a fotossíntese
DESCRIPTION
Pequeno livro de apontamentos sobre fotossínteseTRANSCRIPT

Apontamentos de Fisiologia Vegetal (PLANTAS VASCULARES)
A Fotossíntese
Alexandra Rosa da Costa
Universidade de Évora Dezembro de 2001




A Fotossíntese
v
Notas prévias
1. Este trabalho é uma colectânea de apontamentos, sem pretensões a trabalho de revisão bibliográfica sobre o tema. Apenas se foram coligindo e modernizando os apontamentos para as aulas ao longo dos anos de ensino. Assim, por vezes é uma mera tradução de partes de livros, mas que se espera ser útil a alunos de língua portuguesa. 2. O trabalho foi paginado para ser impresso em frente e verso, com margens espelhadas. 3. Para poder melhorar, agradeço que o eventual leitor me envie as suas críticas, sugestões e correcções de erros que possa detectar para:

A Fotossíntese
vi

A Fotossíntese
vii
ÍNDICE
Página
Notas prévias v
1. INTRODUÇÃO 1
1.1. A RADIAÇÂO SOLAR 2
1.2. O PROCESSO GLOBAL 7
2. AS REACÇÕES DIRECTAMENTE DEPENDENTES DA LUZ 9
2.1. OS CLOROPLASTOS 9
2.2 OS PIGMENTOS RESPONSÁVEIS PELA ABSORÇÃO DA LUZ 11
2.2.1. As clorofilas 12
2.2.2. Os carotenóides 12
2.2.3. As ficobilinas 14
2.3. A ABSORÇÃO DE FOTÕES 15
2.4. ESPECTROS DE ABSORÇÃO 21
2.5. O EFEITO EMERSON 23
2.6. OS FOTOSSISTEMAS 23
2.6.1. A estrutura dos fotossistemas 23
2.6.2. A organização dos complexos nos tilacóides 26
2.6.3. A distribuição de energia entre os fotossistemas 28
2.7. O TRANSPORTE ELECTRÓNICO 31
2.7.1. O esquema em Z 31
2.7.2. A acção do fotossistema II 32
2.7.2.1. A redução das quinonas 32
2.7.2.2. A oxidação da água 34
2.7.3. A acção do citocrómio b6f 35
2.7.4. A acção da plastocianina 36
2.7.5. A acção do fotossistema I 37
2.7.6. O transporte cíclico de electrões 39
2.8. A SÍNTESE DE ATP 39
2.8.1. O acoplamento do transporte electrónico e da síntese de ATP in vivo
39
2.8.2.O mecanismo quimiosmótico da síntese de ATP 41
2.8.2.1. A força motriz protónica (pmf) 41
2.8.2.2. A ATPsintase 42
2.8.2.3. O mecanismo de síntese do ATP 44

A Fotossíntese
viii
ÍNDICE (Continuação)
Página
3. A REDUÇÃO FOTOSSINTÉTICA DO CARBONO 45
3.1. O CICLO DE CALVIN-BENSON 45
3.1.1. A descrição do Ciclo 47
3.1.2. A regulação do Ciclo 50
3.1.2.1. Activação pelo ião Mg2+ e pelo pH 51
3.1.2.2. A regulação pela luz 51
3.1.3. A actividade de oxigenase da Rubisco e a fotorrespiração 53
3.2. O METABOLISMO FOTOSSINTÉTICO EM C4 55
3.2.1. A anatomia de Kranz 55
3.2.2. A ultraestrutura cloroplastidial 57
3.2.3. O metabolismo básico 58
3.2.4. As variações ao metabolismo básico 59
3.2.5. A regulação enzimática 62
3.2.6. O significado biológico desta via fotossintética 63
3.3. O METABOLISMO ÁCIDO DAS CRASSULÁCEAS (CAM) 65
4. A FOTOSSÍNTESE E A DIFUSÃODO CO2 68
5. COMPARAÇÃO ENTRE OS TRÊS TIPOSDE METABOLISMO 76
5.1. EXEMPLOS DE ESPÉCIES EM C3, C4 E CAM 76
5.2. CONDIÇÕES AMBIENTAIS NATURAIS 77
5.3. ANATOMIA FOLIAR 77
5.4. ESTRUTURA DOS CLOROPLASTOS 77
5.5. ENZIMAS FIXADORAS DE CO2 77
5.6. PRODUTOS PRIMÁRIOS DA FIXAÇÃO DO CO2 77
5.7. NECESSIDADES FOTOQUÍMICAS POR CO2 FIXO 77
5.8. EFEITOS DA INTENSIDADE DA LUZ 78
5.9 DISPONIBILIDADE EM CO2 81
5.10. TEMPERATURA 83
5.11. SENSIBILIDADE À CONCENTRAÇÃO EM O2 85
5.12. FOTORRESPIRAÇÃO 86
5.13. VALORES MÁXIMOS PARA A FOTOSSÍNTESE LÍQUIDA 86
5.14. TAXA DE CRESCIMENTO MÁXIMO 86
5.15. PRODUÇÃO ANUAL DE MATÉRIA SECA 86
5. 16. TAXA DE SAÍDA DE FOTOASSIMILADOS DAS FOLHAS 87
5.17. COEFICIENTE TRANSPIRATÓRIO 87
5.18. INFLUÊNCIADA IDADE DAS FOLHAS 88
5.19. TRANSLOCAÇÃO DE CARBOIDRATOS 88
5.20. CARÊNCIA HÍDRICA 89
BIBLIOGRAFIA 91

1
A FOTOSSÍNTESE
1. INTRODUÇÃO
A fotossíntese é o processo através do qual organismos vivos convertem a energia
da luz na energia química de moléculas orgânicas. Este processo aumenta a energia livre
total disponível para os organismos e directa ou indirectamente fornece a energia para todo
o mundo vivo (Lawlor, 1987).
A radiação solar é a fonte de energia para todos os processos biológicos através da
fotossíntese. Sem a energia da luz para mudar a matéria dum estado energético mais baixo
para outro mais elevado a vida não seria possível. Esta mudança de estado energético
corresponde ao rearranjar de electrões em moléculas e à criação de ligações químicas.
De acordo com as leis da termodinâmica os processos biológicos tendem a ir dum
estado de maior para outro de menor energia, excepto se houver energia disponível para
levar a reacção em direcção contrária. Os organismos vivos estão num estado
termodinâmico instável e requerem energia para manterem os constituintes químicos numa
certa ordem e para realizarem trabalho contra o gradiente de energia termoquímico, ao
acumularem matéria, tal como iões ou gases provenientes do ambiente, ou ao crescerem,
ou ao moverem-se, etc. (Lawlor, 1987).
O movimento da matéria, a interconversão química ou as mudanças de estado
energético não ocorrem com eficiência absoluta, pelo que existe uma certa perda de
energia, geralmente na forma de calor. Assim que um sistema biológico tiver acumulado
energia livre, pode convertê-la a diferentes formas químicas, ou em energia física, ou ainda
permutá-la entre organismos, mas com o tempo a energia útil será perdida e atingir-se-á o
equilíbrio, isto é, a morte (Lawlor, 1987).
Apenas organismos capazes de utilizar a energia proveniente do Sol podem
aumentar a energia total do mundo vivo e serem independentes das limitações impostas por
outras fontes de energia. A fotossíntese realizada por certas bactérias, e pelas plantas é
levada a cabo por um mecanismo capaz de captar a energia fugaz duma partícula de luz e
torná-la disponível para reacções bioquímicas (Lawlor, 1987).

A Fotossíntese
2
1.1. A RADIAÇÃO SOLAR:
Através duma série de reacções nucleares que ocorrem dentro do Sol, a sua
massa é convertida em energia de acordo com a relação formulada por Einstein:
2mcE
Devido a esta conversão de massa em energia, o Sol mantém uma
temperatura superficial extremamente elevada, e assim, irradia uma grande
quantidade de energia para o espaço. Parte dessa energia incide na Terra, mas
apenas uma pequena fracção da energia incidente é absorvida pelas plantas. Esta
absorção inicia um fluxo de energia através da biosfera, isto é, todos os seres vivos e
a parte da Terra que habitam (Nobel, 1991).
A radiação solar é uma radiação electromagnética emitida quando um dípolo
(um par de cargas, uma negativa e outra positiva separadas por uma pequena
distância) num átomo oscila causando uma mudança no campo de força. O dípolo
produz um vector eléctrico e um magnético (figura 1), que estão em fase, mas
formando um ângulo recto. A onda electromagnética é caracterizada quer pelo
comprimento de onda, (em metros) que é a distância entre dois picos sucessivos,
quer pela frequência, , que é o número de oscilações por unidade de tempo, em s-1
(Lawlor, 1987).
A frequência é determinada pelas oscilações do dípolo. O comprimento de
onda e a frequência estão relacionados com a velocidade de propagação da onda, v
(em m s-1). NOTA: A velocidade da radiação no vácuo (c) é de 3 x 10-8 m s-1:
v ou c
Os comprimentos de onda entre os 350 e os 750 nm são visíveis ao olho
humano, e são chamados luz (figura 2). A radiação que interessa à fotossíntese
(PAR = Photosynthetic Active Radiation) tem comprimentos de onda entre os 400 e
os 700 nm. Quando falamos de luz estamos a falar da radiação que causa uma
reacção no olho humano, e o termo deveria ser usado apenas nesse contexto. No
entanto, a simplicidade da palavra e a semelhança entre os comprimentos de onda,
justifica que se use o termo “luz” para descrever a PAR (Lawlor, 1987).
A luz para além de ter natureza ondulatória tem natureza corpuscular (em
partículas). Em 1900 Planck resolveu o conflito entre o conceito ondulatório e o
corpuscular, considerando que a luz se comporta como pequenas porções de
energia, chamadas quanta que apenas podem ser absorvidas ou emitidas pela

A Fotossíntese
3
matéria em unidades indivisíveis. A partícula que transporta o quantum de energia é
chamada fotão, e não tem massa.
Figura 1: A luz pode ser representada por uma onda electromagnética correspondente às oscilações
dos campos eléctricos e magnéticos locais. O comprimento de onda () é a distância entre dois pontos sucessivos na mesma fase, ao longo da onda. Retirado de Nobel (1991), figura 4.1, página 193
Quantum e fotão são conceitos diferentes, o primeiro é a energia carregada pelo
fotão. As transições moleculares causadas pela luz envolvem um “salto” ou transição
no estado energético de electrões e só podem ocorrer se toda a energia dum fotão
for captada; se o quantum for maior ou mais pequeno que a energia necessária para
a transição, então o fotão não é absorvido. Assim, os processos que envolvem luz
são processos “tudo ou nada”. A energia dum fotão () depende da frequência da
onda electromagnética, que está relacionada com o comprimento de onda, e é dada
pela relação (Lawlor, 1987):
/hch
Em que h é a constante de Planck (6.62 x 10-34 J s-1).
Pela equação podemos ver que quanto maior for a frequência (), maior é a
energia () e menor o comprimento de onda ().
Um único fotão é em termos biológicos uma unidade pequena; a meio dum
dia ensolarado, a superfície terrestre recebe um máximo de 1,3 x 1021 fotão m-2 s-1.
Assim, é necessário utilizar uma unidade de radiação maior: o mole de fotões ou
mais correctamente um mole de quanta. Isto é frequentemente chamado um Einstein,
embora esta unidade não seja aceite pelo Sistema Internacional (SI) (Lawlor, 1987).

A Fotossíntese
4
Existe alguma confusão sobre as várias maneiras de se medir a luz, porque
tanto o número de quanta, ou a sua energia, ou ainda ambos, podem ser
determinados. Para estudar os aspectos quantitativos ou cinéticos da resposta à luz
de processos químicos ou biológicos só é aceite o número de fotões em determinada
região espectral (ou a energia correspondente). O número de fotões incidente
perpendicularmente numa superfície é dado pela densidade de fluxo fotónico
(mol m-2 s-1, ou em termos energéticos tendo em consideração a gama de c.d.o. J m-2
s-1 , como J s-1 = 1 Watt (W), teremos W m-2).
Figura 2: Representação esquemática do espectro electromagnético.
Retirado de Jones (1992), figura 2.1, página 10
A iluminância, isto é, a impressão visual no olho humano, é dada pelo fluxo
luminoso (lux = lx = lúmen m-2). Nos trabalhos mais antigos se utilizava-se muito o
“foot candle” (fc = 10 764 lx), mas actualmente esta unidade já não é aceite (Lawlor,
1987).
A densidade de fluxo radiante que atinge perpendicularmente a camada mais
externa da atmosfera terrestre, isto é, a constante solar é de cerca de 1368 W m-2.
Baseados na constante solar e feita a média para todas as latitudes, a densidade de
fluxo radiante médio por dia que atinge perpendicularmente a camada mais externa
da atmosfera terrestre é de 29.6 MJ m-2 dia-1. A presença de nuvens faz com que, em
média, apenas cerca de 58 % atinja a superfície terrestre, ou seja, 17.0 MJ m-2 dia-1.
No Verão em dias sem nuvens, a uma latitude média, este valor pode ser de cerca
de 30 MJ m-2 dia-1 . A irradiância instantânea na região do visível, a meio dum dia
sem nuvens com o Sol perpendicular, pode ser de 420 W m-2 (Lawlor, 1987).
A figura 3 mostra o número relativo dos fotões solares que chegam à
atmosfera terrestre (a vermelho) e à sua superfície em função do comprimento de
onda. Cerca de 5 % dos fotões que chegam à atmosfera terrestre estão no
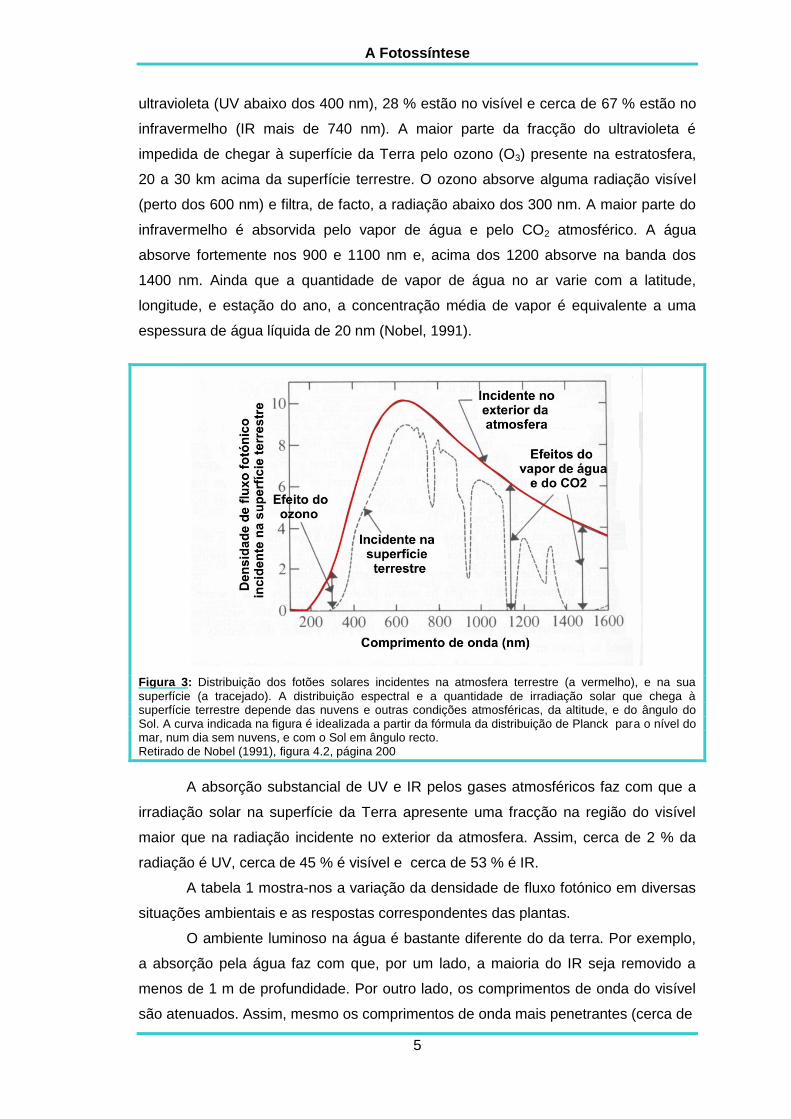
A Fotossíntese
5
ultravioleta (UV abaixo dos 400 nm), 28 % estão no visível e cerca de 67 % estão no
infravermelho (IR mais de 740 nm). A maior parte da fracção do ultravioleta é
impedida de chegar à superfície da Terra pelo ozono (O3) presente na estratosfera,
20 a 30 km acima da superfície terrestre. O ozono absorve alguma radiação visível
(perto dos 600 nm) e filtra, de facto, a radiação abaixo dos 300 nm. A maior parte do
infravermelho é absorvida pelo vapor de água e pelo CO2 atmosférico. A água
absorve fortemente nos 900 e 1100 nm e, acima dos 1200 absorve na banda dos
1400 nm. Ainda que a quantidade de vapor de água no ar varie com a latitude,
longitude, e estação do ano, a concentração média de vapor é equivalente a uma
espessura de água líquida de 20 nm (Nobel, 1991).
Figura 3: Distribuição dos fotões solares incidentes na atmosfera terrestre (a vermelho), e na sua
superfície (a tracejado). A distribuição espectral e a quantidade de irradiação solar que chega à superfície terrestre depende das nuvens e outras condições atmosféricas, da altitude, e do ângulo do Sol. A curva indicada na figura é idealizada a partir da fórmula da distribuição de Planck para o nível do mar, num dia sem nuvens, e com o Sol em ângulo recto. Retirado de Nobel (1991), figura 4.2, página 200
A absorção substancial de UV e IR pelos gases atmosféricos faz com que a
irradiação solar na superfície da Terra apresente uma fracção na região do visível
maior que na radiação incidente no exterior da atmosfera. Assim, cerca de 2 % da
radiação é UV, cerca de 45 % é visível e cerca de 53 % é IR.
A tabela 1 mostra-nos a variação da densidade de fluxo fotónico em diversas
situações ambientais e as respostas correspondentes das plantas.
O ambiente luminoso na água é bastante diferente do da terra. Por exemplo,
a absorção pela água faz com que, por um lado, a maioria do IR seja removido a
menos de 1 m de profundidade. Por outro lado, os comprimentos de onda do visível
são atenuados. Assim, mesmo os comprimentos de onda mais penetrantes (cerca de

A Fotossíntese
6
Tabela 1: Variação da densidade de fluxo fotónico e as correspondentes respostas das plantas.
Retirado de Hay & Fitter (2002), Tab.2.2, página8
500 nm) e com a água mais transparente possível, apenas cerca de 1 % do fluxo
radiante incidente na superfície da água consegue penetrar a 200 m (Nobel, 1991).
A distribuição dos comprimentos de onda dos fotões que atingem a superfície
terrestre influencia fortemente a vida. Por exemplo, a forte absorção do UV pelo
ozono reduz o efeito mutagénico desta radiação. Antes de existir ozono nas camadas
superiores da atmosfera a radiação UV afectou fortemente os processos genéticos.

A Fotossíntese
7
Mesmo agora, a exposição prolongada a radiação UV causa inibição da fotossíntese
e um decréscimo da expansão foliar. É igualmente interessante notar que a radiação
que atinge a superfície terrestre tem um pico nos 680 nm, que é como veremos
mais adiante, coincidente com o pico de absorção das clorofilas. A visão humana
também utiliza a região de comprimentos de onda que chega em maior proporção à
Terra. Podemos portanto, pensar que a pressão selectiva favoreceu a evolução de
sistemas fotoquímicos capazes de utilizar os comprimentos de onda mais
abundantes, evitando os UV que são demasiados energéticos, e os IR que são
demasiado fracos (Nobel, 1991).
1.2. O PROCESSO GLOBAL:
A fotossíntese, que ocorre em todos os organismos contendo clorofila, é a
síntese de carboidratos utilizando luz, dióxido de carbono e água, provenientes do
meio ambiente, com libertação de oxigénio. neste processo há um ganho de energia
nas ligações químicas dos carboidratos:
química energia 22clorofila
22 O)OCH(OHCO luz da energia
Esta reacção requer a energia na forma de “ligações de alta energia” do
composto adenosina trifosfato (ATP), que se pode observar na figura 4, e do poder
redutor da nicotinamida adenina dinucleótido fosfato na forma reduzida (NADPH)
que se pode observar na figura 5. Estes dois compostos são sintetizados por
processos bioquímicos conduzidos pela energia da luz. As transformações de
energia e material envolvem muitos passos (possivelmente milhares), se forem
considerados todos os processos necessários para formar o organismo inteiro
(Lawlor, 1987).
Todos os processos fotossintéticos envolvem oxidações e reduções. Uma
redução é a transferência de um electrão (e-) ou de um electrão e um protão (H+) de
uma molécula dadora (D) para uma molécula aceitadora (A). O dador é oxidado e o
aceitador é reduzido. Quando se transferem electrões para um aceitador
electricamente neutro, este ao reduzir-se fica carregado negativamente e pode
aceitar um protão para restabelecer a neutralidade eléctrica. Na água, ou nas
soluções aquosas diluídas onde ocorrem a fotossíntese e as restantes reacções
metabólicas, existem protões disponíveis em grande quantidade (Lawlor, 1987):
D + A D+ + A + H+ + e- D+ +A
Se o aceitador for um composto oxidado a adição de um electrão e- reduz o
aceitador sem transferência de um protão:

A Fotossíntese
8
D + A+ D+ + A+ + e- D+ + A
Estas reacções de oxidação redução (redox) que ocorrem nos sistemas
biológicos são geralmente catalisadas por enzimas.
As reacções de oxidação-redução têm uma importância fundamental na
compreensão dos mecanismos da fotossíntese. A reacção primária da fotossíntese,
que liga a energia física das moléculas de clorofila excitadas pela absorção de fotões
com os processos bioquímicos, é a transferência de electrões duma clorofila de tipo
especial para um aceitador de electrões. O aceitador fica reduzido e a molécula
especial de clorofila fica oxidada. Então, electrões provenientes doutro composto são
transferidos para a molécula de clorofila oxidada, reduzindo-a e permitindo que o
processo se repita (Lawlor, 1987).
Figura 4: Estrutura da adenosina 5´-trifosfato (ATP) e dos nucleótidos com ela relacionados (ADP =
adenosina 5´-difosfato; AMP 0 adenosina 5´-monofosfato). Retirado de Goodwin e Mercer (1983), figura 6.1, página 163
As reacções de oxidação-redução têm uma importância fundamental na
compreensão dos mecanismos da fotossíntese. A reacção primária da fotossíntese,
que liga a energia física das moléculas de clorofila excitadas pela absorção de fotões
com os processos bioquímicos, é a transferência de electrões duma clorofila de tipo
especial para um aceitador de electrões. O aceitador fica reduzido e a molécula

A Fotossíntese
9
especial de clorofila fica oxidada. Então, electrões provenientes doutro composto são
transferidos para a molécula de clorofila oxidada, reduzindo-a e permitindo que o
processo se repita (Lawlor, 1987).
A fotossíntese pode ser dividida em duas etapas: as reacções
directamente dependentes da luz - fase à luz - em que a energia da luz vai reduzir
o NADP+ a NADPH e permitir a fosforilação do ADP em ATP; e as reacções
indirectamente dependentes da luz - fase às escuras - em que os compostos
formados na primeira fase vão reduzir uma variedade de compostos orgânicos e
inorgânicos.
Figura 5: Estrutura do NADP+ (à esquerda) e de NADPH (à direita). A parte da molécula de NADP
+ que
sofre redução, o anel nicotinamida, está no interior da linha a tracejado. Neste processo há passagem dum electrão para o átomo de azoto da nicotinamida, neutralizando a sua carga positiva, e o segundo electrão é adicionado ao átomo de carbono superior, como parte dum átomo de hidrogénio. Retirado de Salisbury e Ross (1991), figura 10.11, página 220
2. AS REACÇÕES DIRECTAMENTE DEPENDENTES DA LUZ - FASE À
LUZ
2.1. OS CLOROPLASTOS:
A maior parte dos cloroplastos pode ser vista por microscopia óptica, mas a
sua ultraestrutura só pode ser estudada por microscopia electrónica. Cada
cloroplasto está rodeado por uma membrana dupla que controla o movimento de
A Fotossíntese
10
moléculas para dentro e para fora do organito. Dentro do cloroplasto está outro
sistema de membranas que contém os pigmentos fotossintéticos.
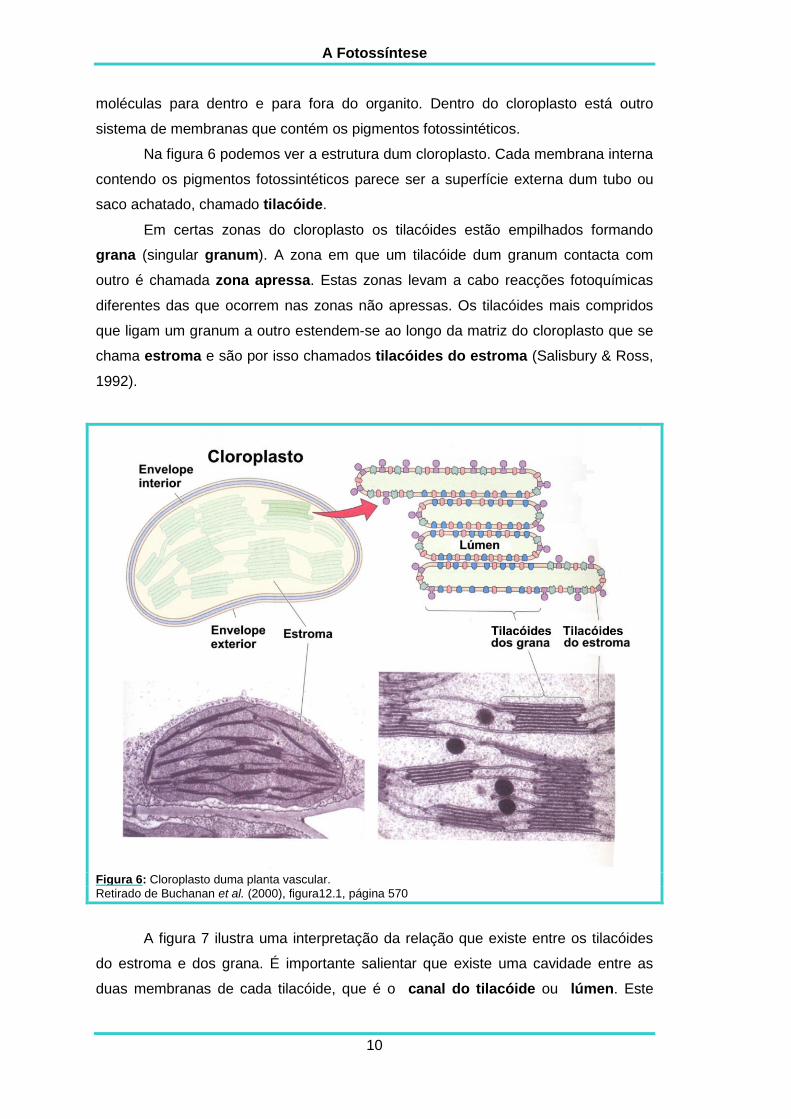
Na figura 6 podemos ver a estrutura dum cloroplasto. Cada membrana interna
contendo os pigmentos fotossintéticos parece ser a superfície externa dum tubo ou
saco achatado, chamado tilacóide.
Em certas zonas do cloroplasto os tilacóides estão empilhados formando
grana (singular granum). A zona em que um tilacóide dum granum contacta com
outro é chamada zona apressa. Estas zonas levam a cabo reacções fotoquímicas
diferentes das que ocorrem nas zonas não apressas. Os tilacóides mais compridos
que ligam um granum a outro estendem-se ao longo da matriz do cloroplasto que se
chama estroma e são por isso chamados tilacóides do estroma (Salisbury & Ross,
1992).
Figura 6: Cloroplasto duma planta vascular.
Retirado de Buchanan et al. (2000), figura12.1, página 570
A figura 7 ilustra uma interpretação da relação que existe entre os tilacóides
do estroma e dos grana. É importante salientar que existe uma cavidade entre as
duas membranas de cada tilacóide, que é o canal do tilacóide ou lúmen. Este

A Fotossíntese
11
canal está preenchido com água e sais minerais e desempenha uma função muito
importante na fotossíntese (Salisbury & Ross, 1992).
2.2. OS PIGMENTOS RESPONSÁVEIS PELA ABSORÇÃO DA LUZ:
Há vários tipos de pigmentos capazes de absorver a luz. Apenas algumas
formas especiais de clorofila a formam centros de reacção. Todos os outros
pigmentos são pigmentos acessórios, formando “antenas” para captar os fotões
que permitirão a excitação dos centros de reacção. Um grupo de muitas moléculas,
de vários tipos, duma “antena” fornece energia a um único centro de reacção. Os
vários pigmentos absorvem luz em diferentes partes do espectro o que permite que
um dado organismo absorva luz de vários comprimentos de onda. Isto é muito
importante do ponto de vista ecológico, pois permite um melhor aproveitamento da
energia disponível e o crescimento em ambientes com recursos luminosos diferentes.
Por exemplo, as ficobilinas presentes nas algas vermelhas absorvem luz azul que é a
côr predominante nas águas profundas onde estas algas crescem.
Figura 7: Interpretação tridimensional da disposição das membranas internas dum cloroplasto,
realçando a relação entre os tilacóides do estroma e os dos grana. É importante notar o canal ou lúmen em ambos os tipos de tilacóides. Retirado de Salisbury e Ross (1992), figura10.3, página 209

A Fotossíntese
12
Nas plantas superiores a clorofila b é um pigmento auxiliar que passa energia
à clorofila a. Em todos os organismos que libertam O2 a excitação move-se para
centros de reacção, que são constituídos por clorofila a, responsável por passar a
energia na forma de um electrão para reacções químicas.
2.2.1. AS CLOROFILAS:
As clorofilas (figura 8) são, provavelmente, os pigmentos biológicos mais
abundantes e, se o mundo aparece à vista humana muito rico em tons de verde, isto
deve-se ao facto das clorofilas absorverem no azul e no vermelho. As folhas podem
conter mais de 1g de clorofila por m2 de superfície, mas este valor varia com as
espécies, com a idade e com a nutrição (particularmente com a fertilização em
azoto).
A molécula de clorofila é uma molécula lipofílica que se encontra apenas nas
membranas. Quimicamente as clorofilas são constituídas por quatro grupos pirrólicos
ligados por pontes CH (grupos meténicos) formando um anel porfirínico plano, o
qual contém no centro um átomo de magnésio. A maioria das clorofilas contém uma
molécula de fitol (C20H39OH) ligada a um dos anéis pirrólicos, ficando aquela
molécula num plano perpendicular ao plano do anel porfirínico (figura 8) (Teixeira e
Ricardo, 1983).
Para além da clorofila a e b existem na natureza outros tipos de clorofila,
quimicamente distintos. A clorofila c encontra-se juntamente com a clorofila a, nas
diatomáceas, nas algas castanhas e nos dinoflagelados. A clorofila d ocorre,
juntamente com a clorofila a, em certo número de algas vermelhas (Teixeira e
Ricardo, 1983).
2.2.2. OS CAROTENÓIDES:
Os carotenóides constituem um grupo de pigmentos que ocorrem tanto nos
vegetais como nos animais. Estes pigmentos estão presentes em praticamente todas
as plantas, embora em concentrações variáveis. São pigmentos geralmente

A Fotossíntese
13
Figura 8: Estrutura de moléculas de clorofila. Estas moléculas têm um anel pofirínico que contém um
ião de Mg2+
no centro dos quatro anéis pirrólicos. As clorofilas também contêm uma “cauda” de fitol que tornam a molécula hidrofóbica. As várias clorofilas diferem nos substituintes à volta da estrutura em anel. No caso das clorofila a existe um grupo metilo, enquanto que na clorofila b está presente na mesma posição um grupo formil. Retirado de Buchanan et al. (2000), figura 12.4, página 575
amarelados ou alaranjados podendo ter colorações acastanhadas ou avermelhadas.
Os carotenóides formam moléculas lineares. Alguns são constituídos exclusivamente
por carbono e hidrogénio pertencendo, portanto, aos hidrocarbonetos. Tais
carotenóides têm o nome de carotenos. Os carotenóides que possuem átomos de
oxigénio nas suas moléculas denominam-se xantófilas (figura 9) (Salisbury & Ross,
1992).

