1. introduÇÃo 1.1. o sistema nervoso É capaz de …livros01.livrosgratis.com.br/cp068937.pdf ·...
TRANSCRIPT

11
1. INTRODUÇÃO
1.1. O SISTEMA NERVOSO É CAPAZ DE REAGIR À LESÃO
O sistema nervoso periférico (SNP) permite o crescimento de axônios
após lesão por longas distâncias, restabelecendo as conexões com seus alvos.
No entanto, o sistema nervoso central (SNC) apresenta um crescimento axonal
que geralmente se limita a poucos milímetros e assim, não restaura suas
conexões (AGUAYO et al., 1981; NICHOLLS et al., 2001). Vários estudos
foram realizados para compreender as diferenças entre os mecanismos de
regeneração do SNC e SNP.
Na década de 80, Aguayo e colaboradores realizaram uma série de
experimentos transplantando células da glia do SNC em axônios periféricos
lesados. Estes verificaram que o implante bloqueava o brotamento neuronal.
Realizaram o processo inverso, enxertando células gliais do SNP na região
medular lesada e verificaram que os axônios lesados cresciam nas regiões
onde havia enxerto. Porém o crescimento cessava quando entravam em
contato com o tecido nervoso central. Isto levou a concluir que a regeneração
do sistema nervoso tem maior relação com o substrato que o cerca do que com
as propriedades inerentes aos tecidos nervoso central e periférico (AGUAYO et
al., 1981). Anos mais tarde foi observado que células de Schwann poderiam
remielinizar axônios após uma lesão medular induzida fotoquimicamente em
ratos (BUNGE et al., 1994).
Keirstead e colaboradores (1992, 1995), em estudos comparando o
desenvolvimento embrionário de pintos e a capacidade de regeneração após
lesão do SNC, demonstraram que durante o desenvolvimento embrionário,
antes da formação da mielina e de oligodendrócitos, o SNC possui grande
capacidade de regeneração. Após a mielinização, essa capacidade era
suprimida. Aparentemente haveria um período crítico de transição durante o
desenvolvimento embrionário, no qual o SNC passaria de um estado favorável
ao crescimento neuronal, para outro desfavorável e não permissivo a esse
crescimento. Este período foi observado em mamíferos por Nicholls (1999). Em
1985, Schwab e Thoenen propuseram a existência de fatores inibidores do
crescimento neuronal no SNC, provavelmente relacionados à mielina. Estes
fatores inibiam o crescimento neuronal mesmo na presença de fatores

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

12
neurotróficos. Dentre eles foram isolados os fator inibidor do crescimento de
prolongamentos de neurônio-35 (neurite inhibitor, NI-35), o NI-250 (Nogo) e a
glicoproteína associada à milelina (Myelin Associated Glycoprotein, MAG -
TATAGIBA et al., 1997; GRANDPRÉ et al., 2000; GAO et al., 2003).
Ramón y Cajal, desde 1928, já atribuía a falha da regeneração do SNC à
falta de fatores tróficos ou promotores do crescimento (apud NICHOLLS et al.,
2001). Na década de 40, Levi-Montalcini e colaboradores, investigando o
desenvolvimento neuronal, descobriram a existência de uma família de
proteínas essenciais à sobrevivência de classes particulares de neurônios,
presentes tanto no desenvolvimento embrionário quanto no indivíduo adulto
após lesão. Dentre estas proteínas, o fator de crescimento de nervos (nerve
growth factor, NGF), foi um dos primeiros identificados. Em 1976, eles
observaram que os axônios de neurônios dos gânglios sensoriais de pintos em
cultura, proliferavam quando colocados em contato com o NGF.
Adicionalmente, a cadeia simpática torácica de gânglios de camundongos que
receberam injeção de um anticorpo para NGF 5 dias após o nascimento, era
muito menor em tamanho que a cadeia dos animais controle, sugerindo que o
NGF atuaria auxiliando o crescimento e a sobrevivência do sistema nervoso.
Um segundo fator foi identificado em extratos do SNC, o fator neurotrófico
derivado do cérebro (brain-derived neurotrophic factor, BDNF), cuja purificação
e caracterização revelaram alto grau de semelhança com a estrutura do NGF.
O BDNF faz parte da família dos fatores de crescimento ou neurotrofinas (apud
NICHOLLS et al., 2001). Hoje, existem outras substâncias endógenas
reconhecidamente relacionadas ao desenvolvimento, crescimento e
sobrevivência do sistema nervoso, tais como as neurotrofinas -3, -4, -5, -6 e -7
(BOYD; GORDON, 2002), o fator neurotrófico ciliar, o fator neurotrófico
derivado da glia (TATAGIBA et al., 1997), aminoácidos excitatórios (glutamato,
por exemplo), e neurotransmissores, como a serotonina e o óxido nítrico (NO).
O papel individual destas substâncias nos processos pós-lesão é
caminho importante para se entender os processos envolvidos na regeneração
do SNC.

13
1.2. A LESÃO MEDULAR
Para o indivíduo com lesão medular, os problemas iniciais são o
rompimento dos tratos axonais desconectando o SNC com as estruturas alvo,
que progride com a degeneração, a lesão progressiva da microvasculatura e a
cascata de eventos bioquímicos que leva à morte celular. Estes processos são
especialmente importantes, pois contribuem para instalação de deficiências
permanentes. O estudo da lesão medular se focaliza na elucidação dos
processos fisiopatológicos, no desenvolvimento de terapêutica para interrupção
do processo secundário de lesão e reversão dos fatores que impedem o
crescimento axonal no SNC. Com este intuito as seguintes questões são
bastante freqüentes: Quais os eventos que disparam a degeneração axonal no
SNC e especificamente na medula espinal? Quais são os mecanismos
moleculares envolvidos na cascata de lesão secundária? Como os diversos
eventos se interrelacionam? Como seria possível parar a cascata de eventos
deletérios ativados pela lesão inicial? Através de procedimentos que minimizam
a lesão primária ou da modulação de moléculas críticas na lesão secundária?
E, estas intervenções levariam à recuperação funcional perdida?
A lesão da medula espinal é um problema clínico sério que impõe
enormes responsabilidades e custos à sociedade, pois atinge principalmente
uma população jovem, no auge de sua produtividade. O Brasil não possui
dados epidemiológicos nacionais a respeito da lesão medular, apesar de
existirem estudos isolados em centros de atendimentos regionais e municipais.
Os dados mais utilizados na literatura são do National Spinal Cord Injury
Statistical Center (2006). Este estudo anual estima que a lesão medular ocorra
principalmente em homens (81%) entre 16-30 anos de idade. A paraplegia
acomete 48% desses pacientes e as lesões torácicas representam 34,4%
desses casos. E as principais causas são acidentes automobilísticos, quedas e
ferimentos por armas de fogo.
Até o momento não existem terapias disponíveis para a cura das vítimas
de lesão medular. Conseqüentemente é necessário o desenvolvimento de
novas estratégias terapêuticas para aumentar as possibilidades de
regeneração dos axônios medulares lesionados, melhorando a função
sensorial e motora e, portanto a qualidade de vida dos pacientes com lesão
medular (SHARMA, 2005).

14
Vários métodos são utilizados para mimetizar a lesão medular incluindo
clips para aneurisma, cateteres com balões, contusão por queda de peso,
placas compressivas, transecção medular, cabos circunferenciais e
estreitamento do canal vertebral, dentre outros (RIVLIN; TATOR, 1977; KHAN;
GRIEBEL, 1983; ALLEN, 1911 apud NYSTROM; BERGLUND, 1988;
DELAMARTER; SHERMAN; CARR, 1995; DIMAR et al., 1999; CARLSON et
al., 2003a,b). Estes métodos são testados em diferentes animais como ratos,
cães, coelhos, gatos, camundongos e macacos (NYSTROM; BERGLUND,
1988; WRATHALL, 1992; DELAMARTER; SHERMAN; CARR, 1995; BEATTIE
et al., 1997; DIMAR et al., 1999; KWON; TETZLAFF, 2001; CARLSON et al.,
2003a,b; SOUZA et al., 2006). Estes estudos são de fundamental importância,
pois fornecem informações essenciais para o entendimento da fisiopatologia da
lesão medular e o desenvolvimento de estratégias terapêuticas.
A causa primária da lesão medular é freqüentemente o deslocamento ou
fratura vertebral que pode causar compressão ou secção medular (TATOR;
FEHLINGS, 1991; DELAMARTER et al., 1995; ROSENBERG et al., 1997).
Como conseqüência imediata à lesão mecânica segue-se uma cascata de
eventos não restritos ao sítio da lesão (epicentro) que evoluem ao longo de
horas e dias, expandindo a lesão vertical e horizontalmente (TATOR;
FEHLINGS, 1991; SCHWAB et al., 2006; YANG; KIM; LEE, 2007). A lesão
medular resultará no acentuado rompimento e reorganização de vias neurais
ascendentes e descendentes, criando modificações substanciais nos aferentes
primários, interneurônios e motoneurônios, influenciando dramaticamente a
interação sensório-motora (FRIGON; ROSSIGNOL, 2006). Com a ruptura ou
contusão dos axônios ocorre o desenvolvimento de hemorragia, isquemia e
edema. A severidade e extensão da lesão secundária dependem da magnitude
do insulto inicial e de um vasto número de fatores que tem influência sobre o
fluxo sanguíneo, reação celular e do microambiente medular (TATOR;
FEHLINGS, 1991; SHARMA et al.,1996; AMAR; LEVY, 1999). No centro da
lesão formam-se cavitações inicialmente preenchidas por fluído e circundadas
por tecido cicatricial (BEATTIE et al., 1997; SCHWAB et al., 2006). Este
consiste na cicatriz glial formada por astrócitos reativos, micróglia e
fibroblastos. Ainda há a cicatriz fibrosa, formada por componentes
extracelulares, como depósitos de matriz extracelular (FAWCETT; ASHER,

15
1999). No SNC, este tecido cicatricial é capaz de produzir moléculas que
inibem o crescimento axonal e de servir como obstáculo físico ao crescimento
dos axônios (LU; WAITE, 1999).
Após o trauma inicial a medula espinal está sujeita a uma série de
eventos neurotóxicos, como influxo de íons, alterações no fluxo sanguíneo
local, e processos inflamatórios que contribuem para a evolução da lesão
neurológica. O evento mais imediato da cascata de lesão secundária é a
indução mecânica da despolarização e a conseqüente abertura de canais
iônicos voltagem-dependentes (canais de cálcio, sódio e potássio). Isto faz com
que haja liberação massiva de neurotransmissores no espaço extracelular, o
que desencadeia a cascata de reações denominada lesão excitotóxica.
A liberação destes neurotransmissores excitatórios (em especial o
glutamato) causa a abertura de canais iônicos operados por receptores
dependentes de ligantes, como os receptores N-metil-D-aspartato (NMDA -
SCHWAB et al., 2006). A resultante mais importante destes eventos é o
acúmulo de cálcio intracelular, que inicia uma série de efeitos deletérios. Dentre
estes efeitos estão a ativação das sintases do óxido nítrico e a disfunção
mitocondrial, que leva à falha do metabolismo energético aeróbico e acúmulo
de lactato (HALL; SPRINGER, 2004). Uma das conseqüências destes dois
eventos é a formação de espécies oxigênio reativas como o ânion peróxido
nitrito (ONOO-). As espécies oxigênio-reativas agem como moléculas
sinalizadoras que iniciam a progressão da inflamação pós-traumática e a
apoptose (morte celular programada - HALL; SPRINGER, 2004; SCHWAB et
al., 2006). Embora o peróxido nitrito possa disparar diferentes mecanismos de
dano celular, a peroxidação de lipídios da membrana celular parece ser o mais
importante deles. A peroxidação de lipídios ocorre em neurônios e vasos
sanguíneos, causando prejuízos diretos sobre a integridade e função da
membrana axonal, danificando a microvasculatura o que causa isquemia e,
indiretamente, contribui para a acentuação da lesão secundária (AMAR; LEVY,
1999; HALL; BRAUGHLER, 1993 apud HALL; SPRINGER, 2004).
Concomitantemente a essa cascata de eventos iniciada com a abertura
dos canais iônicos, o trauma inicial causa dano ao endotélio vascular da
medula espinal e conseqüente redução ou interrupção do fluxo sanguíneo
local. O dano vascular é o principal responsável pela necrose hemorrágica que

16
ocorre após a lesão medular. Inicialmente ocorre espasmo de artérias e veias,
seguida de agregação plaquetária e de fibrina, levando à formação de um
trombo. Este pode causar oclusão vascular, estase venosa, distensão, perda
da auto-regulação vascular, ruptura de pequenos vasos e conseqüente
hemorragia petequial (SONNTAG; DOUGLAS, 1992; AMAR; LEVY, 1999).
Com a perda de regulação vascular, o fluxo sanguíneo medular sofre
alterações hemodinâmicas que podem levar à hipotensão e hipóxia, que
compõem o quadro de isquemia focal (AMAR; LEVY, 1999). A perda da
microcirculação sanguínea se estende consideravelmente nas direções
proximal e distal do sítio de lesão (TATOR; FEHLINGS, 1991).
1.3. DESENVOLVIMENTO DE TERAPÊUTICA PARA A LESÃO MEDULAR
Os achados fisiopatológicos permitem o desenvolvimento de um grande
repertório de abordagens terapêuticas, cada uma voltada para uma fase da
recuperação da lesão. Diante de um trauma raquimedular com fratura e
deslocamento ósseo, as estratégias de tratamento têm objetivos diferentes ao
longo do tempo, de acordo com as características de cada caso. No que tange
a estabilização e a consolidação óssea, existe uma concordância sobre os
benefícios da imobilização e redução do deslocamento/fratura com tração
(WAGNER; CHEHRAZI, 1982; DUH et al., 1994). Todavia, frente à compressão
medular sem transecção completa, outras questões se tornam importantes, tais
como: quão rapidamente deve ser realizada a redução? Diante de uma
redução não invasiva falha, deve ser realizada ou não uma redução cirúrgica?
Esta intervenção melhoraria o prognóstico funcional? (WAGNER; CHEHRAZI,
1982).
No intuito de responder estas questões, muitos pesquisadores
investigaram os efeitos da descompressão cirúrgica em diferentes modelos
animais e métodos de lesão. Nystrom e Berglund (1988) observaram que o
prejuízo neurológico era diretamente proporcional à força e duração da
compressão aplicada sobre a medula da região torácica média de ratos.
Delamarter, Sherman e Carr (1995) mostraram que a descompressão imediata
ou uma hora após a compressão medular por cabo circunferencial em cães era
capaz de melhorar a função motora e o controle de funções autonômicas, bem
como favorecer o retorno de respostas somatosensoriais a potenciais

17
evocados. Quando a descompressão foi realizada após períodos maiores (6,
24 horas ou 1 semana) esses efeitos não foram mais observados. Resultado
semelhante foi encontrado por Carlson et al. (2003). Contudo, neste estudo foi
utilizado um método de compressão por pistão e o período de estudo foi restrito
à 30 ou 180 minutos de compressão.
Com o intuito de analisar a influência do tempo até a descompressão
sobre a recuperação locomotora de ratos com lesão medular, Dimar e
colaboradores (1999) propuseram um novo método. Este método utiliza
espaçadores de diferentes tamanhos para provocar estreitamento do canal
vertebral e conseqüente lesão medular por compressão. Eles demonstraram
que a recuperação motora é inversamente proporcional ao tempo até a
descompressão.
A despeito dos vários resultados favoráveis ao uso da descompressão
medular cirúrgica para aliviar os efeitos deletérios da compressão medular em
animais, ainda não existe um consenso sobre os efeitos da descompressão, ou
qual a janela temporal para obter resultados positivos (DELAMARTER;
SHERMAN; CARR, 1995; DIMAR et al., 1999; BELANGER; LEVI, 2000;
CARLSON et al., 2003; LIM; TOW, 2007). Esta falta de consenso se intensifica
quando se considera os resultados obtidos em ensaios clínicos.
Num estudo em humanos, Chen e colaboradores (1998) mostraram que
uma intervenção cirúrgica como a laminectomia, para descompressão da
medula edematosa, teria efeito neuroprotetor se realizada num período inferior
a 6 horas. Intervenções realizadas em fase subaguda (24 – 72 horas pós-
lesão) não mostraram melhora. Outro estudo mostrou que pacientes com lesão
medular cervical submetidos imediatamente ao protocolo de descompressão
tiveram melhores resultados em sua avaliação de função motora e diminuíram
o período de cuidados intensivos e permanência no hospital, quando
comparados com aqueles submetidos mais tardiamente ao protocolo de
descompressão (PAPADOPOULOS et al., 2002). No entanto, Pollard e Apple
(2003), num estudo retrospectivo com 412 casos de lesão cervical traumática
incompleta não encontraram melhoras significativas nas funções neurológicas
após intervenção cirúrgica imediata.
Segundo Fehlings e Perrin (2006), para se obter resultados mais
conclusivos com relação aos benefícios da descompressão e o tempo para

18
intervenção em humanos, seriam necessários estudos multicentro,
prospectivos, bem delineados e com controles aleatórios, levando a um
protocolo padronizado. Apesar das dificuldades de realização deste tipo de
investigação, principalmente diante de conceitos éticos e logísticos, esta
afirmação visa encorajar a rigidez de critérios no desenho experimental de
testes clínicos, bem como, estimular a troca de informações entre os diferentes
centros de estudos e a realização de novos estudos experimentais nesta área.
Todavia, as opções de tratamento não estão limitadas ao controle da
lesão inicial por meio do tratamento cirúrgico. Grande parte das abordagens
terapêuticas se concentra nas conseqüências agudas e crônicas da lesão
medular e em terapias de reabilitação. Devido à complexidade da cascata da
reação bioquímica envolvida na patogenia da lesão medular, numerosas
abordagens farmacológicas têm sido desenvolvidas. Os principais objetivos
destas estratégias na promoção da regeneração após lesão medular são:
minimizar a progressão da lesão secundária (neuroproteção), favorecer a
condução axonal (neurorestauração) e fornecer ambiente permissivo ao
crescimento axonal (neuroregeneração - SCHWAB et al., 2006).
Intervenções farmacológicas são importantes para reduzir a lesão
secundária que acentua os prejuízos funcionais, danos celulares e teciduais
inicialmente disparados pela injúria mecânica (MALLEI et al., 2005). Dentre
essas intervenções a de maior impacto clínico até o momento é a aplicação do
glucocorticosteróide metilprednisolona (LIM; TOW, 2007). A droga mostrou
benefícios após lesão medular aguda, tanto em animais quanto em humanos,
agindo provavelmente como antiinflamatório e inibidor de peroxidação lipídica
(AMAR; LEVY, 1999; HALL; SPRINGER, 2004). O uso clínico da
metilprednisolona se consolidou após bons resultados em dois grandes ensaios
multicentro denominados National Acute Spinal Cord Injury Study (NASCIS) II e
III (AMAR; LEVY, 1999; BELANGER; LEVI, 2000). Também em fase inicial de
ensaios clínicos está o tratamento agudo com anticorpo contra o Nogo, um
conhecido fator inibidor do crescimento neuronal. Este tratamento já obteve
sucesso em testes pré-clínicos em camundongos, ratos e macacos, levando à
regeneração e recuperação funcional após lesão medular (BUCHLI et al.,
2007).

