001). Acredita-se que no
Atlântico Sul Ocidental os corais evoluíram em condições naturais de alta turbidez,
sendo assim considerados tolerantes a níveis relativamente altos de sedimentação
(LEÃO et al., 2003). No entanto, com o aumento da sedimentação em áreas recifais,
devido a desmatamento costeiro (LEÃO E DOMINGUEZ, 2000; LEÃO et al., 2003,
SILVA et al., No prelo), sobrecarga de nutrientes (COSTAJr et al., 2000), destruição
de habitats e redução de espécies de peixes herbívoros (FRANCINI-FILHO E
MOURA, 2008b), esses ambientes iniciam um processo de redução da capacidade
de recuperação após perturbação, abrindo caminho para um estado de mudança de
fase (da dominância de corais para dominância de algas e/ou outros organismos não
construtores) (DONE, 1992; SCHEFFER, 2001; BELLWOOD et al., 2004;
NORSTRÖM et al., 2009; FRANCINI-FILHO et al., 2013).
Cnidários da ordem Zoantharia (Anthozoa: Hexacorallia) são comuns em
recifes rasos (SUCHANEK E GREEN, 1981), tanto em áreas entre-marés como em
áreas permanentemente submersas, e em recifes tropicais e subtropicais (SEBENS,
1982). Zoantídeos são responsáveis por grande parte do fluxo de energia em
ambientes costeiros (GILI E COMA, 1998). No entanto, eles podem causar efeitos
deletérios, como a diminuição da complexidade estrutural do substrato (HAYWICK E
MUELLER, 1997; MENDONÇA-NETO et al., 2008) e deslocamento competitivo de
outras espécies (SUCHANEK E GREEN, 1981).
A ordem Zoantharia é formada por organismos não-construtores (i.e. sem
esqueleto de CaCO3) que tipicamente apresentam dois ciclos de tentáculos e
incorporam partículas de sedimento no tecido (SINNIGER et al., 2005). Existem
cerca de 360 espécies de zoantídeos reconhecidas em todo o mundo (FAUTIN,

BASILIO, P. S. 15
2006). No entanto, este número não é confiável, pois a taxonomia desse grupo tem
sido historicamente considerada caótica, principalmente devido à limitação na
utilização de dados morfológicos (BURNETT et al., 1997; SINNIGER et al., 2005;
REIMER et al., 2006a; REIMER et al., 2010). Um exemplo emblemático é o
zoantídeo “gold coral” Kulamanamana haumeaae, endêmico do Havaí. Esta espécie
forma um esqueleto esclero-protéico e tem sido explorada para a confecção de jóias
desde a década de 70. No entanto, ela foi descrita formalmente pela ciência apenas
em 2013 (SINNIGER et al., 2013).
Zoantídeos coloniais do gênero Palythoa podem discriminar características
dos sedimentos incorporados em seus tecidos, tais como composição mineral, forma
dos grãos e tamanho do sedimento, com preferência por grãos ≤ 125 µm, tamanho
menor do que aquele de grãos encontrados em zonas de alta energia (HAYWICK E
MUELLER, 1997).
Palythoa caribaeorum (DUCHASSAING E MICHELOTTI, 1860) é uma
espécie colonial pertencente à família Sphenopidae Herteig, 1882. Esta espécie é
comum em ambientes recifais ao longo do Atlântico ocidental, onde ocorre desde o
sul da Flórida, EUA, até Santa Catarina, Brasil (SUCHANEK E GREEN, 1981;
MAIDA E FERREIRA, 1997; KEMP et al., 2006), sendo particularmente abundante
na costa e ilhas oceânicas brasileiras (MAIDA E FERREIRA, 1997; FRANCINI-
FILHO et al., 2013). Palythoa caribaeorum ocorre em profundidades máximas de
cerca de 15 m, provavelmente devido a sua simbiose com zooxantelas dependentes
de luz (SEBENS, 1977, 1982), geralmente dominando ambientes com elevado
hidrodinamismo (HAYWICK E MUELLER, 1997).
Zoantídeos coloniais como P. caribaeorum são importantes em ambientes
recifais, pois contribuem com a produção primária e servem como recurso alimentar
para diversas espécies (SUCHANEK E GREEN, 1981; SEBENS, 1982; STAMPAR
et al., 2007; FRANCINI-FILHO E MOURA, 2010; LONGO et al., 2012). Eles também
podem ser úteis na produção de fármacos (SOARES et al., 2006), além de agir
como indicadores de eventos de branqueamento (i.e. perda de zooxantelas
simbiontes devido a distúrbios ambientais), já que estão entre um dos primeiros a
sofrer com este fenômeno (WILLIAMS E BUNKLEY-WILLIAMS, 1988).
A obtenção de energia por P. caribaeorum pode ocorrer de duas formas: 1)
através de simbiose com dinoflagelados viventes em seu tecido gastrodérmico

BASILIO, P. S. 16
(zooxantelas), os quais fornecem açucar e outros nutrientes em em troca de abrigo
(SEBENS, 1977) e 2) por heterotrofia, capturando partículas orgânicas na coluna
d’água, principalmente durante o entardecer e o amanhecer (SEBENS, 1977, 1982).
Através desta primeira estratégia, P. caribaeorum obtém principalmente fosfatos,
nitrogênio e outros nutrientes que não são produzidos pelas zooxantelas (cf.
SEBENS, 1977; TANNER, 2002).
A captura e encaminhamento do alimento a boca é realizado pelos
tentáculos, presentes em pólipos que permanecem unidos firmemente pelo
cenénquima (estrutura formada pela mesoglea expandida) (BOSCOLO E SILVEIRA,
2005; SEBENS, 1977). Cada pólipo pode medir entre 5 e 10 mm de diâmetro
(BOSCOLO ESILVEIRA, 2005), com essa variação de tamanho estando
potencialmente ligada ao tipo do habitat (SEBENS, 1982). Os pólipos formam um
tapete que geralmente cobre grandes áreas (MEDONÇA-NETO E GAMA, 2009) e
serve de proteção e refúgio para diversos organismos (PÉREZ et al., 2005).
Sebens (1982) mostrou através de um estudo experimental que colônias de
P. caribaeorum, fora d’água, sofrem danos parciais ou morte após exposição
contínua de cinco horas ao sol e sete horas à sombra. Ao contrário de outros
zoantídeos, P. caribaeorum não tem capacidade de reter água entre seus pólipos.
Portanto, uma das estratégias de proteção desta espécie é produzir muco que
protege a colônia contra a dessecação durante a maré baixa. Associadas a este
muco são encontradas substâncias com atividade analgésica (SOARES et al., 2006)
e tóxica (MOORE E SCHEUER, 1971; SUCHANEK E GREEN, 1981), além de
microbiota bastante diversificada (CHIMETTO et al., 2009).
Além de tolerar alta variabilidade ambiental (SEBENS, 1982; KEMP et al.,
2006), P. caribaeorum apresenta a maior taxa de crescimento entre os antozoários
(até 0,4 mm.dia-1) (SUCHANEK E GREEN, 1981, RABELO et al., 2013). Esta
capacidade de crescimento acelerado, associada a estratégias elaboradas de
competição por espaço, garantem a superioridade competitiva de P. caribaeorum
sobre a maioria dos outros invertebrados bentônicos coloniais recifais (SUCHANEK
E GREEN, 1981; PÉREZ et al., 2005; MEDONÇA-NETO E GAMA, 2009).
Palythoa caribaeorum domina espaço matando ou impedindo o crescimento
de seus concorrentes por investidas físicas, com o uso de tentáculos, filamentos
mesentéricos, galgamento e interferência química por alelopatia (SUCHANEK E

BASILIO, P. S. 17
GREEN, 1981; FAUTIN, 2009). Este zoantídeo apresenta uma das toxinas mais
poderosas conhecidas, a Palytoxina, a qual é considerada uma das moléculas
biologicamente ativas mais potentes (MOORE E SCHEUER, 1971; SUCHANEK E
GREEN, 1981; AMIR et al., 1997).
A palytoxina é uma forte arma na competição, proteção e defesa agindo
diretamente no bloqueio da bomba de sódio-potássio, formando canais permeáveis
a cátions na membrana plasmática e promovendo assim a lise celular nos
organismos competidores (FRELIN E VAN RENTERGHEM, 1995). Organismos
recifais bentônicos registrados previamente sendo suprimidos competitivamente por
P. caribaeorum incluem corais escleractínios dos gêneros Favia, Meandrina,
Siderastrea, Porites e Montastrea, além de octocorais, outros zoantídeos, hidrocorais
calcáreos do gênero Millepora (Hidrozoa), poríferos e algas (SUCHANEK E GREEN,
1981; ACOSTA, 2001; PÉREZ et al., 2005; MENDONÇA-NETO E GAMA, 2009).
Palythoa caribaeorum possui ainda elevadas taxas de fissão que ocorrem
durante todo o ano, a qual pode acontecer através de dois processos: 1) fissão da
borda da colônia e liberação do aglomerado de pólipos e 2) liberação de aglomerado
de pólipos após brotamento do tecido (ACOSTA et al., 2005). A reprodução
sexuada também ocorre em P. caribaeorum. Segundo Acosta et al. (2005), no
sudeste do Brasil (costa de São Paulo) ela é periódica, ocorrendo principalmente
entre abril e maio, sendo mais baixa no inverno.
Apesar da toxicidade e a presença de sedimento no tecido de P.
caribaeorum, muitos trabalhos vêm mostrando o consumo deste zoantídeo por
várias espécies, o qual leva ao armazenamento da palytoxina nos tecidos dos
predadores (MOORE E SCHEUER, 1971; FRANCINI-FILHO E MOURA, 2010). São
conhecidos dentre os predadores de P. caribaeorum a tartaruga-de-pente
Eretmochelys imbricata (LINNAEUS, 1766) (STAMPAR et al., 2007), o poliqueta-de-
fogo Hermodice carunculata (PALLAS, 1766) (SUCHANEK E GREEN, 1981;
SEBENS, 1982), várias espécies de ouriços (SEBENS, 1982) e algumas espécies de
peixes recifais no Caribe (SUCHANEK E GREEN, 1981) e no Brasil (FRANCINI-
FILHO E MOURA, 2010; LONGO et al., 2012). Portanto, além da influência de
variáveis ambientais e geográficas na abundância de P. caribaeorum (SEBENS,
1982), a predação também pode ser um fator importante na dinâmica deste
zoantídeo (FRANCINI-FILHO E MOURA, 2010).

BASILIO, P. S. 18
A identificação de processos envolvidos em padrões de competição e
sucessão de organismos bentônicos de ecossistemas recifais depende geralmente
da realização de estudos em longo prazo, devido principalmente à complexidade na
dinâmica desses ambientes (CÔTÉ et al., 2005; SCHUTTE et al., 2010; FRANCINI-
FILHO et al., 2013). Além disso, abordagens experimentais são muitas vezes
necessárias para identificação precisa de processos ecológicos responsáveis pelos
padrões observados (TANNER 1997, 1999).

BASILIO, P. S. 19
2. OBJETIVOS

BASILIO, P. S. 20
2. OBJETIVOS
2.1. Objetivos gerais
Estudar, no Banco dos Abrolhos, Bahia, os padrões espaço-temporais da cobertura
de P. caribaeorum avaliando a influência da distância da costa, profundidade,
latitude e biomassa de peixes predadores de zoantídeos nos padrões de cobertura
observados e descrever e a sucessão de organismos bentônicos através de um
estudo experimental.
2.2. Objetivos específicos
Descrever os padrões espaciais e temporais na cobertura relativa de P.
caribaeorum considerando 25 sítios distribuídos ao longo do Banco dos Abrolhos e
dois habitats (topo e parede; cf. FRANCINI-FILHO et al., 2013) ao longo de três anos
(2006-2008);
Avaliar a influência relativa da distancia da costa, profundidade, latitude
e biomassa de peixes predadores de zoantídeos nos padrões espaciais e temporais
de cobertura relativa de P. caribaeorum;
Descrever os padrões de sucessão bentônica, após experimento de
remoção de colônia de P. caribaeorum, em duas áreas uma próxima a costa e com
elevada sedimentação e outra afastada da costa e com baixa sedimentação. Com
foco na dinâmica de recolonização de P. caribaeorum.
Estudar a dinâmica espacial e temporal na vitalidade de P. caribaeorum
nas áreas experimentais;
Avaliar o efeito da exclusão de peixes predadores de zoantídeos na
cobertura e recolonização de P. caribaeorum nas áreas experimentais;

BASILIO, P. S. 21
3. MATERIAL E MÉTODOS

BASILIO, P. S. 22
3. MATERIAL E MÉTODOS
3.1. Área de Estudo
Este estudo foi conduzido no Banco dos Abrolhos (16º 40’/19º 40’S – 39º
10’/37º20’W), Bahia, o qual cobre uma área de aproximadamente 46.000 km2 em
uma plataforma continental relativamente rasa, raramente excedendo 20 m de
profundidade, que se extende até 300 km de distância da costa (FRANCINI-FILHO
et al., 2008) (Figura 1).
As formações recifais na região são diversas, incluindo: 1) bancos recifais
adjacentes à costa, os quais são geralmente alongados e paralelos à costa, 2)
pináculos biogênicos colunares com topo expandido lateralmente, conhecidos
popularmente como chapeirões e 3) recifes mesofóticos submersos (“afogados”)
com diferentes formas e tamanhos (LEÃO et. al., 2003; FRANCINI-FILHO et al.,
2013; MOURA et al., 2013). Os chapeirões podem atingir 5 a 25m de altura e 20 a
300 m de extensão (LEÃO et. al., 2003; FRANCINI-FILHO E MOURA, 2008a;
FRANCINI-FILHO et al., 2013).
Os principais construtores dos recifes da região são corais escleractíneos
dos gêneros Mussismilia, Siderastrea e Montastraea, além de hidrocorais do gênero
Millepora (LEÃO et al., 2003; FRANCINI-FILHO et al., 2013) e algas calcárias
incrustantes dos gêneros Lithothaminion, Lithophyllum, Sporolithon e Porolithon
(FIGUEIREDO, 1997). A fauna de peixes é composta por cerca de 260 espécies,
das quais 20% são endêmicas do Brasil (MOURA E FRANCINI-FILHO, 2006). O
complexo recifal dos Abrolhos é reconhecido como um hotspot de biodiversidade
marinha, concentrando a mais alta biodiversidade do Atlântico Sul (MOURA, 2000).
A região abriga uma grande variedade de ambientes, e.g. bancos de algas e gramas
marinhas, mangues, recifes de corais e recifes rochosos, estes últimos encontrados
ao redor das ilhas do Arquipélago dos Abrolhos (FRANCINI-FILHO E MOURA,
2008a; AMADO-FILHO et al., 2012; FRANCINI-FILHO et al., 2013; MOURA et al.,
2013).
A precipitação média anual da área é de 1392 milímetros, com temperaturas
médias do ar de 26º C no verão (dezembro a abril) e 23º C no inverno (junho a

BASILIO, P. S. 23
agosto) (DUTRA et al., 2006). Os principais rios que influenciam a região de
Abrolhos são o Rio Jequitinhonha ao norte, Rio Doce ao sul, e os Rios Caravelas e
Peruípe na porção central (MOURA et al., 2011). O Rio Caravelas, apesar de
apresentar a menor drenagem entre os principais rios, representa o segundo maior
sistema estuarino do Nordeste brasileiro, com 66 km2 de área, e é o principal ponto
de partida para o turismo no Parque Nacional Marinho dos Abrolhos e via de acesso
ao terminal de barcaças da Aracruz Celulose, com operações de dragagem
periódicas e descarte de sedimentos próximo aos recifes costeiros (PEREIRA et.al.,
2010; Obs. Pessoal).
Sedimentos terrígenos transportados por rios e sazonalmente
ressuspendidos predominam em áreas costeiras, ao passo que sedimentos
carbonáticos predominam nos recifes mais afastados da costa (Segal et al., 2008).
Os sedimentos terrígenos originam-se através da erosão das falésias e do transporte
de descargas fluviais, sendo caracterizada por uma composição siliciclástica com 30
a 70% de grãos de quartzo, mica raros e lama (DUTRA et al., 2006). O recife de
Timbebas (Figura 1b), apesar de localizado relativamente próximo a costa, média de
17,8 Km, apresenta uma taxa de sedimentação menor do que aquela em outros
recifes costeiros (e.g. Parcel das Paredes, figura 1c), provavelmente pela sua
morfologia pinacular, que favorece a movimentação das ondas, e pela sua posição
mais ao norte da foz do rio Caravelas, contrária a direção das correntes (DUTRA et
al., 2006). Evidências recentes indicam que sedimentos terrígenos estão avançando
para recifes cada vez mais afastados da costa (SILVA et al., No prelo).
Atualmente existem quatro Unidades de Conservação (UCs) que incluem o
meio marinho no Banco dos Abrolhos: 1) a Reserva Extrativista Marinha (RESEX) de
Corumbau, 2) a RESEX de Cassurubá, 3) o Parque Nacional Marinho (PARNAM) de
Abrolhos e 4) a Área de Proteção Ambiental (APA) Ponta da Baleia/Abrolhos. Esta
última engloba recifes desprotegidos contra a pesca (Parcel das Paredes e
Sebastião Gomes), pois não apresenta Plano de Manejo ou qualquer tipo de
regulamentação. Apenas duas destas UCs foram efetivamente implantadas até o
momento, a RESEX de Corumbau (criada em 2000) e o PARNAM de Abrolhos
(criado em 1983), esta última formada por duas áreas descontínuas (Timbebas, mais
próximo à costa, e Arquipélago de Abrolhos e o Parcel dos Abrolhos, mais distantes
da costa), as quais somam 882 km². A RESEX Corumbau possui uma área de 895

BASILIO, P. S. 24
km² e o uso direto de recursos pesqueiros é permitido apenas para as comunidades
locais, existindo também uma área de exclusão total da pesca nos Recifes
Itacolomis, o principal complexo recifal abrangido pela RESEX (FRANCINI-FILHO E
MOURA 2008a, b; MOURA et al., 2009, 2011; FRANCINI-FILHO et al., 2013).

BASILIO, P. S. 25
Figura 1. Mapa do Banco dos Abrolhos mostrando os sítios estudados e áreas marinhas
protegidas. a) Recifes Itacolomis, b) Timbebas, c) Parcel das Paredes, d) Sebastião Gomes, e) Parcel dos Abrolhos e f) Arquipélago dos Abrolhos.