A Fotossíntese
14
Figura 9: Estruturas químicas de A - - caroteno, um pigmento amarelo avermelhado com a fórmula empírica C40 H56; B – luteína, uma xantófila amarela com a fórmula empírica C40 H56 O2; C – licopeno, um
caroteno avermelhado com a fórmula empírica C40 H56 , que não se encontra nos cloroplastos, mas que se encontra no tomate. Retirado de Salisbury & Ross (1992), figura10-4, página210
Os três carotenos mais comuns na natureza são o -caroteno, o -
caroteno; -caroteno. O -caroteno é o mais importante nas plantas e encontra-se
geralmente acompanhado do -caroteno em proporções variáveis.
As xantófilas são dos carotenóides mais abundantes na natureza podendo,
nas folhas em desenvolvimento, atingir concentrações duas vezes superiores às dos
carotenos. A luteína é a xantófila principal nas folhas verdes e nas algas verdes e
vermelhas (Teixeira & Ricardo, 1983).
2.2.3. AS FICOBILINAS:
São pigmentos que existem apenas nos organismos aquáticos, onde a sua
concentração pode igualar ou mesmo ultrapassar a da clorofila a. As ficobilinas
encontram-se sempre associadas a proteínas, formando complexos que se
denominam ficobiliproteínas.
As ficobilinas podem ser de dois tipos: as ficoeritrinas que são pigmentos
vermelhos, muito importantes nas algas vermelhas; e as ficocianinas que são
pigmentos azuis, mais importantes nas cianobactérias mas que também se
encontram nas algas vermelhas. Do ponto de vista químico, as ficobilinas têm

A Fotossíntese
15
semelhanças às clorofilas, mas não possuem cadeia lateral de fitol, não contêm
magnésio e o grupo tetrapirrólico não é cíclico (figura 10) (Teixeira & Ricardo, 1983).
Figura 10: Estrutura das ficobilinas.
Retirado Teixeira e Ricardo (1983), figura 7, página 27
2.3. A ABSORÇÃO DE FOTÕES:
A absorção da radiação visível depende do estado electrónico dos átomos e
moléculas da substância absorvente. As substâncias capazes de absorver radiação
visível têm o nome de pigmentos e a sua cor depende da radiação absorvida.
Assim, a clorofila tem cor verde porque absorve essencialmente no vermelho e no
azul.
Quando um fotão atinge um átomo ou uma molécula, a energia do fotão é
transmitida a determinados electrões, dizendo-se que o átomo ou molécula passou
de um estado basal ou de repouso a um estado de excitação. Note-se que um
átomo ou molécula só é excitável por fotões de determinado comprimento de onda. O
estado de excitação corresponde a uma deslocação dum electrão duma orbital para
outra. A energia necessária para passar um electrão duma orbital de menor energia
para outra de maior energia depende da diferença de energia entre as duas orbitais.
Dado que a energia dum fotão apenas pode ser utilizada na sua totalidade, a
fracção dum quantum não pode ser utilizada, só aqueles fotões com energia igual à
requerida para efectuar a deslocação de um electrão para orbitais mais externas
podem ser absorvidos. Assim, um determinado átomo absorverá selectivamente
radiação de comprimentos de onda correspondentes às energias de transição
electrónica características desse átomo. Ou seja, a absorção da luz só pode ocorrer
quando a energia h de um quantum for igual à diferença entre as energias
correspondentes aos estados energéticos inicial (E0) e final (E1) (Teixeira & Ricardo,
1983).

A Fotossíntese
16
hEE 01
Se o vector electrónico oscilante dum fotão faz com que um electrão numa
molécula entre em ressonância (isto é, vibre com a mesma frequência do fotão),
então a energia do fotão será absorvida.
As moléculas possuem outros estados de excitação energética além do
electrónico, os quais resultam de rotações e vibrações internas. Embora os diferentes
níveis rotacionais e vibracionais difiram pouco em conteúdo energético (quando
comparados com as diferenças de energia que se observam entre os estados de
repouso e excitação electrónica), eles também absorvem radiação luminosa. Note-se
que nas moléculas qualquer electrão está relacionado com mais de um núcleo
atómico, podendo ocupar várias orbitais em torno de dois ou mais núcleos. Esta
situação aumenta bastante o número de níveis de energia possíveis, o que diminui as
energias de transição entre eles. Disto resulta que as moléculas absorvam radiação
visível e infravermelha, enquanto que os átomos apenas absorvem radiação
ultravioleta.
Uma molécula contém numerosos electrões. Uns estão mais próximos dos
núcleos atómicos, outros estão mais afastados. Estes últimos (electrões de valência)
são os que normalmente participam nas reacções fotoquímicas. Existem vários tipos
de electrões de valência (figura 11) (Teixeira & Ricardo, 1983):
electrões n: não envolvidos em ligações químicas, isto é, fazem parte de
pares não compartilhados;
electrões : envolvidos nas ligações químicas do tipo (em que os
electrões estão simetricamente distribuídos em torno do eixo de ligação, isto é, têm
uma simetria cilíndrica relativamente ao eixo longitudinal que liga dois núcleos
atómicos);
electrões : envolvidos nas ligações do tipo (os electrões não se
encontram simetricamente distribuídos em torno do eixo que liga os núcleos
atómicos).
Em condições normais e à temperatura ambiente, a maioria dos electrões de
uma molécula encontra-se no estado de repouso. Quando uma molécula absorve
energia luminosa e passa a um estado excitado, um dos seus electrões passa da
orbital correspondente ao seu estado de repouso a uma orbital mais energética (mais
afastada do núcleo). Este electrão pode ser de qualquer uma das espécies acima
referidas. Assim, assinalando o estado de excitação por um asterisco, podem
verificar-se os seguintes tipos de transição electrónica: n *; *; *.
Porém, os electrões estão fortemente ligados, sendo preciso uma energia elevada

A Fotossíntese
17
para os excitar. A diferença de energia entre o nível basal e o primeiro estado de
excitação de praticamente todos os electrões é tão elevada que a transição *
só tem lugar com radiação ultravioleta de comprimento de onda curto (o chamado UV
longínquo). São, pois, as excitações dos electrões n e , menos firmemente ligados
que têm importância nas reacções fotoquímicas biológicas. De facto, as transições
electrónicas * e n * podem ocorrer por acção da luz visível (Teixeira &
Ricardo, 1983).
Figura 11: Distribuição das nuvens electrónicas em relação aos núcleos atómicos A e B das ligações
e .
Retirado de Teixeira e Ricardo (1983), figura 24, página 73
Na transição electrónica n *, um electrão de um par não compartilhado
passa, ao ser excitado, para uma orbital instável (dita antiligante), como acontece,
por exemplo, no seguinte caso:
\ \
C = Ö: C = O: / /
Na transição electrónica *, um electrão estável passa para uma orbital
instável (antiligante). Por exemplo:
\ \ :
C = Ö: C O: / /
Transições do tipo * podem ocorrer mesmo em moléculas relativamente
simples como o etileno, mas neste caso só o UV de menor comprimento de onda é
absorvido. Porém no caso de moléculas menos simples, que possuam duplas
ligações conjugadas a energia necessária para que se dê a transição é mais baixa e
a absorção terá lugar a comprimentos de onda mais longos. Se o número de ligações
duplas conjugadas for relativamente elevado, a absorção deslocar-se-á para a região
do visível e as moléculas absorventes aparecerão coradas. É o que acontece com os
pigmentos (Teixeira & Ricardo, 1983).
Cada electrão possui, portanto, uma certa energia relacionada com a
distância ao núcleo. Mas uma outra porção da energia do electrão provém do

A Fotossíntese
18
movimento de rotação em torno do seu eixo. Este movimento giratório é designado
por spin. O spin tem dois sentidos possíveis, da esquerda para a direita e da
direita para a esquerda . De acordo com o princípio de exclusão de Pauli, cada
orbital não pode ter mais de dois electrões e quando existem dois, eles deverão ter
spins opostos. Dois electrões de spins opostos (antiparalelos) dizem-se
emparelhados.
Quando um electrão é excitado, ele pode manter o seu spin ou invertê-lo.
Note-se que a inversão do spin não tem lugar logo após a absorção do fotão, mas
durante a permanência no estado excitado, isto é, o electrão imediatamente após a
excitação mantém o seu spin original (oposto ao do seu par não excitado), podendo
mudar de spin ao transitar para outro nível de energia. Os estados de excitação
correspondentes a spins antiparalelos e paralelos designam-se respectivamente por
singleto e tripleto (ver figura 12) (Teixeira & Ricardo, 1983).
Uma molécula é mais estável no seu estado de repouso. O estado de
excitação resultante da absorção de um fotão é instável, a molécula permanece
nesta situação durante períodos de tempo relativamente curtos. A energia de
excitação adquirida por uma molécula A pode ser dissipada de diferentes maneiras
(figura 13).
1. A energia de excitação é libertada sob a forma de calor através da
ocorrência de colisões com outras moléculas. A perda de energia por libertação de
calor processa-se muito rapidamente; a molécula permanece no estado de excitação
durante muito pouco tempo, retomando o seu estado de repouso (ou um estado
energético intermédio) dentro de 10-12 10-11 s (Teixeira & Ricardo, 1983).
2. A molécula excitada retoma o seu estado de repouso emitindo radiação. O
comprimento de onda da radiação emitida é sempre superior ao da radiação
absorvida, a diferença representa as perdas de energia vibratória. A emissão de
radiação mais corrente é a que resulta da passagem singleto de excitação ao estado
de repouso, processo que se denomina fluorescência. A emissão de energia a
partir do estado tripleto, que envolve a inversão de spin, chama-se fosforescência e
tem lugar a um comprimento de onda mais longo que o da fluorescência
característico do sistema.
Enquanto a fluorescência decorre dentro de períodos de tempo próximos de
10-9 s, a fosforescência tem uma duração que varia entre 10-3 s e 10 s, para a maioria

A Fotossíntese
19
das moléculas orgânicas. Esta longa permanência no estado tripleto de excitação é
devida à baixa probabilidade da ocorrência da inversão do spin (Teixeira & Ricardo,
1983).
3. A molécula excitada pode retomar o seu estado de repouso passando a
sua energia de excitação a uma outra molécula. Esta transferência de energia
processa-se sem a ocorrência de colisões. Para que este mecanismo se verifique de
maneira eficiente, as moléculas envolvidas têm de estar muito próximas umas das
outras (distâncias da ordem de 5 a 10 nm). Este mecanismo conhecido como
ressonância indutiva, não envolve transferência de electrões de molécula a
molécula, nem emissão de fotões intermediários, verifica-se apenas uma resposta da
molécula receptora ao campo eléctrico da molécula transmissora (Teixeira & Ricardo,
1983).
Figura 12: Diagrama representativo dos spins dos electrões nos estados singleto e tripleto.
Retirado de Teixeira e Ricardo (1983), figura 25, página 74
4. Neste processo, a molécula excitada perde a sua energia participando
numa reacção química. Se em muitas destas reacções a molécula é modificada,
existem casos em que a sua estrutura não é alterada. Na fotossíntese, por exemplo,
as moléculas excitadas dos pigmentos participam em reacções químicas sem
sofrerem alterações nas suas estruturas moleculares. Por outro lado, a energia
luminosa pode só fornecer a “energia de activação” necessária à reacção ou pode,
como acontece na fotossíntese, ser também parcialmente conservada na energia
química dos produtos.
A figura 13 ajuda a perceber porque razão a luz azul é sempre menos
eficiente na fotossíntese que a luz vermelha. Após excitação com um fotão azul, o
electrão decai sempre muito rapidamente, por perda de calor, para um nível

A Fotossíntese
20
energético mais baixo correspondente ao do fotão vermelho. Deste nível mais baixo
pode ocorrer perda adicional de calor, fluorescência ou fotossíntese (Teixeira &
Ricardo, 1983).
Entre estes quatro processos será favorecido e predominará o que tiver a taxa
mais rápida. Para muitos pigmentos do aparelho fotossintético a fluorescência ocorre
em nanosegundos (10-9 s), enquanto que o processo fotoquímico ocorre em pico
segundos (10-12 s). Assim, quando a via fotoquímica está disponível, observar-se-á
pouca fluorescência e a fotossíntese ocorrerá com muita eficiência. O estado de
singleto da clorofila participa na fotoquímica da transferência de energia. A duração
do estado tripleto da clorofila leva a pensar que não é um estado intermediário dos
acontecimentos que levam à separação de uma carga na fotossíntese (Buchanan et
al, 2000).
Figura 13: Esquema da absorção de fotões (h) por um átomo e a excitação de um electrão. O calor (H)
pode também causar a deslocação dum electrão para uma orbital de maior nível energético. Os processos principais de dissipação de energia são: R transição sem emissão de radiação; F fluorescência; P fosforescência; por reacções químicas na fotossíntese; transferência, por exemplo da
energia do estado tripleto para oxigénio ou carotenóides; ou ainda da energia de excitação para clorofila ou outros pigmentos. Retirado de Lawlor (1987), figura 2.1, página 21
A eficiência do processo fotoquímico pode ser determinado pelo rendimento
quântico que se pode expressar pela seguinte equação:
absorvidos quanta de nº
amentefotoquimic formados produtos de nº
De acordo com esta equação um rendimento quântico igual a 1 indica que
todos os fotões absorvidos são convertidos num produto. Este tanto pode ser CO2
fixo, como O2 libertado, como ainda o início dum processo fotoquímico. Valores
baixos indicam que outras formas de dissipação da energia diminuem a eficiência da
reacção fotoquímica (Buchanan et al., 2000)
A Fotossíntese
21
2.4. ESPECTROS DE ABSORÇÃO E DE ACÇÃO:
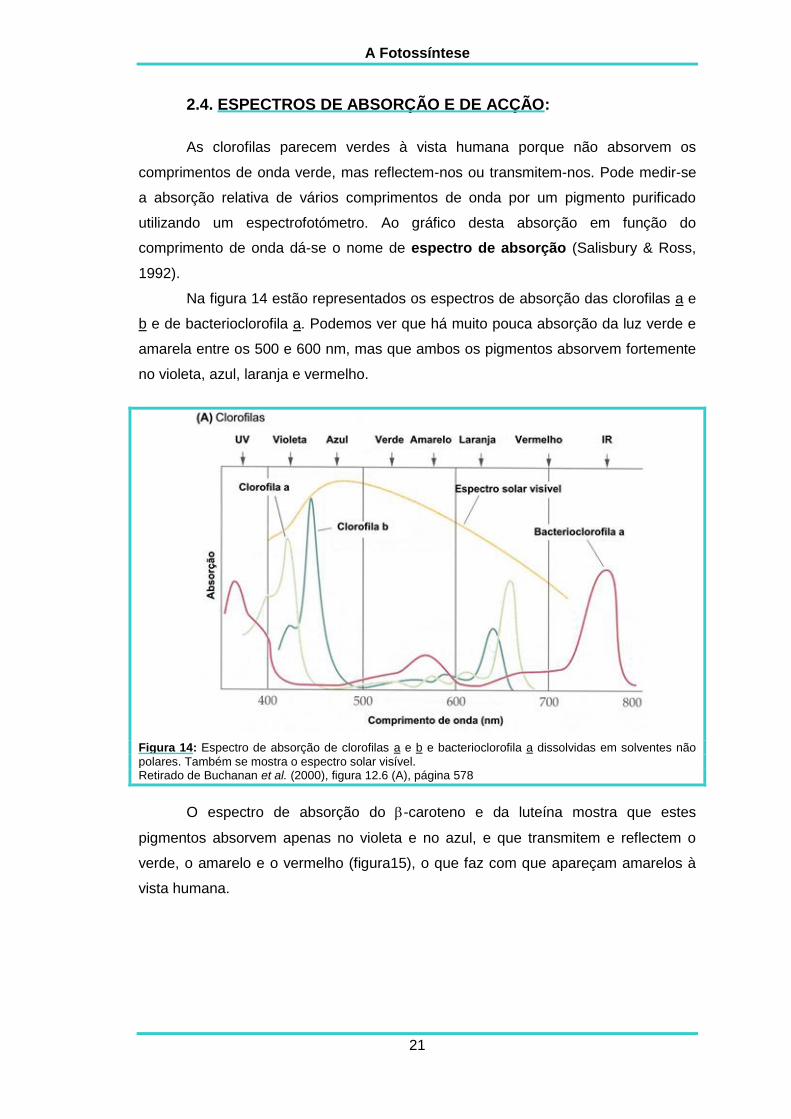
As clorofilas parecem verdes à vista humana porque não absorvem os
comprimentos de onda verde, mas reflectem-nos ou transmitem-nos. Pode medir-se
a absorção relativa de vários comprimentos de onda por um pigmento purificado
utilizando um espectrofotómetro. Ao gráfico desta absorção em função do
comprimento de onda dá-se o nome de espectro de absorção (Salisbury & Ross,
1992).
Na figura 14 estão representados os espectros de absorção das clorofilas a e
b e de bacterioclorofila a. Podemos ver que há muito pouca absorção da luz verde e
amarela entre os 500 e 600 nm, mas que ambos os pigmentos absorvem fortemente
no violeta, azul, laranja e vermelho.
Figura 14: Espectro de absorção de clorofilas a e b e bacterioclorofila a dissolvidas em solventes não
polares. Também se mostra o espectro solar visível. Retirado de Buchanan et al. (2000), figura 12.6 (A), página 578
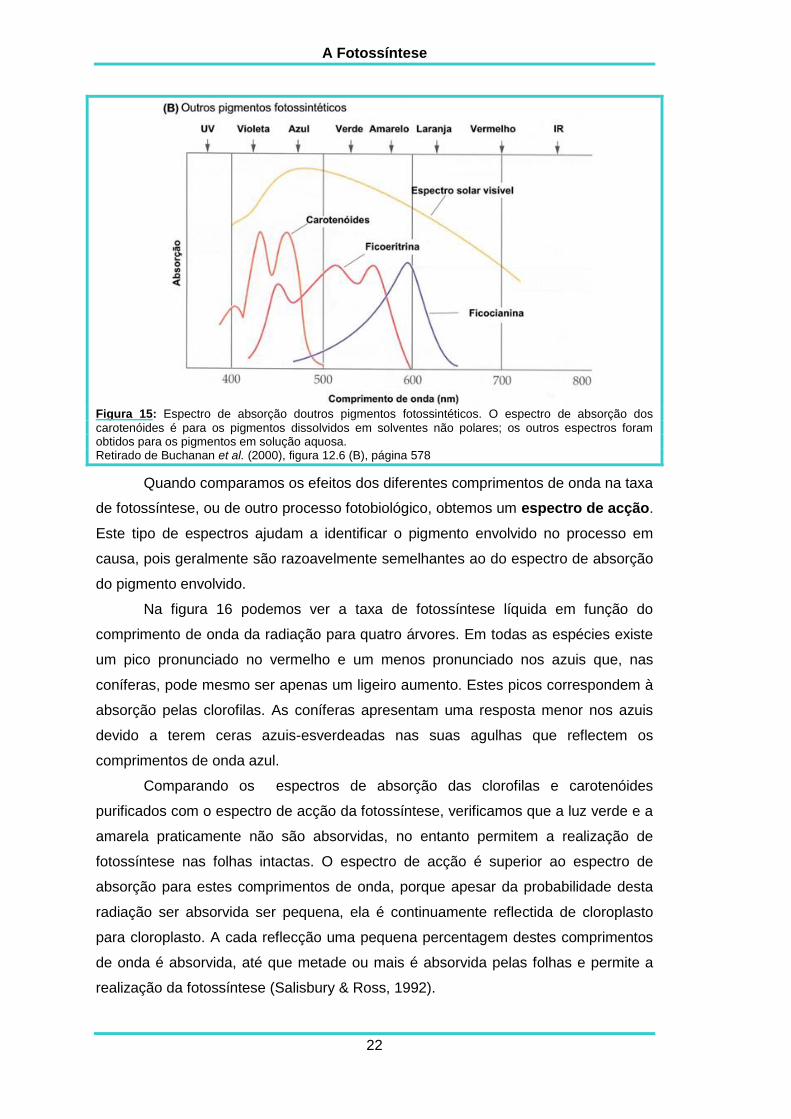
O espectro de absorção do -caroteno e da luteína mostra que estes
pigmentos absorvem apenas no violeta e no azul, e que transmitem e reflectem o
verde, o amarelo e o vermelho (figura15), o que faz com que apareçam amarelos à
vista humana.
A Fotossíntese
22
Figura 15: Espectro de absorção doutros pigmentos fotossintéticos. O espectro de absorção dos
carotenóides é para os pigmentos dissolvidos em solventes não polares; os outros espectros foram obtidos para os pigmentos em solução aquosa. Retirado de Buchanan et al. (2000), figura 12.6 (B), página 578
Quando comparamos os efeitos dos diferentes comprimentos de onda na taxa
de fotossíntese, ou de outro processo fotobiológico, obtemos um espectro de acção.
Este tipo de espectros ajudam a identificar o pigmento envolvido no processo em
causa, pois geralmente são razoavelmente semelhantes ao do espectro de absorção
do pigmento envolvido.
Na figura 16 podemos ver a taxa de fotossíntese líquida em função do
comprimento de onda da radiação para quatro árvores. Em todas as espécies existe
um pico pronunciado no vermelho e um menos pronunciado nos azuis que, nas
coníferas, pode mesmo ser apenas um ligeiro aumento. Estes picos correspondem à
absorção pelas clorofilas. As coníferas apresentam uma resposta menor nos azuis
devido a terem ceras azuis-esverdeadas nas suas agulhas que reflectem os
comprimentos de onda azul.
Comparando os espectros de absorção das clorofilas e carotenóides
purificados com o espectro de acção da fotossíntese, verificamos que a luz verde e a
amarela praticamente não são absorvidas, no entanto permitem a realização de
fotossíntese nas folhas intactas. O espectro de acção é superior ao espectro de
absorção para estes comprimentos de onda, porque apesar da probabilidade desta
radiação ser absorvida ser pequena, ela é continuamente reflectida de cloroplasto
para cloroplasto. A cada reflecção uma pequena percentagem destes comprimentos
de onda é absorvida, até que metade ou mais é absorvida pelas folhas e permite a
realização da fotossíntese (Salisbury & Ross, 1992).
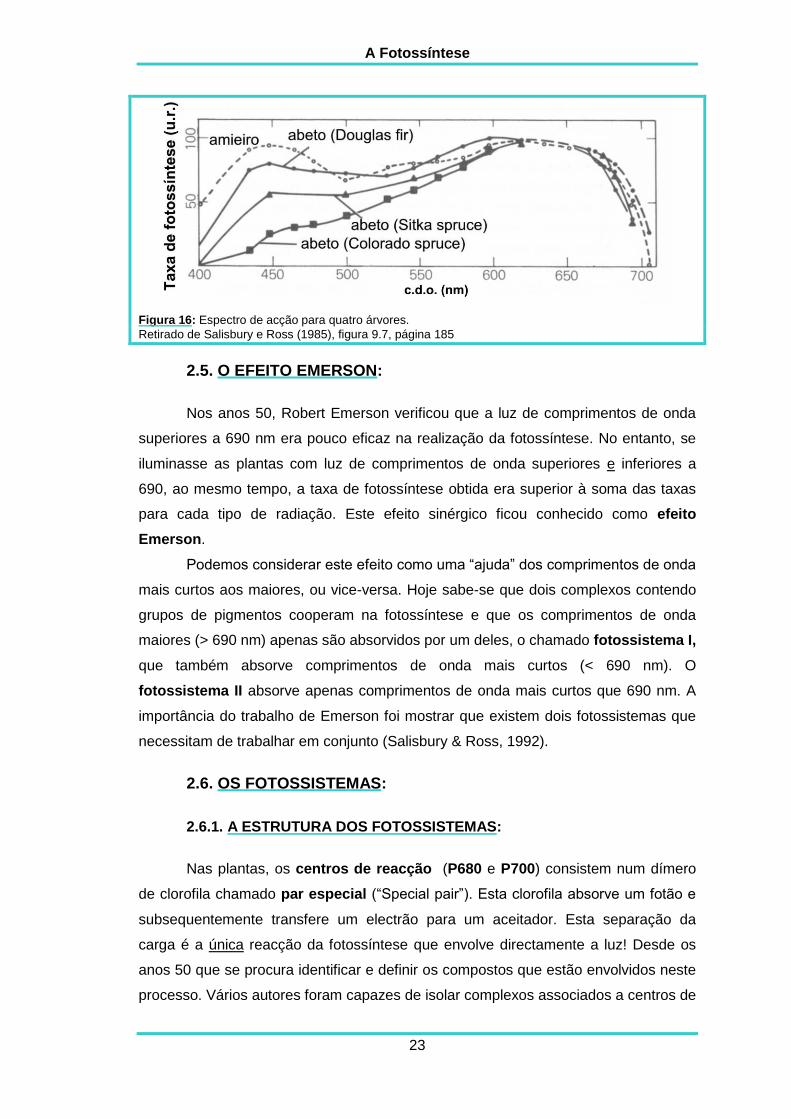
A Fotossíntese
23
Figura 16: Espectro de acção para quatro árvores.
Retirado de Salisbury e Ross (1985), figura 9.7, página 185
2.5. O EFEITO EMERSON:
Nos anos 50, Robert Emerson verificou que a luz de comprimentos de onda
superiores a 690 nm era pouco eficaz na realização da fotossíntese. No entanto, se
iluminasse as plantas com luz de comprimentos de onda superiores e inferiores a
690, ao mesmo tempo, a taxa de fotossíntese obtida era superior à soma das taxas
para cada tipo de radiação. Este efeito sinérgico ficou conhecido como efeito
Emerson.
Podemos considerar este efeito como uma “ajuda” dos comprimentos de onda
mais curtos aos maiores, ou vice-versa. Hoje sabe-se que dois complexos contendo
grupos de pigmentos cooperam na fotossíntese e que os comprimentos de onda
maiores (> 690 nm) apenas são absorvidos por um deles, o chamado fotossistema I,
que também absorve comprimentos de onda mais curtos (< 690 nm). O
fotossistema II absorve apenas comprimentos de onda mais curtos que 690 nm. A
importância do trabalho de Emerson foi mostrar que existem dois fotossistemas que
necessitam de trabalhar em conjunto (Salisbury & Ross, 1992).
2.6. OS FOTOSSISTEMAS:
2.6.1. A ESTRUTURA DOS FOTOSSISTEMAS:
Nas plantas, os centros de reacção (P680 e P700) consistem num dímero
de clorofila chamado par especial (“Special pair”). Esta clorofila absorve um fotão e
subsequentemente transfere um electrão para um aceitador. Esta separação da
carga é a única reacção da fotossíntese que envolve directamente a luz! Desde os
anos 50 que se procura identificar e definir os compostos que estão envolvidos neste
processo. Vários autores foram capazes de isolar complexos associados a centros de

A Fotossíntese
24
reacção de um grande número de organismos fotossintéticos. A caracterização
desses complexos permitiu estabelecer a sua natureza proteica. (Buchanan et al.,
2000).
Para além dos dímeros de clorofila, os complexos dos centros de reacção
contêm vários aceitadores de electrões. Em todos os complexos dos centros de
reacção ocorre uma transferência de um electrão do “par especial” para uma
molécula de outro de pigmento, ou mesmo outra molécula de clorofila. A esta
reacção seguem-se outras transferências de electrões para moléculas que não são
pigmentos, como as quinonas ou os centros Fe-S. Esquematicamente teremos:
ClorA0A1A2 h Clor*A0A1A2
h Clor+A0-A1A2
h Clor+A0A1-A2
h
Clor+A0A1- A2
em que Clor representa a molécula de clorofila e A0, A1 e A2 representam vários
aceitadores de electrões (Buchanan et al., 2000).
Propostas actuais de modelos do fotossistema II (PSII) mostram os
transportadores de electrões organizados numa estrutura em X, contendo duas
moléculas de feofitina, um átomo de ferro e duas moléculas de plastoquininona
designadas por QA e QB que actuam como aceitadores terminais dos electrões (figura
17). Uma proposta de modelo para o fotossistema I (PSI) pode ser analisada na
figura 18. Embora a resolução actual do complexo do PSI ainda seja insuficiente para
identificar todos os transportadores e definir as suas interacções com as subunidades
proteicas, o modelo apresentado já dá uma ideia da organização dos componentes
deste fotossistema (Buchanan et al., 2000).
Para além do centro de reacção e dos seus componentes de transferência de
electrões, todos os fotossistemas têm um conjunto de pigmentos que absorvem a
radiação e que constituem a antena. Dados de vários autores parecem indicar que
em cada fotossistema existem cerca de 250 moléculas de clorofila associadas com o
respectivo centro de reacção. A transferência de energia de uma molécula de
pigmento para outra, por ressonância (ou transferência de Föster) não requer, como
vimos anteriormente, emissão ou reabsorção de fotões (figura 19). A proximidade das
moléculas dadoras e do aceitadoras é fundamental, uma vez que a eficiência da
transferência energética é inversamente proporcional à sexta potência da distância
entre as duas moléculas.
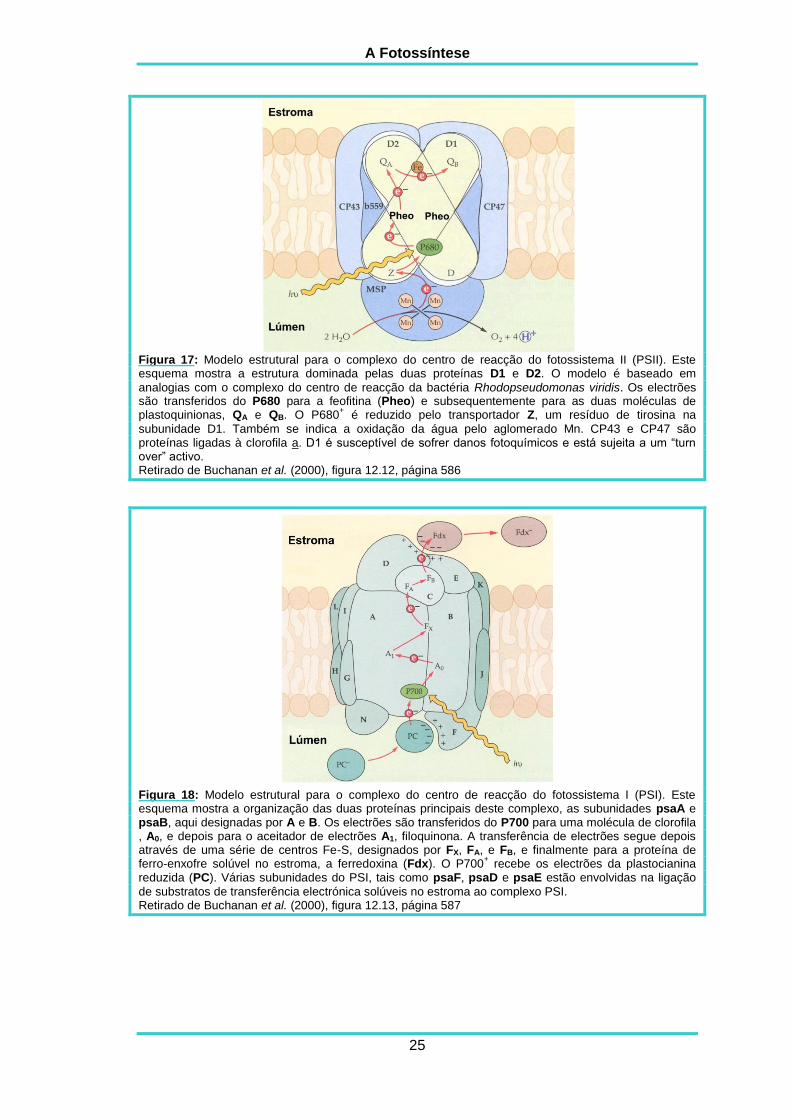
A Fotossíntese
25
Figura 17: Modelo estrutural para o complexo do centro de reacção do fotossistema II (PSII). Este esquema mostra a estrutura dominada pelas duas proteínas D1 e D2. O modelo é baseado em
analogias com o complexo do centro de reacção da bactéria Rhodopseudomonas viridis. Os electrões são transferidos do P680 para a feofitina (Pheo) e subsequentemente para as duas moléculas de plastoquinionas, QA e QB. O P680
+ é reduzido pelo transportador Z, um resíduo de tirosina na
subunidade D1. Também se indica a oxidação da água pelo aglomerado Mn. CP43 e CP47 são proteínas ligadas à clorofila a. D1 é susceptível de sofrer danos fotoquímicos e está sujeita a um “turn over” activo. Retirado de Buchanan et al. (2000), figura 12.12, página 586
Figura 18: Modelo estrutural para o complexo do centro de reacção do fotossistema I (PSI). Este esquema mostra a organização das duas proteínas principais deste complexo, as subunidades psaA e psaB, aqui designadas por A e B. Os electrões são transferidos do P700 para uma molécula de clorofila , A0, e depois para o aceitador de electrões A1, filoquinona. A transferência de electrões segue depois através de uma série de centros Fe-S, designados por FX, FA, e FB, e finalmente para a proteína de ferro-enxofre solúvel no estroma, a ferredoxina (Fdx). O P700
+ recebe os electrões da plastocianina
reduzida (PC). Várias subunidades do PSI, tais como psaF, psaD e psaE estão envolvidas na ligação
de substratos de transferência electrónica solúveis no estroma ao complexo PSI. Retirado de Buchanan et al. (2000), figura 12.13, página 587