19
Outras estratégias em estudo envolvem o uso de fatores neurotróficos
(neurotrofinas, BDNF e fator de crescimento ligado à insulina – IGF),
antagonistas opióides (como dinorfina, hormônio liberador de tireotropina e
naloxone), gangliosídeos, purinas (como a inosina), antagonistas de receptores
de aminoácidos excitatórios endógenos (em especial antagonistas de
receptores NMDA), bloqueadores de canal de cálcio, hormônio de crescimento,
agentes varredores de radicais livres e antioxidantes (HOUWELING et al.,
1998; AMAR; LEVY, 1999; WADA et al., 1999; BELANGER; LEVI, 2000;
NOVIKOVA; NOVIKOV; KELLERTH, 2002; NYBERG; SHARMA, 2002;
CARLSON et al., 2003b; HALL; SPRINGER, 2004; LIU et al., 2006). Além
destas, existem as terapias de transplante de células (células de Schwann -
lemócitos, medula óssea, macrófagos ativados, precursores de
oligodendrócitos e células tronco), terapia oxigênio hiperbárica, terapias para
eliminação de inibidores do crescimento axonal; enxertos de nervo periférico,
tecido de medula espinal fetal e de material biocompatível livre de células
(AMAR; LEVY, 1999; LU; WAITE, 1999; KWON; TETZLAF, 2001; GELLER;
FAWCETT, 2002; REIER, 2004; SCHWAB et al., 2006).
Embora algumas terapias experimentais tenham exibido sucesso sobre
alguns obstáculos à regeneração agindo em diferentes pontos da cascata de
lesão, é evidente que uma única abordagem não seria suficiente para alcançar
a completa recuperação após lesão medular (NOVIKOVA; NOVIKOV;
KELLERTH, 2002). Além do trauma inicial e conseqüente lesão medular,
muitos fatores participam da lesão secundária incluindo neuropeptídios,
monoaminas, células imunes, modificações em concentrações de cátions,
aminoácidos e produtos da hidrólise de fosfolipídios como ácidos graxos
insaturados, eucosanóides e radicais livres, dentre eles, o NO (FADEN;
SALZMAN, 1992; YANG; KIM; LEE, 2007).
1.4. ÓXIDO NÍTRICO E A PLASTICIDADE NEURONAL
O NO é um radical livre, um gás que se difunde rapidamente pela
membrana celular. As duas primeiras ações fisiológicas descritas para o NO
foram a vasodilatação e inibição da agregação plaquetária (IGNARRO, 1996;
SNYDER, 1992). No entanto, no sistema nervoso ele atua como uma molécula
regulatória ou um neurotransmissor. Diferentemente da descrição clássica para

20
neurotransmissores, o NO não é estocado em vesículas e não é liberado por
exocitose. Sua ação não depende da ligação em receptores na membrana pós-
sináptica por que sendo como um gás se difunde de um neurônio para o outro
e também para células vizinhas através da bicamada fosfolipídica. Além disso,
após ação não é recaptado ou degradado, mas inativado pela reação com
outras moléculas e conseqüente formação de novos compostos. Uma vez
dentro das células, o NO pode ligar-se a diferentes alvos dentre eles a enzima
guanilato ciclase presente no citosol das células de diversos órgãos. O NO se
liga à porção heme da enzima e aumenta sua função de catalisar a conversão
de guanosina trifosfato (GTP) a guanilato monofosfato cíclico (GMPc). O GMPc
por sua vez promove o relaxamento de músculos lisos, inibição da agregação e
adesão plaquetária e bloqueio da adesão de células brancas do sangue nas
paredes vascular (POULOS; RAMAN; LI, 1998; PAGLIARO, 2003; VALLANCE,
2003).
O NO é produzido estequiometricamente com a citrulina por uma família
de enzimas, as sintases do óxido nítrico (NOS – BREDT, 2003), a partir da
arginina. O processo requer entre outros co-fatores, a nicotinamida adenina
dinucleotídeo ou NADPH (GRIFFITH; STUEHR, 1995; WU, 2000), cálcio,
calmodulina, tetrahidrobiopterina e oxigênio (DAWSON; DAWSON; SNYDER,
1992; GRIFFITH; STUEHR, 1995). A necessidade de vários co-fatores para
que ocorram as reações catalisadas pela NOS demonstra a alta regulação de
sua ação e reflete a importância da síntese de quantidades precisas de seus
produtos.
Existem três isoformas de NOS identificadas. A primeira, NOS neuronal
(NOSn), foi purificada no SNC de ratos; pouco mais tarde, outras duas
isoformas foram identificadas, a NOS endotelial (NOSe), presente em células
do endotélio vascular e a NOS induzida (NOSi), derivada de macrófagos
(BREDT; SNIDER, 1991; IGNARRO; JACOBS, 2000). Todas as isoformas
possuem sítios de ligação para co-fatores: NADPH, flavina adenina
dinucleotídeo (FAD), flavina mononucleotídeo (FMN), Ca2+/Calmodulina. As
NOS endotelial e neuronal são expressas constitutivamente, podem ser
controladas por meio de retroalimentação negativa pelo NO e produzem baixas
concentrações de NO (nanomolar – VALLANCE, 2003). A NOSi é induzida, por
exemplo após processos lesivos. Esta isoforma não está sujeita à

21
retroalimentação negativa pelo NO e o produz na faixa de concentração
micromolar (PAGLIARO, 2003).
Ao longo da última década, o papel do NO no organismo foi
extensivamente estudado nos sistemas imune, vascular e nervoso, tanto em
condições fisiológicas quanto em condições patológicas (MONCADA, 1992;
GARTHWAITE, 1995; WU, 2000). Em condições normais o NO desempenha
vários papéis importantes na regulação do fluxo sanguíneo cerebral, formação
da memória, combate a infecções e tumores, modulação da neurotransmissão
e da função neuroendócrina (MONCADA, 1992; WOLF, 1997; WU, 2000). O
NO ganhou a reputação de ser um mediador da sinalização de diversas
atividades biológicas, freqüentemente antagônicas. Durante as duas últimas
décadas houve muitos debates sobre os mecanismos desta dicotomia e se o
NO seria um agente benéfico ou deletério. Resultados contraditórios
levantaram a participação do NO em repostas fisiopatológicas (IGNARRO,
1996). Enquanto muitos estudos mostraram a implicação do óxido nitrico
endógeno como molécula tóxica, outras mostraram sua ação protetora. (WINK
et al., 1996; WINK; MITCHELL; 1998; GRISHAM; JOURD'HEUIL; WINK, 1999).
As ações do NO no SNC incluem controle da atividade de canais iônicos e
liberação de transmissores (FAGNI; BOCKAERT, 1996; MEFFERT et al.,
1996).
Várias evidências demonstram que a produção excessiva de NO no
sistema nervoso pode levar a toxicidade. Em condições patológicas o NO pode
estar envolvido em processos isquêmicos, no acidente vascular cerebral,
doenças degenerativas, como a doença de Parkinson (MONCADA, 1994),
lesão e morte neuronal (MONCADA, 1992; SNIDER, 1992; DAWSON, 1995;
BREDT; WU, 2000). Quando produzido excessivamente o NO pode se tornar
um fator indutor de neurotoxicidade (DAWSON; DAWSON, 1996). A atividade
da NOSn é primariamente regulada pelo aumento intracelular de cálcio, o qual
estimula a NOSn através de interação com calmodulina (BREDT; SNYDER,
1990). No cérebro, a biossíntese do NO pela ativação da NOSn é
predominantemente regulada pelo influxo de cálcio na sinapse, que ocorre por
meio da ligação do glutamato com seus receptores tipo NMDA. Quando este
mecanismo se torna exacerbado, por exemplo, diante de lesão do sistema
nervoso, pode ocorrer a formação excessiva de NO (LIPTON et al., 1996). Um

22
dos mecanismos de citotoxidade do NO está na sua rápida ligação com o
radical peróxido (O2-2), formando peróxido nitrito (ONOO-). Tanto o NO como o
peróxido-nitrito podem causar dano mitocondrial e do DNA, podendo levar à
morte celular. Em contraste, o NO pode se isomerizar para formar nitrato
(NO3-), uma substância inofensiva (VALLANCE, 2003; RADI, 2004; BISHOP;
ANDERSON, 2005).
O nitroprussiato de sódio, um doador de NO, retarda a recuperação de
reflexos polissinápticos após isquemia medular transitória (SUZUKI et al., 1995;
NEMOTO et al., 1997). Contudo, Wainwright e colaboradores (2007)
mostraram que outro doador de NO pode reduzir a lesão após isquemia
cerebral em ratos recém-nascidos, provavelmente agindo como um pivô na
regulação na perfusão sanguínea cerebral. Lipton e colaboradores (1993)
forneceram evidências de que o papel do NO depende do seu estado oxidativo.
O NO+ reage com proteínas levando à regulação fisiológica no SN, enquanto
NO- leva a efeitos neurotóxicos. Adicionalmente, pequenas quantidades de NO
estão mais relacionadas a efeitos fisiológicos, e a superprodução, com
neurotoxicidade (DAWSON et al., 1991). Além disso, o NO pode agir como
seqüestrador de radicais livres (PLUTA et al., 2001).
O crescente interesse tem se focado no papel do NO no sítio de lesão e
em áreas localizadas próximas ao epicentro de impacto e sua conexão com o
aumento da expressão de NOSn em neurônios (MARSALA, 2007). Na medula
espinal, a NOSn é predominantemente localizada nos neurônios sensoriais do
corno dorsal, na região ao redor do canal central e na coluna intermediolateral
(REUSS; REUSS, 2001). A estimulação transitória da produção de NO ocorre
imediatamente após a lesão, seguida por outras ondas 24 horas e 3 dias mais
tarde (NAKAHARA et al., 2002). Apesar da existência de evidências de que o
NO tenha papel importante no processo plástico neuronal, seu papel na
fisiopatologia da lesão ainda é controverso.
O estudo do sistema do NO tem fornecido novas perspectivas no
entendimento dos mecanismos de degeneração e regeneração de neurônios,
bem como no desenvolvimento de novos procedimentos terapêuticos em
pacientes que sofreram lesão de vias neurais, tanto do SNC, quanto do SNP.

23
1. OBJETIVOS
2.1. Objetivo Geral:
O objetivo geral deste estudo foi analisar o efeito da compressão da
medula espinal de ratos seguida de cirurgia para descompressão associada ou
não à injeção intratecal do doador de óxido nítrico, N-Ethyl-2-(1-ethyl-2-
hydroxy-2-nitrosohydrazino)ethanamine (NOC12).
2.2. Objetivos específicos:
1 – Analisar as funções sensorial e motora dos animais nos períodos
pré-operatório, 1, 5 e 7 dias após o procedimento experimental, por meio do
teste de retirada de cauda e da escala de avaliação locomotora BBB;
2 - Analisar as modificações no tecido medular após o estreitamento do
canal, somado ou não à descompressão cirúrgica e aplicação da droga. A
extensão da lesão e a porcentagem de tecido preservado foram estimadas
utilizando-se a coloração de hematoxilina-eosina 7 dias após a lesão;
3 - Examinar possíveis modificações na expressão da NOS após injeção
intratecal de doador de óxido nítrico exógeno nos grupos descompressão por
meio de imunohistoquímica para NOS neuronal 7 dias após a lesão.

24
3. MATERIAL
3.1. Sais, drogas e reagentes
Os produtos utilizados são descritos na lista abaixo. Aqueles que não
aparecem na lista estão descritos no corpo do texto, no item “4. Métodos”.
Todos os sais e reagentes utilizados neste estudo foram de qualidade para
análise (p.a.).
• 3, 3’ - Diaminobenzidina (DAB; 3,3’,4,4’ - Tetraaminobiphenyl;
tetrahydrochloride) - Sigma;
• ácido clorídrico (HCl) - Merck;
• álcool absoluto (C2H5OH) - Merck;
• alúmen de potássio (AIK(SO4)2 . 12H2O) - Sigma;
• cloreto de sódio (NaCl) - Merck;
• cloreto de sódio (0,9% - 10ml) - Samtec Biotecnologia;
• cloridrato de cetamina 10% - Ketamina Agener;
• diafanizador vegetal a base de D-limonene – Bio Optica Milano;
• eosina (amarela) Y – (C20H6Br4Na2O5) - Sigma;
• floxina B (C20H2Br4Cl4O5Na2) - Sigma;
• fosfato dissódico (Na2HPO4) - Merck;
• fosfato monossódico 1-Hidratado (NaH2PO4.H2O) - Merck;
• gelatina tipo A - Sigma;
• hematoxilina-cristais (C16H14O6) - Sigma;
• hidróxido de amônia (NH3) - Merck;
• meio de inclusão para microscopia – Entellan, Merck;
• N,N-dimetilformamida (C3H7NO) - Sigma;
• N-Ethyl-2-(1-ethyl-2-hydroxy-2-nitrosohydrazino)ethanamine – NOC12
Calbiochem;
• nitrobluetetrazolium (C40H30Cl2N10O6) - Sigma;
• paraformaldeído - Sigma;
• pentabiótico 1.200.000 UI– Fort Dodge;
• peróxido de hidrogênio (H2O2) - Sigma;
• reagentes A e B - Vectastain, ABC Kit, Vector Laboratories, Inc.;

25
• sabão Sigma CleanTM - Sigma;
• sacarose (C12H22O11) - Merck;
• soro albumina bovina (BSA) - Sigma;
• triton X-100 (t-octylphenoxypoly-ethoxyethanol) - Sigma;
• uretana 99% (H2NCO2C2H5) - Merck;
• xilasina 200mg (Dopaser) – Calier S.A.;
• xilol (C8H10) - Merck;
3.2. Equipamentos
3.2.1. Microscópio
Os cortes histológicos foram analisados em microscópio óptico da marca
Zeiss, modelo Axioskop 2 plus.
3.2.2. Analisador de imagens
Para a mensuração da densidade de células positivamente marcadas
nas regiões de interesse da medula espinal, coradas pela reação de
hematoxilina-eosina (HE) e imunocitoquímica para a proteína NOSn, as
lâminas foram analisadas em um microscópio Zeiss Axioskop 2 plus, o qual
possuía uma câmera de captura de imagem em tempo real da marca Zeiss,
modelo AxioCam HRc. A câmera era conectada a um sistema de edição e de
transferência de imagem. Este era conectado a um computador o qual possui
um monitor LCD Waytec 19’’ de alta resolução (1240x1240 dpi). O “software”
utilizado, para a análise das imagens, foi o “AxioVision v.4.2” (Zeiss).
3.2.3. Estereomicroscópio
Os procedimentos cirúrgicos bem como as fotografias do ato cirúrgico,
foram feitos por meio do auxílio de um estereomicroscópio Leica, modelo
M651, com as objetivas de 10X e 16X, com um câmera fotográfica Olympus,
modelo DP11, acoplada a ele.
3.2.4. Digitalizador de imagens
As imagens dos cortes histológicos foram digitalizadas, utilizando-se um
microscópio Leica, modelo DMRB, conectado a uma câmera fotográfica

26
colorida Olympus, modelo DP 11, a um monitor colorido de vídeo Sansung,
modelo SyncMaster 700b e a um microcomputador. As imagens foram
capturadas e processadas no aplicativo Nikon View 6.0 e Adobe Photoshop
CS2.
3.2.5. Balanças
Os animais foram pesados em uma balança da marca Gehaka, modelo
BG1000, com divisão de 0,01g, e as drogas em uma balança da marca
Scientech, modelo SA210, com divisão de 0,0001g.
3.2.6. Outros equipamentos
• Criostato marca Leica, modelo CM1850
• agitador eletrônico com aquecimento da marca Barnstead/Thermolyne,
modelo Nuova SP18425;
• agitador/incubadora orbital refrigerada da marca Amerex Instruments, Inc.,
modelo GyromaxTM 703R;
• banho-maria da marca Precision Scientific, modelo “shallow form shaking
bath”, nº de categoria: 66799;
• bidestilador e deionizador da marca Sheldon Manufacturing, Inc., modelo
FistreemTM III e sistema de estocagem de água destilada, modelo A56290-857;
• estufa da marca Sheldon Manufacturing Inc., modelo 1500E;
• freezer –20° C da marca Brastemp, modelo Clean 280;
• freezer -70°C da marca So-Low Environmental Equipment, modelo U85- 22;
• mesa agitadora da marca Tecnal, modelo TE – 141, orbital;
• refrigerador da marca Brastemp, modelo Clean 340.
4. MÉTODOS
4.1. Animais
Neste estudo foram utilizados ratos Wistar com peso corporal variando
entre 270-300g. Os animais foram trazidos do Biotério Central da Universidade
de São Paulo (USP), campus de Ribeirão Preto e alojados no Biotério da
Faculdade de Odontologia de Ribeirão Preto (FORP-USP) até ao término do

27
experimento. Foram mantidos em caixas contendo 4 animais com livre acesso
à água e dieta padrão do biotério, em sala com temperatura controlada (23±2°
C) e com um ciclo claro/escuro de 12 x 12 horas, sendo o início do período de
claro às 7:00 horas.
Previamente ao início do experimento, todos os animais foram pesados
e identificados por meio de um código de marcas em suas caudas. No total
foram utilizados 95 ratos durante a pesquisa. Todo esforço foi despendido para
minimizar o sofrimento dos animais. Os experimentos foram realizados de
acordo com o Comitê de Ética em Experimentação Animal da Universidade de
São Paulo (CETEA-USP). Protocolo de aprovação nº 054/2004.
4.2. Espaçador para estreitamento do canal vertebral em 35%
As dimensões do espaçador foram definidas a partir de medidas médias
do diâmetro ântero-posterior do canal vertebral de ratos de peso e idade
semelhantes. Este método permitiu determinar o tamanho do espaçador
necessário para produzir um grau preciso de estreitamento do canal vertebral
de ratos Wistar (DIMAR et al., 1999; modificado por SCHIAVETO DE SOUZA
et al., 2006). Os espaçadores foram manufaturados em policarbonato de modo
a diminuir o diâmetro do canal vertebral do rato em 35%
(3.6mmX2.2mmX0.78mm - comprimento X largura X espessura - Figura 1). Em
cada espaçador foi fixado um fio de sutura de seda 4.0 para facilitar sua
retirada 6h após sua inserção.
4.3. Cirurgia
Os ratos foram anestesiados com uma mistura de ketamina (100mg/Kg)
e xilazina (14mg/Kg) via intraperitoneal (i.p.). Em seguida foi realizada a
tricotomia da região dorsal torácica e uma incisão na região correspondente à
coluna torácica inferior. A musculatura foi então divulsionada, expondo as
vértebras T8 – T11, identificadas pela palpação das apófises espinhosas. Os
procedimentos foram realizados de acordo com o grupo cirúrgico, como segue
abaixo:
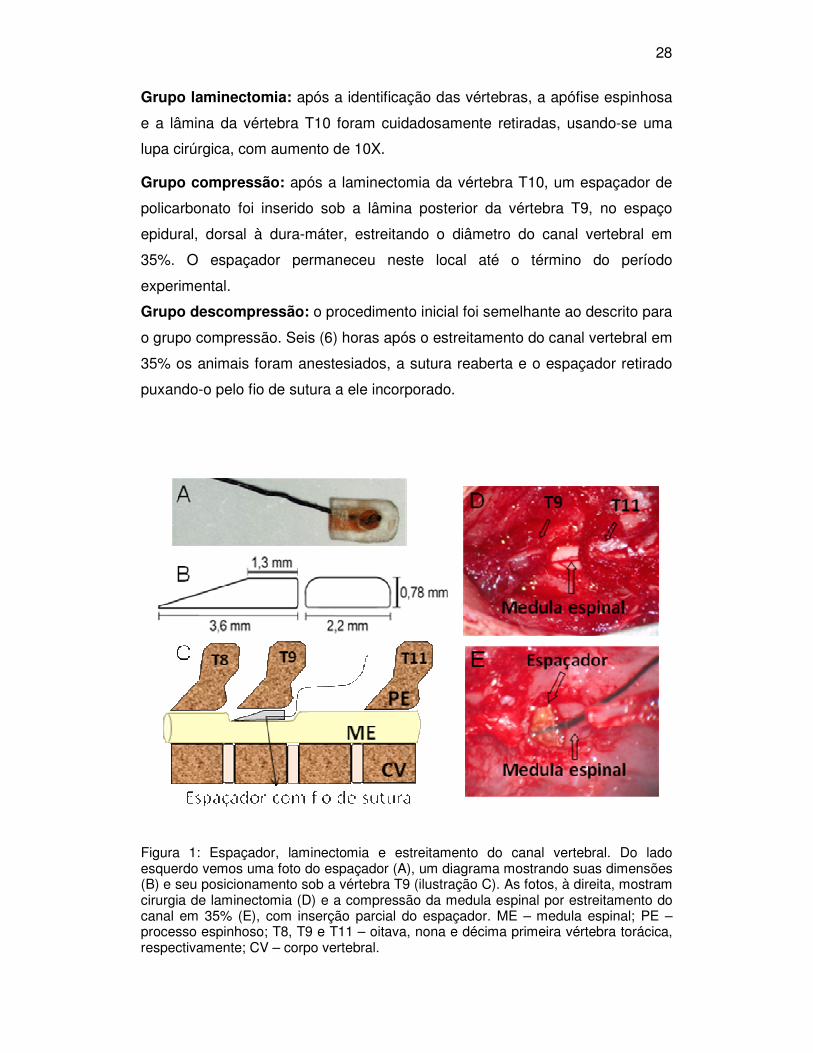
28
Grupo laminectomia: após a identificação das vértebras, a apófise espinhosa
e a lâmina da vértebra T10 foram cuidadosamente retiradas, usando-se uma
lupa cirúrgica, com aumento de 10X.
Grupo compressão: após a laminectomia da vértebra T10, um espaçador de
policarbonato foi inserido sob a lâmina posterior da vértebra T9, no espaço
epidural, dorsal à dura-máter, estreitando o diâmetro do canal vertebral em
35%. O espaçador permaneceu neste local até o término do período
experimental.
Grupo descompressão: o procedimento inicial foi semelhante ao descrito para
o grupo compressão. Seis (6) horas após o estreitamento do canal vertebral em
35% os animais foram anestesiados, a sutura reaberta e o espaçador retirado
puxando-o pelo fio de sutura a ele incorporado.
Figura 1: Espaçador, laminectomia e estreitamento do canal vertebral. Do lado esquerdo vemos uma foto do espaçador (A), um diagrama mostrando suas dimensões (B) e seu posicionamento sob a vértebra T9 (ilustração C). As fotos, à direita, mostram cirurgia de laminectomia (D) e a compressão da medula espinal por estreitamento do canal em 35% (E), com inserção parcial do espaçador. ME – medula espinal; PE – processo espinhoso; T8, T9 e T11 – oitava, nona e décima primeira vértebra torácica, respectivamente; CV – corpo vertebral.