BASILIO, P. S. 26
Tabela 1. Características ambientais e biomassa de predadores de zoantídeos nos sítios
estudados.
RECIFE/SÍTIO Distância da Costa
(km)
Profundidade Média
(m) Latitude
Biomassa de
Predadores (g.m-2)
Recifes Itacolomis (protegido)
AMP1 5,8 1,7 16,909 89,4
AMP2 8,3 1,6 16,912 314,1
Recifes Itacolomis (uso-múltiplo)
B1 5,2 1,5 16,896 31,0
C1 5,2 1,4 16,892 76,7
A1 5,5 1,3 16,900 18,5
B2 7,5 1,5 16,901 72,0
C2 7,8 2,6 16,898 345,1
A2 7,9 2,2 16,905 35,4
Recifes costeiros desprotegidos
Sebastião Gomes (SG) 13,7 3,1 17,910 240,5
Pedra de Leste (PLEST) 14,3 2,3 17,780 135,6
* Arenguera (ARENG) 14,9 5,2 17,660 421,9
PA2 31,4 4,5 17,880 308,9
Recife de Timbebas (protegido)
TIM1 19 4,7 17,482 334,2
TIM2 17,4 5,9 17,478 572,6
TIM3 17 5,6 17,460 485,6
Arquipélago de Abrolhos (protegido)
Paredão Do Farol (FAROL) 58,1 5,8 17,965 150,0
GUARITA 58,1 5,4 17,960 335,9
Mato Verde (MVERDE) 58,1 4,1 17,964 229,1
Porto Norte (PNORTE) 58,1 5,4 17,960 293,7
SIRIBA 58,1 3,1 17,968 650,3
Parcel dos Abrolhos (protegido)
PAB1 63,4 5,9 17,991 401,2
AB2 61,4 7,3 17,983 577,1
PAB3 61,7 6,9 17,998 598,8
* PAB4 61,7 7,7 17,959 324,4
PAB5 60,1 6,6 17,944 585,6
*Realização do experimento de exclusão

BASILIO, P. S. 27
3.2. Delineamento amostral
3.2.1. Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no
Banco dos Abrolhos e a influência de variáveis bióticas e abióticas
Para avaliar variações espaciais e temporais na cobertura relativa de P.
caribaeorum, e a influência de características ambientais e da abundância de peixes
predadores de zoantídeos, foram amostrados 25 sítios distribuídos ao longo da
região de Abrolhos: 1) Parcel dos Abrolhos (dentro da porção mais afastada da costa
do PARNAM; cinco sítios: PAB1, PAB2, PAB3, PAB4 e PAB5), 2) Arquipélago de
Abrolhos (dentro da porção mais afastada da costa do PARNAM; cinco sítios: Siriba,
Portinho Norte, Mato Verde, Guarita e Paredão do Farol), 3) Recifes de Timbebas
(dentro da porção mais costeira do PARNAM; três sítios: TIM1, TIM2 e TIM3), 4)
Recifes Itacolomis, RESEX de Corumbau (seis sítios na área de uso múltiplo: A1,
A2, B1, B2, C1 e C2, e dois sítios na área de proteção integral: AMP1 e AMP2), 5)
Recifes desprotegidos costeiros (Parcel das Paredes; três sítios: Pedra de Leste,
PA2 e Arenguera e Sebastião Gomes; um sítio: SG) (Figura 1 e Tabela 1).
Em cada sitio foram amostrados os habitats topo (mais raso e horizontal) e
parede (mais fundo e vertical). No Arquipélago apenas a “frente recifal” foi
amostrada, não havendo distinção entre topo e parede. Um estudo recente indica
que as assembléias bentônicas do Arquipélago de Abrolhos são mais similares às
assembléias do topo do que da parede dos chapeirões (FRANCINI-FILHO et al.,
2013).
Os dados foram coletados entre os anos de 2006 e 2008. Para a
caracterização da cobertura relativa de P. caribaeorum foi utilizada à metodologia de
foto-quadrados (cf. FRANCINI-FILHO et al., 2013). Nessa metodologia foi utilizado
um retângulo de moldura de PVC com 76 x 88 cm, subdividido em 15 retângulos
menores de 22 x 15 cm cada. São obtidas fotografias em cada subdivisão, gerando
assim 15 imagens que compõem uma amostra (Figura 2). Em cada sítio, foram
obtidos 10 foto-quadrados fixos (delimitados permanentemente por pinos metálicos
fixados ao substrato) em dois habitats (topo e parede), os quais foram distribuídos
ao acaso antes da fixação.

BASILIO, P. S. 28
Figura 2. Mergulhador coletando dados sobre cobertura do substrato através do método de foto-quadrado (cf. FRANCINI-FILHO et al., 2013). Foto: Pablo Pita Ortunda.
A cobertura relativa de organismos bentônicos , observada nas análises das
fotos, foi estimada com a utilização do programa Coral Point Count with Excel
extensions (CPCe) (KOHLER E GILL, 2006), tendo sido alocados 20 pontos
aleatórios por imagem, totalizando assim 300 pontos por amostra (i.e. quadrado).
Abaixo de cada ponto foi identificado o organismo bentônico pelo grupo funcional
(Figura 3).

BASILIO, P. S. 29
Figura 3. Painel de análises do programa Coral Point Count with Excel extensions 4.1 (CPCe) (KOHLER E GILL, 2006). Cada letra representa um dos 20 pontos aleatórios utilizados na identificação dos organismos (imagem amplificada).
Dados sobre a abundância de peixes foram obtidos entre 2006 e 2008 nos
mesmos sítios e habitats mencionados acima. Ao todo, 907 censos visuais
estacionários (CVE) foram obtidos (entre 10 e 15 censos por habitat / síto / ano),
conforme metodologia descrita por MINTE-VERA et al. (2008). Neste método são
utilizados dois raios diferentes de amostragem (2 m e 4 m) (Figura 4). Inicialmente
todas as espécies dentro de um raio de 4 m (definido por trena demarcada) são
identificadas e listadas durante o período de 5 minutos. Depois disso, as espécies
passam a ser registradas quantitativamente, sendo os indivíduos com o
comprimento total (CT) menor ou igual a 10 cm (registrados dentro das categorias: ≤
2 e 2-10 cm) contabilizados dentro do raio de 2 m, e indivíduos com CT > 10 cm
(registrados dentro das categorias: 10-20, 20-30, 30-40, ≥40 cm) registrados no raio
de 4 m (MINTE-VERA et al., 2008; FRANCINI-FILHO E MOURA, 2008a).

BASILIO, P. S. 30
Figura 4. Metodologia de censo visual estacionário (cf. MINTE-VERA et al., 2008). No raio
de dois metros são contabilizados os peixes menores que 10 cm CT e no raio de quatro metros são contabilizados indivíduos >10 cm CT.
Foram incluídas nas análises quantitativas todas as espécies previamente
registradas como predadoras de P. caribaeorum: Abudefduf saxatilis (LINNAEUS,
1758), Aluterus scriptus (OSBECK, 1765), Cantherhines macrocerus (HOLLARD,
1853), Chaetodon striatus Linnaeus, 1758, Chaetodon sedentarius Poey, 1860,
Holacanthus ciliares (LINNAEUS, 1758), Pomacanthus arcuatus (LINNAEUS, 1758)
e Pomacanthus paru (BLOCH, 1787) (RANDALL, 1967; SEBENS, 1982, FRANCINI-
FILHO E MOURA, 2010; LONGO et al.,2012) (Figura 5).
As contagens de peixes foram convertidas em biomassa através da
utilização de relações peso-comprimento (FROESE E PAULY, 2011). A estimativa
foi feita multiplicando-se o peso (valor médio por classe) pelo número de peixes em
cada categoria e somando-se todas as categorias de tamanho (cf. MCCLANAHAN E
KUANDA-ARARA, 1996; FRANCINI-FILHO E MOURA, 2008a).

BASILIO, P. S. 31
Figura 5. Peixes predadores de Palythoa caribaeorum registrados no Banco dos Abrolhos, BA: a) Abudefduf saxatilis, b) Aluterus scriptus, c) Cantherhines macrocerus, d) Chaetodon striatus, e) Chaetodon sedentarius, f) Holacanthus ciliaris, g) Pomacanthus arcuatus e h) Pomacanthus paru. Fotos: R.B. Francini-Filho.
Os dados referentes à distância da costa e latitude foram obtidos a partir de
cartas náuticas digitais. A profundidade de cada sítio foi estimada através da média

BASILIO, P. S. 32
das profundidades obtidas em cada censo visual estacionário de peixes com o uso
de computador de mergulho.
3.2.2. Experimento de exclusão de Palythoa caribaeorum
O experimento foi realizado entre 2006 e 2009 no topo de dois sítios:
Arenguera (norte do Parcel das Paredes e próximo à costa, aproximadamente 15
km) e PAB4 (no Parcel dos Abrolhos, mais afastado da costa, aproximadamente 62
km) (Figura 1 e Tabela 1). Foram selecionados quatro áreas de 5 x 5 m dominadas
por P. caribaeorum em cada sitio, com distâncias mínimas de 15 m entre elas
(Figura 6).
Dentro de cada sítio, duas áreas de 5 x 5 m passaram pelo processo de
manipulação (retirada de P. caribaeorum) (Figura 6 e 7) e outras duas áreas foram
mantidas como controle (i.e. sem exclusão de P. caribaeorum) (Figura 6). Ao todo
foram despendidas cerca 15 h de trabalho individual em cada área manipulada (i.e.
três mergulhadores trabalhando por cinco horas). Apenas P. caribaeorum foi
retirada durante o processo, tendo sido evitada a remoção ou dano a outros
organismos.
Dentro de cada área experimental de 5 x 5 m foram alocados quatro foto-
quadrados (75 x 66 cm) fixos, os quais foram demarcados permanente com
vergalhões de ferro para acompanhamento de longo prazo. Três tratamentos foram
aplicados aos foto-quadrados: 1) gaiola fechada para exclusão de peixes predadores
de P. caribaeorum (1 foto-quadrado) (Figura 8), 2) gaiola aberta no topo (controle
para o efeito da gaiola; 1 foto-quadrtado) e 3) sem gaiola (2 foto-quadrados). As
gaiolas (base com 75 x 66 cm e altura de 50 cm) foram confeccionadas de metal
soldado revestido por malha plástica (5 x 5 cm). Em cada visita o material
depositado sobre a gaiola (sedimento e algas) foi retirado para evitar sombreamento
(Figura 8).
Os foto-quadrados fixados dentro das áreas experimentais dos dois sítios
foram visitados periodicamente (Tabela 2) e o substrato fotografado através da
metodologia apersentada por Francini-Filho et al. (2013). O delineamento completo
é apresentado na Figura 6. As escalas de tempo serão tratadas no decorrer do

BASILIO, P. S. 33
trabalho como tempo corrido em dias (ex.: coleta de dados realizada ápos passados
39 dias de manipulação o tempo seria T39) (Tabela 2).
Tabela 2. Caracterização temporal das coletas realizadas durante o experimento de exclusão de Palythoa caribaeorum. T0: dados de cobertura obtidos antes da retirada da Palythoa caribaeorum, T1: dados de cobertura coletados imediatamente após a retirada da Palythoa caribaeorum. Os demais tempos são referentes aos dias passados desde o inicio do experimento (i.e. retirada da P.caribaeorum).
ARENG
PAB4
Tempo corrido Tempo real
Tempo corrido
Tempo real
Mês/Ano Estação Mês/Ano Estação
T0 JUL/2006 Inverno T0 SET/2006 Primavera
T1 JUL/2006 Inverno
T1 SET/2006 Primavera
T39 AGO/2006 Inverno
T35 OUT/2006 Primavera
T61 SET/2006 Primavera
T55 OUT/2006 Primavera
T88 OUT/2006 Primavera
T86 NOV/2006 Primavera
T183 JAN/2007 Verão
T151 FEV/2007 Verão
T957 MAR/2009 Verão
T903 FEV/2009 Verão
Ao todo, foram realizadas quatro coletas no ano de 2006, uma no ano de
2007 e uma no ano de 2009, totalizaram 3001 imagens. Não foi possível realizar a
coleta de dados de cobertura referente a duas áreas controle (não manipulados) em
PAB4 em 2009 (Tabela 2).

BASILIO, P. S. 34
Figura 6. Esquema representativo do experimento conduzido em dois sítio (Arenguera e
PAB4). GF – gaiola fechada, SG- sem gaiola, GA- gaiola aberta. À direita a delimitação dos foto-quadrados utilizados para captura de imagens digitais. Método de foto-quadrado cf. Francini-Filho et al. (2013).
Figura 7. Área manipulada correspondente ao foto-quadrado fixado dentro da área
experimental. (a) Antes da retirada da Palythoa caribaeorum, Tempo 0 e (b) após a retirada da cobertura de Palythoa caribaeorum, Tempo 1.

BASILIO, P. S. 35
Figura 8. Gaiola de ferro revestida por malha plástica (5 x 5 cm) utilizada no experimento de exclusão de predadores de Palythoa caribaeorum. Foto:Ronaldo Bastos Francini-Filho.
a. Análise de recolonização e vitalidade de Palythoa caribaeorum
Os dados referentes ao experimento de exclusão foram obtidos ao longo de
957 dias, através de fotografias de alta resolução (Figura 7). Inicialmente cada
imagem foi analisada com a utilização dos programas UTHSCSA ImageTool 3.0
para cálculo do número de pixels por cm. Este último valor é necessário para
estabelecimento da escala e análise final no programa eCognition Developer 8.7
(DEFINIENS, 2009), o qual permite a segmentação da imagem por multi-resolução
dos pixels seguida por classificação baseada em similaridade de cores.
Para a análise de recolonização foram calculdas as áreas totais da P.
caribaeorum em cm2 para cada imagem e a porcentagem de cobertura foi calculada
em função da área total de cada imagem analisada (Figura 9).

BASILIO, P. S. 36
Figura 9. Sequência de passos para cálculo da área da Palythoa caribaeorum no programa eCognition Developer 8.7 (DEFINIENS, 2009). Fases da análise: (a) imagem original, (b)
imagem fragmentada (c) seleção dos fragmentos referentes ao organismo e (d) classificação hierárquica.
Para a análise da vitalidade do tecido de P. caribaeorum foi estabelecido o
critério de classificação visual seguinte: branqueado (BRANQ), recoberto por outros
organismos (REC), saudável (SAU) e não-saudável (N-SAU) (Figura 10). A categoria
não-saudavel representa o somatório das categorias branqueada e recoberta.
A área do tecido referente a cada classe foi calculada individualmente por
foto e a porcentagem de cada classe foi calculada em função da área total de
cobertura de P. caribaeorum por imagem analisada.

BASILIO, P. S. 37
Figura 10. Categorias de vitalidade dos tecidos da Palythoa caribaeorum consideradas no presente estudo: a) tecido saudável, b) tecido branqueado e c) tecido recoberto por outros organismos bentônicos.
b. Análise da sucessão bentônica nas áreas experimentais
Para a análise de cobertura relativa de todos os organismos bentônicos na
área do experimento foram utilizadas as mesmas 3001 imagens utilizadas para
cálculo da área de cobertura de P. caribaeorum ao longo do tempo. No entanto, para
esta análise mais abrangente das assembléias bentônicas, foi aplicada a
metodologia de pontos aleatórios (cf. FRANCINI-FILHO et al., 2008), ao invés da
análise de área absoluta, uma vez que esta última é mais custosa. A cobertura
relativa de organismos bentônicos foi estimada com a utilização do programa Coral
Point Count with Excel extensions (CPCe) (KOHLER E GILL, 2006), metodologia
descrita anteriormente (Figura 3).

BASILIO, P. S. 38
3.3. Análises estatísticas
3.3.1. Variações espaciais e temporais na cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos e a influência de variáveis bióticas e abióticas.
Todas as análises foram conduzidas utilizando-se o programa STATISTICA
10.0 (Statsoft Inc.). Os dados foram submetidos aos testes de Kolmogorov-Smirnov
e Levene para atestar a normalidade e a homocedasticidade, respectivamente (ZAR,
1999). Devido a sua distribuição não normal, os dados de cobertura relativa foram
transformados em arcseno √ x.
Primeiramente foi realizado um teste t para amostras independentes
comparando a cobertura relativa de P. caribaeorum entre os habitats topo e parede
dos chapeirões. Uma vez que o resultado deste teste demosntrou coberturas
inexpressivas de P. caribaeorum na parede, todas as análises posteriores foram
realizadas considerando apenas dados obtidos para o topo dos recifes.
Para testar o efeito dos anos e dos sítios na cobertura de P. caribaeorum foi
usada uma Análise de Variância (ANOVA) fatorial. O teste de comparação a
posteriori de Student-Newman-Keuls foi aplicado para identificação de grupos
homogêneos (ZAR, 1999).
Foram realizadas regressões lineares múltiplas para avaliar a influência das
variáveis ambientais e bióticas (profundidade, biomassa de peixes predadores de P.
caribaeorum, distância da costa e latitude) na cobertura relativa de P. caribaeorum,
utilizando os sítios como réplicas (valores médios).
3.3.2. Experimento de exclusão de Palythoa caribaeorum
a. Recolonização e vitalidade da Palythoa caribaeorum
Os dados de porcentagem foram transformados em arcsen √x. Para
testar o efeito dos fatores [sítio, tempo, raspagem (raspado ou não raspado) e
tratamento (gaiola aberta, gaiola fechada ou sem gaiola)] na cobertura total de P.
caribaeorum e nas diferentes categorias de vitalidade (tecido saudável, branqueado,
recoberto e não saudável) foram usadas Análises de Variância (ANOVA) fatorial e
testes de comparação a posteriori de Student-Newman-Keuls (SNK) (ZAR, 1999).

BASILIO, P. S. 39
Para melhor avaliar a variação temporal na porcentagem de P. caribaeorum
total para os sitios, foram padronizados três tempos: TA - inicio do experimento,
antes da exclusão, TB - no meio do período de estudo e TC - final do experimento.
Com esses dados, foram realizadas Análises de Variância (ANOVA) fatorial com os
fatores: sítio x tempo x raspagem x tratamento e teste de comparação a posteriori de
Student-Newman-Keuls (SNK).
b. Sucessão de organismos bentônicos
Os dados foram transformados para √ x, sendo posteriormente calculada
uma matriz de similaridade entre amostras, utilizando-se a distância de Bray-Curtis.
Análises de similaridade (ANOSIM) foram realizadas com o propósito de identificar
diferenças significativas entre grupos dentro de cada fator (CLARKE, 1993). Para
identificar as variáveis responsáveis pelas diferenças entre os fatores observados foi
aplicado o método de porcentagem de similaridade (SIMPER) (CLARKE, 1993).
Todas as análises referentes aos dados de sucessão de organismos bentônicos
foram realizadas através do software PRIMER 6.0 (CLARKE E WARWICK, 2001).

BASILIO, P. S.
40
4. RESULTADOS
BASILIO, P.S.
41
4. RESULTADOS
4.1. Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no
Banco dos Abrolhos e a influência de variáveis bióticas e abióticas
A cobertura relativa de P. caribaeorum variou significativamente entre os
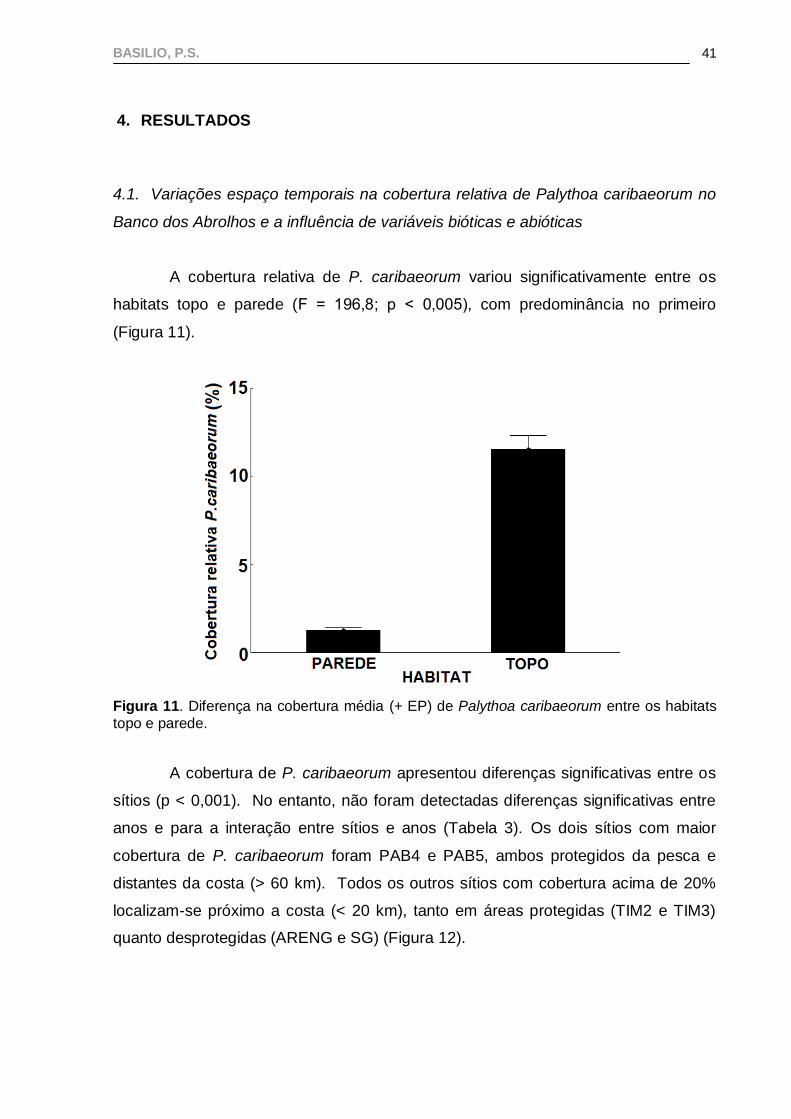
habitats topo e parede (F = 196,8; p < 0,005), com predominância no primeiro
(Figura 11).
Figura 11. Diferença na cobertura média (+ EP) de Palythoa caribaeorum entre os habitats
topo e parede.
A cobertura de P. caribaeorum apresentou diferenças significativas entre os
sítios (p < 0,001). No entanto, não foram detectadas diferenças significativas entre
anos e para a interação entre sítios e anos (Tabela 3). Os dois sítios com maior
cobertura de P. caribaeorum foram PAB4 e PAB5, ambos protegidos da pesca e
distantes da costa (> 60 km). Todos os outros sítios com cobertura acima de 20%
localizam-se próximo a costa (< 20 km), tanto em áreas protegidas (TIM2 e TIM3)
quanto desprotegidas (ARENG e SG) (Figura 12).