A Fotossíntese
26
Figura 19: Esquema representativo do conceito básico de transferência energética que ocorre na
fotossíntese. Um conjunto de pigmentos funciona como antena, absorvendo a luz e transferindo a energia para o centro de reacção, onde reacções químicas armazenam alguma desta energia transferindo electrões de clorofilas para uma molécula de aceitador. Uma molécula dadora fornece um electrão que reduz de novo a clorofila. a transferência de excitação na antena é apenas é um fenómeno puramente físico e não envolve trocas químicas. Retirado de Taiz & Zeiger (1998), figura 7.7, página 163
A proteína principal de ligação de pigmentos nas membranas dos cloroplastos
constitui o “complexo de colheita de luz II” (light harvesting complex II, LHC–II).
Pensa.se que esta proteína representa cerca de metade da proteína total na
membrana do tilacóide. Por microscopia electrónica pode observar-se a estrutura do
LCH-II, em que as três hélices transmembranares desta proteína ligam cerca de 12
moléculas de proteína a e b. Para além disso duas moléculas de carotenóides
servem de armação para as hélices A e B (figura 20-A). As proteínas LHC estão
frequentemente organizadas em estruturas triméricas (figura 20-B) (Buchanan et al.,
2000). A figura 21 apresenta um esquema da organização das clorofilas nos
respectivos fotossistemas.
2.6.2. A ORGANIZAÇÃO DOS COMPLEXOS NOS TILACÓIDES:
Como vimos anteriormente os tilacóides apresentam duas zonas, a apressa
em que um tilacóide dum granum contacta com outro, e zona não apressa a que
está em contacto directo com o estroma. Uma característica importante da
organização dos tilacóides é que os fotossistemas PSI e PSII não estão distribuídos
ao acaso no sistema membranar (figura 22). O PSI está essencialmente nas zonas
não apressas e portanto em contacto com o estroma, enquanto o PSII se encontra
essencialmente nas zonas apressas (Buchanan et al., 2000).
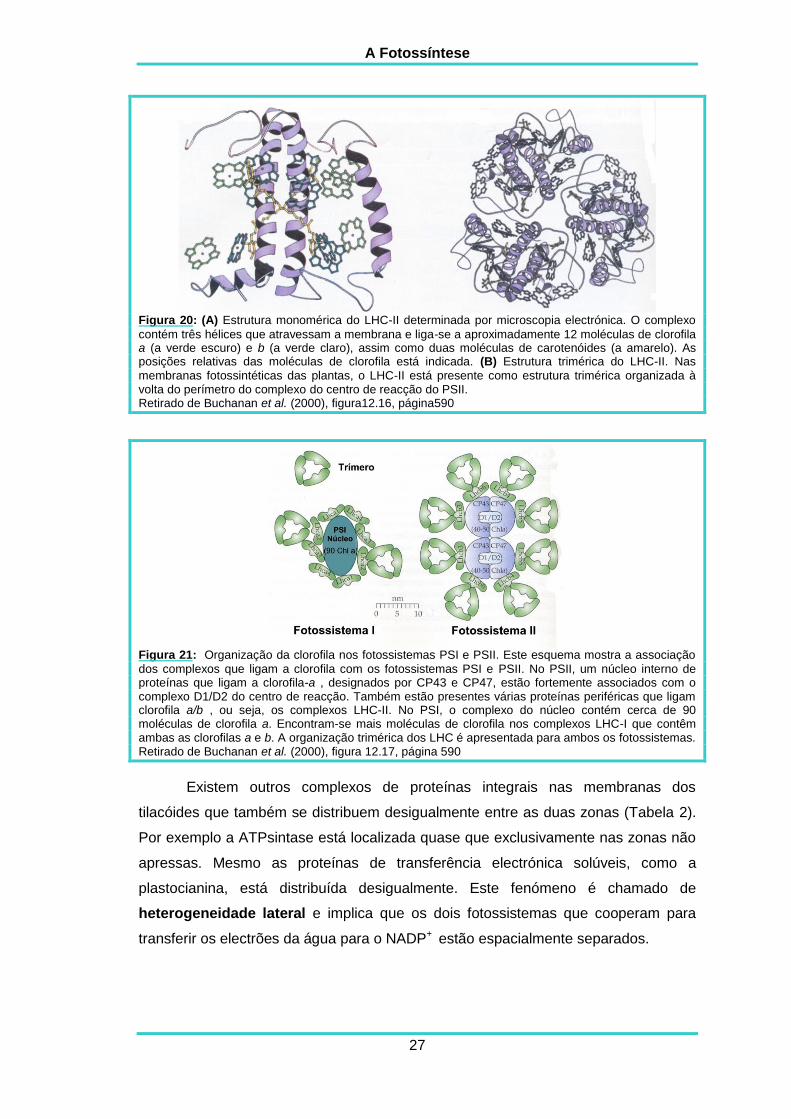
A Fotossíntese
27
Figura 20: (A) Estrutura monomérica do LHC-II determinada por microscopia electrónica. O complexo
contém três hélices que atravessam a membrana e liga-se a aproximadamente 12 moléculas de clorofila a (a verde escuro) e b (a verde claro), assim como duas moléculas de carotenóides (a amarelo). As posições relativas das moléculas de clorofila está indicada. (B) Estrutura trimérica do LHC-II. Nas
membranas fotossintéticas das plantas, o LHC-II está presente como estrutura trimérica organizada à volta do perímetro do complexo do centro de reacção do PSII. Retirado de Buchanan et al. (2000), figura12.16, página590
Figura 21: Organização da clorofila nos fotossistemas PSI e PSII. Este esquema mostra a associação
dos complexos que ligam a clorofila com os fotossistemas PSI e PSII. No PSII, um núcleo interno de proteínas que ligam a clorofila-a , designados por CP43 e CP47, estão fortemente associados com o complexo D1/D2 do centro de reacção. Também estão presentes várias proteínas periféricas que ligam clorofila a/b , ou seja, os complexos LHC-II. No PSI, o complexo do núcleo contém cerca de 90 moléculas de clorofila a. Encontram-se mais moléculas de clorofila nos complexos LHC-I que contêm ambas as clorofilas a e b. A organização trimérica dos LHC é apresentada para ambos os fotossistemas. Retirado de Buchanan et al. (2000), figura 12.17, página 590
Existem outros complexos de proteínas integrais nas membranas dos
tilacóides que também se distribuem desigualmente entre as duas zonas (Tabela 2).
Por exemplo a ATPsintase está localizada quase que exclusivamente nas zonas não
apressas. Mesmo as proteínas de transferência electrónica solúveis, como a
plastocianina, está distribuída desigualmente. Este fenómeno é chamado de
heterogeneidade lateral e implica que os dois fotossistemas que cooperam para
transferir os electrões da água para o NADP+ estão espacialmente separados.

A Fotossíntese
28
Tabela 2: Distribuição dos vários complexos nas duas zonas das membranas dos tilacóides. Retirado de Buchanan et al.(2000), tabela 12.4, página 592
Tilacóides (%)
Componente Zona Apressa Zona não apressa
PSII 85 15
PSI 10 90
Complexo do citocrómio b6f 50 50
LHC-II 90 10
ATP sintase 0 100
Plastocianinaa 40 60
aA percentagem indica a distribuição relativa de cada componente nas zonas apressa ou não
apressa das membranas, com excepção da plastocianina que se refere ao lúmen da respectiva zona da membrana.
De acordo com este modelo têm de existir mecanismos para transferência
electrónica a longa distância. É de notar que nem todas as proteínas membranares
estão distribuídas desigualmente nos tilacóides. Assim, o complexo do citocrómio b6f
que transfere electrões entre os dois fotossistemas está distribuído com bastante
uniformidade. (Buchanan et al, 2000)
2.6.3. A DISTRIBUIÇÂO DA ENERGIA ENTRE OS FOTOSSISTEMAS:
O complexo proteico do LHC-II que, como vimos, funciona como antena para
o fotossistema II encontra-se quase exclusivamente nas zonas apressas. É possível
que a função do empilhamento das membranas nos cloroplastos, isto é, a existência
de grana, esteja relacionada com uma distribuição eficiente da energia entre os
complexos PSI e o PSII. É fundamental que um fotossistema não receba mais fotões
que o outro, uma vez que para haver um máximo de eficiência no transporte
electrónico é necessário que ambos os fotossistemas sofram uma excitação
equilibrada. A separação espacial dos dois fotossistemas pode ajudar nesta
regulação. Além disso, os cloroplastos são capazes de ajustar a associação do LHC-
II ao PSII para regular a distribuição de quanta entre os fotossistemas (Buchanan et
al., 2000).
Uma pequena porção do total do LHC-II sofre uma fosforilação reversível à
luz. Esta fosforilação muda a carga à superfície da proteína. A molécula de LHC-II
depois da fosforilação fica carregada negativamente e é deslocada do núcleo
hidrofóbico das zonas apressas para a zona não apressa que é menos hidrofóbica.
Esta migração duma porção do complexo antena do PSII diminui a absorção da luz
pelo PSII nas zonas apressas ao diminuir a antena associada a este fotossistema
(figura 23).

A Fotossíntese
29
Figura 22: Heterogeneidade lateral dos complexos das membranas dos cloroplastos. O PSII está
essencialmente localizado nas zonas apressas, enquanto que o PSI e a ATPsintase estão quase exclusivamente situados nas zonas não apressas. O complexo do citocrómio b6f está distribuído uniformemente em ambas as zonas. A separação dos fotossistemas implica transportadores electrónicos móveis, tais como as plastocianinas, que transferem electrões entre os complexos da membrana separados no espaço. Retirado de Buchanan et al. (2000), figura12.19, página 591
É evidente que os cloroplastos sentem um desequilíbrio entre a excitação dos
fotossistemas e respondem fosforilando o LHC-II. O modelo mais aceite para explicar
isto envolve uma cinase activada por uma reacção redox (“redox-activated kinase”).
Esta cinase é activada quando o conjunto das plastoquinonas, que são
transportadores electrónicos que conduzem os electrões entre os dois fotossistemas
(como veremos mais adiante), se encontram altamente reduzidas. Isto acontece
quando o PSII recebe mais luz que o PSI e causa uma activação da LHC-II cinase.
Esta cinase fosforila o LHC-II que causa a migração do LHC-II para fora da zona
apressa, diminuindo a absorção de luz pelo PSII. As plastoquinonas vão, de seguida,
ser oxidadas pela activação do PSI e a cinase fica menos activa. Então uma
fosfatase desfosforila o LHC-II que migra de novo para as zonas apressas
aumentando a quantidade de luz absorvida pelo PSII. Este sistema de “feedback”
permite um controlo muito subtil da distribuição da luz entre os dois fotossistemas
apesar de eles se encontrarem em zonas diferentes dos tilacóides (Buchanan et al.,
2000).
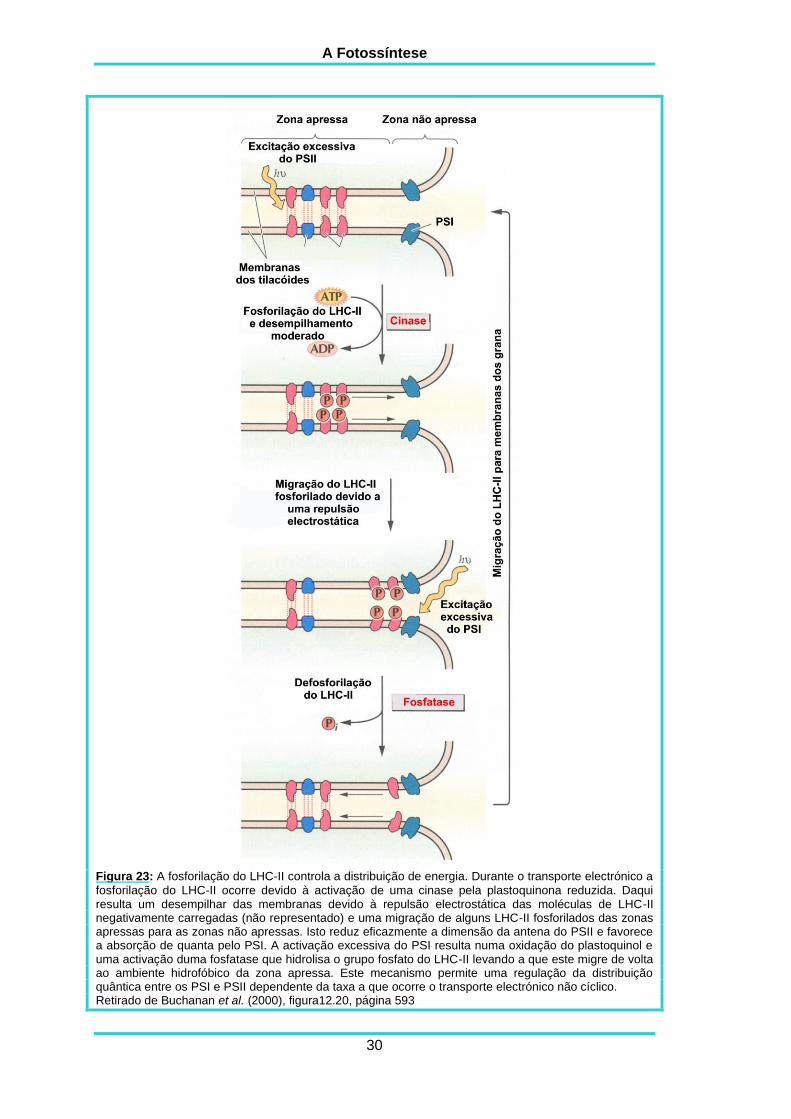
A Fotossíntese
30
Figura 23: A fosforilação do LHC-II controla a distribuição de energia. Durante o transporte electrónico a
fosforilação do LHC-II ocorre devido à activação de uma cinase pela plastoquinona reduzida. Daqui resulta um desempilhar das membranas devido à repulsão electrostática das moléculas de LHC-II negativamente carregadas (não representado) e uma migração de alguns LHC-II fosforilados das zonas apressas para as zonas não apressas. Isto reduz eficazmente a dimensão da antena do PSII e favorece a absorção de quanta pelo PSI. A activação excessiva do PSI resulta numa oxidação do plastoquinol e uma activação duma fosfatase que hidrolisa o grupo fosfato do LHC-II levando a que este migre de volta ao ambiente hidrofóbico da zona apressa. Este mecanismo permite uma regulação da distribuição quântica entre os PSI e PSII dependente da taxa a que ocorre o transporte electrónico não cíclico. Retirado de Buchanan et al. (2000), figura12.20, página 593

A Fotossíntese
31
2.7. O TRANSPORTE ELECTRÓNICO:
2.7.1. O ESQUEMA EM Z:
O esquema em Z resulta de estudos levados a cabo desde os anos 40 até
aos anos 60, e ainda é o modelo preferido pela maioria dos autores para o transporte
electrónico não cíclico (figura 24).
Figura 24: O esquema em Z mostrando os valores de Em dos transportadores electrónicos. A colocação
vertical de cada transportador da cadeia de transporte electrónico não cíclico corresponde ao ponto médio do seu potencial redox. Retirado de Buchanan et al. (2000), figura 12.22 (A), página 595
À luz a primeira separação de carga que ocorre no PSII produz um oxidante
muito forte, o P680+ e um redutor razoavelmente estável que é uma
plastosemiquinona (QA-). No PSI a separação da carga produz um redutor forte e
estável (um centro Fe-S, Fx-) e um oxidante fraco (P700+). O P680+ produzido pelo
PSII tem poder oxidante suficiente para remover electrões da água, enquanto que o
QA- fornece o poder redutor que vai dar electrões ao P700+ através de uma série
decrescente de transferências electrónicas que envolvem outros transportadores
electrónicos, incluindo o complexo proteico transmembranar do citocrómio b6f
(Buchanan et al., 2000).
A figura 25 representa um esquema em Z que ilustra de forma mais detalhada
o movimento dos electrões e protões durante o transporte electrónico acíclico ao
longo dos vários complexos dos tilacóides.

A Fotossíntese
32
Figura 25: A organização na membrana do esquema em Z. Os componentes da cadeia de transporte
electrónico e o complexo sintetizador de ATP encontram-se representados nas membranas dos tilacóides. Indicam-se quatro complexos membranares: o PSI, o PSII, o complexo do citocrómio b6f, e a ATP sintase (CF1 – CF0). Os electrões são transferidos da água para o NADP
+, ao mesmo tempo que se
stabelece um gradiente protónico através da membrana. Este gradiente electroquímico é utilizado para a formação de ATP pela ATPsintase. A transferência de electrões é representada por linhas vermelhas e a translocação de protões por linhas azuis. Fd x = ferredoxina; FNR = ferredoxina-NADP
+ redutase.
Retirado de Buchanan et al. (2000), figura 12.22, página 595
2.7.2. A ACÇÃO DO FOTOSSISTEMA II (PSII):
Pode dizer-se que à luz, o PSII funciona como uma oxiredutase água-
plastoquinona transferindo electrões da água para as plastoquinonas (figura 26).
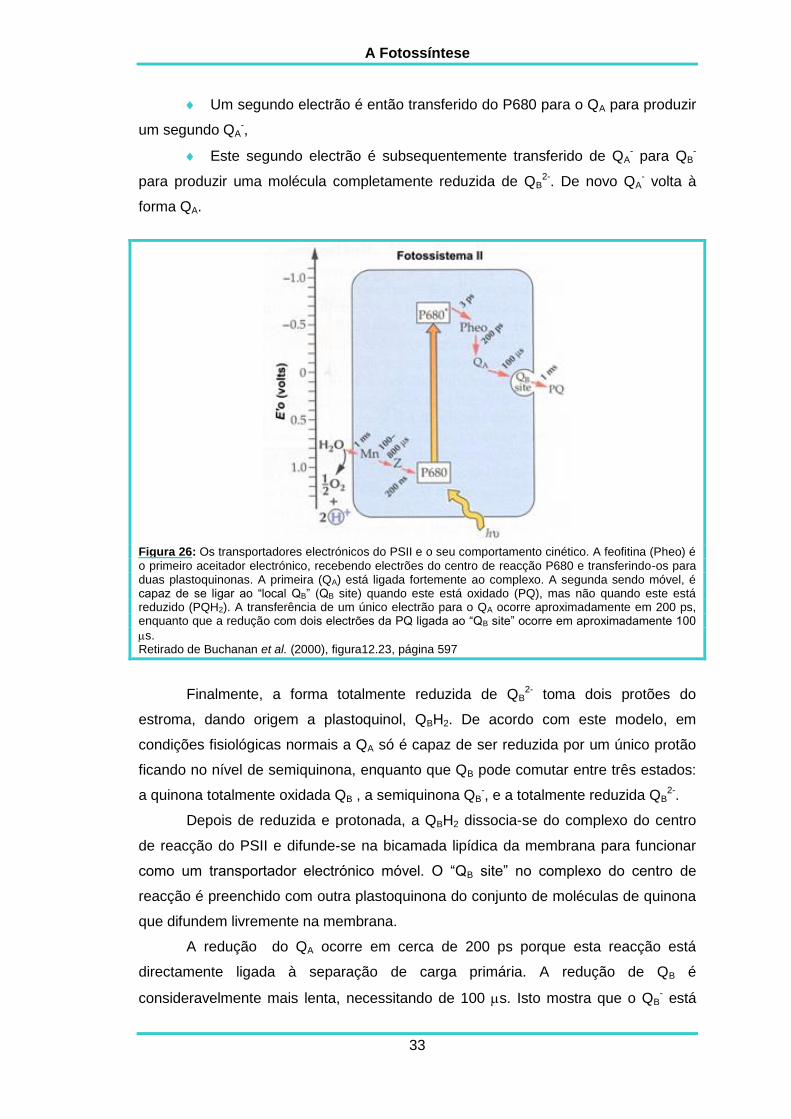
2.7.2.1. A redução das quinonas:
O complexo do centro de reacção do PSII tem ligadas a si duas quinonas: a
QA e a QB. A QA está fortemente ligada e funciona como o primeiro aceitador de
electrões estável. A QB está ligada de forma mais fraca e funciona como um
aceitador de electrões secundário. A redução das duas quinonas decorre num
processo que envolve 4 etapas:
O primeiro electrão é libertado do P680 e é transferido para a QA para
produzir a plastosemiquinona QA-,
Este electrão é então transferido para QB para originar uma semiquinona
QB-. a perda do electrão reverte QA
- a QA,
A Fotossíntese
33
Um segundo electrão é então transferido do P680 para o QA para produzir
um segundo QA-,
Este segundo electrão é subsequentemente transferido de QA- para QB
-
para produzir uma molécula completamente reduzida de QB2-. De novo QA
- volta à
forma QA.
Figura 26: Os transportadores electrónicos do PSII e o seu comportamento cinético. A feofitina (Pheo) é
o primeiro aceitador electrónico, recebendo electrões do centro de reacção P680 e transferindo-os para duas plastoquinonas. A primeira (QA) está ligada fortemente ao complexo. A segunda sendo móvel, é capaz de se ligar ao “local QB” (QB site) quando este está oxidado (PQ), mas não quando este está reduzido (PQH2). A transferência de um único electrão para o QA ocorre aproximadamente em 200 ps, enquanto que a redução com dois electrões da PQ ligada ao “QB site” ocorre em aproximadamente 100
s. Retirado de Buchanan et al. (2000), figura12.23, página 597
Finalmente, a forma totalmente reduzida de QB2- toma dois protões do
estroma, dando origem a plastoquinol, QBH2. De acordo com este modelo, em
condições fisiológicas normais a QA só é capaz de ser reduzida por um único protão
ficando no nível de semiquinona, enquanto que QB pode comutar entre três estados:
a quinona totalmente oxidada QB , a semiquinona QB-, e a totalmente reduzida QB
2-.
Depois de reduzida e protonada, a QBH2 dissocia-se do complexo do centro
de reacção do PSII e difunde-se na bicamada lipídica da membrana para funcionar
como um transportador electrónico móvel. O “QB site” no complexo do centro de
reacção é preenchido com outra plastoquinona do conjunto de moléculas de quinona
que difundem livremente na membrana.
A redução do QA ocorre em cerca de 200 ps porque esta reacção está
directamente ligada à separação de carga primária. A redução de QB é
consideravelmente mais lenta, necessitando de 100 s. Isto mostra que o QB- está

A Fotossíntese
34
firmemente ligado ao complexo do centro de reacção enquanto não recebe um
segundo electrão. pelo contrário, a ligação do QBH2 é relativamente fraca, e este
quinol é facilmente deslocado do seu local de ligação por uma quinona totalmente
oxidada (Buchanan et al., 2000).
2.7.2.2. A oxidação da água:
A oxidação da água não é um processo directo, mas envolve uma série
complexa de reacções no lado do lúmen do tilacóide. A oxidação da água envolve a
transferência de quatro electrões:
2H2O O2 + 4H+ + 4e-
O modelo mais aceite para as reacções de oxidação da água, o “S-state
model”, é descrito na figura 27. Este modelo postula um mecanismo de acumulação
de cargas conduzido pela luz através do qual o complexo libertador de oxigénio do
PSII progride ao longo de cinco estádios sucessivos de oxidação crescente, do S0 ao
S4, sendo S4 um oxidante muito forte capaz de oxidar a água. Cada separação de
carga no centro de reacção P680 rende um P680+ que oxida o acumulador,
avançando-o para o estado-S seguinte e aumentando a carga deste em +1. O único
estado em que há libertação de O2 é o S4 (Buchanan et al., 2000).
Figura 27: O complexo onde ocorre libertação de O2 apresenta 5 estados de oxidação (de S0 a S4). O
ciclo avança sequencialmente cada vez que um fotão é absorvido pelo PSII até se atingir o estado S4 altamente oxidado (carregado positivamente). O S4 é o único estado capaz de produzir a oxidação da água. Retirado de Buchanan et al. (2000), figura 12.30, página 603

A Fotossíntese
35
Pensa-se que o acumulador de cargas do modelo “S-state” é constituído por
átomos de manganês que sofrem oxidações sucessivas originando um complexo
oxidante no estado S4 que é capaz de oxidar a água.
Para além do manganês já foram identificados mais dois cofactores: iões
cloro e iões cálcio, cuja função ainda não foi elucidada.
A ligação entre o P680+ e o complexo S-state envolvendo manganês não é
directa. Existe um transportador electrónico intermediário, chamado Z, e que é um
resíduo de tirosina na subunidade D1 do complexo do centro de reacção do PSII
(Buchanan et al., 2000).
2.7.3. A ACÇÃO DO COMPLEXO DO CITOCROMO b6f:
O complexo citocromo b6f funciona como uma oxiredutase plastoquinol-
plastocianina, transferindo electrões do plastoquinol para a plastocianina que é uma
proteína contendo cobre que se encontra no lúmen do tilacóide. Esta transferência de
electrões é acompanhada pela translocação de protões através da membrana, desde
o estroma até o lúmen. O mecanismo de acção deste complexo, com características
for a do comum, permite a passagem de dois protões por cada electrão transferido
para a plastocianina (Buchanan et al., 2000). A estrutura e as reacções da
plastoquinona podem ser observadas na figura 28.
Figura 28: Estrutura e reacções das plastoquinonas. (A) A plastoquinona consiste numa “cabeça”
quinoide e uma longa “cauda” não-polar que segura a molécula na membrana. (B) Descrição das reacções redox da plastoquinona. Estão aqui representadas a forma totalmente oxidada - quinona (Q); a
forma aniónica - semiquinona (Q-), e a forma reduzida - quinol (QH2); R representa a cadeia lateral.
Retirado de Taiz & Zeiger (1998), figura7.26, página 178
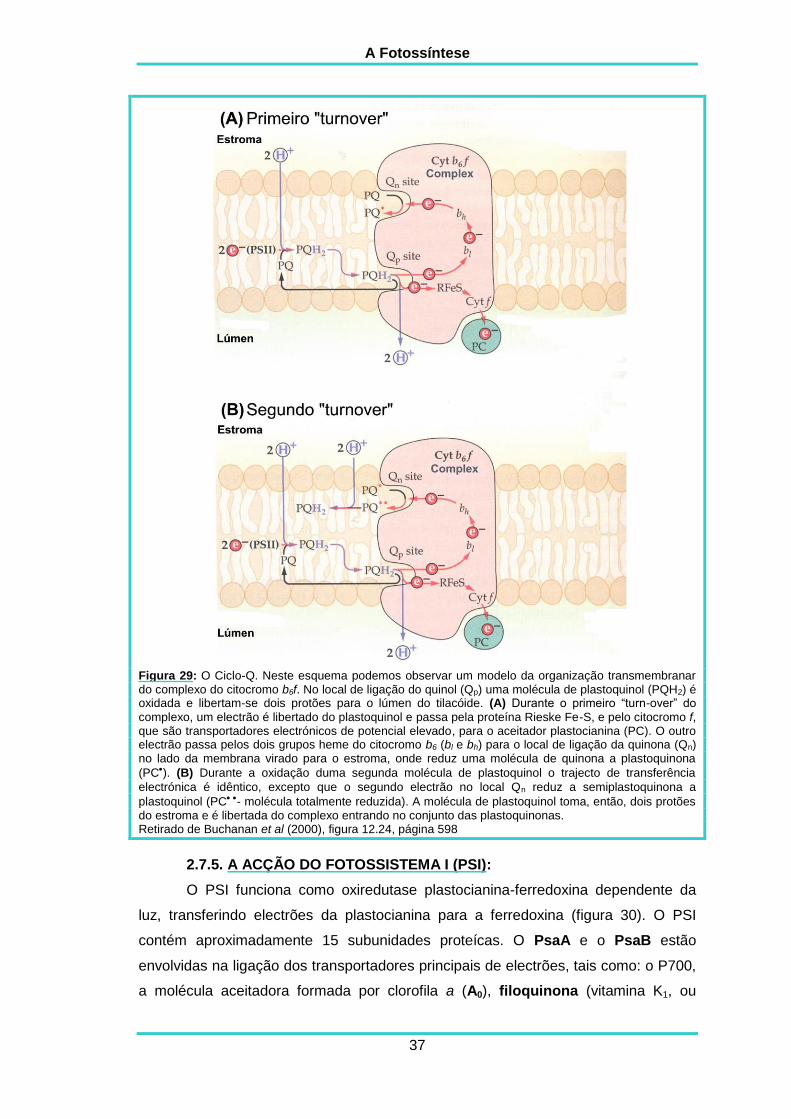
O mecanismo mais aceite para descrever as reacções do complexo do
citocrómo é o Ciclo-Q (figura 29). De acordo com este modelo o complexo do