29
Após cada cirurgia os músculos e a pele foram suturados
separadamente em camadas. Após o procedimento cirúrgico todos os animais
receberam aplicação de salina 0,9% (3ml) e analgésico (Flunixin Meglumine,
2,5mg/Kg) via subcutânea, e antibiótico profilático via intramuscular (dose única
de pentabiótico -240000 UI das penicilinas por kg e 100mg de estreptomicina e
diidroestreptomicina por kg, volume 0,1ml/100g). Em seguida, foram deixados
em caixas aquecidas para recuperação. Nos dias seguintes, os animais foram
manipulados diariamente com massagem abdominal, para facilitar a eliminação
de fezes; e esvaziamento de bexiga duas a três vezes por dia até que
recuperassem controle voluntário. Os animais eram inspecionados diariamente
à procura de sinais de autotomia e infecções.
4.4. Cateterização subaracnóide e injeção intratecal
Para proceder à injeção intratecal se fez necessária a cateterização do
espaço subaracnóide. Para tanto, os animais foram anestesiados e colocados
em decúbito ventral sobre um objeto cilíndrico para que a região lombar ficasse
hiperfletida. Foi realizada uma incisão na pele da região lombar, utilizando
como referência o ponto central de uma linha horizontal traçada entre as
espinhas ilíacas póstero-superiores do animal. Esta incisão permitiu a
visualização dos músculos e fáscias que serviram de referência anatômica para
localização do espaço intervertebral entre L5-L6. Neste ponto, foi introduzida
uma agulha Weiss (20 G) até o nível subaracnóideo, o que provoca um abalo
leve da cauda. Este foi considerado um sinal positivo de posicionamento da
agulha. Um pequeno pedaço de tubo de polietileno PE10 (Becton-Dickinson
and Company) foi inserido por dentro da agulha até projetar-se 1,5 cm dentro
do espaço subaracnóide, de modo a posicioná-lo próximo à intumescência
lombossacra. A agulha foi então cuidadosamente retirada e o cateter fixado à
musculatura mais externa. Para a injeção intratecal, foi adaptado a este cateter
um tubo PE20 previamente calibrado com uma seringa Hamilton de forma que
o volume administrado correspondesse ao deslocamento do volume conhecido
(SILVEIRA, 2006).
Foi injetado o doador de NO, NOC12 ou seu veículo. Este doador se
caracteriza por ter uma vida média de liberação de NO em PBS pH 7.4 de 327
minutos a 22°C e se manter relativamente estável em solução alcalina. Sendo

30
assim, para evitar sua liberação antes da administração, este foi diluído em
Tampão Tris 1M (pH 10.5 - veículo). Foram utilizadas duas concentrações de
NOC12 30 ou 300nM, ou veículo, injetado no espaço subaracnóide (injeção
intratecal) em dose única (volume de 5µl em 1 minuto), um dia após os
procedimentos cirúrgicos. O cateter foi removido dez minutos após o fim do
processo. Os animais tiveram a pele suturada e foram deixados em local
apropriado para recuperação da anestesia. Veículo e solução de NOC12 foram
preparados imediatamente antes da injeção.
4.5. Grupos Experimentais
Os animais foram aleatoriamente distribuídos nos seguintes grupos
experimentais:
1) laminectomia + veículo (n=13) - neste grupo, um dia após a laminectomia,
os animais receberam uma injeção intratecal de veículo;
2) laminectomia + NOC12-30nM (n=7) - os animais deste grupo foram
submetidos à laminectomia seguida, com o intervalo de um dia, pela injeção
intratecal de NOC12 na concentração de 30nM;
3) laminectomia + NOC12-300nM (n=5) - neste grupo, os ratos foram
submetidos à laminectomia seguida, com o intervalo de um dia, pela injeção
intratecal de NOC12 na concentração de 300nM;
4) compressão + veículo (n=10) - os animais deste grupo foram submetidos
ao estreitamento do canal medular em 35% e um dia após, à injeção intratecal
de veículo;
5) compressão + NOC12-30nM (n=10) - neste grupo, os ratos foram
submetidos ao estreitamento de 35% do canal medular seguida, com o
intervalo de um dia, pela injeção intratecal de NOC12 na concentração de
30nM;
6) compressão + NOC12-300nM (n=7) - neste grupo, um dia após o
estreitamento do canal medular em 35%, os animais receberam uma injeção
intratecal de NOC12 na concentração de 300nM;
7) descompressão + veículo (n=17) - os animais deste grupo foram
submetidos à laminectomia, estreitamento do canal medular em 35% e
descompressão seis horas após a inserção do espaçador. No dia seguinte os
animais receberam uma injeção intratecal de veículo;

31
8) descompressão + NOC12-30nM (n=11) - neste grupo, os ratos foram
submetidos ao estreitamento de 35% do canal medular e descompressão
medular 6 horas após. Um dia depois o NOC12 foi injetado via intratecal na
concentração de 30nM;
9) descompressão + NOC12-300nM (n=12) - neste grupo, os ratos foram
submetidos à descompressão após seis horas de estreitamento do canal
medular em 35%, e um dia depois receberam uma injeção intratecal de NOC12
na concentração de 300nM.
4.6. Avaliação Funcional
A avaliação sensorial dos animais foi realizada por meio do teste de
retirada da cauda (D´AMOUR; SMITH, 1941) no pré-teste, bem como 5 e 7 dias
após procedimento cirúrgico. O comportamento motor foi avaliado por meio da
escala de avaliação locomotora de Basso, Beattie e Bresnahan (1995, 1996).
Os testes de comportamento motor foram realizados no pré-teste, 1, 5 e 7 dias
após procedimento cirúrgico. Todos os testes foram realizados no mesmo
horário e local, como descritos a seguir.
4.6.1. Escala Basso, Beattie & Bresnahan (BBB)
Esta escala tem por objetivo avaliar a evolução da recuperação funcional
motora dos animais. Para isso, os animais são colocados um a um em uma
arena de testes idêntica a arena do teste de Campo Aberto e observados
durante 4 minutos, enquanto se locomovem livremente. A avaliação é realizada
através da observação de alguns parâmetros comportamentais: o movimento
do membro, a posição da pata, o tipo do passo, a coordenação da passada, a
abertura dos dedos, a rotação predominante da pata, a estabilidade do tronco e
a posição da cauda. Os parâmetros colhidos em cada teste foram anotados em
tabelas individuais (figura 2) e posteriormente classificados por meio de uma
escala que varia de 0 a 21 pontos (tabela 1). Os pontos refletem a condição
motora dos animais, onde zero representa paralisia total do membro e 21
pontos representa função motora normal do membro (BASSO; BEATTIE;
BRESNAHAN, 1995). Pontuações entre 0 e 7 indicam o retorno de movimentos
isolados de até três articulações (quadril, joelho e tornozelo). Pontuações entre
8 e 13 indicam o retorno dos passos plantares e coordenação dos movimentos

32
entre patas posteriores e anteriores. Por fim, pontuações entre 14 e 21
mostram o retorno da abertura dos dedos durante a passada, posição
predominante da pata em paralelo ao tronco, estabilidade de tronco e
levantamento da cauda. O teste foi realizado às cegas sempre pelo mesmo
examinador. Durante o teste os animais foram gentilmente estimulados a
caminhar continuamente.
Rato # _____ Data: ___/___/___
TRC: ____/____/ ____ DPO: ____ Score D _____ E _____
Movimento do membro Posicionamento
da pata Passo
Coo
rden
ação
Abertura
dedos
Posição
predominante
da pata
Inst
abili
dade
de
tro
nco
Cau
da
Coxa Joelho Tornoz.
Var
redu
ra
Suporte
plantar Dorsal Plantar Inicial Final
D E D E D E s/ c/ D E D E D E D E D E
ø ø ø ø ø ø D D
D
Es Mo
N N N N N N N I I I I Sim ↑
Di Di Di Di Di Di O O O O O O O
Ex Ex Ex Ex
E E E
Es Mo
F F F* F* F F* F* Não ↓
A A A A A A P P P P C C C C C C C
ø - s/ movimento D - direito N - nunca (0%) I - interna
Di - movim. discreto E - esquerdo O - ocasional (≤ 50%) Ex - externa
A - movim. amplo Es - estacionário F - frequente (51-94%) P - paralela
Mo - movimento C - consistente (95-100%) * mais que 4 repetições
Figura 2. Planilha de avaliação sensório-motora. Modelo de planilha utilizado para anotação dos dados observados durante a realização dos testes de avaliação locomotora pela escala BBB (segundo BASSO, BEATTIE e BRESNAHAN, 1995; modificado de SCHIAVETO DE SOUZA, 2004) e sensorial pelo teste de retirada de cauda (TRC).

33
Tabela 1. Categorias de pontuação da Escala BBB de avaliação locomotora*.
00 - Sem movimento do membro posterior; 01 - Movimento discreto de uma ou duas articulações 02 - Movimento amplo de uma articulação, ou movimento amplo de uma articulação e discreto de outra articulação;
03 - Movimento amplo de duas articulações; 04 - Movimento discreto das três articulações do membro posterior; 05 - Movimento discreto de duas articulações e amplo da terceira; 06 - Movimento amplo de duas articulações e discreto da terceira; 07 - Movimento amplo das três articulações do membro posterior; 08 - Movimento em “varredura” do membro posterior, ou posicionamento plantar da pata, ambos sem sustentação de peso; 09 - Posicionamento plantar da pata com sustentação do peso na posição estática, ou passos dorsais com sustentação de peso ocasional, freqüente ou constante e sem passos plantares; 10 - Passos plantares ocasionais, com sustentação de peso, mas sem coordenação; 11 - Passos plantares com sustentação de peso de freqüente a constante e sem coordenação;
12 - Passos plantares com sustentação de peso de freqüente a constante e coordenação ocasional; 13 - Passos plantares com sustentação de peso de freqüente a constante e coordenação freqüente; 14 - Passos plantares com sustentação de peso constante, com coordenação constante; posição da pata predominantemente rodada (externamente ou internamente) no contato inicial ou no final da fase de apoio, ou passos plantares freqüentes, coordenação constante e passos dorsais ocasionais; 15 - Passos plantares e coordenação constantes, sem abertura dos dedos, ou abertura ocasional dos dedos; posição da pata predominantemente paralela ao corpo no contato inicial; 16 - Passos plantares e coordenação constantes; abertura dos dedos freqüente; posição da pata predominantemente paralela ao contato inicial, e rodada ao final da fase de apoio; 17 - Passos plantares e coordenação constantes; abertura dos dedos freqüente; posição da pata predominantemente paralela ao contato inicial e ao final do apoio; 18 - Passos plantares e coordenação constantes; abertura dos dedos constante; posição da pata predominantemente paralela ao contato inicial e rodada ao final da fase de apoio; 19 - Passos plantares e coordenação constantes; abertura dos dedos constante; posição da pata predominantemente paralela ao contato inicial e ao final da fase de apoio, cauda abaixada durante a caminhada; 20 - Passos plantares e coordenação constantes; constante abertura dos dedos; posição da pata predominantemente paralela ao contato inicial e ao final da fase de apoio; cauda constantemente erguida e tronco instável; 21 - Passos plantares, coordenação e abertura dos dedos constantes; posição da pata predominantemente paralela em todas as posições; estabilidade do tronco e cauda erguida. Obs.: O item coordenação refere-se à coordenação entre os membros anteriores e posteriores. * BASSO; BEATTIE; BRESNAHAN, 1996.

34
4.6.2. Teste de retirada de cauda
O teste foi realizado segundo o método descrito por D’Amour e Smith
(1941). Para a realização do teste, cada rato foi gentilmente imobilizado. A
cauda do animal era então colocada sobre uma espiral feita com fio de níquel-
cromo de 0,3mm de diâmetro, a qual era aquecida pela passagem de uma
corrente elétrica por ela. A taxa de aquecimento da espiral de níquel cromo,
pela passagem da corrente elétrica, foi padronizada em 9oC/s. Neste teste é
medido o tempo de latência para a retirada da cauda de uma superfície
aquecida. No início do experimento, foi feito um pequeno ajuste da corrente
para que a latência de retirada da cauda de um animal inocente ficasse entre
2,5 e 3,5 segundos, em três medidas consecutivas (GIGLIO et al., 2006). Um
tempo máximo de corte de 6 segundos foi estipulado, sendo então o
aquecimento interrompido automaticamente, a fim de evitar a ocorrência de
lesão da cauda do animal. Cada teste consistiu da média de três medidas,
tomadas em intervalos de 5 minutos. A média da linha de base para latência de
retirada de cauda (média LBLRC) foi calculada para cada animal como a média
das medidas do pré-teste. Cada latência de retirada de cauda (LRC) foi
normalizada pelo índice de anti-nocicepção (IA) usando a fórmula IA= [(LRC −
média LBLRC) / (6 − média LBLRC)] (D'AMOUR; SMITH, 1941; ECHEVERRY
et al., 2004). Valores positivos indicam antinocicepção ou aumento do limiar
para dor; enquanto valores negativos indicam hipernocicepção, ou redução do
limiar de dor.
4.7. Sacrifício, retirada e conservação da medula
No sétimo dia após os procedimentos experimentais, os animais foram
anestesiados com Uretana na dose de 1,5 g/kg de peso do animal. Após a
anestesia os animais foram perfundidos com o objetivo de lavar o sangue e
pré-fixar os tecidos do animal. Isso foi feito por meio da infusão de 200 mL de
solução fosfato salina tamponada (PBS) a 0,01M, pH 7,4, seguida de 200mL de
paraformaldeído (PFA) 4% em PB 0,1M, pH 7,4. Após a perfusão, o dorso dos
animais foi dissecado, sendo retirados os processos espinhosos e transversos
das vértebras, expondo a medula (ME). A região medular contendo o epicentro
da lesão e regiões rostral e caudal adjacentes (aproximadamente 2,5 cm) foi

35
cuidadosamente dissecada em cada animal. A medula foi fixada a um palito de
madeira, para manter a posição anatômica e colocada em uma solução de PFA
4%, pH 7,4 por 2 horas para pós-fixação seguida de solução de sacarose
tamponada 30% por 18 horas (4 °C) para crioproteção dos tecidos. Decorridas
18 horas, as amostras foram mergulhadas em meio de inclusão para tecido
congelado (Tissue-Tek®), congelados em gelo seco e mantidos em freezer à -
70°C para posterior análise.
4.8. Secção da medula espinal
A medula espinal de cada animal foi levada do freezer à -70ºC para o
criostato, a uma temperatura de –20ºC. A seguir foram feitos cortes
transversais de 20 µm de espessura, no sentido rostro-caudal. Os cortes foram
colhidos diretamente em lâminas para microscópio, previamente gelatinizadas.
Foram colhidos 3 conjuntos de 7 lâminas de cada medula espinal, contendo
aproximadamente 24 cortes em cada uma. As secções foram colhidas de forma
serial. Foi colocada uma secção em cada uma das sete lâminas do conjunto e
descartadas dez secções. Em seguida, foi colocada outra secção em cada
lâmina, ao lado daquela já colhida. Após o preenchimento de um conjunto, um
novo conjunto foi iniciado. Desse modo, o primeiro conjunto recebeu os cortes
rostrais à lesão, o segundo, do centro da lesão e o terceiro da região caudal à
lesão. Após a coleta dos cortes, as lâminas foram mantidas em caixas
apropriadas em freezer a –20°C até o dia do ensaio.
4.9. Análise histológica.
As modificações histológicas decorrentes do estreitamento do canal
vertebral e da sua associação com a injeção intratecal de NOC12 foram
analisadas por meio da coloração de HE (EMERY et al., 1998; WADA et al.,
1999) e pela imunocitoquímica para a enzima NOS neuronal. As modificações
foram avaliadas 7 dias após o procedimento experimental.
As quantificações foram realizadas por análise de imagem
computadorizada utilizando microscópio acoplado à câmera e aplicativo
Axiovision software (Zeiss, Alemanha).

36
Para que a análise histológica fosse realizada às cegas, os códigos de
identificação de cada lâmina foram cobertos por outro experimentador antes do
início da análise e revelados após o término.
4.10. Coloração Hematoxilina-eosina
Nesta coloração, a hematoxilina, age como corante natural, por meio de
seu produto de oxidação, a hemateína. A oxidação química produz-se pelo uso
de iodato de potássio ou óxido de mercúrio. Para corar o tecido, a hemateína
necessita de um mordente (sais de alumínio, ferro, cromo, cobre ou
tungstênio), prévio ou incorporado na própria solução de hematoxilina.
Convencionalmente, para que seja dado o contraste à coloração é usada uma
solução de eosina logo após a aplicação de hematoxilina. Como resultado
dessa coloração, os núcleos são corados em azul (hematoxilina), enquanto o
citoplasma e a maioria dos outros tecidos ficam corados de rosado a vermelho
(eosina; PROPHET et al., 1995).
A técnica da coloração consistiu em deixar as secções de tecido em
contato com a hematoxilina de Harris por 90 segundos, lavar em água corrente
por 5 minutos, colocar para azular, na solução de água amoniacal, por 15
segundos, lavar em água corrente por 5 minutos e corar com a solução de
Eosina por 30 segundos. A seguir as lâminas passavam por uma bateria de
álcoois em concentração crescente, uma mistura de álcool e xilol e xilol puro (3
minutos cada) para então serem montadas com Entellan.
Por meio desta coloração foram avaliadas a densidade de células
marcadas para HE e a extensão longitudinal da lesão medular.
4.11. Análise da densidade de células marcadas para HE
A densidade de células marcadas para HE foi medida na medula
espinal, bilateralmente nos cornos dorsal (lâminas I, II e III) e ventral (lâminas
VIII e IX) dos animais dos grupos descompressão+veículo,
descompressão+NOC12-30 e 300nM. As regiões foram identificadas de acordo
com ilustrações apresentadas por Molander e colaboradores (1989 – T10 na
figura 3A). Para análise, cada amostra de medula espinal foi dividida em quatro
segmentos (figura 3B): dois rostrais e dois caudais ao sítio de lesão (4 mm de
comprimento cada – modificado de TRUDRUNG; WIRTH; MENSE, 2000). Em
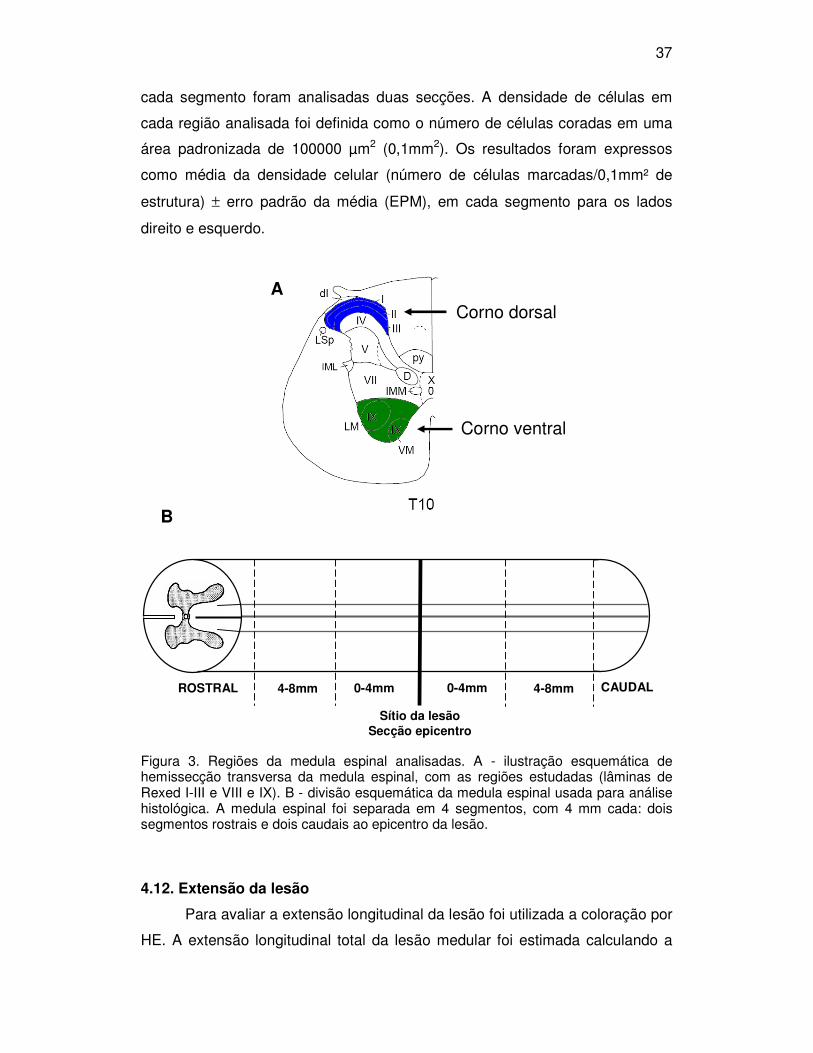
37
cada segmento foram analisadas duas secções. A densidade de células em
cada região analisada foi definida como o número de células coradas em uma
área padronizada de 100000 µm2 (0,1mm2). Os resultados foram expressos
como média da densidade celular (número de células marcadas/0,1mm² de
estrutura) ± erro padrão da média (EPM), em cada segmento para os lados
direito e esquerdo.
Figura 3. Regiões da medula espinal analisadas. A - ilustração esquemática de hemissecção transversa da medula espinal, com as regiões estudadas (lâminas de Rexed I-III e VIII e IX). B - divisão esquemática da medula espinal usada para análise histológica. A medula espinal foi separada em 4 segmentos, com 4 mm cada: dois segmentos rostrais e dois caudais ao epicentro da lesão.
4.12. Extensão da lesão
Para avaliar a extensão longitudinal da lesão foi utilizada a coloração por
HE. A extensão longitudinal total da lesão medular foi estimada calculando a
Sítio da lesão Secção epicentro
0-4mm 0-4mm 4-8mm 4-8mm ROSTRAL CAUDAL
B
A Corno dorsal
Corno ventral

38
distância entre a primeira e a última secção contendo sinais de lesão (KIGERL;
MCGAUGHY; POPOVICH, 2006). O epicentro da lesão foi definido como a
secção que contivesse menor área de preservação. Os seguintes sinais de
lesão foram considerados para o cálculo: hemorragia, cavitação, áreas de
hipercelularidade, fragmentação tecidual ou arquitetura anormal do tecido,
desorganização da substância cinzenta ou branca e oclusão moderada ou
severa do canal central. Para observar possíveis discrepâncias na distribuição
rostro-caudal da lesão também foi calculada a extensão parcial da lesão: 1) a
extensão rostral da lesão - a distância entre a secção considerada como
epicentro e a primeira secção (rostral) que contivesse sinais de dano tecidual; e
2) a extensão caudal da lesão - a distância entre a secção considerada como
epicentro e a última secção (caudal) com sinais de dano tecidual.
Para que o cálculo da distância entre as secções fosse mais acurado,
durante a secção da medula no criostato foram anotados os cortes perdidos/
dispensados e sua localização ao longo da amostra. Esses dados foram
considerados nos cálculos de extensão parcial ou total da medula. Os
resultados foram expressos como a extensão da lesão para rostral e caudal a
partir do epicentro da lesão, bem como, a extensão total da lesão em
micrômetros (µm).
4.13. Porcentagem de tecido preservado
Secções coradas para HE foram utilizadas para quantificar a
porcentagem de tecido poupado da lesão. O tecido poupado foi definido como
aquele livre de sinais de lesão, sendo considerados: hemorragia, cavitação,
áreas de hipercelularidade, fragmentação tecidual ou arquitetura anormal do
tecido, desorganização da substância cinzenta ou branca e oclusão moderada
ou severa do canal central. A análise morfológica se concentrou na medida do
parênquima medular no sítio da lesão (epicentro). As medidas de área total e
área poupada (em µm) foram transformadas em porcentagem e expressa em
porcentagem de área poupada em relação à área total de cada secção
analisada (BEATTIE; BRESNAHAN, 1995, 1996; MA et al., 2001; BASSO;
PLEMEL et al., 2008).