BASILIO, P.S.
42
Tabela 3. Análise de variância (ANOVA) fatorial testando o efeito dos fatores sítio e ano na
cobertura relativa de Palythoa caribaeorum no Banco dos Abrolhos.
Fator Cobertura relativa
F P
Sítio 21,6 *** Ano 0,2 ns
Sítio X Ano 0,3 ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
Figura 12. Cobertura de Palythoa caribaeorum em diferentes sítios no Banco dos Abrolhos
e seus respectivos níveis de proteção e distância da costa. As letras indicam grupos homogêneos de sítios com base em comparações múltiplas a posteriori de Student-
Newman-Keuls.Veja código dos sítios na Tabela 1.
As cinco espécies de predadores de P. caribaeorum mais abundantes em
termos de biomassa foram A. saxatilis (2,95 ± 0,20 EP), P. paru (2,23 ± 0,27 EP), P.
arcuatus (1,13 ± 0,17 EP), H. ciliares (0,48 ± 0,06 EP) e C. striatus (0,47 ± 0,03)
(Figura 13). Foram registradas diferenças significativas entre sítios para a biomassa
total e para a maioria das espécies, exceto A. scriptus (Figura 13). Os cinco sítios
que apresentaram os maiores valores de biomassa total, em ordem decrescente,
foram TIM2, PAB5, TIM3, PAB3 e SIRIBA (Figura13).

BASILIO, P.S.
43
Figura 13. Variação na biomassa (g.m-2) (MÉDIA + EP) das espécies de peixes predadores de Palythoa caribaeorum no topo dos sítios estudados. *P < 0,05, **P < 0,01, ***P < 0,001 e
ns- não significativo. Veja código dos sítios na Tabela1.

BASILIO, P.S.
44
A cobertura de P. caribaeorum foi influenciada positivamente pela
profundidade (P < 0,01; beta = 0,98) e negativamente pela distância da costa (P <
0,05; beta = -0,79), com o efeito positivo da profundidade sendo mais forte que o
efeito negativo da distância da costa. Não foram registrados efeitos significativos da
latitude e da biomassa de predadores (Tabela 4).
Tabela 4. Resultado da regressão múltipla mostrando a influência relativa da distancia da
costa, profundidade, latitude e biomassa de peixes predadores de zoantídeos na cobertura de Palythoa caribaeorum.
Espécie r2 P
Parcial r2
Distância da costa (km)
Profundidade (m)
Latitude Predadores
(g.m-2)
P. caribaeorum 0,44 ** -0,79* 0,98** 0,32ns -0,05ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
4.2. Experimento de exclusão
4.2.1. Recolonização da Palythoa caribaeorum
Como esperado, a ANOVA evidenciou diferenças significativas na cobertura
de P. caribaeorum nas áreas manipuladas, entre os tempos t0 e t1 (F = 1119,1; P <
0,001) antes e imediatamente após a exclusão de P. caribaeorum em ambos os
sítios (Figura 14). A redução na cobertura total de P. caribaeorum nas áreas
manipuladas foi de 99,99% em ARENG e 99,93% em PAB4.

BASILIO, P.S.
45
Figura 14. Eficiência na exclusão de Palythoa caribaeorum nas áreas manipuladas, antes (T0) e imediatamente após a exclusão (T1). Sítios: Arenguera (ARENG) e Parcel dos Abrolhos (PAB4).
A cobertura de P. caribaeorum variou em relação a quase todos os fatores,
exceto para a interação entre sítio x tempo x tratamento e sítio x raspagem x
tratamento (Tabela 5). A falta de significância entre a interação tratamento e tempo
indica que as trajetórias temporais foram similares em todos os tratamentos, ou seja,
não foi registrado efeito da exclusão de predadores de P. caribaeorum (Tabela 5). A
interação entre sítio x tempo x raspagem foi significativa devido, principalmente, ao
declínio óbvio após a raspagem (Tabela 5 e Figura 15).
Tabela 5. Análise de variância (ANOVA) fatorial testando os efeitos de sítio, tempo, raspagem (raspado ou não raspado) e tratamento (com gaiola, com gaiola aberta e sem gaiola) na cobertura de Palythoa caribaeorum.
Fatores Palythoa caribaeorum total
F P
Sítio 158,38 ***
Sítio x tempo 37,82 ***
Sítio x raspagem 29,28 ***
Sítio x tratamento 4,97 ***
Sítio x tempo x raspagem 25,00 ***
Sítio x tempo x tratamento 1,95 ns
Sítio x raspagem x tratamento 2,27 ns
Sítio x tempo x raspagem x tratamento 3,14 *
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
BASILIO, P.S.
46
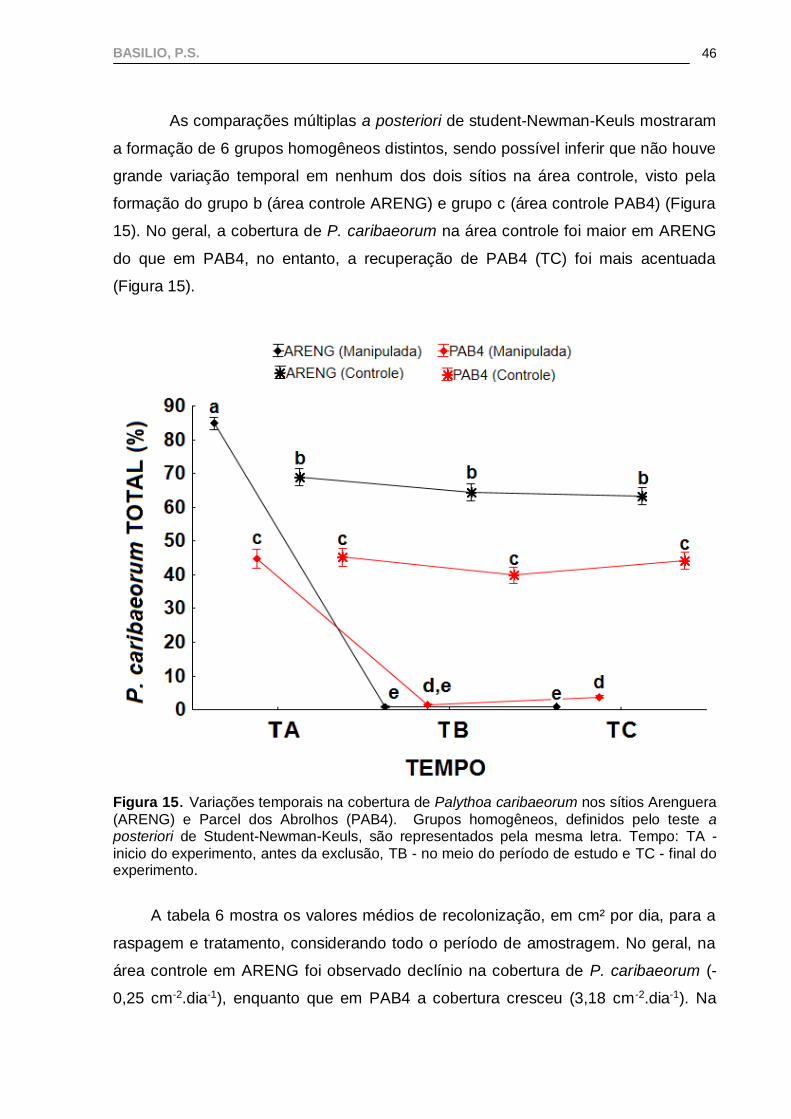
As comparações múltiplas a posteriori de student-Newman-Keuls mostraram
a formação de 6 grupos homogêneos distintos, sendo possível inferir que não houve
grande variação temporal em nenhum dos dois sítios na área controle, visto pela
formação do grupo b (área controle ARENG) e grupo c (área controle PAB4) (Figura
15). No geral, a cobertura de P. caribaeorum na área controle foi maior em ARENG
do que em PAB4, no entanto, a recuperação de PAB4 (TC) foi mais acentuada
(Figura 15).
Figura 15. Variações temporais na cobertura de Palythoa caribaeorum nos sítios Arenguera
(ARENG) e Parcel dos Abrolhos (PAB4). Grupos homogêneos, definidos pelo teste a posteriori de Student-Newman-Keuls, são representados pela mesma letra. Tempo: TA -
inicio do experimento, antes da exclusão, TB - no meio do período de estudo e TC - final do experimento.
A tabela 6 mostra os valores médios de recolonização, em cm² por dia, para a
raspagem e tratamento, considerando todo o período de amostragem. No geral, na
área controle em ARENG foi observado declínio na cobertura de P. caribaeorum (-
0,25 cm-2.dia-1), enquanto que em PAB4 a cobertura cresceu (3,18 cm-2.dia-1). Na

BASILIO, P.S.
47
área manipulada, a taxa de crescimento média total foi maior em ARENG (0,23 cm -
2.dia-1) do que em PAB4 (0,12 cm-2.dia-1) (Tabela 6).
Observando separadamente as áreas de raspagem, pode-se perceber que em
PAB4 a maior taxa de crescimento foi observada na gaiola fechada (GF), tanto na
área controle (1,93 cm-2.dia-1), quanto na área manipulada (0,07 cm-2.dia-1). Já em
ARENG, as maiores taxas foram observadas na Gaiola aberta (GA), sendo de 0,13
cm-2.dia-1 na área manipulada e de 0,31 cm-2.dia-1 na área controle (Tabela 6).
Tabela 6. Taxas de crescimento da P.caribaeorum (cm².dia-1) para cada tratamento nas
áreas manipuladas e controle,nos sítios PAB4 e ARENG.
Raspagem Tratamento Crescimento (cm².dia-1)
PAB4 ARENG
Manipulado GF 0,07 0,01
Manipulado GA 0,05 0,13
Manipulado SG -0,01 0,10
Manipulado Total 0,12 0,23
Controle GF 1,93 -0,14
Controle GA 1,02 0,31
Controle SG 0,24 -0,43
Controle Total 3,18 -0,25
Tratamento: Gaiola aberta (GA), gaiola fechada (GF) e sem gaiola (SG).
a. Arenguera (ARENG)
As análises realizadas separadamente para ARENG demonstraram que a
cobertura de P. caribaeorum variou significativamente em relação a quase todos os
fatores, exceto nas interações tempo x tratamento e tempo x tratamento x raspagem,
indicando que não houve efeito da exclusão de predadores pelas gaiolas (Tabela 7).
A interação significativa entre os fatores tempo x raspagem indica que a variação
temporal na cobertura de P. caribaeorum ocorreu de forma diferenciada nas áreas
controle e nas áreas manipuladas (Tabela 7).
BASILIO, P.S.
48
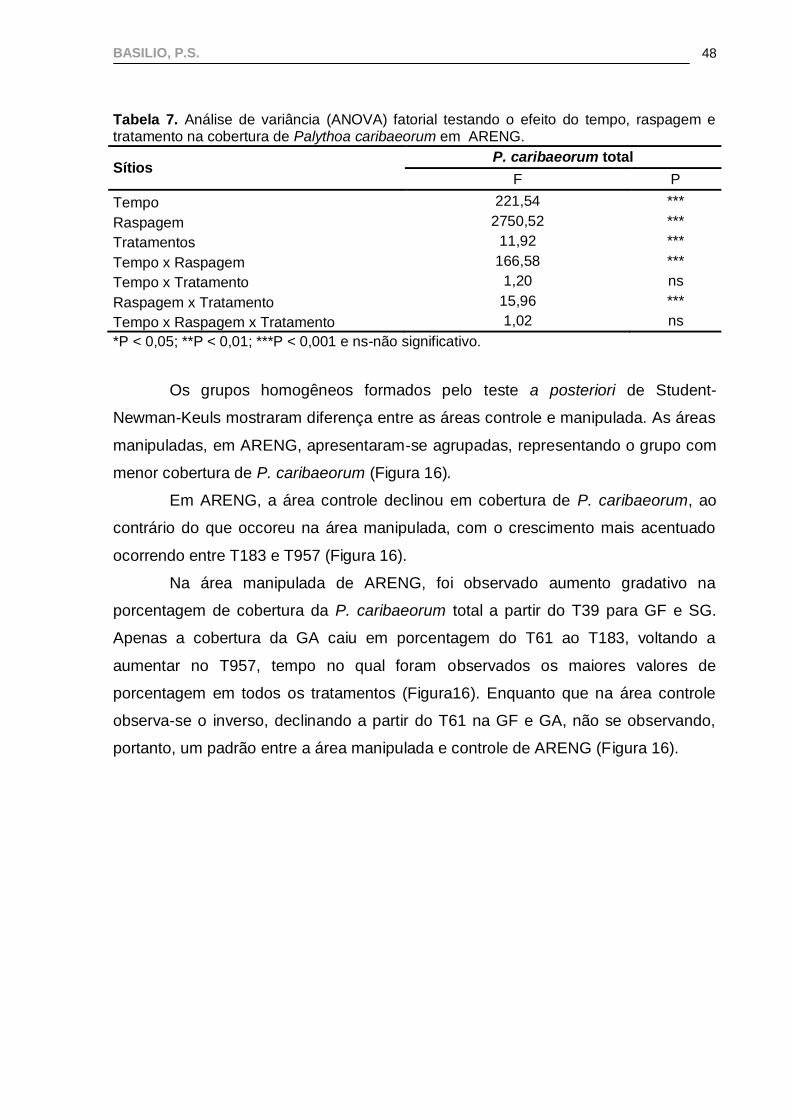
Tabela 7. Análise de variância (ANOVA) fatorial testando o efeito do tempo, raspagem e tratamento na cobertura de Palythoa caribaeorum em ARENG.
Sítios P. caribaeorum total
F P
Tempo 221,54 ***
Raspagem 2750,52 ***
Tratamentos 11,92 ***
Tempo x Raspagem 166,58 ***
Tempo x Tratamento 1,20 ns
Raspagem x Tratamento 15,96 ***
Tempo x Raspagem x Tratamento 1,02 ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
Os grupos homogêneos formados pelo teste a posteriori de Student-
Newman-Keuls mostraram diferença entre as áreas controle e manipulada. As áreas
manipuladas, em ARENG, apresentaram-se agrupadas, representando o grupo com
menor cobertura de P. caribaeorum (Figura 16).
Em ARENG, a área controle declinou em cobertura de P. caribaeorum, ao
contrário do que occoreu na área manipulada, com o crescimento mais acentuado
ocorrendo entre T183 e T957 (Figura 16).
Na área manipulada de ARENG, foi observado aumento gradativo na
porcentagem de cobertura da P. caribaeorum total a partir do T39 para GF e SG.
Apenas a cobertura da GA caiu em porcentagem do T61 ao T183, voltando a
aumentar no T957, tempo no qual foram observados os maiores valores de
porcentagem em todos os tratamentos (Figura16). Enquanto que na área controle
observa-se o inverso, declinando a partir do T61 na GF e GA, não se observando,
portanto, um padrão entre a área manipulada e controle de ARENG (Figura 16).
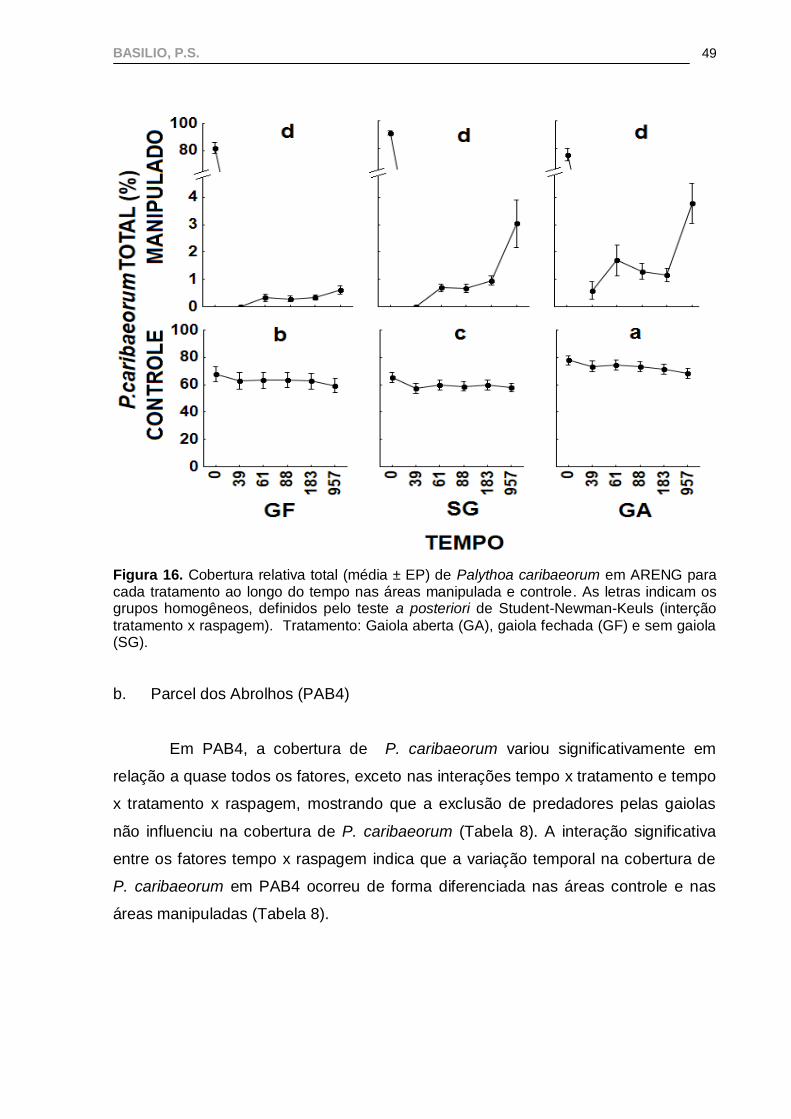
BASILIO, P.S.
49
Figura 16. Cobertura relativa total (média ± EP) de Palythoa caribaeorum em ARENG para cada tratamento ao longo do tempo nas áreas manipulada e controle. As letras indicam os grupos homogêneos, definidos pelo teste a posteriori de Student-Newman-Keuls (interção
tratamento x raspagem). Tratamento: Gaiola aberta (GA), gaiola fechada (GF) e sem gaiola (SG).
b. Parcel dos Abrolhos (PAB4)
Em PAB4, a cobertura de P. caribaeorum variou significativamente em
relação a quase todos os fatores, exceto nas interações tempo x tratamento e tempo
x tratamento x raspagem, mostrando que a exclusão de predadores pelas gaiolas
não influenciu na cobertura de P. caribaeorum (Tabela 8). A interação significativa
entre os fatores tempo x raspagem indica que a variação temporal na cobertura de
P. caribaeorum em PAB4 ocorreu de forma diferenciada nas áreas controle e nas
áreas manipuladas (Tabela 8).