A Fotossíntese
36
citocrómo contém um local de ligação a quinol (Qp) e outro de ligação a quinona (Qn)
em lados opostos da membrana do tilacóide. A oxidação do quinol ocorre em dois
passos descontínuos no local Qp, situado no lado do lúmen. No Qp o quinol é oxidado
para semiquinona por um centro Rieske Fe-S e o electrão libertado passa do centro
Fe-S para o citocrómo f e depois para a plastocianina. A plastosemiquinona é então
oxidada por uma das duas hemes, a bl (“b-low” baixo potencial) localizado no
citocromo b6 perto do lado do lúmen. A acompanhar esta oxidação, os protões são
libertados do quinol para o lúmen. O electrão na heme bl é transferido através da
membrana para a segunda heme, a bh (“b-high alto potencial). Este electrão é então
transferido para a molécula de quinona que está ligada no local (Qn) no lado do
estroma da membrana, produzindo uma semiquinona. Este ciclo repete-se outra vez
para oxidar o segundo plastoquinol, com um electão a passar para a plastocianina e
o segundo a ser transferido para o Qn para produzir uma molécula de quinol
totalmente reduzida (PQH2). O quinol totalmente reduzido toma dois protões do
estroma e dissocia-se do local Qn. O resultado líquido deste ciclo é de uma
molécula de plastoquinol oxidada a quinona (PQ; no local Qp), dois electrões são
transferidos para a plastocianina (PC)
e quatro protões são transferidos do estroma para o lúmen do cloroplasto (Buchanan
et al.,2000), ou seja:
2PQH2 + 2PCox + 2H+ PQH2 + PQ + 2PCred + 4 H+
As reacções do complexo do citocromo b6f estão entre as mais importantes,
em termos de limitação da taxa a que ocorre o fluxo de electrões.
2.7.4. A ACÇÃO DA PLASTOCIANINA:
Como vimos o complexo citocromo b6f reduz o transportador de electrões
móvel chamado plastocianina que é uma proteína de baixo peso molecular,
contendo cobre. A transferência de electrões envolvendo a plastocianina é muito
rápida: a sua redução pelo complexo do citocromo b6f ocorre em cerca de 100 a 200
s e a sua oxidação pelo PSI requer cerca de 10 s. Isto levou alguns autores a
pensar que os três componentes deverão estar organizados numa espécie de
“supercomplexo”, o que eliminaria a necessidade para um movimento de longa
distância da plastocianina.
A Fotossíntese
37
Figura 29: O Ciclo-Q. Neste esquema podemos observar um modelo da organização transmembranar do complexo do citocromo b6f. No local de ligação do quinol (Qp) uma molécula de plastoquinol (PQH2) é oxidada e libertam-se dois protões para o lúmen do tilacóide. (A) Durante o primeiro “turn-over” do
complexo, um electrão é libertado do plastoquinol e passa pela proteína Rieske Fe-S, e pelo citocromo f, que são transportadores electrónicos de potencial elevado, para o aceitador plastocianina (PC). O outro electrão passa pelos dois grupos heme do citocromo b6 (bl e bh) para o local de ligação da quinona (Qn) no lado da membrana virado para o estroma, onde reduz uma molécula de quinona a plastoquinona
(PC). (B) Durante a oxidação duma segunda molécula de plastoquinol o trajecto de transferência
electrónica é idêntico, excepto que o segundo electrão no local Qn reduz a semiplastoquinona a
plastoquinol (PC - molécula totalmente reduzida). A molécula de plastoquinol toma, então, dois protões do estroma e é libertada do complexo entrando no conjunto das plastoquinonas. Retirado de Buchanan et al (2000), figura 12.24, página 598
2.7.5. A ACÇÃO DO FOTOSSISTEMA I (PSI):
O PSI funciona como oxiredutase plastocianina-ferredoxina dependente da
luz, transferindo electrões da plastocianina para a ferredoxina (figura 30). O PSI
contém aproximadamente 15 subunidades proteícas. O PsaA e o PsaB estão
envolvidas na ligação dos transportadores principais de electrões, tais como: o P700,
a molécula aceitadora formada por clorofila a (A0), filoquinona (vitamina K1, ou

A Fotossíntese
38
aceitador A1), e o centro Fe-S (Fx). A subunidade de baixo peso molecular PsaC liga
os centros FA e FB que são centros Fe-S. Tal como acontece no PSII, existem no PSI
um grande número de subunidades proteicas sem grupos prostéticos e cujas
funções, em muitos casos, são desconhecidas. As subunidades PsaD e PsaF têm
sido associadas à ligação ao complexo da ferredoxina e da plastocianina,
respectivamente. (Buchanan et al., 2000).
A via da transferência electrónica no PSI é esquematizada na figura 30. O
dador de electrões para o PSI é a plastocianina e o aceitador final dos electrões é a
proteína solúvel chamada ferredoxina. Esta proteína Fe-S (2Fe-2S) está localizada
no estroma e é um agente redutor forte com um potencial redox de -420 mV, capaz
de reduzir o NADP+ numa reacção termodinamicamente favorável na gama
fisiológica de pH. Este transportador não transfere os electrões directamente para o
NADP+, mas através da acção de uma enzima intermediária chamada ferredoxina –
NADP+ reductase (FNR) (Buchanan et al., 2000).
Figura 30: Os transportadores electrónicos do PSI.O trajecto da transferência electrónica através do complexo do PSI é aqui representada envolvendo o P700, um monómero de clorofila a, (A0), uma filoquinona (A1) e uma série de transportadores electrónicos adicionais que incluem três centros Fe-S diferentes (FX, FA e FB). A transferência alectrónica através destes transportadores ocorre na gama dos pico a nanosegundos, com o aceitador de electrões terminal (Fdx) a ser reduzido em aproximadamente
2 s. Buchanan et al. (2000), figura12.28, página 601
Existem dados que indicam que a ferredoxina e a FNR formam um complexo
graças a interacções electrostáticas entre si. A FNR é uma enzima contendo FAD,
fracamente associada à membrana do tilacóide de onde é facilmente dissociada e
que pode ser reduzida em duas etapas. Na primeira um electrão reduz a FNR ao
estado flavina-semiquinona, na segunda etapa outro electrão leva-a ao estado

A Fotossíntese
39
totalmente reduzido FADH2. A FNR, então, transfere os dois electrões para o NADP+
(Buchanan et al., 2000).
2.7.6. O TRANSPORTE CÍCLICO DE ELECTRÕES:
Como se descreveu anteriormente a cadeia de transporte acíclico de
electrões liga a oxidação da água (libertação do O2) com a redução do NADP+ e a
produção de ATP. No entanto, os cloroplastos também levam a cabo um processo de
transporte electrónico que só envolve o PSI e produz apenas ATP (Buchanan et al.,
2000).
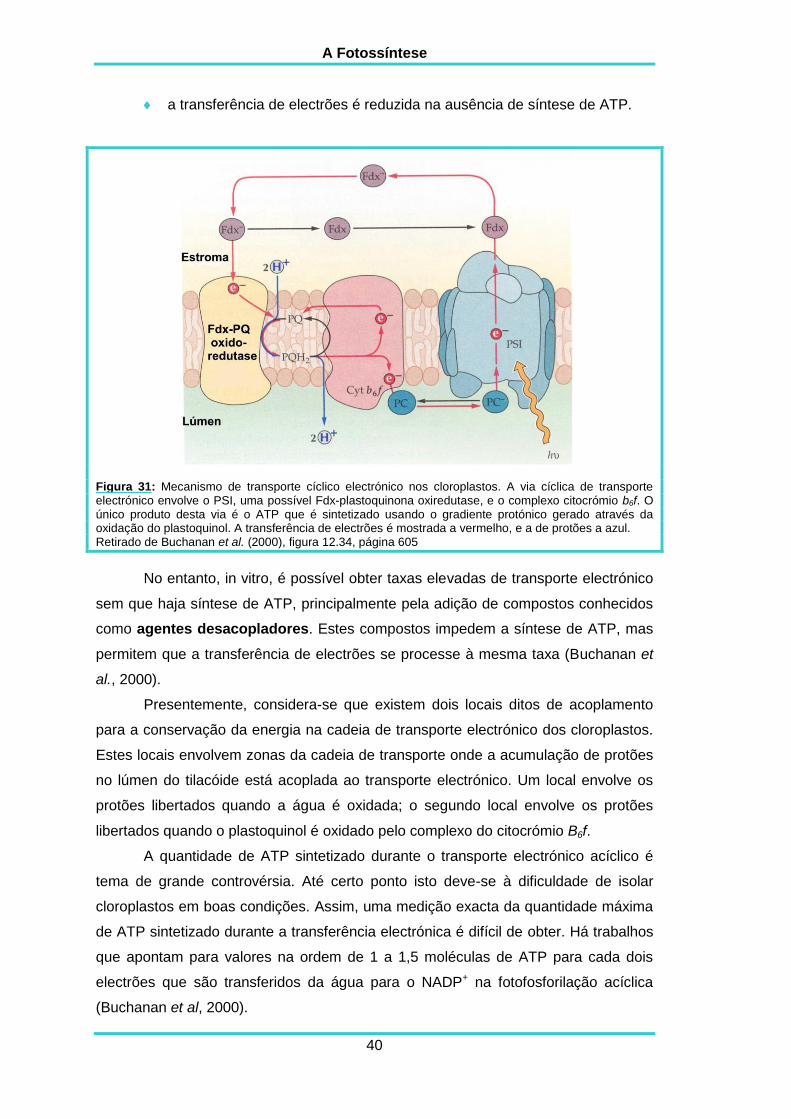
No modelo mais comum para descrever este processo o PSI reduz a
ferredoxina à luz, mas esta, em vez de passar os electrões para NADP+, interactua
com uma Fdx-plastoquinona oxidoredutase que permite a transferência dos electrões
para as quinonas. Moléculas de plastoquinol podem, então, ser oxidadas no
complexo do citocromo b6f permitindo a translocação de protões através da
membrana possivelmente através do Ciclo-Q (figura 31).
Ainda que se possa demonstrar in vitro o transporte cíclico de electrões e a
concomitante síntese de ATP, a função in vivo desta via ainda suscita muita
controvérsia (Buchanan et al, 2000). Experiências em que se mediu a fixação do CO2
nos cloroplastos intactos irradiados com luz de c.d.o. vermelho e vermelho-longo e
em que se avaliou a resposta deste sistema a inibidores deram indicações que o
transporte electrónico cíclico permite formar ATP utilizado para a fixação de CO2. No
entanto, dados bioquímicos sobre este ciclo são ainda escassos, particularmente
ainda não se caracterizou a enzima chave de todo o processo, a Fdx- plastoquinona
oxiredutase (Buchanan et al., 2000).
2.8. A SÍNTESE DE ATP:
A síntese de ATP à luz é chamada fotofosforilação e foi estudada nos anos
50 por Arnon e colaboradores. Os cloroplastos podem sintetizar ATP graças ao
transporte de electrões acíclico com a consequente libertação de O2 e formação de
NADPH, ou graças ao transporte de electrões cíclico no qual só se forma ATP.
2.8.1. O ACOPLAMENTO DO TRANSPORTE ELECTRÓNICO E DA SÍNTESE DE ATP in vivo:
In vivo, na fotofosforilação, assim como na fosforilação oxidativa que ocorre
na respiração, a síntese de ATP está energeticamente acoplada ao transporte
electrónico. Isto quer dizer que:
não pode ocorrer fotofosforilação sem que ocorra transporte electrónico;
A Fotossíntese
40
a transferência de electrões é reduzida na ausência de síntese de ATP.
Figura 31: Mecanismo de transporte cíclico electrónico nos cloroplastos. A via cíclica de transporte electrónico envolve o PSI, uma possível Fdx-plastoquinona oxiredutase, e o complexo citocrómio b6f. O único produto desta via é o ATP que é sintetizado usando o gradiente protónico gerado através da oxidação do plastoquinol. A transferência de electrões é mostrada a vermelho, e a de protões a azul. Retirado de Buchanan et al. (2000), figura 12.34, página 605
No entanto, in vitro, é possível obter taxas elevadas de transporte electrónico
sem que haja síntese de ATP, principalmente pela adição de compostos conhecidos
como agentes desacopladores. Estes compostos impedem a síntese de ATP, mas
permitem que a transferência de electrões se processe à mesma taxa (Buchanan et
al., 2000).
Presentemente, considera-se que existem dois locais ditos de acoplamento
para a conservação da energia na cadeia de transporte electrónico dos cloroplastos.
Estes locais envolvem zonas da cadeia de transporte onde a acumulação de protões
no lúmen do tilacóide está acoplada ao transporte electrónico. Um local envolve os
protões libertados quando a água é oxidada; o segundo local envolve os protões
libertados quando o plastoquinol é oxidado pelo complexo do citocrómio B6f.
A quantidade de ATP sintetizado durante o transporte electrónico acíclico é
tema de grande controvérsia. Até certo ponto isto deve-se à dificuldade de isolar
cloroplastos em boas condições. Assim, uma medição exacta da quantidade máxima
de ATP sintetizado durante a transferência electrónica é difícil de obter. Há trabalhos
que apontam para valores na ordem de 1 a 1,5 moléculas de ATP para cada dois
electrões que são transferidos da água para o NADP+ na fotofosforilação acíclica
(Buchanan et al, 2000).

A Fotossíntese
41
2.8.2. O MECANISMO QUIMIOSMÓTICO DA SÍNTESE DE ATP:
2.8.2.1. A força motriz protónica (pmf):
Peter Mitchell propôs, nos anos 60, o modelo quimiosmótico para explicar a
síntese de ATP quer nos cloroplastos, quer nas mitocôndrias. De acordo com este
modelo, a força motriz energética para a síntese de ATP é um gradiente iónico
através duma membrana selectivamente permeável. Nos cloroplastos e nas
mitocôndrias, este gradiente é um gradiente protónico formado em consequência da
transferência electrónica. Este gradiente estabelece-se devido a uma diferença na
concentração de protões e pode também resultar numa diferença de potencial
eléctrico através da membrana. Estas fontes de energia potencial podem ser usadas
para a fosforilação da ADP pela ATP sintase, que é uma enzima que liga este fluxo
de protões energeticamente favorável à síntese de ATP.
Várias características importantes do modelo quimiosmótico foram verificadas
experimentalmente:
este mecanismo requer um sistema membranar intacto com uma
permeabilidade aos protões intrinsecamente baixa;
os componentes de transporte electrónico devem estar dispostos
vectorialmente na membrana de forma a permitir que os protões a atravessem em
locais determinados e no sentido do lúmen;
à medida que os electrões passam ao longo da série de transportadores,
os protões são translocados através da membrana nos locais de acoplamento.
Nos cloroplastos, na via de transporte electrónico acíclica, os protões são
“depositados” no lúmen no complexo libertador de oxigénio do PSII (complexo de
oxidação da água) e no local Qp (local de oxidação do plastiquinol) perto da face
virada para o lúmen do complexo do citocromo b6f. Esta acumulação unidireccional
de protões resulta numa concentração muito elevada de protões no lado da
membrana virado para o lúmen e uma concentração menor no lado do estroma. O
gradiente que assim se forma fornece a força motriz protónica, pmf – “proton
motive force” (Buchanan et al., 2000).
No entanto, ainda existe uma certa controvérsia sobre se os protões são
móveis (deslocalizados) no interior da totalidade do canal do tilacóide, ou se estão
localizados (compartimentados) em certas regiões próximas da superfície interior da
membrana do tilacóide (Sebanek, 1992).
Estes possíveis locais de “compartimentação” deverão ter uma grande
capacidade de tamponização fornecida, em parte por resíduos de lisina com uma

A Fotossíntese
42
constante de ionização ácido-base anormalmente baixa1 (pKa 7,5) e, também por
grupos carboxilo. Estes grupos com grande capacidade de tamponização pertencem
a proteínas associadas às membranas, incluindo as do complexo de oxidação da
água (Ewy & Dilley, 2000)
Para alguns autores o gradiente de H+ localizado pode comutar para um
equilíbrio livre com o lúmen, e os protões que são assim dirigidos para o lúmen
parecem ser capazes de permitir a formação de ATP com uma eficiência idêntica à
do gradiente protónico localizado. Esta comutação reversível entre um modo de
acoplamento localizado e deslocalizado parece estar sob controlo de uma proteína
que liga Ca2+. O fluxo de H+ dependente do Ca2+ (“calcium gating”) tem uma função
fisiológica muito importante ao regular o pH do lúmen do tilacóide que por sua vez
regula a actividade de uma enzima – a violaxantina de-epoxidase – que desempenha
uma função chave na resposta foto-protectora dos cloroplastos a uma intensidade
luminosa excessiva (Pan & Dilley, 2000)
Independentemente da questão da compartimentalização dos protões a força
motriz protónica tem dois componentes que estão ligados entre si:
um componente pH que resulta da diferença de concentração em protões
através da membrana (pH);
um componente de potencial eléctrico que resulta de uma diferença de
cargas através da membrana ().
-1-1
HH mol VkJ 5.96/pmf e pH
No caso dos cloroplastos o contribuinte principal para o pmf é essencialmente
o gradiente de pH (pH), mas nas mitocôndrias é o potencial eléctrico (Buchanan
et al., 2000).
2.8.2.2. A ATPsintase:
Para além do gradiente protónico é ainda necessário a presença na
membrana da ATPsintase que é capaz de utilizar a força motriz protónica para
converter ADP em ATP. Este complexo enzimático foi estudado inicialmente pela sua
capacidade em hidrolisar ATP (ATPase). No entanto, à luz, a força motriz protónica
(pmf) que se estabelece através da membrana causa a activação da ATPsintase e
inibe a actividade de ATPase que seria contraproducente. A ATP sintase dos
tilacóides apresenta dois segmentos (Buchanan et al.,2000):
um grupo hidrofílico (“cabeça”) no lado do estroma da membrana (CF1)
1 O pKa da cadeia lateral da lisina é 10.0
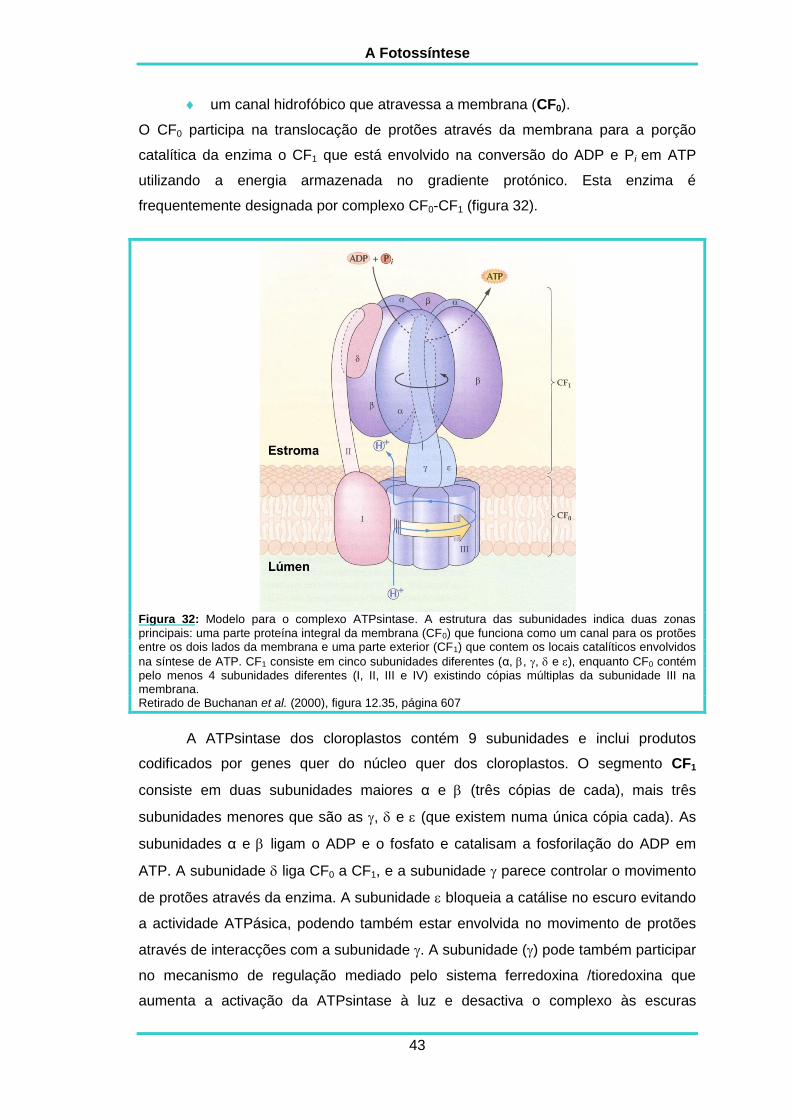
A Fotossíntese
43
um canal hidrofóbico que atravessa a membrana (CF0).
O CF0 participa na translocação de protões através da membrana para a porção
catalítica da enzima o CF1 que está envolvido na conversão do ADP e Pi em ATP
utilizando a energia armazenada no gradiente protónico. Esta enzima é
frequentemente designada por complexo CF0-CF1 (figura 32).
Figura 32: Modelo para o complexo ATPsintase. A estrutura das subunidades indica duas zonas
principais: uma parte proteína integral da membrana (CF0) que funciona como um canal para os protões entre os dois lados da membrana e uma parte exterior (CF1) que contem os locais catalíticos envolvidos
na síntese de ATP. CF1 consiste em cinco subunidades diferentes (α, , , e ), enquanto CF0 contém pelo menos 4 subunidades diferentes (I, II, III e IV) existindo cópias múltiplas da subunidade III na membrana. Retirado de Buchanan et al. (2000), figura 12.35, página 607
A ATPsintase dos cloroplastos contém 9 subunidades e inclui produtos
codificados por genes quer do núcleo quer dos cloroplastos. O segmento CF1
consiste em duas subunidades maiores α e (três cópias de cada), mais três
subunidades menores que são as , e (que existem numa única cópia cada). As
subunidades α e ligam o ADP e o fosfato e catalisam a fosforilação do ADP em
ATP. A subunidade liga CF0 a CF1, e a subunidade parece controlar o movimento
de protões através da enzima. A subunidade bloqueia a catálise no escuro evitando
a actividade ATPásica, podendo também estar envolvida no movimento de protões
através de interacções com a subunidade . A subunidade () pode também participar
no mecanismo de regulação mediado pelo sistema ferredoxina /tioredoxina que
aumenta a activação da ATPsintase à luz e desactiva o complexo às escuras

A Fotossíntese
44
bloqueando uma possível hidrólise de ATP que seria contraproducente (Buchanan et
al, 2000).
A composição do segmento CF1 está já bem definida, mas a composição do
segmento CF0 é menos conhecida, ainda que se possa pensar conter quatro
subunidades (I, II, III e IV). Provavelmente estas subunidades estão presentes em
cópias únicas com excepção da subunidade III que existe em doze cópias por
complexo. Pensa-se que as subunidades I, II e IV estão envolvidas na ligação dos
segmentos CF0 e CF1 enquanto a subunidade III do CF0 forma uma via para a
translocação de protões desde o lúmen para o estroma (Buchanan et al, 2000).
Pensa-se que deverão passar quatro H+ do lúmen para o estroma por cada ATP
sintetizado (Groth & Strotmann, 1999).
2.8.2.3. O mecanismo de síntese do ATP:
O mecanismo mais aceite para a síntese do ATP é chamado de “binding
change mechanism” ou literalmente “mecanismo de mudança de ligação” foi
proposto por Boyer e colegas em 1993. A característica principal deste mecanismo é
o conjunto de mudanças conformacionais do complexo proteico. De acordo com este
mecanismo a energia armazenada no gradiente protónico não é usada directamente
para levar a cabo a síntese do ATP mas antes para libertar uma forma de ATP
fortemente ligada ao sítio catalítico da enzima (Buchanan et al., 2000).
O segmento CF1 da enzima tem três locais de ligação distintos, cada um
deles pode existir em três estados conformacionais diferentes (figura 33):
ligação fraca ao nucleótido (L – loose);
ligação forte ao nucleótido (T – tight);
local de ligação aberto (O – open).
Os três estados conformacionais estão sempre presentes no complexo CF1,
cada um associado com um dos três centros catalíticos da enzima. De acordo com
este modelo o ADP e o Pi ligam-se a um local desocupado no estado O (open). À
medida que os protões se movem do lúmen para o estroma através do canal do CF0
liberta-se energia de que resulta uma rotação da subunidade do CF1. Esta rotação
causa alterações conformacionais nos três locais de ligação a nucleótidos. O local T
(tight) que tem ligado um ATP converte-se em local O (open) por libertação do ATP,
enquanto que o local L (loose) que contém ADP e Pi é convertido em local T. Isto
facilita a síntese de ATP, sem que seja necessária mais energia para esta conversão
(Buchanan et al, 2000).

A Fotossíntese
45
Figura 33: Esquema do “binding change mechanism” da síntese de ATP pelo complexo CF0 – CF1. Os três locais de ligação de nucleótidos estão representados na enzima: o local O (open), disponível para ligar ADP e Pi; o local L (loose), no qual o ADP e o Pi estão fracamente ligados; e o local T (tight) onde
o nucleótido fica fortemente ligado. Alterações conformacionais levadas a cabo pelo movimento de
protões através da membrana no passo (1) têm como resultado uma rotação na subunidade que causa interconversão destes locais e mudança de afinidade dos locais para os nucleótidos. A formação do ATP no local T é esquematizado no passo (2), mas esta condensação do ADP e Pi não requer mais
energia. Retirado de Buchanan et al. (2000), figura 12.36, página610
Embora este mecanismo seja o mais aceite é, no entanto, posto em causa por
alguns autores como Groth & Strotmann (1999) e McCarty et al.(2000) que
consideram que a rotação da subunidade observada in vitro pode não ocorrer in
vivo.
3. AS REACÇÕES INDIRECTAMENTE DEPENDENTES DA LUZ
- FASE ÀS ESCURAS – A REDUÇÃO FOTOSSINTÉTICA DO CARBONO
Vimos no ponto anterior que, os produtos das reacções fotossintéticas
directamente dependentes da luz são o ATP e o NADPH. Cerca de 95 % destes
compostos vão ser usados na redução do CO2 a hidratos de carbono, os restantes 5
% vão ser utilizados na redução fotossintética do nitrato e do sulfato. Como as
reacções conducentes à fixação do CO2 não dependem directamente da luz são
muitas vezes chamadas “fase às escuras da fotossíntese”. É, no entanto, importante
realçar que esta designação é errada, uma vez que sem a presença da luz não
poderia ocorrer redução do carbono. Assim, alguns autores preferem chamar a esta
etapa “as reacções fotossintéticas do carbono” (Taiz & Zeiger, 1998).
3.1. O CICLO DE CALVIN-BENSON:
A via bioquímica da incorporação do CO2 em carboidratos só começou a ser
compreendida quando em meados dos anos 40 os isótopos radioactivos de carbono
puderam ser utilizados em análises laboratoriais. O isótopo 14C que decai com

A Fotossíntese
46
emissão de raios foi utilizado como marcador por Melvin Calvin, Andrew Benson e
James Bassham, na Universidade de Berkeley, na Califórnia.
Estes autores desenvolveram algas, sobretudo do género Chlorella, na
presença de 14CO2 durante diferentes períodos de tempo, de seguida matavam
rapidamente as algas e extraiam os produtos da fotossíntese com solventes (figura
34).
Estes extractos eram, então, separados por cromatografia em papel, primeiro
numa direcção com um solvente e depois na direcção perpendicular à primeira com
outro solvente. O papel cromatográfico era colocado sob um filme próprio para raios-
X. Quando se revelava o filme apareciam manchas negras nos locais onde no croma-
tograma estavam os compostos que continham carbono radioactivo.
Muitos dos compostos marcados formavam-se rapidamente e, se o período
de exposição ao 14CO2 fosse reduzido para apenas meio segundo, então o único
composto marcado era o ácido 3-fosfoglicérico (3-PGA). Parecia, assim, que o
primeiro composto estável resultante da fixação do CO2 na fotossíntese era o 3-PGA,
que é uma molécula com 3 átomos de carbono (Hall & Rao, 1981).
Figura 34: Representação esquemática do aparelho (“lollipop”) utilizado para estudar a fixação de CO2
em algas fotossintéticas.
Retirado de Hall e Rao (1981), figura 6.1, página 52
Por degradação controlada do 3-PGA radioactivo estes autores mostraram
que o carbono terminal do grupo carboxilo ( COOH) era radioactivo. Como a
molécula de 3-PGA tem apenas 3 átomos de carbono, poder-se-ia pensar que o

A Fotossíntese
47
14CO2 se fixaria a um composto contendo 2 átomos de carbono, no entanto, nunca tal
composto foi encontrado.
Calvin e Benson procuraram um composto que se acumulasse quando se
removesse a fonte de CO2 após um período de fotossíntese, considerando que tal
composto seria o aceitador de CO2. Verificaram, então, que este composto é a
ribulose-1,5-bisfosfato (RuBP), que por adição de CO2 se decompõe em duas
moléculas de 3-PGA (figura 35). A enzima que catalisa esta reacção é a RuBP
carboxilase-oxigenase (RUBISCO – figura 36) que é a proteína mais abundante
nos tecidos verdes, e mesmo, segundo alguns autores, a proteína mais abundante na
Terra (Salisbury & Ross, 1992).
Figura 35: Esquema da reacção de fixação do CO2 na ribulose-1,5-bisfosfato (RuBP) catalisada pela
enzima Rubisco. Retirado de Salisbury e Ross (1991), página 226
Calvin e Benson verificaram que a assimilação do CO2 era um processo
cíclico e autocatalítico. Consoante os autores este ciclo pode ser chamado de “Ciclo
de Calvin-Benson”, “Via redutora da pentose fosfato (RPP)” ou ainda “Ciclo da
redução fotossintética do carbono (PCR)”. Este ciclo é o processo fundamental de
assimilação de carbono em todos os organismos fotossintéticos, incluindo os
procariotas (Salisbury & Ross, 1992).
3.1.1. A DESCRIÇÃO DO CICLO:
Arbitrariamente podemos considerar que o Ciclo de Calvin (figura 37)
começa pela carboxilação da RuBP originando duas moléculas de 3-PGA, reacção 1,
catalisada pela rubisco. A reacção 2 utiliza uma molécula de ATP para fosforilar o 3-
PGA a ácido-1,3-bisfosfato (1,3-bisPGA), que é reduzido a 3-fosfogliceraldeído (3-
GAP, ou 3-PGald)) pela NADP gliceraldeído-3-deshidrogenase. Esta última reacção é
a única redução do ciclo e é compreensivelmente de grande importância (Salisbury &
Ross, 1992).