39
4.14. Imunocitoquímica para a enzima NOS neuronal
A imunocitoquímica pode ser considerada como a demonstração de
antígenos em secções de tecidos, por meio do uso da interação antígeno-
anticorpo, culminando com a ligação de um marcador ao antígeno. O objetivo
principal é alcançar reprodutibilidade e demonstração consistente de antígenos
específicos, com um mínimo de coloração de fundo, preservando a integridade
do tecido.
4.15. Análise da densidade de células positivas para NOS neuronal
Para análise da expressão da proteína NOSn foram utilizadas secções
dos grupos descompressão+veículo, descompressão+NOC12-30 e 300nM
adjacentes àquelas coradas para HE. A técnica consistiu em incubar as
secções em uma solução contendo anticorpo primário para NOS1 ou neuronal
(rabbit polyclonal IgG – doado por P.C. Emson, Cambridge, UK), na diluição
1:2000 (em PBS 0,1M); soro de cabra 1% e Triton X-100 0,2%, a 18°C, por
24h, seguida pela incubação com o anticorpo secundário (anti-rabbit), diluição
1:400, a 25°C, por 1h. O tecido foi então incubado com o complexo avidina-
biotina-peroxidase (diluição 1:150), a 25°C, por 1h. A reação foi revelada pela
adição de 3,3-diaminobenzidina (DAB) na presença de peróxido de hidrogênio,
por 10 minutos. As lâminas com os tecidos foram então desidratadas,
diafanizadas e montadas com lamínulas e Entellan.
A densidade de neurônios imunorreativos para a NOS foi quantificada
por análise de imagem computadorizada assim como descrito para a análise do
HE. Foram analisadas as lâminas I-III e as lâminas VIII-IX de Rexed em dois
segmentos rostrais e dois caudais ao epicentro da lesão (distantes 4 mm – Fig.
3). A densidade de neurônios imunorreativos em cada região analisada foi
definida como o número de células positivamente marcadas em uma área
padronizada de 100000 µm2 (0,1mm2) e expressa como média ± EPM da
densidade de células NOSn+/0,1mm2.
4.16. Análise estatística
Os resultados obtidos por meio dos testes de comportamento, bem
como daqueles obtidos pela análise histológica, foram analisados

40
estatisticamente pelos “softwares” SPSS para Windows, na versão 8.0 ou pelo
SigmaStat para Windows, na versão 2.0. Em todos os testes utilizados foram
consideradas estatisticamente significantes as diferenças na qual o valor de p
foi menor ou igual a 0,05.
4.16.1. Comportamento
Os resultados de comportamento motor não mostraram diferença
estatística entre escores obtidos para cada lado, por este motivo foram feitas
médias dos escores dos lados. Os dados da escala BBB e do teste de retirada
de cauda foram comparados por meio de análise de variança (ANOVA) de três
vias de medidas repetitivas, sendo considerados os fatores: lesão, tratamento e
tempo. Após este teste, se houve efeito e/ou interação entre os fatores, as
médias de cada fator foram comparadas por ANOVA de uma/duas vias
(independente e/ou de medidas repetitivas), seguido pelo teste de múltiplas
comparações de Duncan.
4.16.2. Resultados da análise histológica
Inicialmente, os resultados da densidade de células marcadas para HE e
imunorreativas para NOSn foram comparados por ANOVA de medidas
repetitivas e não mostraram diferença estatística entre os lados. Por este
motivo estes dados foram agrupados como média dos lados e comparados por
ANOVA de uma via, seguida pelo teste de múltiplas comparações de Duncan,
sendo o tratamento o fator analisado. Os resultados da extensão de lesão total
e área de tecido poupado foram comparados por meio de ANOVA de duas
vias, sendo considerados os fatores: lesão e tratamento. Após este teste, se
houve efeito e/ou interação entre os fatores, as médias de cada fator foram
comparadas por ANOVA de uma via, seguida pelo teste de múltiplas
comparações de Duncan. Para análise dos resultados da extensão parcial de
lesão foi utilizada ANOVA de três vias, sendo considerados os fatores: lesão
(laminectomia, compressão e descompressão), tratamento (veículo, NOC12 30
e 300nM) e direção (rostral e caudal). Esta análise foi seguida pela ANOVA de
duas vias e teste de múltiplas comparações de Duncan.
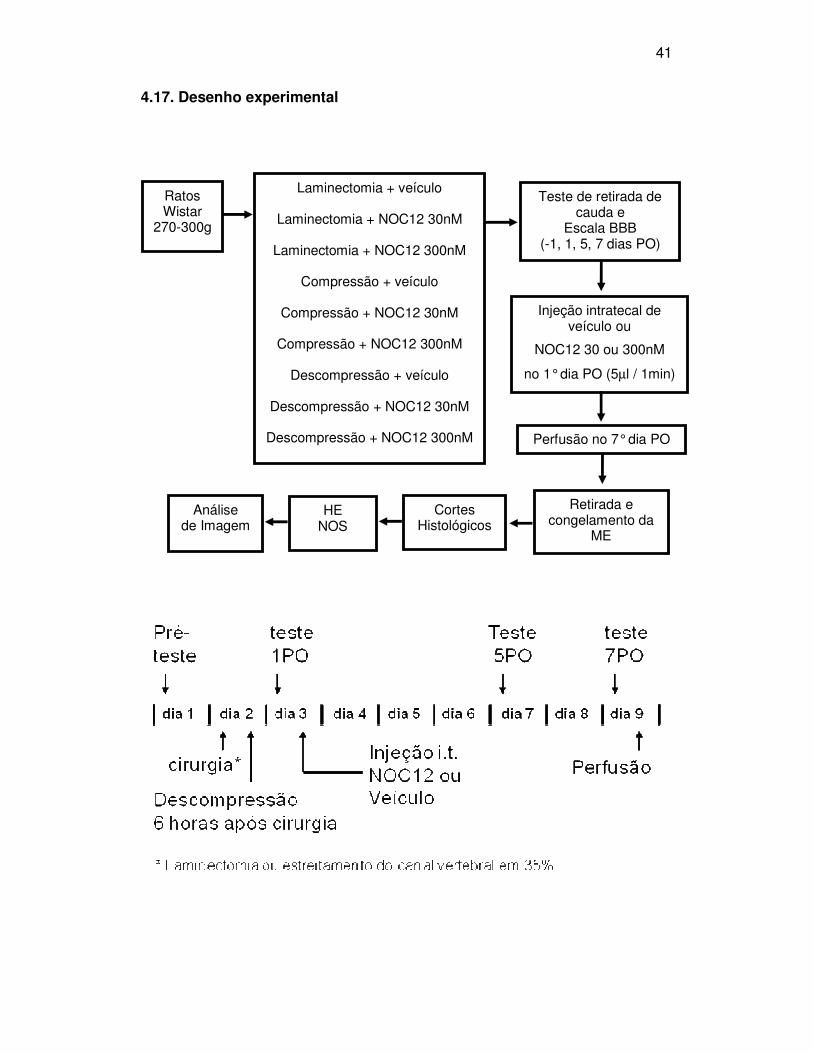
41
4.17. Desenho experimental
Ratos Wistar
270-300g
Perfusão no 7° dia PO
Retirada e congelamento da
ME
Cortes Histológicos
HE NOS
Análise de Imagem
Teste de retirada de cauda e
Escala BBB (-1, 1, 5, 7 dias PO)
Injeção intratecal de veículo ou
NOC12 30 ou 300nM
no 1° dia PO (5µl / 1min)
Laminectomia + veículo
Laminectomia + NOC12 30nM
Laminectomia + NOC12 300nM
Compressão + veículo
Compressão + NOC12 30nM
Compressão + NOC12 300nM
Descompressão + veículo
Descompressão + NOC12 30nM
Descompressão + NOC12 300nM

42
5. RESULTADOS
5.1. Escala BBB
As alterações da função motora decorrentes do estreitamento do canal
vertebral, da descompressão e da injeção intratecal de NOC12, avaliadas por
meio da escala BBB, estão apresentadas na figura 4.
Os ratos do grupo laminectomia + veículo apresentaram uma média de
21 pontos na BBB, indicando função locomotora normal. A administração de
NOC12-30 ou 300nM, per si, não modificou os escores dos animais
submetidos à laminectomia em todos os testes realizados.
De modo geral, a compressão medular por meio do estreitamento do
canal vertebral em 35% induziu importante déficit locomotor em todos os dias
pós-operatórios (F(3,22) = 90,524, p<0,001, teste de Duncan, p<0,05). No
primeiro dia pós-operatório, os animais submetidos à compressão
apresentaram déficit motor acentuado, com pontuação BBB entre 7 (movimento
amplo das três articulações do membro posterior) e 10 (passos plantares
ocasionais, com sustentação de peso, mas sem coordenação entre o membro
anterior e posterior).
Em todos os dias analisados, os grupos compressão que receberam
injeção intratecal de veículo ou NOC12-30nM apresentaram pontuação BBB
significativamente menor que os grupos laminectomia e descompressão que
receberam as mesmas drogas (veículo- F(6,68) = 7,681, p<0,001; NOC12-30nM -
F(6,44) = 10,631, p<0,001; teste de Duncan, p<0,05; figura 4). O grupo
compressão + NOC12-300nM obteve pontuação BBB menor que o grupo
laminectomia nos tempos 1 e 7 dias pós-operatório (F(6,36) = 5,647, p<0,001;
teste de Duncan, p<0,05). No quinto dia pós-operatório este grupo apresentou
pontuação significativamente menor que os grupos laminectomia e
descompressão que receberam NOC12-300nM (teste de Duncan, p<0,05). Não
houve diferença significativa entre os grupos compressão submetidos à injeção
intratecal de NOC12-30 ou 300nM e aqueles que receberam veículo. Todos os
grupos compressão apresentaram pontuação BBB significativamente maior no
sétimo dia pós-operatório, quando comparado com o primeiro dia (teste de
Duncan, p<0,05). Não houve diferença entre o quinto e o primeiro dia pós-
operatório para nenhum dos grupos compressão.

43
A cirurgia para descompressão medular reduziu o déficit locomotor no
primeiro dia pós-operatório, independentemente da injeção intratecal (F(2,83) =
46,314, p<0,001, teste de Duncan, p<0,05). Com isso os ratos passaram a
exibir no primeiro dia pós-operatório uma pontuação BBB de 13 a 14 (passos
plantares com suporte de peso freqüente a constante; coordenação entre o
membro anterior e posterior de freqüente a constante; e posição da pata
durante locomoção predominantemente rodada). Adicionalmente, o grupo
descompressão + veículo mostrou uma recuperação significativa cinco e sete
dias após a lesão, comparado com 1° dia pós-operatório (F(3,67)=17,076,
p<0,001, teste de Duncan, p<0.05).
No primeiro dia pós-operatório, os animais submetidos à descompressão
medular (ainda sem a administração de NOC12-30nM) mostraram pontuação
BBB significativamente menor que a do grupo laminectomia e maior que a do
grupo compressão (teste de Duncan, p<0,05). Nos dias 5 e 7 pós-operatório,
eles apresentaram pontuação BBB similar à do grupo laminectomia + NOC12
30nM (F(6,64)=10,631, p<0.001, teste de Duncan, p>0.05). Comparada com o
grupo que recebeu veículo, a administração de NOC12-30nM após a
descompressão induziu aumento na pontuação da escala BBB nos dias 5 e 7
pós-operatório. A pontuação passou de 17 (freqüente) para 18 e 19 (abertura
dos dedos constante).
A pontuação BBB dos animais do grupo descompressão + NOC12
300nM no primeiro dia pós-operatório foi significativamente menor que a do
grupo laminectomia + NOC12-300nM (F(6,36)= 5,647, p<0.001,teste de Duncan,
p<0,05). No quinto e sétimo dia pós-operatório o grupo descompressão +
NOC12-300nM apresentou pontuação BBB semelhante a do grupo
laminectomia. Houve um aumento significativo da pontuação BBB do grupo
descompressão + NOC12-300nM no quinto dia pós-operatório, quando
comparado ao grupo compressão + NOC12-300nM (teste de Duncan, p<0,05).
O grupo descompressão que recebeu NOC12-300nM não foi diferente do que
recebeu veículo em nenhum dos tempos analisados. Adicionalmente,
pontuação dos dias 5 e 7 pós-operatório foi significativamente maior que a do
primeiro dia, quando ainda não haviam recebido injeção de NOC12-300nM
(teste de Duncan, p<0,05).
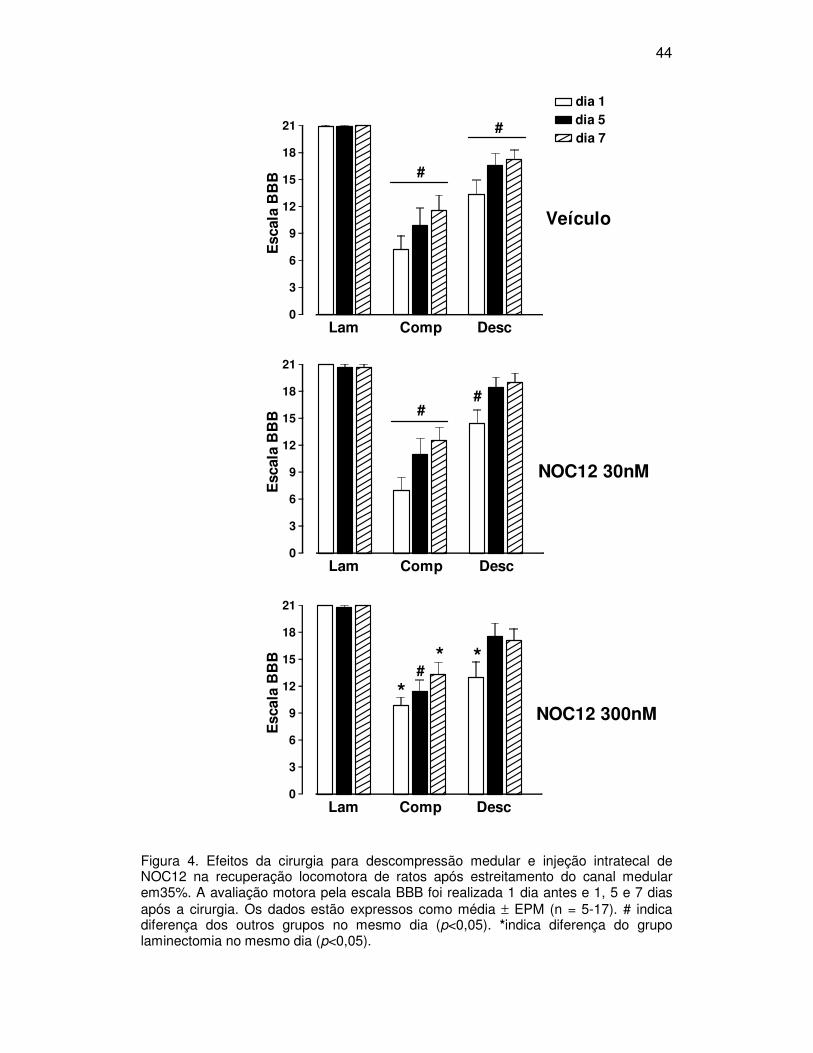
44
0
3
6
9
12
15
18
21
Esc
ala
BB
B
dia 7dia 5dia 1
#
#
Veículo
Lam Comp Desc
0
3
6
9
12
15
18
21
Esc
ala
BB
B ##
Lam Comp
NOC12 30nM
Desc
0
3
6
9
12
15
18
21
Esc
ala
BB
B
*
Lam Comp Desc
*#
*
NOC12 300nM
Figura 4. Efeitos da cirurgia para descompressão medular e injeção intratecal de NOC12 na recuperação locomotora de ratos após estreitamento do canal medular em35%. A avaliação motora pela escala BBB foi realizada 1 dia antes e 1, 5 e 7 dias após a cirurgia. Os dados estão expressos como média ± EPM (n = 5-17). # indica diferença dos outros grupos no mesmo dia (p<0,05). *indica diferença do grupo laminectomia no mesmo dia (p<0,05).
45
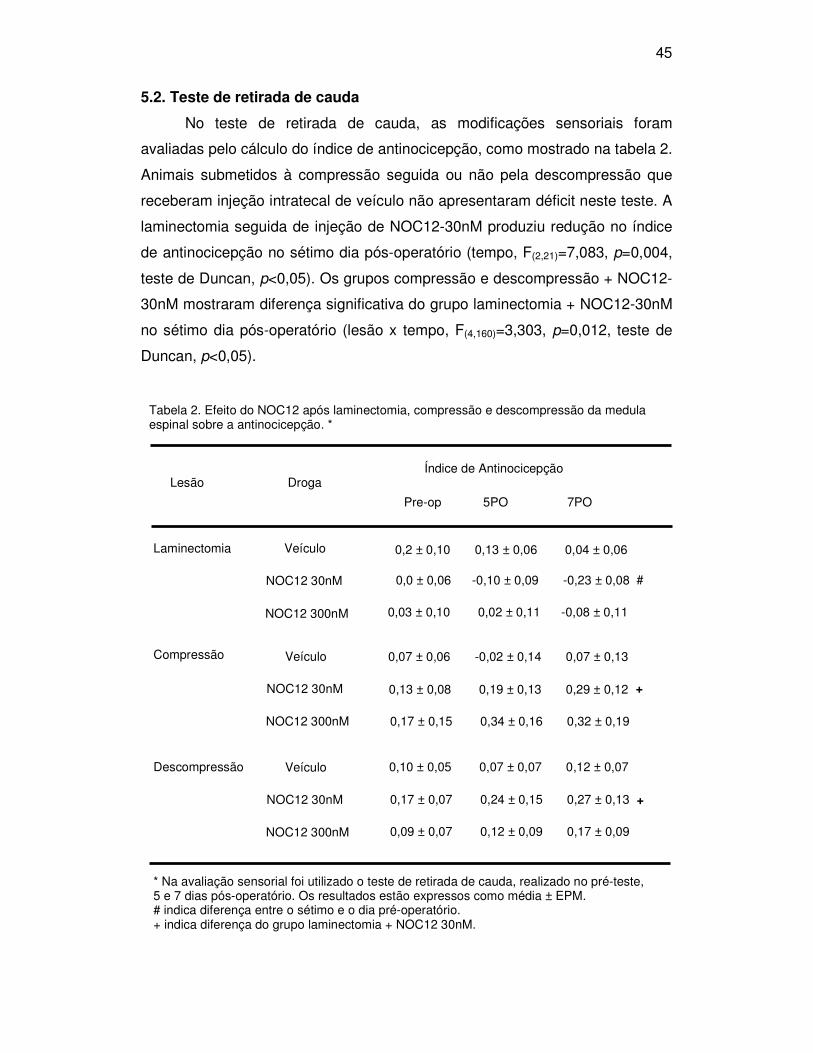
5.2. Teste de retirada de cauda
No teste de retirada de cauda, as modificações sensoriais foram
avaliadas pelo cálculo do índice de antinocicepção, como mostrado na tabela 2.
Animais submetidos à compressão seguida ou não pela descompressão que
receberam injeção intratecal de veículo não apresentaram déficit neste teste. A
laminectomia seguida de injeção de NOC12-30nM produziu redução no índice
de antinocicepção no sétimo dia pós-operatório (tempo, F(2,21)=7,083, p=0,004,
teste de Duncan, p<0,05). Os grupos compressão e descompressão + NOC12-
30nM mostraram diferença significativa do grupo laminectomia + NOC12-30nM
no sétimo dia pós-operatório (lesão x tempo, F(4,160)=3,303, p=0,012, teste de
Duncan, p<0,05).
Tabela 2. Efeito do NOC12 após laminectomia, compressão e descompressão da medula espinal sobre a antinocicepção. *
Droga Lesão Índice de Antinocicepção
Laminectomia
Pre-op 5PO 7PO
Veículo
NOC12 30nM
NOC12 300nM
Veículo
NOC12 30nM
NOC12 300nM
Veículo
NOC12 30nM
NOC12 300nM
Compressão
Descompressão
0,2 ± 0,10 0,13 ± 0,06 0,04 ± 0,06
0,0 ± 0,06 -0,10 ± 0,09 -0,23 ± 0,08
0,03 ± 0,10 0,02 ± 0,11 -0,08 ± 0,11
0,07 ± 0,06 -0,02 ± 0,14 0,07 ± 0,13
0,13 ± 0,08 0,19 ± 0,13 0,29 ± 0,12
0,17 ± 0,15 0,34 ± 0,16 0,32 ± 0,19
0,10 ± 0,05 0,07 ± 0,07 0,12 ± 0,07
0,17 ± 0,07 0,24 ± 0,15 0,27 ± 0,13
0,09 ± 0,07 0,12 ± 0,09 0,17 ± 0,09
+
+
#
* Na avaliação sensorial foi utilizado o teste de retirada de cauda, realizado no pré-teste, 5 e 7 dias pós-operatório. Os resultados estão expressos como média ± EPM. # indica diferença entre o sétimo e o dia pré-operatório. + indica diferença do grupo laminectomia + NOC12 30nM.