BASILIO, P.S.
50
Tabela 8. Análise de variância (ANOVA) fatorial testando o efeito do tempo, raspagem e tratamento na cobertura de Palythoa caribaeorum em PAB4.
Sítios P. caribaeorum total
F P
Tempo 60,95 ***
Raspagem 747,59 ***
Tratamentos 12,41 ***
Tempo x Raspagem 50,61 ***
Tempo x Tratamento 0,93 ns
Raspagem x Tratamento 3,06 *
Tempo x Raspagem x Tratamento 1,36 ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
Os grupos homogêneos formados pela interação raspagem x tratamento
mostraram diferenças entre as áreas controle e manipulada, sendo a area
manipulada representada pelos menores valores em porcentagem de cobertura de
P. caribaeorum (Figura 17).
Em PAB4, o valor máximo de cobertura de P. caribaeorum foi registrado no
T151 e o mínimo no T55, tanto na área manipulada quanto na área controle (Figura
17).
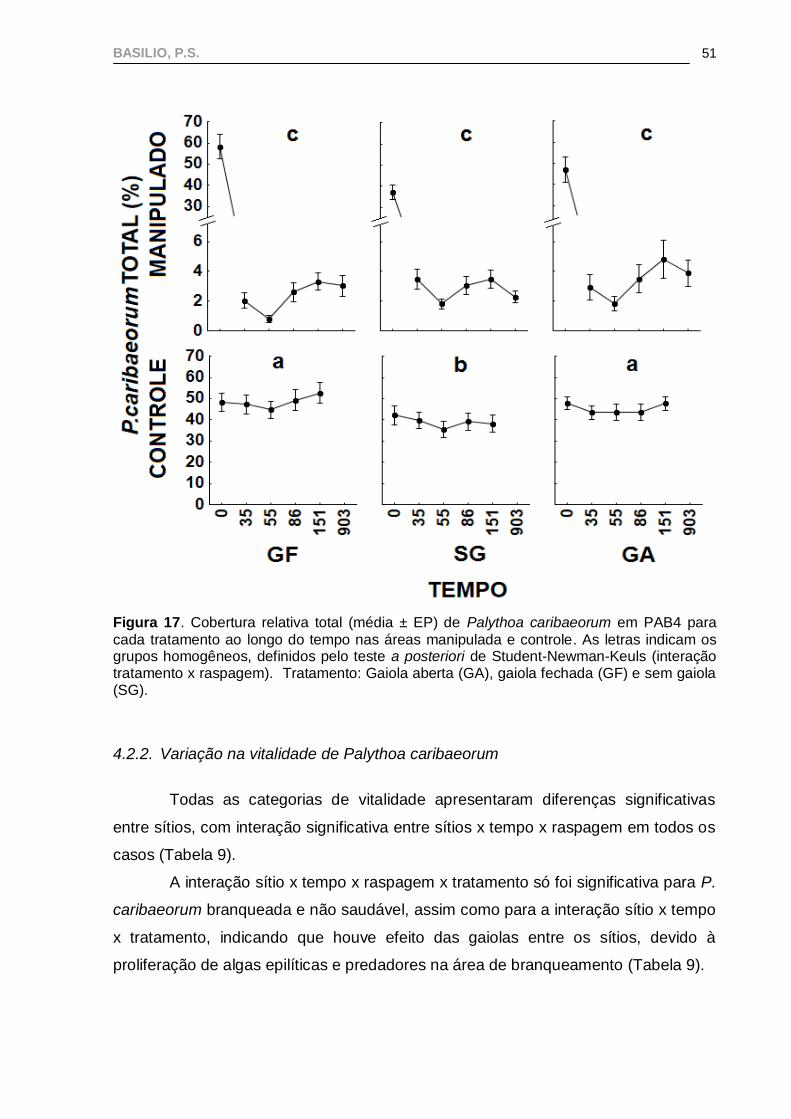
BASILIO, P.S.
51
Figura 17. Cobertura relativa total (média ± EP) de Palythoa caribaeorum em PAB4 para
cada tratamento ao longo do tempo nas áreas manipulada e controle. As letras indicam os grupos homogêneos, definidos pelo teste a posteriori de Student-Newman-Keuls (interação tratamento x raspagem). Tratamento: Gaiola aberta (GA), gaiola fechada (GF) e sem gaiola (SG).
4.2.2. Variação na vitalidade de Palythoa caribaeorum
Todas as categorias de vitalidade apresentaram diferenças significativas
entre sítios, com interação significativa entre sítios x tempo x raspagem em todos os
casos (Tabela 9).
A interação sítio x tempo x raspagem x tratamento só foi significativa para P.
caribaeorum branqueada e não saudável, assim como para a interação sítio x tempo
x tratamento, indicando que houve efeito das gaiolas entre os sítios, devido à
proliferação de algas epilíticas e predadores na área de branqueamento (Tabela 9).
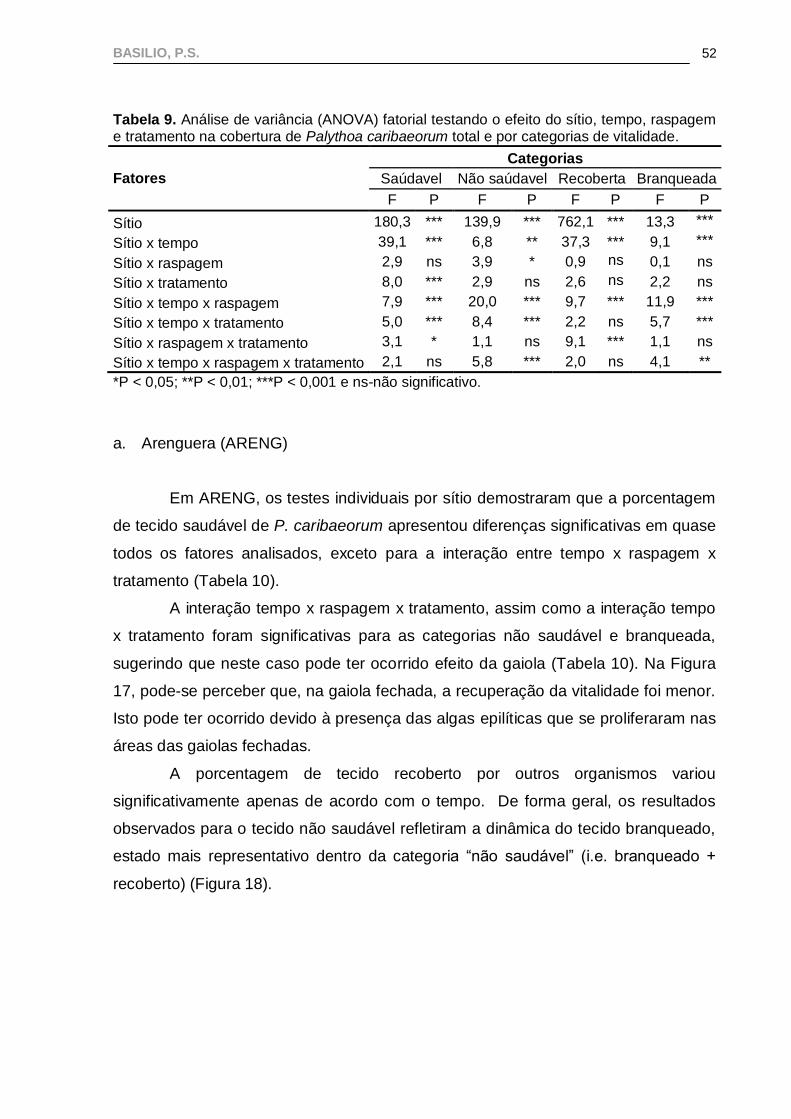
BASILIO, P.S.
52
Tabela 9. Análise de variância (ANOVA) fatorial testando o efeito do sítio, tempo, raspagem e tratamento na cobertura de Palythoa caribaeorum total e por categorias de vitalidade.
Fatores
Categorias
Saúdavel Não saúdavel Recoberta Branqueada
F P F P F P F P
Sítio 180,3 *** 139,9 *** 762,1 *** 13,3 ***
Sítio x tempo 39,1 *** 6,8 ** 37,3 *** 9,1 ***
Sítio x raspagem 2,9 ns 3,9 * 0,9 ns 0,1 ns
Sítio x tratamento 8,0 *** 2,9 ns 2,6 ns 2,2 ns
Sítio x tempo x raspagem 7,9 *** 20,0 *** 9,7 *** 11,9 ***
Sítio x tempo x tratamento 5,0 *** 8,4 *** 2,2 ns 5,7 ***
Sítio x raspagem x tratamento 3,1 * 1,1 ns 9,1 *** 1,1 ns
Sítio x tempo x raspagem x tratamento 2,1 ns 5,8 *** 2,0 ns 4,1 **
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
a. Arenguera (ARENG)
Em ARENG, os testes individuais por sítio demostraram que a porcentagem
de tecido saudável de P. caribaeorum apresentou diferenças significativas em quase
todos os fatores analisados, exceto para a interação entre tempo x raspagem x
tratamento (Tabela 10).
A interação tempo x raspagem x tratamento, assim como a interação tempo
x tratamento foram significativas para as categorias não saudável e branqueada,
sugerindo que neste caso pode ter ocorrido efeito da gaiola (Tabela 10). Na Figura
17, pode-se perceber que, na gaiola fechada, a recuperação da vitalidade foi menor.
Isto pode ter ocorrido devido à presença das algas epilíticas que se proliferaram nas
áreas das gaiolas fechadas.
A porcentagem de tecido recoberto por outros organismos variou
significativamente apenas de acordo com o tempo. De forma geral, os resultados
observados para o tecido não saudável refletiram a dinâmica do tecido branqueado,
estado mais representativo dentro da categoria “não saudável” (i.e. branqueado +
recoberto) (Figura 18).
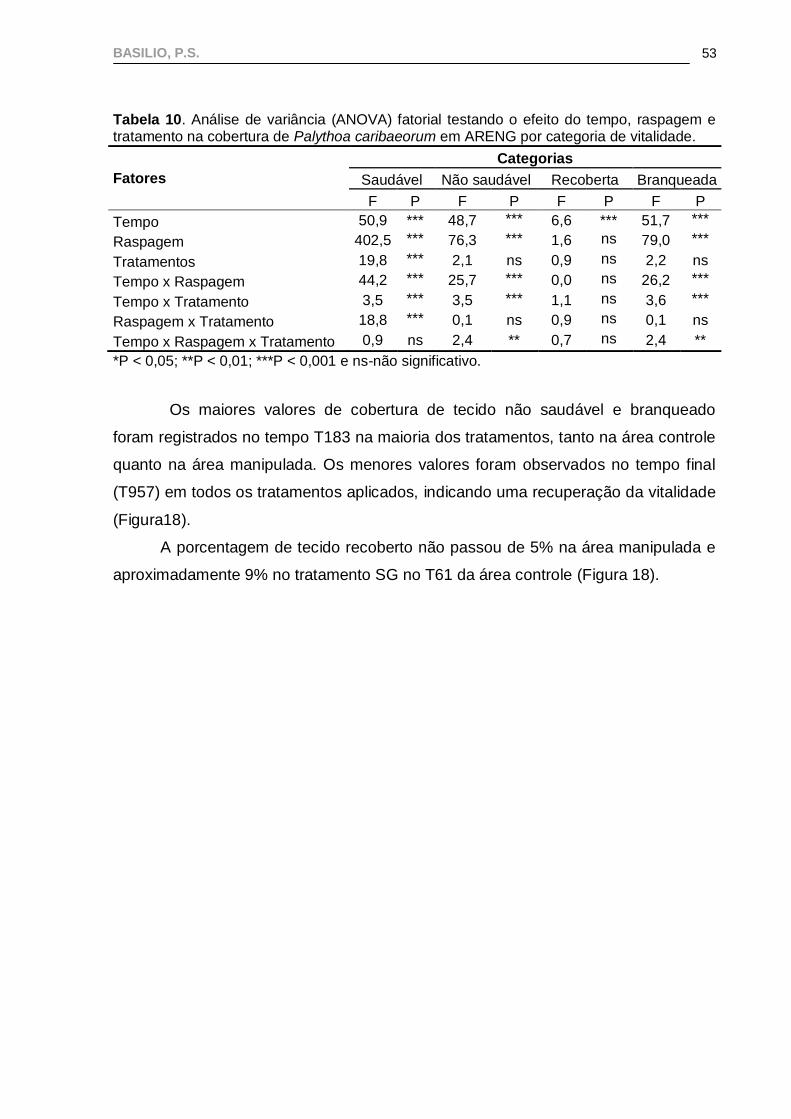
BASILIO, P.S.
53
Tabela 10. Análise de variância (ANOVA) fatorial testando o efeito do tempo, raspagem e tratamento na cobertura de Palythoa caribaeorum em ARENG por categoria de vitalidade.
Fatores
Categorias
Saudável Não saudável Recoberta Branqueada
F P F P F P F P
Tempo 50,9 *** 48,7 *** 6,6 *** 51,7 ***
Raspagem 402,5 *** 76,3 *** 1,6 ns 79,0 ***
Tratamentos 19,8 *** 2,1 ns 0,9 ns 2,2 ns
Tempo x Raspagem 44,2 *** 25,7 *** 0,0 ns 26,2 ***
Tempo x Tratamento 3,5 *** 3,5 *** 1,1 ns 3,6 ***
Raspagem x Tratamento 18,8 *** 0,1 ns 0,9 ns 0,1 ns
Tempo x Raspagem x Tratamento 0,9 ns 2,4 ** 0,7 ns 2,4 **
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
Os maiores valores de cobertura de tecido não saudável e branqueado
foram registrados no tempo T183 na maioria dos tratamentos, tanto na área controle
quanto na área manipulada. Os menores valores foram observados no tempo final
(T957) em todos os tratamentos aplicados, indicando uma recuperação da vitalidade
(Figura18).
A porcentagem de tecido recoberto não passou de 5% na área manipulada e
aproximadamente 9% no tratamento SG no T61 da área controle (Figura 18).

BASILIO, P.S.
54
Figura 18. Porcentagem (média ± EP) de diferentes categorias de vitalidade de Palythoa caribaeorum durante o tempo na área manipulada (raspada) e controle em ARENG. As letras indicam os grupos homogêneos definidos pelo teste a posteriori de Student-Newman-
Keuls. Tratamento: Gaiola aberta (GA), gaiola fechada (GF) e sem gaiola (SG).
b. Parcel dos Abrolhos (PAB4)
A interação tempo x raspagem x tratamento foi significativa para as
categorias saudável e não saudável (Tabela 11), indicando trajetórias diferentes nas
gaiolas (Figura 19).
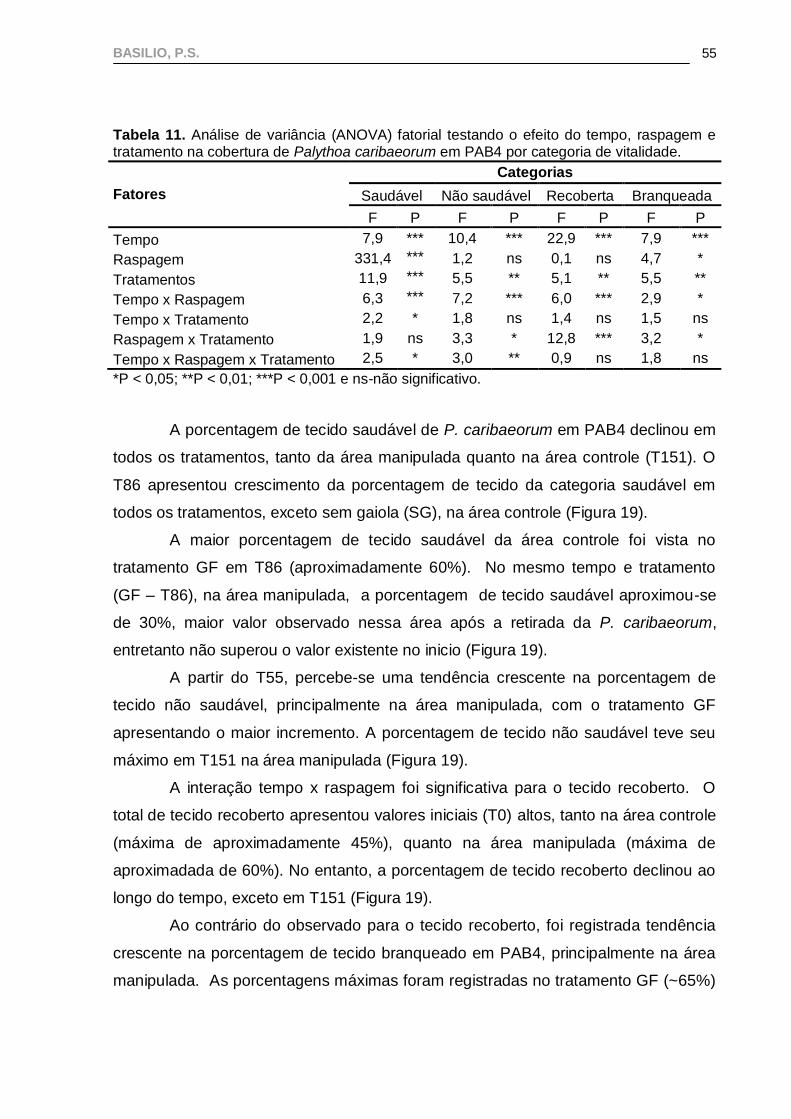
BASILIO, P.S.
55
Tabela 11. Análise de variância (ANOVA) fatorial testando o efeito do tempo, raspagem e tratamento na cobertura de Palythoa caribaeorum em PAB4 por categoria de vitalidade.
Fatores
Categorias
Saudável Não saudável Recoberta Branqueada
F P F P F P F P
Tempo 7,9 *** 10,4 *** 22,9 *** 7,9 ***
Raspagem 331,4 *** 1,2 ns 0,1 ns 4,7 *
Tratamentos 11,9 *** 5,5 ** 5,1 ** 5,5 **
Tempo x Raspagem 6,3 *** 7,2 *** 6,0 *** 2,9 *
Tempo x Tratamento 2,2 * 1,8 ns 1,4 ns 1,5 ns
Raspagem x Tratamento 1,9 ns 3,3 * 12,8 *** 3,2 *
Tempo x Raspagem x Tratamento 2,5 * 3,0 ** 0,9 ns 1,8 ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
A porcentagem de tecido saudável de P. caribaeorum em PAB4 declinou em
todos os tratamentos, tanto da área manipulada quanto na área controle (T151). O
T86 apresentou crescimento da porcentagem de tecido da categoria saudável em
todos os tratamentos, exceto sem gaiola (SG), na área controle (Figura 19).
A maior porcentagem de tecido saudável da área controle foi vista no
tratamento GF em T86 (aproximadamente 60%). No mesmo tempo e tratamento
(GF – T86), na área manipulada, a porcentagem de tecido saudável aproximou-se
de 30%, maior valor observado nessa área após a retirada da P. caribaeorum,
entretanto não superou o valor existente no inicio (Figura 19).
A partir do T55, percebe-se uma tendência crescente na porcentagem de
tecido não saudável, principalmente na área manipulada, com o tratamento GF
apresentando o maior incremento. A porcentagem de tecido não saudável teve seu
máximo em T151 na área manipulada (Figura 19).
A interação tempo x raspagem foi significativa para o tecido recoberto. O
total de tecido recoberto apresentou valores iniciais (T0) altos, tanto na área controle
(máxima de aproximadamente 45%), quanto na área manipulada (máxima de
aproximadada de 60%). No entanto, a porcentagem de tecido recoberto declinou ao
longo do tempo, exceto em T151 (Figura 19).
Ao contrário do observado para o tecido recoberto, foi registrada tendência
crescente na porcentagem de tecido branqueado em PAB4, principalmente na área
manipulada. As porcentagens máximas foram registradas no tratamento GF (~65%)