A Fotossíntese
48
Figura 36: Estrutura da Rubisco. Nos cloroplastos das plantas a Rubisco é constituída por oito
subunidades maiores (L – large) e oito subunidades menores (S – small). Os quatro lobos visíveis na estrutura contêm cada um subunidades maiores e menores. As subunidades menores, das quais se podem ver quatro, estão representadas a vermelho. As subunidades maiores estão representadas a azul e a verde de forma a mostrar os limites dos dímeros. A Rubisco é considerada por muitos autores como sendo a proteína mais frequente na Terra e constitui cerca de metade doas proteínas do estroma cloroplastidial. Retirado de Buchanan et al. (2000), figura12.39, página 613
O 3-GAP é convertido a dihidroxiacetona fosfato (DHAP) na reacção 4. Estes
dois compostos são utilizados em reacções que conduzem à regeneração da RuBP.
Esta regeneração resulta da interconversão de compostos fosforilados com 3, 4, 5, 6
e 7 átomos de carbono, utilizando um ATP na passagem da ribulose-5-fosfato a
ribulose-1,5-bisfosfato (Salisbury & Ross, 1992).
O Ciclo de Calvin, que ocorre no estroma dos cloroplastos, pode ser resumido
a 3 etapas essenciais (figura 38):
carboxilação - envolve a fixação do CO2 na ribulose-1,5-bisfosfato
(RuBP) para formar duas moléculas de ácido 3-fosfoglicérico (3-PGA);
redução - envolve a redução das duas moléculas de 3-PGA a duas
moléculas de 3-fosfogliceraldeído (3-GAP), com consumo de 2 ATP e 2 NADPH (1
ATP e 1 NADPH por cada 3-PGA);
regeneração - envolve a formação de nova RuBP a partir de compostos
fosforilados de 3 a 7 átomos de carbono, e que consome outro ATP na fosforilação
da ribulose-5-fosfato a RuBP.
A adição de três moléculas de CO2 a três moléculas de RuBP (C5) leva à
formação de seis moléculas de 3-PGA cada uma das quais é fosforilada e reduzida
para formar o açúcar em três átomos de carbono GAP. Cinco GAP e três moléculas
de ATP são usadas para regenerar três moléculas de RuBP. A molécula de GAP
remanescente é o produto líquido da fixação de carbono e pode ser usada para
construir carboidratos e outros constituintes celulares.
As necessidades energéticas para a síntese de uma triose a partir de três
CO2 é de nove moléculas de ATP e seis moléculas de NADPH. Em condições
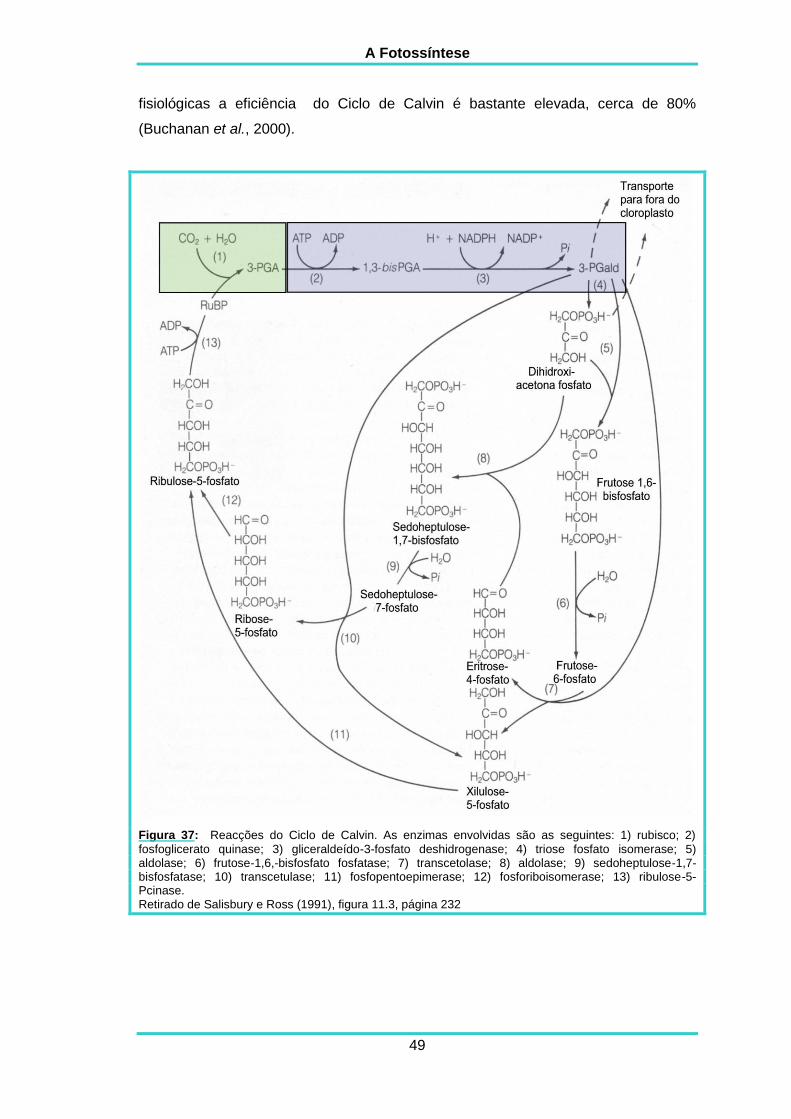
A Fotossíntese
49
fisiológicas a eficiência do Ciclo de Calvin é bastante elevada, cerca de 80%
(Buchanan et al., 2000).
Figura 37: Reacções do Ciclo de Calvin. As enzimas envolvidas são as seguintes: 1) rubisco; 2)
fosfoglicerato quinase; 3) gliceraldeído-3-fosfato deshidrogenase; 4) triose fosfato isomerase; 5) aldolase; 6) frutose-1,6,-bisfosfato fosfatase; 7) transcetolase; 8) aldolase; 9) sedoheptulose-1,7-bisfosfatase; 10) transcetulase; 11) fosfopentoepimerase; 12) fosforiboisomerase; 13) ribulose-5-Pcinase. Retirado de Salisbury e Ross (1991), figura 11.3, página 232

A Fotossíntese
50
Figura 38: O Ciclo de Calvin pode ser dividido em três fases: carboxilação, redução e regeneração. A
fixação de uma molécula de CO2 requer duas moléculas de NADPH e três de ATP. 3-PGA = 3-fosfoglicerato; GAP = gliceraldeído fosfato = fosfogliceraldeído. Retirado de Buchanan et al. (2000), figura 12.40, página 613
3.1.2. A REGULAÇÂO DO CICLO:
Muitas das enzimas do Ciclo de Calvin que catalisam reacções reversíveis
(como por exemplo a aldolase, a transcetolase, a gliceraldeído-3-fosfato
desidrogenase), são comuns à via glicolítica da degradação de carboidratos. Uma
vez que as enzimas envolvidas quer na síntese, quer na degradação de carboidratos
estão presentes no cloroplasto é fundamental que, à luz, o aparelho para a síntese
esteja “ligado” e o aparelho para a degradação esteja “desligado”. Isto é, são
necessários mecanismos regulatórios específicos para evitar uma ciclização inútil e
assegurar uma actividade óptima (Buchanan et al., 2000).

A Fotossíntese
51
3.1.2.1. Activação pelo ião Mg2+ e pelo pH:
Alterações na concentração em Mg2+ e no pH do estroma são reguladores
importantes de enzimas como a Rubisco, a frutose-1,6-bisfosfatase e
fosforibulocinase. A activação da Rubisco envolve a formação dum complexo
carbamato-Mg2+ num resíduo lisina no sítio activo (figura 39). A concentração em
Mg2+ do estroma é um factor importante nesta activação. À medida que a luz causa o
aumento de H+ no lúmen do tilacóide, são transportados iões Mg2+ daí para o estroma
para compensar o influxo de cargas positivas, e a concentração em Mg2+ passa de 1-
3 mM para cerca de 3-6 mM. A frutose-1,6-bisfosfatase e, em menor escala, outras
enzimas são activadas pelo aumento de pH na solução do estroma devido à luz
(Buchanan et al., 2000).
Figura 39: Activação da Rubisco por carbamilação. A Rubisco é activada pela ligação de CO2 e de
Mg2+
. A reacção de activação, que liberta protões, é promovida pelo aumento do pH e da concentração de Mg
2+ no estroma associado à iluminação do cloroplasto.
Retirado de Buchanan et al. (2000), figura 12.42, página 616
3.1.2.2. A regulação pela luz:
A modificação covalente de várias enzimas no Ciclo de Calvin constitui um
mecanismo independente de regulação pela luz. A activação de certas enzimas do
cloroplasto pela luz envolve um sistema regulador dependente de reduções (“redox-
dependent”) que inclui a ferredoxina, a tiorredoxina (uma proteína de baixo peso
molecular contendo radicais dissulfito) e a enzima ferredoxina-tiorredoxina redutase
(figura 40). A ferredoxina reduzida é produzida pelas reacções “à luz” da fotossíntese
e reage com a tiorredoxina f ou m oxidada numa reacção catalisada pela ferredoxina-
tiorredoxina redutase (FTR). A tiorredoxina pode por sua vez reduzir ligações
dissulfito intramoleculares de enzimas alvo, alterando, assim, a sua conformação e
modulando a sua actividade. No escuro os grupos sulfidrilo da enzima ficam
oxidados. As enzimas do Ciclo de Calvin sujeitas a regulação pela tiorredoxina

A Fotossíntese
52
reduzida são activadas pela redução à luz, e desactivadas pela oxidação às escuras.
Esta sequência de reacções liga as reacções à luz às reacções de fixação do CO2,
garantindo que a síntese de carboidratos decorre à luz. Ao mesmo tempo a
tiorredoxina reduzida formada à luz inibe os processos catabólicos que ocorrem
essencialmente às escuras, como por exemplo a via oxidativa das pentoses fosfato –
PPP (Buchanan et al., 2000).
Muitos dos mecanismos de regulação da Rubisco estão relacionados com a
sua função central na fixação de CO2. Ainda que se saiba que, por um lado a Rubisco
tem de estar completamente activada para funcionar correctamente na fixação de
CO2 e que por outro lado, a sua activação está dependente da sua ligação ao CO2,
há muitos aspectos que não estão esclarecidos. Por exemplo, in vitro a reacção de
carbamilação requer CO2 na ordem do milimolar, enquanto in vivo essa concentração
é na ordem do micromolar. Além disso, há moléculas de açúcar-fosfato que se ligam
à Rubisco afectando a reacção de activação com o CO2 (Buchanan et al., 2000).
Figura 40: O sistema ferredoxina-tiorredoxina constitui um mecanismo dependente da luz para a
activação e desactivação de enzimas. O transporte electrónico fotossintético que ocorre no PSI leva à redução da ferredoxina que é uma proteína Fe-S. A ferredoxina reduzida por sua vez reduz a tiorredoxina que é uma proteína regulatória, graças à acção da ferredoxina-tiorredoxina redutase. A tiorredoxina reduzida vai, então, reduzir as ligações dissulfito de numerosas proteínas alvo, modulando a sua actividade. A via de transferência electrónica está representada a vermelho. No escuro, o O2 oxida quer a enzima alvo, quer a tiorredoxina (não está representado neste esquema). Retirado de Buchanan et al. (2000), figura 12.43, página616
A enzima Rubisco activase está especificamente envolvida na activação da
Rubisco através da carbamilação. Embora não se conheçam os detalhes da sua
actividade, a Rubisco activase parece promover a dissociação de moléculas de

A Fotossíntese
53
açúcar fosfato ligadas ao sítio activo da Rubisco, permitindo assim que ocorra a
carbamilação. Aparentemente, ATP é hidrolisado como parte da reacção de
activação, ainda que esta reacção não esteja completamente esclarecida (figura 41).
Nalgumas plantas a actividade da Rubisco de folhas adaptadas ao escuro é
muito baixa e não pode ser reactivada pela adição de CO2 e de Mg2+; em
contrapartida a Rubisco de folhas da mesma planta destacadas ao meio dia é
activada por aquele tratamento. A diferença na activação está relacionada com a
presença de um inibidor fortemente ligado, o 2-carboxiarabinitol-1-fosfato (CA1P),
que é um análogo do intermediário em C6 do mecanismo catalítico da Rubisco.
Aparentemente existem dois mecanismos para a remoção do CA1P da Rubisco, uma
Figura 41: A Rubisco activase remove RuBP ligado à Rubisco inactiva e descarbamilada numa reacção
que envolve ATP. A Rubisco livre pode, então, ser activada por carbamilação, ligação de CO2 e Mg2+
numa reacção como a que é esquematizada na figura 39. A Rubisco activase pode também ser activada pela luz, via tioredoxina. Retirado de Buchanan et al. (2000), figura12.44, página 617
exige a acção da Rubisco activase e a outra envolvendo a degradação do inibidor
pela luz. Este facto explicaria os diferentes graus de inactivação da Rubisco
observada nas plantas adaptadas ao escuro ou à luz. Estes mecanismos de
regulação múltipla da Rubisco parecem indicar o papel chave que esta enzima
desempenha no metabolismo dos cloroplastos (Buchanan et al., 2000).
3.1.3. A ACTIVIDADE DE OXIGENASE DA RUBISCO E A FOTORRESPIRAÇÃO: A enzima rubisco mostra não apenas actividade como carboxilase, mas
também como oxigenase. Isto quer dizer que além de catalisar a reacção de fixação
do CO2 pela ribulose-1,5-bisfosfato, catalisa igualmente a fixação de O2 àquele
substrato. A descoberta deste processo permitiu explicar a libertação de CO2
associada ao consumo de O2, que se observa nas plantas expostas à luz. A
libertação de CO2 pelos orgãos verdes das plantas tem o nome de fotorrespiração.
No entanto, é importante realçar que este processo nada tem a ver com a respiração

A Fotossíntese
54
mitocondrial, também chamada respiração às escuras, excepto que, em ambos os
casos, há libertação de CO2 e consumo de O2.
Em condições atmosféricas normais, isto é, 21 % de O2 e 0.033 % de CO2, a
razão de ambos os processos é de 3:1. Nestas condições a actividade de oxigenase
da rubisco leva à formação de ácido fosfoglicólico que é desfosforilado para glicolato.
A formação do glicolato ocorre nos cloroplastos de plantas que apenas apresentam
Ciclo de Calvin (Plantas em C3), e o seu metabolismo é realizado pela chamada via
do glicolato.
A fotorrespiração no seu todo ocorre em três organelos celulares, os
cloroplastos, os peroxisomas e as mitocôndrias (figura 42). Dos cloroplastos o
glicolato é transportado para os peroxisomas onde é oxidado para formar glioxilato
com consumo de oxigénio. O glioxilato é, então, transaminado para glicina que
passa para as mitocôndrias. Duas moléculas de glicina produzem uma de serina com
libertação de CO2 e NH3. Esta reacção mitocondrial é a fonte do CO2 libertado pela
fotorrespiração (figura 43). É uma reacção importante porque o NH3 libertado deve
ser reencorporado em amino ácidos, de modo a que a formação de glicina possa
continuar. Este processo requer ATP e ferredoxina reduzida. A serina é então
convertida a 3-PGA por uma série de reacções que envolve a perda do seu grupo
amino e o ganho dum grupo fosfato a partir de ATP. Parte do 3-PGA é convertido a
RuBP e outra parte é convertida em sacarose e amido nos cloroplastos.
A equação geral para a fotorrespiração (parando o fluxo de carbono no 3-PGA
e abreviando ferredoxina para Fd) pode ser escrita da seguinte forma:
32
222 Fe Fd 2 Pi 3 ADP2PGA-3 3 CO Fe Fd 2OH ATP2O 3RUBP2
Podemos, então, ver que a fotorrespiração conserva em média 3/4 dos
carbonos retirados da RuBP (na forma de fosfoglicolato) quando o O2 reage com ele
(um carbono perdido por cada par de moléculas de ácido de dois carbonos formados
e por três O2 absorvidos. É importante notar que a fotorrespiração utiliza ATP e água
em vez de os formar, e também que requer um redutor (ferredoxina reduzida).

A Fotossíntese
55
Figura 42: Microfotografia que mostra a associação muito próxima de cloroplastos, peroxisomas (P) e
mitocôndrias (M) numa célula foliar. A matriz aparentemente cristalina do peroxisoma deve-se à enzima catalase, embora muitos peroxisomas com catalase não apresentem esta estrutura. Retirado de Salisbury & Ross (1992), figura 11.8, página 239
3.2. O METABOLISMO FOTOSSINTÉTICO EM C4:
Em 1965, KORTSHAK e colegas, no Hawaii, utilizando a cana de açúcar
(Saccharum officinarum), que é uma espécie em que a fotossíntese é invulgarmente
rápida e eficiente, verificaram que a maioria do CO2 é fixo inicialmente em ácidos
orgânicos com quatro átomos de carbono. Após cerca de um segundo a
fotossintetizar em presença de 14C, esta espécie apresentava 80 % do 14C fixo em
ácido málico e aspártico, e apenas 10 % em 3-PGA. Isto mostrava que o primeiro
produto da fotossíntese nesta espécie não era o 3-PGA. Estes resultados foram
rapidamente confirmados e ampliados para outras espécies por HATCH e SLACK
que trabalhavam na Austrália. A estas espécies foi dado o nome global de “plantas
em C4” por oposição às que formam em primeiro lugar 3-PGA e que são portanto
“plantas em C3” (Salisbury & Ross, 1992).
3.2.1. A ANATOMIA DE KRANZ:
As plantas que se seguem esta via fotossintética apresentam características
anatómicas próprias (figura 44). Assim, à volta dos feixes apresentam células
clorofilinas que são as chamadas células da bainha dos feixes à volta das quais se
dispõem as células do mesófilo. A esta anatomia dá-se o nome de anatomia de
Kranz (em alemão Kranz quer dizer coroa). As células das bainhas dos feixes
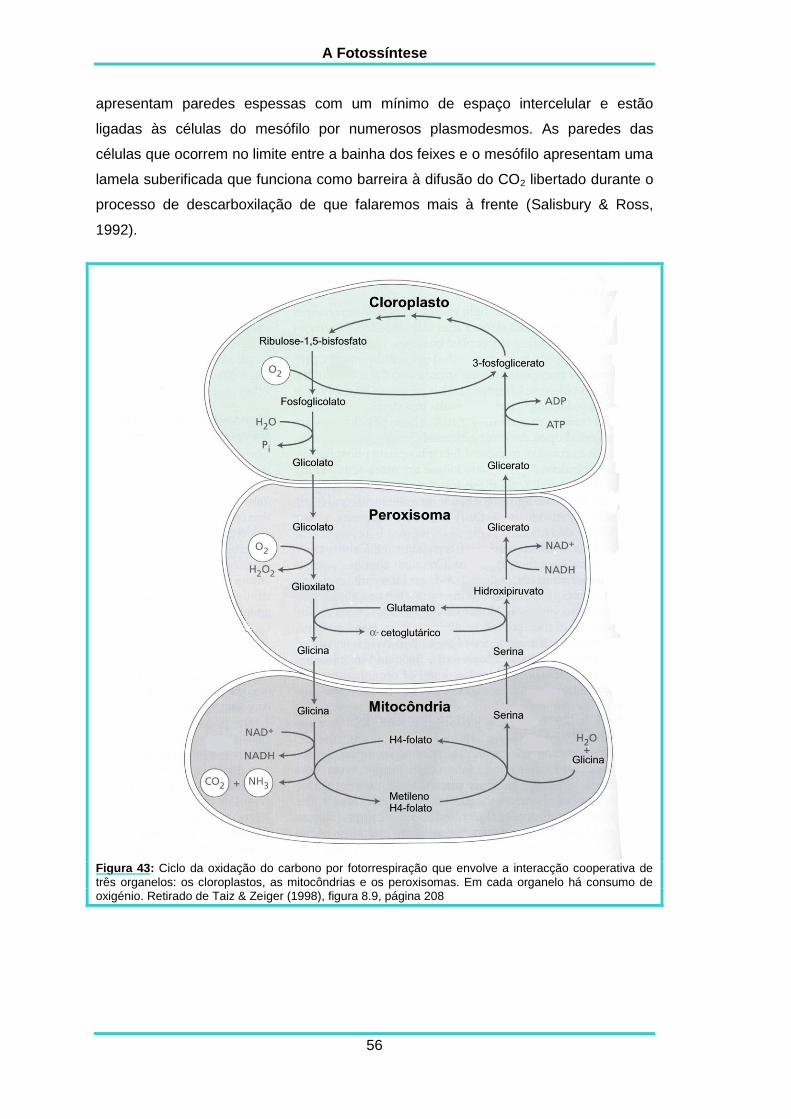
A Fotossíntese
56
apresentam paredes espessas com um mínimo de espaço intercelular e estão
ligadas às células do mesófilo por numerosos plasmodesmos. As paredes das
células que ocorrem no limite entre a bainha dos feixes e o mesófilo apresentam uma
lamela suberificada que funciona como barreira à difusão do CO2 libertado durante o
processo de descarboxilação de que falaremos mais à frente (Salisbury & Ross,
1992).
Figura 43: Ciclo da oxidação do carbono por fotorrespiração que envolve a interacção cooperativa de
três organelos: os cloroplastos, as mitocôndrias e os peroxisomas. Em cada organelo há consumo de oxigénio. Retirado de Taiz & Zeiger (1998), figura 8.9, página 208
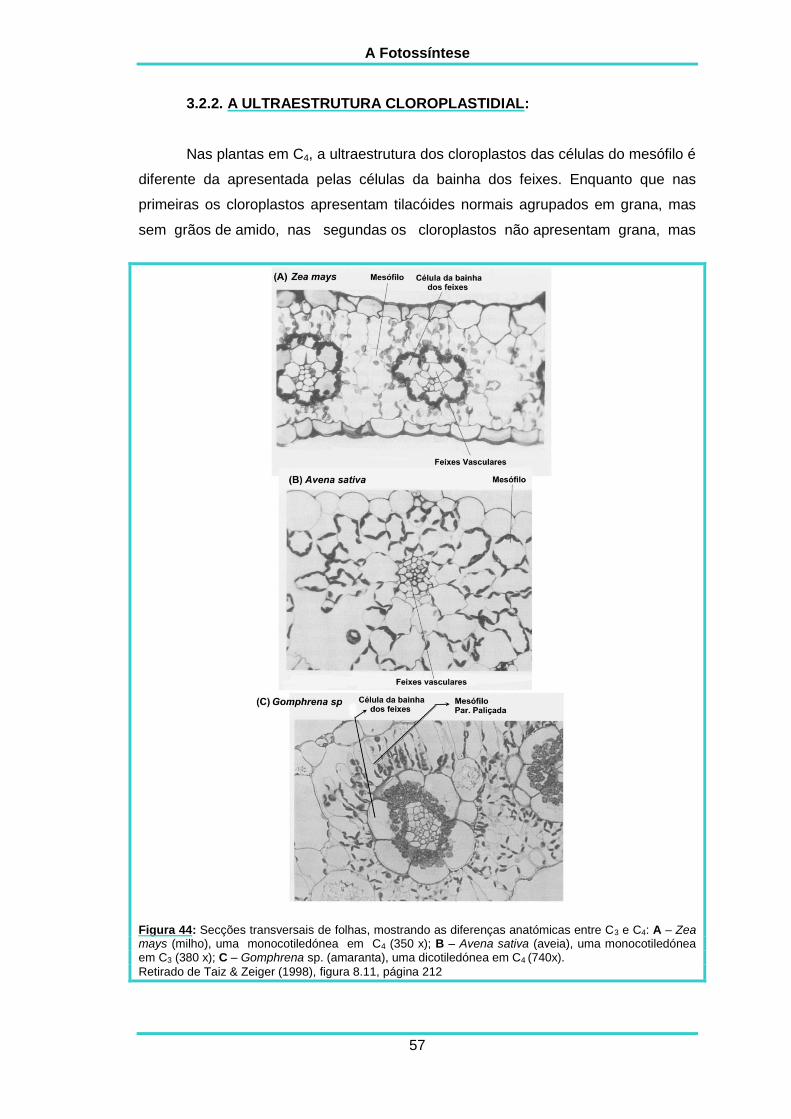
A Fotossíntese
57
3.2.2. A ULTRAESTRUTURA CLOROPLASTIDIAL:
Nas plantas em C4, a ultraestrutura dos cloroplastos das células do mesófilo é
diferente da apresentada pelas células da bainha dos feixes. Enquanto que nas
primeiras os cloroplastos apresentam tilacóides normais agrupados em grana, mas
sem grãos de amido, nas segundas os cloroplastos não apresentam grana, mas
Figura 44: Secções transversais de folhas, mostrando as diferenças anatómicas entre C3 e C4: A – Zea mays (milho), uma monocotiledónea em C4 (350 x); B – Avena sativa (aveia), uma monocotiledónea em C3 (380 x); C – Gomphrena sp. (amaranta), uma dicotiledónea em C4 (740x).
Retirado de Taiz & Zeiger (1998), figura 8.11, página 212
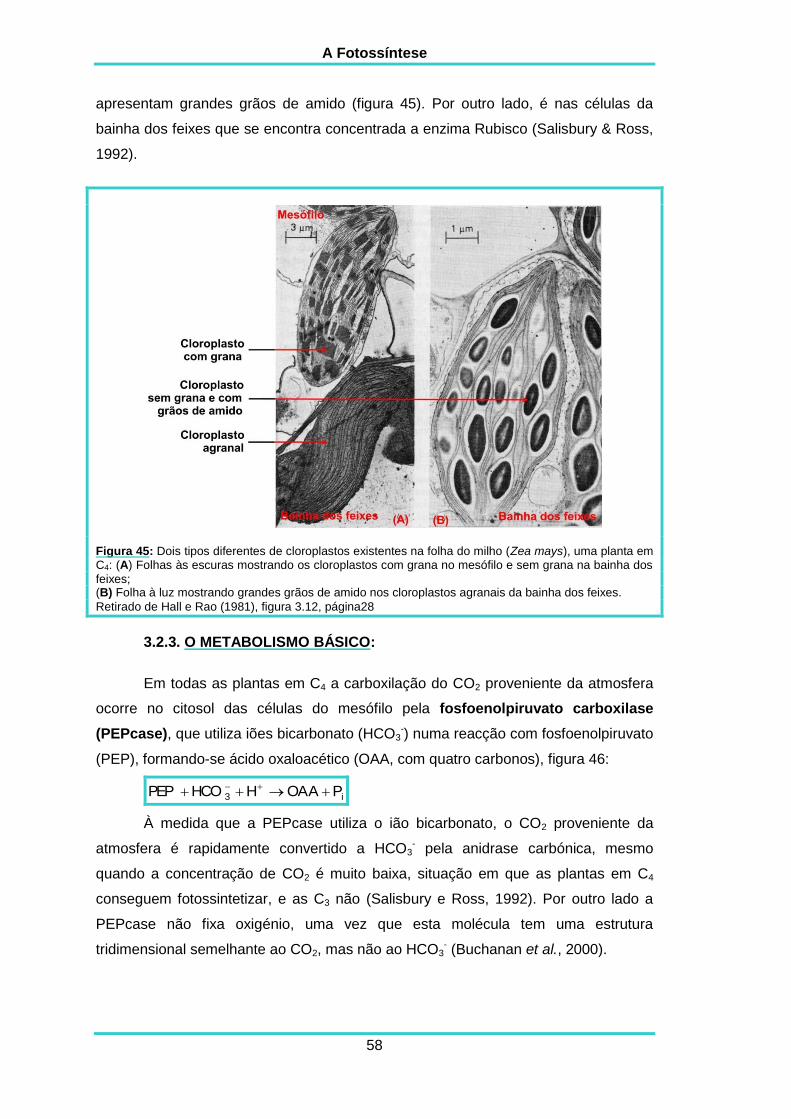
A Fotossíntese
58
apresentam grandes grãos de amido (figura 45). Por outro lado, é nas células da
bainha dos feixes que se encontra concentrada a enzima Rubisco (Salisbury & Ross,
1992).
Figura 45: Dois tipos diferentes de cloroplastos existentes na folha do milho (Zea mays), uma planta em C4: (A) Folhas às escuras mostrando os cloroplastos com grana no mesófilo e sem grana na bainha dos
feixes; (B) Folha à luz mostrando grandes grãos de amido nos cloroplastos agranais da bainha dos feixes.
Retirado de Hall e Rao (1981), figura 3.12, página28
3.2.3. O METABOLISMO BÁSICO:
Em todas as plantas em C4 a carboxilação do CO2 proveniente da atmosfera
ocorre no citosol das células do mesófilo pela fosfoenolpiruvato carboxilase
(PEPcase), que utiliza iões bicarbonato (HCO3-) numa reacção com fosfoenolpiruvato
(PEP), formando-se ácido oxaloacético (OAA, com quatro carbonos), figura 46:
i3 POAAHHCOPEP
À medida que a PEPcase utiliza o ião bicarbonato, o CO2 proveniente da
atmosfera é rapidamente convertido a HCO3- pela anidrase carbónica, mesmo
quando a concentração de CO2 é muito baixa, situação em que as plantas em C4
conseguem fotossintetizar, e as C3 não (Salisbury e Ross, 1992). Por outro lado a
PEPcase não fixa oxigénio, uma vez que esta molécula tem uma estrutura
tridimensional semelhante ao CO2, mas não ao HCO3- (Buchanan et al., 2000).
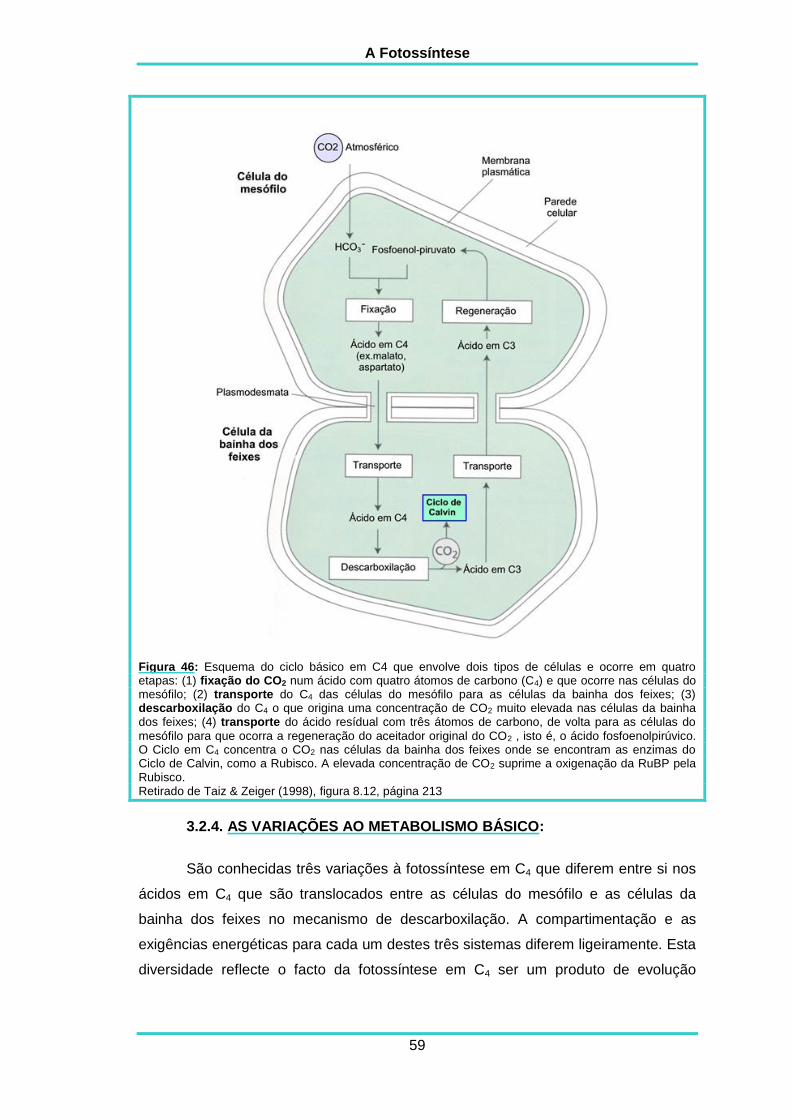
A Fotossíntese
59
Figura 46: Esquema do ciclo básico em C4 que envolve dois tipos de células e ocorre em quatro etapas: (1) fixação do CO2 num ácido com quatro átomos de carbono (C4) e que ocorre nas células do mesófilo; (2) transporte do C4 das células do mesófilo para as células da bainha dos feixes; (3) descarboxilação do C4 o que origina uma concentração de CO2 muito elevada nas células da bainha dos feixes; (4) transporte do ácido resídual com três átomos de carbono, de volta para as células do
mesófilo para que ocorra a regeneração do aceitador original do CO2 , isto é, o ácido fosfoenolpirúvico. O Ciclo em C4 concentra o CO2 nas células da bainha dos feixes onde se encontram as enzimas do Ciclo de Calvin, como a Rubisco. A elevada concentração de CO2 suprime a oxigenação da RuBP pela Rubisco. Retirado de Taiz & Zeiger (1998), figura 8.12, página 213
3.2.4. AS VARIAÇÕES AO METABOLISMO BÁSICO:
São conhecidas três variações à fotossíntese em C4 que diferem entre si nos
ácidos em C4 que são translocados entre as células do mesófilo e as células da
bainha dos feixes no mecanismo de descarboxilação. A compartimentação e as
exigências energéticas para cada um destes três sistemas diferem ligeiramente. Esta
diversidade reflecte o facto da fotossíntese em C4 ser um produto de evolução