46
5.3. Exame histológico
Fotomicrografias de medula espinal dos grupos laminectomia,
compressão e descompressão coradas com HE são apresentadas nas figuras
5A, B e C, respectivamente. No exame microscópico dos grupos laminectomia
não foram encontrados sinais de lesão ao longo da medula (Fig. 5A e 5A’). Nas
secções do epicentro da lesão, a compressão induziu uma redução na
distância dorso-ventral da medula (barra, Fig. 5B). Esta redução foi
parcialmente restaurada pela descompressão (barra, Fig. 5C). Foram ainda
observadas mudanças na arquitetura do tecido com deslocamento da
substância branca e cinzenta (Fig. 5B’ and 5C’). Com a compressão, o canal
central foi ocluído (Fig. 5B’), o que foi atenuado pela descompressão (Fig. 5C’).
Cavitações difusas foram encontradas com maior incidência nos grupos
descompressão (cabeça de seta na Fig. 5C’).
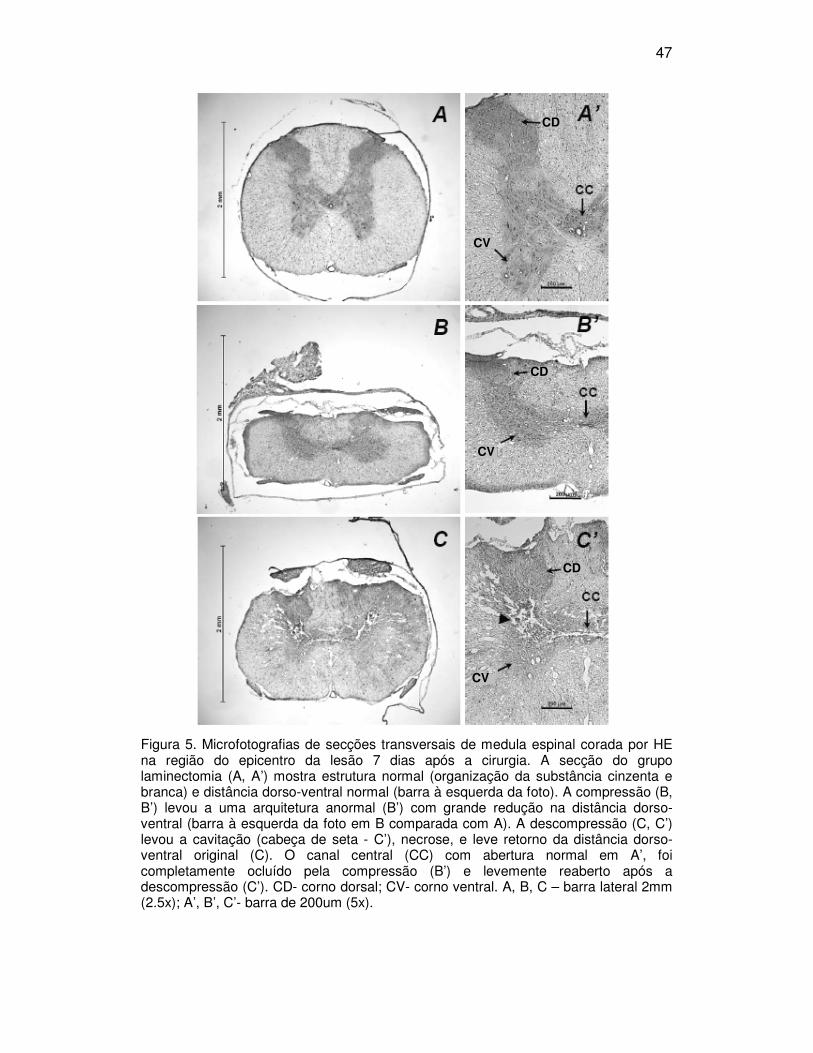
47
Figura 5. Microfotografias de secções transversais de medula espinal corada por HE na região do epicentro da lesão 7 dias após a cirurgia. A secção do grupo laminectomia (A, A’) mostra estrutura normal (organização da substância cinzenta e branca) e distância dorso-ventral normal (barra à esquerda da foto). A compressão (B, B’) levou a uma arquitetura anormal (B’) com grande redução na distância dorso-ventral (barra à esquerda da foto em B comparada com A). A descompressão (C, C’) levou a cavitação (cabeça de seta - C’), necrose, e leve retorno da distância dorso-ventral original (C). O canal central (CC) com abertura normal em A’, foi completamente ocluído pela compressão (B’) e levemente reaberto após a descompressão (C’). CD- corno dorsal; CV- corno ventral. A, B, C – barra lateral 2mm (2.5x); A’, B’, C’- barra de 200um (5x).
CD
CV
CD
CV
CV
CD

48
5.4. Extensão da lesão
A extensão total da lesão (porção rostral+caudal) estimada em secções
medulares dos grupos laminectomia, compressão e descompressão com
injeção intratecal de NOC12 ou veículo é apresentada na figura 6. Os grupos
compressão e descompressão que receberam veículo, NOC12-30 ou 300nM
mostraram lesão mais extensa que os animais do respectivo grupo
laminectomia (F(2,34)=39,7, p<0,01, teste de Duncan, p<0,05). A extensão de
lesão do grupo descompressão + veículo não foi diferente do grupo
compressão + veículo.
A extensão da lesão do grupo descompressão + NOC12-30nM foi
reduzida significativamente quando comparada com o grupo descompressão +
veículo (F(4,34)=3,52, p=0,016, teste de Duncan, p<0,05). Houve ainda um efeito
marginal da injeção de NOC12-30nM com redução na extensão da lesão do
grupo descompressão quando comparado com o grupo compressão + NOC12
30nM (teste de Duncan, p=0,06). O grupo descompressão que recebeu NOC12
300nM apresentou redução na extensão da lesão quando comparado com o
grupo compressão + NOC12-300nM (F(2,11)=22,845, p<0,001, teste de Duncan,
p<0,05).
O avanço da lesão para o sentido rostral e caudal foi avaliado
comparando-se as medidas de lesão a partir do epicentro da lesão até o
extremo rostral e caudal onde houvesse sinal de lesão. Os resultados da
extensão parcial da lesão (mostrando a porção rostral e caudal da lesão
separadamente) são mostrados na figura 7, 8 e 9. A análise estatística mostrou
que os grupos compressão e descompressão que receberam veículo, NOC12-
30nM ou 300nM apresentaram extensão de lesão rostral e distal
significativamente maior que os grupos laminectomia, exceto o grupo
descompressão NOC12 30nM no sentido rostral ao epicentro (veículo -
F(2,12)=17,982, p<0,001; NOC12 30nM - F(2,11)=8,323, p=0,006; NOC12-300nM -
F(2,11)=22,845, p<0,001, teste de Duncan, p<0,05). Os grupos descompressão
que receberam NOC12-30 e 300nM apresentaram redução na extensão caudal
da lesão medular quando comparados com o grupo descompressão + veículo
(efeito da droga, F(2,12)=3,722, p=0,055, teste de Duncan, p<0,05). O grupo
descompressão + NOC12-300nM mostrou ainda redução significativa na
extensão da lesão na região caudal ao epicentro da lesão quando comparado
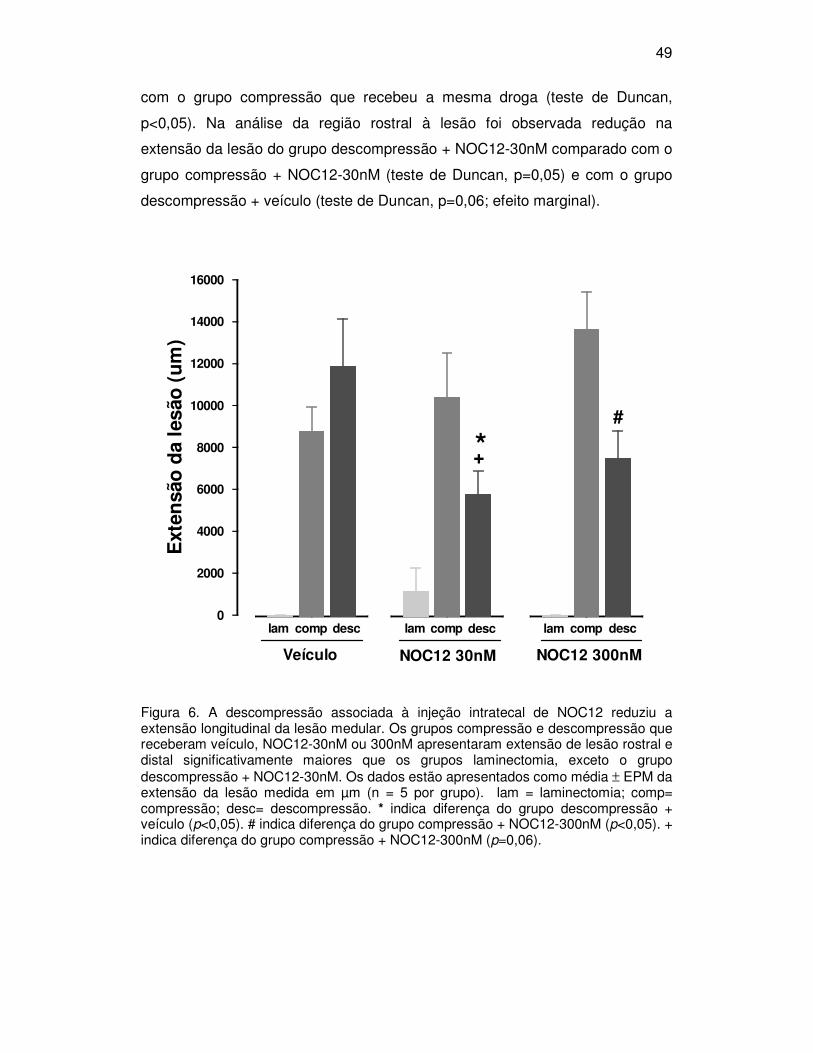
49
com o grupo compressão que recebeu a mesma droga (teste de Duncan,
p<0,05). Na análise da região rostral à lesão foi observada redução na
extensão da lesão do grupo descompressão + NOC12-30nM comparado com o
grupo compressão + NOC12-30nM (teste de Duncan, p=0,05) e com o grupo
descompressão + veículo (teste de Duncan, p=0,06; efeito marginal).
0
2000
4000
6000
8000
10000
12000
14000
16000
NOC12 300nM
Ext
ensã
o d
a le
são
(u
m)
Veículo NOC12 30nM
lam lamcomp lamcomp comp
+
desc desc desc
#
*
Figura 6. A descompressão associada à injeção intratecal de NOC12 reduziu a extensão longitudinal da lesão medular. Os grupos compressão e descompressão que receberam veículo, NOC12-30nM ou 300nM apresentaram extensão de lesão rostral e distal significativamente maiores que os grupos laminectomia, exceto o grupo descompressão + NOC12-30nM. Os dados estão apresentados como média ± EPM da extensão da lesão medida em µm (n = 5 por grupo). lam = laminectomia; comp= compressão; desc= descompressão. * indica diferença do grupo descompressão + veículo (p<0,05). # indica diferença do grupo compressão + NOC12-300nM (p<0,05). + indica diferença do grupo compressão + NOC12-300nM (p=0,06).
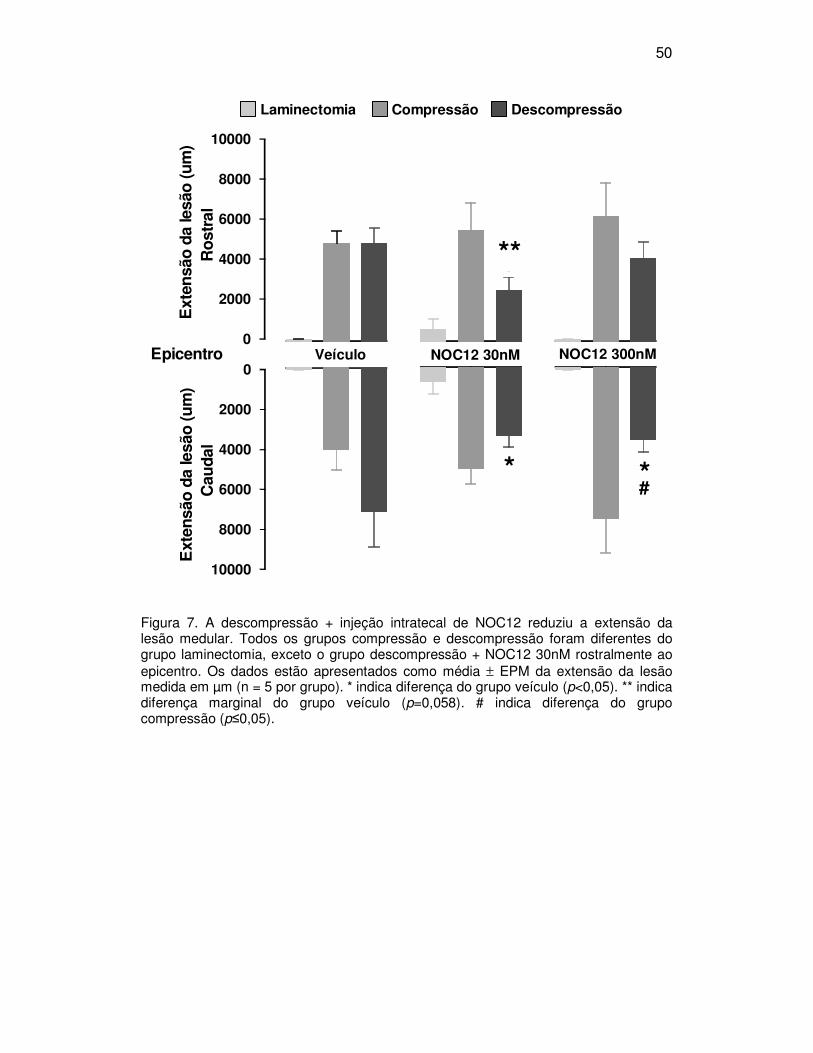
50
0
2000
4000
6000
8000
10000
Ext
ensã
o d
a le
são
(u
m)
10000
8000
6000
4000
2000
0
Ext
ensã
o d
a le
são
(u
m)
Laminectomia Compressão Descompressão
Veículo NOC12 30nM NOC12 300nM
*
Epicentro
Ro
stra
lC
aud
al
+**
*#
Figura 7. A descompressão + injeção intratecal de NOC12 reduziu a extensão da lesão medular. Todos os grupos compressão e descompressão foram diferentes do grupo laminectomia, exceto o grupo descompressão + NOC12 30nM rostralmente ao epicentro. Os dados estão apresentados como média ± EPM da extensão da lesão medida em µm (n = 5 por grupo). * indica diferença do grupo veículo (p<0,05). ** indica diferença marginal do grupo veículo (p=0,058). # indica diferença do grupo compressão (p≤0,05).

51

52

53
Figura 9. Microfotografias de secções transversais de medula espinal de animais dos grupos descompressão + veículo ou NOC12 30 ou 300nM coradas por HE 7 dias após a cirurgia. As secções dos grupos descompressão+veículo (A, D e G); descompressão+NOC12 30nM (B, E e H) e descompressão+NOC12 300nM (C, F e I) mostram a extensão da lesão nos diferentes segmentos analisados: epicentro (D, E e F), rostral (A, B e C) e caudal à lesão (G, H e I). As setas indicam regiões onde pode-se observar infiltrado, regiões de fragmentação tecidual, desorganização da substância branca e cinzenta. Rostral = 3500-4500µm do epicentro. Caudal = 4500-5500µm do epicentro. Na foto A (5x) a barra indica 0,3mm; na microfotografia panorâmica (2.5x) a barra indica 0,5mm.
5.5. Porcentagem de tecido preservado
A porcentagem de tecido preservado da lesão no epicentro de cada
medula dos animais dos grupos laminectomia, compressão e descompressão
que receberam veículo, NOC12-30 ou 300nM sete dias após os procedimentos
é mostrada na figura 10. Os grupos compressão e descompressão
apresentaram porcentagem de tecido preservado significativamente menor que
os grupos laminectomia, independente do tratamento farmacológico recebido
(efeito da lesão, veículo -F(2,14)=6,521, p=0,012; compressão - F(2,15)=8,458,
p=0,004; descompressão - F(2,13)=11,853, p=0,002, teste de Duncan, p<0,05.
Não houve efeito do tratamento (F(2,36)=0,672, p=0,517) ou interação entre
lesão e tratamento (F(2,36)=0,271, p=0,895).
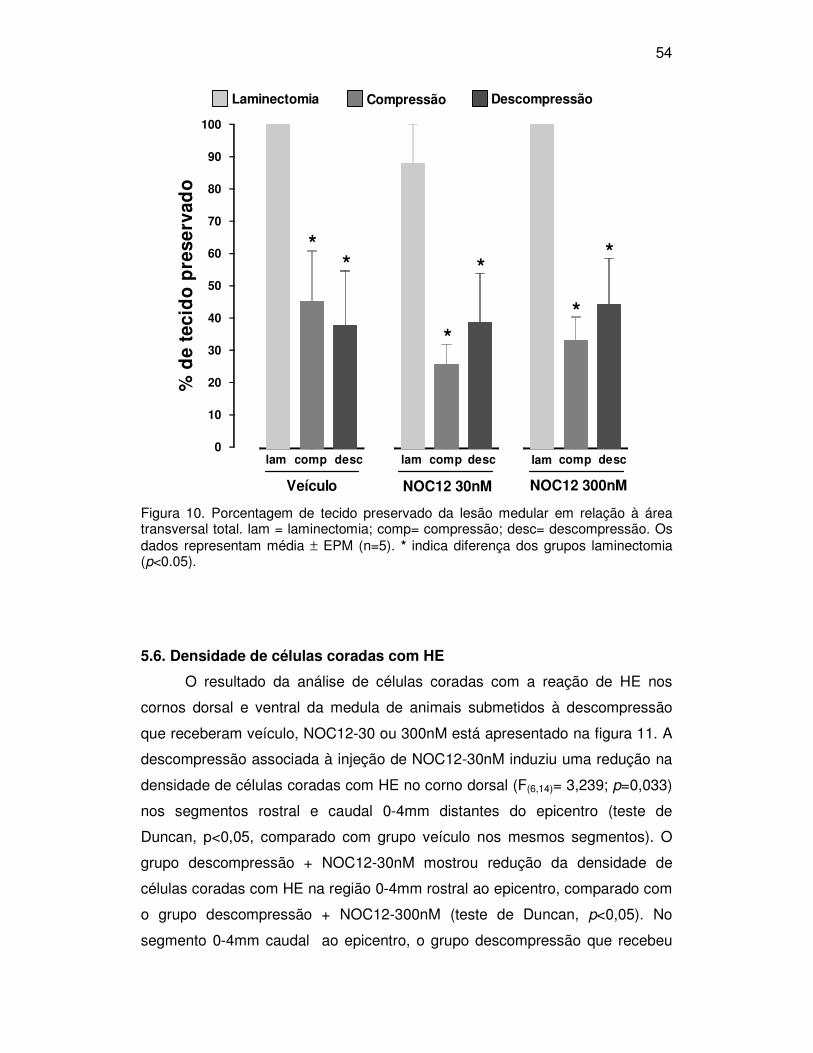
54
Veículo
0
10
20
30
40
50
60
70
80
90
100
% d
e te
cid
o p
rese
rvad
o
NOC12 30nM NOC12 300nM
lam lamcomp lamcomp compdesc desc desc
Laminectomia Compressão Descompressão
**
*
*
*
*
Figura 10. Porcentagem de tecido preservado da lesão medular em relação à área transversal total. lam = laminectomia; comp= compressão; desc= descompressão. Os dados representam média ± EPM (n=5). * indica diferença dos grupos laminectomia (p<0.05).
5.6. Densidade de células coradas com HE
O resultado da análise de células coradas com a reação de HE nos
cornos dorsal e ventral da medula de animais submetidos à descompressão
que receberam veículo, NOC12-30 ou 300nM está apresentado na figura 11. A
descompressão associada à injeção de NOC12-30nM induziu uma redução na
densidade de células coradas com HE no corno dorsal (F(6,14)= 3,239; p=0,033)
nos segmentos rostral e caudal 0-4mm distantes do epicentro (teste de
Duncan, p<0,05, comparado com grupo veículo nos mesmos segmentos). O
grupo descompressão + NOC12-30nM mostrou redução da densidade de
células coradas com HE na região 0-4mm rostral ao epicentro, comparado com
o grupo descompressão + NOC12-300nM (teste de Duncan, p<0,05). No
segmento 0-4mm caudal ao epicentro, o grupo descompressão que recebeu
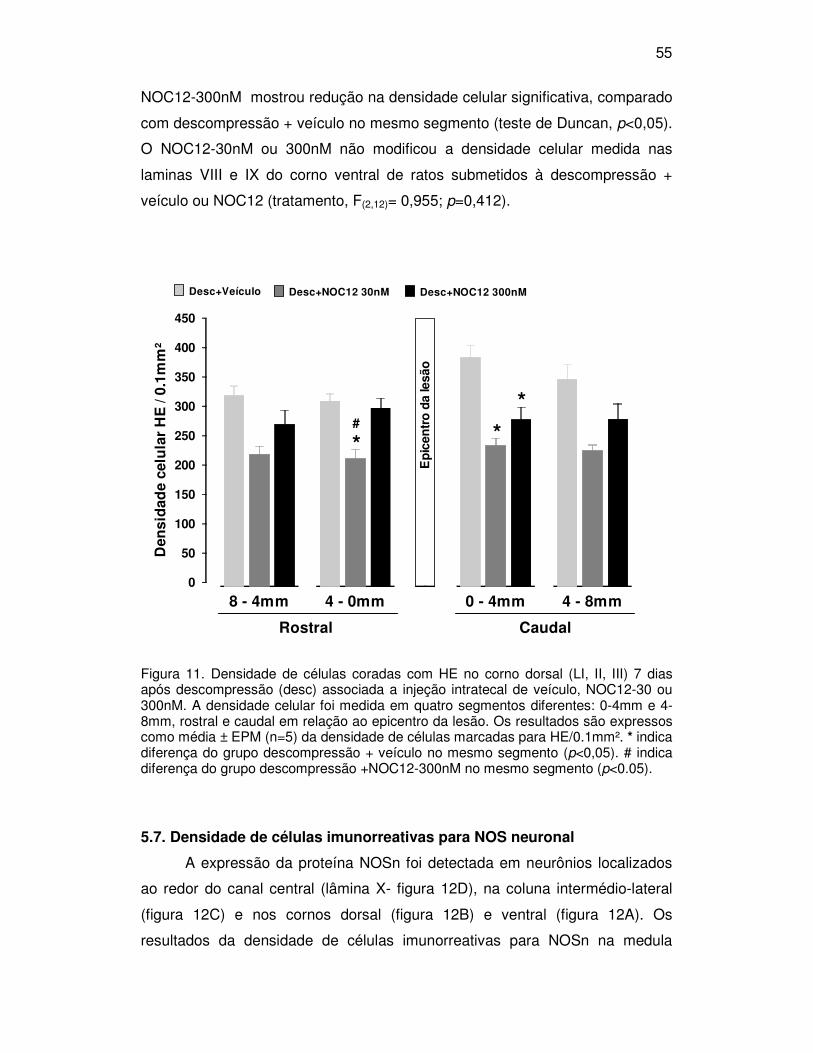
55
NOC12-300nM mostrou redução na densidade celular significativa, comparado
com descompressão + veículo no mesmo segmento (teste de Duncan, p<0,05).
O NOC12-30nM ou 300nM não modificou a densidade celular medida nas
laminas VIII e IX do corno ventral de ratos submetidos à descompressão +
veículo ou NOC12 (tratamento, F(2,12)= 0,955; p=0,412).
8 - 4mm0
50
100
150
200
250
300
350
400
450
Den
sid
ade
celu
lar
HE
/ 0.
1mm
²
4 - 0mm 0 - 4mm 4 - 8mm
Desc+Veículo Desc+NOC12 30nM Desc+NOC12 300nM
Ep
icen
tro
da
lesã
o
Rostral Caudal
***
#
Figura 11. Densidade de células coradas com HE no corno dorsal (LI, II, III) 7 dias após descompressão (desc) associada a injeção intratecal de veículo, NOC12-30 ou 300nM. A densidade celular foi medida em quatro segmentos diferentes: 0-4mm e 4-8mm, rostral e caudal em relação ao epicentro da lesão. Os resultados são expressos como média ± EPM (n=5) da densidade de células marcadas para HE/0.1mm². * indica diferença do grupo descompressão + veículo no mesmo segmento (p<0,05). # indica diferença do grupo descompressão +NOC12-300nM no mesmo segmento (p<0.05).
5.7. Densidade de células imunorreativas para NOS neuronal
A expressão da proteína NOSn foi detectada em neurônios localizados
ao redor do canal central (lâmina X- figura 12D), na coluna intermédio-lateral
(figura 12C) e nos cornos dorsal (figura 12B) e ventral (figura 12A). Os
resultados da densidade de células imunorreativas para NOSn na medula