BASILIO, P.S.
56
da área manipulada e SG (~50%) da área controle, valores estes que superaram em
até 80% a cobertura observada inicialmente (T0) (Figura 19).
Figura19. Porcentagem (média ± EP) de diferentes categorias de vitalidade de Palythoa caribaeorum durante o tempo na área manipulada e controle em PAB4. As letras indicam grupos homogêneos definidos pelo teste a posteriori de Student-Newman-Keuls.
Tratamento: Gaiola aberta (GA), gaiola fechada (GF) e sem gaiola (SG).
4.3 Sucessão de organismos bentônicos
As Análises de Similaridade (ANOSIM) demonstraram diferenças
significativas para todos os fatores, exceto tratamento (Tabela 12). Portanto, não foi
detectada influência das gaiolas nestas análises. O diagrama da ordenação nMDS

BASILIO, P.S.
57
(Figura 20) permitiu visualizar diferenças entre as áreas manipulada e controle de
ambos os sítios e a diferenças em relação aos tempos.
Foi registrada tendência, após a manipulação, de aproximação entre as
assembleias bentônicas dos dois sítios ao longo do tempo, convergindo para um
estado diferente do inicial em ambos os sítios (Figura 20).
Tabela 12. Análise de Similaridade (ANOSIM) testando o efeito do tempo, raspagem e
tratamento nas categorias de substrato entre os sítios.
FATORES
R-Global P
Sitio 0,369 ***
Tempo 0,095 ***
Raspagem 0,393 ***
Tratamento 0,027 ns
*P < 0,05; **P < 0,01; ***P < 0,001 e ns-não significativo.
Figura 20. Escalonamento multidimensional não-métrico (NMDS) mostrando a similaridade
(Bray -Curtis) dos sítios (PAB4 e ARENG), tempo e raspagem em relação as categorias de substrato.
As categorias de substrato responsáveis pela dissimilaridade (40,50%) entre
os grupos de sítios, em ordem decrescente, foram: P. caribaeorum, algas epilíticas,
corais, algas calcárias incrustantes, algas calcárias articuladas e Halimeda spp., com

BASILIO, P.S.
58
as duas primeiras categorias listadas representando mais de 50% do percentual de
contribuição (Tabela 13).
Tabela 13. Porcentagens obtidas pelo SIMPER referentes às espécies que mais
contribuíram para as diferenças encontradas entre os sítios.
Categorias de substrato ARENG PAB4
Abundância Média Abundância Média Dissimilaridade %
Palythoa caribaeorum 6,76 5,95 27,76 Alga Epilítica 5,56 9,76 27,34
Coral 0,63 2,91 14,28 Alga calcária incrustante 3,23 2,07 10,52
Alga calcária articulada 1,53 0,26 8,48
Halimeda spp. 1,07 0,11 6,37
O padrão de cobertura observado para P. caribaeorum foi muito semelhante
ao visto nas análises anteriores (i.e. análises de área). De modo geral, foi registrado
aumento acentuado na cobertura de algas epilíticas ao longo do tempo na área
manipulada em ambos os sítios, mas principalmente em ARENG (Figura 21).
A cobertura de alga calcárea incrustante apresentou, após a manipulação da área,
um aumento imediato (~15%) em ARENG e posterior declínio de mesmo valor,
enquanto que em PAB4 observa-se um leve declínio imediato (menor que 1%) e
aumento posterior de aproximadamente 5% (Figura 21).
A cobertura de coral apresenta um pequeno aumento (~4%) em PAB4 na área
manipulada. Em ARENG a variação de cobertura é muito baixa (menor que 0,4%)
(Figura 21).

BASILIO, P.S.
59
Figura 21. Porcentagem (média ±EP) dos principais componentes bentônicos responsáveis
pela dissimilaridade entre os sítios (ARENG e PAB4) durante o tempo na área manipulada e controle. Para melhor visualização da dinâmica de recolonização os gráficos apresentam escala diferenciada.

BASILIO, P.S.
60
5. DISCUSSÃO

BASILIO, P.S.
61
5. DISCUSSÃO
5.1 Variações espaço temporais na cobertura relativa de Palythoa caribaeorum no
Banco dos Abrolhos e a influência de variáveis bióticas e abióticas
Os sítios com maior cobertura relativa de P. caribaeorum foram PAB5 e
PAB4 (distantes da costa) e ARENG (próximo à costa). Entretanto, não foram
observadas diferenças significativas na cobertura relativa ao longo do tempo (2006-
2008). O mesmo resultado foi obtido por Francini-Filho et. al. (2013).
O Banco dos Abrolhos, situado sobre a plataforma continental, é conhecido
como um ambiente relativamente estável. Os distúrbios naturais que influenciam
essa área dizem respeito às oscilações do nível do mar que ocorreram na escala de
milhares de anos e a eventos ligados ao branqueamento de organismos bentônicos,
principalmente corais escleractíneos (Leão et al., 2003). Mesmo que os estudos
feitos no Banco dos Abrolhos tenham sido geralmente realizados em períodos pós-
impactos (e.g.; sobrepesca, sedimentação e branqueamento), ainda é possível
sugerir que este ambiente mantém relativa estabilidade, com componentes bióticos
adaptados às dinâmicas locais, fazendo com que as mudanças temporais de curto
prazo não sejam tão marcantes.
Foram observadas variações significativas entre os sítios de amostragem,
com os maiores valores de cobertura de Palythoa caribaeorum sendo encontrados
no topo dos recifes. Além disso, a regressão múltipla mostrou que a influência
positiva da profundidade foi o fator mais forte sobre distribuição da espécie.
Contudo, apesar da importante diferença na profundidade entre os sítios de
amostragens, acredita-se que apenas este fator não seja suficiente para explicar a
diferença marcante na cobertura de P. caribaeorum (SEGAL E CASTRO, 2011).
As profundidades médias dos sítios estudados ficaram entre 1,3m e 7,7 m.
Estes valores encontram-se dentro da área de distribuição característica de P.
caribaeorum, que pode variar entre 0 a 15 m de profundidade (SEBENS,1977; 1982;
BENAYAHU E LOYA, 1981; SUCHANEK E GREEN, 1981; DALY et al., 2007;
MENDONÇA-NETO, et al., 2008). Os sítios com maior cobertura de P. caribaeorum
foram os que apresentaram as maiores profundidades de topo, inclusive nos recifes
próximos da costa como Arenguera e Timbebas. A distância da costa foi outro fator

BASILIO, P.S.
62
que influenciou a distribuição de P. caribaeorum no Banco dos Abrolhos, porém, de
forma negativa. As áreas de topo favorecem ao desenvolvimento da P.caribaeorum
devido à sua inclinação horizontal do substrato, além de receberem maior incidência
de Luz. Em resumo, P. caribaeorum domina topos fundos e próximos a costa.
De acordo com a regressão múltipla, a biomassa de peixes predadores de
zoantídeos não influenciou na distribuição da P. caribaeorum. Trabalhos anteriores
sugerem que a predação em ambientes recifais pode agir no controle da abundância
de zoantídeos (SUCHANEK E GREEN, 1981; SEBENS, 1982; TANNER, 1997;
FRANCINI-FILHO E MOURA, 2010), mesmo considerando que eles possuem,
morfologia e compostos bioquímicos que representam impedimentos ao predador
(SUCHANEK E GREEN, 1981). No entanto, os resultados do presente estudo, o
qual representa a primeira avaliação abrangente da influência da predação na
dinâmica de P. caribaeorum, indicam que variáveis ambientais, como profundidade e
distância da costa, são mais importantes do que variáveis bióticas (i.e predação) na
dinâmica de P. caribaeorum.
5.2 Experimento de exclusão: Recolonização e vitalidade da Palythoa caribaeorum.
Palythoa caribaeorum é conhecida como uma espécie de crescimento
acelerado, podendo atingir taxas de crescimento na ordem de 0,4 mm.dia-1, a maior
entre os antozoários (SUCHANEK E GREEN, 1981; RABELO et al., 2013). Na área
do experimento essa taxa foi de 0,23 cm². dia-1 em ARENG e 0,12 cm².dia-1 em
PAB4, na área manipulada, valores tambem inferiores ao encontrado por Rabelo et
al. (2013) (11.05 ± 0.74 cm2 .mês-1) em recifes de arenito no nordeste brasileiro.
Estas diferenças nos valores podem está relacionadas ao fato de que, neste
trabalho foi medido a recolonização de uma área (i.e. recrutamento seguido de
crescimento) e não o crescimento linear das colônias de P. caribaeorum como visto
nos trabalhos acima citados.
Em PAB4, (sítio protegido e afastado da costa) o crescimento na área
manipulada foi maior que o visto em ARENG (desprotegido e próximo a costa),
porém, ainda muito pequeno. A cobertura de P. caribaeorum não passou de 2,7% e
4% na área manipulada, em ARENG e PAB4 respectivamente, mesmo após 900

BASILIO, P.S.
63
dias do experimento. Na área controle, PAB4 também apresentou um leve
crescimento, enquanto que em ARENG foi registrado declínio.
Após 183 dias do inicio do experimento (verão), PAB4 apresentou picos de
cobertura em todos os tempos e tratamentos. O período do ano pode ter interferido
nesse resultado. De modo geral, o experimento demostrou aumentos na cobertura
em períodos próximos ao verão em ambos os sítios, sendo mais acentuado em
PAB4. Na região onde está localizado o Banco dos Abrolhos, durante os meses de
verão, as taxas de sedimentação e turbidez são reduzidas e a temperatura da água
é mais elevada (LEÃO, 2002; SEGAL E CASTRO, 2011), o que pode favorecer o
crescimento de P. caribaeorum.
Na área do experimento, observou-se que os maiores declínios na cobertura
de P. caribaeorum, tanto na área manipulada quando na controle, ocorreram no
inverno, período no qual a taxa de sedimentação aumenta, assim como a turbidez
da água. Abrolhos, no período de inverno, passa por frentes frias e chuvas que são
trazidas pelo aumento do vento sul, além de ondas que ressuspendem o sedimento
fino do fundo, o que acarreta em águas mais turvas (DUTRA et al., 2006; SEGAL et
al., 2008).
Em PAB4, o padrão de recuperação e declínio descrito acima foi mais
acentuado. A explicação para isso pode estar relacionada a morfologia dos recifes.
No Banco dos Abrolhos, os recifes afastados da costa possuem uma formação
pinacular, a qual permite maior fluxo de água entre os pináculos. Por este motivo as
perturbações meteorológicas não são tão intensas (DUTRA et al., 2006). Em
contrapartida, a morfologia de recifes próximos à costa atua como uma armadilha de
sedimento, impedindo o fluxo de água nessas áreas, onde, pelo menos uma vez por
ano, distúrbios meteorológicos abrem espaços para novas espécies bentônicas se
estabelecerem (DUTRA et al., 2006).
Apesar da resistência da P. caribaeorum à sedimentação (REIMER et al.
2006b), alguns fatores em seu desenvolvimento podem ser limitados em altos níveis
de sedimento ou taxas temporariamente persistentes, principalmente devido à
redução da luminosidade (ROGERS, 1990) e de substratos estáveis para o
assentamento e fixação das larvas (WOOLFE E LARCOMBE, 1999). Além disso, o
excesso de sedimento pode promover a permanência da larva na coluna d’água (TE,
1992), afetar a capacidade fotossintética por declínio das trocas metabólicas e

BASILIO, P.S.
64
gasosas (ABDEL-SALAM E PORTER, 1988; WEBER et al., 2012) e sufocar a
colônia (ROGERS, 1990).
Além da sedimentação, o tipo de reprodução e a ação da densidade
populacional podem ter influenciado na dinâmica de crescimento e recolonização da
P. caribaeorum na área manipulada do experimento. Palythoa caribaeorum adquire
espaço no recife através da competição com outros organismos bentônicos,
reprodução assexuada e assentamento de novas larvas. As colônias de Palythoa
aumentam de tamanho por fusão de colônias filhas e podem diminuir por fissão de
uma colônia, morte parcial e reabsorção dos pólipos (TANNER, 2000). É conhecido
que, em altas densidades, muitas espécies (e.g. briozoários e zoantídeos) tendem a
resistir melhor a perturbações, reduzindo a mortalidade e podendo se beneficiar com
as taxas de captação de alimentos, crescimento e/ou sobrevivência (BUSS, 1981;
OKAMURA, 1985; TANNER, 1997).
Em colônias pequenas (i.e. colônias recém-assentadas ou geradas por
reprodução assexuada), a energia assimilada é direcionada principalmente para o
crescimento e reprodução a assexuada (SHIROMA E REIMER, 2010). No entanto,
seria necessário o estabelecimento inicial da espécie na área manipulada, através
da colonização do substrato nu, neste caso, a reprodução sexuada e dispersão de
larvas seria a maneira mais provável de se chegar ao novo habitat.
Ainda não é claro o que promove a ativação da gametogênese, da desova e
nem o período reprodutivo entre as espécies de Palythoa, no entanto, sabe-se que
fatores abióticos locais podem atuar na frequência de pólipos férteis (BOSCOLO E
SILVEIRA, 2005). Shiroma e Reimer (2010) registraram periodicidade na
reprodução da P. tuberculosa, a qual apresenta ovos do inicio de junho ao início de
agosto, no Japão. No sudeste do Brasil existem registros de maiores percentuais de
pólipos férteis de P. caribaeorum nos meses de agosto, setembro e março, com
contínua liberação de oócitos durante o ano (BOSCOLO E SILVEIRA, 2005). No
Banco dos Abrolhos, a reprodução foi documentada no mês de março (LONGO
et.al., 2012). Após a liberação dos ovos é provável que o gasto energético passe a
ser direcionado ao estabelecimento da larva em um ambiente propício ao seu
desenvolvimento.
A taxa de recrutamento em cnidários coloniais recifais é geralmente
influênciada pelo suprimento de larvas, substrato adequado e número de mortes nas

BASILIO, P.S.
65
fases iniciais (CONNELL et al., 1997). Na maioria dos sistemas biológicos, o
assentamento tem sido maior que recrutamento real, pois, ao se estabelecer em
ambiente adverso, a larva pode mudar subtamente de habitat, através de
metamorfose invertida, ou morrer devido a predação e outros fatores (TE, 1992).
É de se esperar, portanto, que mesmo existindo recrutamento na área
manipulada (observação pessoal), a baixa densidade e o tamanho,
consideravelmente pequeno das novas colônias que conseguiram se estabelecer,
tenham sido desfavoráveis para um processo mais rápido de recolonização. Tanner
(2002) sugere que variações na cobertura devido ao recrutamento só são
detectáveis após um nível de 10 colônias.mês-1. Outros fatores agravantes na
recolonização de um ambiente é o fato de que as regiões de borda de colónias de P.
caribaeorum (proporcionalmente maiores em colônias pequenas) mostram uma
porcentagem mais elevada de pólipos estéreis, em comparação com os centros
(BOSCOLO E SILVEIRA, 2005) e a taxa de mortalidade de classes de tamanho
menores são relativamente altas, variando de 22 a 44% (TANNER, 1997).
O tamanho reduzido das novas colônias tambem faz com que ações de
predadores possam eliminar mais facilmente a espécie em determinada área. Com o
experimento, constatou-se que houve influência da gaiola na cobertura de P.
caribaeorum, quando comparados os sítios, mas não para os sítios individualmente.
Um dos motivos que pode explicar essa influência é a diferença existente entre os
sítios em relação à biomassa de peixes predadores, sendo maior nos recifes
protegidos, como é o caso de PAB4. Outra explicação seria a proliferação
diferenciada de algas epilíticas, principalmente dentro das gaiolas.
A maior taxa de crescimento observada entre os tratamentos foi, como
esperado, na área com gaiola fechada (exclusão de predadores) em PAB4,
considerando as áreas, manipulada e controle, juntas. Para os sítios
individualmente, observou-se que houve variação temporal em relação à raspagem
para ambos os sítios, entretanto não se observou efeito da exclusão dos
predadores, mostrando que a predação não influenciou diretamente a recolonização.
Para melhor compreender os fatores responsáveis pela dinâmica de
recolonização da P. caribaeorum na área do experimento, analises de vitalidades
também foram realizadas, no intuito de acrescentar informações importantes sobre o

BASILIO, P.S.
66
porquê da não recuperação da P. caribaeorum em cobertura nas áreas
manipuladas, como dito anteriormente.
No geral a porcentagem de tecido saudável em PAB4 foi menor do que em
ARENG, em todos os tempos, tanto na área manipulada quanto na área controle.
Muitos fatores podem ter influenciado a diferença na vitalidade da P. caribaeorum
entre os sítios, como: (1) variações temporais na temperatura, pluviosidade,
salinidade e turbidez; (2) diferentes características do sedimento (cf. DUTRA et al.,
2006; WEBER et al., 2012); (3) maior cobertura de corais competidores como a
Mussismilia braziliensis (VERRILL, 1868), Favia gravida Verrill, 1868 e Siderastrea
spp De Blalnville, 1830, espécies predominantes nos recifes protegidos no Banco
dos Abrolhos (cf. FRANCINI-FILHO et al., 2013) e (4) a variação na biomassa de
predadores.
Uma colônia com vitalidade reduzida desloca a energia adquirida para a
sobrevivência em vez do crescimento. Karlson (1988) sugere que a distribuição da
energia dependerá das estratégias de sobrevivência. Em ambientes com alta taxa
de sedimentação existe o comprometimento da sobrevivência, pois, além das
funções vitais, a espécie tem que administrar a retirada do sedimento de sua
superfície (DUTRA et al., 2006).
O branqueamento, assim como várias doenças, vem atingindo os ambientes
recifais (GLYNN 1993; BROWN 1997; DOUGLAS 2003; HOEGH-GULDBERG,
1999). Em ARENG a porcentagem de tecido branqueado foi o mais representativo
dentro da categoria não saudável, com picos de porcentagem no verão, quando as
temperaturas superficiais da água são mais altas (FRANCINI-FILHO et al. 2010). As
principais causas do branqueamento envolvem o aumento da radiação solar e
consequente elevação da temperatura em águas superficiais; excesso de
sedimento; redução da salinidade e agentes infeciosos (HOEGH-GULDBERG E
SMITH 1989; ROGERS 1990; GLYNN 1991; KOBLUK E LYSENKO 1994; LESSER
1996).
ARENG, por está mais próximo à costa, também está mais susceptível a
interferências antrópicas (e.g. poluição), que fazem com que a vitalidades dos recifes
decline e essas perturbações crônicas se tornem ainda mais prejudiciais
(RICHMOND, 1993; HUGHES, 1994; CONNELL et al., 1997), reduzindo, inclusive, a
capacidade do recife de se recuperar após distúrbios naturais (i.e. resiliência;

BASILIO, P.S.
67
HOLLING, 1973; BROWN, 1997; BELLWOOD et al., 2004). As altas taxas de
sedimento e descarga de nutrientes podem ser os principais responsáveis pela
grande porcentagem de tecido branqueado nesse sítio.
Ao final do experimento, após 957 dias, foi observada recuperação de tecido
saudável tanto na área manipulada quanto na área controle de ARENG. No entanto,
a recuperação da vitalidade foi relativamente menor na área dentro da gaiola
fechada. A explicação seria o sombreamento ocasionado pelo acúmulo de algas na
estrutura externa da gaiola, dificultando a assimilação de energia e/ou pela
competição direta entre algas e zoantídeos. O mesmo fator pode ter interferido em
PAB4, que apresentou trajetórias diferenciadas para os tratamentos, com aumento
claro do tecido não saudável na área com a gaiola fechada. Outro fator que pode ter
atuado nesse resultado é o fato de que em PAB4 a biomassa de predadores de
zoantídeos é maior do que em ARENG. Sebens (1982) observou que alguns
predadores de P. caribaeorum predam preferencialmente colónias fisicamente
danificadas.
A porcentagem de tecido recoberto em ARENG foi bem inferior àquela vista
em PAB4. Portanto, em PAB4, tecido branqueado e recoberto estão mais
representados dentro da categoria “não saudável”, com recobrimento principalmente
por sedimentos. Entretanto, o tipo de sedimento que recobre as colônias neste sítio
é de origem biogênica, composto por grãos finos e derivados do revolvimento do
substrato do fundo. No geral, componentes mais finos e sem enriquecimento de
matéria orgânica seriam rejeitados facilmente, evitando o dano temporário ou
permanente ao organismo, diferentemente do que ocorre em componentes com
carga orgânica alta e grãos maiores (WEBER et al., 2012).
Dutra et al. (2006) apontam que a porcentagem de CaCO3 acumulado no
sedimento em Abrolhos é proporcional a distância da costa, com sedimentos
carbonáticos (grãos finos) quase puros dominantes em recifes mais distantes da
costa (e.g. PAB4). Entretanto o que observamos em PAB4 foi a formação de uma
fina camada de sedimento sobre as colônias. Uma hipótese levantada aqui é que a
origem carbonária do sedimento em contato com o muco excretado pela P.
caribaeorum forme uma película que favorece a proliferação de microalgas e
cianobactérias (observação pessoal), com esse crescimento sendo dependente do
tempo durante o qual o sedimento se mantém sobre a colônia. O ambiente gerado

BASILIO, P.S.
68
por essa aglomeração impediria o crescimento por asfixia, redução da vitalidade,
perda das zooxantelas e até a morte da área recoberta, todas essas consequências
já foram documentadas na literatura (cf. ABDEL-SALAM E PORTER, 1988;
YOSHIOKA E YOSHIOKA, 1989; ROGERS, 1990; TE, 1992; WOOLFE E
LARCOMBE, 1999; WEBER et al., 2012). Diaz-Pulido e McCook (2002) citam as
cianobactérias como colonizadoras dominantes após eventos de branqueamento.
PAB4 apresentou uma tendência contraria na porcentagem de tecido
branqueado e recoberto, ou seja, na medida em que a porcentagem de tecido
branqueado aumentou a porcentagem de tecido recoberto diminuiu. Esse padrão
pode ser explicado pelo fato de que o tecido imediatamente abaixo das áreas
recobertas (principalmente por sedimento) já estariam branqueadas. Em corais,
Nemeth e Nowlis (2001) encontraram uma forte correlação positiva entre taxas de
sedimentação e o branqueamento.
A porcentagem de tecido saudável e não saudável da Palythoa caribaeorum
apresentou diferenças significativas durante o tempo em relação à raspagem (área
manipulada e controle) em ARENG e PAB4. Em ARENG os maiores valores de
cobertura de tecido não saudável foram registrados no tempo T183 na maioria dos
tratamentos, tanto na área controle quanto na área manipulada. Em PAB4
observou-se tendência crescente na porcentagem de tecido branqueado,
principalmente na área manipulada. Essa diferença sugere que a densidade exerce
influência na vitalidade (OKAMURA, 1985; TANNER, 1997).
As áreas manipuladas, diferente dos controles, passaram por modificações
em seu equilíbrio com a retirada da P. caribaeorum. Neste caso elas iniciam um
processo de recolonização que sofre interferência de vários fatores. A competição,
por exemplo, atua limitando o tamanho das populações de zoantídeos (FRECHETTE
et al., 1992; TANNER, 1997, 2000). Em áreas com altas densidades populacionais
as colônias são favorecidas na captura de alimento, no crescimento, na taxa de
sobrevivência (OKAMURA, 1985) e, possivelmente, a exclusão de sedimento.
Entretanto, em altas densidades, a prevalência de doenças como o branqueamento
também é maior.
A cobertura de P. caribaeorum presente nos substratos nus das áreas
manipuladas foi formada, basicamente, por recrutas e pequenas colônias. Neste

BASILIO, P.S.
69
caso, a eficiência fisiológica de algumas funções básicas à sobrevivência podem ter
sido comprometidas, ocasionado assim baixa vitalidade.
As porcentagens de tecido saudável e não saudável de P. caribaeorum
também apresentaram diferenças significativas em relação ao tempo. Algas
epilíticas, fortes competidoras por espaço, apresentam variação sazonal na
biomassa, com picos no final do verão (ATEWEBERHAN et al., 2006). Neste caso,
alterações sazonais na abundância de organismos competidores, bem como na
precipitação, temperatura e sedimentação, poderiam ter ocasionado variações na
vitalidade de P. caribaeorum.
5.3 Sucessão de organismos bentônicos
A estrutura das comunidades bentônicas nas áreas manipuladas do
experimento variou de acordo com duas fases principais: uma na qual os sítios
diferiram um do outro (inicio do experimento) e outra na qual os sítios convergiram a
um novo estado sucessional semelhante entre eles (fim do experimento) (veja Fig.
19). Neste caso, um estudo de maior duração seria necessário para avaliar se esta
convergência permaneceria ao longo do tempo ou se as comunidades de cada sítio
retornariam ao estado original registrado no início do experimento.
A cobertura de organismos bentônicos não variou significativamente em
relação ao tratamento, indicando que a estrutura das comunidades bentônicas como
um todo não foi influenciada pela exclusão dos predadores. Portanto, outros fatores
devem ter ditado a dinâmica sucessional dos locais estudados.
A cobertura de P. caribaeorum foi dominante nas áreas de estudo de ambos
os sítios, sendo maior em ARENG. Em PAB4, além da P. caribaeorum, foi registrada
uma cobertura relativamente alta de corais hermatípicos. Padrão semelhante foi
observado em estudos pretéritos realizados no Banco dos Abrolhos (FRANCINI
FILHO et al., 2010, SEGAL E CASTRO, 2011; FRANCINI-FILHO et al., 2013).
Após a manipulação, novos espaços foram gerados para colonização de
outros organismos em ambos os sítios. Segundo Mendonça-Neto et al. (2008), a
abertura de novos espaços após retirada de P. caribaeorum favorece o aumento da
riqueza em organismos bentônicos. Fatores ambientais e/ou agentes biológicos
específicos podem atuar na formação de zonas ou mosaicos (SEBENS, 1982).

BASILIO, P.S.
70
Os padrões de assentamento de larvas e a competição por espaço e
alimento após o assentamento vão agir de modo a determinar a estrutura da
comunidade em um novo habitat. Os limites de tamanho dessas populações não são
definidos exclusivamente pela área disponível, mas também pela competição
(FRECHETTE et al., 1992, TANNER, 1997, 2000).
Na competição por alimento, a reserva nutricional exerce um efeito imediato
sobre a densidade (TANNER, 2000). Portanto, muitas espécies utilizam formas
diferentes de aquisição de energia (TANNER, 2002), como é o caso da P.
caribaeorum e dos corais hermatípicos que são parcialmente autotróficos.
Os principais organismos bentônicos registrados na área do experimento
foram respectivamente: Palythoa caribaeorum, espécie alvo do estudo; algas
epilítica; algas calcárias incrustantes e corais hermatípicos, estes dois últimos sendo
os mais importantes construtores de recifes na região de Abrolhos. Essas categorias
bentônicas estão entre as mais abundantes no Banco dos Abrolhos (FRANCINI
FILHO et al., 2013).
A cobertura de coral em PAB4 se manteve superior àquela registrada em
ARENG, apresentando um pequeno aumento ao fim do experimento. Esse aumento
pode ser explicado tanto pela disponibilidade de áreas para recrutamento, como pela
maior cobertura de coral já existente na área. Connell et al. (1997) sugerem que
tanto os organismos bentônicos residentes quanto o substrato rígido disponível (i.e.
não colonizado), são fundamentais para o assentamento de larvas e sobrevivência
de recrutas de corais (HUGHES, 1985; CONNELL et al., 1997).
Muitos estudos em sistemas marinhos enfatizam a ação do recrutamento na
regulação de populações (UNDERWOOD E FAIRWEATHER 1989; HUGHES 1990)
e na recuperação do ecossistema recifais após distúrbios (HUGHES E CONNELL
1999; HUGHES E TANNER 2000). Tanner (1997) mostra que a influência do
recrutamento é pequena para zoantídeos, principalmente em parcelas com substrato
disponível e de baixa densidade, assim, o recrutamento em densidades
intermediárias de corais, como é o caso de PAB4, poderia influenciar no aumento da
população. Portanto, a menor cobertura de corais em ARENG do que em PAB4
pode ter influenciado no padrão de maior recuperação de corais em PAB4 do que
em ARENG.

BASILIO, P.S.
71
Na área manipulada de ARENG, foi notório o aumento inicial na cobertura de
algas calcárias incrustante, o qual esteve claramente ligado a exposição das
mesmas após a exclusão da P. caribaeorum. No decorrer do experimento ocorreu
um declino acentuado de algas calcárias incrustantes, provavelmente devido à
predação intensa por budiões (peixes da família Labridae) (observação pessoal;
Francini-Filho et al. 2008). Nas áreas controle, no decorrer do experimento, também
foi registrada redução da cobertura da alga calcária incrustante. De acordo com
Francini-Filho et al. (2013), a cobertura de alga calcária incrustante diminuiu
drasticamente nos topos de recifes costeiros desprotegidos. Estes mesmos autores
registraram relação negativa entre a cobertura de alga calcaria incrustante e P.
caribaeorum, sugerindo uma concorrência entre eles. Em PAB4, a tendência
observada foi de pequeno aumento da cobertura dessas algas, o que sugere uma
melhor condição de equilíbrio competitivo neste último sítio.
Com o tempo, foi registrado aumento acentuado na cobertura de algas
epilíticas, principalmente em ARENG, na área manipulada, este fator pode estar
ligado a não proteção do sítio e ao elevado fluxo de nutrientes (SEGAL E CASTRO,
2011), devido a sua maior proximidade com a costa. Ambientes desprotegidos estão
susceptíveis a perturbações antrópicas, como descarga de nutrientes e sobrepesca
de herbívoros (FRANCINI-FILHO et al., 2013), o que pode ser crítico para a
manutenção da estrutura e função dos recifes de coral, pois a presença dos
herbívoros reduz a abundância de algas, deixando o substrato livre para a
colonização de outros organismos (BURKEPILE E HAY, 2008) (ex.: organismos
construtores).
A persistência na proliferação de algas, principalmente macroalgas, leva à
redução da capacidade de recuperação de ecossistemas recifais após perturbações
intensas (BELLWOOD et al., 2004; FRANCINI-FILHO et al., 2013). Essa é conhecida
como uma das primeiras fases de degradação nos ambientes recifais, visto que a
área disponível para o recrutamento é ocupada por algas e sedimento associado,
reduzindo as chances dos recrutas se estabelecerem no substrato (HUGHES 1994).
Bellwood et al. (2004) destacam que várias outras transições já foram
documentadas, mas a questão a ser resolvida é: até que ponto cada estado
alternativo é estável ou reversível? Na elaboração de estratégias de conservação e
gestão, esses modelos podem agir de maneira a auxiliar ao retorno do estado mais

BASILIO, P.S.
72
estável do ambiente em questão, voltando ao domínio desejável por corais e outros
organismos construtores (FRANCINI FILHO et al., 2010).
Francini-Filho et al. (2013) observaram que no período de 2006 a 2008 foi
visto um aumento significativo na cobertura de algas epilítica para todo o Banco de
Abrolhos, incluindo parte do período de execução dos experimentos presentes
(2006-2009). O crescimento excessivo de algas pode causar a mortalidade parcial
da P. caribaeorum (SOTO, 2007). Os resultados, aqui encontrados mostraram que
na área manipulada do experimento, a cobertura de alga epilítica foi superior em
relação às demais categorias nos dois sítios, sobretudo em ARENG. Os principais
motivos disso foram a disponibilidade de substrato presente nas áreas pós-exclusão
da P. caribaeorum e a capacidade competitiva e colonizadora das algas epilítica em
áreas abertas.
Outro fator que pode ter influenciado no padrão sucessional das
comunidades bentônicas nas áreas experimentadas foram as variações nas taxas de
sedimentação e o tipo de sedimento presente e cada sítio. Tanto o soterramento de
outros organismos bentônicos como a redução na disponibilidade de luz
(consequências da sedimentação), podem influencia no crescimento dos organismos
bentônicos (ACEVEDO E MORELOCK, 1988).
No Banco dos Abrolhos, estudos já mostraram que a sedimentação pode ser
determinante no desenvolvimento dos recifes e que a distribuição das espécies
bentônicas está intimamente ligada à variação sazonal nas taxas ressuspensão e
aporte de sedimentos (DUTRA et al., 2006; SEGAL E CASTRO, 2011).
Recifes de coral saudáveis apresentam normalmente taxas de sedimentação
de 10 mg.cm-2.dia-1 (ROGERS, 1983, 1990), neste sentido, pode-se dizer que
Abrolhos está dentro dos limites esperados para recifes saudáveis, mostrando taxa
sedimentar máxima de (10.7 ± 2.0 mg cm-2 day -1) e mínima de (3.3 ± 1.9 mg cm-2
day -1) (SEGAL E CASTRO, 2011).
Alguns corais hermatípicos, por exemplo, em ambientes com alta taxa
sedimentar, tem a respiração reduzida e grande parte de sua energia é utilizada na
produção de muco, metabolicamente muito caro, para defesa contra sedimentação
(RIEGL E BRANCH, 1995), reduzindo assim o crescimento. Estes corais são,
portanto, predominantes nos recifes mais afastados da costa, com é o caso de PAB4

BASILIO, P.S.
73
(SEGAL E CASTRO, 2011), que sofre menor influência do aporte sedimentar
terrígeno.
Não apenas a taxa sedimentar, mas a composição do sedimento (SEGAL E
CASTRO, 2011), e o modo como se apresentam, seja em suspensão ou
transportado rentes ao substrato (SOTO, 2007), podem afetar de maneiras
diferentes as comunidades bentônicas. Os recifes próximos da costa em Abrolhos
são predominantes em sedimento terrígeno carregados de nutrientes (VILLAÇA E
PITOMBO, 1997) e elevada concentração de metais (AMADO-FILHO et al., 1997)
provenientes das descargas fluviais. Nos recifes mais afastados da costa, a
predominância é de sedimento biogênico (SEGAL E CASTRO, 2011), ou seja,
provenientes de carapaças e esqueletos de plantas e animais marinhos.
Estudos com radioisótopos solúveis mostram que este sedimento presente
nas áreas distantes da costa em Abrolhos é proveniente da ressuspensão de
sedimento do fundo e não do escoamento costeiro, como os encontrados em recifes
próximos à costa (SEGAL et al., 2008). Neste caso, a perturbação é inferior em
intensidade e em frequência (DUTRA et al., 2006).
A turbidez proveniente de sedimentos em suspensão pode reduzir a
diversidade e impedir o crescimento dos organismos bentônicos (TORRES, 2001).
Em Abrolhos, essa taxa aumenta no inverno e é inversamente proporcional à
distância da costa (SEGAL E CASTRO, 2011). Nesses ambientes a alocação de
energia pode representar redução na sobrevivência de organismos bentônicos, pois
interfere na assimilação de nutrientes tanto por fotossíntese como pela captura pelos
pólipos, a pouca energia assimilada terá que ser administrada, além das funções
vitais, na remoção de sedimento (DUTRA et al., 2006).
Apesar de não aparecer nas analises, pois geralmente estavam associadas
às algas epilítica, as cianobactérias estavam representadas na área de estudo,
portanto é importante mostrar a ação dessa categoria nos recifes. Assim como as
algas epilíticas, as cianobactéria conseguem se estabelecer em camadas de
sedimento formadas no substrato, algumas espécies podem agir como epífitas ou
psâmicas (CRISPINO E SANT’ANNA, 2006), proliferando-se como tapetes em
ambientes enriquecidos de nutrientes (GLIBERT et al., 2004; PAUL et al., 2005).
Em Abrolhos, as cianobactérias foram comumente vistas associadas ao
sedimento depositado, inclusive sobre colônias de P. caribaeorum recobertas,

BASILIO, P.S.
74
principalmente em PAB4 (observação pessoal). Nesse caso, elas podem promover
a morte do organismo imediatamente abaixo pela formação de um ambiente anóxico
em seu talo (JANSON et al., 1994; KUFFNER E PAUL, 2004), além de inibir o
assentamento de larvas em um raio de até 0,55m (MAIDA et al., 1995; KUFFNER E
PAUL, 2004). Como característica positiva, algumas espécies podem agir como
simbiontes fixadores de nitrogênio em corais (LESSER et al., 2004).
No sítio PAB4, em decorrência de sua condição de ambiente protegido,
encontramos uma área mais equilibrada, o que favorece o desenvolvimento da
biodiversidade de organismos, acarretando em dinâmicas ecológicas que já se
encontram bem estabelecidas, diferente do que ocorre em ARENG.

BASILIO, P.S.
75
6. CONCLUSÃO

BASILIO, P.S.
76
6. CONCLUSÃO
P. caribaeorum domina topos fundos e próximos a costa.
Variáveis ambientais, como profundidade e distância da costa, são mais
importantes do que variáveis bióticas (i.e predação) na dinâmica de P.
caribaeorum.
Apesar da cobertura de P. caribaeorum apresentar relação negativa com a
distância da costa, as taxas de recolonização não foram maiores no sítio
mais próximo a costa.
A cobertura de P. caribaeorum aumenta em períodos próximos ao verão, em
ambos os sítios.
A baixa densidade de Palythoa foi desfavorável para um processo mais
rápido de recolonização.
Foi identificado duas fases sucessionais principais: uma na qual os sítios
diferiram um do outro (inicio do experimento) e outra na qual os sítios
convergiram a um novo estado sucessional semelhante (fim do experimento).
Neste caso, um estudo de maior duração seria necessário para avaliar
se esta convergência permaneceria ao longo do tempo ou se as
comunidades de cada sítio retornariam ao estado original registrado
no início do experimento.
Em abrolhos, inúmeros fatores agiram de maneira interligada, atuando tanto
na dinâmica de recolonização da P. caribaeorum como na dinâmica
sucessional de ambientes nus, diferenciando os ambientes e os processos
que neles ocorrem.

BASILIO, P.S.
77
7. REFERÊNCIAS
ABDEL-SALAM, H. A.; PORTER, J. W. Physiological effects of sediment rejection on photosynthesis and respiration in three Caribbean reef corals. Proceedings of the 6th International Coral Reef Symposium, v. 2, p.
285-292 (1988). ACEVEDO, R.; MORELOCK J. Effects of terrigenous sediment influx on coral
reefs zonation in southwestern Puerto Rico. Proceedings of the 6th International Coral Reef Symposium, Australia, 1988. v.2, p. 189-194 (1988).
ACOSTA, A. Disease in Zoanthids: Dynamics in space and time.
Hydrobiologia, v. 460, p. 113-130 (2001). ACOSTA, A.; SAMMARCO, P.; W. DUARTE L. F. New fission processes in the
zoanthid Palythoa caribaeorum: Description and quantitative aspects. Bulletin of Marine Sciences, v.76, n.1, p. 1-26 (2005).
AMADO-FILHO, G. M.; ANDRADE, L. R.; REIS, R. P.; BASTOS, W. and W. C.
Heavy metal concentrations in seaweed species from the Abrolhos reef region, Brazil. Proceedings of the 8 International Coral Reef Symposium (Panamá), p. 1843-1846 (1997).
AMADO-FILHO, G. M.; MOURA, R. L.; BASTOS, A. C.; SALGADO, L. T.;
SUMIDA, P. Y.; GUTH, A. Z. ; FRANCINI-FILHO, R. B.; PEREIRA-FILHO, G. H.; ABRANTES, D. P.; BRASILEIRO, P. S. ; BAHIA, R. G.; LEAL, R. N.; KAUFMAN, L. ; KLEYPAS, J. A.; FARINA, M.; THOMPSON, F. L. Rhodolith beds are major CaCO3 bio-factories in the Tropical South West Atlantic. PLoS ONE, v. 7, n. 4 (2012).
AMIR, I.; HARRIS, J.B.; ZAR, M. A. The effect of palytoxin on neuromuscular
junctions in the anococcygeus muscle of the rat. Journal of Neurocytology, v. 26, p. 367–376 (1997).
ATEWEBERHAN, M.; BRUGGEMANN, J. H.; BREEMAN, A. M. Effects of
extreme seasonality on community structure and functional group dynamics of coral reef algae in the southern Red Sea (Eritrea). Coral Reefs, v. 25, p. 391–406 (2006).
AULT, T. R.; C. R. JOHNSON. Spatially and temporally predictable fish
communities on coral reefs. Ecological Monographs, n. 68, p. 25-50 (1998).
BASTIDAS, C.; BONE, D. Competitive strategies between Palythoa
caribaeorumand Zoanthus sociatus (Cnidaria: Anthozoa) at a reef flat environment in Venezuela.Bulletin of Marine Science, v. 59, n. 3, p. 543-

BASILIO, P.S.
78
555 (1996). BAUMAN, A. G.; BURT, J. A.; FEARY, D.A.; MARQUIS, E.; USSEGLIO, P.
Tropical harmful algal blooms: An emerging threat to coral reef communities? Marine Pollution Bulletin. 60, 2117–2122 (2010).
BELLWOOD, D. R.; HUGHES, T. P.; FOLKE, C.; NYSTRÖM, N. Confronting
the coral reef crisis. Nature, v. 429, n. 24, p. 827-833 (2004). BENAYAHU, Y.; LOYA, Y. Competition for space among coral-reef sessile
organisms at eilat, Red Sea. Bulletin of Marine Science, v. 31, n. 3, p.
514-522 (1981). BOSCOLO, H. K; SILVEIRA, F. L. Reproductive biology of Palythoa
caribaeorum and Protopalythoa variabilis (Cnidaria, Anthozoa, Zoanthidea) from the southeastern coast of Brazil. Brazilian Journal of Biology, v. 65, p. 29-41 (2005).
BRADBURY, R. H.; YOUNG, P. C. The effects of a major forcing function,
wave energy, on a coral reef ecosystem. Marine Ecology Progress Series, v. 5, p. 229-241 (1981).
BROWN, B. E. Coral bleaching: causes and consequences. Coral Reefs v. 16,
p. 129-138 (1997).
BRUNO, J.F.; SELIG, E.R. Regional decline of coral cover in the Indo-Pacific: timing, extent, and subregional comparisons. PLoS ONE 2(8): e711 (2007). [doi:10.1371/journal.pone.0000711]
BURKEPILE, D. E.; HAY, M. E. Impact of herbivore identity on algal
succession and coral growth on a Caribbean reef. PLoS ONE, v. 5, n. 1 (2010).
BURNETT, W. J.; BENZIE, J. A. H.; BEARDMORE, J. A.; RYLAND, J. S.
Zoanthids (Anthozoa, Hexacorallia) from the Great Barrier Reef and Torres Strait, Australia: systematics, evolution and a key to species. Coral Reefs,
v. 16, p. 55-68 (1997). BUSS, L. W. Group living, competition, and the evolution of cooperation in a
sessile invertebrate. Science, v. 213, p. 1012-1014 (1981).
CLARKE, K. R.; WARWICK, R. M. Change in marine communities: an
approach to statistical analysis and interpretation, 2nd edition. PRIMER-E, Plymouth (2001).
CONNELL, J. H. Diversity in tropical rain forests and coral reefs. Science. v.
199, n. 24, p. 1302-1310 (1978). CONNELL, J. H.; HUGHES, T. P.; WALLACE, C. C. A 30-year study of coral

BASILIO, P.S.
79
abundance, recruitment, and disturbance at several scales in space and time. Ecological Monographs, v. 67, p. 461-488 (1997).
CONNELL, S. D. The relationship between large predatory fish and recruitment
and mortality of juvenile coral reef-fish on arti-ficial reefs. Journal of Experimental Marine Biology and Ecology, v. 209, p. 261-278 (2002).
COSTA JR, O. S.; LEÃO, Z. M. A. N; NIMMO, M; ATTRIL, M. Nutrification
impacts on coral reefs from northern Bahia, Brazil. Hydrobiologia v. 440,
p. 307-315 (2000). CÔTÉ, I. M.; GILL, J. A.; GARDNER, T. A.; WATKINSON, A. R. Measuring
coral reef decline through meta-analyses. Philosophical Transactions of the Royal Society B., v. 360, p. 385–395 (2005).
CRISPINO, L. M. B.; SANT’ANNA C. L. Cianobactèrias marinhas bentùnicas
de ilhas costeiras do Estado de São Paulo, Brasil. Revista Brasileira de Botânica, v.29, n.4, p.639-656 (2006).
DALY, M.; BRUGLER, M. B.; CARTWRIGHT, P.; COLLINS, A. G.; DAWSON,
M. N.; FAUTIN, D. G.; FRANCE, S. C.; MCFADDEN, C. S.; OPRESKO, D. M.; RODRIGUEZ, E.; ROMANO, S. L.; STAKE, J. L. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa, v. 1668 p. 127–182 (2007).
DE’ATH, G.; LOUGH, J. M.; FABRICIUS, K. E. Declining coral calcification on
the Great Barrier Reef. Science, v. 323 (2009).
DEFINIENS DEVELOPER GmbH. eCognition user guide. Definiens Imaging
GmbH, Munich, Germany, (2009). DIAZ-PULIDO, G.; MCCOOK, L. J. The fate of bleached corals: patterns and
dynamics of algal recruitment. Marine Ecology Progress, v. 232, p. 115–
128 (2002). DONE, T. J. Phase shifts in coral reef communities and their ecological
significance. Hydrobiologia 247,121–132 (1992).
DONNER, S. D. ; SKIRVING, W. J.; LITTLE, C. M.; OPPENHEIMER, M.;
HOEGH-GULDBERG, O. Global assessment of coral bleaching and required rates of adaptation under climate change Global Change Biology, v. 11, p. 2251–2265, (2005).
DOUGLAS, A.E. Coral bleaching––how and why? Marine Pollution Bulletin,
v. 46, p. 385–392 (2003). DUTRA, L. X. C. Os efeitos do aporte de sedimento na vitalidade dos
recifes de corais de Abrolhos, Bahia. 2003. 91p. Dissertação de (Mestrado em Geologia), Instituto de Geociências, Universidade Federal

BASILIO, P.S.
80
da Bahia, Salvador (2003). DUTRA L. X. C.; KIKUCHI R. K. P.; LEÃO Z. M. A. N. Effects of sediment
accumulation on reef corals from Abrolhos, Bahia, Brazil.Journal of Coastal Research Special, v. 39, p. 633-638 (2006).
FABRICIUS, K. E. Effects of terrestrial runoff on the ecology of corals and coral
reefs: review and synthesis. Marine Pollution Bulletin, v. 50, p. 125-146 (2005).
FABRICIUS, K. E.; WILD, C.; WOLANSKI, E.; ABELE, D. Effects of
transparent exopolymer particles and muddy terrigenous sediments on the survival of hard coral recruits. Estuarine, Coastal, and Shelf Science, v.
57, p. 613-621 (2003). FAUTIN, D.G. Hexacorallians of the world (2006). Disponível em:
<http://geoportal.kgs.ku.edu/hexacoral/anemone2/index.cfm> Acesso em: 17 fev 2013.
FAUTIN, D. G. Structural diversity, systematics, and evolution of cnidae.
Toxicon, v. 54, p. 1054–1064 (2009).
FERREIRA, C. E. L; FLOETER, S. R.; GASPARINI, J. L.; JOYEUX, J. C.;
FERREIRA, B. P. Trophic structure patterns of brazilian reef fishes: a latitudinal comparison. Journal of Biogeography, v. 31, p. 1093–1106
(2004). FIGUEIREDO, M. A. O. Colonization and growth of crustose coralline algae in
Abrolhos, Brazil. Proceedings of the 8th International Coral Reef Symposium, v. 1, p. 689-694 (1997).
FRANCINI-FILHO, R. B.; MOURA, R. L. Dynamics of fish assemblages on
coral reefs subjected to different management regimes in the Abrolhos Bank, eastern Brazil. Aquatic Conservation: Marine and Freshwater Ecosystems v. 18, p. 166-1179 (2008a).
FRANCINI-FILHO, R. B.; MOURA, R. L. Evidence for spillover of reef fishes
from a no-take marine reserve: An evaluation using the before-after control-impact (BACI) approach. Fisheries Research, v.93, p. 346-356
(2008b). FRANCINI-FILHO, R. B.; MOURA, R. L. Predation on the toxic zoanthid
Palythoa Caribaeorum by reef fishes in the Abrolhos Bank, eastern Brazil. Brazilian Journal Of Oceanography, v. 58, n. 1, p. 77-79 (2010).
FRANCINI-FILHO, R. B.; MOURA, R. L.; FERREIRA, C. M.; CONI, E. O. C.
Live coral predation by parrotfishes (Perciformes: Scaridae) in the Abrolhos Bank, eastern Brazil, with comments on the classification of species into functional groups. Neotropical Ichthyology, v.6, p. 191-200 (2008).

BASILIO, P.S.
81
FRANCINI-FILHO, R.B. ; REIS, R. ; MEIRELLES, P. ; MOURA, R. ;
THOMPSON, F. ; KIKUCHI, R. ; KAUFMAN, L. . Seasonal prevalence of white plague like disease on the endemic Brazilian reef coral Mussismilia braziliensis. Latin American Journal of Aquatic Research, v. 38, p. 292-
296, 2010. FRANCINI-FILHO, R. B.; FERREIRA, C. M.; CONI, E. O. C.; MOURA, R. L.;
KAUFMAN, L. Foraging activity of roving herbivorous reef fish (Acanthuridae and Scaridae) in eastern Brazil: influence of resource availability and interference competition. Journalof the Marine Biological Associationof the United Kingdom. v. 90, p. 481-492 (2010).
FRANCINI-FILHO, R. B.; CONI, E. O. C.; MEIRELLES, P. M.; AMADO-FILHO,
G. M.; THOMPSON, F. L.; PEREIRA-FILHO, G. H.; BASTOS, A. C.; ABRANTES, D. P.; FERREIRA, C. M.; GIBRAN, F. Z.; GÜTH, A. Z.; SUMIDA, P. Y. G.; OLIVEIRA, N. L.; KAUFMAN, L.; MINTE-VERA, C. V.; MOURA, R. L. Dynamics of coral reef benthic assemblages of the Abrolhos Bank, Eastern Brazil: inferences on natural and anthropogenic drivers. PLoS ONE, v. 8, n. 1 (2013).
FRECHETTE, M.; AITKEN, A. E.; PAGE, L. Interdependence of food and
space limitation of a benthic suspension feeder: Consequences for self-thinning relationships. Marine Ecology Progress Series, v. 83, p. 55-62 (1992).
FRELIN, C.; VAN RENTERGHEM, C. Palytoxin Recent electrophysiological
and pharmacological evidence for several mechanisms of action. General Pharmacology, v, 26, p. 33-37 (1995).
FROESE, R.; PAULY, D. FishBase (2011). Disponível em:
<www.fishbase.org> Acesso em: 04 nov 2012. GARDNER, T. A.; CÔTÉ I. M.; GILL, J. A.; GRANT, A.; WATKINSON, A. R.
Long-term region-wide declines in Caribbean corals. Science, v. 301, p.
958–960 (2003). GHERARDI, D. F. M. & BOSENCE, D. W. J., Composition and community
structure of the coralline algal reef from Atol das Rocas, South Atlantic, Brazil. Coral Reefs, v. 19, p. 205-219 (2001).
GILI, J. M.; COMA, R. Benthic suspension feeders: their paramount role in
littoral marine food webs. Trends in Ecology & Evolution, v. 13, p. 316-
321 (1998). GILMOUR, J. Experimental investigation into the effects of suspended
sediment on fertilisation, larval survival and settlement in a scleractinian coral. Marine Biology, v. 135, p. 451–462 (1999).

BASILIO, P.S.
82
GLIBERT, P. M.; HEIL, C. A.; HOLLANDER, D.; REVILLA, M.; HOARE, A.; ALEXANDER, J.; MURASKO, S. Evidence for dissolved organic nitrogen and phosphorus uptake during a cyanobacterial bloom in Florida Bay. Marine Ecology Progress Series, v. 280, p. 78–83 (2004).
GLYNN, P. W. Coral reef bleaching in the 1980s and possible connections with
global warming. Trends in Ecology & Evolution, v. 6, p. 175-179 (1991).
GRATWICKE, B.; SPEIGHT, M. R. The relationship between fish species
richness, abundance and habitat complexity in a range of shallow tropical marine habitats. Journal of Fish Biology, v. 66, p. 650-667 (2005).
HAYWICK, D. W.; MUELLER, E. M. Sediment retention in encrusting
Palythoaspp. – a biological twist to a geological process. Coral Reefs, v. 16 p. 39–46 (1997).
HODGSON, G. Tetracycline reduces sedimentation damage to corals. Marine
Biology, v. 104, p. 493–496 (1990). HOEGH-GULDBERG, O. Climate change, coral bleaching and the future of the
worldís coral reefs. Marine and Freshwater Research, v. 50, p. 839-66
(1999). HOEGH-GULDBERG, O.; SMITH, G. J. The effect of sudden changes in
temperature, irradiance and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata (Esper 1797) and Seriatopora hystrix (Dana 1846). Journal of Experimental Marine Biology and Ecology, v. 129, p. 279-303 (1989).
HOEGH-GULDBERG, O.; MUMBY, P. J.; HOOTEN, A. J.; STENECK, R. S.;
GREENFIELD, P.; GOMEZ, E.; HARVELL, C. D.; SALE, P. F.; EDWARDS, A. J.; CALDEIRA, K.; KNOWLTON, N.; EAKIN, C. M.; IGLESIAS-PRIETO, R.; MUTHIGA, N.; BRADBURY, R.H.; DUBI, A.; HATZIOLOS, M. E. Coral reefs under rapid climate change and ocean acidification. Science 318:1737-1742 (2007).
HOLLING, C.S., Resilience and stability of ecological sys-tems. The Annual
Review of Ecology, Evolution, and Systematics, v. 4, p. 1-23 (1973). HUBBARD, J. A. E. B.; POCOCK, Y. P. Sediment rejection by recent
scleractinian corals: a key to palaeo-environmental reconstruction. Geologische Rundschau, v. 61, n. 2, p. 598-626 (1972). Disponivel em :<http://link.springer.com/article/10.1007%2FBF01896337>
HUGHES,T. P. Life histories and population dynamics of early successional
corals. Proceedings of the 6th International Coral Reef Symposium, v. 4. p. 101-106 (1985).
HUGHES, T. P. Recruitment limitation, mortality, and population regulation in

BASILIO, P.S.
83
open systems: A case study. Ecology, v. 71, p. 12-20 (1990).
HUGHES, T. P. Catastrophes, phase shifts, and large-scale degradation of a
Caribbean coral reef. Science, v. 265, p. 1547 –1551 (1994). HUGHES, T. P.; CONNELL, J. H. Multiple stressors on coral reefs: a long-term
perspective. limnology and oceanography, part 2: The effects of multiple stressors on freshwater and marine ecosystems. American Society of Limnology and Oceanography, v. 44, n. 3, p. 932-940 (1999).
HUGHES, T. P.; TANNER, J. E. Recruitment failure, life histories, and long-
term decline of caribbean corals. Ecological Society of America, v. 81, n. 8, p. 2250-2263 (2000).
HUGHES, T. P.; BAIRD, A. H.; BELLWOOD, D. R.; CARD, M.; CONNOLLY, S.
R.; FOLKE, C. Climate change, human impacts, and the resilience of coral reefs. Science, v. 301, p. 929–933 (2003).
JACKSON, J. B. C.; KIRBY, M. X.; BERGER, W. H.; BJORNDAL, K. A.;
BOTSFORD, L. W.; BOURQUE, B. J.; BRADBURY, R. H.; COOKE, R.; ERLANDSON, J. ; ESTES, J. A. ; HUGHES, T. P.; KIDWELL, S.; LANGE, C. B.; LENIHAN, H. S.; PANDOLFI, J. M.; PETERSON, C. H.; STENECK, R. S.; TEGNER, M. J.; WARNER, R. R. Historical overfishing and the recent collapse of coastal ecosystems. Science, v. 293, p. 629-638 (2001).
JANSON, S.; CARPENTER, E. J.; BERGMAN, B. Compartmentalisation of
nitrogenase in a non-heterocystous cyanobacterium: Trichodesmium contortum. FEMS Microbiology Letters, v. 118, p. 9–14 (1994).
KARLSON, R. H. Size-dependant growth in two zoanthid species: a contrast in
clonal strategies. Ecology, v. 69, p. 1219-1232 (1988).
KEMP, D. W.; COOK, C. B.; LAJEUNESSE, T. C.; BROOKS, W. R. A
comparison of the thermal bleaching responses of the zoanthid Palythoa caribaeorum from three geographically different regions in south Florida. Journal of Experimental Marine Biology and Ecology, v. 335, p. 266–276, (2006).
KLEYPAS, J. A. Coral reef development under naturally turbid conditions:
fringing reefs near Broad Sound, Australia. Coral Reefs, v. 15, p. 153-167, 1996.
KLIPPEL, S.; OLAVO, G.; COSTA, P. A. S.; MARTINS, A. S.; PERES, M. B.
Avaliação dos estoques de lutjanídeos da costa central do Brasil: análise de coortes e modelo preditivo de Thompson e Bell para comprimentos. In: COSTA, P. A. S.; MARTINS, A. S.; OLAVO, G.(Eds.) Pesca e potenciais de exploração de recursos vivos na região central da Zona Econômica Exclusiva brasileira. Rio de Janeiro: Museu Nacional. p. 83-98 (Série Livros n.13) (2005).

BASILIO, P.S.
84
KOBLUK, D. R.; LYSENKO, M. A. ‘‘Ring’’ bleaching in southern Caribbean
Agaricia agaricites during a rapid water cooling. Bulletin of Marine Science, v. 54, p. 142–150 (1994).
KOHLER, K. E.; GILL, S. M. Coral point count with excel extensions (CPCe): A
visual basic program for the determination of coral and substrate coverage using random point count methodology. Computers Geoscices, v. 32, p. 1259-1269 (2006).
KUFFNER, I. B.; PAUL, V. J. Effects of the benthic cyanobacterium Lyngbya
majusculaon larval recruitment of the reef corals Acropora surculosa and Pocillopora damicornis. Coral Reefs, v. 23, p. 455–458 (2004).
LEÃO, Z. M. A. N. Abrolhos, BA - O complexo recifal mais extenso do Atlântico
Sul. Em: SCHOBBENHAUS, C.; CAMPOS, D.A.; QUEIROZ, E.T.; WINGE, M.; BERBERT-BORN, M.L.C (Eds.). Sátios Geolágicos e Paleontologicos do Brasil. 1ed. Brasília: DNPM/CPRM - Comissão Brasileira de Sítios Geológicos e Paleobiologicos (SIGEP), v. 1, p. 345-359 (2002).
LEÃO, Z. M. A. N. ; DOMINGUEZ, J. M. L. Tropical coast of Brazil. Marine
Pollution Bulletin, v. 41, n. 1-6, p. 112-122 (2000).
LEÃO, Z. M. A. N.; KIKUCHI, R. K. P.; MAIA, M. P.; LAGO, R. A. L. A
catastrophic coral cover decline since 3000 years. Northern Bahia, Brazil. Proceedings 8th International Coral Reef Symposium, v. 1, p. 583–588
(1997). LEÃO, Z. M. A. N.; KIKUCHI, R. K. P.; TESTA, V. Corals and coral reefs of
Brazil. In: CORTÉS, J. (Ed.). Latin America Coral Reefs. Amsterdam:
Elsevier Publisher, p. 9-52 (2003). LESSER, M. P. Exposure of symbiotic dinoflagellates to elevated temperatures
and ultraviolet radiation causes oxidative stress and inhibits photosynthesis. Limnology and oceanography, v. 41, p. 271–283 (1996).
LESSER , M. P.; MAZEL, C. H.; GORBUNOV, M. Y.; FALKOWSKI, P. G.
Discovery of symbiotic nitrogen-fixing cyanobacteria in corals. Science, v.
305, p. 997–1000 (2004). LONGO, G.O.; KRAJEWSKI, J.P.; SEGAL, B.; FLOETER, S.R. First record of
predation on reproductive Palythoa caribaeorum (Anthozoa: Sphenopidae): insights on the trade-off between chemical defences and nutritional value. Marine Biodiversity. v. 5; p. 1-3 (2012).
LOYA, Y. Effects of water turbidity and sedimentation on the community
structutre of Puerto Rican corals. Bulletin of Marine Science, v. 26, n. 4, p. 450-466, (1976).

BASILIO, P.S.
85
LOYA, Y.; KRAMARSKY-WINTER, E. In situ eutrophication caused by fish
farms in the northern Gulf of Eilat (Aqaba) is beneficial for its coral reefs: a critique. Marine Ecology Progress Series, v. 261, p. 299–303 (2003).
MAIDA, M.; FERREIRA, B. P. Coral reefs of Brazil: overview and field guide.
Proceedings 8th International Coral Reef Symposium, p. 263–274
(1997). MAIDA, M.; SAMMARCO, P. W.; COLL, J. C. Effects of soft corals on
scleractinian coral recruitment. I. Directional allelopathy and inhibition of settlement. Marine Ecology Progress Series, v. 121, p. 191-202 (1995).
MCCLANAHAN, T. R.; KAUNDA-ARARA, B. Fishery recovery in a coral-reef
marine park and its effect on the adjacente fishery. Conservation Biology,
v. 10, p. 1187–1199 (1996). MENDONÇA-NETO, J. P.; GAMA, B. A. P.; The native Palythoa caribaeorum
overgrows on invasive species in the intertidal zone. Coral Reefs, v. 28, p.
497 (2009). MENDONÇA-NETO, J. P.; FERREIRA, C. E. L.; CHAVES, L. C. T.; PEREIRA,
R. C. Influence of Palythoa caribaeorum (Anthozoa, Cnidaria) zonation on site-attached reef fishes. Anais da Academia Brasileira de Ciências, v. 80, n. 3. p. 495-513 (2008).
MINTE-VERA, C. V.; MOURA, R. L.; FRANCINI-FILHO, R. B.; Nested
sampling: an improved visual-census technique for studying reef fish assemblages. Marine Ecology Progress Series, v. 367 p. 283-293
(2008). MOBERG, F; FOLKE, C. Ecological goods and services of coral reef
ecosystems. Ecological Economics, v. 29, p. 215-233 (1999).
MOORE, R. E.; SCHEUER, P. J. Palytoxin: A new marine toxin from a
coelenterate. Science, v. 172, p. 495-496 (1971). MOURA, R. L. Brazilian reefs as priority areas for biodiversity conservation in
the Atlantic Ocean. Proceedings of the 9th International Coral Reef
Symposium, v. 2, p. 917-920 (2000). MOURA, R. L.; FRANCINI-FILHO, R. B. Reef and shore fishes of the Abrolhos
Region, Brazil. In: DUTRA, G. F.; ALLEN, G. R.; WERNER, T.; MCKENNA, A. S. (eds) A rapid marine biodiversity assessment of the Abrolhos Bank, Bahia, Brazil. RAP Bulletin of Biological Assessment 38, Washington, p.
45-55 (2006). MOURA, R. L., MINTE-VERA, C. V., CURADO, I. B., FRANCINI-FILHO, R. B.,
RODRIGUES, H. C. L., DUTRA, G. F., ALVES, D. C., SOUTO, F. J. B.,

BASILIO, P.S.
86
Challenges and prospects offisheries co-management under a marine extractive reserve framework in Northeastern Brazil. Coastal Management, v. 37, p. 617–632 (2009).
MOURA, R. L.; FRANCINI-FILHO, R. B. ; CHAVES, E. M. ; MINTE-VERA, C.
V. ; LINDEMAN, K. C. Use of riverine through reef habitat systems by dog snapper (Lutjanus jocu) in eastern Brazil. Estuarine, Coastal and Shelf Science, v. 95, p. 274-278 (2011)
MOURA, R. L.; SECCHIN, N. A.; AMADO-FILHO, G. M.; FRANCINI-FILHO, R.
B.; FREITAS, M. OLIVEIRA; MINTE-VERA, C. V.; TEIXEIRA, J. B.; THOMPSON, F. L.; DUTRA, G. F.; SUMIDA, P. Y. G.; GUTH, A. Z.; LOPES, R. M.; BASTOS, ALEX C. Spatial patterns of benthic megahabitats and conservation planning in the Abrolhos Bank, Continental Shelf Research (2013).
NEMETH, S. R.; NOWLIS, J. S. Monitoring the effects of land development on
the near-shore reef environment of St. Thomas, USVI. Bulletin of Marine Science, v. 69, p. 759 -775 (2001).
NORSTRÖM, A. V.; NYSTRÖM, M.; LOKRANTZ, J.; FOLKE, C. Alternative
states on coral reefs: beyond coral–macroalgal phase shifts. Marine Ecology Progress Series, v. 376, p. 295–306 (2009).
NZALI, L. M.; JOHNSTONE, R. W.; MGAYA, Y. D. Factors affecting
scleractinian coral recruitment on a nearshore reef in Tanzania. Ambio, v. 27, n. 8, p. 717-72, (1998).
OKAMURA, B. The effects of ambient flow velocity, colony size and upstream
colonies on the feeding success of Bryozoa. 11.Conopeum reticulum, an encrusting species. Journal of Experimental Marine Biology and Ecology, v. 89, p. 69-80 (1985).
PANDOLFI, J. M.; BRADBURY, R. H.; SALA, E.; HUGHES, T. P.; BJORNDAL,
K. A.; COOKE, R. G.; MCARDLE, D.; MCCLENACHAN, L.; NEWMAN, J. H.; PAREDES, G.; WANER, R. R.; JACKSON, J. B. C. Gobal trajectories of the long-term decline of coral reef ecosystems. Science, v. 30, n. 1, p.
955-958 (2003). PASTOROK, R. A.; BILYARD, G. R. Effects of sewage pollution on coral-reef
communities. Mar Marine Ecology Progress Series, v. 21, p. 175-189
(1985). PAUL, V. J. ; THACKER, R. W. ; BANKS, K. GOLUBIC, S. Benthic
cyanobacterial bloom impacts the reefs of South Florida (Broward County, USA). Coral Reefs, v. 24, p. 693–697 (2005).
PEREIRA, M. D.; SIEGLE, E.; MIRANDA, L. B.; SCHETTINI, C. A. F.;
Hidrodinâmica e transporte de material particulado em suspensão sazonal

BASILIO, P.S.
87
em um estuário dominado por maré: Estuário De Caravelas (Ba). Revista Brasileira de Geofísica, v. 28, n. 3, p. 427-444 (2010).
PÉREZ, C. D.; VILA-NOVA, D. A.; SANTOS, A. M. Associated community with
the zoanthid Palythoa caribaeorum (Duchassaing & Michelotti, 1860) (Cnidaria, Anthozoa) from littoral of Pernambuco, Brazil. Hydrobiologia, v. 548, p. 207-215 (2005).
PHILIPP, E.; FABRICIUS, K. Photophysiological stress in scleractinian corals
in response to short-term sedimentation. Journal of Experimental Marine Biology and Ecology, v. 287, p. 57–78 (2003).
QUINN, F., Competitive hierarchies in marine benthic communities.
Oecologia, v. 54, p. 129-135 (1982). RABELO, E. F.; SOARES, M. D. O.; MATTHEWS-CASCON, H. Competitive
interactions among zoanthids (cnidaria: zoanthidae) in an intertidal zone of northeastern Brazil. Brazilian Journal of Oceanography, v. 61, p. 35-42 (2013).
RANDALL, J. E.; Food habits of reef Wshes of the West Indies. Studie in
Tropical Oceanography, v. 5, p. 665–847 (1967). REIMER, J. D.; ONO, S.; TAKISHITA, K.; TSUKAHARA, J.; MARUYAMA, T.
Molecular evidence suggesting species in the zoanthid genera Palythoa and Protopalythoa (Anthozoa: Hexacorallia) are congeneric. Zoological science, v. 23, p. 87-94 (2006a).
REIMER, J. D.; TAKISHITA, K.; MARUYAMA, T. Molecular identification of
symbiotic dinoflagellates (Symbiodinium spp.) from Palythoa spp. (Anthozoa: Hexacorrallia) in Japan. Coral Reefs, v. 25, p, 521-527
(2006b). REIMER, J. D.; ONO, S.; TSUKAHARA, J.; IWASE, F. Molecular
characterization of the zoanthid genus Isaurus (Anthozoa: Hexacorallia) and associated zooxanthellae (Symbiodinium spp.) from Japan. Marine Biology, v. 153, p. 351–363 (2008).
REIMER, J. D.; NAKACHI, S.; HIROSE, M.; HIROSE, E.; HASHIGUCHI, S.
Using hydrofluoric acid for morphological investigations of zoanthids (Cnidaria: Anthozoa): a critical assessment of methodology and necessity. Marine Biotechnology, v. 12, p. 605-617 (2010).
RIEGL, B. Effects of sand deposition on scleractinian and alcyonacean corals.
Marine Biology, v. 121, p. 517-526 (1995).
RIEGL, B.; BRANCH G. M. Effects of sediment on the energy budgets of
four scleractinian (Bourne 1900) and five alcyonacean (Lamouroux 1816) corals Bernhard. Journal of Experimental Marine Biology and

BASILIO, P.S.
88
Ecology, v. 186, p. 259-275 (1995).
ROBERTS, C. M. Selecting marine reserves locations: optimality versus
opportunism. Bulletin of Marine Science, v. 66, p. 581-592 (2000). ROBERTS, C. M.; POLUNIN, N. V. C. “Marine Reserves: Simple Solutions To
Managing Complex Fisheries?”.Ambio, v. 22, p. 363–368 (1993).
ROGERS, C. S. Sublethal and lethal effects of sediments applied to common
Caribbean reef corals in the field. Marine Pollution Bulletin, v. 4, n. 3, p. 378-382 (1983).
ROGERS, C. S. Responses of coral reefs and reef organisms to
sedimentation. Marine Ecology Progress Series, v. 62, p. 185-202 (1990).
SANDIN, S. A.; SMITH, J. E.; DEMARTINI, E. E.; DINSDALE, E. A.; DONNER,
S. D.; FRIEDLANDER, A. M.; KONOTCHICK, T.; MALAY, M.; MARAGOS, J. E.; OBURA, D.; PANTOS, O.; PAULAY, G.; RICHIE, M.; ROHWER, F.; SCHOROEDER, R. E.; WALSH, S.; JACKSON, J. M.; KNOWLTON, N.; SALA, E. Baselines and Degradation of Coral Reefs in the Northern Line Islands. PLoS ONE, v.3, n. 2 (2008).
SCHEFFER, M.; CARPENTER, S.; FOLEY, J, A.; FOLKE, C.; WALKER, B.
Catastrophic shifts in ecosystems. Nature 413,591–596 (2001).
SCHEFFER, M.; CARPENTER, S.; YOUNG, B. Cascading effects of
overfishing marine systems. Tree v. 20, p. 579-581 (2005). SCHUTTE, V.G.W; SELIG, E.R.; BRUNO, J.F. Regional spatio-temporal trends
in Caribbean coral reef benthic communities. Marine Ecology Progress Series, v. 402, p. 115–122 (2010).
SEBENS, K. P. Autotrophic and heterotrophic nutrition of coral reef zoanthids.
Proceedings 3rd International Coral Reef Symposium. v. 1, p. 397-404
(1977). SEBENS, K. P. Intertidal distribution of zoanthids on the Caribbean coast of
Panama: effects of predation and desiccation. Bulletin of Marine Sciences, v. 32, p. 316–335 (1982).
SEGAL, B.; CASTRO, C. B.; Coral community structure and sedimentation at
different distances from the coast of the Abrolhos Bank, Brazil. Brazilian Journal of Oceanography, v. 59, n. 2, p. 119-129 (2011).
SEGAL, B.; EVANGELISTA, H.; KAMPEL, M.; GONCALVES, A. C.; POLITO,
P. S.; SANTOS, E. A. Potential impacts of polar fronts on sedimentation processes at Abrolhos coral reef (South-West Atlantic Ocean/Brazil). Continental Shelf Research, v. 28, p. 533-544 (2008).

BASILIO, P.S.
89
SHEPPARD, C. R. C. Coral populations on reef slopes and their major
controls. Marine Ecology Progress Series, v. 7, p. 83-115 (1982).
SHIROMA, E.; REIMER, J. D. Investigations into the Reproductive Patterns,
Ecology, and Morphology in the zoanthid genus Palythoa (Cnidaria: Anthozoa: Hexacorallia) in Okinawa, Japan. Zoological Studies, v. 49, n.
2, p. 182-194 (2010).
SILVA, A.S. ; LEÃO, Z.M.A.N; KIKUCHI, R.K.P. ; COSTA, A.B. ; SOUZA,
J.R.B. Sedimentation in the coastal reefs of Abrolhos over the last decades. Continental Shelf Research (No Prelo),
http://dx.doi.org/10.1016/j.csr.2013.06.002i SINNIGER, F.; OCAÑA, O. V.; BACO, A. R. Diversity of Zoanthids (Anthozoa:
Hexacorallia) on Hawaiian Seamounts: Description of the Hawaiian Gold Coral and Additional Zoanthids. PLoS ONE, v. 8, n.1 (2013).
SOARES, C. L. S.; PÉREZ, C. D.; MAIA, M. B. S.; SILVA, R. S.; MELLO, L. F.
A. Avaliação da atividade antiinflamatória e analgésica do extrato bruto hidroalcoólico do zoantídeo Palythoa caribaeorum (Duchassaing & Michelotti, 1860). Revista Brasileira de Farmacognosia, v. 16, n. 4 (2006).
SOROKIN, Y. I. Biomass, metabolic rates and feeding of some common reef
zoantharians and octocorals. Australian Journal of Marine & Freshwater Research, v. 42, p. 729–741 (1991).
SOTO, V. A. Effects of sedimentation on the distribution and ecology of
the reef zoanthid Palythoa caribaeorum. Tese University Of Puerto Rico. 83p. (2007).
SPALDING, S. M.; RAVILIOUS, C.; GREEN, E. P. World Atlas of Coral Reefs.
California: University of California Press, 424 p. (2001). STAMPAR, S. N.; SILVA, P. FRANCISCO DA; LUIZ-JR., O. J. Predation on
the Zoanthid Palythoa caribaeorum (Anthozoa, Cnidaria) by a Hawksbill Turtle (Eretmochelys imbricata) in Southeastern Brazil. Symposium on Sea Turtle Biology and Conservation. n. 117, p. 3 (2007).
SUCHANEK, T. H.; GREEN, D. J. Interespecific competition between Palythoa
caribaeorum and other sessile invertebrates on St. Croix reefs, U.S. Virgin Islands. Proceedings of the 4th International Coral Reef Symposium, v.
2, p. 679-684 (1981) TANNER, J. E. The effects of density on the zoanthid Palythoa caesia.
Journalof Animal Ecology, v. 66, p. 793–810 (1997).
TANNER, J. E. Density dependent population dynamics in clonal organisms: a

BASILIO, P.S.
90
modelling approach. Journal of Animal Ecology, v. 66, p. 390–399
(1999). TANNER, J. E. Stochastic density-dependence regulates population size in a
benthic marine invertebrate. Oecologia, v. 122, p. 514–520 (2000).
TANNER, J. E. Consequences of density-dependent heterotrophic feeding for
a partial autotroph. Marine Ecology Progress Series, v. 227, p. 293–304 (2002).
TE, F. T. Response to higher sediment loads by Pocillopora damicornis
planulae. Coral Reefs, v. 11, p. 131-134 (1992). TITLYANOV, E. A.; T. V. TITLYANOVA. Reef-building corals - symbiotic
autotrophic organisms: 1. General structure, feeding pattern, and light-dependent distribution in the shelf. The Russian Journal of Marine Biology, v. 28, p. S1–S15 (2002).
TORRES, J. L. Impacts of sedimentation on the growth rates of Montastrea
annularis in Southwest Puerto Rico. Bulletin of Marine Science, v. 69, p. 631-637 (2001).
UNDERWOOD, A. J.; FAIRWEATHER, P. G. Supply side ecology and benthic
marine assemblages. Trends in Ecology & Evolution, v. 4, p. 16-19 (1989).
VAN KATWIJK, M. M.; MEIER N. E; VAN LOON, R.; VAN HOVE, E. M.;
GIESEN, W. B. J. T.; VAN DER VELDE, G.; DEN HARTOG, C. Sabaki River sediment load and coral stress: correlation between sediments and condition of the Malindi-Watamu reefs in Kenya (Indian Ocean). Marine Biology, v. 117, p. 675-683 (1993).
VILLAÇA, R.; PITOMBO, F. B. Benthic communities of shallow-water reefs of
Abrolhos, Brazil. Revista Brasileira de Oceanografia, v. 45, p. 35-43 (1997).
WEBER, M.; BEER, D.; LOTT, C.; POLERECKY L.; KOHLS, K.; ABED, R. M.
M.; FERDELMAN, T. G.; FABRICIUS, K. E. Mechanisms of damage to corals exposed to sedimentation. Proceedings of the National Academy of Sciences, v. 109, n. 24 p. 1558–1567 (2012).
WILLIAMS, E. H.; BUNKLEY-WILLIAMS, L. Bleaching of Caribbean coral reef
symbionts. Proceedings of the 6th International Coral Reef Symposium, v. 3, p. 313-318 (1988).
WOOLFE, K. J.; LARCOMBE, P. Terrigenous sedimentaion and coral reef
growth: a conceptual framework. Marine Geology, v. 155, p. 331-345
(1999).

BASILIO, P.S.
91
ZAR, J.H. Biostatistical Analysis, 4th edn. Prentice-Hall: New Jersey, p 363,
(1999).