A Fotossíntese
60
convergente tendo sido desenvolvido em várias ocasiões separadas e em taxa muito
diferentes (Buchanan et al., 2000):
Formadora de malato ou “NADP-ME” ou enzima málico dependente
do NADP+:
Neste tipo o OAA é reduzido a malato nos cloroplastos das células do
mesófilo utilizando-se NADPH, e a reacção é catalisada pela enzima málica
dependente do NADP+ (figura 47). O malato formado é então translocado para os
cloroplastos das células da bainha dos feixes onde é descarboxilado para piruvato, e
o CO2 libertado é carboxilado pela rubisco num Ciclo de Calvin normal, produzindo
duas moléculas de 3-PGA. O piruvato formado volta para as células do mesófilo onde
é reconvertido a fosfoenolpiruvato (PEP) com gasto de ATP (Lawlor, 1987).
Figura 47: Esquema do metabolismo fotossintético em C4 do tipo “NADP-ME”. Exemplos deste tipo:
Zea mays (milho); Sorghum bicolor (sorgo). Enzimas: 1. PEP carboxilase; 2. NADP+-malato
desidrogenase; 3. Enzima NADP+-málico; 4. Piruvato-ortofosfato dicinase (PPDK).
Retirado de Buchanan et al. (2000), figura 12.48(A), página 623
Formadoras de aspartato (I), ou “PCK”, ou ainda, fosfoenolpiruvato carboxicinase :
Neste tipo o OAA é convertido a aspartato no citosol das células do mesófilo.
O aspartato é então exportado para o citosol das células das bainhas dos feixes onde
é descarboxilado pela PEPcarboxicinase (PCK) para PEP com gasto de um ATP
(figura 48). De novo o CO2 libertado é utilizado no Ciclo de Calvin, nos cloroplastos.
O PEP é convertido a piruvato que é transaminado para alanina. A alanina é
exportada para o citosol das células do mesófilo onde é desaminada para piruvato
que é fosforilado para PEP (Lawlor, 1987).
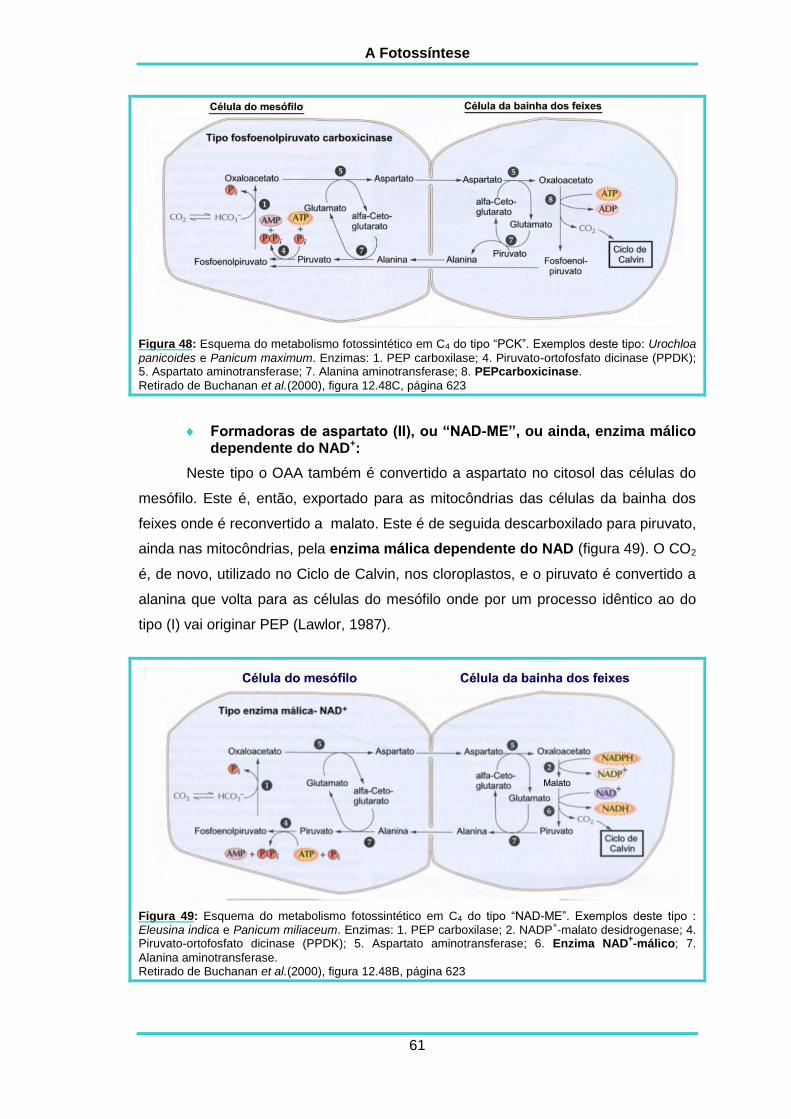
A Fotossíntese
61
Figura 48: Esquema do metabolismo fotossintético em C4 do tipo “PCK”. Exemplos deste tipo: Urochloa
panicoides e Panicum maximum. Enzimas: 1. PEP carboxilase; 4. Piruvato-ortofosfato dicinase (PPDK); 5. Aspartato aminotransferase; 7. Alanina aminotransferase; 8. PEPcarboxicinase.
Retirado de Buchanan et al.(2000), figura 12.48C, página 623
Formadoras de aspartato (II), ou “NAD-ME”, ou ainda, enzima málico dependente do NAD+:
Neste tipo o OAA também é convertido a aspartato no citosol das células do
mesófilo. Este é, então, exportado para as mitocôndrias das células da bainha dos
feixes onde é reconvertido a malato. Este é de seguida descarboxilado para piruvato,
ainda nas mitocôndrias, pela enzima málica dependente do NAD (figura 49). O CO2
é, de novo, utilizado no Ciclo de Calvin, nos cloroplastos, e o piruvato é convertido a
alanina que volta para as células do mesófilo onde por um processo idêntico ao do
tipo (I) vai originar PEP (Lawlor, 1987).
Figura 49: Esquema do metabolismo fotossintético em C4 do tipo “NAD-ME”. Exemplos deste tipo : Eleusina indica e Panicum miliaceum. Enzimas: 1. PEP carboxilase; 2. NADP
+-malato desidrogenase; 4.
Piruvato-ortofosfato dicinase (PPDK); 5. Aspartato aminotransferase; 6. Enzima NAD+-málico; 7.
Alanina aminotransferase. Retirado de Buchanan et al.(2000), figura 12.48B, página 623

A Fotossíntese
62
3.2.5. A REGULAÇÃO ENZIMÁTICA:
A NADP+malato desidrogenase, a PEP carboxilase e a PPDK que são
enzimas do ciclo em C4, são reguladas pela luz. Esta regulação é fundamental para
manter a coordenação entre as actividades das células do mesófilo e as da bainha
dos feixes e para garantir que há ácidos em C4 disponíveis para as células da bainha
dos feixes para a fixação do CO2 pela Rubisco no Ciclo de Calvin (Buchanan et al,
2000).
Os mecanismos de regulação são diferentes para cada enzima. A activação
pela luz da NADP+-malato desidrogenase ocorre no cloroplasto por via do sistema
ferredoxina-tiorredoxina que vimos anteriormente. Duas outras enzimas chaves nas
células do mesófilo, a PPDK e a PEPcarboxilase são reguladas por fosforilação das
proteínas.
A PEP carboxilase que está presente no citosol de folhas adaptadas ao
escuro é inibida por baixas concentrações de malato e tem uma afinidade baixa para
o seu substrato, isto é, o fosfoenol piruvato. Assim, esta forma da enzima está
essencialmente inactiva no escuro. À luz, uma serina cinase é activada que fosforila
especificamente a PEPcase (figura 50). A PEPcase fosforilada é muito menos
sensível à concentração em malato o que permite que esta enzima seja capaz de
funcionar como carboxilase mesmo em presença de concentrações elevadas de
malato nas células do mesófilo. O mecanismo que activa a cinase reguladora à luz
ainda não é compreendido.
A regulação da PPDK dos cloroplastos depende de uma proteína reguladora
específica e envolve uma série de reacções complexas que originam uma enzima
fosforilada e inactiva no escuro ou uma enzima desfosforilada e activa à luz. A fonte
do grupo fosfato é uma molécula de ADP em vez de ATP. A concentração do ADP no
escuro aumenta por não haver produção de ATP, o que aumenta a fosforilação da
PPDK . À luz a concentração em ADP diminui uma vez que há produção de ATP, o
que aumenta a proporção de PPDK desfosforilada e activa (figura 51). A proteína
reguladora é bicatalítica, catalisando quer a fosforilação quer a desfosforilação do
PPDK. Além disto, as concentrações em metabolitos, como o piruvato e o fosfato,
afectam a activação desta enzima, o que dá origem a um passo regulador altamente
sensível na via em C4. Tal como para o sistema de activação da PEPcase não se
conhece, ainda, a forma como a luz actua nos mecanismos de activação PPDK.
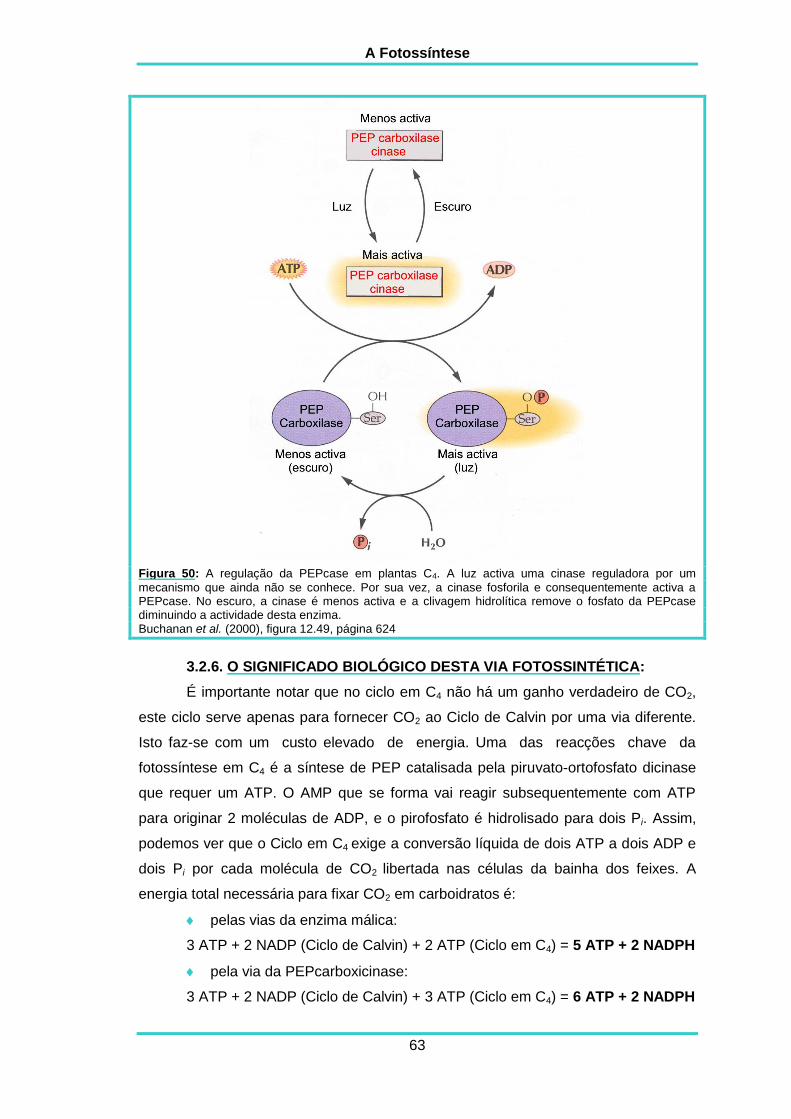
A Fotossíntese
63
Figura 50: A regulação da PEPcase em plantas C4. A luz activa uma cinase reguladora por um
mecanismo que ainda não se conhece. Por sua vez, a cinase fosforila e consequentemente activa a PEPcase. No escuro, a cinase é menos activa e a clivagem hidrolítica remove o fosfato da PEPcase diminuindo a actividade desta enzima. Buchanan et al. (2000), figura 12.49, página 624
3.2.6. O SIGNIFICADO BIOLÓGICO DESTA VIA FOTOSSINTÉTICA:
É importante notar que no ciclo em C4 não há um ganho verdadeiro de CO2,
este ciclo serve apenas para fornecer CO2 ao Ciclo de Calvin por uma via diferente.
Isto faz-se com um custo elevado de energia. Uma das reacções chave da
fotossíntese em C4 é a síntese de PEP catalisada pela piruvato-ortofosfato dicinase
que requer um ATP. O AMP que se forma vai reagir subsequentemente com ATP
para originar 2 moléculas de ADP, e o pirofosfato é hidrolisado para dois Pi. Assim,
podemos ver que o Ciclo em C4 exige a conversão líquida de dois ATP a dois ADP e
dois Pi por cada molécula de CO2 libertada nas células da bainha dos feixes. A
energia total necessária para fixar CO2 em carboidratos é:
pelas vias da enzima málica:
3 ATP + 2 NADP (Ciclo de Calvin) + 2 ATP (Ciclo em C4) = 5 ATP + 2 NADPH
pela via da PEPcarboxicinase:
3 ATP + 2 NADP (Ciclo de Calvin) + 3 ATP (Ciclo em C4) = 6 ATP + 2 NADPH

A Fotossíntese
64
Figura 51: A regulação da actividade da piruvato-ortofosfato dicinase (PPDK) é modulada por uma
proteína reguladora. Esta proteína promove a fosforilação do PPDK no escuro o que torna a enzima inactiva. No escuro, a concentração em ADP que é o dador de fosfato aumenta deviso à ausência da fotofosforilação. À luz o PPDK é desfosforilado e , assim, activado. Buchanan et al. (2000), página 12.50, página 625
Apesar das plantas em C3 consumirem menos ATP por CO2 fixo, a
fotorrespiração pode oxidar uma fracção significativa dos fotossintetisados. Por outro
lado, a afinidade da PEPcase para o CO2 é muito superior ao da Rubisco, pelo que
se pode pensar que este ciclo actua como concentrador de CO2 nas células da
bainha dos feixes onde está a Rubisco. Principalmente porque tudo aponta para que
o transporte de metabolitos entre o mesófilo e a bainha dos feixes ocorra por difusão
e não por transporte activo que consumiria energia.
Podemos perguntar qual é o “interesse biológico” do metabolismo em C4, uma
vez que exige maior dispêndio de energia e exige o movimento de metabolitos
através das membranas (embora se possa pensar que este movimento se faz por
difusão simples). De facto a fotossíntese em C4 é superior à C3 porque permite
uma assimilação eficiente do CO2, mesmo quando este está em concentrações
muito diluídas. A condutância estomática é mais pequena nas espécies em C4 que
nas espécies em C3, o que ajuda a conservar a água sem que a consequente
diminuição na concentração interna de CO2 afecte grandemente a fotossíntese. Isto
explica-se porque, por um lado a PEPcase é muito mais eficaz a fixar o CO2 que a
rubisco, e por outro lado a estrutura dos cloroplastos está modificada de forma a
optimizar a formação de ATP e NADPH (Lawlor, 1987).
A maioria das plantas em C4 são características de ambientes ensolarados e
quentes, muitas vezes com poucos recursos hídricos, embora não desérticos.
Existem também algumas espécies em C4 em habitats salinos, em que o

A Fotossíntese
65
metabolismo C4 pode ser uma vantagem na sobrevivência ao “stress” osmótico e
salino (Salisbury & Ross, 1992).
3.3. O METABOLISMO ÁCIDO DAS CRASSULÁCEAS (CAM):
Há já muito tempo que se sabe que as folhas de plantas pertencentes à
família das Crassuláceas (ex: Kalanchoë sp., Sedum sp., etc) apresentam um ritmo
diário de formação de ácidos orgânicos. Durante o dia o conteúdo total em ácidos
diminui na solução celular das folhas, enquanto durante a noite a noite, o conteúdo
em ácidos aumenta. Este ritmo de acidificação encontra-se também em plantas de
outras famílias como por exemplo, as Cactáceas e as Bromeliáceas. Todas as
plantas que seguem este ritmo de acidificação têm o chamado Metabolismo Ácido
das Crassuláceas (CAM).
Para além do metabolismo ácido das folhas, as plantas CAM apresentam
suculência das folhas e frequentemente dos pecíolos e dos caules, isto é, grande
espessura com uma razão superfície/volume baixa. Tanto quanto se sabe, todas as
plantas CAM apresentam uma certa suculência, embora existam espécies suculentas
que não seguem a via CAM, como é o caso das plantas halófitas (de habitats
salinos). Outra característica das plantas CAM é a estrutura das folhas, nos casos em
que estas se encontram bem desenvolvidas. Estas folhas apresentam células do
mesófilo de maiores dimensões, as mais exteriores contêm cloroplastos, as mais
interiores apresentam grandes vacúolos e não têm cloroplastos constituindo o
chamado hidrênquima. O tecido vascular não apresenta bainha à sua volta como
acontece com as C4 (figura 52).
O PEP utilizado é proveniente de carbohidratos de armazenagem,
provavelmente glucanos (amido e dextrano), que são mobilizados para produzir 3-
PGA e PEP. O ATP é utilizado, mas também é formado nestas reacções pelo que o
balanço é zero.
As plantas CAM assimilam CO2 e sintetizam ácidos orgânicos,
essencialmente malato, durante a noite, mas assimilam muito pouco CO2 durante o
dia (figura 53). Durante a noite os estomas abrem, oferecendo menor resistência à
difusão do CO2 para o interior das folhas. O CO2 é fixo no fosfoenolpiruvato (PEP),
pela fosfoenolpiruvato carboxilase (PEPcase), originando oxaloacetato (OAA),
provavelmente no citosol das células (figura 54). O OAA é reduzido para malato, com
dispêndio de NADH pela NAD malato deshidrogenase. O ácido málico formado é
transportado na forma hidrogenomalato através do tonoplasto e acumula-se no
vacúolo.

A Fotossíntese
66
Com a luz, os estomas fecham, e a fixação do CO2 da atmosfera diminui
muito rapidamente para uma taxa muito baixa (figura 53). O malato que estava
armazenado no vacúolo sai e é descarboxilado para piruvato pela enzima málica
NADP dependente e possivelmente pela enzima málica NAD dependente, ou ainda
pela PEP carboxicinase. O CO2 assim libertado entra nos cloroplastos e é assimilado
no Ciclo de Calvin utilizando ATP e NADPH provenientes do transporte electrónico. O
piruvato formado é fosforilado a PEP e reciclado para trioses fosfato e outros
carbohidratos que são armazenados (figura54).
Figura 52: Corte transversal duma folha duma planta CAM (Salsoli kali). Neste corte podemos observar
o mesófilo que é constituido por parênquima clorofilino em paliçada e por hidrênquima. Retirado de Fahn (1974), figura 108, página 126
No início do período de luz há pouca fixação de CO2 porque a PEPcase é
inibida pelo malato, mas à medida que este é descarboxilado a PEPcase fixa mais
CO2. Após uma exposição prolongada à luz e consequentemente uma
descarboxilação mais prolongada, o CO2 proveniente da atmosfera é assimilado
directamente pela rubisco, no Ciclo de Calvin, originando hidratos de carbono para
armazenagem.
O metabolismo CAM permite uma acumulação de CO2 durante o período
escuro, quando os estomas estão abertos, sem que haja uma perda substancial de
água. Esta característica é particularmente importante, uma vez que a maior parte
das plantas que exibem este tipo de metabolismo são características de habitats
quentes e secos (desérticos). Assim, estas plantas conseguem ter uma razão CO2fixo
/ água perdida muito elevada em condições que seriam letais para a maioria das
plantas em C3.
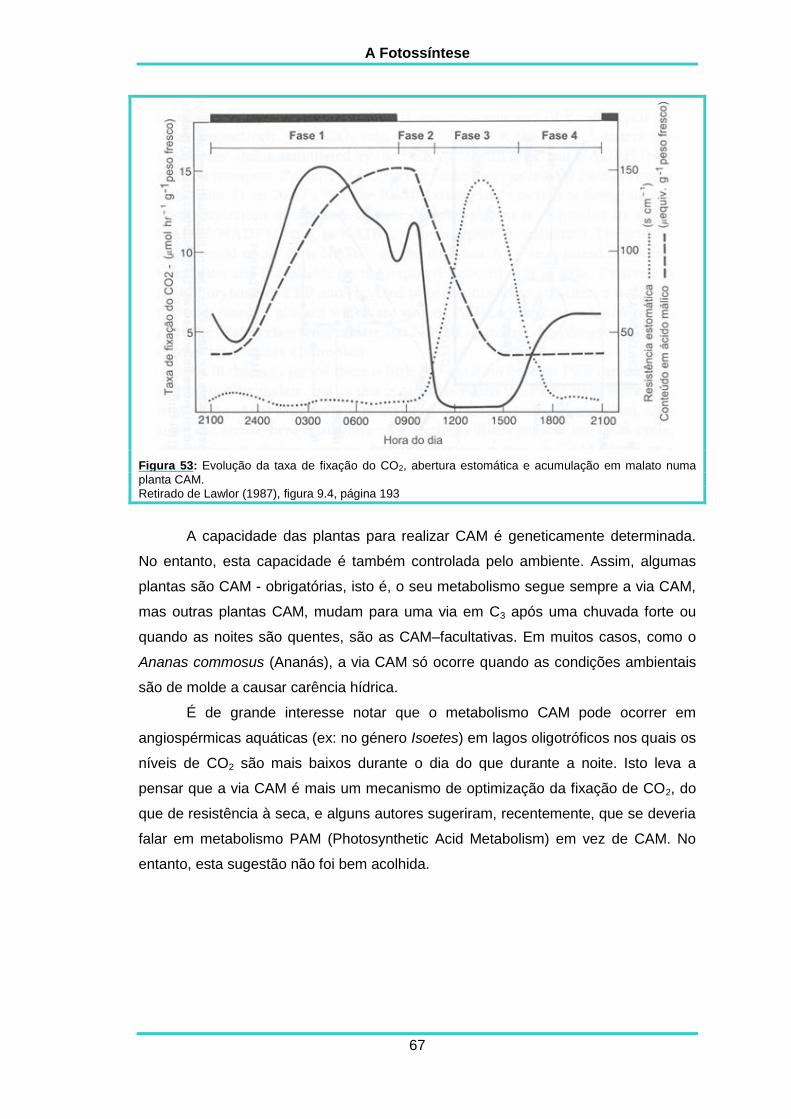
A Fotossíntese
67
Figura 53: Evolução da taxa de fixação do CO2, abertura estomática e acumulação em malato numa
planta CAM.
Retirado de Lawlor (1987), figura 9.4, página 193
A capacidade das plantas para realizar CAM é geneticamente determinada.
No entanto, esta capacidade é também controlada pelo ambiente. Assim, algumas
plantas são CAM - obrigatórias, isto é, o seu metabolismo segue sempre a via CAM,
mas outras plantas CAM, mudam para uma via em C3 após uma chuvada forte ou
quando as noites são quentes, são as CAM–facultativas. Em muitos casos, como o
Ananas commosus (Ananás), a via CAM só ocorre quando as condições ambientais
são de molde a causar carência hídrica.
É de grande interesse notar que o metabolismo CAM pode ocorrer em
angiospérmicas aquáticas (ex: no género Isoetes) em lagos oligotróficos nos quais os
níveis de CO2 são mais baixos durante o dia do que durante a noite. Isto leva a
pensar que a via CAM é mais um mecanismo de optimização da fixação de CO2, do
que de resistência à seca, e alguns autores sugeriram, recentemente, que se deveria
falar em metabolismo PAM (Photosynthetic Acid Metabolism) em vez de CAM. No
entanto, esta sugestão não foi bem acolhida.

A Fotossíntese
68
Figura 54: No metabolismo ácido das crassuláceas (CAM), as reacções de fotossíntese e as de
absorção de CO2 estão separadas no tempo: a absorção e a fixação de CO2 ocorrem de noite; a descarboxilação e a refixação do CO2 libertado no interior das células ocorrem durante o dia. O CAM é uma adaptação que permite essencialmente minimizar a quantidade de vapor de água que é perdido quando os estomas estão abertos para permitir a entrada de CO2. Nas plantas CAM os estomas estão abertos durante a noite, quando está mais fresco. Então o CO2 é fixo em ácido málico e armazenado no vacúolo. À medida que o ácido málico se acumula os vacúolos das folhas acidificam. Em presença da luz os estomas fecham e as folhas desacidificam. O ácido málico sai dos vacúolos e sofre uma descarboxilação. O CO2 que é libertado não sai uma vez que os estomas estão fechados e é assimilado através do Ciclo de Calvin utilizando o ATP e o NADPH que se formou nos processos fotoquímicos. Retirado de Taiz & Zeiger (1998), figura8.14, página 217
4. A FOTOSSÍNTESE E A DIFUSÃO DO CO2
Se pensarmos numa folha como um todo, a fotossíntese pode ser
considerada não só como a produção de assimilados, isto é, a produção de
biomassa, mas também como fluxo de CO2 para o interior da folha. Este fluxo ocorre
quase exclusivamente na forma de difusão.
Como vimos anteriormente, a difusão é um processo espontâneo que resulta
no movimento de substâncias de um local para outro, onde se encontram em
concentração mais baixa. Este processo ocorre quer na fase líquida, quer na gasosa.
A difusão resulta do movimento térmico ao acaso das moléculas. A transferência das
moléculas em movimento duma zona onde se encontram em concentração mais
elevada para uma de concentração mais baixa, tem maior probabilidade de acontecer
que o contrário. Por outras palavras, o número de moléculas por unidade de volume
é mais elevado nas zonas de maior concentração, de modo que, há mais moléculas a
difundirem-se para as zonas de menor concentração, que no sentido contrário.

A Fotossíntese
69
Quando o sistema se encontra isolado de efeitos exógenos, a difusão tende a
estabelecer um equilíbrio de concentrações.
Em 1855, FICK investigou os princípios quantitativos da difusão. A migração
de moléculas ao longo dum gradiente negativo de concentrações é semelhante ao
fluxo térmico ao longo dum gradiente negativo de temperaturas. Baseado na analogia
com o fluxo térmico, FICK foi capaz de deduzir que a força que dá origem ao
movimento de moléculas depende do gradiente de concentrações. Este gradiente é a
medida da mudança dum parâmetro com o aumento da distância, o que quer dizer
que o gradiente da concentração da substância j, na direcção x, pode ser expresso
na forma cj /x. Geralmente, o fluxo duma certa substância é proporcional à força
correspondente. Assim, o valor negativo da concentração, ou gradiente da substância
j, representa a força motriz e este processo pode ser expresso como -cj / x para a
difusão numa direcção, isto é, numa dimensão.
O sinal negativo indica que a difusão ocorre na direcção das concentrações
mais baixas, de modo que o fluxo da substância j, Jj, pode ser expresso como
(Sebanek, 1992):
x
c j
jj DJ
em que, Dj representa o coeficiente de difusão da substância j. Esta equação exprime
a primeira lei de FICK da difusão.
Em certos casos, é possível considerar, sem incorrer num erro muito grande
que, o coeficiente de difusão Dj é constante. Por convenção, o fluxo em direcção a
valores de x superiores é chamado fluxo positivo. No entanto, como o fluxo se
dirige no sentido das concentrações mais baixas é necessário usar o sinal (-) na
equação referida. Esta lei foi demonstrada experimentalmente.
A via seguida pelas moléculas de CO2 no decurso da fotossíntese é bastante
complexa. As moléculas de CO2 que se encontram no ar que rodeia as folhas devem
passar, em primeiro lugar, pela camada muito fina (décimos de milímetro) de ar
parado que rodeia as folhas, a chamada “camada adjacente” (ou em inglês
“boundary layer”), e assim, têm de ultrapassar a resistência que esta camada oferece
ao seu movimento, resistência da “camada adjacente” (ra). Depois, têm de passar
através dos estomas, ultrapassando a resistência estomática (rs). Finalmente, têm
de penetrar as paredes das células, o plasmalema, o citosol, e as membranas dos
cloroplastos, para se deslocarem para os locais de carboxilação, ou seja, o estroma
cloroplastidial. Esta última resistência tem o nome de resistência do mesófilo (rm), e

A Fotossíntese
70
envolve quer a resistência física ao transporte, quer a resistência química que
caracteriza a eficiência das reacções de carboxilação (figura 55).
O conceito de resistência à difusão, ou mais exactamente, da soma de
resistências à transferência de CO2 por difusão, pode ser ilustrado usando a primeira
parte da via de transporte do CO2 descrita anteriormente, isto é, o movimento através
da “camada adjacente”. A contribuição da resistência desta camada ao transporte de
CO2 é muito pequena, e é muitas vezes desprezada. Apesar disto, a sua análise
permite ilustrar os efeitos, fisiologicamente importantes, de alguns factores físicos do
ambiente sobre as plantas.
De acordo com a primeira lei de FICK, o transporte duma substância por
difusão (J), é dado pelo gradiente de concentrações (c), pelo coeficiente de difusão
D, e pela espessura da camada (a) através da qual a difusão está a decorrer
(Sebanek, 1992):
a
cDJ
Por analogia com a lei de OHM, em que o fluxo (J) é directamente
proporcional à diferença de potencial (E), e indirectamente proporcional à resistência
(R), ou seja, J = E / R, é possível escrever:
D
RR
cJ a
ou, ,
Baseado em estimativas da espessura da “camada adjacente”, é possível
exprimir a resistência contra a difusão de CO2, ou vapor de água.
Por exemplo, para a = 0.25 cm; e DH2O = 0.25 cm2 s-1 (coeficiente de difusão
do vapor de água no ar a 20 ºC) podemos escrever:
R = 0.25 (cm) / 0.25 (cm2 s-1) = 1 s cm-1
Analogamente, para uma “camada adjacente” 10 vezes mais fina (a = 0.025
cm),
R = 0.1 s cm-1.
Estes exemplos mostram o efeito decisivo que a espessura da “camada
adjacente” tem na grandeza da resistência correspondente. Assim, é importante
conhecermos os factores principais que afectam a sua espessura. Os mais
importantes são a velocidade do vento (u), e as dimensões da folha, expressas pelo
comprimento (l) na direcção do vento. Estes factores podem relacionar-se da
seguinte forma (Sebanek, 1992):
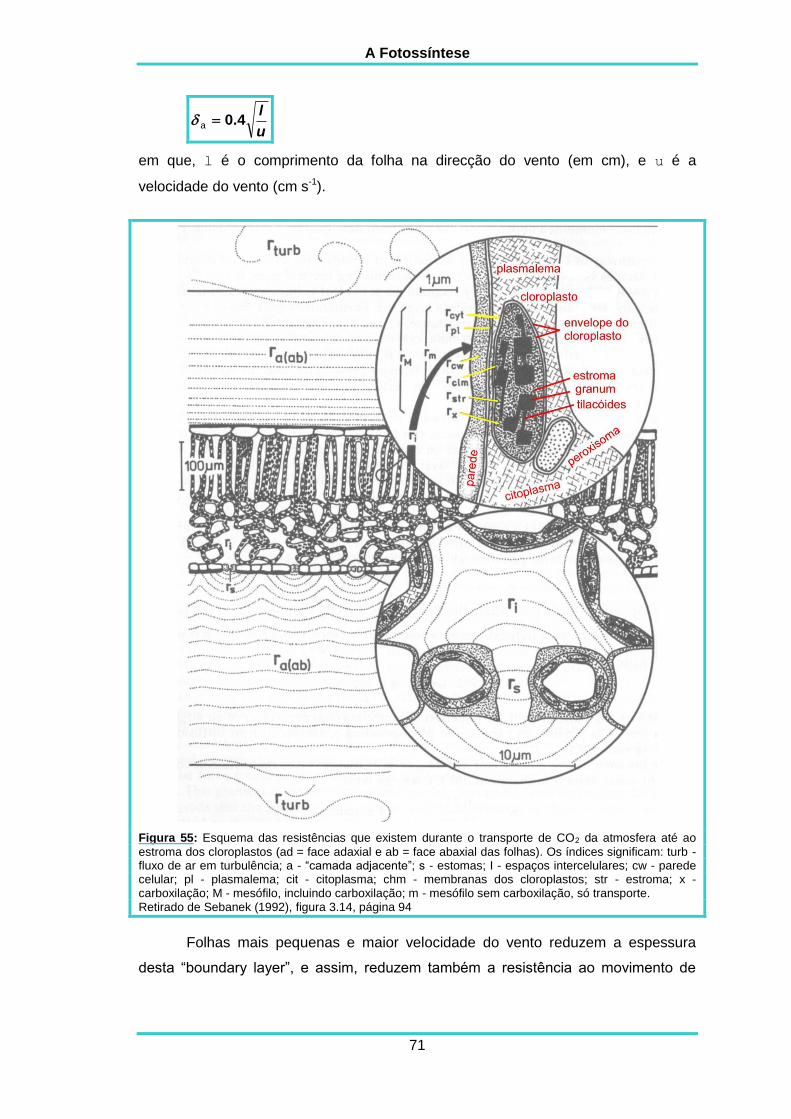
A Fotossíntese
71
u
l4.0a
em que, l é o comprimento da folha na direcção do vento (em cm), e u é a
velocidade do vento (cm s-1).
Figura 55: Esquema das resistências que existem durante o transporte de CO2 da atmosfera até ao
estroma dos cloroplastos (ad = face adaxial e ab = face abaxial das folhas). Os índices significam: turb - fluxo de ar em turbulência; a - “camada adjacente”; s - estomas; I - espaços intercelulares; cw - parede celular; pl - plasmalema; cit - citoplasma; chm - membranas dos cloroplastos; str - estroma; x - carboxilação; M - mesófilo, incluindo carboxilação; m - mesófilo sem carboxilação, só transporte. Retirado de Sebanek (1992), figura 3.14, página 94
Folhas mais pequenas e maior velocidade do vento reduzem a espessura
desta “boundary layer”, e assim, reduzem também a resistência ao movimento de

A Fotossíntese
72
vapor de água, CO2 e calor. As trocas com o ambiente, nestas circunstâncias, são
muito maiores do que para folhas grandes ou baixa velocidade do vento.
A análise da resistência à difusão contribui muito para o estudo da fisiologia
da fotossíntese, particularmente para a determinação da chamada taxa de
fotossíntese líquida (PN), isto é, a taxa de absorção do CO2 por unidade de área
foliar e por unidade de tempo (mol m-2 s-1):
R
COPN
2
em que, CO2 é a diferença de concentrações entre a atmosfera que rodeia a folha e
o local de carboxilação, e R é o valor da resistência à difusão. Este último valor pode
ser calculado a partir da equação anterior:
NP
COR 2
em que CO2 e PN podem ser obtidos experimentalmente.
Com base na via da difusão para o CO2, descrita anteriormente, é possível
dividir a resistência total à difusão em três componentes:
ra - representa a resistência à difusão pela “bounday layer”;
rs - caracteriza a resistência à difusão através dos poros estomáticos e
espaços intercelulares. Tendo em conta a grande variabilidade na largura dos poros
estomáticos, esta resistência exprime acima de tudo a abertura dos referidos poros;
rm - envolve a componente de transporte real, isto é, o transporte através da
parede celular, plasmalema, citosol, membrana cloroplastidial e estroma.
Todas estas resistências podem ser dispostas em série, de modo que a
resistência total é:
msa rrrR
e, do mesmo modo a taxa de fotossíntese pode ser dada por:
msa
claN
rrr
COCOP
)()( 22
em que, (CO2)a é a concentração de CO2 na atmosfera que rodeia a folha, e (CO2)cl é
a concentração nos cloroplastos, que varia de 0 a , ou seja, a concentração mínima
de CO2 nos espaços intercelulares (Sebanek, 1992).
Duma maneira geral, as folhas da maioria das plantas são anfiestomáticas,
isto é, têm estomas em ambas as faces. Anteriormente, expressámos as trocas
gasosas em termos de unidade de área da folha inteira e, as diferenças de anatomia
e morfologia existentes entre as duas faces não foram tomadas em consideração. A

A Fotossíntese
73
absorção de CO2 através de ambos os lados duma folha pode ser comparado com
um esquema de resistências eléctricas. As resistências da face superior ou da face
inferior estão em série, mas em conjunto representam um arranjo em paralelo.
Assim, a resistência total pode ser descrita por2:
um
us
ua
lm
ls
la
lm
ls
la
um
us
ua
COrrrrrr
rrrrrrR
2
em que os expoentes u e l indicam a resistência nas faces superior (“upper”) e
inferior (“lower”), respectivamente.
Este procedimento não é uma complicação desnecessária. De facto, verifica-
se experimentalmente que os resultados obtidos quando se considera a folha como
um todo, ou quando se consideram ambas as faces em paralelo, são
significativamente diferentes.
Na figura 56 podemos ver o esquema de uma análise mais detalhada dos
fluxos de CO2 existentes entre a atmosfera e o local de carboxilação, com as
resistências encontradas em plantas em C3 e C4.
Na análise feita até agora, não tivemos em consideração outros tipos de
fluxos de CO2 que ocorrem na folha, isto é, os que são condicionados pela respiração
mitocondrial e fotorrespiração (figura 57). Desprezar esta reassimilação de CO2
resulta numa sobrestimação da chamada fotossíntese total, ou taxa de carboxilação
real. Além disto, a apreciação correcta destes fluxos permite uma formulação mais
realística das taxas e direcções dos fluxos de CO2 nas folhas.
Utilizando modelos matemáticos baseados no conhecimento dos aspectos
bioquímicos da fotossíntese tem sido possível obter muitos dados sobre as
interacções que se estabelecem entre as taxas de fotossíntese, respiração e
fotorrespiração (Sebanek, 1992).
2
lm
ls
la
um
us
ua
um
us
ua
lm
ls
la
COlm
ls
la
um
us
ualuCO rrrrrr
rrrrrrR
rrr
1
rrr
1
R
1
R
1
R
12
2

A Fotossíntese
74
Figura 56: Esquema das principais resistências (r) e concentrações em CO2 (c) na via de transporte do
CO2, entre a atmosfera e o local de carboxilação. Os índices são os mesmos que na figura 3.36. F - fotossíntese líquida; RL - fotorrespiração; RD - respiração; PG - fotossíntese total. Retirado de Sebanek (1992), figura 3.15, página 96
Quando se estuda os componentes individuais da resistência total à
transferência de CO2 durante a fotossíntese, é possível obter dados sobre a
localização do efeito causado por um determinado factor. Assim, a avaliação dos
efeitos de diferentes irrigações, fertilizantes, reguladores de crescimento, etc.,
permite revelar o mecanismo destes efeitos e realizar uma generalização de
resultados concretos. É evidente que uma alteração em rs indica um efeito a nível dos
estomas, enquanto mudanças no rm sugere alterações na estrutura anatómica ou nas
reacções bioquímicas.

A Fotossíntese
75
Figura 57: Relações mútuas entre fotossíntese total (PG), fotorrespiração (RL) e respiração mitocondrial
(RD), expressa em fluxos de oxigénio. Retirado de Sebanek (1992), figura 3.16, página97
A taxa de fotossíntese é afectada por numerosos factores externos, como
veremos no ponto a seguir. No entanto, é evidente que a taxa de fixação do CO2 é
igualmente afectada pela estrutura dos orgãos de assimilação, ou seja, número e
tamanho dos cloroplastos, forma das células, estrutura dos espaços intercelulares,
espessura da folha e o número dos estomas. O pressuposto fluxo de CO2 apenas
numa direcção representa uma grande desvantagem do conceito de resistência à
difusão como foi apresentado mais acima. A descrição quantitativa da estrutura da
folha apresentada é evidentemente insuficiente para a formulação dum modelo mais
realístico, tridimensional, do transporte de CO2 dentro da folha. No entanto, é
possível derivar intuitivamente a existência duma dependência na estrutura
anatómica da folha, do fluxo de CO2 para os cloroplastos. Estudaremos, a seguir,
dois exemplos (Sebanek, 1992):
Vários autores consideram que a área interna duma folha (Ames) é mais de dez
vezes superior à sua área externa (A). Pode considerar-se que a maior parte dessa
superfície interna está “coberta” por uma camada de cloroplastos. A taxa de
fotossíntese total (PG) relaciona-se com a taxa de fluxo de CO2 para os cloroplastos,
cujo volume total é dcl. A taxa de fotossíntese total pode também ser expressa como
a taxa de fotossíntese por unidade de volume de cloroplastos (Pcl) em mol (CO2) s-1
cm-3. Se a média da espessura da camada de cloroplastos for cl, então a taxa de
fotossíntese por unidade de área da folha pode ser expressa como:

A Fotossíntese
76
A/APP mesclclG
Esta relação indica que a mudança da superfície interna da folha afecta
consequentemente a taxa de fotossíntese por unidade de área.
O segundo exemplo está associado com a relação entre a frequência dos
estomas (n) e a grandeza da resistência estomática (rs). Alguns autores deduziram a
seguinte equação:
)./(.
)/.4ln(
..nD
a
ba
ba
drs
em que, a é metade do comprimento da abertura estomática, b é metade da largura
da abertura estomática, d é a profundidade do poro estomático, D é o coeficiente de
difusão.
Nas folhas de plantas jovens de cevada (Hordeum vulgare) foram observados
os seguintes valores para o poro estomático:
a = 14.4 x10-6 m; b = 1.0 x 10-6 m; d = 15.8 x 10-6 m; n = 11.2 x 10-7.
Substituindo estes valores na equação anterior obtemos o valor de rs = 207 s
m-1. Este valor está de acordo com os que se obtiveram por meio de métodos que
utilizam as trocas gasosas. Esta relação indica também a dependência da resistência
estomática com o número de estomas.
5. COMPARAÇÃO ENTRE OS TRÊS TIPOS DE METABOLISMO
A taxa de fotossíntese líquida (PN) é afectada por muitos factores externos
(como a luz, a concentração em CO2, a temperatura, etc.). Além disso, a história da
planta, isto é, o ambiente onde se desenvolveu influencia as suas características
fotossintéticas. No entanto, as diferenças resultantes do tipo de metabolismo
fotossintético, isto é, C3, C4 e CAM, por vezes sobrepõem-se às restantes.
Assim, é importante estudar-se a forma como os vários factores ambientais e
internos influenciam os vários tipos de metabolismo fotossintético.
5.1. EXEMPLOS DE ESPÉCIES EM C3, C4 E CAM:
C3: Beta vulgaris, Chenopodium album, Spinacea oleracea, Convolvulus arvensis, Raphanus sativus, Curcubita pepo, Triticum aestivum, Hordeum vulgare, Avena sativa, Pisum sativum, Nicotiana tabacum. C4: Zea mays, Saccharum officinarum, Atriplex sabulosa, Cyperus esculentus, Panicum miliaceum, Sorghum bicolor, Digitaria sanguinalis. CAM: Agave americana, Kalanchoe daigremontina, Euphorbia grandidens, Germanium pratensis. (Sebanek, 1992)

A Fotossíntese
77
5.2. CONDIÇÕES AMBIENTAIS NATURAIS:
C3: Clima temperado. C4: Condições de carência hídrica moderada, elevada intensidade de radiação solar, humidade relativa baixa. CAM: Condições áridas, isto é, seca, elevada intensidade luminosa, temperaturas diurnas elevadas e nocturnas baixas (Sebanek, 1992).
5.3. ANATOMIA FOLIAR:
C3: Mesófilo geralmente diferenciado em parênquima esponjoso e parênquima em paliçada. C4: Mesófilo com bainha dos feixes (Anatomia de Kranz). CAM: Células do mesófilo com grandes vacúolos (Sebanek, 1992).
5.4. ESTRUTURA DOS CLOROPLASTOS:
C3: Cloroplastos típicos com grana, grãos de amido, e uma razão de clorofila a : b = 3 : 1. C4: Dimorfismo cloroplastidial: nas células do mesófilo os cloroplastos apresentam grana, mas não apresentam grãos de amido, e têm uma razão de clorofilas a : b = 3 : 1; nas células das bainhas dos feixes os cloroplastos são agranais com grandes grãos de amido e uma razão de clorofilas a : b = 5 : 1. CAM: Cloroplastos com pouco grana, com numerosos grãos de amido e uma razão de clorofilas a : b = 3 : 1 (Sebanek, 1992).
5.5. ENZIMAS FIXADORAS DE CO2:
C3: Rubisco (Ribulose-1,5-bisfosfato carboxilase oxigenase) C4: PEPcase (Fosfoenolpiruvato carboxilase) e Rubisco, separadas no espaço. CAM: PEPcase e Rubisco, separadas no tempo (Sebanek, 1992).
5.6. PRODUTOS PRIMÁRIOS DA FIXAÇÃO DO CO2:
C3: Ácido 3-fosfoglicérico (3-PGA). C4: Ácido oxaloacético (OAA). CAM: Ácido oxaloacético (OAA) (Sebanek, 1992).
5.7. NECESSIDADES FOTOQUÍMICAS POR MOLÉCULA DE CO2
FIXA:
C3: 3 ATP e 2 NADPH + H+ C4: 5 ATP e 2 NADPH + H+ CAM: 5 ATP e 2 NADPH + H+ (Sebanek, 1992).

A Fotossíntese
78
5.8. EFEITOS DA INTENSIDADE DA LUZ:
A taxa de fotossíntese líquida duma folha, numa atmosfera de 350 mol mol-1
CO2, varia em função da intensidade luminosa (figura 58). No escuro não existe
fixação de CO2, excepto para as CAM. Para intensidades luminosas muito baixas a
perda de CO2 através da respiração é superior ao ganho através da fotossíntese, o
que permite explicar os valores negativos para a fotossíntese líquida que se
observam na parte esquerda do gráfico (Taiz & Zeiger, 1998)
A partir de certo valor de irradiância, mesmo que esta aumente não há
aumento da taxa de fotossíntese líquida (setas a vermelho no gráfico), é o ponto de
saturação para a luz (Salisbury & Ross, 1992). Entre a escuridão e o ponto de
saturação existe uma intensidade luminosa para a qual o CO2 fixo na fotossíntese é
igual ao CO2 libertado na respiração, é o chamado ponto de compensação para a
luz. Este ponto varia com as espécies, com a irradiância durante o crescimento, com
a temperatura e a concentração de CO2 quando se fazem as medições. Apenas pode
haver crescimento da planta quando a irradiância está acima do ponto de
compensação (Salisbury & Ross, 1992).
Na figura 58 podemos ver as respostas que alterações na irradiância numa
folha, causam em três dicotiledóneas: uma C4 do deserto, Tridestromia oblongifolia,
uma C3 da orla marítima, Atriplex hastata e por fim, uma planta que se encontra no
solo duma floresta tropical húmida, Alocasia macrorrhiza.
As respostas da Alocasia são típicas de muitas espécies de ambientes
sombrios - plantas de sombra - incluindo a maioria das plantas de interior, e
caracterizam-se por:
1. quando expostas a elevadas intensidades luminosas, estas espécies
exibem taxas de fotossíntese muito inferiores às das outras espécies que se
desenvolveram em ambientes mais luminosos;
2. o processo fotossintético satura para intensidades luminosas muito baixas;
3. para níveis de irradiância baixos estas espécies fotossintetisam com taxas
muito mais elevadas que as restantes espécies (a título meramente exemplificativo
podemos indicar valores na ordem dos 1 a 5 mol m-2 s-1 para plantas de
sombra e de 10 a 20 mol m-2 s-1 para plantas de Sol. Taiz & Zeiger, 1998);
4. apresentam pontos de compensação para a luz muito baixos.
Estas características permitem-lhes crescer lentamente no seu ambiente
natural, sobrevivendo onde espécies com pontos de compensação mais elevados
morreriam (Salisbury & Ross, 1992)
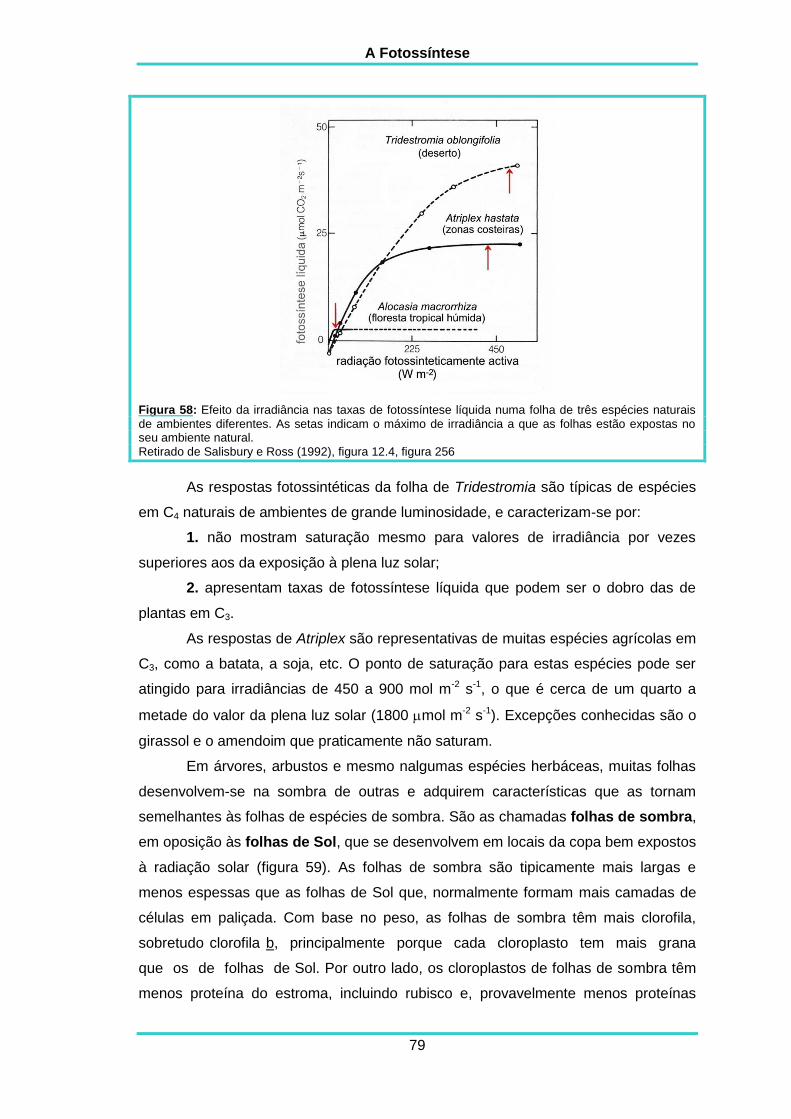
A Fotossíntese
79
Figura 58: Efeito da irradiância nas taxas de fotossíntese líquida numa folha de três espécies naturais
de ambientes diferentes. As setas indicam o máximo de irradiância a que as folhas estão expostas no seu ambiente natural.
Retirado de Salisbury e Ross (1992), figura 12.4, figura 256
As respostas fotossintéticas da folha de Tridestromia são típicas de espécies
em C4 naturais de ambientes de grande luminosidade, e caracterizam-se por:
1. não mostram saturação mesmo para valores de irradiância por vezes
superiores aos da exposição à plena luz solar;
2. apresentam taxas de fotossíntese líquida que podem ser o dobro das de
plantas em C3.
As respostas de Atriplex são representativas de muitas espécies agrícolas em
C3, como a batata, a soja, etc. O ponto de saturação para estas espécies pode ser
atingido para irradiâncias de 450 a 900 mol m-2 s-1, o que é cerca de um quarto a
metade do valor da plena luz solar (1800 mol m-2 s-1). Excepções conhecidas são o
girassol e o amendoim que praticamente não saturam.
Em árvores, arbustos e mesmo nalgumas espécies herbáceas, muitas folhas
desenvolvem-se na sombra de outras e adquirem características que as tornam
semelhantes às folhas de espécies de sombra. São as chamadas folhas de sombra,
em oposição às folhas de Sol, que se desenvolvem em locais da copa bem expostos
à radiação solar (figura 59). As folhas de sombra são tipicamente mais largas e
menos espessas que as folhas de Sol que, normalmente formam mais camadas de
células em paliçada. Com base no peso, as folhas de sombra têm mais clorofila,
sobretudo clorofila b, principalmente porque cada cloroplasto tem mais grana
que os de folhas de Sol. Por outro lado, os cloroplastos de folhas de sombra têm
menos proteína do estroma, incluindo rubisco e, provavelmente menos proteínas

A Fotossíntese
80
associadas ao transporte electrónico que os cloroplastos das folhas de Sol. Assim, as
folhas de sombra investem mais energia na produção de pigmentos colectores de
radiação, enquanto as mais expostas investem essencialmente nas enzimas
responsáveis pela fixação de CO2 (Salisbury & Ross, 1992).
As respostas fotossintéticas dos três tipos de metabolismo, a diferentes
intensidades luminosas podem ser sintetizadas da seguinte forma:
C3: a fotossíntese satura para um quarto a um meio da intensidade luminosa da luz plena do Sol, em condições de Verão de climas temperados; C4: a fotossíntese geralmente não satura, mesmo em condições de máxima irradiância; CAM: as plantas saturam mesmo para intensidades luminosas de cerca de um décimo da irradiância solar (Sebanek, 1992).
Figura 59: Corte transversal de folhas de Acer saccharinum. a) folha de Sol, do lado Sul de uma árvore
isolada. É de notar a cutícula espessa na epiderme superior; b) folha de sombra do meio duma copa duma árvore isolada; c) e d) folhas de sombra, da parte de baixo da copa de árvores numa floresta.
Retirado de Salisbury e Ross (1992), figura 12.5, página 256
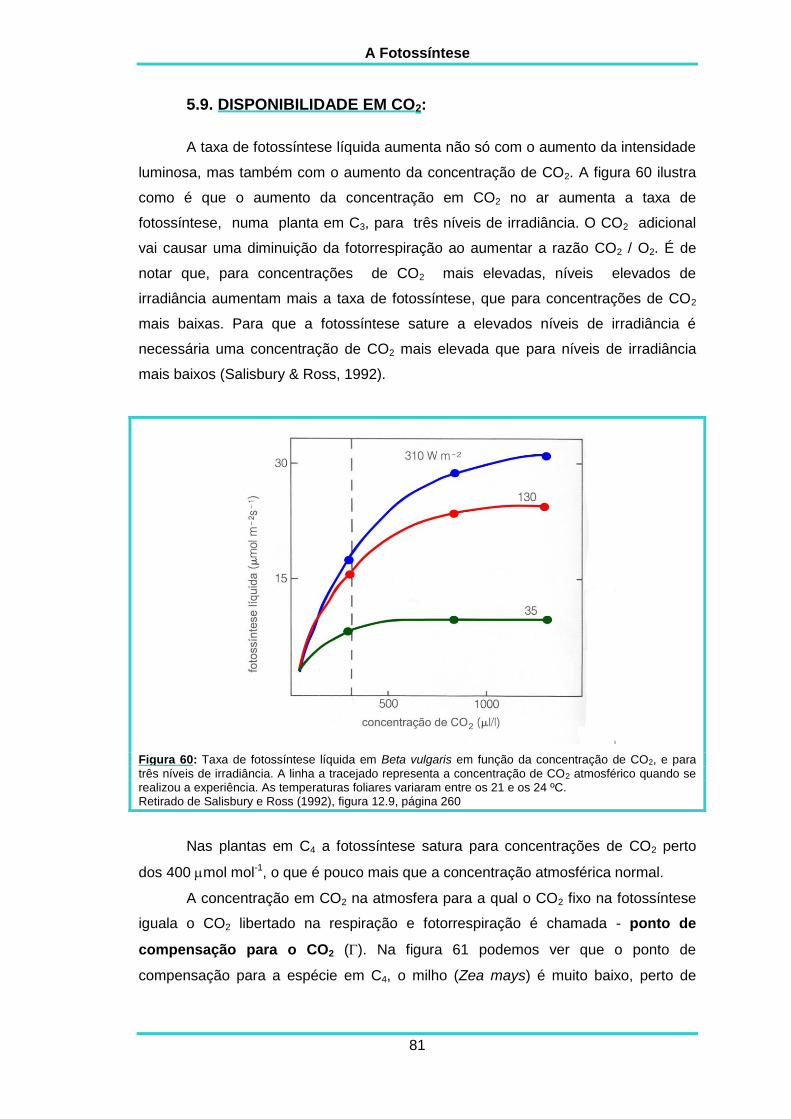
A Fotossíntese
81
5.9. DISPONIBILIDADE EM CO2:
A taxa de fotossíntese líquida aumenta não só com o aumento da intensidade
luminosa, mas também com o aumento da concentração de CO2. A figura 60 ilustra
como é que o aumento da concentração em CO2 no ar aumenta a taxa de
fotossíntese, numa planta em C3, para três níveis de irradiância. O CO2 adicional
vai causar uma diminuição da fotorrespiração ao aumentar a razão CO2 / O2. É de
notar que, para concentrações de CO2 mais elevadas, níveis elevados de
irradiância aumentam mais a taxa de fotossíntese, que para concentrações de CO2
mais baixas. Para que a fotossíntese sature a elevados níveis de irradiância é
necessária uma concentração de CO2 mais elevada que para níveis de irradiância
mais baixos (Salisbury & Ross, 1992).
Figura 60: Taxa de fotossíntese líquida em Beta vulgaris em função da concentração de CO2, e para
três níveis de irradiância. A linha a tracejado representa a concentração de CO2 atmosférico quando se realizou a experiência. As temperaturas foliares variaram entre os 21 e os 24 ºC. Retirado de Salisbury e Ross (1992), figura 12.9, página 260
Nas plantas em C4 a fotossíntese satura para concentrações de CO2 perto
dos 400 mol mol-1, o que é pouco mais que a concentração atmosférica normal.
A concentração em CO2 na atmosfera para a qual o CO2 fixo na fotossíntese
iguala o CO2 libertado na respiração e fotorrespiração é chamada - ponto de
compensação para o CO2 (). Na figura 61 podemos ver que o ponto de
compensação para a espécie em C4, o milho (Zea mays) é muito baixo, perto de
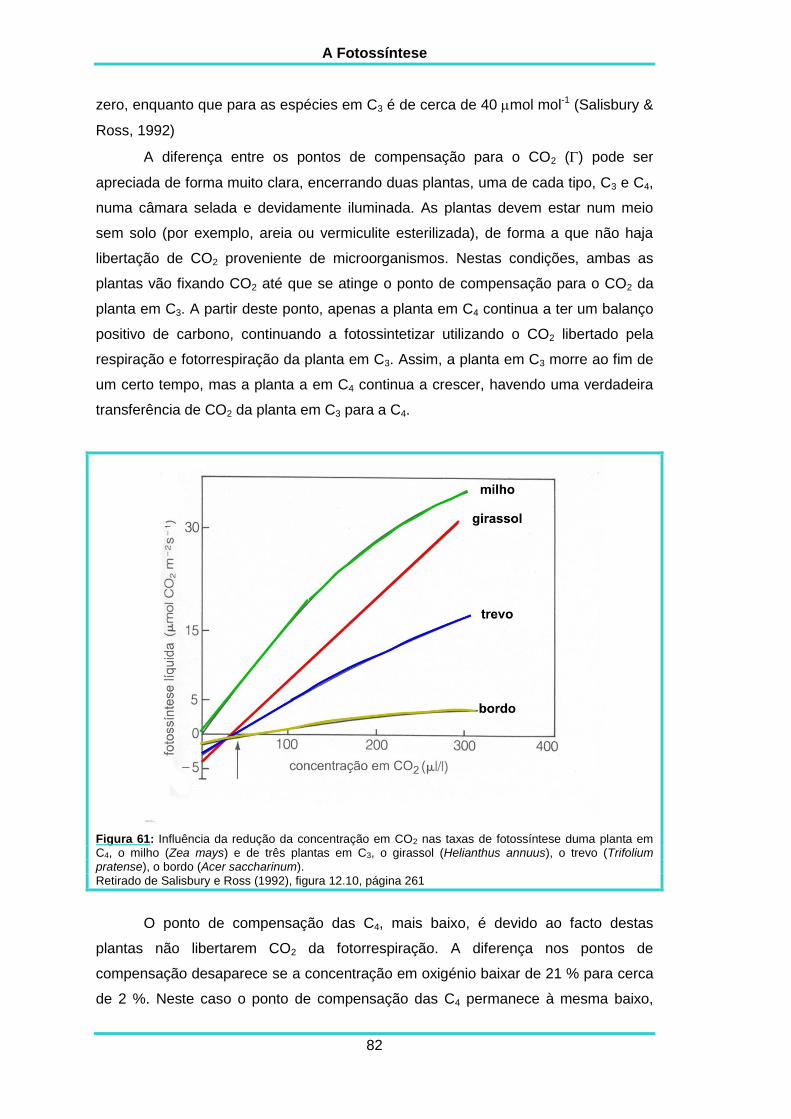
A Fotossíntese
82
zero, enquanto que para as espécies em C3 é de cerca de 40 mol mol-1 (Salisbury &
Ross, 1992)
A diferença entre os pontos de compensação para o CO2 () pode ser
apreciada de forma muito clara, encerrando duas plantas, uma de cada tipo, C3 e C4,
numa câmara selada e devidamente iluminada. As plantas devem estar num meio
sem solo (por exemplo, areia ou vermiculite esterilizada), de forma a que não haja
libertação de CO2 proveniente de microorganismos. Nestas condições, ambas as
plantas vão fixando CO2 até que se atinge o ponto de compensação para o CO2 da
planta em C3. A partir deste ponto, apenas a planta em C4 continua a ter um balanço
positivo de carbono, continuando a fotossintetizar utilizando o CO2 libertado pela
respiração e fotorrespiração da planta em C3. Assim, a planta em C3 morre ao fim de
um certo tempo, mas a planta a em C4 continua a crescer, havendo uma verdadeira
transferência de CO2 da planta em C3 para a C4.
Figura 61: Influência da redução da concentração em CO2 nas taxas de fotossíntese duma planta em C4, o milho (Zea mays) e de três plantas em C3, o girassol (Helianthus annuus), o trevo (Trifolium pratense), o bordo (Acer saccharinum).
Retirado de Salisbury e Ross (1992), figura 12.10, página 261
O ponto de compensação das C4, mais baixo, é devido ao facto destas
plantas não libertarem CO2 da fotorrespiração. A diferença nos pontos de
compensação desaparece se a concentração em oxigénio baixar de 21 % para cerca
de 2 %. Neste caso o ponto de compensação das C4 permanece à mesma baixo,

A Fotossíntese
83
mas o das C3 baixa para valores perto de zero, porque como há pouco O2 para
competir com o CO2 para a enzima rubisco e para o substrato, a fotorrespiração
torna-se negligível (Salisbury & Ross, 1992).
Durante a estação do crescimento, Primavera - Verão, as plantas em C3
apresentam muitas vezes uma taxa de fotossíntese abaixo do óptimo devido
essencialmente a uma concentração em CO2 baixa, sobretudo para folhas expostas a
irradiâncias elevadas. Por vezes brisas ligeiras permitem aumentar a fotossíntese,
porque permitem substituir o ar desprovido do CO2 da “boundary layer”. Se
pensarmos em termos de factor limitante para a fotossíntese, chegamos à conclusão
que, quer a luz, quer o CO2 podem ser factores limitantes, mas não para as mesmas
folhas duma mesma planta. Assim, as folhas superiores, mais expostas à radiação, o
CO2 pode ser o factor limitante, enquanto que as folhas mais interiores da copa
podem estar saturadas de CO2, mas necessitarem de mais luz.
Nas plantas de estufa por vezes o factor limitante pode ser o CO2, isto é
particularmente grave no Inverno, quando as estufas têm de estar fechadas por
causa do frio. Alguns agricultores minimizam este problema aumentando o teor em
CO2 das estufas a partir de tanques de CO2 pressurizado, ou através de outras fontes
de CO2, o que lhes permite aumentar a produção, sobretudo de plantas ornamentais.
No entanto, o nível de CO2 não deve exceder os 1000 a 1200 mol mol-1, porque
acima destas concentrações o CO2 torna-se tóxico ou causa o fecho estomático
(Salisbury & Ross, 1992).
Comparando o efeito da disponibilidade em CO2 nos três tipos de
metabolismo fotossintético podemos concluir o seguinte:
C3: a concentrações baixas de CO2 mostram um valor de fotossíntese líquida (PN)
inferior às plantas em C4; a partir de 1000 mol mol-1 estas diferenças não são tão evidentes; C4: a uma concentração nula de CO2 estas plantas praticamente não libertam CO2; CAM: apresentam um ponto de compensação para o CO2 muito baixo no escuro (Sebanek, 1992) Quanto ao ponto de compensação para o CO2, teremos:
C3: 45 a 60 mol mol-1;
C4: abaixo dos 5 mol mol-1;
CAM: variável, pode ser de cerca de 200 mol mol-1 (Sebanek, 1992)
5.10. TEMPERATURA:
A gama de temperaturas nas quais pode existir fotossíntese é muito grande.
Algumas algas azuis podem fotossintetizar a 70 ºC, enquanto que algumas coníferas
podem fotossintetizar a temperaturas por vezes inferiores a - 6 ºC.
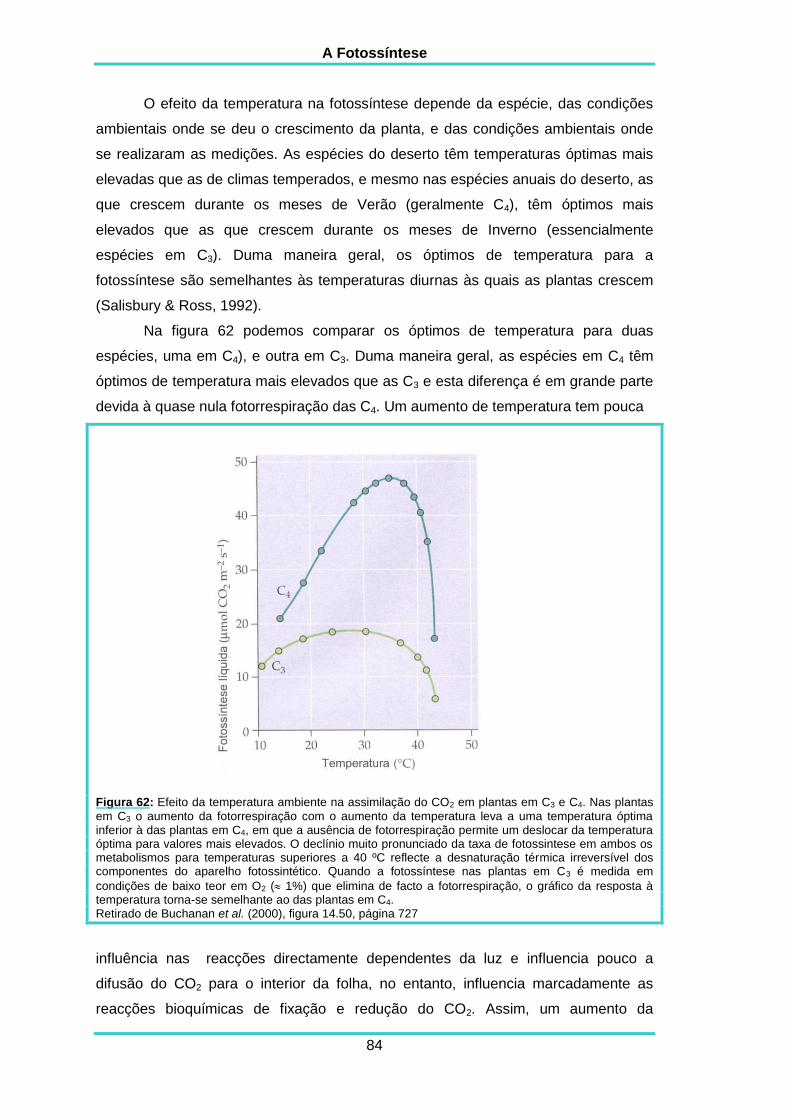
A Fotossíntese
84
O efeito da temperatura na fotossíntese depende da espécie, das condições
ambientais onde se deu o crescimento da planta, e das condições ambientais onde
se realizaram as medições. As espécies do deserto têm temperaturas óptimas mais
elevadas que as de climas temperados, e mesmo nas espécies anuais do deserto, as
que crescem durante os meses de Verão (geralmente C4), têm óptimos mais
elevados que as que crescem durante os meses de Inverno (essencialmente
espécies em C3). Duma maneira geral, os óptimos de temperatura para a
fotossíntese são semelhantes às temperaturas diurnas às quais as plantas crescem
(Salisbury & Ross, 1992).
Na figura 62 podemos comparar os óptimos de temperatura para duas
espécies, uma em C4), e outra em C3. Duma maneira geral, as espécies em C4 têm
óptimos de temperatura mais elevados que as C3 e esta diferença é em grande parte
devida à quase nula fotorrespiração das C4. Um aumento de temperatura tem pouca
Figura 62: Efeito da temperatura ambiente na assimilação do CO2 em plantas em C3 e C4. Nas plantas
em C3 o aumento da fotorrespiração com o aumento da temperatura leva a uma temperatura óptima inferior à das plantas em C4, em que a ausência de fotorrespiração permite um deslocar da temperatura óptima para valores mais elevados. O declínio muito pronunciado da taxa de fotossintese em ambos os metabolismos para temperaturas superiores a 40 ºC reflecte a desnaturação térmica irreversível dos componentes do aparelho fotossintético. Quando a fotossíntese nas plantas em C3 é medida em
condições de baixo teor em O2 ( 1%) que elimina de facto a fotorrespiração, o gráfico da resposta à temperatura torna-se semelhante ao das plantas em C4. Retirado de Buchanan et al. (2000), figura 14.50, página 727
influência nas reacções directamente dependentes da luz e influencia pouco a
difusão do CO2 para o interior da folha, no entanto, influencia marcadamente as
reacções bioquímicas de fixação e redução do CO2. Assim, um aumento da

A Fotossíntese
85
temperatura geralmente aumenta a taxa de fotossíntese até que haja desnaturação
das enzimas e destruição dos fotossistemas. No entanto, a perda de CO2 por
respiração também aumenta com a temperatura, e isto é particularmente evidente
para a fotorrespiração, em grande parte devido ao aumento da solubilidade do O2
quando comparado com o CO2. Devido à competição pelo O2, a fixação líquida de
CO2 nas plantas em C3 não aumenta tanto com a temperatura como seria de esperar.
O efeito promotor dum aumento da temperatura é praticamente contrabalançado pelo
aumento da respiração e fotorrespiração, e assim, a curva de resposta da
fotossíntese à temperatura, nas plantas em C3, aparece larga e achatada entre os 15
e os 30 ºC. Uma vez que nas plantas em C4 a fotorrespiração tem pouca importância
estas apresentam um óptimo de temperatura na ordem dos 30 a 40 ºC (Salisbury &
Ross, 1992).
Podemos resumir o efeito da temperatura nos três tipos de metabolismo
fotossintético do seguinte modo:
C3: a temperatura mínima é da ordem dos 0 ºC, e nalgumas espécies pode ocorrer fotossíntese mesmo a -6 ºC; há um óptimo pouco definido entre os 15 e os 25 ºC e um máximo de temperatura perto dos 30 ºC. C4: a fotossíntese líquida diminui muito para temperaturas abaixo dos 10 ºC, e o óptimo de temperatura varia com as espécies entre os 25 e os 40 ºC. CAM: o óptimo de temperatura é de cerca de 40 ºC (Sebanek, 1992).
5.11. SENSIBILIDADE À CONCENTRAÇÃO EM O2:
Otto WARBURG, um bioquímico alemão de grande importância que se
dedicou em grande parte ao estudo da fotossíntese em algas, notou em 1920 que a
fotossíntese é inibida pelo O2. Esta inibição ocorre em todas as espécies em C3
estudadas até hoje, e é designada por efeito Warburg. A figura 63 ilustra este efeito
para folhas de feijão (Phaseolus vulgaris) expostas a duas concentrações diferentes
de CO2, uma é ligeiramente inferior à concentração atmosférica normal, e outra é
bastante inferior. É importante notar que mesmo uma concentração de O2 perto do
normal, isto é, 21 %, é inibitória da fotossíntese quando comparada com uma
concentração de zero, para ambos os níveis de CO2. Além disso, a inibição pelo O2 é
maior para concentrações menores de CO2. Este efeito foi mais tarde relacionado
com o processo da fotorrespiração que já vimos anteriormente (Salisbury & Ross,
1992).
Podemos sintetizar o efeito da passagem duma atmosfera de 1 % para 21 %
O2 na fotossíntese líquida (PN) dos vários tipos de metabolismo do seguinte modo:
C3: PN diminui; C4: PN não se altera;

A Fotossíntese
86
CAM: PN praticamente não se altera (Sebanek, 1992).
5.12. FOTORRESPIRAÇÃO:
C3: pode atingir cerca de 1/3 da fotossíntese ilíquida (total); C4: é muito baixa ou mesmo inexistente; CAM: é muito baixa (Sebanek, 1992).
A ausência de fotorrespiração nas plantas em C4 deve-se, por um lado, ao
facto da enzima responsável pela primeira carboxilação, no mesófilo, a
fosfoenolpiruvato carboxilase (PEPcase), não ter afinidade para o oxigénio. Por outro
lado, nas células onde existe rubisco, ou seja, nas células da bainha dos feixes a
concentração é muito elevada devido à descarboxilação do ácido com quatro átomos
de carbono proveniente do mesófilo (malato ou aspartato), o que aumenta a relação
CO2 / O2, evitando a competição entre o CO2 e o O2 pela rubisco e pelo substrato.
Mesmo o CO2 proveniente da respiração mitocondrial para sair para o exterior tem de
passar pelas células do mesófilo, e aí a PEPcase que tem uma grande afinidade
para o CO2 fixa-o antes deste chegar à câmara estomática.
5.13. VALORES MÁXIMOS PARA A FOTOSSÍNTESE LÍQUIDA:
Valores obtidos para uma irradiância perto da saturação e uma concentração
de CO2 perto da atmosférica:
C3: 15 a 30 mol (CO2) m-2 s-1;
C4: 35 a 45 mol (CO2) m-2 s-1;
CAM: 1 a 5 mol (CO2) m-2 s-1 (Sebanek, 1992)
5.14. TAXA DE CRESCIMENTO MÁXIMO:
C3: 0.5 a 2.0 g (matéria seca) dm-2 dia-1; C4: 4 a 5 g (matéria seca) dm-2 dia-1; CAM : 0.012 a 0.020 g (matéria seca) dm-2 dia-1 (Sebanek, 1992)
5.15. PRODUÇÃO ANUAL DE MATÉRIA SECA:
C3: 22.0 3.3 t ha-1 ano-1;
C4: 38.6 16.9 t ha-1 ano-1; CAM: existe uma grande variabilidade nos dados disponíveis (Sebanek, 1992)
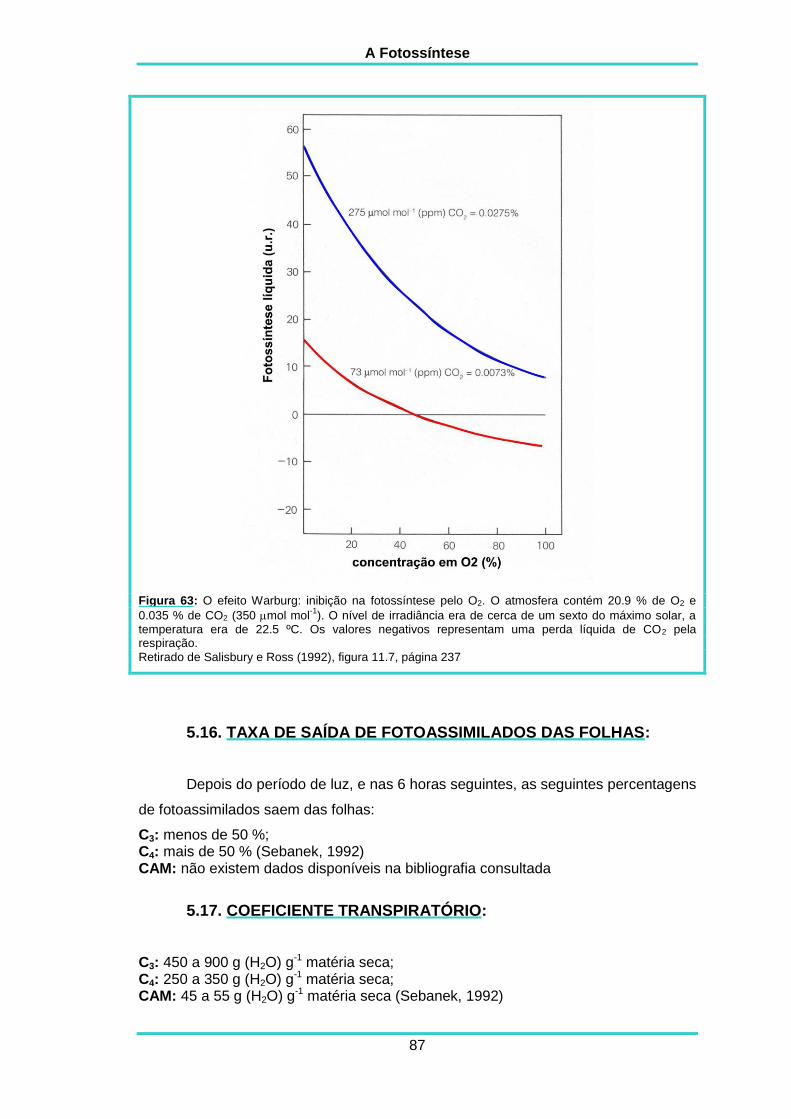
A Fotossíntese
87
Figura 63: O efeito Warburg: inibição na fotossíntese pelo O2. O atmosfera contém 20.9 % de O2 e
0.035 % de CO2 (350 mol mol-1
). O nível de irradiância era de cerca de um sexto do máximo solar, a temperatura era de 22.5 ºC. Os valores negativos representam uma perda líquida de CO2 pela respiração.
Retirado de Salisbury e Ross (1992), figura 11.7, página 237
5.16. TAXA DE SAÍDA DE FOTOASSIMILADOS DAS FOLHAS:
Depois do período de luz, e nas 6 horas seguintes, as seguintes percentagens
de fotoassimilados saem das folhas:
C3: menos de 50 %; C4: mais de 50 % (Sebanek, 1992) CAM: não existem dados disponíveis na bibliografia consultada
5.17. COEFICIENTE TRANSPIRATÓRIO:
C3: 450 a 900 g (H2O) g-1 matéria seca; C4: 250 a 350 g (H2O) g-1 matéria seca; CAM: 45 a 55 g (H2O) g-1 matéria seca (Sebanek, 1992)
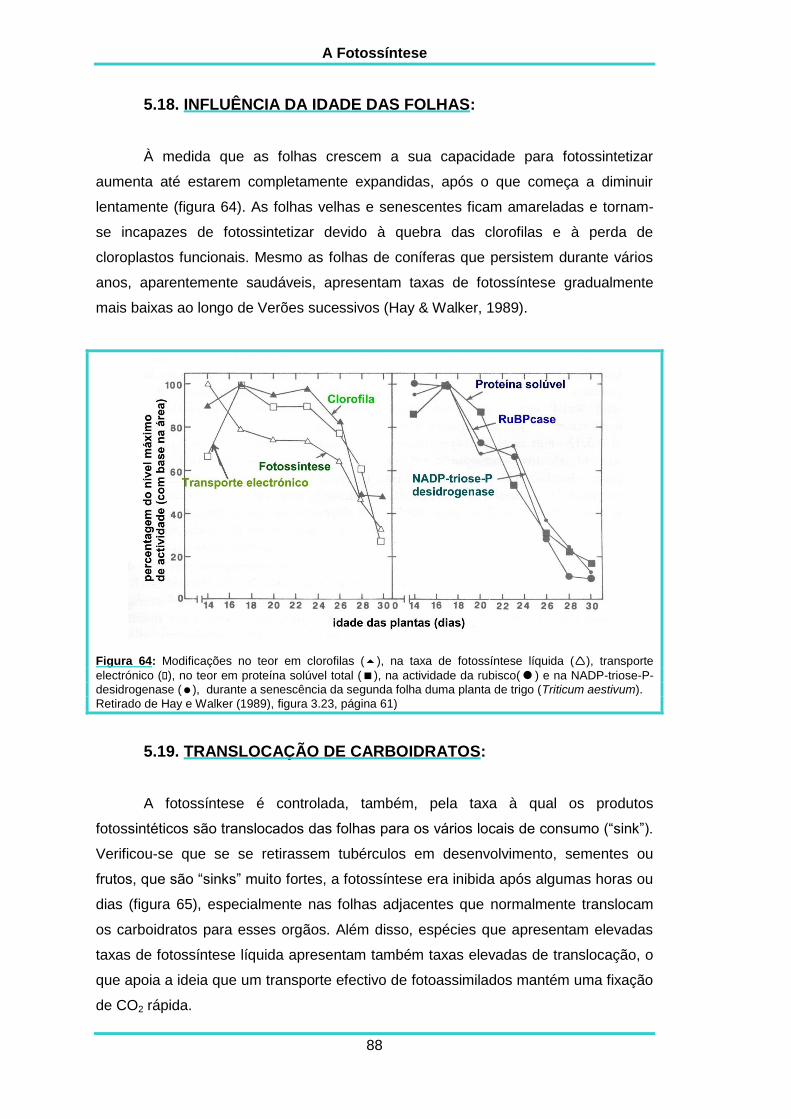
A Fotossíntese
88
5.18. INFLUÊNCIA DA IDADE DAS FOLHAS:
À medida que as folhas crescem a sua capacidade para fotossintetizar
aumenta até estarem completamente expandidas, após o que começa a diminuir
lentamente (figura 64). As folhas velhas e senescentes ficam amareladas e tornam-
se incapazes de fotossintetizar devido à quebra das clorofilas e à perda de
cloroplastos funcionais. Mesmo as folhas de coníferas que persistem durante vários
anos, aparentemente saudáveis, apresentam taxas de fotossíntese gradualmente
mais baixas ao longo de Verões sucessivos (Hay & Walker, 1989).
Figura 64: Modificações no teor em clorofilas (), na taxa de fotossíntese líquida (), transporte
electrónico (), no teor em proteína solúvel total (), na actividade da rubisco() e na NADP-triose-P-
desidrogenase (), durante a senescência da segunda folha duma planta de trigo (Triticum aestivum).
Retirado de Hay e Walker (1989), figura 3.23, página 61)
5.19. TRANSLOCAÇÃO DE CARBOIDRATOS:
A fotossíntese é controlada, também, pela taxa à qual os produtos
fotossintéticos são translocados das folhas para os vários locais de consumo (“sink”).
Verificou-se que se se retirassem tubérculos em desenvolvimento, sementes ou
frutos, que são “sinks” muito fortes, a fotossíntese era inibida após algumas horas ou
dias (figura 65), especialmente nas folhas adjacentes que normalmente translocam
os carboidratos para esses orgãos. Além disso, espécies que apresentam elevadas
taxas de fotossíntese líquida apresentam também taxas elevadas de translocação, o
que apoia a ideia que um transporte efectivo de fotoassimilados mantém uma fixação
de CO2 rápida.
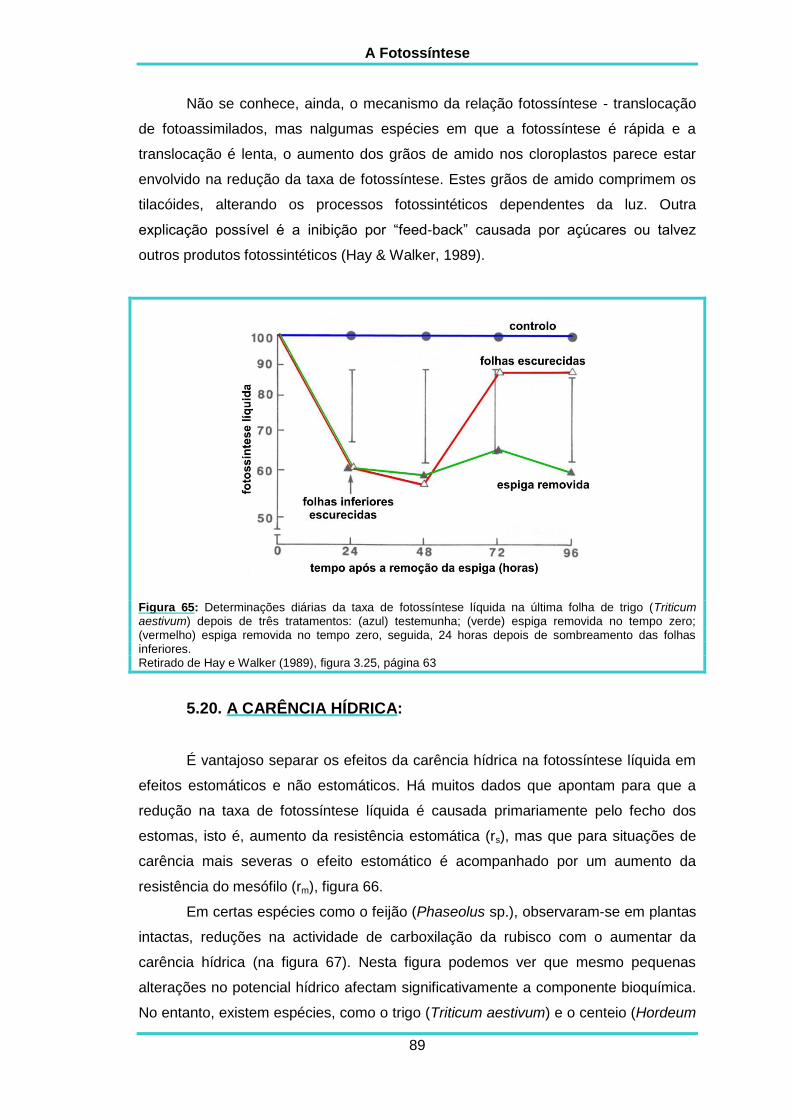
A Fotossíntese
89
Não se conhece, ainda, o mecanismo da relação fotossíntese - translocação
de fotoassimilados, mas nalgumas espécies em que a fotossíntese é rápida e a
translocação é lenta, o aumento dos grãos de amido nos cloroplastos parece estar
envolvido na redução da taxa de fotossíntese. Estes grãos de amido comprimem os
tilacóides, alterando os processos fotossintéticos dependentes da luz. Outra
explicação possível é a inibição por “feed-back” causada por açúcares ou talvez
outros produtos fotossintéticos (Hay & Walker, 1989).
Figura 65: Determinações diárias da taxa de fotossíntese líquida na última folha de trigo (Triticum aestivum) depois de três tratamentos: (azul) testemunha; (verde) espiga removida no tempo zero; (vermelho) espiga removida no tempo zero, seguida, 24 horas depois de sombreamento das folhas inferiores. Retirado de Hay e Walker (1989), figura 3.25, página 63
5.20. A CARÊNCIA HÍDRICA:
É vantajoso separar os efeitos da carência hídrica na fotossíntese líquida em
efeitos estomáticos e não estomáticos. Há muitos dados que apontam para que a
redução na taxa de fotossíntese líquida é causada primariamente pelo fecho dos
estomas, isto é, aumento da resistência estomática (rs), mas que para situações de
carência mais severas o efeito estomático é acompanhado por um aumento da
resistência do mesófilo (rm), figura 66.
Em certas espécies como o feijão (Phaseolus sp.), observaram-se em plantas
intactas, reduções na actividade de carboxilação da rubisco com o aumentar da
carência hídrica (na figura 67). Nesta figura podemos ver que mesmo pequenas
alterações no potencial hídrico afectam significativamente a componente bioquímica.
No entanto, existem espécies, como o trigo (Triticum aestivum) e o centeio (Hordeum
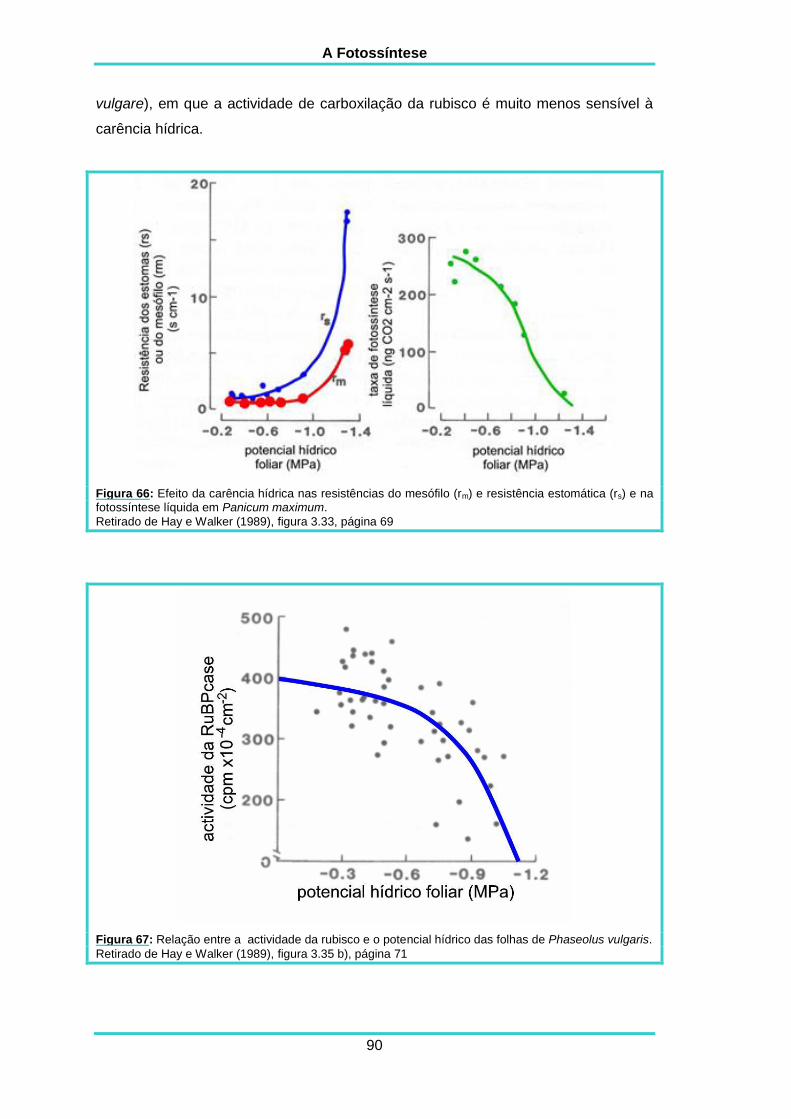
A Fotossíntese
90
vulgare), em que a actividade de carboxilação da rubisco é muito menos sensível à
carência hídrica.
Figura 66: Efeito da carência hídrica nas resistências do mesófilo (rm) e resistência estomática (rs) e na fotossíntese líquida em Panicum maximum.
Retirado de Hay e Walker (1989), figura 3.33, página 69
Figura 67: Relação entre a actividade da rubisco e o potencial hídrico das folhas de Phaseolus vulgaris.
Retirado de Hay e Walker (1989), figura 3.35 b), página 71

A Fotossíntese
91
BIBLIOGRAFIA:
BUCHANAN, B.; W. GRUISSEM & R. JONES (2000) - “Biochemistry and Molecular Biology of Plants” - American Society of Plant Physiologists EWY, R.G. & R.A. DILLEY (2000) – “Distinguishing between luminal localized proton buffereing pools in thylakoid membranes” Plant Physiology 122:583-595 FAHN, A. (1974) – “Plant Anatomy” 2nd Ed. Pergamon Press, Oxford FITTER, A.H. & R.K.M. HAY (2002) – “Environmental Physiology of Plants” – 3rd Ed. Academic Press, London GOODWIN, T.W. & E.I. MERCER (1983) – “Introduction to Plant Biochemistry” - 2nd
Ed. Pergamon Press, Oxford GROTH, G. & H. SHOTMANN (1999) – “New Results about structure function and regulation of the chloroplast ATPsynthase(CF0 – CF1)” Physiologia Plantarum 106:142-148 HALL, D.O. & K.K. RAO (1981) – “Photosynthesis” 3rd Ed. – Edward Arnold Pub.,
U.K. HAY, R.K.K. & A.J. WALKER (1989) – “An introduction to the physiology of crop
yield” Longman Scientific and Technical, New York LAWLOR, D.W. (1987) – “Photosynthesis: metabolism, control and physiology” Longman Scientific and Tachnical, New York NOBEL, P.S. (1991) – “Physicochemical and Environmental Plant Physiology” Academic Press, London SALISBURY, F.D. & C.W.ROSS (1985 and1992) – “Plant Physiology” 3rd and 4th Eds. Wadsworth Pub. Comp. California SEBANEK, J. (1992) – “Plant Physiology” – Elsevier, Amsterdam TAIZ, L. & E. ZEIGER (1998) – “Plant Physiology” 2nd Ed. Sinauer Ass. Inc., Pub,
USA TEIXEIRA, A.R.N. & C.P.P. RICARDO (1983) – “Fotossíntese” Didáctica Ed., Lisboa