56
espinal de animais dos grupos descompressão, tratados com veículo ou
NOC12, sacrificados 7 dias após o procedimento experimental são mostrados
na tabela 3.
A densidade de células NOSn+ nos cornos dorsal e ventral dos animais
submetidos a descompressão associada à microinjeção intrathecal de NOC12
não foi estatisticamente diferente dos animais que receberam veículo ( Dorsal:
droga, F(2,12)= 0,791; p=0,255; Ventral: droga, F(2,12)= 0,136; p=0,874).
Figura 12. Expressão da NOSn em neurônios da medula espinal. Fotomicrografia panorâmica da imunocitoquímica para NOSn da medula espinal (A). Neurônios NOSn+ estão presentes no corno ventral (seta sólida em B), no corno dorsal (setas sólidas em C), na coluna intermediolateral (IML – setas sólidas à esquerda em D) e ao redor do canal central (lâmina X – setas sólidas à direita em D), com projeções para a lâmina X (cabeças de seta em D). Em B as setas largas indicam neurônios negativos para NOSn no corno ventral da medula espinal.
57
* Resultados são expressos como média ± SEM (n=5) da densidade de células NOSn+ em 0.1mm².
3,5 ± 1,1 4,3 ± 1,7 2,4 ± 0,8 3,8 ± 0,9
Tabela 3. Densidade de células NOSn+ /mm² nos cornos dorsal e ventral da medula espinal de ratos após descompressão e injeção intratecal de de NOC12*.
Tratamento
Rostral
4-8mm 0-4mm 0-4mm 4-8mm
Veículo
NOC12 30nM
NOC12 300nM
2,9 ± 1,2 4,7 ± 0,5 3,8 ± 1,2 2,4 ± 1,2
2,9 ± 0,6 3,0 ± 1,0 2,7 ± 0,5 2,8 ± 1,3
Caudal
62,4 ± 4,7 70,6 ± 5,4 59,7 ± 4,5 66,3 ± 6,5
69,9 ± 9,6 68,2 ± 5,4 63,7 ± 3,1 70,1 ± 4,0
Corno Dorsal
Veículo
NOC12 30nM
NOC12 300nM
Corno Ventral
53,5 ± 3,7 53,7 ± 2,8 59,0 ± 3,0 64,5 ± 3,5
Região

58
6. DISCUSSÃO
Neste estudo foi investigado o efeito da compressão da medula espinal
de ratos por estreitamento do canal em 35%, seguida por descompressão
cirúrgica associada ou não à administração intratecal de NOC12. O
comportamento sensório-motor foi avaliado por meio da escala BBB e teste de
retirada de cauda. A resposta tecidual à lesão foi analisada por meio da análise
da coloração com HE e imunocitoquímica para NOSn.
Os resultados mostraram que o estreitamento do canal vertebral em
35% produziu déficit motor de moderado a grave, com melhora produzida por
meio da descompressão cirúrgica 6 horas após a lesão. A descompressão
associada ao doador de NO 24h após a lesão promoveu melhora na
recuperação motora e redução da extensão da lesão. A descompressão
acompanhada da aplicação de NOC12 não produziu modificação na expressão
da enzima NOS neuronal.
Por várias décadas a avaliação do comportamento locomotor de ratos
foi enfocada na escala criada por Tarlov e Klinger (1954) e suas modificações
ao longo dos anos. Em meados dos anos 90, Basso, Beattie e Bresnahan
desenvolveram uma nova escala de avaliação motora. Esta nova escala foi
criada com o intuito de ampliar a escala de Tarlov de 0-5 pontos para uma
escala de 0-21 pontos, avaliando um maior repertório de movimentos. A escala
BBB faz combinações de movimentos de modo a formar categorias que
representem os estágios seqüenciais de recuperação após lesão medular em
ratos (BASSO et al., 1996). Na avaliação do comportamento motor pela escala
BBB foi observada a habilidade motora do animal para caminhar livremente por
uma arena semelhante a do Teste do Campo Aberto, porém com dimensões
maiores. Este sistema de pontuação usa uma escala ordinal não linear onde a
parte inicial de recuperação concerne a aspectos grosseiros da locomoção,
enquanto as partes superiores incluem aspectos discretos da habilidade motora
dos animais (METZ et al., 2000). A escala poderia ser dividida em três fases: 1)
o ganho de movimentos das articulações dos membros posteriores, mas
incapacidade de caminhar; 2) capacidade de dar passadas e ganho de
qualidade na caminhada; e 3) ganho de posicionamento correto das patas e
cauda e estabilidade de tronco (BASSO; BEATTIE; BRESNAHAN, 1995). Além
disso, a escala BBB demonstrou correlação com o grau de lesão e

59
comprometimento histológico (BASSO; BEATTIE; BRESNAHAN, 1996); e alta
reprodutibilidade tanto intra quanto inter-observador (BASSO; BEATTIE;
BRESNAHAN, 1995, 1996; METZ et al., 2000). Desse modo, tornou-se muito
difundida e proporcionou parâmetro de comparação entre estudos de diferentes
laboratórios no estudo da lesão medular (DIMAR et al., 1999; FOUAD et al.,
2000 METZ et al., 2000; BROWN; WOLFE; WRATHALL, 2005; MALLEI et al.,
2005; SCHIAVETO DE SOUZA et al., 2006; YANG; KIM; LEE, 2007;
SAGANOVÁ et al., 2008).
É recomendado o uso de estatística paramétrica para avaliar os
resultados da escala BBB (BASSO; BEATTIE; BRESNAHAN, 1995, 1996). A
unificação de metodologia facilita a interpretação e correlação dos dados com
outros estudos (SCHEFF; SAUCIER; CAIN, 2002).
Os resultados da escala BBB neste estudo indicaram que o
estreitamento do canal espinal em 35% induz lesão medular em ratos (7-10
pontos). Estes resultados estão em oposição aos de Dimar et al. (1999) que
não observaram déficit motor após o estreitamento do canal espinal de 35% em
ratos. Neste estudo o déficit motor só foi encontrado após a associação do
estreitamento com contusão medular. Esta discrepância pode ser devida à
diferença de espécie utilizada para o estudo (Wistar x Sprage Dawley) ou às
dimensões do espaçador de 35% adequadas para ratos adultos Wistar
(SCHIAVETO DE SOUZA et al., 2006).
Estudo anterior em nosso laboratório padronizou as medidas de
espaçador para estreitamento de canal espinal de 35% e 50% para ratos
Wistar adultos. As medidas foram conseguidas a partir de medidas médias do
diâmetro ântero-posterior do canal vertebral de ratos de peso e idade
semelhantes (SCHIAVETO DE SOUZA et al., 2006). Utilizando a mesma
técnica deste estudo, Schiaveto de Souza (2004) e Nascimento et al. (não
publicado) também mostraram déficit locomotor na pontuação da escala BBB
no primeiro dia após estreitamento do canal espinal em 35%. Encontraram
ainda alta correlação entre os resultados da escala BBB e o teste do plano
inclinado.
O procedimento de descompressão cirúrgica 6 horas após a
compressão melhorou a recuperação motora dos animais, como mostrado pela
pontuação BBB. Os grupos compressão, independente do tratamento

60
farmacológico recebido, apresentaram pontuação BBB entre 7 (parte inicial da
escala) e 10, significando movimento das três articulações posteriores
preservado. A descompressão aumentou a pontuação BBB para 13-14, que
representa a capacidade de dar passos plantares com suporte de peso. A parte
média da escala (8-14 pontos) representa o ganho de posicionamento plantar
nas passadas e a coordenação entre os membros anteriores e posteriores,
fundamentais para uma caminhada de razoável a boa. Adicionalmente, a
associação da descompressão cirúrgica com o tratamento com NOC12 nas
doses 30 e 300nM levou a pontuação BBB destes grupos a valores
semelhantes aos do grupo laminectomia, onde não foi observado déficit. Este
efeito foi observado nos dias 5 e 7 pós-operatório.
Existe uma correlação positiva entre o estreitamento do canal espinal e a
severidade do déficit neurológico em humanos (MEVES; AVANZI, 2005). Tem
sido demonstrado que a descompressão precoce pode reduzir o grau de lesão
secundária em humanos (WAGNER; CHEHRAZI, 1982; SHIELDS et al., 2005),
coelhos (JARMUNDOWICZ et al., 1997) e cães (CARLSON et al., 2003a,
2003b; DELAMARTER; SHERMAN; CARR, 1995).
O efeito protetor atribuído à descompressão cirúrgica pode estar ligado à
redução do dano isquêmico. Segundo Zhang e Guth (1997) traumas medulares
lesam a vasculatura local, causando redução da perfusão apropriada do tecido,
o que é fundamental para a manutenção da estrutura e da função medular. Na
lesão medular por estreitamento do canal espinal os vasos permanecem
comprimidos. Dessa forma os tecidos adjacentes irrigados por estes vasos vão
sofrer enquanto durar a compressão dos vasos.
Em um trabalho prévio do nosso laboratório, Schiaveto de Souza et al.
(2006) mostraram que a lesão medular induzida tanto por contusão quanto por
estreitamento do canal espinal reduz a vascularização no sítio de lesão
proporcionalmente à intensidade da lesão. Carlson et al. (2003a,b)
encontraram retorno significativo do fluxo sanguíneo medular após a
descompressão em cães. Eles sugerem uma associação do mecanismo
isquêmico de lesão com o deslocamento sustentado do parênquima medular.
Durante a compressão sustentada da medula ocorre um relaxamento
viscoelástico do tecido e seu retorno se torna mais difícil quanto maior for o
período de compressão, levando a um aumento das modificações patológicas,

61
redução no retorno da atividade eletrofisiológica e maior prejuízo funcional.
Nosso estudo mostra que com a compressão tanto a substância cinzenta
quanto a substância branca sofrem deslocamento da região dorsal para central
e ventral, com conseqüente oclusão do canal central. Após a descompressão
há uma redução no deslocamento medular que ainda pode ser visualizada sete
dias após o procedimento.
A análise morfológica do tecido medular revelou ainda uma lesão com
aparência consistente com a descrição de outros estudos sobre contusão
medular. Estes descreveram que a substância cinzenta, em especial a região
central da medula, parece ser mais suscetível à lesão inicial, levando a áreas
de hemorragia, necrose, cavitação, invasão celular, aspecto esponjoso da
substância cinzenta e fragmentação da substância branca (BASSO; BEATTIE;
BRESNAHAN, 1996; SHARMA et al., 1996; BEATTIE et al., 1997; LIU et al.,
1997; CARLSON et al., 2003a). Sete dias após o estreitamento e
descompressão, foi observada a presença de infiltrado celular na região central
do tecido e espalhando-se radialmente. Nos grupos compressão onde houve
grande deformação das regiões analisadas e áreas de destruição tecidual, a
análise histológica das regiões muito próximas ao epicentro da lesão foi
dificultada.
A descompressão associada à administração intratecal de NOC12-30nM
induziu redução significativa na densidade de células marcadas pela coloração
de HE no corno dorsal próximo ao sítio de lesão (0-4mm). Este mesmo grupo
exibiu redução na extensão de lesão. Seria, então, plausível considerar que
esta redução possa ser devida a uma redução no número de células infiltrantes
nas lâminas superficiais de Rexed. Tem sido descrita a presença de neutrófilos,
macrófagos e células gliais em secções transversais de medula muitos dias
após a lesão medular (BASSO; BEATTIE; BRESNAHAN, 1996; LIU et al.,
1997; IADECOLA, 1997; KIGERL; MCGAUGHY; POPOVICH, 2006).
Várias medidas foram desenvolvidas para correlacionar a intensidade de
lesão e os danos teciduais na medula. Dentre elas estão medidas de volume
de lesão por ressonância magnética (CARLSON et al., 2003a), por
reconstrução tridimensional (BEATTIE et al., 1996; KIGERL; MCGAUGHY;
POPOVICH, 2006;), medidas de tecido medular poupado da lesão (BASSO;
BEATTIE; BRESNAHAN, 1996; KIGERL; MCGAUGHY; POPOVICH, 2006),

62
medidas da extensão longitudinal da lesão (KIGERL; MCGAUGHY;
POPOVICH, 2006), além de avaliações qualitativas de secções transversais e
longitudinais (DIMAR et al., 1999; BEATTIE et al., 1996).
Nos resultados da extensão total da lesão, ou seja, da soma das
extensões rostral e caudal, observamos que a descompressão per si não foi
capaz de reduzir significativamente a extensão longitudinal total da lesão
medular em ratos. Porém, a administração intratecal do doador de NO, NOC12,
24 horas após a descompressão reduziu a extensão da lesão total. Neste
estudo foi realizada medida da lesão para análise da sua progressão para as
regiões caudais e rostrais ao epicentro da lesão. Os resultados mostraram uma
redução significativa na extensão caudal da lesão nestes grupos, quando
comparados com seu veículo, o que não ocorreu para a região rostral. Uma
possível explicação para este fato seria um leve aumento observado na
extensão caudal da lesão no grupo descompressão + veículo. Foram
realizadas, ainda medidas de tecido poupado de lesão, a partir de secções
transversais no sítio da lesão. Os resultados não mostraram efeito da
descompressão ou descompressão associada ao uso de doador de NO
(comparado com os grupos compressão) como nos resultados anteriores.
Diferente dos nossos resultados, Carlson e colaboradores (2003a),
utilizando um modelo de compressão sustentada, encontraram redução do
volume de lesão inversamente proporcional ao tempo de compressão. A lesão
foi realizada por meio de pressão/peso mecanicamente controlado em cães
durante 30 ou 180 minutos. Neste estudo foi observado ainda que quanto maior
a duração da compressão, maior o volume de lesão e o prejuízo funcional.
Não existem dados de nosso conhecimento na literatura, que mostrem
resultados com relação à associação da descompressão cirúrgica com a
administração de doador de NO e a redução de extensão da lesão. No entanto
existem resultados interessantes referentes à aplicação isolada de
moduladores do sistema nitrérgico.
Já foi demonstrado que a administração sistêmica aguda ou subcrônica
de inibidores da NOS e doadores de NO após isquemia cerebral poderia induzir
neuroproteção. Zhang et al. (2001), encontrou uma melhora funcional e
neurogênese após a aplicação do doador de NO, (Z)-1-[N-(2-aminoetil)-N-(2-
amonioetil)aminio] diazen-1-ium-1,2-diolate (DETA/NONOate). A administração

63
foi realizada de forma aguda ou subcrônica (1h - 6 dias) na dose de 0,4mg/kg
por dia. Wainwright et al. (2007) demonstrou redução neuropatológica da lesão
com a administração i.p. de DETANONOate na dose de 0,4mg/kg, 1 – 24 horas
antes da lesão hipóxica-isquêmica. Redução no volume de lesão também foi
mostrado utilizando outro doador de NO, o 1-substituído-diazen-1-ium-1,2-
diolate [NONOate], ou ProliNO (PLUTA et al., 2001). A inibição da NOS
induzida pode reduzir o volume de infarto produzido pela oclusão da artéria
cerebral média (ZHANG; XU, 1995; ZHANG; IADECOLA, 1998; IADECOLA).
Em contraste, resultados semelhantes foram descritos após aplicação de Nω-
nitro-l-arginina methyl ester (L-NAME), um inibidor da NOS, na dose de
30mg/kg , 30 minutos antes da lesão.
Com relação ao papel do NO na lesão medular, os estudos mostram
resultados contraditórios. Hamada e colaboradores (1996) mostraram que o
tratamento subcrônico (5 minutos a 72 horas) com L-NAME, em doses baixas
(i.v.) auxilia na recuperação motora de ratos após lesão medular por
compressão. Em contraste, a injeção intravenosa de L-NAME em alta dose
(30mg/kg) 15 minutos antes da lesão inibiu a recuperação do distúrbio motor
pós-compressão medular (HAMADA et al., 1996). Além disso, Mallei e
colaboradores (2005) descreveram que um doador de NO derivado da
prednisolona, o NCX1015 (prednisolone 21-((4'-nitrooxymethyl)benzoate))
quando aplicado subcutâneo é capaz de melhorar a função motora de roedores
após lesão medular. Em conjunto estes resultados sugerem que os efeitos
protetores ou tóxicos do NO após lesão do SNC dependem do tempo de
exposição, da dose e da forma de administração dos doadores, precursores ou
inibidores de NO (PLUTA et al., 2001; ZHANG et al., 2001; WAINWRIGHT et
al., 2007).
Algumas teorias explicam o mecanismo de neuroproteção induzido por
NO pela facilitação do fluxo sanguíneo, limitação da excitotoxicidade e
seqüestro de espécies oxigênio-reativas (IADECOLA, 1997). Lipton e
colaboradores (LIPTON et al., 1993) forneceram evidências de que o papel do
NO dependeria do seu estado oxidativo. O NO+ reagiria com as proteínas
levando à regulação fisiológica no SNC. Enquanto, o NO- levaria a efeitos
neurotóxicos. Adicionalmente, a concentração do NO parece ser determinante
pra sua função biológica. Respostas celulares precisas são diferentemente

64
reguladas por concentrações específicas de NO. Thomas e colaboradores
(2008) propuseram níveis de concentração envolvendo diferentes atividades.
De modo geral, baixas concentrações promoveriam sobrevivência celular e
proliferação, enquanto altos níveis favoreceriam apoptose e senescência
celular. Um vasto número de fatores determina a formação do NO e sua
concentração, tais como, difusão, consumo e disponibilidade de substrato.
Estes são parâmetros químicos e bioquímicos que moldam as respostas
celulares ao NO.
Uma rota alternativa para o mecanismo protetor do NO, baseada na sua
interação com receptores NMDA, tem sido descrita como de vasta importância
(STAMLER et al., 2001). O óxido nitrico quando gerado nas sinapses do SNC
pela ativação de receptors NMDA, além de exercer efeitos fisiológicos pela
modificação nos níveis de GMPc, pode promover retroalimentação negativa
sobre os receptores NMDA. Isto ocorre por meio de uma modificação química
da função da proteína pela transferência de um grupamento NO para o grupo tiol
(S-H), formando um nitrosotiol (S-NO), mecanismo denominado S-nitrosação
(STAMLER et al., 2001). O receptor NMDA é um exemplo de proteína
alegadamente regulada por S-nitrosação (STAMLER et al., 2001; AHERN
et al., 2002; LIPTON et al., 2002). O resultado é a inibição da função do
receptor NMDA. Funcionalmente, este efeito inibitório serve para proteção
contra dano neuronal que pode resultar da superativação de receptores NMDA
(LEI et al., 1992; MANZONI et al., 1992).
Existem evidências de que a expressão de NOS pode ser modificada
pela lesão medular. A NOS é induzida em neurônios sensoriais do corno dorsal
após lesão da medula espinal ou lesão de nervos periféricos (MARSALA et al.,
1997; SHARMA et al., 1996; YU et al., 1994; WU, 1993). Hemisecção medular
ou lesão por impacto podem resultar em aumento da expressão de NOS em
interneurônios, especialmente na região rostral à lesão (VINCENT, 1994; WU,
2000; SHARMA; ALM, 2004). Xu e colaboradores (2000) demonstraram a
indução da expressão de NOS neuronal no nucleus dorsalis sete dias após a
hemissecção medular e observaram um pico 3 semanas após a lesão. Eles
ainda descreveram um aumento no número de neurônios positivos para NOS
neuronal com intensificação da marcação imunocitoquímica no nucleus dorsalis
ipsilateral à lesão após o tratamento com L-arginina, um precursor da NOS. A

65
imunorreatividade para NOS neuronal induzida pela lesão pode ser revertida
pela aplicação tópica de anticorpo contra NOS (SHARMA et al., 1996). Em
contraste, nosso estudo não detectou alteração significativa na expressão de
NOSn em neurônios da medula espinal após administração intratecal de
doador de NO em ratos submetidos à descompressão, comparado com
aqueles submetidos à injeção intratecal de veículo. Para avaliar se houve
alteração na expressão da NOSn após compressão e descompressão seria
necessário quantificar estes grupos.
Além do prejuízo motor, a lesão medular freqüentemente produz
paresia espástica e condições dolorosas (CHRISTENSEN; HULSEBOSCH,
1997; HAO et al., 1998). É sugerido que alterações na produção de NO após
transecção medular poderiam alterar a percepção da dor, produzindo dor
espontânea e hiper-reflexia motora e autonômica (TRUDRUNG; WIRTH;
MENSE, 2000; GONZALEZ et al., 2001). Acredita-se que o papel do NO na
hiperalgesia ocorra via GMP cíclico (HALEY et al., 1992; MELLER et al., 1994;
LUO; CIZKOVA, 2000; DUARTE; FERREIRA, 2000; SAKURADA et al., 2001).
Foi mostrado que doadores de óxido nitrico administrados intratecal ou
sistematicamente podem produzir hiperalgesia (MACHELSKA et al., 1998). No
entanto, existem estudos que apontam para a uma ação antinociceptiva do NO
(BUDZINSKI et al., 2000; JAIN et al., 2001).
Nesta pesquisa a função reflexa (reflexo nociceptivo) foi avaliada pelo
teste de retirada de cauda após exposição a estímulo térmico. O teste de
retirada de cauda tem sido utilizado para avaliar o reflexo nociceptivo após
lesão medular (ADVOKAT, 2002; SCHIAVETO DE SOUZA, 2004). Nossos
resultados não mostraram modificação na nocicepção de animais submetidos à
compressão ou descompressão. Resultado semelhante foi encontrado por
Merkler e colaboradores (2001), utilizando o mesmo teste após hemissecção
dorsal da medula espinal. Em nosso estudo, isto pode ser explicado analisando
a natureza anatômica do reflexo. O reflexo de retirada de cauda frente a um
estímulo térmico depende de um reflexo medular que envolve os segmentos
sacrais da medula espinal. Como a compressão medular foi realizada a nível
torácico, é provável que os segmentos responsáveis pelo reflexo de retirada de
cauda tenham sido mantidos intactos.

66
O circuito neural que participa na resposta ao teste de retirada de cauda
envolve a entrada de estímulos sensoriais do local de estímulo para neurônios
do corno dorsal da medula (podendo sofrer modulação por mediadores
periféricos) e seguem no sistema nervoso sob controle inibitório. Os três
maiores sistemas inibitórios utilizam glicina, ácido gama aminobutírico (GABA),
e peptídeos opióides endógenos como transmissores (WILLIS; COGGESHALL,
1991). Na via sensorial, o NO participa no processamento da entrada
nociceptiva (MELLER; GEBHART, 1993), bem como, na via descendente para
transmissão da resposta (FURST, 1999).
Nossos resultados mostraram ainda que a laminectomia associada à
aplicação intratecal de NOC12-30nM induziu uma hipernocicepção térmica
tardia. A hipernocicepção induzida por NO tem sido demonstrada, dentro de
poucas horas após a administração intratecal de doadores de NO (INOUE et
al., 1997; TAO; JOHNS, 2000; SOUZA; PRADO, 2001) ou inibidores da NOS
(FAIRBANKS et al., 2000; SOUSA; PRADO, 2001; TANG et al., 2007). Um
mecanismo potencial pelo qual o NO poderia ser pró-nociceptivo seria
modulando a sensibilização dos neurônios do corno dorsal da medula que
participam da transmissão nociceptiva para os neurônios dos centros
superiores, pela geração de GMPc e modulação da expressão gênica
(MILLAN, 1999). Estes resultados podem ser provenientes de uma interação
entre a intervenção cirúrgica e a adição de NO exógeno que modulariam os
mecanismos de nocicepção em longo prazo. A indução da nNOS no corno
dorsal aumentaria o NO produzido, e este participaria de respostas
nociceptivas prolongadas subseqüentes a estas lesões (WIESENFELD-HALLIN
et al., 1993). Existem evidências de que a indução da expressão de NOS
mediada por fibras do tipo C está envolvida na sensibilização de neurônios do
corno dorsal, os quais causam estados de dor devido a dano tecidual
inflamatório ou lesão de fibra aferente primária (MELLER; GEBHART, 1992;
MILLAN, 1999).
Jerosch-herold (2005) revisou a validade, confiabilidade e
responsividade dos testes de sensibilidade disponíveis para humanos. O teste
de monofilamento de Semmes–Weinstein foi um dos únicos testes que
alcançaram os critérios-padrão. Este teste também pode ser utilizado em
estudos experimentais utilizando roedores. Diferentemente do teste de retirada

67
de cauda, este não visa avaliar a nocicepção, mas a presença ou não de
alodinia (sensação dolorosa causada por uma estimulação não nociceptiva).
Consiste na aplicação perpendicular de monofilamentos de diferentes
gramaturas na região inferior da pata dos animais (estímulo inócuo). O sinal
positivo de alodinia é a reação de retirada da pata. Este teste vem sendo
utilizado na avaliação sensorial pós-lesão medular em roedores (referencia). O
teste do monofilamento não pôde ser aplicado fidedignamente neste estudo,
pois muitos animais com estreitamento de canal espinal estavam com os
membros posteriores completa ou parcialmente paraplégicos durante o período
de execução dos testes. Não podendo, assumir a posição plantar das patas
traseiras, necessária para a realização do teste. Desse modo, este teste não foi
incluído na análise do efeito da descompressão ou administração de NOC12.
No entanto, os animais foram cuidadosamente inspecionados e nenhum animal
apresentou sinais de autotomia (sinal de dor intensa que gera comportamento
de autofagia).
7. CONCLUSÕES
Este estudo demonstrou que:
� O estreitamento do canal espinal de 35% em ratos induz déficit motor e
que a descompressão aguda (6 horas) melhorou a recuperação neurológica;
� A descompressão associada à liberação de NO auxiliou a recuperação
neurológica como revelado pelo seletivo aumento na pontuação da BBB e pela
redução da extensão da lesão;
� Não houve modificação na densidade de células positivas para NOS
neuronal no corno ventral da medula espinal.
� Estes resultados indicam que a descompressão associada à liberação
aguda de NO promoveu melhora na recuperação após compressão medular
por estreitamento do canal vertebral em 35%.

68
REFERÊNCIAS
ADVOKAT, C. Spinal transection increases the potency of clonidine on the tail-flick and hindlimb flexion reflexes. Eur J Pharmacol, v. 437, n. 1-2, p. 63-67, 2002. AGUAYO, A. J.; DAVID, S.; BRAY, G. M. Influences of the glial environment on the elongation of axons after injury: transplantation studies in adult rodents. J Exp Biol, v. 95, p. 231-240, 1981. AHERN, G.P.; KLYACHKO, V. A.; JACKSON, M. B. cGMP and S-nitrosylation: two routes for modulation of neuronal excitability by NO. Trends Neurosci, v. 25, n. 10, p. 510-517, 2002. AMAR, A. P.; LEVY, M. L. Pathogenesis and pharmacological strategies for mitigating secondary damage in acute spinal cord injury. Neurosurgery, v. 44, n. 5, p. 1027-1039, 1999. ANDERSON, T. E; STOKES, B. T. Experimental models for spinal cord injury research: physical and physiological considerations. J Neurotrauma, v. 9, n. S. 1, p. S135-142, 1992. BASSO, D. M,; BEATTIE, M. S.; BRESNAHAN, J. C. Graded histological and locomotor outcomes after spinal cord contusion using the NYU weight-drop device versus transection. Exp Neurol, v. 139, n. 2, p. 244-256, 1996. BASSO, D. M.; BEATTIE, M. S.; BRESNAHAN, J. C. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma, v. 12, n. 1, p. 1-21, 1995. BASSO, D. M.; BEATTIE, M. S.; BRESNAHAN, J. C.; ANDERSON, D. K.; FADEN, A. I.; GRUNER, J. A.; HOLFORD, T. R.; HSU, C. Y.; NOBLE, L. J.; NOCKELS, P. L.; PEROT, P. L.; SALZMAN, S. K.; YOUNG, W. MASCIS evaluation of open field locomotor scores: effects of experience and teamwork on reliability. J Neurotrauma, v. 13, n. 7, p. 343-359, 1996. BEATTIE, M.S.; BRESNAHAN, J. C.; KOMON, J.; TOVAR, C. A.; VAN METER, M.; ANDERSON, D. K.; FADEN, A. I.; HSU, C. Y.; NOBLE, L. J.; SALZMAN, S.; YOUNG, W. Endogenous repair after spinal cord contusion injuries in the rat. Exp Neurol, v. 148, n. 2, p. 453-463, 1997. BELANGER, E.; LEVI, A. D. The acute and chronic management of spinal cord injury. J Am Coll Surg. v. 190, n. 5, p. 603-618, 2000. BISHOP, A.; ANDERSON, J. E. NO signaling in the CNS: from the physiological to the pathological. Toxicology, v. 208, p. 193-205, 2005.

69
BOYD, J. G.; GORDON, T. A dose-dependent facilitation and inhibition of peripheral nerve regeneration by brain-derived neurotrophic factor. Eur J Neurosci, v. 15, p. 613-626, 2002. BREDT, D. S. Nitric oxide signaling specificity – the heart of the problem. J Cell Sci, v. 116, p. 9-15, 2003. BREDT, D. S.; SNYDER, S. H. Nitric oxide as a neuronal messenger. TIPS, v. 12, p. 125-128, 1991. BROWN, K. M.; WOLFE, B. B.; WRATHALL, J. R. Rapid functional recovery after spinal cord injury in young rats. J Neurotrauma, v. 22, n. 5, p. 559-574, 2005. BUCHLI, A. D.; ROUILLER, E.; MUELLER, R.; DIETZ, V.; SCHWAB, M. E. Repair of the injured spinal cord: a joint approach of basic and clinical research. Neurodegener Dis, v. 4, n. 1, p. 51-56, 2007. BUDZINSK, M.; MISTEREK, K.; GUMULKA, W.; DOROCIAK, W. Inhibition of inducible nitric oxide synthase in persistent pain. Life Sci, v. 66, p. 301–305, 2000. BUNGE, M. B.; HOLETS, V. R.; BATES, M. L.; CLARKE, T. S.; WATSON, B. D. Characterization of photochemically induced spinal cord injury in the rat by light and electron microscopy. Exp Neurol, v. 127, n. 1, p. 76-93, 1994. CARLSON, G. D.; GORDEN, C. D.; NAKAZAWA, S.; WADA, E.; SMITH, J. S.; LAMANNA, J. C. Sustained spinal cord compression: part II: effect of methylprednisolone on regional blood flow and recovery of somatosensory evoked potentials. J Bone Joint Surg Am, v. 85-A, n. 1, p. 95-101, 2003. CARLSON, G. D.; GORDEN, C. D.; OLIFF, H. S.; PILLAI, J. J.; LAMANNA, J. C. Sustained spinal cord compression: part I: time-dependent effect on long-term pathophysiology. J Bone Joint Surg Am, v. 85-A, n. 1, p. 86-94, 2003. CHEN, T.Y.; DICKMAN, C. A.; ELERAKY, M.; SONNTAG, V. K. The role of decompression for acute incomplete cervical spinal cord injury in cervical spondylosis. Spine, v. 23, n. 22, p. 2398-2403, 1998. CHRISTENSEN, M. D.; HULSEBOSCH, C. E. Spinal cord injury and anti-NGF treatment results in changes in CGRP density and distribution in the dorsal horn in the rat. Exp Neurol, v. 147, n. 2, p. 463-475, 1997. D’AMOUR, F. E.; SMITH, D. L. A method for determining loss of pain sensation. J Pharmacol Ther, v. 72, p. 74-79, 1941. DAWSON, T. M.; DAWSON, V. L.; SNYDER, S. H. A novel neuronal messenger molecule in brain: the free radical, nitric oxide. Ann Neurol, v. 32, n. 3, p. 297-311, 1992.

70
DAWSON, V. L. Nitric oxide: role in neurotoxicity. Clin Exp Pharmacol Physiol, v. 22, n. 4, p. 305-308, 1995. DAWSON, V. L.; DAWSON, T. M. Nitric oxide neurotoxicity J Chem Neuroanat, v. 10, n.3-4, p. 179-190, 1996. DAWSON, V. L.; DAWSON, T. M.; LONDON, E. D.; BREDT, D. S.; SNYDER, S. H. Nitric oxide mediates glutamate neurotoxicity in primary cortical cultures (endothelium-derived relaxing factor/N-methyl-n-aspartate). Proc Natl Acad Sci USA, v. 88, p. 6368-6371, 1991. DELAMARTER, R. B.; SHERMAN, J.; CARR, J.B. Pathophysiology of spinal cord injury. Recovery after immediate and delayed decompression. J Bone Joint Surg Am, v. 77, n. 7, p. 1042-1049, 1995. DIMAR, J. R. 2ND; GLASSMAN, S. D.; RAQUE, G. H.; ZHANG, Y. P.; SHIELDS, C. B. The influence of spinal canal narrowing and timing of decompression on neurologic recovery after spinal cord contusion in a rat model. Spine, v. 24, n. 16, p. 1623-1633, 1999. DUARTE, I.D.; FERREIRA, S.D. L-NAME causes antinociception by stimulation of the arginine-NO-cGMP pathway. Mediators Inflamm, v.9, p. 25–30, 2000. DUH, M. S.; SHEPARD, M. J.; WILBERGER, J. E.; BRACKEN, M. B. The effectiveness of surgery on the treatment of acute spinal cord injury and its relation to pharmacological treatment. Neurosurgery, v. 35, n. 2, p. 240-248, 1994. ECHEVERRY, M. B.; GUIMARAES, F. S.; DEL BEL, E. A. Acute and delayed restraint stress-induced changes in nitric oxide producing neurons in limbic regions. Neuroscience, v. 125, n. 4, p. 981-993, 2004. EMERY, E.; ALDANA, P.; BUNGE, M. B.; PUCKETT, W.; SRINIVASAN, A.; KEANE, R. W.; BETHEA, J.; LEVI, A. D. Apoptosis after traumatic human spinal cord injury. J Neurosurg, v. 89, n. 6, p. 911-920, 1998. FAGNI, L.; BOCKAERT, J. Effects of nitric oxide on glutamate-gated channels and other ionic channels. J Chem Neuroanat, v.10, n. 3-4, p. 231-240, 1996. FAIRBANKS, C. A.; SCHREIBER, K. L.; BREWER, K. L.; YU, C. G.; STONE, L. S.; KITTO, K. F.; NGUYEN, H. O.; GROCHOLSKI, B. M.; SHOEMAN, D. W.; KEHL, L. J.; REGUNATHAN, S.; REIS, D. J.; YEZIERSKI, R. P.; WILCOX, G. L. Agmatine reverses pain induced by inflammation, neuropathy, and spinal cord injury. Proc Natl Acad Sci USA, v. 97, n. 19, p. 10584-10589, 2000. FAWCETT, J. W.; ASHER, R. A. The glial scar and central nervous system repair. Brain Res Bull, v. 49, n. 6, p. 377-391, 1999.

71
FEHLINGS, M. G.; PERRIN, R. G. The role and timing of early decompression for cervical spinal cord injury: update with a review of recent clinical evidence. Injury, v. 36, n. S.2, p. B13-26. 2005. FEHLINGS, M. G.; PERRIN, R. G. The timing of surgical intervention in the treatment of spinal cord injury: a systematic review of recent clinical evidence. Spine, v. 31, n. S.11, p. S28-35, 2006. FOUAD, K.; METZ, G. A.; MERKLER, D.; DIETZ, V.; SCHWAB, M. E. Treadmill training in incomplete spinal cord injured rats. Behav Brain Res, v. 115, n. 1, p. 107-113, 2000. FRIGON, A.; ROSSIGNOL, S. Functional plasticity following spinal cord lesions. Prog Brain Res, v. 157, p. 231-260, 2006. FÜRST, S. Transmitters involved in antinociception in the spinal cord. Brain Res Bull, v. 48, n. 2, p. 129-41, 1999. GAO, Y.; NIKULINA, E.; MELLADO, W.; FILBIN, M. T. Neurotrophins elevate cAMP to reach a threshold required to overcome inhibition by MAG through extracellular signal-regulated kinase dependent inhibition of phosphodiesterase. J. Neuroscience, v. 23, n. 37, p. 11770-11777, 2003. GARTHWAITE, J.; BOULTON, C. L. Nitric oxide signaling in the central nervous system. Annu Rev Physiol, v. 57, p. 683-706, 1995. GELLER, H. M.; FAWCETT, J. W. Building a bridge: engineering spinal cord repair. Exp Neurol, v. 174, n. 2, p. 125-136, 2002. GIGLIO, C. A.; DEFINO, H. L.; DA-SILVA, C. A.; DE-SOUZA, A. S.; DEL BEL, E. A. Behavioral and physiological methods for early quantitative assessment of spinal cord injury and prognosis in rats. Braz J Med Biol Res, v. 39, n. 12, p. 1613-1623, 2006. GONZÁLEZ, S.; LABOMBARDA, F.; GONZALEZ DENISELLE, M. C.; SARAVIA, F. E.; ROIG, P.; DE NICOLA, A. F. Glucocorticoid effects on Fos immunoreactivity and NADPH-diaphorase histochemical staining following spinal cord injury. Brain Res, v. 912, n. 2, p. 144-153, 2001. GRANDPRÉ, T.; NAKAMURA, F.; VARTANIAN, T.; STRITTMATTER, S. M. Identification of the Nogo inhibitor of axon regeneration as a reticulon protein. Nature, v. 403, p. 439-444, 2000. GRIFFITH, O. W.; STUEHR, D. J. Nitric oxide synthases: properties and catalytic mechanism. Annu Rev Physiol, v. 57, p. 707-736, 1995. GRISHAM, M. B.; JOURD'HEUIL, D.; WINK, D. A. Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites: implications in inflammation. Am J Physiol, v. 276, p. G315–G321, 1999.

72
HALEY, J. E.; DICKENSON, A. H.; SCHACHTER, M. Electrophysiological evidence for a role of nitric oxide in prolonged chemical nociception in the rat. Neuropharmacol, v. 31, p. 251–258, 1992. HALL, E. D.; SPRINGER, J. E. Neuroprotection and acute spinal cord injury: a reappraisal. NeuroRx, v. 1, n. 1, p. 80-100, 2004. HAMADA, Y.; IKATA, T.; KATOH, S.; TSUCHIYA, K.; NIWA, M.; TSUTSUMISHITA, Y.; FUKUZAWA, K. Roles of nitric oxide in compression injury of rat spinal cord. Free Radic Biol Med, v. 20, n. 1, p. 1-9, 1996. HAO, J. X.; XU, I. S.; WIESENFELD-HALLIN, Z.; XU, X. J. Anti-hyperalgesic and anti-allodynic effects of intrathecal nociceptin/orphanin FQ in rats after spinal cord injury, peripheral nerve injury and inflammation. Pain, v. 76, n. 3, p. 385-393, 1998. HOPPER, R.; LANCASTER, B.; GARTHWAITE, J. On the regulation of NMDA receptors by nitric oxide. Eur J Neurosci, v.19, n. 7, p. 1675-82, 2004. HOUWELING, D. A.; VAN ASSELDONK, J. T.; LANKHORST, A. J.; HAMERS, F. P.; MARTIN, D.; BÄR, P. R.; JOOSTEN, E. A.. Local application of collagen containing brain-derived neurotrophic factor decreases the loss of function after spinal cord injury in the adult rat. Neurosci Lett, v. 251, n. 3, p. 193-196, 1998. IADECOLA, C. Bright and dark sides of nitric oxide in ischemic brain injury. Trends Neurosci, v. 20, n. 3, p. 132-139, 1997. IADECOLA, C.; ZHANG, F.; XU, X. Inhibition of inducible nitric oxide synthase ameliorates cerebral ischemic damage. Am J Physiol, v. 268, n. 1Pt 2, p. 286-292, 1995. IGNARRO, L. J. Physiology and pathophysiology of nitric oxide. Kidney Int, v. 55, p. S2–S5, 1996. IGNARRO, L. J.; JACOBS, A., Nitric oxide synthase and production of nitric oxide. In STEINBUSCH, H. M. V.; DE VENTE, J.; VINCENT, S. R. (Ed.), Handbook of chemical neuroanatomy, v. 17: Functional neuroanatomy of the nitric oxide system. Elsevier Science B. V., Amsterdan, p. 1-17, 2000. INOUE, T.; MASHIMO, T.; SHIBUTA, S.; YOSHIYA, I. Intrathecal administration of a new nitric oxide donor, NOC-18, produces acute thermal hyperalgesia in the rat. J Neurol Sci, v. 153, p. 1-7, 1997. JARMUNDOWICZ, W.; TOSIK, D.; CHLEBINSKI, J.; GORKIEWICZ, Z. The effect of early decompression on the extent of changes in spinal cord microcirculation in experimental traumatic injury to the cord in rabbits. Neurol Neurochir Pol, v. 31, n. 6, p. 1167-1175, 1997 (resumo).

73
JEROSCH-HEROLD, C. Assessment of sensibility after nerve injury and repair: a systematic review of evidence for validity, reliability and responsiveness of tests. J Hand Surg [Br], v. 30, n. 3, p. 252-264, 2005. KEIRSTEAD, H. S.; DYER, J. K.; SHOLOMENKO, G. N.; MCGRAW, J. DELANEY, K. R.; STEEVES, J. D. Axonal regeneration and physiological activity following transection and immunological disruption of myelin within the hatchling chick spinal cord. J. Neuroscience, v. 15, n. 10, p. 6963-6974, 1995. KEIRSTEAD, H. S.; HASAN, S. J.; MUIR, G. D.; STEEVES, J. D. Supression of the onset of myelination extends the permissive period for the functional repair of embryonic spinal cord. Proc Natl Acad Sci USA, v. 89, p. 11664-11668, 1992. KHAN, M.; GRIEBEL, R. Acute spinal cord injury in the rat: comparison of three experimental techniques. Can.J.Neurol.Sci., v. 10, n.3, p. 161-165, Aug. 1983. KIGERL, K. A.; MCGAUGHY, V. M.; POPOVICH, P. G. Comparative analysis of lesion development and intraspinal inflammation in four strains of mice following spinal contusion injury. J Comp Neurol, v. 494, n. 4, p. 578-594, 2006. KWON, B. K.; TETZLAFF, W. Spinal cord regeneration: from gene to transplants. Spine, v. 26, n. S24, p. S13-22, 2001. LEI, S. Z.; PAN, Z. H.; AGGARWAL, S. K.; CHEN, H. S.; HARTMAN, J.; SUCHER, N. J.; LIPTON, S. A. Effect of nitric oxide production on the redox modulatory site of the NMDA receptor-channel complex. Neuron, v. 8, n. 6, p. 1087-1099, 1992. LIM, P. A.; TOW, A. M. Recovery and regeneration after spinal cord injury: a review and summary of recent literature. Ann Acad Med Singapore, v. 36, n. 1, p. 49-57, 2007. LIPTON, S. A.; CHOI, Y. B.; PAN, Z. H.; LEI, S. Z.; CHEN, H. S.; SUCHER, N. J.; LOSCALZO, J.; SINGEL, D. J.; STAMLER, J. S. A redox-based mechanism for the neuroprotective and neurodestructive effects of nitric oxide and related nitroso-compounds. Nature, v. 364, n. 6438, p. 626-632, 1993. LIPTON, S. A.; CHOI, Y. B.; SUCHER, N. J.; PAN, Z. H.; STAMLER, J. S. Redox state, NMDA receptors and NO-related species. Trends Pharmacol Sci, v. 17, n. 5, p. 186-187, 1996. LIPTON, S. A.; CHOI, Y. B.; TAKAHASHI, H.; ZHANG, D.; LI, W.; GODZIK, A.; BANKSTON, L. A. Cysteine regulation of protein function--as exemplified by NMDA-receptor modulation. Trends Neurosci, v. 25, n. 9, p. 474-480, 2002. LIU, F.; YOU, S. W.; YAO, L. P.; LIU, H. L.; JIAO, X. Y.; SHI, M.; ZHAO, Q. B.; JU, G. Secondary degeneration reduced by inosine after spinal cord injury in rats. Spinal Cord, v. 44, n. 7, p. 421-426, 2006.

74
LIU, X. Z.; XU, X. M.; HU, R.; DU, C.; ZHANG, S. X.; MCDONALD, J. W.; DONG, H. X.; WU, Y. J.; FAN, G. S.; JACQUIN, M. F.; HSU, C. Y.; CHOI, D. W. Neuronal and Glial Apoptosis after Traumatic Spinal Cord Injury. J Neurosci, v. 17, n. 14, p. 5395–5406, 1997. LU, J.; WAITE, P. Advances in spinal cord regeneration. Spine, v. 24, n. 9, p. 926-930, 1999. LUO, Z. D.; CIZKOVA, D. The role of nitric oxide in nociception. Curr Rev Pain, v. 4, p. 459–466, 2000. MA, M.; BASSO, D. M.; WALTERS, P.; STOKES, B. T.; JAKEMAN, L. B. Behavioral and histological outcomes following graded spinal cord contusion injury in the C57Bl/6 mouse. Exp Neurol, v. 169, n. 2, p. 239-254, 2001. MACHELSKA, H.; PRZEWLOCKI, R.; RADOMSKI, M. W.; PRZEWLOCKA, B. Differential effects of intrathecally and intracerebroventricularly administered nitric oxide donators on noxious mechanical and thermal stimulation. Pol J Pharmacol v. 50, p. 407–415, 1998 (abstr). MALLEI, A.; ADEN, S. A.; BACHIS, A.; BRANDOLI, C.; ONGINI, E.; MOCCHETTI, I. The nitrosteroid NCX1015, a prednisolone derivative, improves recovery of function in rats after spinal cord injury. Brain Res, v. 1062, n. 1-2, p. 16-25, 2005. MANZONI, O.; PREZEAU, L.; MARIN, P.; DESHAGER, S.; BOCKAERT, J.; FAGNI, L. Nitric oxide-induced blockade of NMDA receptors. Neuron, v. 8, n. 4, p. 653-662, 1992. MARSALA, J.; KLUCHOVA, D.; MARSALA, M. Spinal cord gray matter layers rich in NADPH diaphorase-positive neurons are refractory to ischemia-reperfusion-induced injury: a histochemical and silver impregnation study in rabbit. Exp Neurol, v. 145, n. 1, p. 165-179, 1997. MARSALA, J.; ORENDÁCOVÁ, J.; LUKÁCOVÁ, N.; VANICKÝ, I. Traumatic injury of the spinal cord and nitric oxide. Prog Brain Res, v. 161, p. 171-183, 2007. MEFFERT, M. K.; CALAKOS, N. C.; SCHELLER, R. H.; SCHULMAN, H. Nitric oxide modulates synaptic vesicle docking fusion reactions. Neuron, v. 16, n. 6, p. 1229-1236, 1996. MELLER, S. T.; CUMMINGS, C. P.; TRAUB, R. J.; GEBHART, G. F. The role of nitric oxide in the development and maintenance of the hyperalgesia produced by intraplantar injection of carrageenan in the rat. Neuroscience, v. 60, p. 347–367, 1994. MELLER, S. T.; GEBHART, G. F. Nitric oxide (NO) and nociceptive processing in the spinal cord. Pain, v. 52, n. 2, p. 127-136, 1993.

75
MELLER, S. T.; PECHMAN, P. S.; GEBHART, G. F.; MAVES, T. J. Nitric oxide mediates the thermal hyperalgesia produced in a model of neuropathic pain in the rat. Neuroscience, v. 50, n. 1, p. 7-10, 1992. MERKLER, D.; METZ, G. A. S.; RAINETEAU, O.; DIETZ, V.; SCHWAB, M. E.; FOUAD, K. Locomotor recovery in spinal cord-injured rats treated with an antibody neutralizing the myelin-associated neurite growth inhibitor nogo-A. J Neurosci, v. 21, n. 10, p. 3665–3673, 2001. METZ, G. A.; MERKLER, D.; DIETZ, V.; SCHWAB, M. E.; FOUAD, K. Efficient testing of motor function in spinal cord injured rats. Brain Res, v. 883, n. 2, p. 165-177, 2000. MEVES, R.; AVANZI, O. Correlation between neurological deficit and spinal canal compromise in 198 patients with thoracolumbar and lumbar fractures. Spine, v. 30, n. 7, p. 787-791, 2005. MILLAN, M. J. The induction of pain: an integrative review. Prog Neurobiol, v. 57, n. 1, p. 1-164, 1999. MOLANDER, C.; XU, Q.; RIVERO-MELIAN, C.; GRANT, G. Cytoarchitectonic organization of the spinal cord in the rat: II. The cervical and upper thoracic cord. J Comp Neurol, v. 289, n. 3, p. 375-385, 1989. MONCADA, S. Nitric oxide gas: mediator, modulator, and pathophysiologic entity. J Lab Clin Med, v. 120, n. 2, p. 187-191, 1992. MONCADA, S. Nitric oxide. J. Hipertens, v. 12, p. S35-S39, 1994. NAKAHARA, S.; YONE, K.; SETOGUCHI, T.; YAMAURA, I.; ARISHIMA, Y.; YOSHINO, S.; KOMIYA, S. Changes in nitric oxide and expression of nitric oxide synthase in spinal cord after acute traumatic injury in rats. J Neurotrauma, v. 19, n. 11, p. 1467-1474, 2002. NEMOTO, T.; SUZUKI T. S. T.; MORIYA, H.; NAKAYA, H. Inhibition of nitric oxide synthesis accelerates the recovery of polysynapitc reflex potentials after transient spinal cord ischemia in cats. Naunyn-Schmiedeberg’s Arch Pharmacol, v. 355, p. 447–451, 1997. NICHOLLS, J. G.; MARTIN, A. R.; WALLACE, B. G; FUCHS, P.A. From neuron to brain. Fourth edition, Sunderland, Sinauer associates, Inc, 2001. NOVIKOVA, L. N.; NOVIKOV, L. N.; KELLERTH, J. O. Differential effects of neurotrophins on neuronal survival and axonal regeneration after spinal cord injury in adult rats. J Comp Neurol, v. 452, n. 3, p. 255-263, 2002. NYBERG, F.; SHARMA, H. S. Repeated topical application of growth hormone attenuates blood-spinal cord barrier permeability and edema formation following spinal cord injury: an experimental study in the rat using Evans blue, ([125])I-sodium and lanthanum tracers. Amino Acids, v. 23, n. 1-3, p. 231-239, 2002.

76
NYSTRÖM, B.; BERGLUND, J. E. Spinal cord restitution following compression injuries in rats. Acta Neurol Scand, v. 78, n. 6, p. 467-472, 1988. PAGLIARO, P. Differential biological effects of products of nitric oxide (NO) synthase: it is not enough to say NO. Life Sci., v. 73, n.17, p. 2137-2149, 2003. PAPADOPOULOS, S. M.; SELDEN, N. R.; QUINT, D. J.; PATEL, N.; GILLESPIE, B.; GRUBE, S. Immediate spinal cord decompression for cervical spinal cord injury: feasibility and outcome. J Trauma, v. 52, n. 2, p. 323-332, 2002. PLEMEL, J. R.; DUNCAN, G.; CHEN, K. W.; SHANNON, C.; PARK, S.; SPARLING, J. S.; TETZLAFF. W. A graded forceps crush spinal cord injury model in mice. J Neurotrauma, v. 25, n. 4, p. 350-370, 2008. PLUTA, R. M.; RAK, R.; WINK, D. A.; WOODWARD, J. J.; KHALDI, A.; OLDFIELD, E. H.; WATSON, J. C. Effects of nitric oxide on reactive oxygen species production and infarction size after brain reperfusion injury. Neurosurgery, v. 48, n. 4, p. 884-892, 2001. POLLARD, M. E., APPLE, D. F. Factors associated with improved neurologic outcomes in patients with incomplete tetraplegia. Spine. v. 28, n. 1, p. 33-39, 2003. POULOS, T. L.; RAMAN, C. S.; LI, H. NO news is good news. Structure, v. 6, n. 3, p. 255-258, 1998. RADI, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc Natl Acad Sci U S A, v. 101, n. 12, p. 4003-4008, 2004. REIER, P. J. Cellular transplantation strategies for spinal cord injury and translational neurobiology. NeuroRx, v. 1, n. 4, p. 424-451, 2004. REUSS, M. H.; REUSS, S. Nitric oxide synthase neurons in the rodent spinal cord: distribution, relation to Substance P fibers, and effects of dorsal rhizotomy. J Chem Neuroanat, v. 21, n. 2, p. 181-196, 2001. RIVLIN, A. S.; TATOR, C. H. Objective clinical assessment of motor function after experimental spinal cord injury in the rat. J Neurosurg, v. 47, n. 4, p. 577-581, 1977. ROSENBERG, N.; LENGER, R.; WEISZ, I.; STEIN, H. Neurological deficit in a consecutive series of vertebral fracture patients with bony fragments within spinal canal. Spinal Cord, v. 35, p. 92-95, 1997. SAGANOVÁ, K.; ORENDÁCOVÁ, J.; CÍZKOVÁ, D.; VANICKÝ, I. Limited minocycline neuroprotection after balloon-compression spinal cord injury in the rat. Neurosci Lett, v. 433, n. 3, p. 246-249, 2008.

77
SAKURADA, C.; SUGIYAMA, A.; NAKAYAMA, M.; YONEZAWA, A.; SAKURADA, S.; TAN-NO, K.; KISARA, K.; SACURADA, T. Antinociceptive effect of spinally injected l-NAME on the acute nociceptive response induced by low concentrations of formalin. Neurochem Int, v. 38, p. 417–423, 2001. SCHEFF, S. W.; SAUCIER, D. A.; CAIN, M. E. A statistical method for analyzing rating scale data: the BBB locomotor score. J Neurotrauma, v. 19, n. 10, p. 1251-1260, 2002. SCHIAVETO DE SOUZA, A.; NEVES DO NASCIMENTO, L.; DA SILVA, C. A.; DEFINO, H. L. A.; DEL BEL, E. A. Vascular changes induced in the spinal cord by mechanical traumatism and permanent compression of the medullary canal in rats. Rev Bras Ortop, v. 41, n. 6, p. 221-226, 2006. SCHIAVETO-DE-SOUZA, A. Estudo da lesão medular em ratos produzida pelo estreitamento do canal vertebral. 2004. 219 f. Tese (Doutorado em Ciências) Departamento de Fisiologia, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 2004. SCHWAB, J. M.; BRECHTEL, K.; MUELLER, C. A.; FAILLI, V.; KAPS, H. P.; TULI, S. K.; SCHLUESENER, H. J. Experimental strategies to promote spinal cord regeneration--an integrative perspective. Prog Neurobiol, v. 78, n. 2, p. 91-116, 2006. SHARMA, H. S. Neuroprotective effects of neurotrophins and melanocortins in spinal cord injury: an experimental study in the rat using pharmacological and morphological approaches. Ann N Y Acad Sci, v. 1053, p. 407-421, 2005. SHARMA, H. S.; ALM. P. Role of nitric oxide on the blood-brain and the spinal cord barriers, em The Blood-Spinal Cord and Brain Barriers in Health and Disease. San Diego, Elsevier Academic Press, p. 191-230, 2004. SHARMA, H. S.; WESTMAN, J.; OLSSON, Y.; ALM, P. Involvement of nitric oxide in acute spinal cord injury: an immunocytochemical study using light and electron microscopy in the rat. Neurosci Res, v. 24, n. 4, p. 373-384, 1996. SHIELDS, C. B.; ZHANG, Y. P.; SHIELDS, L. B.; HAN, Y.; BURKE, D. A.; MAYER, N. W; The therapeutic window for spinal cord decompression in a rat spinal cord injury model. J Neurosurg Spine, v. 3, n. 4, p. 302-307, 2005. SNYDER, S. H. Nitric oxide: first in a new class of neurotransmitters. Science, v. 257, n. 5069, p. 494-496, 1992. SONNTAG, V. K.; DOUGLAS, R. A. Management of cervical spinal cord trauma. J Neurotrauma, v. 9, n. S1, p. S385-396, 1992. SOUSA, A. M.; PRADO, W. A. The dual effect of a nitric oxide donor in nociception. Brain Res, v. 897, p. 9-19, 2001.

78
SOUZA, A. S.; NASCIMENTO, L. N.; SILVA, C. A.; DEFINO, H. L. A.; DEL BEL, E. A. Estudo experimental das alterações vasculares da medula espinhal induzidas por traumatismo mecânico e compressão do canal vertebral. Rev Bras Ortop, v. 41, n. 6, p. 221-226, 2006. STAMLER, J. S.; LAMAS, S.; FANG, F. C. Nitrosylation. the prototypic redox-based signaling mechanism. Cell, v. 106, n. 6, p. 675-83, 2001. SUZUKI, T.; SEKIKAWA, T.; NEMOTO, T.; MORIYA, H.; NAKAYA, H. Effects of nicorandil on the recovery of reflex potentials after spinal cord ischaemia in cats. Br J Pharmacol, v. 116, n. 2, p. 1815-1820, 1995. TANG, Q.; SVENSSON, C. I.; FITZSIMMONS, B.; WEBB, M.; YAKSH, T. L.; HUA, X. Inhibition of spinal constitutive NOS-2 by 1400W attenuates tissue injury and inflammation-induced hyperalgesia and spinal p38 activation. Eur J Neurosci, v. 25, n. 10, p. 2964–2972, 2007. TAO, Y. X.; JOHNS, R. A. Activation of cGMP-dependent protein kinase Ialpha is required for N-methyl-D-aspartate- or nitric oxide-produced spinal thermal hyperalgesia. Eur J Pharmacol, v. 392, n. 3, p. 141-145, 2000. TARLOV, I. M.; KLINGER, H. Spinal cord compression studies. II. Time limits for recovery after acute compression in dogs. AMA Arch Neurol Psychiatry, v. 71, n. 3, p. 271-290, 1954. TATAGIBA, M.; BRÖSAMLE, C.; SCHWAB, M. E. Regenerationof injured axons in the adult mammalian central nervous system. Neurosurgery, v. 40, n. 3, p. 541-547, 1997. TATOR, C. H.; FEHLINGS, M. G. Review of the secondary injury theory of acute spinal cord trauma with emphasis on vascular mechanisms. J Neurosurg, v. 75, n. 1, p. 15-26, 1991. THOMAS, D. D.; RIDNOUR, L. A.; ISENBERG, J. S.; FLORES-SANTANA, W.; SWITZER, C. H.; DONZELLI, S.; HUSSAIN, P.; VECOLI, C.; PAOLOCCI, N.; AMBS, S.; COLTON, C. A.; HARRIS, C. C.; ROBERTS, D. D.; WINK, D. A. The chemical biology of nitric oxide: Implications in cellular signaling. Free Radic Biol Med, v. 45, n. 1, p. 18-31, 2008. TRUDRUNG, P.; WIRTH, U.; MENSE, S. Changes in the number of nitric oxide-synthesizing neurones on both sides of a chronic transection of the rat spinal cord. Neurosci Lett, v. 287, p. 125-128, 2000. VALLANCE, P. Nitric oxide: therapeutic opportunities. Fundam Clin Pharmacol, v. 17, n. 1, p. 1-10, 2003. VINCENT, S.R. Nitric oxide: a radical neurotransmitter in the central nervous system. Prog. Neurobiol, v. 42, p. 129-160, 1994.

79
WADA, S.; YONE, K.; ISHIDOU, Y.; NAGAMINE, T.; NAKAHARA, S.; NIIYAMA, T.; SAKOU, T. Apoptosis following spinal cord injury in rats and preventative effect of N-methyl-D-aspartate receptor antagonist. J Neurosurg, v. 91, n. S1, p. 98-104, 1999. WADA, S.; YONE, K.; ISHIDOU, Y.; NAGAMINE, T.; NAKAHARA, S.; NIIYAMA, T.; SAKOU, T. Apoptosis following spinal cord injury in rats and preventative effect of N-methyl-D-aspartate receptor antagonist. J Neurosurg, v. 91, n.1, p. S98-104, 1999. WAGNER, F. C. J. R,; CHEHRAZI, B. Early decompression and neurological outcome in acute cervical spinal cord injuries. J Neurosurg, v. 56, n. 5, p. 699-705, 1982. WAINWRIGHT, M. S.; GRUNDHOEFER, D.; SHARMA, S.; BLACK, S. M. A nitric oxide donor reduces brain injury and enhances recovery of cerebral blood flow after hypoxia-ischemia in the newborn rat. Neurosci Lett, v. 415, n. 2, p. 124-129, 2007. WIESENFELD-HALLIN, Z.; HAO, J. X.; XU, X. J.; HOKFELT, T. Nitric oxide mediates ongoing discharges in dorsal root ganglion cells after peripheral nerve injury. J Neurophysiol, v. 70, n. 6, p. 2350-2353, 1993. WILLIS, W. D.; COGGESHALL, R. E. Sensory mechanisms of spinal cord. 2ª ed. Plenum, New York, 1991. WINK, D. A.; HANBAUER, I.; GRISHAM, M. B.; LAVAL, F.; NIMS, R. W.; LAVAL, J.; COOK, J.C.; PACELLI, R.; LIEBMANN, J.; KRISHNA, M. C.; FORD, M. C.; JB, M. The chemical biology of NO: insights into regulation, protective and toxic mechanisms of nitric oxide. Curr Top Cell Regul, v.34, p. 159–187, 1996. WINK, D.A.; MITCHELL, J. B. The chemical biology of nitric oxide: insights into regulatory, cytotoxic and cytoprotective mechanisms of nitric oxide. Free Radic Biol Med, v. 25, p. 434–456, 1998. WOLF, G. Nitric oxide and Nitric oxide synthase: biology, pathology, localization. Histol. Histopathol., v. 12, p. 251-261, 1997. WRATHALL, J. R. Spinal cord injury models. J Neurotrauma, v. 9, n. S1, p. S129-34. WU, W. Expression of nitric oxide synthase (NOS) in injured CNS neurons as shown by NADPH-diaphorase histochemistry. Exp Neurol, v. 120, p. 153-159, 1993. WU, W. Response of nitric oxide synthase to neuronal injury, em Functional Neuroanatomy of the Nitric Oxide System. Handb Chem Neuroanat. Amsterdam/New York, Elsevier, v. 17, p. 315-353, 2000.

80
WU, W. Response of nitric oxide synthase to neuronal injury. . In STEINBUSCH, H. M. V.; DE VENTE, J.; VINCENT, S. R. (Ed.), Handbook of chemical neuroanatomy, v. 17: Functional neuroanatomy of the nitric oxide system. Elsevier Science B. V., Amsterdan, p. 315-353, 2000. YANG, J. Y.; KIM, H. S.; LEE, J. K. Changes in nitric oxide synthase expression in young and adult rats after spinal cord injury. Spinal Cord, v. 45, n. 11, p. 731-738, 2007. YANG, J. Y.; KIM, H. S.; LEE, J. K. Changes in nitric oxide synthase expression in young and adult rats after spinal cord injury. Spinal Cord, v. 45, n. 11, p. 731-738, 2007. YU, W. A. Nitric oxide synthase in motor neurons after axotomy. J Histochem Cytochem, v. 42, n. 4, p. 451-457, 1994. ZHANG, F.; IADECOLA, C. Temporal characteristics of the protective effect of aminoguanidine on cerebral ischemic damage. Brain Res, v. 802, n. 1-2, p. 104-110, 1998. ZHANG, R.; ZHANG, L.; ZHANG, Z.; WANG, Y.; LU, M.; LAPOINTE, M.; CHOPP, M. A. Nitric oxide donor induces neurogenesis and reduces functional deficits after stroke in rats. Ann Neurol, v. 50, n. 5, p. 602-611, 2001. ZHANG, Z.; GUTH, L. Experimental spinal cord injury: Wallerian degeneration in the dorsal column is followed by revascularization, glial proliferation, and nerve regeneration. Exp Neurol, v. 147, n. 1, p. 159-171, 1997.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo