ufrrjr1.ufrrj.br/adivaldofonseca/wp-content/uploads/2014/06/catro-a-a... · os quais nove como...
TRANSCRIPT

UFRRJ
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM
CIÊNCIAS VETERINÁRIAS
TESE
Distribuição e Longevidade de Larvas Infectantes
de Nematóides Gastrintestinais de Caprinos
(Capra hircus) em Solo e Pastagem Irrigados e Não-
irrigados no Município de Seropédica, RJ, Brasil
Abisair Andrade de Castro
2004

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
PARASITOLOGIA VETERINÁRIA
DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES DE NEMATÓIDES
GASTRINTESTINAIS DE CAPRINOS (Capra hircus) EM SOLO E PASTAGEM
IRRIGADOS E NÃO-IRRIGADOS NO MUNICÍPIO DE SEROPÉDICA, RJ, BRASIL
ABISAIR ANDRADE DE CASTRO
Sob a Orientação do Professor
Dr. Adivaldo Henrique da Fonseca
E Co-orientação do Professor
Dr. Laerte Grisi
Tese submetida como requisito parcial
para obtenção do grau de Philosophiae
Doctor em Ciências Veterinárias, Área
de concentração em Parasitologia
Veterinária.
Seropédica, RJ
Março de 2004

Verificar na Biblioteca Central.
...........
.....................
Castro, Abisair Andrade
Distribuição e longevidade de larvas infectantes de nematóides
gastrintestinais de caprinos (Capra hircus) em solo e pastagem irrigados e
não-irrigados no município de Seropédica, RJ, Brasil. Seropédica. RJ.
UFRRJ. Instituto de Veterinária. 2004.
xv. nº f: il..............................
Orientador: Adivaldo Henrique da Fonseca

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE VETERINÁRIA
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS –
PARASITOLOGIA VETERINÁRIA
ABISAIR ANDRADE DE CASTRO
Tese submetida ao Curso de Pós-Graduação em Ciências Veterinárias, como requisito
parcial para obtenção do grau de Philosophiae Doctor, em Ciências Veterinárias, área de
concentração em Parasitologia Veterinária.
TESE APROVADA EM 25/03/2004
Adivaldo Henrique da Fonseca. Dr. UFRRJ
(Orientador)
Laerte Grisi. Dr. UFRRJ
Maria de Lurdes de Azevedo Rodrigues. Dra. UFRRJ
Débora Henrique da Silva Anjos. Dra. UFRJ
Denise Botelho de Oliveira Braga. Dra. UFF

“O temor ao Senhor é o princípio da ciência... A sabedoria é cousa principal: adquire pois a sabedoria; sim, com tudo o que possuis adquire o conhecimento. Exalta-a, e ela te exaltará; e abraçando-a tu, ela te honrará. Dará à tua cabeça um diadema de graça, e uma coroa de glória te entregará.” (Provérbios, 1:7; 4:7-9.)

A Deus, pois está escrito...
“... sem Mim, nada podeis fazer.” (João 15:5)
Ao meu esposo, João Carlos,
pelo apoio, compreensão e carinho...
Aos meus pais, Othniel e Maria Auxiliadora, cujos ensinamentos são minha maior e mais rica herança...
Dedico...
Amo vocês.

AGRADECIMENTOS
Primeiramente, a Deus, por ter me dado condições físicas e psicológicas, e ainda
por proporcionar os meios para o desenvolvimento e término deste trabalho.
Ao prof. Dr. Adivaldo Henrique da Fonseca pela amizade, apoio e orientação
deste estudo.
Ao pesquisador Dr. Manoel Pimentel Neto pelo carinho, amizade e orientação
na escolha do tema deste estudo.
Ao meu esposo, João Carlos; aos meus pais, Othniel e Maria Auxiliadora; ao
meu irmão, Asriel, a minha cunhada, Cristiane, e ao pequeno Abhner Lucas por estarem
sempre ao meu lado.
A amiga M. Sc.Erika Izaac de Ornelas por todo incentivo, apoio e ajuda na parte
inicial e ao longo deste estudo.
A amiga e futura Dra. Cátia Marques da Costa pela assistência no cuidado com
os animais em várias ocasiões, e pelos valiosos e enriquecedores momentos
compartilhados no dia a dia.
Aos mestrandos Daniel S. Guedes Junior e Carlo J. F. Oliveira pela amizade,
disponibilidade e ajuda em vários momentos.
Aos bolsistas de iniciação científica Fábio J. M. Silva e Patrícia F. Magalhães
pelo importante auxílio durante a fase final deste estudo.
A doutoranda Alessandra Scofield pela amizade e grandiosa ajuda no
processamento das imagens.
Ao técnico de laboratório Luiz Carlos Ribeiro da Paz e ao estagiário André Luis
C. de Azevedo pela amizade e toda ajuda disponibilizada em vários momentos.
Aos demais colegas do Laboratório de Doenças Parasitárias - Projeto Sanidade
Animal/ Embrapa: Isis Abel, Renata C. Madureira, Nathalie C. da Cunha, Luciana de
Almeida, Raquel Lisboa, Jaime da S. Pena, Fabíola N. Corrêa, e Charles P. Rangel pela
amizade, apoio e por compartilharem de muitos momentos na realização deste trabalho.
A professora Dra Maria de Lurdes de Azevedo Rodrigues pela amizade,
incentivo e preciosos conselhos, nos primeiros passos da minha carreira profissional.
A professora Dra. Débora H. Anjos pela amizade desde os tempos de
graduação...

Ao professor Dr. Rafael de La Veja Ruíbal pelo apoio técnico na parte estatística
deste estudo e ao técnico veterinário Andrés Carrejo Hernández pelos momentos
compartilhados na fase final deste estudo.
A Sra. Sandra Sanchez, coordenadora do setor de agropecuária do Colégio
Técnico da Universidade Rural, por gentilmente ceder os animais para realização do
presente estudo.
Ao Sr. Josué L. Castro, responsável pelo setor de caprinocultura do Colégio
Técnico da Universidade Rural, pelas orientações práticas no cuidado com os animais.
Ao professor e químico Wanderley M. Passos por fornecer o detergente não-
iônico, utilizado neste estudo.
Ao engenheiro agrônomo Ednaldo Silva por elaborar o projeto de irrigação.
A todos os funcionários e técnicos do Prédio de Sanidade Animal,
PSA/Embrapa, pela amizade demonstrada ao longo do curso.
A todos os professores do Curso de Pós-Graduação em Ciências Veterinárias -
Parasitologia Veterinária, e ainda, a todos aqueles que contribuíram direta ou
indiretamente na elaboração deste estudo.
Ao CNPq e FAPERJ pelo auxílio financeiro.

BIOGRAFIA
ABISAIR ANDRADE DE CASTRO, filha de Othniel Rodrigues de Castro e Maria
Auxiliadora da Silva Andrade de Castro, nasceu em 25 de março de 1973, na cidade do
Rio de Janeiro, (R.J.) onde cursou o ensino fundamental na Escola Municipal Pracinha
João da Silva. Concluiu o curso normal em 1990, ao nível de ensino médio, no Instituto
de Educação Sarah Kubitschek.
Ingressou no curso de Ciências Físicas e Biológicas em 1993, na Universidade
Federal Rural do Rio de Janeiro, onde obteve os graus de Licenciatura e Bacharelado
em 1997 e 1998, respectivamente.
Foi bolsista de Iniciação Científica do Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq), no período de 01/08/1995 a 31/12/1997, participando
de trabalhos de pesquisa no Laboratório de Helmintologia da Estação para Pesquisas
Parasitológicas W. O. Neitz, sob orientação da Profa Dra Maria de Lurdes de Azevedo
Rodrigues, do Departamento de Parasitologia Animal, Instituto de Veterinária, da
Universidade Federal Rural do Rio de Janeiro.
Em março de 2000, defendeu a dissertação de Mestrado, também sob orientação
da Dra. Maria de Lurdes de Azevedo Rodrigues, pelo Curso de Pós-graduação em
Medicina Veterinária – Parasitologia Veterinária, do DPA/IB-UFRRJ; tendo sido no
mesmo ano, aprovada e matriculada para o Doutorado pelo Curso de Pós-graduação em
Ciências Veterinárias – Parasitologia Veterinária, CPGCV-PV, da UFRRJ, agora sob
orientação do Dr. Adivaldo Henrique da Fonseca.
Durante o período acadêmico registraram-se 54 publicações científicas, dentre
os quais nove como artigos em revistas científicas indexadas e 45 em congressos e
eventos científicos nacionais e internacionais. E nesta data, apresenta e defende esta tese
como requisito parcial para a obtenção do título de Philosophiae Doctor.

SUMÁRIO
Pág.
1. INTRODUÇÃO................................................................................................................... 1
2. REVISÃO DE LITERATURA................................................................................................. 4
2.1 TÉCNICAS PARA RECUPERAÇÃO DE LARVAS INFECTANTES DA PASTAGEM.............. 4
2.2 DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES NA PASTAGEM, NO
EXTERIOR......................................................................................................................
8
2.2 DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES NA PASTAGEM, NO
BRASIL..........................................................................................................................
15
2.3 ESTUDOS COM PASTAGEM IRRIGADA………………………………...................... 16
2.4 DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES NO SOLO....................... 18
3. CAPÍTULO I: “COMPARAÇÃO ENTRE AS TÉCNICAS DE BAERMANN MODIFICADA E
DONALD UTILIZADAS PARA RECUPERAR LARVAS INFECTANTES DE NEMATÓIDES
GASTRINTESTINAIS DE RUMINANTES DA PASTAGEM”............................................................
21
RESUMO............................................................................................................................... 22
ABSTRACT............................................................................................................................ 23
3.1. INTRODUÇÃO........................................................................................................ 24
3.2. MATERIAL E MÉTODOS......................................................................................... 25
3.2.1. LOCAL...................................................................................................... 25
3.2.2. AMOSTRAS................................................................................................ 25
3.2.3. COLETA DE DADOS.................................................................................... 25
3.2.4. DESCRIÇÃO DA TÉCNICA DE BAERMANN................................................... 25
3.2.5. DESCRIÇÃO DA TÉCNICA DE DONALD........................................................ 26
3.2.6. ANÁLISE ESTATÍSTICA............................................................................... 26
3.3. RESULTADOS E DISCUSSÃO................................................................................... 28
3.4. CONCLUSÕES......................................................................................................... 32

4. CAPÍTULO II: “DISTRIBUIÇÃO SAZONAL E LONGEVIDADE DE LARVAS INFECTANTES DE
NEMATÓIDES GASTRINTESTINAIS DE CAPRINOS (Capra hircus) EM PASTAGEM IRRIGADA E
NÃO-IRRIGADA”....................................................................................................................
33
RESUMO............................................................................................................................... 33
ABSTRACT............................................................................................................................ 34
4.1. INTRODUÇÃO......................................................................................................... 35
4.2. MATERIAL E MÉTODOS.......................................................................................... 38
4.2.1. LOCAL....................................................................................................... 38
4.2.2. ANIMAIS E AMOSTRAS DE FEZES................................................................ 38
4.2.3. CLIMA....................................................................................................... 38
4.2.4. PIQUETES................................................................................................... 38
4.2.5. COLETA DE DADOS.................................................................................... 40
4.2.6. ESTIMATIVA DOS DADOS EM MATÉRIA SECA.............................................. 40
4.2.7. ANÁLISE ESTATÍSTICA............................................................................... 40
4.3. RESULTADOS E DISCUSSÃO................................................................................... 43
4.3.1. DADOS APRESENTADOS MÊS A MÊS........................................................... 49
4.3.2. DADOS APRESENTADOS EM QUATRO ESTAÇÕES......................................... 56
4.3.3.DADOS APRESENTADOS NOS PERÍODOS CHUVOSO E SECO........................... 59
4.4. CONCLUSÕES......................................................................................................... 63
5. CAPÍTULO III: “DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES DE
NEMATÓIDES GASTRINTESTINAIS DE CAPRINOS (Capra hircus) EM SOLO E PASTAGEM
IRRIGADOS E NÃO-IRRIGADOS”.............................................................................................
65
RESUMO............................................................................................................................... 66
ABSTRACT............................................................................................................................ 67
5.1. INTRODUÇÃO......................................................................................................... 68
5.2. MATERIAL E MÉTODOS......................................................................................... 69
5.2.1. LOCAL....................................................................................................... 69
5.2.2. ANIMAIS E AMOSTRAS DE FEZES................................................................ 69
5.2.3. CLIMA E SOLO............................................................................................ 69
5.2.4. PIQUETES................................................................................................... 69
5.2.5. COLETA DE DADOS.................................................................................... 69
5.2.6. ESTIMATIVA DOS DADOS EM MATÉRIA SECA.............................................. 70

5.2.7. ANÁLISE ESTATÍSTICA............................................................................... 70
5.3. RESULTADOS E DISCUSSÃO................................................................................... 73
5.3.1. DADOS APRESENTADOS MÊS A MÊS........................................................... 75
5.3.2. DADOS APRESENTADOS EM QUATRO ESTAÇÕES......................................... 79
5.3.3.DADOS APRESENTADOS NOS PERÍODOS CHUVOSO E SECO........................... 83
5.4. CONCLUSÕES........................................................................................................ 87
6. REFERÊNCIAS BIBLIOGRÁFICAS........................................................................................ 88

LISTA DE FIGURAS
FIGURAS Pág.
1. Técnica de Baermann processando gramínea.................................................................. 27
2. Etapas da técnica de Donald; A - início da etapa de lavagem; B - cálices de
decantação; C - funis de separação.................................................................................
27
3. Número total de larvas infectantes de nematóides gastrintestinais de bovinos, por
kg/pasto, recuperadas pelas técnicas de Baermann e Donald..............................................
31
4. A – coletor plástico cortado em forma de bisel recebendo glicerina para facilitar a
coleta; B – amostras de fezes sendo coletadas diretamente do reto com auxílio de
coletor..................................................................................................................................
39
5. A - Piquetes de experimento; A () piquete não-irrigado, “A” com amostras; B ()
piquete irrigado, “B” com amostras....................................................................................
39
6. Coprocultura, técnica utilizada para obtenção de larvas infectantes. () Setas
indicam as L3 migrando pela parede do copo de vidro.......................................................
42
7. Larvas infectantes encontradas no presente estudo. São respectivamente: A e B:
extremidade anterior e posterior de Trichostrongylus sp.; C e D: extremidade anterior e
posterior de Haemonchus sp. e E e F: extremidade anterior e posterior de
Oesophagostomum sp..........................................................................................................
44
8: Curvas gráficas dos números de L3/kg de pasto de Trichostrongylus, Haemonchus e
Oesophagostomum, apresentados à esquerda, em matéria seca (ms) e à direita, em
matéria verde (mv)..............................................................................................................
46

9: A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos entre
os piquetes A, não-irrigado e B, irrigado. B. Longevidade de larvas infectantes de
nematóides gastrintestinais de caprinos entre os piquetes A, não-irrigado e B,
irrigado................................................................................................................................
47
10. Pastagens: A Amostra no piquete A, não-irrigado; B. Amostra no piquete B, irrigado 48
11: Estimativa do número de fêmeas, do número de ovos por gênero, distribuição de
larvas por kg/ms nos piquetes A e B, para Trichostrongylus e Haemonchus.....................
50
12: Dados climáticos. Distribuição por mês, do número larvas infectantes de
Trichostrongylus, Haemonchus e Oesophagostomum por Kg/ms e seus respectivos
dados transformados, em Seropédica..................................................................................
52
13: Dados climáticos. Longevidade por mês, do número larvas infectantes de
Trichostrongylus, Haemonchus e Oesophagostomum em semanas e seus respectivos
dados transformados, em Seropédica..................................................................................
53
14: A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos em
quatro estações. B. Longevidade de larvas infectantes de nematóides gastrintestinais de
caprinos em quatro estações. (Letras, minúsculas, idênticas não diferem ao nível de 5%
de probabilidade.)................................................................................................................
57
15. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, em quatro estações, obtidos na Estação experimental da
Pesagro-Rio, Seropédica, RJ................................................................................................
58
16. A. Distribuição nos períodos chuvoso e seco, de larvas infectantes de nematóides
gastrintestinais de caprinos, expressos número de L3/Kg matéria seca e em logaritmo do
número de L3. B. Longevidade nos períodos chuvoso e seco, de larvas infectantes de
nematóides gastrintestinais de caprinos expressos em semanas e seus respectivos dados
transformados em logaritmo (Letras, minúsculas, idênticas não diferem ao nível de 5%
de probabilidade.).................................................................................................................
60

17. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, nos períodos chuvoso e seco, obtidos na Estação
experimental da Pesagro-Rio, Seropédica, RJ.....................................................................
61
18. Trado, equipamento utizado para coleta de solo. () Seta indica o comprimento de
10cm; limite de profundidade utilizada no experimento......................................................
72
19. Etapas da técnica de Baermann, utilizada para processamento de amostras do solo.
A – funis com água destilada aquecida; B – amostras de solo sendo processadas..............
72
20. A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos, no
pasto e solo, expressos em número de L3/Kg de matéria seca e em Kg de solo. B.
Longevidade de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto e
solo, expressos em semanas. (Letras, minúsculas, distintas diferem ao nível de 5% de
probabilidade.).....................................................................................................................
74
21. A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos, no
pasto, expressos em número de L3/Kg de matéria seca e em logaritmo do número de L3.
B. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos, no solo,
expressos em número de L3/Kg e em logaritmo do número de L3 (Letras, minúsculas,
idênticas não diferem ao nível de 5% de probabilidade.)....................................................
76
22. A. Longevidade de larvas infectantes de nematóides gastrintestinais de caprinos, no
pasto, expressos em semanas e em logaritmo do número de semanas. B. Longevidade de
larvas infectantes de nematóides gastrintestinais de caprinos, no solo, expressos em
semanas e em logaritmo do número de semanas (Letras, minúsculas, idênticas não
diferem ao nível de 5% de probabilidade.)..........................................................................
77
23. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, obtidos na Estação experimental da Pesagro-Rio, Sero., RJ.
78

24. A. Distribuição, nas quatro estações, de larvas infectantes de nematóides
gastrintestinais de caprinos, no pasto, expressos número de L3/Kg matéria seca e em
logaritmo do número de L3. B. Distribuição, nas quatro estações, de larvas infectantes
de nematóides gastrintestinais de caprinos, no solo, expressos em número de L3/Kg
matéria seca e em logaritmo do número de L3 (Letras, minúsculas, idênticas não
diferem ao nível de 5% de probabilidade.)..........................................................................
80
25. A. Longevidade, nas quatro estações, de larvas infectantes de nematóides
gastrintestinais de caprinos, no pasto, expressos em semanas e em logaritmo do número
de semanas. B. Longevidade, nas quatro estações, de larvas infectantes de nematóides
gastrintestinais de caprinos, no solo, expressos em semanas e em logaritmo do número
de semanas (Letras, minúsculas, idênticas não diferem ao nível de 5% de
probabilidade.).....................................................................................................................
81
26. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, em quatro estações, obtidos na Estação experimental da
Pesagro-Rio, Seropédica, RJ...............................................................................................
82
27. A. Distribuição nos períodos chuvoso e seco, de larvas infectantes de nematóides
gastrintestinais de caprinos, no pasto, expressos número de L3/Kg matéria seca e em
logaritmo do número de L3. B. Distribuição nos períodos chuvoso e seco, de larvas
infectantes de nematóides gastrintestinais de caprinos, no solo, expressos em número de
L3/Kg matéria seca e em logaritmo do número de L3 (Letras, minúsculas, idênticas não
diferem ao nível de 5% de probabilidade.)..........................................................................
84
28. A. Longevidade, nos períodos chuvoso e seco, de larvas infectantes de nematóides
gastrintestinais de caprinos, no pasto, expressos em semanas e em logaritmo do número
de semanas. B. Longevidade, nos períodos chuvoso e seco, de larvas infectantes de
nematóides gastrintestinais de caprinos, no solo, expressos em semanas e em logaritmo
do número de semanas (Letras, minúsculas, idênticas não diferem ao nível de 5% de
probabilidade.).....................................................................................................................
85

29. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, nos períodos chuvoso e seco, obtidos na Estação
experimental da Pesagro-Rio, Seropédica, RJ.....................................................................
86

LISTA DE TABELAS
TABELAS Pág.
1: Número das amostras, número total e média de larvas infectantes de nematóides
gastrintestinais de Haemonchus sp e Oesophagostomum sp, recuperadas pelas
técnicas de Baermann e Donald, 21 e 28 dias após o depósito.......................................
29
2: Números médios de larvas infectantes de Trichostrongylus e Haemonchus
recuperadas; período de sobrevivência por mês e seus respectivos valores
transformados.................................................................................................................
51

xvii
RESUMO
CASTRO, Abisair Andrade de. Distribuição e Longevidade de larvas infectantes de
nematóides gastrintestinais de caprinos (Capra hircus) em pastagem no município
de Seropédica, RJ, Brasil. Seropédica: UFRRJ, 2004. 97p. (Tese – Doutorado em
Ciências Veterinárias, Parasitologia Veterinária)
O conhecimento do grau de contaminação das pastagens por larvas infectantes de
trichostrongilídeos é muito útil aos propósitos epidemiológicos. Pode-se determinar o
risco de infecção dos animais e fornecer dados para o estabelecimento de programas de
controle integrado. Assim este estudo foi elaborado em três partes: a primeira diz
respeito a um estudo comparativo entre duas técnicas para recuperar larvas infectantes
da pastagem. Para isto, seis amostras de fezes naturalmente infectadas foram
depositadas a campo. Após 21 e 28 dias ao depósito, amostras de gramínea foram
coletadas e processadas pelas técnicas propostas. A análise estatística demonstrou que
não houve diferença significativa entre os resultados. Assim, as duas técnicas podem ser
utilizadas para recuperação de larvas infectantes da pastagem.
A segunda parte refere-se a distribuição sazonal e longevidade de larvas infectantes em
pastagem irrigada e não-irrigada. Amostras de vegetação, dos dois piquetes, não-
irrigado A, e irrigado B, foram examinadas a cada 14 dias pela técnica de Donald,
durante maio/01 a abril/03. Os resultados indicam que as condições ambientais foram
favoráveis ao desenvolvimento e longevidade de larvas infectantes de nematóides
gastrintestinais de caprinos. Os gêneros Trichostrongylus e Haemonchus ocorreram em
praticamente todos os meses. Trichostrongylus mostrou maior longevidade e
Haemonchus foi o mais abundante. O sistema de irrigação pareceu ser favorável à
longevidade de Haemonchus e Oesophagostomum, no entanto não mostrou influência
positiva no aumento de número de larvas no pasto. A ocorrência de Oesophagostomum
e Cooperia foi rara e inexistente, respectivamente.
A última parte aborda a distribuição e longevidade de larvas no solo e pastagem irrigada
e não-irrigada. Assim, amostras de solo e vegetação não-irrigada e irrigada foram
examinadas a cada 14 dias durante o período de outubro/02 a setembro/03. Amostras de
solo e vegetação foram processadas usando as técnicas de Baermann e Donald,
respectivamente. Os resultados mostram que as larvas de helmintos foram capazes de
migrar para o solo, considerando este um refúgio às condições adversas. No entanto, a
distribuição e longevidade indicaram a irrelevante importância epidemiológica deste
habitat. A vegetação foi o habitat eleito pelas larvas. O sistema de irrigação não
favoreceu a migração de larvas para o solo.
Palavras-Chave: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp.,
vegetação, irrigação, solo, epidemiologia, ruminantes.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Orientador), Laerte Grisi – UFRRJ (Co-orientador) e
Fábio B. Scott – UFRRJ.

18
ABSTRACT
CASTRO, Abisair Andrade de. Distribution and longevity of infective larvae of goats
(Capra hircus) gastrointestinal nematodes in pastures on borough Seropédica, RJ,
Brazil. Seropédica: UFRRJ, 2004. 97p. (Tesis – Doctor Science at Ciências Veterinárias,
Parasitologia Veterinária)
The knowledge of trichostrongyle infective larvae population level present on the pastures
is useful for epidemiological purposes. It enables to estimate parasitic risk for animals and
supply data to set up integrated control programs. This paper was elaborated in three parts:
first, a comparative study between Baermann and Donald techniques was accomplished for
the recovery of ruminants gastrointestinal nematodes infective larvae from pasture. To
develop the assay, six fecal samples naturally infected were placed on lawn. After 21 and
28 days, pastures samples were collected and processed for each proposed technique. The
statistic analysis showed that there is no significant difference between the results. Thus,
both techniques can be used to recover infective larvae from pasture.
The second part, refers to the seasonal distribution and longevity of goats gastrointestinal
nematodes infective larvae in irrigated and non-irrigated pastures. Thus, pasture samples at
two plots non-irrigated, A and irrigated, B were examined every 14 days for Donald’s
technique between mai/01 at april/03. The results indicate that environment conditions were
favorable to development and longevity of nematode infective larvae. The genus
Trichostrongylus and Haemonchus were present on all mouths. Trichostrongylus showed
more longevity and Haemonchus was more density. The irrigation system seems to be
favorable for the longevity of Haemonchus and Oesophagostomum infective larvae.
However, it did not have a positive influence on the increase of infective larvae number in
pasture. The Oesophagostomum and Cooperia occurrence was scarce and non-existent,
respectively.
The last part refers to distribution and longevity nematodes infective larvae in soil and
herbage irrigated and non-irrigated. Soil and pastures samples at two plots non-irrigated
and irrigated were examined every 14 days during october/02 at september/03. The soil and
herbage samples were processed using Baermann and Donald techniques, respectively. The
results showed that helminthes larvae were able to migrate for soil, that is considered a
refuge from the environment adverse condition. However, the distribution and longevity
indicate the low epidemiological importance of this niche. Pastures was the elected habitat
for the larvae. The irrigation system was not favorable to migration of infective larvae for
the soil.
Keywords: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp., pastures,
irrigation, soil, epidemiology, ruminants.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Advisor), Laerte Grisi – UFRRJ (Co-advisor) e Fábio B.
Scott – UFRRJ.

1
1. INTRODUÇÃO
O rebanho caprino da região sudeste do Brasil, calculado em 196.924 animais,
segundo o ANUALPEC (2001), representa a segunda maior região em número de
animais, e é em sua grande maioria constituído basicamente para uma pecuária de
subsistência. Os caprinos são considerados como importante fonte de alimento
fornecendo carne, leite principalmente para pequenos criadores nas áreas tropicais de
todo mundo (DEVENDRA, 1981).
Os nematóides gastrintestinais parasitos de caprinos constituem-se em um dos
fatores relevantes para o comprometimento da saúde e conseqüentemente da
produtividade em detrimento da verminose sub-clínica e morte dos animais
(REINECKE, 1970; ALLOMBY & URQUART, 1975). Eles possuem ciclo de vida
similar, com exceção do gênero Oesophagostomum que apresenta fase tissular. As
fêmeas adultas colocam ovos que são conduzidos juntamente com as fezes ao meio
exterior. Estes eclodem e desenvolvem-se em larvas de primeiro estágio, L1, e em
seguida em larvas de segundo estágio, L2. Elas são rabdiformes e se alimentam de
microorganismos do interior das fezes. De acordo com ROSE (1960) as massas fecais
funcionam como um reservatório e a pastagem, como veículo mais importante na
transmissão da parasitose. A larva de terceiro estágio, L3, é a infectante e migra das
fezes, indo a maioria para a vegetação, e uma pequena parte para o solo. Estas não se
alimentam e podem viver dias ou meses dependendo da espécie do nematóide e das
condições ambientais (LEVINE, 1980). Ao serem ingeridas por hospedeiros
susceptíveis, tornam-se adultas.
Durante o ciclo evolutivo, os estágios pré-parasíticos de nematóides
gastrintestinais no meio ambiente sofrem influência de uma série de fatores tais como:
temperatura, precipitação pluviométrica, umidade, luz solar, evaporação, oxigênio, tipo
de solo e pastagem (REINECKE, 1970). O desenvolvimento das larvas no pasto
depende basicamente da temperatura e da disponibilidade de umidade. Em geral, o
desenvolvimento dos estágios de vida livre é realçado pelo aumento da temperatura,
porém o tempo de sobrevivência diminui. A faixa de temperatura ótima para o
desenvolvimento parece ser entre 200C e 300C. Temperaturas extremas acima de 35-
400C, assim como temperaturas baixas são geralmente prejudiciais a todos os estágios

2
de vida livre e ovos; porém a larva infectante é geralmente mais resistente que outros
estágios (GIBBIS, 1973). Segundo este mesmo autor, fatores do microambiente,
incluindo tipo de solo e densidade da vegetação, também podem ter importante papel da
migração das larvas. Assim como período pré-patente e a fecundidade das fêmeas destes
nematóides. FITZSIMMONS (1970), em seu extenso trabalho de revisão, agrupou
alguns gêneros de helmintos de pequenos ruminantes como Haemonchus, Ostertagia,
Cooperia e Trichostrongylus como helmintos de ciclo de vida curto, tendo várias
gerações por ano, enquanto Nematodirus, e Oesophagostomum aqueles com ciclo de
vida longo, tendo somente uma ou duas gerações por ano. Segundo GIBBIS (1973), a
capacidade de ovopositar das fêmeas destes helmintos relacionada com o controle da
sua respectiva população é notável: a baixa taxa de produção de ovos de Nematodirus,
50 a 100 ovos por fêmea adulta, é compensada pela habilidade de seus estágios de vida
livre sobreviverem no pasto durante os meses do inverno. A alta capacidade de
ovopostura de Oesophagostomum, 3000 ovos por fêmea adulta, é justificada, pois é
longo o período pré-patente e há uma relativa susceptibilidade de seus estágios de vida
livre às condições ambientais.
Para estudar a distribuição e sobrevivência dos estágios pré-parasíticos de
tricostrongilídeos no ambiente, os métodos para recuperação de L3 na pastagem ganham
uma grande importância (DURIE, 1959). Várias técnicas têm sido testadas por vários
pesquisadores para isolar larvas de nematóides de diferentes meios onde as larvas
existem. A técnica mais bem conhecida e mundialmente utilizada é a de Baermann
(CORT et al. 1922). Outras técnicas foram descritas como TAYLOR, 1939; KAUZAL,
1940; DINABURG, 1942. DONALD (1967) descreveu sua técnica e obteve excelentes
percentuais de recuperação de L3 da pastagem. Alguns pesquisadores têm utilizado esta
técnica como método apropriado para suas pesquisas (VIANA, 1999; PIMENTEL
NETO et al., 2000; ALMEIDA, 2003). Segundo LANCASTER (1970) é importante
estabelecer a técnica que será utilizada, comparando um método com outro, com a
finalidade de padronizar a rotina, pois o valor da estimativa de larvas na vegetação e sua
relação com estudos epidemiológicos é indiscutível.
O crescente conhecimento da biologia e epidemiologia de nematóides
gastrintestinais de bovinos e ovinos têm contribuído para controle e conseqüentemente
com o aumento da produtividade. No entanto, pouco se conhece sobre a dinâmica
populacional de nematóides gastrintestinais de caprinos, no ambiente (CHARLES,
1989). Poucos dados também existem sobre a influência da irrigação sobre a

3
transmissão de larvas infectantes de tricostrongilídeos (STEWART & DOUGLAS,
1938; FURMAN, 1944). A introdução do sistema de irrigação pode mudar a
epidemiologia do parasitismo interno de nematóides, segundo afirmaram GRUNER et
al. (1989). BULLICK & ANDERSEN (1978) nos E. U. A. mostraram o positivo efeito
do sistema de irrigação sobre o desenvolvimento e sobrevivência de H. contortus.
Informações a respeito do ambiente sobre o desenvolvimento dos estágios de
vida livre de nematóides de ruminantes é de fundamental importância, pois a partir daí,
pode-se desenvolver medidas estratégicas e táticas de controle, objetivando um manejo
prático ou combate direto às formas pré-parasíticas (KRECEK et al., 1991). Existem
poucas informações disponíveis sobre a influência de solos tropicais albergando larvas
infectantes de nematóides (LYAKU et al. 1988). Na realidade existe uma grande
controvérsia a respeito deste fato. Alguns autores afirmam que o solo pode atuar como
reservatório de larvas infectantes, assim como as fezes, onde num ambiente
desfavorável, as L3 migrariam a determinadas profundidades para se protegerem. E
num momento seguinte, quando as condições ambientais estivessem favoráveis
migrariam de volta a superfície, recontaminando a pastagem (KAUZAL, 1941; REES,
1950; AL SAQUUR et al. 1982; GRUNER et al. 1982; CALLINAN & WESTCOTT,
1986). Por outro lado, existem outros autores que discordam deste fato, pois em seus
trabalhos, recuperaram poucas larvas do solo (ANSERSEN et al. 1970, ROSE &
SMALL, 1985).
O presente trabalho foi elaborado tendo como base os seguintes objetivos:
avaliar métodos para recuperar L3 da pastagem; estudar a distribuição e longevidade de
larvas infectantes de nematóides gastrintestinais de caprinos em pastagem irrigada e
não-irrigada, bem como no solo. O trabalho foi didaticamente dividido em três
capítulos. No primeiro trata-se da escolha da técnica para recuperação de larvas da
vegetação; o segundo, sobre a distribuição sazonal e longevidade de L3 na vegetação
irrigada e não-irrigada durante dois anos e o terceiro capítulo mostra o estudo de
distribuição e longevidade de L3 no pasto e solo, em área irrigada e não-irrigada.

4
2. REVISÃO DE LITERATURA
2.1. Técnicas para recuperação de larvas infectantes da pastagem
Em 1917 foi descrita por Baermann a primeira técnica para isolar larvas de
nematóides do solo, a qual era basicamente constituída por um funil de vidro, com
borracha de látex presa à sua parte posterior e fechada com uma pinça. Dentro do funil
um tamis com malha de 1mm coberto com pano, onde a amostra era colocada.
Primeiramente o funil era completado com água destilada esterilizada até à borda, em
seguida a amostra era mergulhada em água. Esta técnica foi detalhadamente descrita por
CORT et al. (1922); onde os autores testaram o efeito de várias modificações para
recuperar larvas de ancilostomídeos do solo: temperatura da água (acima de 120C), tipos
de substratos (úmido/seco), tamanhos das amostras (grandes/pequenas), tempo de
recuperação (seis horas ou mais). Eles registraram que mesmo nas melhores condições
para a obtenção de larvas, a taxa de recuperação foi alta, porém muito variável.
Utilizando-se a modificada técnica de Baermann segundo CORT et al. (1922),
com acréscimo de um número conhecido de larvas de ancilostomídeos, em 100mg de
amostras de diferentes tipos de solos, STOLL (1923), recuperou entre 90 a 95% de
larvas do húmus, constituído basicamente de vegetação em vários estágios de
decomposição; 80% de larvas da areia e entre 33 a 50% de larvas do solo argiloso.
CORT et al. (1926), usaram rotineiramente água a 450C, e observaram que de 1900-
3750 larvas de Ancylostoma e Necator colocadas em lotes de 200g de areia úmida,
recuperaram uma taxa de 58% de larvas, depois de 24 horas.
MÖNNING (1930) testando esta técnica em amostras de fezes e diferentes tipos
de solos, para recuperação de larvas de Trichostrongylus sp, Haemonchus contortus e
Oesophagostomum columbianum, registrou excelentes resultados comparados àqueles
de CORT et al. (1922) com larvas de ancilostomídeos. Seus melhores resultados foram
provenientes de amostras de solo e com a utilização de pequenos funis com cerca de
6cm de diâmetro.
A técnica de Baermann testada para recuperar larvas infectantes, L3, de
estrongilídeos, a partir de número de L3 conhecido em 40mg fezes estéreis de eqüinos,
resultou em baixos números de larvas, tendo os mesmos alta variação (PARNELL,
1936).

5
O primeiro registro da utilização da técnica de Baermann para recuperar larvas
infectantes das pastagens, foi descrito por TAYLOR em 1939 embora suas pesquisas a
campo tenham sido iniciadas anteriormente. A nova adaptação da técnica consistia na
lavagem e sedimentação de grandes quantidades de vegetação variando de 300 a 500g,
cortadas em zig-zag. Em seguida, as amostras eram tamisadas por papel filtro e a
subseqüente separação das larvas do sedimento era realizado pelo funil de Baermann. A
ineficiência do método e a perda de larvas durante a separação foi reconhecida pelo
autor.
Várias modificações à técnica de Baermann, para avaliar a sua eficácia em
recuperar larvas de H. contortus e Trichostrongylus spp. também foram sugeridas por
KAUZAL (1940). Ele fez vários testes associando o tamanho do funil à quantidade de
amostra e concluiu que, apesar, de um maior número de larvas ter sido recuperado
quando se utilizou água a temperatura ambiente, os mesmos foram baixos e variaram
muito. Dois anos depois, DINABURG (1942) testou novamente a eficiência da técnica
para recuperar L3 de H. contortus de amostras de solo e fezes, e também observou sua
alta variação: com o tipo de material utilizado, o substrato em questão, o número inicial
de larvas, concluindo assim, sua ineficiência.
O método de flutuação, denominado assim por CROFTON (1954), foi
desenvolvido para recuperar larvas infectantes da vegetação. As amostras eram
umedecidas, onde se adicionava lentamente uma solução saturada de sulfato de zinco
(ZnSO4) que permanecia por cinco minutos. Em seguida, as amostras eram tamizadas.
No entanto, seus resultados não foram satisfatórios.
O uso de lavagens da vegetação, seguido por centrifugações e adição de solução
por diferença de densidade, para separar L3 do sedimento proveniente da centrifugação,
foi primeiramente descrito por PARFITT (1955). Ele trabalhou com grandes
quantidades de pastagem e no teste de sua técnica, quando adicionou um número
conhecido de L3 na vegetação, foi observado uma taxa de recuperação em torno de
43%.
A modificada técnica de Baermann foi utilizada para recuperar os estágios de
vida livre de nematóides tricostrongilídeos das pastagens por ROHRBACHER Jr.
(1957), o qual adicionou gotas de detergente não-iônico, a faixa de 0.5ml por litro. O
procedimento resultou no aumento do número de larvas recuperadas da vegetação. No
entanto, o autor concluiu que, os percentuais de recuperação foram baixos; indicando

6
que a estimativa do número de larvas presentes no pasto, por esta técnica inspirava
cautela.
Tendo como objetivo estudar a distribuição e sobrevivência dos estágios pré-
parasíticos de tricostrongilídeos no pasto, DURIE (1959), criou uma outra técnica para
recuperar larvas da vegetação, uma vez que até aquele momento a técnica mais
utilizada, Baermann, juntamente com as outras novas técnicas apresentavam baixa
eficiência. Embora sua contribuição tenha sido considerável, a técnica era constituída
por um equipamento muito complexo. Trabalhando com grandes quantidades
individuais de amostras, sua técnica conseguiu recuperar a taxa média de 74 a 78% de
larvas do pasto e solo, respectivamente.
Visando o estudo da freqüência e distribuição de L3 de tricostrongilídeos em
amostras de pastagem ao acaso, DONALD (1967), desenvolveu uma nova técnica para
recuperar larvas de pequenas amostras, 25 a 35g, da pastagem. A técnica consistia
basicamente em várias lavagens das amostras de vegetação, com a utilização de
detergente não-iônico, duas centrifugações e separação das L3 do sedimento por
ressuspenção do mesmo em solução de iodeto de potássio a 1.63 molar. A avaliação
quanto a eficiência da técnica mostrou que seu desempenho foi independente da
quantidade de larvas presentes; podendo a mesma recuperar de 90% à 94% das larvas
no pasto.
Para estimar o número de larvas na vegetação ao longo do dia, HEATH &
MAJOR (1968), utilizaram ovinos fistulados.
Depois da técnica descrita por Donald, outras técnicas surgiram, baseadas no
mesmo princípio tendo na realidade poucas modificações e apresentando pequenas
diferenças no percentual de recuperação de L3 da pastagem (BAWDEN, 1969;
LANCASTER, 1970).
Uma nova e minuciosa avaliação foi proposta e testada por TODD Jr. et al.
(1970), com o objetivo de comprovar a eficácia da técnica de Baermann; pois eles
consideravam trabalhosas e de alto custo as técnicas até então desenvolvidas, mesmo
algumas demonstrando altos percentuais de recuperação de larvas. Os autores então,
avaliaram o efeito do tempo, tipo de filtro, temperatura, solução, iluminação, tamanho
do funil, peso e comprimento da gramínea, peso e tipo de solo sobre o número de larvas
recuperadas. Diante dos resultados os autores concluíram que a técnica de Baermann era
uma ferramenta útil para determinar o número relativo de larvas presentes no meio
examinado, se a mesma fosse utilizada com cautela.

7
SMEAL & HENDY (1972) propuseram uma técnica de recuperação de larvas,
onde o objetivo era trabalhar várias amostras simultaneamente, otimizando o tempo. O
método usado por esses autores para separar larvas da vegetação foi baseado na técnica
de Donald. No entanto, o mecanismo elaborado para separação das larvas do sedimento
do pasto era extremamente complexo, exigindo muitos equipamentos. Oito anos depois,
uma nova tentativa na elaboração de técnica para recuperar larvas da pastagem foi
idealizada, visando que a mesma tivesse uma alta taxa de recuperação, trabalhasse com
grandes amostras, fosse rápida e de baixo custo. Surgia então a técnica descrita por
YOUNG & TRAJSTMAN (1980) onde os princípios da técnica de Donald, eram
preservados, em termos das lavagens da vegetação e utilização de iodeto de potássio
para separação das larvas do sedimento. No entanto, alguns ajustes permitiram que o
tamanho das amostras fosse maior, sem, contudo perder a eficácia, em torno dos 94%.
No ano seguinte, outras inovações surgiam como tentativa de aperfeiçoamento
dessas técnicas: inclusive a utilização de máquinas de lavar, para a fase inicial de
processamento, onde as gramíneas eram lavadas. Esta referida técnica foi sugerida por
BÜRGER (1981). Apesar da taxa média de recuperação, em torno de 60%, os resultados
mostraram uma boa correlação entre a taxa de quantificação larval e o total da carga
parasitária em bezerros traçadores, indicando que a leitura das larvas no pasto, refletia o
risco de contaminação dos hospedeiros.
CHIEJINA (1982) utilizou os princípios da técnica de Baermann, e com o
volume coletado, resultante desta primeira etapa, fez lavagens por meio de
centrifugações e ressuspendeu o sedimento em solução saturada de sulfato de magnésio.
O autor recomendou este procedimento para rotina da estimativa de população de L3 no
pasto, pois se tratou de método simples, sem a exigência de grandes equipamentos e o
mesmo foi considerado relativamente eficaz.
Vários métodos foram descritos para avaliar a concentração de larvas no pasto.
Prévios estudos (WALLER et al., 1981; CABARET et al., 1982) àquela época,
indicavam que poderia haver correlação entre a quantidade de larvas no pasto e a carga
parasitária de nematóides no hospedeiro. A fim de se confirmar essa hipótese
GETTINBY et al. (1985), compararam duas técnicas: de um lado a recuperação de L3
da pastagem, seguindo a conhecida técnica de Baermann e de um outro lado, a técnica
de fístula esofagiana. Posteriormente introduziram animais traçadores no local do
experimento. As observações dos autores confirmaram a existência de correlação
positiva entre o número de larvas no pasto e a carga parasitária dos hospedeiros. Com as

8
mesmas intenções MARTIN et al. (1990) no sul da Austrália, formularam uma
modificada técnica para recuperar L3 da pastagem, seguindo os princípios de Donald e
compararam com os resultados de animais traçadores. Os autores observaram que
existia correlação entre os dois métodos; e a medida que aumentavam o número de
animais traçadores, aumentava também a correspondência entre os métodos,
confirmando a correlação positiva de larvas na pastagem com a carga parasitária de
nematóides no hospedeiro herbívoro.
ESYKER & KOOYMAN (1993) fizeram adaptações à técnica descrita por
JORGESEN (1975) para recuperar larvas de Dictyocaulus viviparus da pastagem, e a
testaram com a original para recuperar larvas de tricostrongilídeos da vegetação fezes.
Os autores observaram a melhor eficiência para técnica original com larvas da
pastagem. A técnica utilizava 200 a 500g de vegetação e toda a fase inicial era muito
parecida com a técnica de Donald. Os autores também comentaram a falta de
padronização de técnicas nessa área e sugeriram que os pesquisadores não só
escolhessem determinada técnica mais também testassem e comparassem.
Com o objetivo de avaliar o efeito do tempo a partir do momento de coleta da
pastagem até o término total do processamento, para recuperar L3 da vegetação FINE et
al. (1993) elaboraram um estudo onde comprovou-se a importância do tempo de
processamento. Para isto, os autores basearam seus estudos na técnica de Baermann. Os
resultados mostraram que as taxas de recuperação declinavam a medida que o tempo de
processamento aumentava.
COUVILLION (1993) em seu trabalho de revisão enfatizou a padronização da
técnica escolhida pelo período de observação, e também, argumentou a importância da
estimativa do número de larvas infectantes na vegetação como um importante estudo
adjunto a epidemiologia de nematodioses gastrintestinais.
2.2. Distribuição e longevidade de larvas infectantes na pastagem, no exterior
As pesquisas sobre ecologia de larvas de nematóides em pastagens foram
iniciadas por TAYLOR em 1938. Ele contaminou artificialmente pastagens com larvas
de tricostrongilídeos e observou que no início, a mortalidade das larvas foi alta, mas
depois houve uma contaminação residual e as larvas foram encontradas nas pastagens
por um período de até 133 dias ou 19 semanas. A partir deste trabalho, muitos outros
foram feitos em diferentes regiões do mundo. Pôde-se então, observar a influência
climática determinando as diferenças na permanência de larvas nas pastagens, nas

9
estações do ano. O clima também determina as espécies presentes e/ou predominantes
em cada região. As condições ambientais sobre a migração de larvas de
tricostrongilídeos para a vegetação foram estudadas por ROGERS (1940). O autor
registrou que a umidade do solo quando apresentou-se acima de 85%, favoreceu a
migração de larvas para a vegetação; e que a espécie H. contortus foi a mais ativa em
altas temperaturas. Nos Estados Unidos, DINABURG (1944) contaminou
artificialmente as pastagens com larvas de H. contortus e verificou que nos meses do
verão e inverno as condições foram desfavoráveis para a sobrevivência das larvas.
DOLL & HULL (1948) também trabalhando nos Estados Unidos, com larvas de
tricostrongilídeos, encontraram L3 de H. contortus, Cooperia spp. e Oesophagostomum
spp., sendo que somente as larvas de H. contortus foram encontradas na pastagem até
três meses após o início do experimento.
No Quênia, DINNIK & DINNIK (1958) trabalharam com fezes de caprinos
experimentalmente infectados com larvas de H. contortus, e verificaram que o
desenvolvimento, dos estágios pré-parasíticos foram favorecidos quando a temperatura
do ar alcançou a média das máximas de 23.30C ou mais, e a média das mínimas de
12.20C; com precipitação pluviométrica de 25.4mm. Esses mesmos autores, em 1961,
continuaram seus estudos: eles contaminaram artificialmente as pastagens com H.
contortus em diferentes épocas do ano. No período quente e seco constataram que a
sobrevivência máxima das larvas foi de cinco semanas, sendo o mais curto observado
durante o experimento. No período frio, as larvas sobreviveram por um período muito
maior, 14 semanas. Nesse período a temperatura e a umidade foram mais baixas,
havendo presença de névoa durante a noite e orvalho pelas manhãs. O período chuvoso
apresentou dados intermediários entre o seco e o frio. As larvas que ficaram em lugares
sombreados e em vegetação mais densa, tiveram o período de sobrevivência maior. As
que permaneceram em exposição solar direta e em pastagens menos densa,
sobreviveram por menos tempo.
O tempo e o clima na dinâmica das larvas de nematóides de ruminantes foram
discutidos por LEVINE (1963), nos Estados Unidos, o qual definiu como “tempo” o
conjunto de condições climáticas num determinado momento. Já o “clima” foi definido
como a soma destas condições climáticas em um período mais longo. Desta forma, o
clima indicaria quais os nematóides mais prováveis de serem identificados num
determinado local e o tempo determinaria as variações destes parasitos. Portanto, o
clima seria responsável por estabelecer uma situação abrangente, enquanto que o tempo

10
determinaria as minúcias. O autor comentou ainda, que as condições de tempo poderiam
tomar características diferentes de um ano para outro. Com relação aos gêneros
Cooperia e Trichostrongylus, descreveu que estes foram mais resistentes na fase larval
do que os gêneros Oesophagostomum e Haemonchus.
Na Índia, TRIPATHI (1966), observando a ocorrência dos nematóides
gastrintestinais de caprinos, baseado na eliminação de ovos, citou que os maiores
índices ocorreram durante a estação chuvosa, e que a temperatura e umidade foram
fatores essenciais para o desenvolvimento de ovos e larvas. Durante a estação quente, a
baixa produção de ovos foi provavelmente devida à alta temperatura e baixa umidade,
que favoreceu a dessecação e destruição de ovos e larvas na pastagem.
Num levantamento helmintológico realizado por McCULLOCK &
KASIMBALA (1968) na Tanzânia, em 227 caprinos, os autores identificaram os
seguintes parasitos: H. contortus, T. colubriformis, O. columbianum, B.
trigonocephalum, C. pectinata, C. punctata, T. ovis. Os autores comentaram que os
fatores climáticos exerceram notável influência sobre as cargas parasitárias, pois
verificaram que B. trigonocephalum apresentou uma ocorrência bastante baixa,
chegando a zero, nas regiões de baixa precipitação pluviométrica anual; enquanto que
H. contortus apresentou seu pico na época das chuvas. Afirmaram ainda, que o clima
representou um fator de pouca importância nas infecções por T. colubriformis e O.
columbianum. Em 1970, esses mesmos autores, após observarem a importância
econômica e a distribuição dos principais nematóides gastrintestinais em rebanhos
examinados durante um ano, citaram as seguintes ocorrências: H. contortus 68%, T.
colubriformis 84%, O. columbianum 71%, B. trigonocephalum 32.5%, C. pectinata
1.3%.
Em um estudo sobre a distribuição sazonal de larvas de Haemonchus sp. obtidas
de culturas de fezes de caprinos, TRIPATHI (1970a) constatou que a maior recuperação
foi observada no final do período chuvoso e a menor durante a época quente ou seca. Os
resultados obtidos indicaram que a temperatura elevada aliada à alta umidade favoreceu
o desenvolvimento das larvas e que o aumento ocorrido nas culturas de larvas na
estação chuvosa foi devido à infecção adquirida pelos hospedeiros no início deste
período. Em um outro estudo, TRIPATHI (1970b), registrou a maior predominância
para os gêneros Strongyloides, Haemonchus, Trichostrongylus, Oesophagostomum e
raramente Cooperia, durante a estação das chuvas. Observou-se que o período seco ou
quente foi desfavorável ao desenvolvimento e sobrevivência das larvas no ambiente.

11
A prevalência dos principais nematóides gastrintestinais de caprinos foi estudada
na Nigéria, onde se observou a presença de H. contortus, T. colubriformis, O.
columbianum, sendo também rara a presença de nematóides do gênero Cooperia
(FABIYI, 1970). Um levantamento sobre helmintos parasitos de animais domésticos,
VASQUEZ & MARCHINARES (1971) citaram que gêneros como Haemonchus,
Ostertagia, Trichostrongylus e Oesophagostumum são encontrados parasitando
caprinos.
Através de um estudo sobre variação sazonal dos nematóides de caprinos na
Nigéria, FABIYI (1973), observou que os aumentos de infecção ocorriam na estação
úmida ou das chuvas e que estas condições ambientais pareciam favorecer o
desenvolvimento e sobrevivência das fases pré-parasíticas. Altas contagens de
Haemonchus e Strongyloides foram obtidas na época das chuvas e de Gaigeria,
Oesophagostomum e Trichostrongylus só ocorreram no final desta estação.
BAKER (1975), na Califórnia, citou que H. contortus, T. colubriformis, T. axei e
O. circumcincta são as mais importantes espécies parasitas de caprinos. Relatou,
também, que entre os elementos necessários para o completo desenvolvimento dos
estágios pré-parasíticos estão a temperatura e a umidade como essenciais; destacou que
a confecção de bioclimatográficos regionais auxiliam no controle dos nematóides
gastrintestinais.
Em seu trabalho de revisão sobre o desenvolvimento e sobrevivência de larvas
de H. contortus e T. colubriformis, LEVINE & TODD (1975), deram ênfase aos fatores
ambientais e destes, estabeleceram que a temperatura e umidade eram os mais
importantes. Observaram que poucas larvas de H. contortus sobreviveram no inverno, e
que as mesmas permaneceram por mais tempo na primavera e menos no verão. Ao
contrário T. colubriformis sobreviveu bem no inverno; e que o outono foi a melhor
época para a sobrevivência de ambas espécies. Os autores comentaram que as larvas de
H. contortus apresentaram maior resistência quando submetidas a repetidas dessecações
do que as de T. colubriformis.
Os parâmetros de desenvolvimento e sobrevivência de H. contortus foram
estudados na Nigéria por um período de um ano. Observou-se que o desenvolvimento
de ovos a L3 foi dentro de uma semana nos meses de maio a outubro, com exceção do
mês de agosto; e que o maior período de sobrevivência destas larvas foi de 63 dias, nove
semanas a partir do mês de setembro. Os autores concluíram que a precipitação
pluviométrica foi o fator mais importante, influenciando o desenvolvimento e a

12
sobrevivência da L3; pois para o desenvolvimento de ovo a L3, registrou-se média
diária de 3mm ou mais. E para o tempo de sobrevivência das larvas verificou-se que
quando a chuva caía uniformemente por um período, prolongava-se a permanência das
larvas no pasto (OKON & ENYENIHI, 1977).
Para acompanhar a dinâmica populacional dos estágios de vida livre de
nematóides gastrintestinais de caprinos, numa região de clima tropical, AUMONT &
GRUNER (1989) contaminaram piquetes com ovos de helmintos, durante a estação
chuvosa e seca. O acompanhamento de cada depósito durou 56 dias, oito semanas. Eles
recuperaram larvas de Haemonchus e Trichostrongylus e observaram que as larvas
surgiram na vegetação sete a 14 dias depois do depósito. A longevidade das larvas na
vegetação foi de 49 a 56 dias, sete a oito semanas. Os autores comentaram que houve
uma marcante diminuição na taxa de eclosão de ovos e do desenvolvimento de L3 com
algumas paradas na eclosão dos ovos de Trichostrongylus, na estação seca.
Consideraram que os eventos climáticos e o fato da vegetação apresentar-se seca
durante o pastoreio, foram fatores importantes, parecendo interferir na dinâmica
populacional das formas de vida livre destes helmintos.
A variação sazonal da população de nematóides gastrintestinais de caprinos, na
vegetação foi estudada por RAHMAN & COLLINS (1990), na Austrália. Eles
verificaram que o perfil do gráfico de disponibilidade de larvas no pasto foi paralelo
àquele de temperatura e precipitação pluviométrica, sugerindo que os picos de L3
ocorreram quando a temperatura e a disponibilidade de umidade foram consideradas
ótimas. O gênero predominante foi Trichostrongylus, seguido por Haemonchus e
Ostertagia. A alta proporção de larvas de Haemonchus em culturas de fezes foi
observada durante os meses do verão.
O desenvolvimento e sobrevivência de larvas infectantes de H. contortus e T.
colubriformis foram estudadas em pastagens tropicais na Austrália. Piquetes foram
mensalmente contaminados com fezes de caprinos, naturalmente infectados. As duas
espécies de larvas desenvolveram-se bem durante todo o ano na estação úmida e
esporadicamente na estação seca. A contagem de larvas apresentou-se negativa depois
de nove semanas de contaminação, entre setembro e março; porém a longevidade
aumentou durante a estação fria de abril a agosto (BANKS et al. 1990).
CONNOR et al. (1990) investigaram aspectos epidemiológicos e possíveis
métodos de controle para helmintoses em caprinos naturalmente infectados, ao sul da
Tanzânia. Durante a estação chuvosa, contagens de ovos apresentaram-se altas e depois

13
caíram permanecendo baixas durante a estação seca. O tratamento anti-hemíntico
mostrou-se eficiente quando administrado após as chuvas, demonstrando que o
tradicional programa de aplicação de anti-helmínticos pode ser uma importante base
para controles rotacionais.
Um programa de rotação de pastagens para combater nematóides gastrintestinais
de caprinos, em ambiente tropical chuvoso foi estabelecido por BARGER et al. (1994).
Os autores observaram que, a partir da contaminação de ovos de H. contortus, T.
colubriformis e O. columbianum mensalmente durante um ano, o desenvolvimento e
sobrevivência das L3 foi rápido e contínuo, com um período curto para a longevidade
no pasto, de três a sete semanas. Estes resultados indicaram que um sistema rotacional
com 10 piquetes para pasto poderia, ser utilizado com a seqüência de rotação de 3.5 dias
para cada piquete. Este tempo permitiria a redução no tratamento de anti-helmínticos
em caprinos.
A contaminação da pastagem por larvas infectantes de tricostrongilídeos de
caprinos, foi avaliada durante a estação seca, nas ilhas francesas de Guadalupe. Os
autores observaram que a contaminação diminuiu significativamente na estação seca,
porém mantiveram-se em níveis considerados importantes (SIMON et al. 1996).
Com o objetivo de estudar a capacidade de cabras leiteiras desenvolverem
resposta a H. contortus e T. colubriformis depois de um prévio contato, HOSTE &
CHARTIER (1998), elaboraram um estudo e observaram que a resposta desenvolvida
foi incapaz de limitar a população de nematóides, podendo haver repercussões
patofisiológicas e conseqüências na produção.
DIALLO (1998) estudou comparativamente a dinâmica sazonal de nematóides
gastrintestinais nos pequenos ruminantes, ovinos e caprinos, na África, com base na
liberação de ovos. Os resultados mostraram que os ovinos apresentaram um nível de
infecção maior que os caprinos e que todos os animais foram infectados ao longo de um
ano predominantemente pelos gêneros Haemonchus, Oesophagostomum,
Trichostrongylus, Cooperia e Gaigeria e ainda, que houve uma marcante redução nas
intensidades desses parasitos durante a estação seca. O pico de intensidade ficou
registrado entre maio e setembro que coincidiu com a máxima anual de precipitação e
umidade. Dessa forma o autor estabeleceu os melhores períodos para intervenção com
anti-helmínticos: no fim da estação seca, maio; início da estação seca, novembro e no
meio do inverno, agosto.

14
A relação entre o número por ovos de grama de fezes, o.p.g., e carga parasitária
nos caprinos foi estudada por MORALES et al. (1998). Os autores observaram que
houve uma correlação positiva e que o aumento do o.p.g. estava associado à presença de
H. contortus e O. columbianum; enquanto a redução do o.p.g. estava associada com a
presença de Trichostrongylus sp.
No nordeste da África foi conduzido um estudo com helmintos de pequenos
ruminantes. Os autores observaram que os gêneros predominantes foram Haemonchus,
Oesophagostomum, Trichostrongylus e Strongyloides e que, os ovinos apresentaram-se
mais parasitados do que os caprinos. Os autores comentaram ainda a importância de um
estudo epidemiológico para estabelecer um efetivo método de controle
(TCHOUMBOUE et al. 2000). Na Grécia, para um estudo sobre a flutuação estacional
de nematóides gastrintestinais de caprinos, BOUGAS & THEODORIDIS (2000)
sacrificaram cerca de 268 animais ao longo de dois anos e observaram que nematóides
como H. contortus, T. colubriformis, Nematodirus filicollis e Chabertia ovina foram
continuamente encontrados durante todo o período enquanto que C. curticei, e N.
helvetianus foram encontrados esporadicamente.
Um estudo sobre a atuação dos helmintos gastrintestinais sobre a produção de
caprinos no Quênia, mostrou que a contagem de ovos no grupo tratado com anti-
helmínticos permaneceu baixa enquanto que no grupo controle houve um aumento
gradual durante o período de estudo. A contagem esteve alta durante a curta estação
chuvosa. O grupo controle perdeu peso durante a estação seca e num grande esforço,
houve um certo crescimento no curto período chuvoso. Já grupo tratado manteve seu
peso na estação seca e melhorou durante a estação chuvosa (GITHIGIA et al. 2001). A
patogenicidade de H. contortus em caprinos foi estudada durante as diferentes estações
do ano por LAHA et al. (2001). Eles observaram que os maiores percentuais de infecção
foram durante a estação chuvosa.
Na Etiópia, os efeitos ambientais sobre a dinâmica sazonal da liberação de ovos
por helmintos gastrintestinais de caprinos foram avaliados por um período de um ano. O
gênero Haemonchus foi o mais prevalente seguindo por Strongyloides e
Trichostrongylus. A liberação de ovos no mês de julho foi significativamente diferente
que nos outros meses exceto no mês de junho. Os resultados mostraram que infecções
mistas são comuns em caprinos. As altas contagens de ovos coincidiram com a estação
das chuvas. Dessa forma o uso de medicamentos devem ser usados antes e depois da
estação das chuvas como uma estratégia útil para o controle das helmintoses

15
gastrintestinais (DEBELA, 2002). Na Índia, BHOJANE et al. (2002) observaram que
infecções por Haemonchus e Strongyloides em caprinos foram altas e que infecções por
Oesophagostomum foram baixas; e comentaram ainda, que a estação do ano tem um
significante efeito sobre o parasitismo.
2.3. Distribuição e longevidade de larvas infectantes na pastagem, no Brasil
No estudo sobre a incidência de helmintos de caprinos procedentes da Bahia,
GRISI (1975) registrou a presença e os seguintes valores para os períodos de chuva e
seca, respectivamente: H. contortus, 85.79% e 58.82%, T. colubriformis, 61.22 e
37.25% O. columbianum, 61.22% e 54.90% e C. curticei, 4.08% e 0%, dentre outros.
CAVALCANTI (1974) pesquisando a prevalência estacional de nematóides
gastrintestinais de caprinos procedentes de três zonas fisiográficas do Estado de
Pernambuco, verificou que a intensidade do parasitismo foi maior nos lotes de animais
trabalhados na estação das chuvas do que nos lotes distribuídos na estação de estiagem.
As espécies encontradas parasitando caprinos foram: H. contortus, T. colubriformis, O.
columbianum, Strongyloides, B. trigonocephalum sendo as três primeiras mais
prevalentes e as que ocorreram com mais intensidade.
Utilizando caprinos traçadores para estudar a prevalência sazonal de nematóides
gastrintestinais de caprinos, no Estado de Pernambuco, CHARLES (1989) notificou que
as espécies mais prevalentes foram H. contortus, S papillosus e O. columbianum.
Verificou também que a carga parasitária foi considerada alta durante a última estação
chuvosa e início da seca (março-junho) e que a mesma apresentou-se baixa no meio da
estação chuvosa (janeiro-fevereiro). A contaminação dos animais traçadores ocorreu
principalmente em meados da estação chuvosa e no princípio da estação seca (janeiro-
junho).
Em um estudo conduzido no nordeste brasileiro, foi observada a evolução de
nematóides gastrintestinais de caprinos. Os autores concluíram que a idade dos caprinos
não pareceu ser um fator de resistência aos nematóides gastrintestinais, embora a carga
parasitária, dos animais com 11 a 12 meses de idade, tenha sido significativamente
maior que nos grupos com outras idades; sendo as espécies mais prevalentes em ordem
decrescente: T. colubriformis, H. contortus e O. columbianum (SILVA et al. 1998).
A variação sazonal de nematóides gastrintestinais de pequenos ruminantes foi
estudada no semi-árido nordeste brasileiro por AROSEMENA et al. (1999). Os autores
observaram que a prevalência desses nematóides foi maior nos caprinos e que as

16
espécies predominantes foram H. contortus e T. axei. A dinâmica sazonal da carga
parasitária apresentou-se de forma diferente nos ovinos e caprinos, porém a prevalência
foi considerada baixa para os dois hospedeiros no início da estação chuvosa (janeiro,
fevereiro e março).
Em Seropédica, PIMENTEL NETO & FONSECA (no prelo) estudaram a
distribuição das helmintoses pulmonares e gastrintestinais em caprinos traçadores. Os
autores verificaram que H. contortus foi o nematóide mais abundante na pastagem
durante todo período, sugerindo uma possível competição com T. colubriformis. Em
ordem decrescente os autores relataram os helmintos mais prevalentes: H. contortus,
54.6%, T. colubriformis, 41% e Oesophagostomum, 2%. Registraram o achado
esporádico de Cooperia.
2.4. Estudos com pastagem irrigada
Com a necessidade de aumentar a produtividade de pequenos ruminantes em
épocas de estiagem, surgiram os programas de irrigação e com estes os primeiros
estudos sobre a ecologia de nematóides gastrintestinais em pastos irrigados, pois a
freqüente irrigação parecia favorecer o desenvolvimento e sobrevivência de larvas na
vegetação (STEWART & DOUGLAS, 1938). Outros estudos confirmando esta hipótese
foram realizados, FURMAN (1944), BULLISK & ANDERSEN (1978).
Um estudo sobre a incidência sazonal de tricostrongilídeos em pastos irrigados
foi realizado no sul da África, por HORAK & LOUW (1977). Eles observaram que H.
contortus foi a espécie mais prevalente e que o pico de sua carga parasitária foi
registrado nos meses de janeiro - maio. Trichostrongylus sp. apresentou-se com maior
freqüência em abril – agosto. O registro de O. columbianum foi considerado esporádico.
Na Califórnia, a ocorrência sazonal de nematóides gastrintestinais de ruminantes
em pasto irrigado, mostrou que o sistema influencia na ecologia dos tricostrongilídeos
(BAKER et al. 1981).
URIARTE et al. (1985) investigou a distribuição de tricostrongilídeos em
sistema de pastos irrigados no nordeste da Espanha. Eles concluíram que esse sistema, o
qual foi um pouco diferente dos tradicionais, pois nesse era permitido o alagamento da
área, não favoreceu aos tricostrongilídeos.
O efeito da irrigação no desenvolvimento e sobrevivência de L3 de caprinos foi
estudada em Guadalupe, nas ilhas francesas. Segundo os autores a irrigação
proporcionou condições favoráveis ao desenvolvimento dos ovos em larvas. A proposta

17
de rotação de pastagem com 28-35 dias poderia minimizar o crescente risco de infecção
dos caprinos devido à irrigação (GRUNER et al. 1989). Em um estudo in vitro, sobre a
migração horizontal de L3 de tricostrongilídeos, os autores observaram que em
gramínea irrigada o máximo em termos de distância que a larva pôde alcançar foi de
21cm da amostra fecal HOLASOVÁ et al. (1989).
No estudo realizado por KRECEK et al. (1991) para avaliar os efeitos
ambientais sobre a ecologia das fases de vida livre de nematóides de ruminantes,
verificou-se o efeito do tempo ao longo do dia; a sazonalidade e o tipo de extrato:
vegetação e solo sobre a disponibilidade de larvas de Haemonchus em pastos irrigados,
no sul da África. Houve diferença significativa nos resultados apresentados entre as
estações, no tipo de extrato e na interação estação com extrato. No entanto, não foi
registrado diferença significativa no número de larvas quantificadas ao longo do dia. Os
autores também verificaram que larvas de Haemonchus foram recuperadas em maior
número no verão e outono e que o número de larvas recuperadas na vegetação foi maior
que o recuperado no solo. Neste mesmo ano, REINECKE & LOWN (1991) estudam
uma forma de controle das nematodioses de ovinos em pastagens irrigadas, associando
anti-helmínticos com trocas estratégicas de rebanhos de ovinos por bovinos. Os bezerros
permaneciam no piquete de dezembro a fevereiro o que seria relativo a estação do
outono; já os ovinos eram introduzidos em junho e permaneciam até a primavera, no
mês de agosto.
Os efeitos do microclima sobre L3 de H. contortus e H. placei em pastos
irrigados foram estudados por KRECEK et al. (1992). O efeito da temperatura foi
similar para as duas espécies. À medida que a temperatura aumentava, o número de L3
no pasto aumentava; o mesmo foi registrado com relação à umidade relativa.
De acordo com REINECKE et al. (1994), infecção maciça por Trichostrongylus,
Haemonchus e Ostertagia foi adquirida por ovinos depois de um verão com sistema de
irrigação, no sul da África.
A epidemiologia de nematóides gastrintestinais de bovinos foi estudada em duas
áreas: irrigada e não-irrigada, no Zimbábue, África. Foi observado que durante a estação
seca, o número de larvas apresentou-se baixo nos pastos irrigados. H. contortus
sobreviveu nesta estação como L4 inibida. O desenvolvimento de L3 no pasto coincidiu
com pico de liberação de ovos na estação das chuvas (MOYO et al. 1996).
O potencial de áreas irrigadas favorece a pastagem e pode ser usada como
estratégia contra os problemas climáticos aumentando a produtividade de ruminantes

18
nas condições do Mediterrâneo. No entanto, esse sistema favorece grandemente a
contaminação com nematóides, os quais limitam o desenvolvimento e
conseqüentemente baixam a produtividade. VALDERRABANO et al. (2000) sugeriu
que esse sistema fosse associado a controles estratégicos de anti-helmínticos. Seu estudo
mostrou que essa hipótese é viável.
CARMICHAEL & BIEN (2002) estudaram o sistema de irrigação no verão da
Austrália para aumentar a produtividade de ovinos, observando também as helmintoses.
Eles relataram que invernos e primaveras antecedidos por verões irrigados poderiam ser
estações altamente arriscadas para a contaminação dos animais, embora o sistema
favorecesse o desenvolvimento da vegetação, aumentando a produtividade.
Depois de quatro anos de observações sobre a ecologia das larvas de
tricostrongilídeos em pastos irrigados, no sul da África, WYK et al. (2003) concluíram a
que a necessidade da utilização de anti-helmínticos foi baixa no final do inverno e
primavera.
2.5. Distribuição e longevidade de larvas infectantes no solo
Os resultados dos trabalhos de TAYLOR (1938, 1939) estimularam a
investigação da população de larvas infectantes no solo, pois na primeira publicação
Taylor afirmou que houve uma contaminação residual e no segundo o autor descreveu a
técnica por ele utilizada, que originariamente foi descrita para recuperar larvas do solo.
KAUZAL (1941) considerando, então que ovos e larvas de nematóides poderiam
sobreviver por longos períodos e que o mesmo poderia servir como reservatório para
futuras contaminações da vegetação, quando as condições ambientais fossem
favoráveis; iniciou seu estudo examinando vegetação e solo para determinação do
número de larvas infectantes de nematóides. O autor concluiu que examinando apenas a
vegetação para estimar o número de L3 de H. contortus não foi suficiente. Em seu
experimento o máximo de larvas recuperadas da vegetação foi de 16%, enquanto que no
solo foi de 75%. Ele comentou que as consideráveis flutuações no número de larvas
presentes no solo e vegetação podem ser correlacionadas com as condições ambientais.
Os resultados também mostraram que a recuperação de larvas na vegetação é
dependente da quantidade de gramínea, pois essa cobertura vegetal pode possibilitar
boas condições e proteger as larvas.
CROFTON (1948) estudou a distribuição de larvas na vegetação, no solo e na
região superficial solo, a uma polegada de profundidade, durante um ano. Ele observou

19
que nos meses quentes, junho, julho e agosto a maioria das larvas foram encontradas na
vegetação, depois no solo e por fim na região superficial. Já nos meses de abril, maio e
setembro, o maior número de larvas foi encontrado na região superficial e poucas na
vegetação e solo. O autor concluiu que a distribuição de L3 pode ser explicada sem
referências ao geotropismo.
A superfície do solo, 5mm de camada, e a vegetação adjacente foram avaliadas
quanto à distribuição de larvas por REES (1950), ao longo do dia durante um ano.
Observou-se que a maioria das larvas foram encontradas na vegetação, mas houve
registro de larvas na superfície do solo.
ANDERSEN et al. (1970) recuperaram poucas larvas do solo. Eles concluíram
que as larvas migram diretamente dos pellets para a vegetação e que as poucas L3
encontradas no solo eram provenientes da lavagem da gramínea ou da rota de migração
para a vegetação.
A ecologia de Trichostrongylus axei foi estudada por CALLINAN (1978). Ele
encontrou 66% das larvas na vegetação em comparação ao solo. No ano seguinte
resultados semelhantes foram registrados para a espécie T. vitrinus CALLINAN (1979).
A capacidade de migração das larvas foi estudada por FINCHER & STEWART
(1979). Eles enterraram fezes contaminadas, alguns dias depois começaram a recuperar
larvas da vegetação. Os autores observaram que as L3 foram capazes de migrar a partir
de 12,5cm de profundidade.
Em um estudo sobre a migração de larvas de Ostertagia em vegetação e em
camadas do solo, observou-se que as L3 foram recuperadas em todas as ocasiões de
análise a pelo menos 15cm de profundidade. Este estudo sugeriu a possível migração
das larvas para o solo durante o inverno, reemergindo na primavera e verão (AL
SAQUR et al., 1982).
A distribuição de tricostrongilídeos no solo foi estudada a campo. Os autores
observaram que no final do inverno as larvas puderam migrar da superfície a 40cm ou
mais de profundidade. Eles afirmaram que esta migração pode ter sido em decorrência
do carreamento pela água; associando a migração das L3 para o solo com a oferta de
água no ambiente (GRUNER et al., 1982).
ROSE & SMALL (1985) coletaram durante um ano amostras de vegetação e
solo de área utilizada como pastoreio para ovinos. Pouquíssimas larvas foram
recuperadas do solo, no entanto, um número considerável foi recuperado da vegetação.
Na segunda parte deste estudo, os autores enterraram a 10cm de profundidade, fezes

20
com L3 de Trichostrongylus vitrinus e fizeram coletas da vegetação adjacente. Os
resultados indicaram que as larvas migraram para a vegetação, restando poucas no solo.
Os autores concluíram que o solo pode ser um reservatório para L3.
A distribuição de tricostrongilídeos na vegetação e no solo foi estudada em
condições de temperatura e umidade controladas por CALLINAN & WESTCOTT
(1986). Eles recuperaram em média oito vezes mais larvas do solo do que na pastagem.
Concluíram, então que o solo é um importante reservatório de larvas.
A longevidade de lavas infectantes de tricostrongilídeos foi estudada em
diferentes tipos de solos tropicais por LYAKU et al. (1988). Eles observaram que larvas
de Cooperia e Oesophagostomum sobreviveram até nove semanas nos solos e que
Haemonchus e Trichostrongylus sobreviveram até três.
HOLASOVÁ et al. (1989) estudaram a migração de tricostrongilídeos. Os
achados deste estudo mostraram que as larvas foram recuperadas esporadicamente do
solo a uma profundidade de 2cm e que somente uma única vez larvas de Ostertagia
foram recuperadas a 7cm de profundidade.
3. CAPÍTULO I

21
COMPARAÇÃO ENTRE AS TÉCNICAS DE BAERMANN MODIFICADA E DONALD
UTILIZADAS PARA RECUPERAR LARVAS INFECTANTES DE NEMATÓIDES
GASTRINTESTINAIS DE RUMINANTES DA PASTAGEM.
RESUMO

22
CASTRO, Abisair Andrade de. Comparação entre as técnicas de Baermann
modificada e Donald utilizadas para recuperar larvas infectantes de nematóides
gastrintestinais de ruminantes da pastagem. Seropédica: UFRRJ, 2004. 97p. (Tese –
Doutorado em Ciências Veterinárias, Parasitologia Veterinária)
A estimativa de densidade de larvas na vegetação é usada em estudos epidemiológicos
para determinar as variações sazonais ou mensais de contaminação da pastagem cm
larvas infectantes de tricostrongilídeos, e com isso estabelecer o risco de infecção dos
hospedeiros. Sendo assim, um estudo comparativo entre as técnicas de Baermann e
Donald, foi realizado para recuperação de larvas infectantes de nematóides
gastrintestinais de ruminantes da pastagem. Para o desenvolvimento do estudo, seis
amostras de fezes naturalmente infectadas foram depositadas a campo. Após 21 e 28
dias ao depósito, amostras de gramínea foram coletadas e processadas pelas técnicas
propostas. A análise estatística demonstrou que não houve diferença significativa entre
os resultados. Assim, as duas técnicas podem ser utilizadas para recuperação de larvas
infectantes da pastagem.
Palavras-Chave: recuperação de larvas, vegetação, bovinos, Haemonchus sp.,
Oesophagostomum sp.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Orientador), Laerte Grisi – UFRRJ (Co-orientador) e
Fábio B. Scott – UFRRJ.

23
ABSTRACT
CASTRO, Abisair Andrade de. Comparison between modified Baermann and
Donald techniques used for the recovery of gastrointestinal nematode infective
larvae from pasture. Seropédica: UFRRJ, 2004. 97p. (Tesis – Doctor Science at
Ciências Veterinárias, Parasitologia Veterinária)
Estimates of the density of larvae on herbage are used in epidemiologic studies to
determine seasonal or monthly variations in contamination of pastures with third-stage
larvae of the trichostrongyles, and to provide an index of the risk of exposure of grazing
animals. At this, a comparative study between Baermann and Donald techniques were
accomplished for the recovery of gastrointestinal ruminantes infective larvae nematodes
from pasture. To develop the assay, six faecal samples naturally infected were placed on
lawn. After 21 and 28 days, pastures samples were collected and processed for each
proposed technique. The statistic analysis showed that there is no significative
difference among the results. Thus, both techniques can be used to recover infective
larvae from pasture.
Keywords: larvae recovery, herbage, cattle, Haemonchus sp., Oesophagostomum sp.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (advisor), Laerte Grisi – UFRRJ (Co-advisor) e Fábio
Babbur S. – UFRRJ.

24
3. 1. INTRODUÇÃO
As nematodioses gastrintestinais de ruminantes ocorrem comumente em diversos
níveis de infecção, reduzindo a produtividade dos rebanhos. Estes helmintos têm
evolução direta e a fase pré-parasítica ocorre no ambiente. A estimativa do número de
larvas infectantes de tricostrongilídeos na pastagem, é parte integrante do estudo de
ecologia e epidemiologia desses nematóides parasitas de ruminantes. O controle de
nematóides gastrintestinais em ovelhas como proposto por GIBSON & EVERETT
(1968) e LANCASTER (1970) é baseado no estudo da epidemiologia de doenças, onde
a estimativa de larvas infectantes nas pastagens foi parte essencial. Similarmente
THOMAS (1959) em seu estudo sobre distribuição sazonal de Nematodirus têve a
contagem de larvas na vegetação como base. Várias técnicas têm sido descritas a fim de
avaliar a intensidade de L3 na pastagem (TAYLOR, 1939; KAUZAL, 1940;
DINABURG, 1942; CROFTON, 1954; ROHRBACHER Jr. 1957; DURIE, 1959;
DONALD, 1967; BAWDEN, 1969; LANCASTER, 1970; TODD Jr., 1970; SMEAL &
HENDY, 1972; CHIEJINA, 1982; GETTINGY et al., 1985; FINE et al. 1993). Taylor,
acima citado, foi o primeiro pesquisador a utilizar a técnica de Baermann (CORT et al.,
1922), para recuperar larvas infectantes das pastagens. Em seguida, pesquisadores como
KAUZAL (1940), DINABURG (1942), ROHRBACHER Jr. (1957) também a testaram
e como DURIE (1959) afirmaram que a mesma era ineficiente para recuperar L3 da
vegetação. DONALD (1967) descreveu e testou sua técnica verificando excelentes
percentuais de recuperação de larvas nas pastagens. Alguns pesquisadores têm utilizado
esta técnica como método apropriado para suas pesquisas (VIANA, 1999; PIMENTEL
NETO et al., 2000; ALMEIDA, 2003). Outros autores têm preservado seus princípios
como as várias lavagens da vegetação e a passagem pelo iodeto de potássio
(LANCASTER, 1970; SMEAL & HENDY, 1972; YOUNG & TRAJSTMAN, 1980).
O presente estudo foi elaborado com o objetivo de comparar as técnicas de
Baermann (CORT et al., 1922) e Donald (DONALD, 1967) quanto à recuperação de
larvas infectantes da pastagem, visando estabelecer entre as duas, o melhor protocolo
considerando para isso o custo, praticidade e qualidade da leitura.

25
3. 2. MATERIAL E MÉTODOS
3.2.1. Local:
O presente estudo foi conduzido em pastagens próximas ao prédio de Sanidade
Animal e no laboratório de Doenças Parasitárias do Instituto de Veterinária, Convênio
Embrapa - Universidade Federal Rural do Rio de Janeiro; no município de Seropédica,
localizado a 22o 54’ de longitude oeste, à altitude de 33 metros.
3.2.2. Amostras:
Aproximadamente 1,3kg de fezes foram colhidas diretamente da ampola retal de
bezerros naturalmente infectados. As amostras de fezes foram homogeneizadas, das
quais foram retiradas alíquotas para determinação do número de ovos por grama de
fezes, o.p.g., de acordo com a técnica modificada de GORDON & WHITLOCK (1939)
e, para a coprocultura (ROBERTS & O’SULLIVAN, 1959) através da qual, foram
recuperadas larvas infectantes, as quais foram identificadas utilizando-se a chave
descrita por KEITH (1953).
3.2.3. Coleta de Dados:
Para a contaminação da pastagem com larvas infectantes, seis amostras de fezes
previamente homogeneizadas com peso igual a 200g, foram depositadas a campo. A
distância entre as amostras fecais foi de aproximadamente 70cm. O local escolhido para
o depósito foi considerado homogêneo, em relação à irradiação solar, à precipitação
pluviométrica e, à vegetação, constituída por grama-batatais, Paspalum notatum.
Decorridos 21 e 28 dias, a gramínea circundante de cada amostra de fezes foi colhida
em quatro pontos distintos ao redor da mesma, numa área de 10,5 x 5,5cm (SOARES,
1980). Em seguida, as respectivas amostras de gramíneas foram homogeneizadas e
divididas em duas alíquotas de sete gramas, as quais foram processadas de acordo com
as técnicas de Baermann e Donald.
3.2.4. Descrição da técnica de Baermann:
Para o processamento pela técnica de Baermann, utilizaram-se funis de vidro
com diâmetro de aproximadamente 15cm, com borracha de látex presa à sua parte

26
posterior e fechada com uma pinça (Figura 1). Dentro do funil um tamis com diâmetro
de aproximadamente 10cm e 17/18 malhas/polegadas. As amostras de gramíneas
picadas foram envolvidas em gaze, colocadas dentro dos tamises; estes conjuntos foram
mergulhados nos funis contendo água destilada esterilizada aquecida à 50oC. Após 24
horas, cinco mililitros do sedimento foram recuperados em tubos de ensaio. Duas horas
depois, o sobrenadante foi desprezado e o sedimento montado em lâmina com solução
de lugol a 5%, e o número de L3 quantificado e identificado (KEITH, 1953).
3.2.5. Descrição da técnica de Donald:
Para o processamento da técnica de Donald, as amostras de gramínea foram
acondicionadas em potes plásticos, com 2.250ml de solução mamalian ringer’s (Figura
2a). Estas amostras foram picadas e homogeneizadas nesta solução com três gotas de
teepol, detergente não-iônico. Após seis horas, as amostras foram tamisadas em
malhas de 145 micras e deixadas em repouso em cálices de sedimentação por pelo
menos três horas (Figura 2b). A partir daí sucessivas lavagens foram feitas da seguinte
maneira: o sobrenadante foi sifonado e o líquido restante homogeneizado e passado para
cálices menores. Totalizando três passagens de cálices, com intervalo de três horas.
Depois dessa etapa as amostras passaram por duas centrifugações: a primeira com um
volume inicial de 120ml por cinco minutos a 300g. A segunda com volume de 20-30ml
por três minutos a 90g. O sedimento resultante dessa última centrifugação foi
ressuspendido em 100ml de iodeto de potássio cuja densidade foi de 1.63molar, em
cálice de separação, por dez minutos (Figura 2c). Os cinco mililitros recuperados do
sobrenadante foram destinados à leitura onde se acrescentava uma gota de hipossulfito
de sódio e as larvas quantificadas e identificadas (KEITH, 1953).
3.2.4. Análise estatística:
Os valores médios, dos números de larvas infectantes recuperadas, em cada
técnica foram comparados utilizando-se o teste de Wilcoxon, a 5% de probabilidade
(SIEGEL, 1975).
Shell do Brasil

27
Figura 1. Técnica de Baermann processando gramínea.
A
B C
Figura 2. Etapas da técnica de Donald; A - início da etapa de lavagem; B -
cálices de decantação; C - funis de separação.

28
3.3. RESULTADOS E DISCUSSÃO
De acordo com os resultados apresentados na Tabela 1, larvas infectantes do
gênero Haemonchus, e Oesophagostomum foram recuperadas pelas duas técnicas, sendo
os maiores percentuais do gênero Haemonchus. Confirmando os resultados obtidos pela
coprocultura realizada no início do experimento. Observou-se no presente estudo, que
apesar de não haver diferença significativa entre as duas técnicas; a de Baermann
recuperou maior número de larvas em sete amostras das doze testadas em relação ao
Donald (FIGURA 1). Isto se deve provavelmente a distribuição de larvas na vegetação
que foi ao acaso. Não era conhecida a quantidade de larvas na gramínea processada por
uma ou por outra técnica. É importante relatar que as leituras provenientes da técnica de
Baermann, foram, na maioria das vezes, muito difíceis, devido a grande quantidade de
sujidade residual. O mesmo foi relatado por TODD et al. (1970). Embora a técnica de
Baermann tenha apresentado maior facilidade no seu processamento, assim como, baixo
custo em relação a técnica de Donald; esta última pareceu ser mais indicada, pois no
momento da leitura a técnica de Donald foi drasticamente superior. Os resultados
encontrados neste estudo discordam das afirmativas de KAUZAL (1940),
DINAMBURG (1942), DURIE (1959), ROHRBACHER Jr., (1957), os quais atestaram
a ineficiência da técnica de Baermann. TODD et al. (1970) concluíram em seu estudo
que a técnica de Baermann é uma ferramenta útil para determinar o número de L3 no
pasto, se o pesquisador a utilizar com rigor metodológico, levando em consideração
suas limitações. CHIEJINA (1982) registrou relativa eficiência do Baermann para
recuperação de L3 da pastagem; assim como o notificado no presente estudo. Outras
técnicas para recuperação e diferenciação de L3 da pastagem foram descritas
(PARFITT, 1955; HEALTH & MAJOR, 1968; LANCASTER, 1970; SMEAL &
HENDY, 1972;). Todas exigem muito tempo de processamento e necessitam de
equipamentos específicos, difíceis de serem reproduzidos e não possuem uma alta
eficiência para recuperação de L3 da pastagem. De acordo com MARTIN et al. (1990)
entre todas as técnicas já descritas, a de Donald é uma das que conseguiram recuperar
aproximadamente 90% das larvas nas pastagens, onde seu desempenho foi independente
do número de larvas no pasto. O método mais prático para avaliar a eficiência de uma
técnica talvez seja fazer uma direta comparação com método já existente. Assumindo a
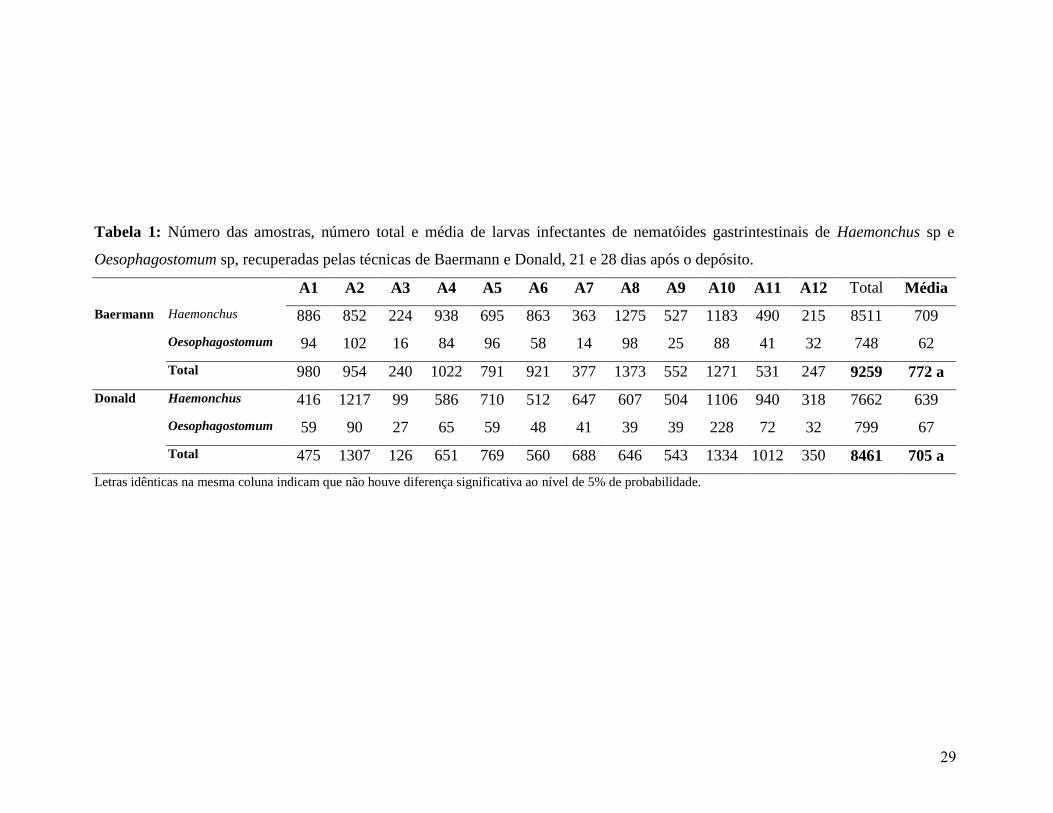
29
Tabela 1: Número das amostras, número total e média de larvas infectantes de nematóides gastrintestinais de Haemonchus sp e
Oesophagostomum sp, recuperadas pelas técnicas de Baermann e Donald, 21 e 28 dias após o depósito.
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 A11 A12 Total Média
Baermann Haemonchus 886 852 224 938 695 863 363 1275 527 1183 490 215 8511 709
Oesophagostomum 94 102 16 84 96 58 14 98 25 88 41 32 748 62
Total 980 954 240 1022 791 921 377 1373 552 1271 531 247 9259 772 a
Donald Haemonchus 416 1217 99 586 710 512 647 607 504 1106 940 318 7662 639
Oesophagostomum 59 90 27 65 59 48 41 39 39 228 72 32 799 67
Total 475 1307 126 651 769 560 688 646 543 1334 1012 350 8461 705 a
Letras idênticas na mesma coluna indicam que não houve diferença significativa ao nível de 5% de probabilidade.

30
padronização da amostragem e fazendo sub-amostras da vegetação coletada, dessa
forma providenciando material com o qual testes de comparação possam ser feitos
(LANCASTER, 1970).
Compreende-se que estudos nessa área devam continuar, visando estabelecer um
protocolo melhor, onde este possa tornar-se padrão preenchendo as exigências quanto à
economia, praticidade e principalmente eficiência.
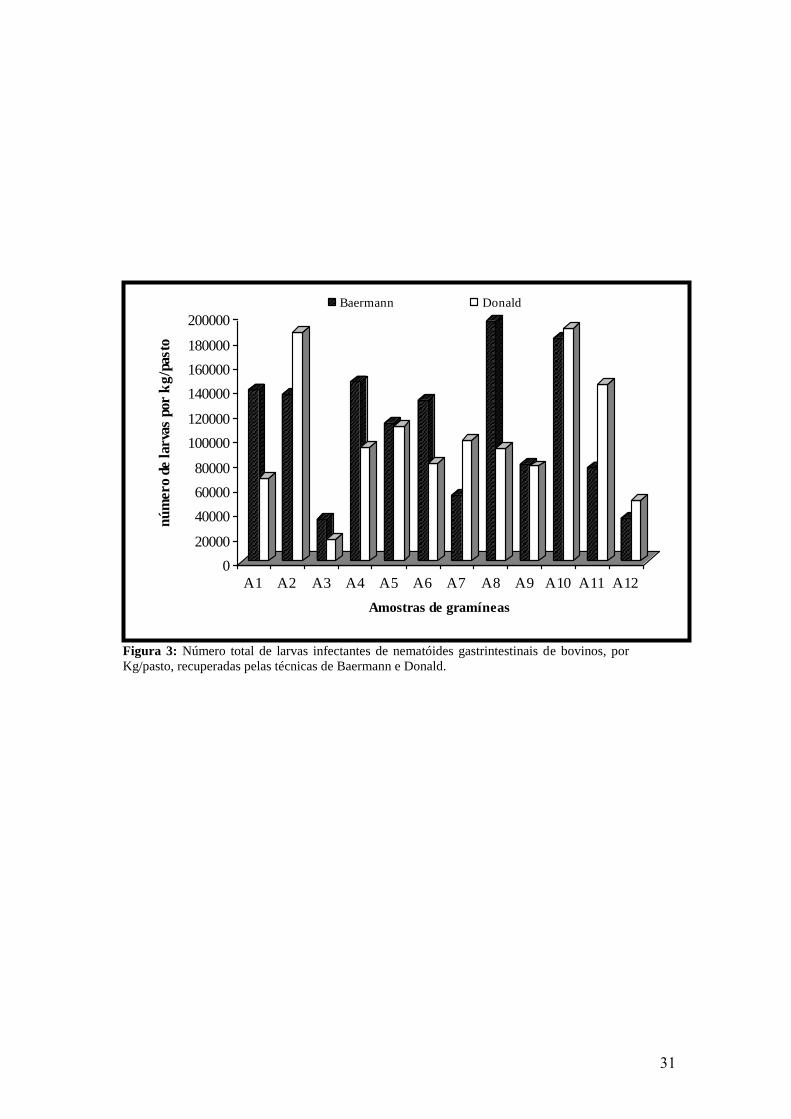
31
Figura 3: Número total de larvas infectantes de nematóides gastrintestinais de bovinos, por
Kg/pasto, recuperadas pelas técnicas de Baermann e Donald.
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
nú
mero d
e l
arva
s p
or k
g/p
asto
A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 A11 A12
Amostras de gramíneas
Baermann Donald

32
3.4. CONCLUSÕES
De acordo com os resultados apresentados, conclui-se que as duas técnicas são
eficientes para recuperação de larvas infectantes da pastagem, dando a estimativa de
larvas no ambiente, favorecendo e contribuindo para estudos de controle com base na
ecologia das larvas. Sendo a técnica de Baermann mais prática e econômica e a de
Donald de maior qualidade quanto à visualização das larvas.

33
4. CAPÍTULO II
DISTRIBUIÇÃO SAZONAL E LONGEVIDADE DE LARVAS INFECTANTES DE NEMATÓIDES
GASTRINTESTINAIS DE CAPRINOS (Capra hircus) EM PASTAGEM IRRIGADA E
NÃO-IRRIGADA.

34
RESUMO
CASTRO, Abisair Andrade de. Distribuição sazonal e Longevidade de larvas
infectantes de nematóides gastrintestinais de caprinos (Capra hircus) em pastagem
irrigada e não-irrigada. Seropédica: UFRRJ, 2004. 97p. (Tese – Doutorado em
Ciências Veterinárias, Parasitologia Veterinária)
O controle das nematodioses em caprinos tem sido realizado principalmente com
produtos anti-helmínticos. No entanto, o alto custo destes produtos e o rápido aumento
de resistência justificam a necessidade de novas alternativas. Dessa forma, o
conhecimento de alguns aspectos da epidemiologia é de extrema importância. A
distribuição sazonal e a longevidade de larvas infectantes de nematóides gastrintestinais
de caprinos em pastagem irrigada e não-irrigada foram estudadas no presente trabalho.
Dessa forma, amostras de vegetação, dos dois piquetes, não-irrigado A, e irrigado B,
foram examinadas a cada 14 dias pela técnica de Donald, durante maio/01 a abril/03. Os
resultados indicam que as condições ambientais foram favoráveis ao desenvolvimento e
longevidade de L3 de caprinos. Os gêneros Trichostrongylus e Haemonchus ocorreram
em praticamente todos os meses. Trichostrongylus mostrou maior longevidade e
Haemonchus foi o que apresentou maior densidade. O sistema de irrigação pareceu ser
favorável a longevidade de Haemonchus e Oesophagostomum, no entanto não mostrou
influência positiva no aumento de número de larvas no pasto. A ocorrência de
Oesophagostomum e Cooperia foi rara e inexistente, respectivamente.
Palavras-Chave: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp.,
irrigação, epidemiologia, pequenos ruminantes.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Orientador), Laerte Grisi – UFRRJ (Co-orientador) e
Fábio B. Scott – UFRRJ.

35
ABSTRACT
CASTRO, Abisair Andrade de. Seasonal distribution and longevity of infective
larvae of goats (Capra hircus) gastrointetinal nematodes in irrigated and non-
irrigated pastures. Seropédica: UFRRJ, 2004. 97p. (Tesis – Doctor Science at Ciências
Veterinárias, Parasitologia Veterinária)
The control of nematodiasis in goat has been usually performed with antihelmintic
agents. However, the high cost of these products and the fast increase of product’s
resistance explains the search for alternative control methods. Therefore, the knowledge
of some aspects in epidemiology is very important. The seasonal distribution and
longevity of goats gastrointestinal nematodes infective larvae in irrigated and non-
irrigated pasture were studied in the present work. Thus, pasture samples of two plots
non-irrigated A, and irrigated B, were examined every 14 days using Donald’s
technique between mai/01 and april/03. The results indicate that environment conditions
were favorable to the development and longevity of nematodes infective larvae. The
genus Trichostrongylus and Haemonchus were present on all months. Trichostrongylus
showed more longevity and Haemonchus showed more density. The irrigation system
seems to be favorable for the longevity of Haemonchus and Oesophagostomum
infective larvae. However, it did not have a positive influence at the increase of
infective larvae number in pasture. The Oesophagostomum and Cooperia occurrence
was scarce and non-existent, respectively.
Keywords: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp., irrigation,
epidemiology, small ruminants.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Advisor), Laerte Grisi – UFRRJ (Co-advisor) e Fábio
Babbur Scott – UFRRJ.

36
4. 1. INTRODUÇÃO
Os helmintos gastrintestinais constituem-se em agentes etiológicos de
significativa importância para a diminuição da produtividade na pecuária caprina
(NGINYI et al. 2001). As variações sazonais na dinâmica das populações dos helmintos
são reguladas, principalmente, pelas condições climáticas sobre os estágios de vida
livre, pela raça e pela susceptibilidade individual do hospedeiro. A interação hospedeiro
versus parasito é influenciada pela precipitação pluviométrica, faixas climáticas
favoráveis, concentração de animais por área, faixa etária e índice nutricional (GIBBS,
1973).
Segundo KATES (1965), o potencial biótico das helmintoses de ruminantes,
quando estudado e conhecido em uma determinada região, torna possível estabelecer o
seu modelo estacional, visando as dosificações estratégicas e táticas de controle.
LEVINE (1963) ao estudar o desenvolvimento e sobrevivência das larvas de H.
contortus nas pastagens, verificaram que 50mm de precipitação e 15 a 370C de
temperatura média mensal indicavam ótimas condições para a transmissão deste
parasito. No entanto, apenas os meses integrados por uma elipse no bioclimatograma
apresentavam melhor período de potencial de transmissão.
As regiões da Baixada do Estado do Rio de Janeiro correspondem à extensa área
fisiográfica, abrangendo a região Metropolitana, Baía da Ilha Grande, Baixadas
Litorâneas e parte das regiões Norte e Nordeste, coincidindo com o tipo climático AW,
segundo a classificação de KÖPPEN (1958). Essa região é caracterizada por inverno
seco e verão chuvoso e quente, com temperatura média das médias do mês mais quente
superior a 330C, e a precipitação média no verão é de 739mm, e no inverno, de 128mm.
O período seco torna-se desfavorável para a vegetação, diminuindo assim a oferta de
volumoso para os animais. Em regiões onde situação semelhante ocorre, foi implantado
sistema de irrigação como na Califórnia (BAKER et al. 1981), Guadalupe, nas ilhas
francesas (GRUNER et al. 1989), Sul da África (KRECEK et al. 1991). De acordo com
GRUNER et al. (1989) a irrigação pode mudar a epidemiologia de nematóides
gastrintestinais.
Os métodos de controle de helmintoses utilizados no Estado do Rio de Janeiro
têm sido baseados no uso de anti-helmínticos, com potencial estabelecimento de

37
populações resistentes aos princípios ativos disponíveis no mercado. Os sistemas de
manejo zootécnicos e sanitários, associados aos estudos epidemiológicos, podem
minimizar o uso de produtos químicos.
Com o objetivo de estudar a distribuição sazonal e longevidade de larvas
infectantes de nematóides gastrintestinais de caprinos, em pastagem irrigada e não-
irrigada, este trabalho foi desenvolvido visando contribuir com a aquisição de
conhecimentos e fornecer dados para futuros projetos de controle estratégico.

38
4.2. MATERIAL E MÉTODOS
4.2.1. Local:
O presente estudo foi conduzido em pastagens próximas ao prédio de Sanidade
Animal e no laboratório de Doenças Parasitárias do Instituto de Veterinária, Convênio
Embrapa - Universidade Federal Rural do Rio de Janeiro; no município de Seropédica,
localizado a 22o 54’ de longitude oeste, à altitude de 33 metros.
4.2.2. Animais e amostras de fezes:
Nove caprinos mestiços, de ambos os sexos, procedentes do Colégio Técnico da
Universidade Rural, portadores de infecção natural foram utilizados como doadores de
fezes. Estes animais permaneceram, num piquete de aproximadamente 3500m2. As
amostras de fezes foram coletadas diretamente do reto com auxílio de coletores (Figura
3A, B). Previamente ao depósito, retiravam-se alíquotas do pool de fezes coletadas, para
determinação do número de ovos por grama de fezes, o.p.g., segundo a modificada
técnica de GORDON & WHITLOCK (1939); e realização de coprocultura (ROBERTS
& O’SULLIVAN, 1950) para obtenção de larvas infectantes (Figura 5) e posterior
identificação de acordo com a chave de KEITH (1953).
4.2.3. Clima:
O clima da região, segundo a classificação de KOPPEN (1958), é do tipo AW,
com inverno seco, verão chuvoso e quente. Os dados de temperatura e precipitação
pluviométrica foram fornecidos pelo posto de meteorologia Ecologia Agrícola do Km
47, distante cerca de um km do local do experimento.
4.2.4. Piquetes:
Formaram-se dois piquetes, “A” e “B” de aproximadamente 81m2 cada,
constituídos por Paspalum notatum, grama batatais (Figura 4A - a, b ). O piquete “A” foi
estudado de acordo com as condições ambientais (Figura 4B). O piquete “B” (Figura 4C)
foi irrigado a taxa de 50mm de água por semana, durante todo o período do experimento,
não obedecendo esta regra apenas quando a precipitação pluviométrica excedia esta taxa.
Quando a mesma ocorria, sem, contudo exceder a taxa, calculava-se a diferença.

39
Figura 4. A – coletor plástico cortado em forma de bisel recebendo glicerina para facilitar a
coleta; B – amostras de fezes sendo coletadas diretamente do reto com auxílio de coletor
Figura 5. A - Piquetes de experimento; A () piquete não-irrigado, “A” com amostras; B ()
piquete irrigado, “B” com amostras.

40
4.2.5. Coleta de Dados:
Duas amostras de fezes com 35g cada, foram depositadas a campo, de 14 em 14
dias, as quais foram identificadas como “massa fecal teste” (MFT) e “massa fecal
controle” (MFC); no período de maio de 2001 a abril de 2003. As amostras distavam
cerca de 80cm umas das outras. Uma semana após o depósito, coletou-se a gramínea da
MFT, rente ao solo, numa área de 5cm de largura por 30cm de comprimento (ROSE,
1970). Logo após a coleta, a gramínea foi processada pela técnica de DONALD (1967)
para recuperação de larvas da pastagem. A MFC foi trabalhada utilizando-se a mesma
metodologia, quando a MFT correspondente apresentou-se negativa por duas vezes
consecutivas (OKON & ENYENIHI, 1977).
4.2.6. Estimativa dos dados em matéria seca:
Ao longo do experimento os dados foram coletados em peso de matéria verde.
Para calcular o correspondente em matéria seca, coletou-se durante um ano, outubro de
2002 a setembro de 2003, três amostras de vegetação de acordo com ROSE (1970).
Essas amostras foram individualmente pesadas, em seguida mantidas em estufa a 80oC
por 48 horas, sendo novamente pesadas. Com os dados médios obtidos empregou-se a
análise de regressão, obtendo-se a equação da reta, y= a + b x .
Donde: y= matéria verde;
a=0.94;
b= 0.202 e
x= matéria seca.
Através da qual estimou-se a matéria seca para todas as amostras coletadas. Para
comprovação da curva dos resultados, uma nova análise de regressão foi realizada,
confrontando-se os dados de matéria verde com os de matéria seca.
4.2.7. Análise estatística:
Para melhor aproveitamento dos resultados, os mesmos foram apresentados sob
análise de três pontos de vista: dados mês a mês; dados em quatro estações e dados em
duas estações. Para os três casos, o número médio de larvas para distribuição e o
número médio de semanas para longevidade foram transformados em Log10 (x + 2).
Com esses dados empregou-se análise de variância (ANOVA) considerando-se um nível
de 5% de probabilidade. Em seguida, para as análises que diferiram significativamente,
foi aplicado o teste de Duncan a 5%, para comparação entre as médias (VIEIRA, 1999)

41
Com a finalidade de estudar a relação entre os gêneros Trichostrongylus e
Haemonchus calculou-se o número de fêmeas, a partir do o.p.g. para cada gênero,
utilizando-se o seguinte cálculo de acordo com UENO & GONÇALVES (1998):
no de fêmeas =
o.p.g x quantidade/fezes/dias
Postura/fêmea/dia
Em seguida, empregou-se análise de correlação ao nível de 5% de probabilidade.
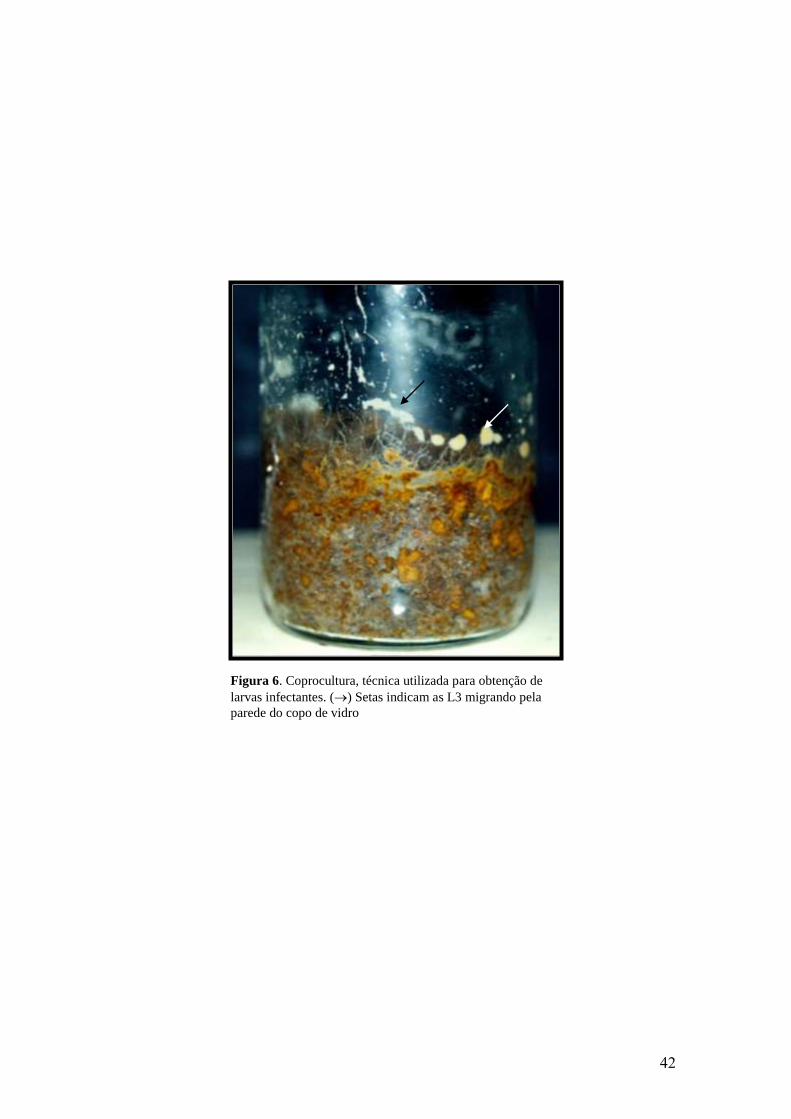
42
Figura 6. Coprocultura, técnica utilizada para obtenção de
larvas infectantes. () Setas indicam as L3 migrando pela
parede do copo de vidro

43
4. 3. RESULTADOS E DISCUSSÃO
As larvas infectantes recuperadas neste estudo foram Trichostrongylus sp,
Haemonchus sp, e Oesophagostomum (Figura 7). Sendo o maior número de larvas
recuperadas 300.612, identificadas como pertencentes ao gênero Haemonchus, seguido
por Trichostrongylus, 201.767, e Oesophagostomum, 4.344. Estes dados concordam
com PIMENTEL NETO & FONSECA (no prelo), pois estes autores trabalhando com
caprinos traçadores em Seropédica recuperaram 54.6% de Haemonchus; 41% de
Trichostrongylus e 2% de Oesophagostomum.
A recuperação de larvas do gênero Oesophagostomum foi considerada baixa nos
dois piquetes, durante todo o período do experimento. Por esse motivo não se aplicou
teste estatístico a esse gênero. Resultados semelhantes foram encontrados por HORAK
& LOWN (1977), no sul da África, OKON & ENYENIHI (1977) na Nigéria,
AUMONT & GRUNER (1989) em Guadalupe, nas ilhas francesas, BOJANE et al.
(2002) na Índia, PIMENTEL NETO & FONSECA (no prelo) em Seropédica, Brasil.
Larvas de Oesophagostomum, no presente estudo, foram recuperadas em sete dos 24
meses de experimento no piquete A: outubro/01, março/02, maio/02, junho/02,
setembro/02, março/03 e abril/03. Já no piquete B ocorreram em oito dos 24 meses:
novembro/01, março/02, maio/02, junho/02, julho/02, agosto/02, março/02 e abril/03. O
maior período de sobrevivência das larvas de Oesophagostomum foi no piquete B, mês
de maio, chegando a seis semanas.
Larvas pertencentes ao gênero Cooperia não foram recuperadas em nenhum dos
meses ao longo deste estudo. TRIPATHI (1970b) registrou a rara ocorrência desse
gênero em caprinos na Índia; assim como FABIYI (1970) na Nigéria, GRISI (1975) na
Bahia, BOUGAS & THEODORIDIS (2000) na Grécia; PIMENTEL NETO &
FONSECA (no prelo) em Seropédica, Brasil. VASQUEZ & MARCHINARES (1971)
não registraram a presença desse gênero, parasitando caprinos no Peru. Da mesma
forma CAVALCANTI (1974) e CHARLES (1989) em Pernambuco, AUMONT &
GRUNER (1989) em Guadalupe, BAKER (1975) na Califórnia, e SILVA et al. (1998)
também no nordeste brasileiro.

44
F E
A
D C
B
Figura 7: Larvas infectantes encontradas no presente estudo. São
respectivamente: A e B: extremidade anterior e posterior de
Trichostrongylus sp.; C e D: extremidade anterior e posterior de
Haemonchus sp. e E e F: extremidade anterior e posterior de
Oesophagostomum sp.

45
Vários pesquisadores como AL SAQUR et al (1982), CALLINAN &
WESTCOTT (1986), AUMONT et al. (1989), AUMONT & GRUNER (1989), BANKS
et al. (1990), KRECEK et al (1992), BARGER et al (1994) entre outros, apresentaram
os dados de seus resultados em número de larvas por kg de matéria seca. No entanto,
autores como HOLOSOVÁ et al. (1989), RAHMAN & COLLINS (1990), PIMENTEL
NETO et al. (2000), ALMEIDA (2003) apresentaram seus resultados em número de
larvas por kg de matéria verde. No presente estudo foi verificada a relação entre as duas
formas de apresentação de resultados e de acordo com os cálculos estatísticos da análise
de regressão (r2= 0.97), constatou-se que os valores transformados de matéria verde
para matéria seca, embora aumentados, mantinham a mesmo modelo de curva gráfica
(figura 8).
Os números de larvas recuperadas da vegetação e o período de sobrevivência em
semanas, não apresentaram diferença significativa entre os dois piquetes (Figura 9A e
B), embora, os valores tenham sido superiores no piquete B, para todos os gêneros
encontrados neste estudo. A única exceção foi descrita ao número total de L3 de
Haemonchus, onde o maior valor registrou-se no piquete B, demonstrando assim, uma
possível atuação da irrigação favorecendo o desenvolvimento destas larvas. No entanto,
quanto à sobrevivência, a irrigação não otimizou o período de longevidade destas.
Mesmo que não se possa afirmar estatisticamente, o sistema de irrigação pareceu
favorecer a distribuição de larvas de Oesophagostomum. De uma forma geral estes
resultados discordam dos achados dos autores STWART & DOUGLAS (1938),
FURMAN (1944), BULLISK & ANDERSEN (1978), HORAK & LOWN (1977),
BAKER et al. (1981), GRUNER et al. (1989), os quais afirmaram que a irrigação
favorecia o desenvolvimento e sobrevivência de larvas de tricostrongilídeos.
VALDERRABANO et al. (2000) observaram que a irrigação favorecia a pastagem,
podendo ser utilizada contra problemas climáticos aumentando a produtividade dos
rebanhos. No presente estudo, a vegetação do piquete B apresentou-se viçosa durante
todo o período do experimento (Figura 10).
GORDON (1967) observou o fenômeno de competição entre espécies do gênero
Trichostrongylus e Haemonchus. PIMENTEL NETO (1976) observou retardamento no
desenvolvimento do ciclo biológico de H. contortus quando ocorreu infecção
concomitante com T. axei. REINECKE (1989) relatou que a contaminação prévia por T.

46
T r i c h o s tr o n g yl u s s p .
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
5 0 0 0 0
6 0 0 0 0
J F M A M J J A S O N DL
3 /
Kg
ms
O e s o p h a g o s to m u m s p .
0
2 0 0
4 0 0
6 0 0
8 0 0
1 0 0 0
1 2 0 0
J F M A M J J A S O N D
L3
/K
g m
s
O e s o p h a g o s to m u m s p .
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
J F M A M J J A S O N D
L3
/ K
g m
v
H a e m o n c h u s s p .
0
2 0 0 0 0
4 0 0 0 0
6 0 0 0 0
8 0 0 0 0
1 0 0 0 0 0
1 2 0 0 0 0
J F M A M J J A S O N D
L3
/K
g m
s
T r i c h o s tr o n g yl u s s p .
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
J F M A M J J A S O N D
L3
/K
g m
v
H a e m o n c h u s s p .
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
J F M A M J J A S O N D
L3
/K
g m
v
Figura 8: Curvas gráficas dos números de L3/kg de pasto de Trichostrongylus, Haemonchus e Oesophagostomum,
apresentados à esquerda, em matéria seca (ms) e à direita, em matéria verde (mv).

47
Figura 9: A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos entre os piquetes A, não-
irrigado e B, irrigado. B. Longevidade de larvas infectantes de nematóides gastrintestinais de caprinos entre os piquetes
A, não-irrigado e B, irrigado.
T r i c h o s tr o n g yl u s s p .
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
5 0 0 0 0
6 0 0 0 0
7 0 0 0 0
8 0 0 0 0
J F M A M J J A S O N D
L3
Kg
/ms
A a
B a
H a e m o n c h u s s p .
0
2 0 0 0 0
4 0 0 0 0
6 0 0 0 0
8 0 0 0 0
1 0 0 0 0 0
1 2 0 0 0 0
J F M A M J J A S O N D
L3
Kg
/ms
A a
B b
O e s o p h a g o s to m u m s p .
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
J F M A M J J A S O N D
L3
Kg
/ms
A
B
T r i c h o s tr o n g yl u s s p .
0
5
1 0
1 5
2 0
J F M A M J J A S O N D
se
ma
na
s
A a
B a
H a e m o n c h u s s p .
0
5
1 0
1 5
2 0
J F M A M J J A S O N D
se
ma
na
s
A a
B a
O e s o p h a g s to m u m s p .
0
1
2
3
4
5
6
7
J F M A M J J A S O N D
se
ma
na
s
A
B
A B

48
Figura 10. Pastagens: A Amostra no piquete A, não-irrigado; B. Amostra no piquete B, irrigado.

49
axei fez com que houvesse supressão de novas infecções pelo H. contortus ou que as L4
sofressem parada de crescimento. PIMENTEL NETO & FONSECA (no prelo)
comentaram a possibilidade de existência de competição entre os gêneros
Trichostrongylus e Haemonchus em caprinos. No presente estudo, verificou-se que não
houve relação entre os dois gêneros, ou seja, que ambos ocorreram independentemente,
quando se analisou a estimativa do número de fêmeas (r2=0.02), embora as curvas de
distribuição do número de L3, o.p.g. e fêmea de Trichostrongylus e Haemonchus, nos
dois piquetes tenha mostrado uma tendência a esta hipótese (Figura 11). Fato este que
pode ser justificado pela simples diferença de ovopostura das respectivas fêmeas
adultas: a de Trichostrongylus é capaz de liberar 200 ovos/dia enquanto que uma fêmea
de Haemonchus libera 5000 ovos/dia (UENO & GONÇALVES, 1998).
4.3.1. Dados apresentados mês a mês:
As larvas infectantes do gênero Trichostrongylus apresentaram distribuição e
longevidade significativamente diferentes entre os meses. Estes dados estão
sucintamente apresentados na tabela 2. A curva gráfica dos números médios totais e dos
dados médios transformados para os parâmetros acima citados, e os dados climáticos
estão representados nas figuras 12 e 13, respectivamente. A distribuição de L3 de
Trichostrongylus entre os meses, foi a seguinte: o número de larvas recuperadas no mês
de janeiro foi significativamente diferente dos meses de junho, julho, agosto, setembro,
outubro, novembro, sendo que nestes foram registrados os maiores números de L3. O
mês de fevereiro foi considerado diferente dos meses de março, agosto e novembro.
Março diferiu de todos com exceção de janeiro. Abril foi considerado diferente de
março, agosto e novembro. O mês de maio diferiu apenas de março. Os meses de junho,
julho, setembro e outubro foram considerados diferentes apenas de janeiro e março.
Agosto e novembro diferiram estatisticamente de janeiro, fevereiro, março e abril.
Finalizando, o mês de dezembro diferiu apenas do mês de março. O mês que no qual
recuperou-se maior quantidade de L3 foi novembro com cerca de 56.484 larvas por kg
de matéria seca.
O período de sobrevivência das larvas de Trichostrongylus apresentou-se da
seguinte forma: o mês de janeiro diferiu significativamente dos meses de abril, maio,
junho, julho quanto ao número de semanas que as larvas sobreviveram; considerando
que estes foram os que apresentaram maior período de longevidade. Fevereiro diferiu
dos meses de março a outubro, inclusive. Março diferiu apenas de fevereiro e maio.
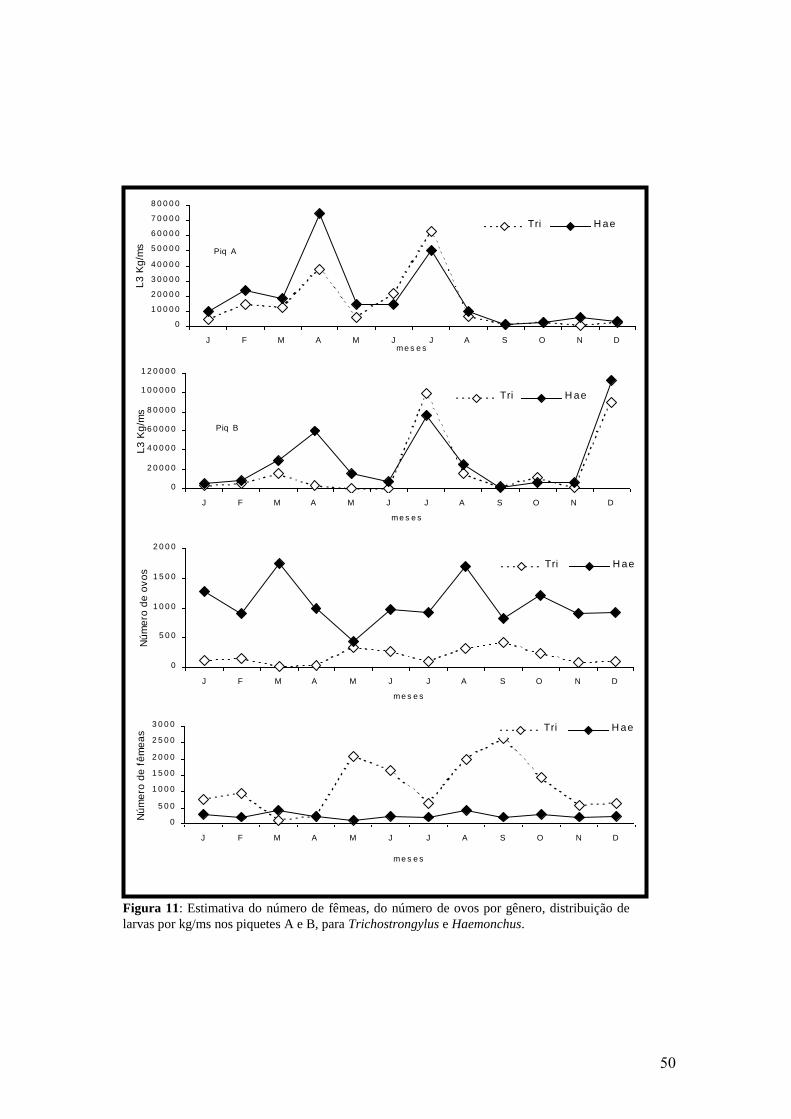
50
Piq A
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
5 0 0 0 0
6 0 0 0 0
7 0 0 0 0
8 0 0 0 0
J F M A M J J A S O N Dm e s e s
L3
Kg
/ms
Tri H ae
Piq B
0
2 0 0 0 0
4 0 0 0 0
6 0 0 0 0
8 0 0 0 0
1 0 0 0 0 0
1 2 0 0 0 0
J F M A M J J A S O N D
m e s e s
L3
Kg
/ms
Tri H ae
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
J F M A M J J A S O N D
m e s e s
Nú
me
ro d
e o
vo
s
Tri H ae
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
2 5 0 0
3 0 0 0
J F M A M J J A S O N D
m e s e s
Nú
me
ro d
e f
êm
ea
s
Tri H ae
Figura 11: Estimativa do número de fêmeas, do número de ovos por gênero, distribuição de
larvas por kg/ms nos piquetes A e B, para Trichostrongylus e Haemonchus.
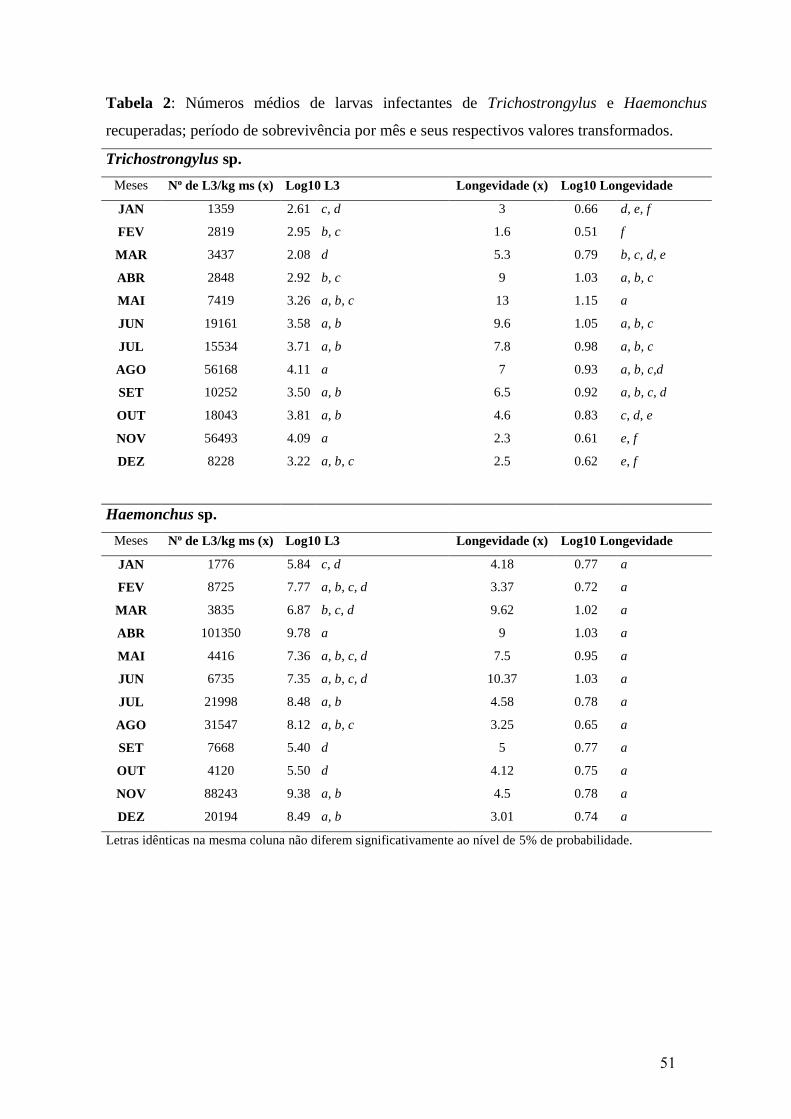
51
Tabela 2: Números médios de larvas infectantes de Trichostrongylus e Haemonchus
recuperadas; período de sobrevivência por mês e seus respectivos valores transformados.
Trichostrongylus sp.
Meses No de L3/kg ms (x) Log10 L3 Longevidade (x) Log10 Longevidade
JAN 1359 2.61 c, d 3 0.66 d, e, f
FEV 2819 2.95 b, c 1.6 0.51 f
MAR 3437 2.08 d 5.3 0.79 b, c, d, e
ABR 2848 2.92 b, c 9 1.03 a, b, c
MAI 7419 3.26 a, b, c 13 1.15 a
JUN 19161 3.58 a, b 9.6 1.05 a, b, c
JUL 15534 3.71 a, b 7.8 0.98 a, b, c
AGO 56168 4.11 a 7 0.93 a, b, c,d
SET 10252 3.50 a, b 6.5 0.92 a, b, c, d
OUT 18043 3.81 a, b 4.6 0.83 c, d, e
NOV 56493 4.09 a 2.3 0.61 e, f
DEZ 8228 3.22 a, b, c 2.5 0.62 e, f
Haemonchus sp.
Meses No de L3/kg ms (x) Log10 L3 Longevidade (x) Log10 Longevidade
JAN 1776 5.84 c, d 4.18 0.77 a
FEV 8725 7.77 a, b, c, d 3.37 0.72 a
MAR 3835 6.87 b, c, d 9.62 1.02 a
ABR 101350 9.78 a 9 1.03 a
MAI 4416 7.36 a, b, c, d 7.5 0.95 a
JUN 6735 7.35 a, b, c, d 10.37 1.03 a
JUL 21998 8.48 a, b 4.58 0.78 a
AGO 31547 8.12 a, b, c 3.25 0.65 a
SET 7668 5.40 d 5 0.77 a
OUT 4120 5.50 d 4.12 0.75 a
NOV 88243 9.38 a, b 4.5 0.78 a
DEZ 20194 8.49 a, b 3.01 0.74 a
Letras idênticas na mesma coluna não diferem significativamente ao nível de 5% de probabilidade.
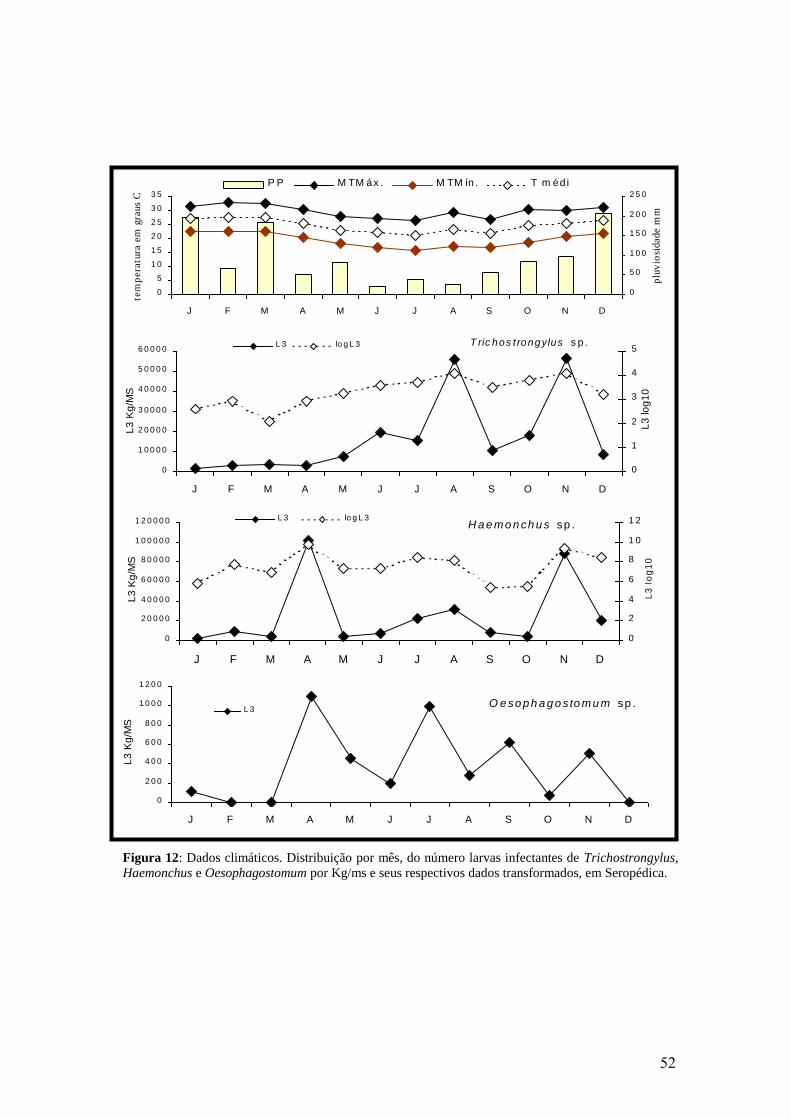
52
H a e m o n ch u s sp .
0
2 0 0 0 0
4 0 0 0 0
6 0 0 0 0
8 0 0 0 0
1 0 0 0 0 0
1 2 0 0 0 0
J F M A M J J A S O N D
L3
Kg
/MS
0
2
4
6
8
1 0
1 2
L3
lo
g1
0
L 3 lo g L 3
T ric hos t rongylus s p .
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
5 0 0 0 0
6 0 0 0 0
J F M A M J J A S O N D
L3
Kg
/MS
0
1
2
3
4
5
L3
lo
g1
0
L 3 lo g L 3
0
5
1 0
1 5
2 0
2 5
3 0
3 5
J F M A M J J A S O N D
tem
pera
tura
em
gra
us
C
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
plu
vio
sidade m
m
P P M TM áx . M TM ín . T m éd i
O e so p h a g o sto m u m sp .
0
2 0 0
4 0 0
6 0 0
8 0 0
1 0 0 0
1 2 0 0
J F M A M J J A S O N D
L3
Kg
/MS
L 3
Figura 12: Dados climáticos. Distribuição por mês, do número larvas infectantes de Trichostrongylus,
Haemonchus e Oesophagostomum por Kg/ms e seus respectivos dados transformados, em Seropédica.
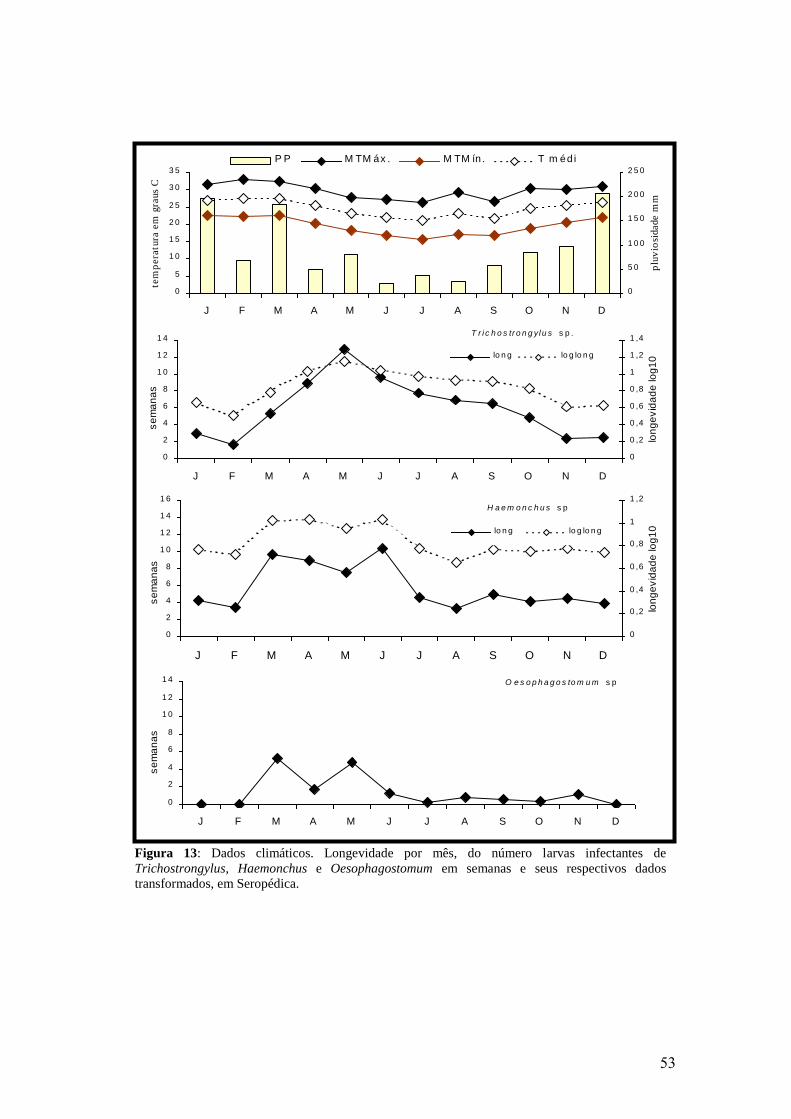
53
H a e m o n c h u s s p
0
2
4
6
8
1 0
1 2
1 4
1 6
J F M A M J J A S O N D
se
ma
na
s
0
0 ,2
0 ,4
0 ,6
0 ,8
1
1 ,2
lon
ge
vid
ad
e lo
g1
0lo n g lo g lo n g
O e s o p h a g o s to m u m s p
0
2
4
6
8
1 0
1 2
1 4
J F M A M J J A S O N D
se
ma
na
s
0
5
1 0
1 5
2 0
2 5
3 0
3 5
J F M A M J J A S O N D
tem
pera
tura
em
gra
us
C
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
plu
vio
sidade m
m
P P M TM áx . M TM ín . T m éd i
T r i c h o s tr o n g yl u s s p .
0
2
4
6
8
1 0
1 2
1 4
J F M A M J J A S O N D
se
ma
na
s
0
0 ,2
0 ,4
0 ,6
0 ,8
1
1 ,2
1 ,4
lon
ge
vid
ad
e lo
g1
0lo n g lo g lo n g
Figura 13: Dados climáticos. Longevidade por mês, do número larvas infectantes de
Trichostrongylus, Haemonchus e Oesophagostomum em semanas e seus respectivos dados
transformados, em Seropédica.

54
Abril foi considerado diferente de janeiro, fevereiro, novembro e dezembro. Maio
diferiu de janeiro, fevereiro, março, outubro, novembro e dezembro. Junho e julho
diferiram dos meses de janeiro, fevereiro, novembro e dezembro. Agosto e setembro
diferiram significativamente dos meses de fevereiro, novembro e dezembro. O mês de
outubro diferiu apenas dos meses de fevereiro e maio. Novembro e dezembro diferiram
dos meses de abril a setembro, inclusive. O mês de depósito em que as larvas
apresentaram maior período de longevidade foi maio, com 13 semanas.
A distribuição do número de larvas infectantes de Haemonchus diferiu
significativamente entre os meses de experimento da seguinte forma: o mês de janeiro
diferiu dos meses de abril, julho, novembro e dezembro. Fevereiro não diferiu de
nenhum dos meses. O mês de março foi diferente apenas para o mês de abril. Abril
apresentou-se significativamente diferente de janeiro, março, setembro e outubro.
Apresentaram distribuição semelhante, os meses de maio e junho, os quais não
diferiram dos demais meses. Julho diferiu apenas de janeiro, setembro e outubro.
Agosto também diferiu dos meses de setembro e outubro. Os meses de setembro e
outubro foram semelhantes, diferindo apenas de abril, julho, agosto, novembro e
dezembro. Também foram semelhantes os meses de novembro e dezembro diferindo
apenas de janeiro, setembro e outubro. O maior número de larvas infectantes de
Haemonchus recuperadas da vegetação foi registrado no mês de abril chegando
aproximadamente 101.350 L3 por kg de matéria seca.
A longevidade das larvas de Haemonchus não apresentou diferença significativa
entre os meses, embora o maior período de sobrevivência, dez semanas, tenha sido
registrado em junho.
A distribuição de Haemonchus no presente estudo reflete que este gênero
ocorreu praticamente durante todo o ano. Na Nigéria, OKON & ENYENIHI (1977)
estudaram o desenvolvimento e sobrevivência de H. contortus. Eles registraram a
presença de L3 no pasto apenas nos meses de outubro/73, maio/74, junho/74, julho/74,
setembro/74 e outubro/74 e nestes meses os respectivos períodos de longevidade quatro,
seis, seis, cinco, nove e sete semanas.
Mesmo com valores baixos, a distribuição do número médio de larvas do gênero
Oesophagostomum nos dois piquetes indicou a ocorrência das larvas durante o
experimento, sendo os meses de abril e julho àqueles em que se recuperou o maior
número de larvas. A longevidade destas L3 foi de no máximo cinco semanas em março
e maio.

55
REINECKE (1970) comentou que as fases pré-parasíticas de tricostrongilídeos
estariam sujeitas a fatores como temperaturas, precipitações pluviométricas, tipo de
solo, vegetação. No presente estudo, a temperatura pareceu ser um fator limitante para o
gênero Trichostrongylus, pois nos meses mais quentes, novembro a abril, cuja média foi
em torno dos 270C, o número de L3 recuperado e a longevidade foi significativamente
menor em relação aos outros meses, quando a temperatura apresentou-se mais amena,
em torno dos 220C. A precipitação não pareceu ter influência clara sobre este gênero.
OKON & ENYENIHI (1977) associaram a distribuição de Haemonchus aos dados de
precipitação, informando ser este o principal fator. A distribuição de larvas de
Haemonchus no presente estudo, não pareceu ser influenciada diretamente pela
precipitação. Estes dados concordam com os achados de AUMONT et al. (1989). Eles
comentaram que não foi clara a relação entre os dados climáticos com a dinâmica da
população de larvas. Provavelmente existiram outros fatores que influenciaram a
distribuição e longevidade das larvas infectantes de tricostrongilídeos, neste estudo. De
acordo com GIBBIS (1973), LEVINE (1963) fatores do microambiente podem ter
importante papel no desenvolvimento e sobrevivência das L3. ROSEMBERG (1974)
definiu microclima como o clima onde vivem animais e plantas. RIBEIRO (1992)
observou que as temperaturas do microclima embora fossem mais elevadas, mantinham
a mesma curva gráfica dos dados do macroclima. Talvez as condições do
microambiente possam ter uma influência mais perceptível, justificando a distribuição e
o período de longevidade das larvas de forma mais clara.
Embora boa parte dos pesquisadores como TRIPATHI (1970a,b), FABIYI
(1973), CAVALCANTI (1974) tenham trabalhado por um período de um ano ou mais,
seus resultados foram comumente apresentados por estações, sejam elas quatro ou duas
de acordo com o ponto de vista de cada um. Por esse motivo não foi possível discutir
esses dados com os dos referidos pesquisadores, pois nesse tópico apresentou-se os
dados mês a mês. No entanto pode-se dizer que estes dados estão de acordo com os
achados de PIMENTEL NETO et al. (2000) e PIMENTEL NETO & FONSECA (no
prelo), pois estes pesquisadores trabalharam com tricostrongilídeos de bovinos e
caprinos, respectivamente na região de Seropédica e observaram que as condições locais
foram consideradas propícias, uma vez que estas larvas foram encontradas o ano todo
assim como no presente estudo.

56
4.3.2. Dados apresentados em quatro estações:
A distribuição de larvas por kg de matéria seca e a longevidade em número de
semanas das larvas recuperadas neste estudo, nas quatro estações convencionais, estão
representadas nas figuras 14 A e B. As estações mais quentes foram o verão e outono
com a média de temperatura das máximas em torno dos 32 e 300C, respectivamente
(Figura 15). Estas estações, no entanto, foram as mais chuvosas, registrando-se 156 e
104 mm de precipitação pluviométrica, respectivamente.
Os três gêneros encontrados neste estudo ocorreram nas quatro estações.
Trichostrongylus apresentou significativo desenvolvimento no inverno e primavera. Já
longevidade das larvas desse gênero foi significativamente superior nas estações do
outono e inverno. O que demonstra que a temperatura talvez tenha sido um fator
limitante para esse gênero, como afirmou LEVINE (1963) sobre o desenvolvimento de
tricostrongilídeos. TRIPATHI (1970b) observou que o período quente foi desfavorável
ao desenvolvimento e sobrevivência das larvas no ambiente. A precipitação
pluviométrica não pareceu ser um fator de importância para esse gênero, pois as
estações chuvosas foram justamente àquelas, nas quais o número de larvas de
Trichostrongylus foi mais baixo.
Foi registrado para o gênero Haemonchus, que os maiores números de larvas
recuperadas foram nas estações do outono e primavera, embora estatisticamente não
tenha ocorrido diferença entre as quatro estações. Observou-se, porém, que houve
diferença estatística para a longevidade dessas larvas entre as estações; sendo o outono
considerado a melhor época para a sobrevivência das larvas. DINABURG (1944)
contaminou artificialmente as pastagens com H. contortus e verificou que nos meses do
verão as condições foram desfavoráveis para a sobrevivência das larvas. Resultados
equivalentes foram registrados no presente estudo. SKINNER & TODD Jr. (1980)
estudaram a migração lateral de H. contortus nas pastagens e concluíram que quando a
temperatura diminuía, a sobrevivência das larvas aumentavam. Talvez seja essa a
justificativa para o outono ser a melhor época para a sobrevivência das larvas.
Os resultados presente estudo estão de acordo com LEVINE & TODD (1975).
Eles estudaram o desenvolvimento e sobrevivência de larvas de H. contortus e T.
colubriformis. Os autores observaram que poucas larvas de H. contortus sobreviveram
no inverno, e ao contrário T. colubriformis sobreviveu bem nesta estação; e que o
outono foi a melhor época para a sobrevivência de ambas espécies. Os autores

57
Figura 14: A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos em quatro estações. B.
Longevidade de larvas infectantes de nematóides gastrintestinais de caprinos em quatro estações. (Letras, minúsculas,
idênticas não diferem ao nível de 5% de probabilidade.).
O e s o p h a g o s to m u m s p .
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
v e r o u t in v p r i
L3
/ k
g m
s
H a e m o n c h u s s p .
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
v e r o u t in v p r i
L3
/ k
g m
s
2 ,6
2 ,8
3
3 ,2
3 ,4
3 ,6
Lo
g1
0-
L3
a
a a
a
T r i c h o s to n g yl u s s p .
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
3 0 0 0 0
3 5 0 0 0
v e r o u t in v p r i
L3
/ k
g m
sb b
a a
O e s o p h a g o s to m u m s p .
0
1
2
3
4
5
v e r o u t in v p r i
se
ma
na
s
T r i c h o s tr o n g yl u s s p .
0
2
4
6
8
1 0
v e r o u t in v p r i
se
ma
na
s
c
a
b
a
H a e m o n c h u s s p .
0
2
4
6
8
1 0
v e r o u t in v p r i
se
ma
na
s
b
a
b
b
A B
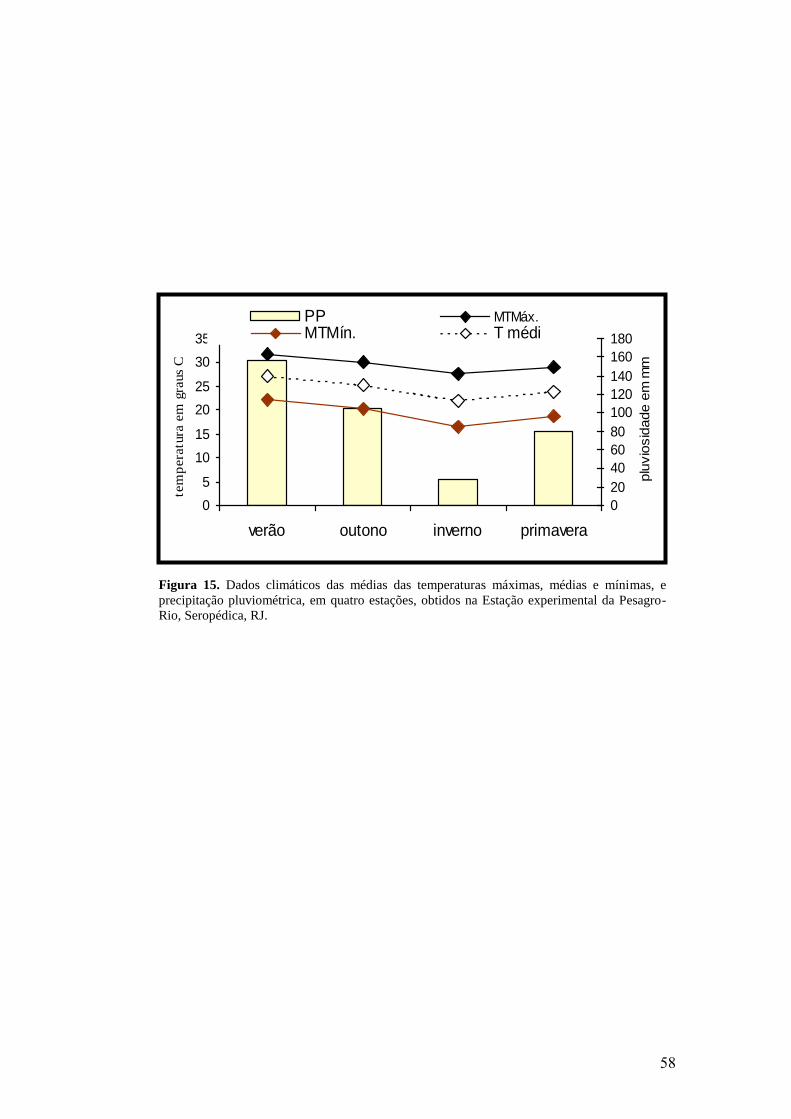
58
0
5
10
15
20
25
30
35
verão outono inverno primavera
tem
pera
tura
em
gra
us
C
0
20
40
60
80
100
120
140
160
180
plu
vio
sid
ade e
m m
m
PP MTMáx.
MTMín. T médi
Figura 15. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, em quatro estações, obtidos na Estação experimental da Pesagro-
Rio, Seropédica, RJ.

59
comentaram que as larvas de H. contortus apresentaram maior resistência quando
submetidas a repetidas dessecações do que as de T. colubriformis.
LEVINE (1963) descreveu que larvas do gênero Trichostrongylus foram mais
resistentes na fase larval do que os gêneros Haemonchus e Oesophagostomum. No
presente estudo essa diferença não foi registrada para os dois primeiros gêneros acima
citados, uma vez que ambos ocorrem durante todas as estações e em praticamente todos
os meses. No entanto, com relação a Oesophagostomum, existe concordância com a
afirmativa acima, uma vez que essas larvas foram recuperadas em maior número apenas
na estação do inverno e o maior período de sobrevivência foi de quatro semanas no
outono.
4.3.3. Dados apresentados nos períodos chuvoso e seco:
Os dados de distribuição e de longevidade das larvas de nematóides
gastrintestinais expressos em dois períodos, chuvoso e seco, estão sucintamente
apresentados nas figuras 16A e B, respectivamente. Os dados climáticos divididos em
duas estações estão representados na figura 17. Os três gêneros ocorreram nos dois
períodos estudados. Não houve diferença significativa entre os períodos para nenhum
dos gêneros estudados, embora Trichostrongylus tenha apresentado maior número de
larvas, e maior tempo de sobrevivência no período seco. Haemonchus ao contrário
apresentou o maior número de larvas recuperadas na época das chuvas e a longevidade
das mesmas foi semelhante nos dois períodos. Autores como TRIPATHI (1970a),
FABIYI (1973), CAVALCANTI (1974), GRISE (1975), CONNOR et al. (1990),
LAHA et al. (2001) registraram a importância do período das chuvas nas infecções por
tricostrongilídeos; assim como McCULLOCK & KASIMBALA (1968) registraram que
H. contortus apresentou seu pico no período chuvoso; este dado não concorda com o
achado neste estudo. McCULLOCK & KASIMBALA (1968) afirmaram ainda, que o
clima representou um fator de pouca importância nas infecções por T. colubriformis e
O. columbianum. No presente estudo verificou-se que as condições podem atuar
limitando ou favorecendo larvas de Trichostrongylus e Oesophagostomum.
Para acompanhar a dinâmica populacional dos estágios de vida livre de
nematóides gastrintestinais de caprinos, numa região de clima tropical, AUMONT &
GRUNER (1989) contaminaram piquetes com ovos de helmintos, durante a estação
chuvosa e seca. Eles recuperaram larvas de Haemonchus e Trichostrongylus e
observaram que as larvas surgiram na vegetação sete a 14 dias depois do depósito. Este

60
Figura 16.A. Distribuição nos períodos chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de
caprinos, expressos número de L3/Kg matéria seca e em logaritmo do número de L3. B. Longevidade nos períodos
chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de caprinos expressos em semanas e seus
respectivos dados transformados em logaritmo (Letras, minúsculas, idênticas não diferem ao nível de 5% de
probabilidade.).
T r i c h o s tr o n g yl u s s p .
0
4 0 0 0
8 0 0 0
1 2 0 0 0
1 6 0 0 0
c h u v o s a s e c a
L3
/ kg
ms
2 ,9
3
3 ,1
3 ,2
3 ,3
3 ,4
3 ,5
3 ,6
Lo
g1
0-L
3
H a e m o n c h u s s p .
1 8 0 0 0
1 9 0 0 0
2 0 0 0 0
2 1 0 0 0
2 2 0 0 0
c h u v o s a s e c a
L3
/ kg
ms
7
7 ,1
7 ,2
7 ,3
7 ,4
7 ,5
7 ,6
7 ,7
7 ,8
Lo
g1
0-L
3
O e s o p h a g o s to m u m s p .
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
c h u v o s a s e c a
L3
/ kg
ms
a
a
a
a
O e s o p h a g o s to m u m s p .
0
0 ,5
1
1 ,5
2
2 ,5
c h u v o s a s e c a
se
ma
na
s
T r i c h o s tr o n g yl u s s p .
0
2
4
6
8
1 0
c h u v o s a s e c a
se
ma
na
s
0
2
4
6
8
1 0
Lo
g1
0-
se
ma
na
s
a
a
H a e m o n c h u s s p .
0
1
2
3
4
5
6
7
c h u v o s a s e c a
se
ma
na
s
A B
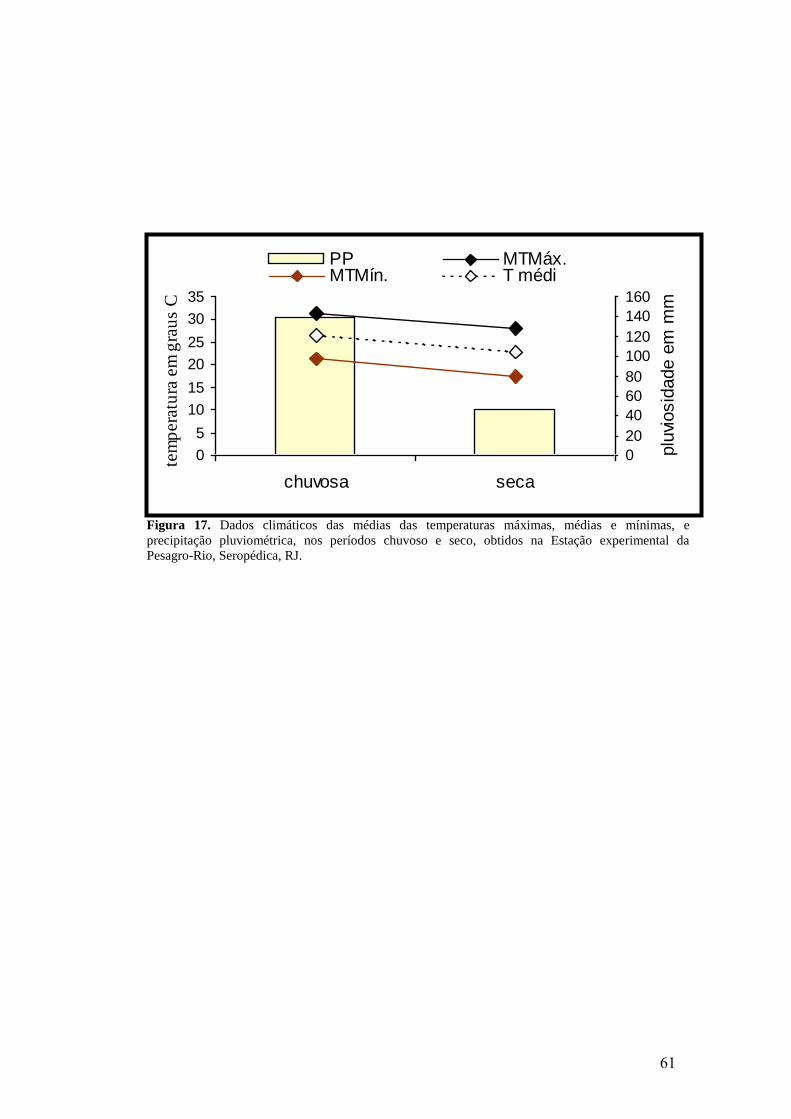
61
0
5
10
15
20
25
30
35
chuvosa seca
tem
pera
tura
em
gra
us C
0
20
40
60
80
100
120
140
160
plu
viosid
ade e
m m
m
PP MTMáx.MTMín. T médi
Figura 17. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, nos períodos chuvoso e seco, obtidos na Estação experimental da
Pesagro-Rio, Seropédica, RJ.

62
fato foi observado no presente estudo. Os autores relataram que a longevidade das
larvas na vegetação foi de 49 a 56 dias, sete a oito semanas. No presente estudo as
larvas sobreviveram até nove semanas. Consideraram que os eventos climáticos e o fato
da vegetação apresentar-se seca durante o pastoreio, foram fatores importantes,
parecendo interferir na dinâmica populacional das formas de vida livre destes helmintos.
O gênero Oesophagostomum apresentou seu pico na época da seca e seu maior
período de sobrevivência foi de duas semanas para os dois períodos. O clima pareceu
interferir na ecologia das formas pré-parasíticas dessas larvas.

63
4. 4. CONCLUSÕES
Pode-se concluir, de acordo com os resultados apresentados que:
1. As condições climáticas de Seropédica foram favoráveis ao desenvolvimento e
longevidade de larvas infectantes de nematóides gastrintestinais de caprinos;
2. Os gêneros Trichostrongylus e Haemonchus ocorreram em praticamente todos os
meses de período do experimento; sendo que em termos de quantidade, Haemonchus foi
o que apresentou maior número, e em longevidade, Trichostrongylus, foi o que
apresentou o maior período de sobrevivência;
3. A ocorrência dos gêneros Oesophagostomum e Cooperia foi considerada,
respectivamente, baixa e inexistente, no período estudado;
4. Não há diferença entre a apresentação dos resultados como matéria verde ou matéria
seca;
5. Foi constatado que o sistema de irrigação não favoreceu o desenvolvimento e a
longevidade de L3 de nematóides gastrintestinais de caprinos, com exceção ao número
total de larvas de Haemonchus, que foi significativamente maior no piquete irrigado.
6. Não houve relação entre os gêneros Trichostrongylus e Haemonchus. Eles ocorrem
independentemente.
7. Os meses de agosto e novembro foram os que apresentaram maior número de larvas
infectantes de Trichostrongylus; e o maior período de sobrevivência foi no mês maio.;
8. Os meses de abril e novembro foram os que apresentaram maior número de larvas
infectantes de Haemonchus; e o maior período de sobrevivência foi registrado nos
meses de março e junho;

64
9. Com análise nas quatro estações Trichostrongylus apresentou maior recuperação de
larvas no inverno e primavera;
10. Não houve diferença entre as estações, para distribuição e longevidade de larvas de
Haemonchus;
11. Outono foi a época mais favorável à sobrevivência das larvas infectantes de
nematóides gastrintestinais de caprinos;
12. Não houve diferença entre os períodos chuvoso e seco, para distribuição e
longevidade das larvas encontradas neste estudo.

65
5. CAPÍTULO III
DISTRIBUIÇÃO E LONGEVIDADE DE LARVAS INFECTANTES DE NEMATÓIDES
GASTRINTESTINAIS DE CAPRINOS (Capra hircus) EM SOLO E PASTAGENS IRRIGADAS E
NÃO-IRRIGADAS.

66
RESUMO
CASTRO, Abisair Andrade de. Distribuição e Longevidade de larvas infectantes de
nematóides gastrintestinais de caprinos (Capra hircus) em solo e pastagem irrigada
e não-irrigada. Seropédica: UFRRJ, 2004. 97p. (Tese – Doutorado em Ciências
Veterinárias, Parasitologia Veterinária)
A distribuição e longevidade de larvas infectantes de nematóides gastrintestinais de
caprinos foi estudada em solo e em vegetação irrigada e não-irrigada com o objetivo de
contribuir para aquisição de conhecimentos e fornecer dados para o estabelecimento de
programas de controle integrado. Assim, amostras de solo e vegetação não-irrigada e
irrigada foram examinadas a cada 14 dias durante o período de setembro/02 a
setembro/03. Os resultados mostram que as larvas de helmintos foram capazes de
migrar para o solo, considerando este um refúgio às condições adversas. No entanto, a
distribuição e longevidade indicaram a irrelevante importância epidemiológica deste
habitat. A vegetação foi o local eleito pelas larvas. O sistema de irrigação não favoreceu
a migração de larvas para o solo.
Palavras-Chave: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp., solo,
vegetação, irrigação, epidemiologia
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Orientador), Laerte Grisi – UFRRJ (Co-orientador) e
Fábio B. Scott – UFRRJ.

67
ABSTRACT
CASTRO, Abisair Andrade de. Distribution and longevity of infective larvae of goats
(Capra hircus) gastrointetinal nematodes in soil and irrigated and non-irrigated
pastures. Seropédica: UFRRJ, 2004. 97p. (Tesis – Doctor Science at Ciências
Veterinárias, Parasitologia Veterinária)
The distribution and longevity of infective larvae of goats gastrointestinal nematodes
was studied in the soil and herbage irrigated and non-irrigated with aim to contribute the
acquisition of knowledge and gives dates to establishment to program of integrated
control. Thus, samples soil and pastures at two plots non-irrigated and irrigated were
examined every 14 days during september/02 at september/03. The samples soil and
herbage were processed for Baermann and Donald techniques, respectively. The results
showed that helminthes larvae were able to migrate for soil, considering this place a
refuge because the adverse condition environment. However, the distribution and
longevity indicate the small epidemiological importance this niche. Pastures was the
elect place for the larvae. The irrigation system was not favorable to migration of
infective larvae for the soil.
Keywords: Trichostrongylus sp., Haemonchus sp., Oesophagostomum sp., soil,
pastures, irrigation, epidemiology.
Comitê: Adivaldo Henrique da Fonseca – UFRRJ (Advisor), Laerte Grisi – UFRRJ (Co-advisor) e Fábio
B. Scott – UFRRJ.

68
5. 1. INTRODUÇÃO
O conhecimento da dinâmica das larvas infectantes de nematóides parasitos de
ruminantes nas pastagens e no solo de determinada região, levando-se em consideração
as condições bióticas e abióticas, é epidemiologicamente importante, uma vez que com
estes dados, pode-se determinar o risco de infecção dos animais. Desta forma,
estabelecendo medidas preventivas de controle como: alternância de pastagens,
dosificações estratégicas.
Existem poucas informações disponíveis sobre a influência de solos tropicais
albergando larvas infectantes de tricostrongilídeos (LYAKU et al. 1988). Na realidade
existe uma grande controvérsia a respeito deste fato. Alguns autores afirmam que o solo
pode atuar como reservatório de larvas infectantes, onde num ambiente desfavorável, as
L3 migrariam a determinadas profundidades para se protegerem. E num momento
seguinte, quando as condições ambientais estivessem favoráveis migrariam de volta a
superfície, recontaminando a pastagem (KAUZAL, 1941; REES, 1950). Por outro lado,
existem outros autores que discordam deste fato, pois em seus trabalhos, recuperaram
poucas larvas do solo (ANDERSON et al. 1970; ROSE & SMALL, 1985). Há, porém
que se levar em consideração os tipos de solos estudados, as condições de temperatura e
umidade, bem como a época do ano, e outros fatores. Dessa forma, com o objetivo de
estudar a distribuição e longevidade de larvas infectantes de nematóides gastrintestinais
de caprinos no solo e pasto, este estudo foi elaborado, visando no futuro propiciar uma
base sólida para programas estratégicos de controle.

69
5.2. MATERIAL E MÉTODOS
5.2.1. Local:
O presente estudo foi conduzido em pastagens próximas ao prédio de Sanidade
Animal e no laboratório de Doenças Parasitárias do Instituto de Veterinária, Convênio
Embrapa - Universidade Federal Rural do Rio de Janeiro; no município de Seropédica,
localizado a 22o 54’ de longitude oeste, à altitude de 33 metros.
5.2.2. Animais:
Nove caprinos mestiços, de ambos os sexos, procedentes do Colégio Técnico da
Universidade Federal Rural do Rio de Janeiro - CTUR, portadores de infecção natural
foram utilizados como doadores de fezes. Estes animais permaneceram, num piquete de
aproximadamente 3500m2. As amostras de fezes foram coletadas diretamente do reto
com auxílio de coletores.
5.2.3. Clima e solo:
O clima da região, segundo a classificação de KOPPEN (1958), é do tipo AW,
com inverno seco, verão chuvoso e quente. Os dados de temperatura, umidade relativa e
precipitação pluviométrica foram fornecidos pelo posto de meteorologia Ecologia
Agrícola do Km 47, distante cerca de um km do local do experimento. O solo é do tipo
franco-argiloso.
5.2.4. Piquetes:
Formaram-se dois piquetes, “A” e “B” de aproximadamente 81m2 cada,
constituídos por Paspalum notatum, grama batatais. O piquete “B” foi irrigado a taxa de
50mm de água por semana, durante todo o período do experimento, não obedecendo esta
regra apenas quando a precipitação pluviométrica excedia esta taxa. Quando a mesma
ocorria, sem, contudo exceder a taxa calculava-se a diferença.
5.2.5. Coleta de Dados:
Depositaram-se a campo, de 14 em 14 dias, quatro amostras de fezes, onde os
pares foram identificados como “massa fecal teste” e “massa fecal controle”, sendo um

70
par para análise de solo e outro para pastagem no período de setembro de 2002 a
setembro de 2003. As amostras distavam cerca de 80cm umas das outras. Previamente
ao depósito, retiravam-se alíquotas do pool de fezes coletadas, para determinação do
número de ovos por grama de fezes, o.p.g., segundo a modificada técnica de GORDON
& WHITLOCK (1939); e realização de coprocultura (ROBERTS & O’SULLIVAN,
1950) para obtenção de larvas infectantes e posterior identificação de acordo com a
chave de KEITH (1953). Alíquotas de solo e pastagem foram coletadas após o sétimo
dia de depósito e a partir daí, de 14 em 14 dias. Para a coleta do solo utilizou-se um
trado (Figura 12) com as seguintes dimensões: 3,5cm de diâmetro por 10cm de
comprimento (ROSE & SMALL, 1985). A vegetação foi coletada numa área de 5cm de
largura por 30cm de comprimento (ROSE, 1970). Primeiramente trabalhava-se a MFT,
quando esta apresentava-se negativa por duas vezes consecutivas, passava-se para a
MFC seguindo o mesmo procedimento. As respectivas amostras foram processadas
pelas técnicas de Baermann (CORT et al., 1922) (Figura 13) e Donald (DONALD,
1967). As L3 recuperadas foram quantificadas e identificadas segundo KEITH (1953).
5.2.6. Estimativa dos dados em matéria seca:
Ao longo do experimento os dados foram coletados em peso de matéria verde.
Para calcular o correspondente em matéria seca, coletou-se durante um ano, outubro de
2002 a setembro de 2003, três amostras de vegetação de acordo com ROSE (1970).
Essas amostras foram individualmente pesadas, em seguida mantidas em estufa a 80oC
por 48 horas, sendo novamente pesadas. Com os dados médios obtidos empregou-se a
análise de regressão, obtendo-se a equação da reta, y= a + b x .
Donde: y= matéria verde;
a=0.94;
b= 0.202 e
x= matéria seca.
Através da qual estimou-se a matéria seca para todas as amostras coletadas. Para
comprovação da curva dos resultados, uma nova análise de regressão foi realizada,
confrontando-se os dados de matéria verde com os de matéria seca.
5.2.7. Análise estatística:
Para melhor aproveitamento dos resultados, os mesmos foram apresentados sob
análise de três pontos de vista: dados mês a mês; dados em quatro estações e dados em

71
duas estações. Para os três casos, o número médio de larvas para distribuição e o
número médio de semanas para longevidade foram transformados em Log10 (x + 2).
Com esses dados empregou-se análise de variância (ANOVA) considerando-se um nível
de 5% de probabilidade. Em seguida, para as análises que diferiram significativamente,
foi aplicado o teste de Duncan a 5%, para comparação entre as médias (VIEIRA, 1999)

72
Figura 12. Trado, equipamento utizado para coleta de solo.
() Seta indica o comprimento de 10cm; limite de
profundidade utilizada no experimento.
A
B
Figura 13. Etapas da técnica de Baermann, utilizada para
processamento de amostras do solo. A – funis com água destilada
aquecida; B – amostras de solo sendo processadas

73
5. 3. RESULTADOS E DISCUSSÃO
As larvas infectantes recuperadas neste capítulo também foram Trichostrongylus
sp, Haemonchus sp, e Oesophagostomum. Os números de larvas recuperadas para os
gêneros acima citados no pasto (kg/ms) e solo (kg) foram respectivamente: 99.243 e
461; 289.594 e 876, e 14.563 e 109 larvas.
A recuperação de larvas do gênero Oesophagostomum foi considerada baixa.
Dessa forma não se aplicou teste estatístico a esse gênero. Como no capítulo anterior,
larvas do gênero Cooperia não foram recuperadas neste estudo.
Não houve diferença significativa (p 0.05) entre os piquetes, irrigado e não-
irrigado com relação à distribuição e longevidade de larvas dos dois gêneros analisados.
Ou seja, a irrigação neste estudo não favoreceu o desenvolvimento e longevidade de
larvas infectantes de nematóides gastrintestinais de caprinos. Contrariamente, autores
como HORAK & LOUW (1977), GRUNER et al. (1989), HOLOSOVÁ et al. (1989)
afirmaram que sistemas irrigados beneficiavam a ecologia das larvas na vegetação. Esta
aparente contradição pode estar relacionada ao tipo de vegetação em questão. A
vegetação utilizada neste estudo, Paspalum notatum, grama batatais é um tipo de
gramínea rasteira e de pequeno porte, aproximadamente 20cm de altura. Talvez por
mais desenvolvida que esta gramínea pudesse estar, não proporcionaria condições
adequadas para o desenvolvimento e sobrevivência das larvas. As vegetações utilizadas
nos estudos dos autores acima citados, Dicanthuim caricosum e Lolium multiflorum
eram de porte maior, aproximadamente 120 e 127cm de altura, respectivamente, que em
sistema de irrigação provavelmente favoreceram a expansão da cobertura vegetal,
criando um microclima favorável ao desenvolvimento das larvas.
ROSE (1960), REES (1950), ANDERSEN et al. (1970), CALLINAN (1978);
ROSE & SMALL (1985), KRECEK et al. (1991) observaram que a maioria das larvas
migraram para a vegetação e uma pequena parte para o solo. Estes resultados são
equivalentes aos achados neste estudo, pois o número de larvas recuperadas na
vegetação foi significativamente maior (p 0.05) que o número de larvas recuperadas
do solo (Figura 20A). Também houve diferença significativa entre pasto e solo quanto
ao período de sobrevivência (Figura 20B). Constatou-se que as larvas sobreviveram por
mais tempo na vegetação que no solo. Estes resultados podem estar intimamente

74
Figura 20.A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto e solo, expressos em
número de L3/Kg de matéria seca e em Kg de solo. B. Longevidade de larvas infectantes de nematóides gastrintestinais
de caprinos, no pasto e solo, expressos em semanas. (Letras, minúsculas, distintas diferem ao nível de 5% de
probabilidade.).
T r i c h o s tr o n g yl u s s p .
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
J F M A M J J A S O N D
L3
/ K
g d
e m
s
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
L3
/ kg
de
so
lo
Pa s to a
S o lo b
H a e m o n c h u s s p .
0
6 0 0 0 0
1 2 0 0 0 0
1 8 0 0 0 0
2 4 0 0 0 0
J F M A M J J A S O N D
L3
/ K
g d
e m
s
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
6 0 0
L3
/ kg
de
so
lo
Pa s to a
S o lo b
O e s o p h a g o s to m m s p .
0
2 0 0 0
4 0 0 0
6 0 0 0
8 0 0 0
J F M A M J J A S O N D
m e s e s
L3
/ K
g d
e m
s
0
2 0
4 0
6 0
8 0
L3
/ kg
de
so
lo
Pa s to
S o lo
O e s o p h a g o s to m m s p .
0
2
4
6
8
J F M A M J J A S O N Dm e s e s
se
ma
na
s -
pa
sto
0
0 ,5
1
1 ,5
2
2 ,5
se
ma
na
s -
so
lo
Pa s to
S o lo
T ric h o s tro n g y lu s s p .
0
2
4
6
8
1 0
1 2
J F M A M J J A S O N D
se
ma
na
s -
pa
sto
0
1
2
3
4
5
6
se
ma
na
s -
so
lo
Pa s to a
S o lo b
H a e m o n c h u s s p .
0
2
4
6
8
1 0
1 2
1 4
J F M A M J J A S O N D
se
ma
na
s -
pa
sto
0
1
2
3
4
5
se
ma
na
s -
so
lo
Pa s to a
S o lo b
A B

75
relacionados ao tipo de solo da região, que é do tipo franco argiloso, o qual possui em
sua composição cerca de 70% de argila e 30% de areia (RAMOS et al. 1973). Isto
provavelmente dificultou a migração e sobrevivência das L3 no solo, uma vez que este
pode produzir uma liga bastante forte com baixa aeração. Autores como KAUZAL
(1941), CALLINAN & WESTCOTT (1986), LYAKU et al. (1988) consideraram que o
solo poderia atuar como um reservatório de larvas infectantes, buscando neste, um
refúgio as condições adversas, pois seus resultados apontaram para essa hipótese, no
entanto, os tipos de solos trabalhados por esses autores eram do tipo arenoso e humífero.
5.3.1. Dados apresentados mês a mês:
Analisando os resultados deste estudo mês a mês, observou-se que não houve
diferença significativa entre os meses tanto para distribuição quanto para longevidade
das larvas infectantes de Trichostrongylus e Haemonchus, na vegetação e solo
respectivamente (Figuras 21 A, B e 22 A, B).
Foi registrado, porém, que a maior quantidade de larvas recuperadas para
Trichostrongylus no pasto foi no mês de julho e no solo no mês de outubro. Meses cujos
valores médios de temperatura e precipitação foram respectivamente 22 e 260C e 10 e
52mm (Figura 23). Na vegetação as L3 foram capazes de sobreviver por um período de
aproximadamente dez semanas e no solo seis, ambos no mês de abril.
No pasto, o gênero Haemonchus teve seu pico de recuperação no mês de março,
onde foi registrada a temperatura média de 260C e 246mm de precipitação; e no solo o
mês de outubro. A longevidade dessas larvas no pasto e solo foi de 13 semanas em
março e cinco semanas em abril, respectivamente. LYAKU et al. (1988) observaram
que larvas de Haemonchus sobreviveram até três semanas em solos, concordando com o
registrado no presente estudo.
Larvas de Oesophagostomum na vegetação foram mais recuperadas em maio,
cujos valores de temperatura e precipitação foram respectivamente 220C e 70.6mm e no
solo, em outubro. O período de sobrevivência dessas larvas foi de oito semanas em
março, na vegetação e de duas semanas no solo, em abril. LYAKU et al. (1988)
verificaram que essas larvas sobreviveram até nove semanas, em seus estudos.
A exemplo do capítulo anterior, a temperatura pareceu ser um fator importante
na distribuição de Trichostrongylus e Oesophagostomum ao longo do ano, pois os dois
gêneros, foram recuperados em meses onde a temperatura apresentou-se mais amena.
Concordando com LEVINE (1963), que afirmou que a temperatura poderia ser um fator

76
Figura 21.A. Distribuição de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto, expressos em
número de L3/Kg de matéria seca e em logaritmo do número de L3. B. Distribuição de larvas infectantes de nematóides
gastrintestinais de caprinos, no solo, expressos em número de L3/Kg e em logaritmo do número de L3 (Letras,
minúsculas, idênticas não diferem ao nível de 5% de probabilidade.).
T ric h o s tro n g y lu s s p . - p a s to
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
L3
/ k
g m
s
0
0 ,5
1
1 ,5
2
2 ,5
3
3 ,5
4
4 ,5
Lo
g1
0-
L3
H a e m o n c h u s s p . - p a s to
0
6 0 0 0 0
1 2 0 0 0 0
1 8 0 0 0 0
2 4 0 0 0 0
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
L3
/ k
g m
s
0
1
2
3
4
5
6
Lo
g1
0-
L3
O e s o p h a g o s to m u m s p . - p a s to
0
2 0 0 0
4 0 0 0
6 0 0 0
8 0 0 0
J F M A M J J A S O N Dm e s e s
L3
/ k
g m
s
T ric h o s tro n g y lu s s p . - s o lo
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
3 5 0
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
L3
/ k
g s
olo
0
0 ,5
1
1 ,5
2
2 ,5
3
Lo
g1
0-
L3
H a e m o n c h u s s p . - s o lo
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
6 0 0
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
L3
/ k
g s
olo
0
0 ,5
1
1 ,5
2
2 ,5
3
Lo
g1
0-
L3
vv
O e s o p h a g o s to m u m s p . - s o lo
0
2 0
4 0
6 0
8 0
J F M A M J J A S O N Dm e s e s
L3
/ k
g s
olo
A B

77
Figura 22.A. Longevidade de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto, expressos em
semanas e em logaritmo do número de semanas. B. Longevidade de larvas infectantes de nematóides gastrintestinais
de caprinos, no solo, expressos em semanas e em logaritmo do número de semanas (Letras, minúsculas, idênticas não
diferem ao nível de 5% de probabilidade.).
T ric hos t rongylus s p . - pas to
0
2
4
6
8
1 0
1 2
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
se
ma
na
s -
Lo
n L
3
0
0 ,2
0 ,4
0 ,6
0 ,8
1
1 ,2
Lo
g1
0 -
lo
ng
L3
H a e m o n c h u s s p . - p a s to
0
2
4
6
8
1 0
1 2
1 4
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
se
ma
na
s -
Lo
n L
3
0
0 ,2
0 ,4
0 ,6
0 ,8
1
1 ,2
1 ,4
Lo
g1
0 -
lo
ng
L3
O e s o p h a g o s to m u m s p . - p a s to
0
2
4
6
8
J F M A M J J A S O N Dm e s e s
se
ma
na
s -
Lo
n L
3
T ric hos t rongylus s p . - s o lo
0
1
2
3
4
5
6
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
se
ma
na
s -
Lo
n L
3
0
0 ,2
0 ,4
0 ,6
0 ,8
1
Lo
g1
0 -
lo
ng
L3
H aem onc hus s p . - s o lo
0
1
2
3
4
5
Jan
a
F ev
a
M ar
a
A br
a
M ai
a
Jun
a
Ju l
a
A go
a
S et
a
O ut
a
N ov
a
D ez
a
se
ma
na
s -
Lo
n L
3
0
0 ,1
0 ,2
0 ,3
0 ,4
0 ,5
0 ,6
0 ,7
0 ,8
0 ,9
Lo
g1
0 -
lo
ng
L3
O e s o p h a g o s to m u m s p . - s o lo
0
0 ,5
1
1 ,5
2
2 ,5
J F M A M J J A S O N Dm e s e s
se
ma
na
s -
Lo
n L
3
A B
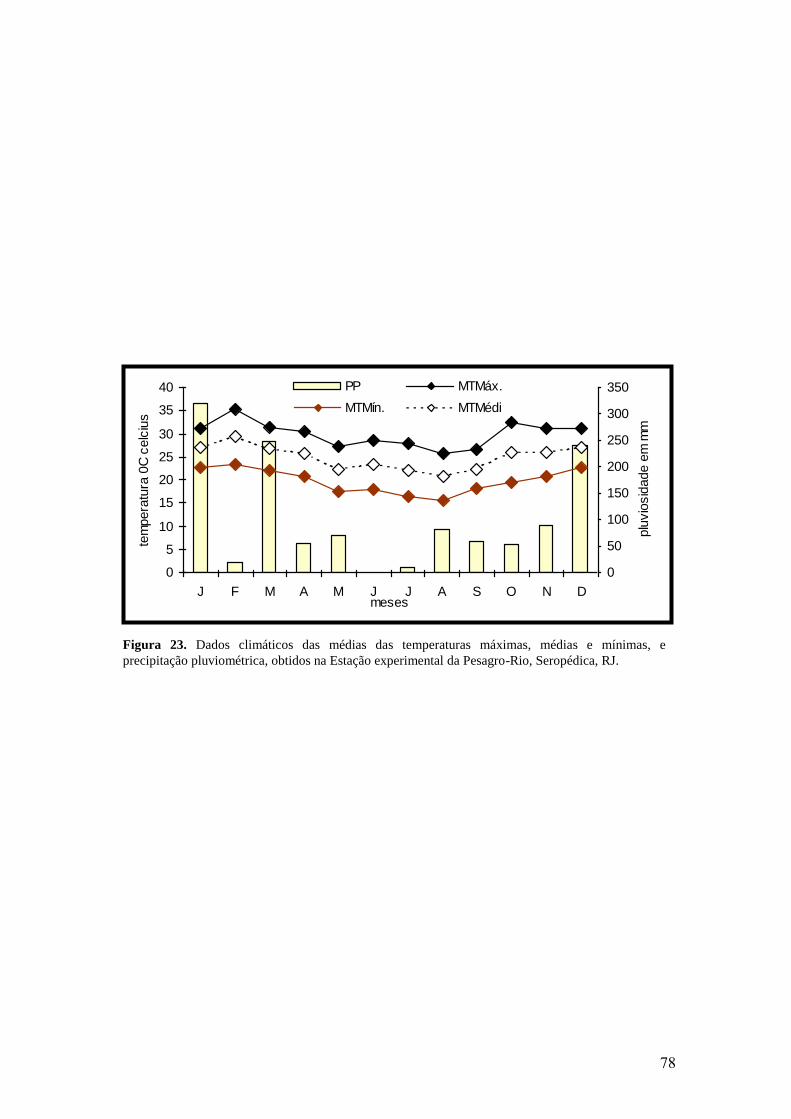
78
0
5
10
15
20
25
30
35
40
J F M A M J J A S O N Dmeses
tem
pera
tura
0C
celc
ius
0
50
100
150
200
250
300
350
plu
vio
sid
ade e
m m
m
PP MTMáx.
MTMín. MTMédi
Figura 23. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, obtidos na Estação experimental da Pesagro-Rio, Seropédica, RJ.

79
limitante. A precipitação não pareceu exercer importância, pois os meses de maior
precipitação não coincidiram com os meses de maior recuperação de larvas. E embora
os maiores números de L3 de Haemonchus tenham sido recuperados num mês chuvoso,
esses eventos pareceram não ter relação direta com a distribuição de larvas; podendo
outros fatores, estarem influenciando de forma mais clara, como sugeriu AUMONT et
al. (1989).
5.3.2. Dados apresentados em quatro estações:
Assim como no tópico anterior, o número de larvas recuperadas e o período
sobrevivência destas foi significativamente maior no pasto (p 0.05) do que solo. A
distribuição de L3 assim como a longevidade na vegetação e solo, para os três gêneros,
estão sucintamente apresentados nas Figura 24 A, B e Figura 25 A, B, respectivamente.
Os gêneros Trichostrongylus e Haemonchus ocorreram na vegetação e no solo,
nas quatro estações. Esses dados concordam com os achados de PIMENTEL NETO &
FONSECA (no prelo), na vegetação. O gênero Oesophagostomum ocorreu nas estações
do outono e inverno, na vegetação e nas quatro estações, no solo. Os dados climáticos
divididos em quatro estações estão apresentados na figura 26.
A distribuição de larvas de Trichostrongylus foi significativamente diferente
entre as estações do verão e as demais, sendo esta a época em que menos se recuperou
larva da vegetação. TRIPATHI (1970b) verificou que o período quente foi desfavorável
à sobrevivência das larvas. Foi no inverno quando registrou-se o maior número de
larvas, 15.498. No solo não foi registrada diferença entre as estações, embora o maior
número de larvas tenha sido recuperado na primavera, duas L3. O outono foi a estação
onde registrou-se significativamente o maior período de sobrevivência para larvas de
Trichostrongylus na vegetação, em relação as demais estações. No solo, não houve
diferença significativa entre as estações, embora também tenha sido no outono,
registrado a maior longevidade.
O gênero Haemonchus foi recuperado na vegetação, em maior número na
estação do outono, diferindo significativamente das demais. DINABURG (1944)
observou que os meses do verão e inverno foram desfavoráveis ao desenvolvimento e
sobrevivência das larvas de Haemonchus. Estes dados estão de acordo com o presente
estudo. No solo, embora não tenha ocorrido diferença estatística entre as estações, a
primavera foi a que se destacou. O período de sobrevivência de Haemonchus na

80
Figura 24.A. Distribuição, nas quatro estações, de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto,
expressos número de L3/Kg matéria seca e em logaritmo do número de L3. B. Distribuição, nas quatro estações, de larvas
infectantes de nematóides gastrintestinais de caprinos, no solo, expressos em número de L3/Kg matéria seca e em
logaritmo do número de L3 (Letras, minúsculas, idênticas não diferem ao nível de 5% de probabilidade.).
H a e m n c h u s s p . - p a s to
0
2 0 0 0 0
4 0 0 0 0
6 0 0 0 0
8 0 0 0 0
1 0 0 0 0 0
v e r o u t in v p r i
L3
/ K
g m
s
T r i c h o s tr o n g yl u s s p . - p a s to
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
v e r o u t in v p r i
L3
/ K
g m
s
O e s o p h a g o s to m u m s p . - p a s to
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
v e r o u t in v p r ie s ta ç õ e s
L3
/ K
g m
s
b
a
a
a
b
a
b b
T r i c h o s tr o n g yl u s s p . - s o lo
0
0 ,5
1
1 ,5
2
v e r o u t in v p r i
L3
/ kg
0
0 ,1
0 ,2
0 ,3
0 ,4
0 ,5
0 ,6
Lo
g -
L3
/ K
g
H a e m n c h u s s p . - s o lo
0
5 0
1 0 0
1 5 0
2 0 0
v e r o u t in v p r i
L3
/kg
0
0 ,5
1
1 ,5
2
2 ,5
Lo
g -
L3
/kg
O e s o p h a g o s to m u m s p . - s o lo
0
5
1 0
1 5
2 0
2 5
3 0
v e r o u t in v p r ie s ta ç õ e s
L3
/ K
g
a
aa
a
aa
a
a
A B

81
Figura 25.A. Longevidade, nas quatro estações, de larvas infectantes de nematóides gastrintestinais de caprinos, no pasto,
expressos em semanas e em logaritmo do número de semanas. B. Longevidade, nas quatro estações, de larvas infectantes de
nematóides gastrintestinais de caprinos, no solo, expressos em semanas e em logaritmo do número de semanas (Letras,
minúsculas, idênticas não diferem ao nível de 5% de probabilidade.).
T r i c h o s tr o n g yl u s s p . - p a s to
0
2
4
6
8
1 0
v e r o u t in v p r is
em
an
as
H a e m n c h u s s p . - p a s to
0
2
4
6
8
1 0
v e r o u t in v p r i
se
ma
na
s
O e s o p h a g o s to m u m s p . - p a s to
0
2
4
6
8
v e r o u t in v p r ie s ta ç õ e s
se
ma
na
s
c
a
b
b , c
b
a
b b
T r i c h o s tr o n g yl u s s p . - s o lo
0
0 ,5
1
1 ,5
2
2 ,5
3
3 ,5
v e r o u t in v p r i
se
ma
na
s
0
0 ,2
0 ,4
0 ,6
0 ,8
Lo
g 1
0 -
se
ma
na
s
H a e m n c h u s s p . - s o lo
0
0 ,5
1
1 ,5
2
2 ,5
3
3 ,5
ve r o u t in v p r i
se
ma
na
s
0
0 ,2
0 ,4
0 ,6
0 ,8
Lo
g1
0 -
se
ma
na
s
O e s o p h a g o s to m u m s p . - s o lo
0
1
2
3
4
v e r o u t in v p r ie s ta ç õ e s
se
ma
na
s
a
a
a a
a
a
a
a
A B
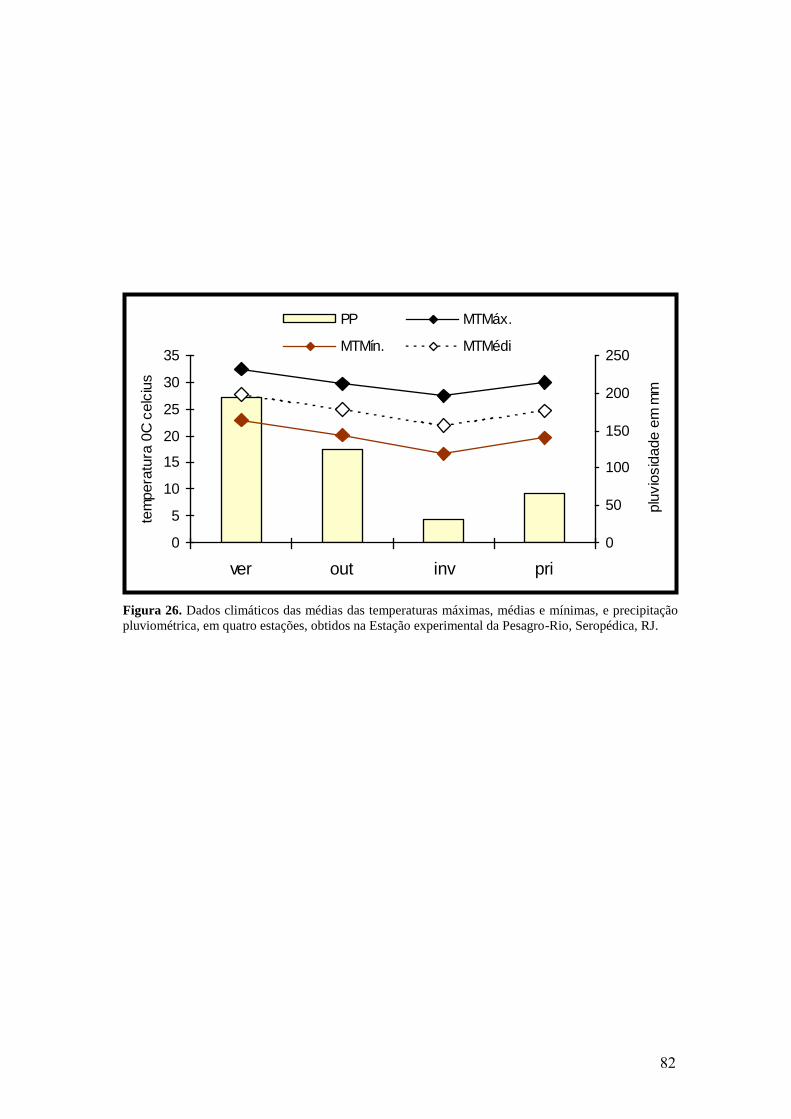
82
0
5
10
15
20
25
30
35
ver out inv pri
tem
pera
tura
0C
celc
ius
0
50
100
150
200
250
plu
vio
sid
ade e
m m
m
PP MTMáx.
MTMín. MTMédi
Figura 26. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e precipitação
pluviométrica, em quatro estações, obtidos na Estação experimental da Pesagro-Rio, Seropédica, RJ.

83
vegetação foi significativamente maior no outono. No solo também não houve diferença
entre as estações.
LEVINE & TODD (1975) observaram que poucas larvas de Haemonchus
sobreviveram no inverno, e ao contrário larvas de Trichostrongylus sobreviveram bem
nesta estação. O mesmo foi verificado neste estudo.
Embora não se tenha empregado teste estatístico para Oesophagostomum,
verificou-se que o outono e a primavera foram estações eleitas para o desenvolvimento
de L3 na vegetação e solo, respectivamente. O melhor período de sobrevivência foi
identificado como a estação do outono na vegetação e no solo, não houve destaque.
5.3.3. Dados apresentados nos períodos chuvoso e seco:
Os dados de distribuição e longevidade dos três gêneros de larvas recuperadas
neste estudo ao nível de vegetação e solo, estão respectivamente apresentados nas
figuras 27A, B e 28A, B. Os dados climáticos estão apresentados na figura 29.
O três gêneros ocorreram na vegetação e no solo, nos dois períodos estudados. A
recuperação e sobrevivência de larvas de Trichostrongylus na vegetação foi
significativamente superior no período seco. Para o solo, o período chuvoso e seco,
foram, respectivamente, as épocas em que se recuperou maior número de larvas e que as
mesmas sobreviveram, embora não tenha sido verificada diferença significativa entre os
períodos. A distribuição e longevidade de larvas de Oesophagostomum foram
semelhantes à de Trichostrongylus.
Não houve diferença estatística entre os dois períodos para a distribuição e
longevidade de larvas de Haemonchus tanto na vegetação quanto no solo. TRIPATHI
(1970a), FABIYI (1973), CAVALCANTI (1974), GRISE (1975), CONNOR et al.
(1990), McCULLOCK & KASIMBALA (1968) registraram que H. contortus
apresentou seu pico no período chuvoso. Embora os resultados na vegetação mostrem
uma tendência a este fato, os mesmos discordam, pois estatisticamente não houve
diferença entre os dois períodos.
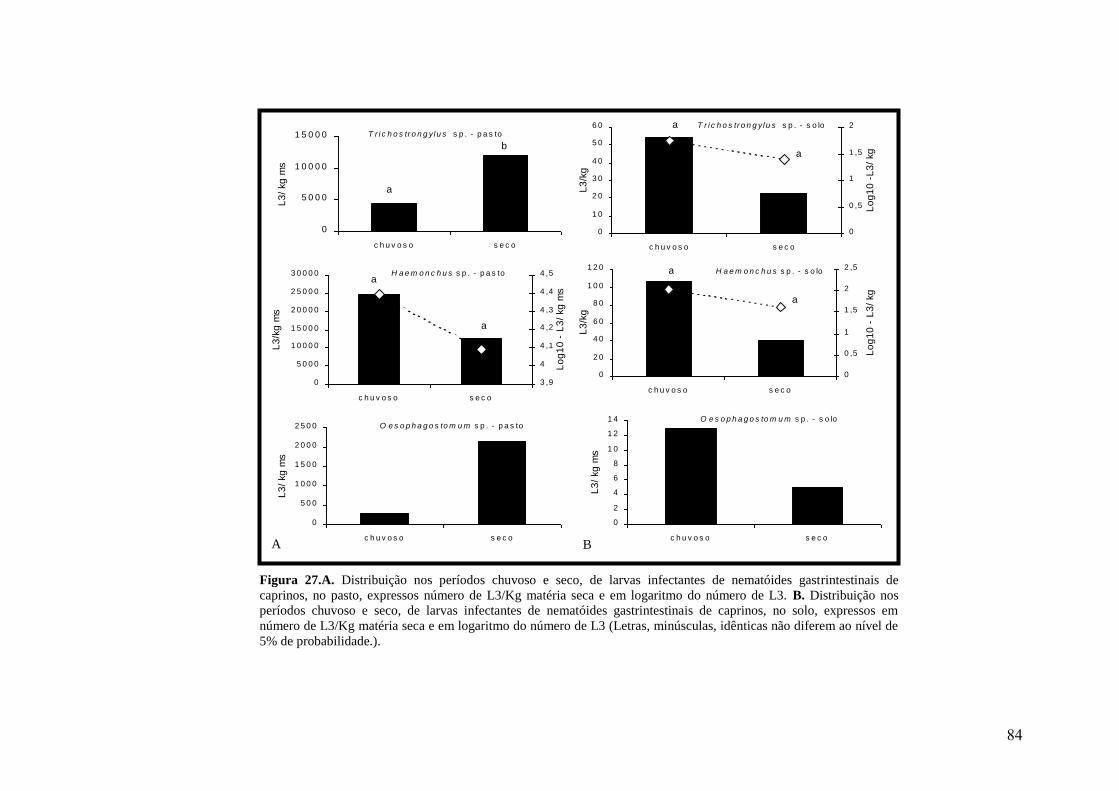
84
Figura 27.A. Distribuição nos períodos chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de
caprinos, no pasto, expressos número de L3/Kg matéria seca e em logaritmo do número de L3. B. Distribuição nos
períodos chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de caprinos, no solo, expressos em
número de L3/Kg matéria seca e em logaritmo do número de L3 (Letras, minúsculas, idênticas não diferem ao nível de
5% de probabilidade.).
H a e m o n c h u s s p . - p a s to
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
3 0 0 0 0
c h u v o s o s e c o
L3
/kg
ms
3 ,9
4
4 ,1
4 ,2
4 ,3
4 ,4
4 ,5
Lo
g1
0 -
L3
/ kg
ms
T r i c h o s tr o n g yl u s s p . - p a s to
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
c h u v o s o s e c o
L3
/ kg
ms
O e s o p h a g o s to m u m s p . - p a s to
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
2 5 0 0
c h u v o s o s e c o
L3
/ kg
ms
a
b
a
a
T r i c h o s tr o n g yl u s s p . - s o lo
0
1 0
2 0
3 0
4 0
5 0
6 0
c h u v o s o s e c o
L3
/kg
0
0 ,5
1
1 ,5
2
Lo
g1
0 -
L3
/ kg
H a e m o n c h u s s p . - s o lo
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
c h u v o s o s e c o
L3
/kg
0
0 ,5
1
1 ,5
2
2 ,5
Lo
g1
0 -
L3
/ kg
O e s o p h a g o s to m u m s p . - s o lo
0
2
4
6
8
1 0
1 2
1 4
c h u v o s o s e c o
L3
/ kg
ms
a
a
a
a
A B
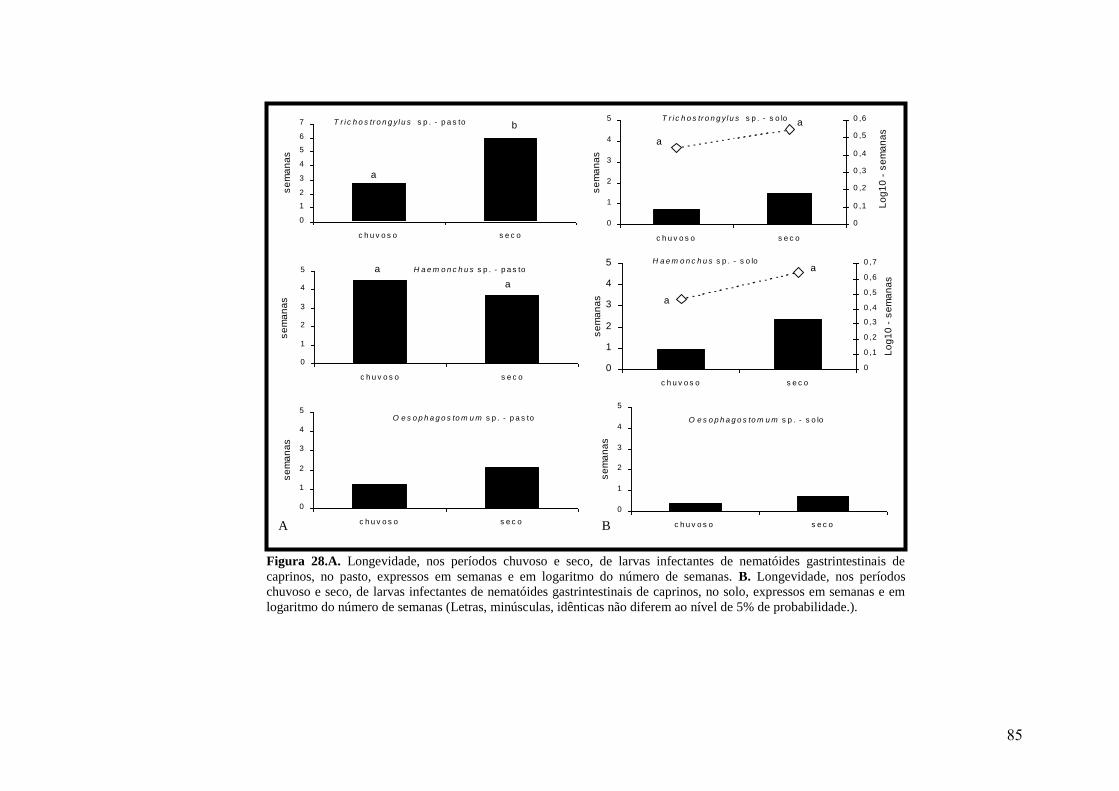
85
Figura 28.A. Longevidade, nos períodos chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de
caprinos, no pasto, expressos em semanas e em logaritmo do número de semanas. B. Longevidade, nos períodos
chuvoso e seco, de larvas infectantes de nematóides gastrintestinais de caprinos, no solo, expressos em semanas e em
logaritmo do número de semanas (Letras, minúsculas, idênticas não diferem ao nível de 5% de probabilidade.).
T r i c h o s tr o n g yl u s s p . - p a s to
0
1
2
3
4
5
6
7
c h u v o s o s e c o
se
ma
na
s
H a e m o n c h u s s p . - p a s to
0
1
2
3
4
5
c h u v o s o s e c o
se
ma
na
s
O e s o p h a g o s to m u m s p . - p a s to
0
1
2
3
4
5
c h u v o s o s e c o
se
ma
na
s
a
b
a
a
T r i c h o s tr o n g yl u s s p . - s o lo
0
1
2
3
4
5
c h u v o s o s e c o
se
ma
na
s
0
0 ,1
0 ,2
0 ,3
0 ,4
0 ,5
0 ,6
Lo
g1
0 -
se
ma
na
s
H a e m o n c h u s s p . - s o lo
0
1
2
3
4
5
c h u v o s o s e c o
se
ma
na
s
0
0 ,1
0 ,2
0 ,3
0 ,4
0 ,5
0 ,6
0 ,7
Lo
g1
0 -
se
ma
na
s
O e s o p h a g o s to m u m s p . - s o lo
0
1
2
3
4
5
c h u v o s o s e c o
se
ma
na
s
a
a
a
a
A B
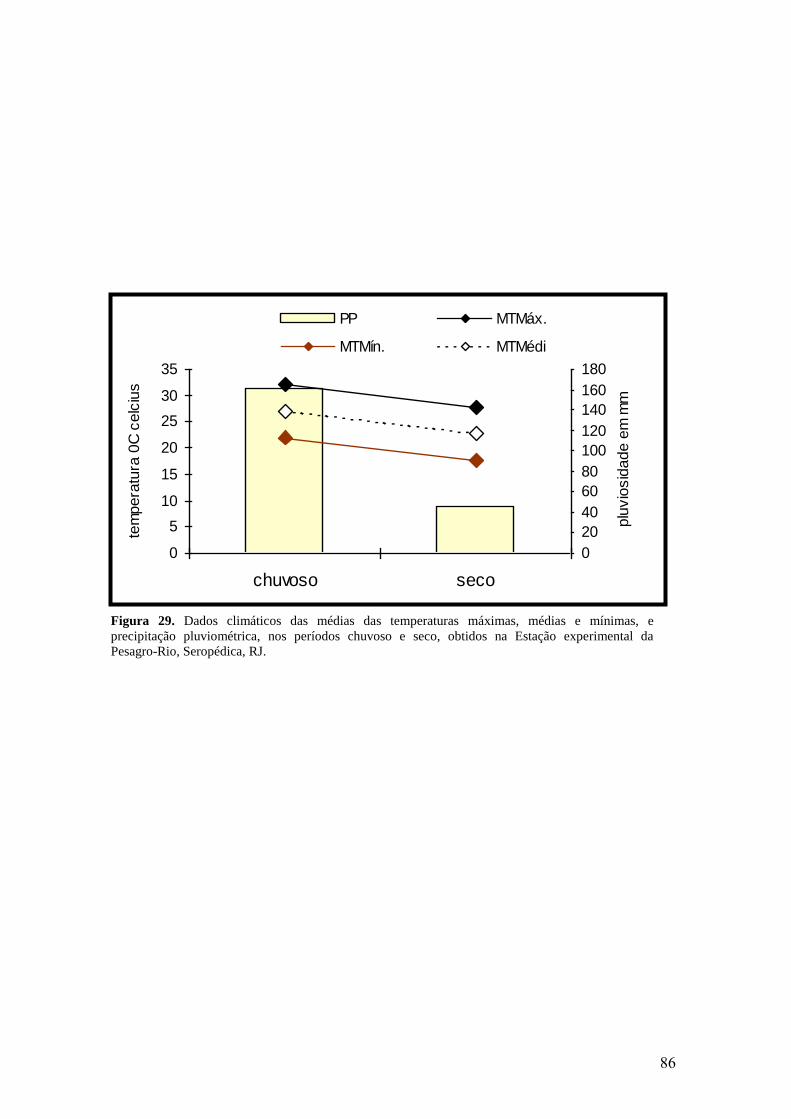
86
0
5
10
15
20
25
30
35
chuvoso seco
tem
pera
tura
0C
celc
ius
0
20
40
60
80
100
120
140
160
180
plu
vio
sid
ade e
m m
m
PP MTMáx.
MTMín. MTMédi
Figura 29. Dados climáticos das médias das temperaturas máximas, médias e mínimas, e
precipitação pluviométrica, nos períodos chuvoso e seco, obtidos na Estação experimental da
Pesagro-Rio, Seropédica, RJ.

87
5. 4. CONCLUSÕES
De acordo com os resultados apresentados neste capítulo, pôde-se concluir que:
1. O sistema de irrigação não favoreceu a migração de larvas infectantes para a
vegetação e solo;
2. As larvas infectantes de nematóides gastrintestinais foram capazes de migrar para o
solo, porém a distribuição e o período de sobrevivência mostraram a irrelevante
importância epidemiológica deste habitat, nesta região.
3. Ao longo de um ano, não houve diferença significativa entre os meses, para
distribuição e longevidade de larvas infectantes dos três gêneros encontrados neste
estudo;
4. Com análise nas quatro estações, o gênero Trichostrongylus obteve o maior e menor
número de larvas recuperadas no inverno e verão, respectivamente. Esses resultados
foram associados à temperatura;
5. O maior número de larvas infectantes de Haemonchus recuperadas foi no outono;
6. O outono foi a época mais favorável para a sobrevivência de larvas infectantes de
nematóides gastrintestinais de caprinos, na vegetação;
7. O período seco foi favorável à distribuição e sobrevivência de larvas de
Trichostrongylus;
8. Não houve diferença significativa entre os períodos chuvoso e seco para distribuição
e longevidade de Haemonchus.

88
6. REFERÊNCIAS BIBLIOGRÁFICAS
AL SAQUR, I.; BAIRDEN, J.; ARMOUR, J & GETTINBY, G. Population study of
bovine Ostertagia spp infective larvae on herbage and soil. Research in Veterinary
Science, v. 32, n. 3, p. 332-337. 1982.
ALMEIDA, L. R. Desenvolvimento, sobrevivência e distribuição de larvas infectantes
de nematódeos gastrintestinais de ruminantes, na estação seca da Baixada Fluminense,
Rio de Janeiro. 2003. 50 p. Dissertação (Mestrado) – Universidade Federal Rural do
Rio de Janeiro, Rio de Janeiro, 2003.
ANDERSEN, F. L.; LEVINE, N. D. & BOATMAN, P. A. Survival of third-stage
Trichostrongylus colubriformis larvae on pasture. Journal of Parasitology, n. 56, v. 2, p.
209-232, 1970.
AROSEMENA, N. A. E.; BEVILAQUA, C. M. L.; MELO, A. C. F. L. & GIRÃO, M.
D. Seasonal variations of gastrointestinal nematodes in sheep and goat from semi-arid
area. Revue de Medecine Veterinaire, v. 150, n. 11, p. 873-876, 1999.
AUMONT, G. & GRUNER, L. Population evolution of free-living stage of goat
gastrointestinal nematodes on herbage under tropical conditions in Guadeloupe (French
west indies). International Journal for Parasitology, v. 19, n. 5, p. 539-546, 1989.
AUMONT, G.; COULAUD, G.; GRUDE, A & GRUNER, L. Pasture populations of
cattle nematode larvae in Guadeloupe (French west indies). International Journal for
Parasitology, v. 19, n. 5, p. 547-554, 1989.
BAERMANN, G. Eine einfache methode zur auffindung von Ankylostomum
(Nematoden) larven in Erdproden. Geneesk Tidjschr Nederl Indie, n. 57, v. 1, p. 131-
137, 1917.
BARGER, I. A.; SIALE, K.; BANKS, D. J. D. & LE JAMBRE, L. F. Rotaional grazing
for control of gastrointestinal nematodes of goats in west tropical environment.
Veterinary Parasitology, v. 53, n. 1, p. 109-116, 1994.
BAKER, W. D. A. Control of parasitic gastroenteritis in goat. Journal of American
Veterinary Medicine of Association, v. 167, n. 12, p. 1069-1075, 1975.
BAKER, N. F.; FISK, R. A.; BUSHNELL, R. B. & OLIVER, M. N. Seasonal
occurrence of infective nematode larvae on irrigated pasture grazed by cattle in
California. American Journal of Veterinary Research, v. 42, n. 7, p. 1188-1191, 1981.
BANKS, D. J. D.; SINGH, R.; BARGER, I. A.; PRATAP, B. & LE JAMBRE, L. F.
Development and survival of infective larvae of Haemonchus contortus on pasture in a
tropical environment. International Journal for Parasitology, v. 20, n. 2, p. 155-160,
1990.

89
BAWDEN, R. J. A rapid technique for the recovery of strongyloid larvae from pasture
samples. Australian Veterinary Journal, v. 45, n. 5, p. 228-230.1969.
BHOJANE, G. R.; DAKSHINKAR, N. P. & KOTEKAR, M. D. Prevalence of
endoparasites in free ranging goats of Nagpur. Indian Journal of Small Ruminants, v. 8,
n. 1, p. 57-59, 2002.
BOUGAS, G. & THEODORIDIS, Y. Gastrointestinal nematodes of goats in Thessalia
region. Bulletin of the Hellenic Veterinary Medical Society, v. 51, n. 4, p. 281-287,
2000.
BULLISK, G. R & ANDERSEN, F. L. Effect of irrigation on survival of third-stage
Haemonchus contortus larvae (Nematoda, Trichostongylidae). Great Basin Nature, v.
38, n. 5, 369-378, 1978.
BÜRGER, H. J. Experiences with our techniques for the recovery of nematode larvae
from herbage. p. 25-30. In: NANSEN, P.; JORGENSEN, R. J. & SOULSBY, E. J. L.
Epidemiology and control of nematodiasis in cattle. Brussel, Luxembourg. 250p. 1981.
CABARET, J. RAYNAUD, J. P. & LE STANG, J. P. veterinary Parasitology, v. 10, n.
1, p. 65-71. 1982.
CALLINAN, A. P. L. The ecology of the free-living stages of Trichostrongylus axei.
International Journal for Parasitology, v. 8, n. 6, p. 453-456, 1978.
CALLINAN, A. P. L. The ecology of the free-living stages of Trichostrongylus vitrinus.
International Journal for Parasitology, v. 9, n. 2, p. 133-136, 1979.
CALLINAN, A. P. L. & WESTCOTT, J. M. Vertical distribution of trichostrongylid
larvae on herbage and soil. International Journal for Parasitology, v. 16, n. 3, p. 241-
244, 1986.
CARMICHAEL, I. H. & BRIEN, F. Influences on internal parasitism of sheep in South-
east Australian: studies of (1) internal parasite control on pívot irrigation systems and
(2) cobalt nutrition and internal parasite interactions. Inaugural Sheep and Wool
Industry Conference, Autralia, v. 50, n. 4, p. 518-533, 2002.
CAVALCANTI, A. M. L. Prevalência estacional de helmintos gastrintestinais de
caprinos nas zonas da Mata, do Agreste e do Sertão de Pernambuco. 1974. 48p.
Dissertação (Mestrado) – Universidade Federal de Minas Gerais, Belo Horizonte, 1974.
CHARLES, T. P. Seasonal prevalence of gastrointestinal nematodes of goats in
Pernambuco State, Brazil. Veterinary Parasitology, v. 30, n. 4, p. 335-343. 1989.
CHIEJINA, S. N. Evaluation of modified Baermann apparatus for the recovery of
infective trichostrongylid larvae from herbage samples. Journal of Helminthology, v.
56, n. 2, p. 105-109, 1982.

90
CONNOR, R. J.; MUNYUKU, A. P.; MACKYAO, E. & HALLIWELL, R. W.
Helminthosis in goats in southern Tanzania: Investigations on epidemiology and
control. Tropical Animal Health Production, v. 22, n. 1, p. 1-6, 1990.
CORT, W.W.; ACKERT, J.E.; AUGUSTINE, D. L. & PAYNE, F. K. Investigations on
the control of hooworm disease. II. The description of an apparatus for isolating
infective hookworm larvae from soil. The American Journal of Hygiene, v. 2, n. 1, p. 1-
16, 1922.
CORT, W. W.; STOLL, N. R & GRANT, J. B. Researches on hookworm in China. I.
Problems and methods of attack. The American Journal of Hygiene, v. 7, n. 1, p. 1-32,
1926.
COUVILLION, C. E. Estimation of the numbers of trichostrongylid larvae on pastures.
Veterinary Parasitology, v. 46, n. 2, p. 197-203, 1993.
CROFTON, H. D. The ecology of the immature phases of Trichostongyle nematodes I.
The vertical distribution of infective larvae of Trichostrongylus retortaeformis in
relation to their habitat. Parasitology, v. 39, n. 1, p. 17-25, 1948.
CROFTON, H. D. The ecology of the immature phases of Trichostongyle parasites. V.
The estimation of pasture infestation. Parasitology, v. 44, n. 1, p. 313-324, 1954.
DEBELA, E. Epidemiology of gastrointestinal helminthiasis of Rift Valley goats under
traditional husbandry system in Adami Tulu district, Ethiopia. Ethipian Journal of
Science, v. 25, n. 1, p. 35-44, 2002.
DEVENDRA, C. Potential of sheep and goats in less developed countries. Journal
Animal Science, v. 51, n. 4, p. 461-473, 1981.
DIALLO, M. B. Seasonal dynamics of gastrointestinal nematodes in small ruminants in
the Kindia prefecture (Guinea). Bulletin of Animal Health and Production in Africa, v.
46, n. 4, p. 251-253, 1998.
DINABURG, A. G. The efficiency of the Baermann apparatus in the recovery of larvae
of Haemonchus contortus. The Journal of Parasitology, v. 28, n. 6, p. 433-440. 1942.
DINABURG, A. G. The survival of the infective larvae of the ruminant stomach worm.
Haemonchus contortus, on outdoor grass plots. American Journal of Veterinary
Research, v. 5, n. 1, p. 32-37, 1944.
DINNIK, J. A. & DINNIK, N. N. Observations on the development of Haemonchus
contortus larvae under field conditions in the Kenya hinghlands. Bulletin of
Epizootiologia Disease of Africa, v. 6, n. 1, p. 11-21, 1958.
DOLL, E. R. & HULL, F. E. Survival of sheep nematode larvae on pasture during
summer. Veterinary Medical, v. 43, n. 4, p. 147-148, 1948.
DONALD, A. B. A technique for the recovery of strongyloid larvae from small sample
units of pasture. Journal of Helminthology, v. 61, n. 1, p. 1-10, 1967.

91
DURIE, P. H. A new technique for the recovery of infective strongyle larvae from soil
and pasture. Journal of Helminthology, v. 33, n. 2, p. 189-196, 1959.
EYSKER, M. & KOOYMAN, F. N. J. Notes on necropsy and herbage processing
techniques for gastrointestinal nematodes of ruminants. Veterinary Parasitology, v. 46,
n. 2, p. 206-213, 1993.
FABIYI, J. P. An investigation into the incidence of goat helminth parasites in the Zaria
of Nigeria. Bulletin Epizootiology Disease of Africa, v. 18, n. 1, p. 29-34, 1970.
FABIYI, J. P. Seasonal fluctuations of nematode infestations in goats in the savannh
belt of Nigeria. Bulletin Epizootiology Disease of Africa, v.21, n. 3, p. 277-286, 1973.
FINCHER, G. T. & STWART, T. B. Vertical migration by nematode larvae of cattle
parasites through soil. Proceedings of the Helminthological Society of Washington, v.
46, n. 1, p. 43-46, 1979.
FINE, A. E.; HARTMAN, R.; KRECEK, R. C. & GROENEVELD, H. T. Effects of
time, from collection to processing, on the recovery of Haemonchus contortus third-
stage larvae from herbage. Veterinary Parasitology, v. 51, n. 1, p. 77-83, 1993.
FITZSIMMONS, W. M. Pathogenesis of the trichostrongyles. Helminthological
Abstracts, v. 38, n. 2, p.139-190. 1970.
FURMAN, D. P. Effects of environment upon the free-living stages of Ostertagia
circumcincta (Stadelmann) Trichostrongylidae. 2. Field experiments. American Journal
of Veterinary Research, n. 5, v. 2, p. 147-153, 1944.
GETTINBY, G.; McKELLAR, Q. A.; BAIRDEN, K.; THEODORIDIS, Y. &
WHITELAW, A. Comparison of two techniques used for the recovery of nematode
infective larvae from pasture. Research in Veterinary Science, v. 39, n. 1, p. 99-102,
1985.
GIBBS, H. C. Transmission of parasites with reference to the strongyles of domestic
sheep and cattle. Canadian Journal of Zoology, v. 51, n. 3, p. 281-289. 1973.
GIBSON, T. E. & EVERETT, G. Experiments on the control of Trichostrongyloisis in
lambs. Journal of Compendium Pathology, v. 78, n. 5, p. 427-434, 1968.
GITHIGIA, S. M.; THAMSBORG, S. M.; MUNYUA, W. K. & MAINGI, N. Impact of
gastrointestinal helmints on production in goats in Kenya. Small Ruminant Research, v.
42, n. 1, p. 21-29, 2001.
GORDON, H. McL. Self-cure reaction. p. 174-190. In: SOULSBY, E. J. L. The
reaction of the host to parasitism. Lyon, France, 490p. 1967.
GORDON, H. McL. & WHITLOCK, H.V. A new technique for counting nematode
eggs in sheep faeces. Journal Council Sience of Induntry and Research, v 12, n.1, p. 50-
52, 1939.

92
GRISI, L. Incidência de helmintos em Capra hircus L. procedentes do Estado da Bahia.
Revista Brasileira de Biologia, v. 35, n.1,p. 101-108, 1975.
GRUNER, L.; BERBIGIER, P.; CORTET, J. & SAUVE, C. Effects of irrigation on
appearance and survival of infective larvae of goats gastro-intestinal nematodes in
Guadeloupe (French West Indies). International Journal for Parasitology, v. 19, n. 4, p.
409-415, 1989.
GRUNER, L.; MAULEON, H. & SAUVE, C. Migrations of Trichostrongyle infective
larvae experiments with ovine parasites in soil. Animal Rech Véterinaire, n. 13, v. 1, p.
51-59, 1982.
HEATH, D. D. & MAJOR, G. W. A technique for the recovery of strongyle larvae from
masticated herbage. Journal of Helminthology, v. 42, n. 3,p. 299-304, 1968.
HORAK, I. G. & LOUW, J. P. Parasites of domestic andwild animals in South Africa.
IV. Helminths in sheep on irrigated pasture on the transvall highveld. Onderstepoort
Journal of Veterinary Research, v. 44, n.4, p. 261-270, 1977.
HOLOSOVÁ, E.; PAVLASEK, I & KOTRL, B. Migration of infective larvae of sheep
gastrointestinal nematodes. Acta Veterinaria Brno, v. 58, n. 4, p. 369-378, 1989.
HOSTE, H. & CHARTIER, C. Response to challenge infection with Haemonchus
contortus and Trichostrongylus colubriformis in dairy goats. Consequences on milk
production. Veterinary Parasitology, v. 74, n. 1, p. 43-54, 1998.
IBGE. Banco de Dados Agregados. Sistema IBGE de recuperação automática – SIDRA
– 2001. Disponível em: http://www.sidra.ibge.gov.br/bda/tabela/protabl.asp?z=t&o=1.
Acesso em: 27 out. 2003, 14:40.
JORGENSEN, R. J. Isolation of infective Dictyicaulus larvae from herbage. Veterinary
Parasitology, v. 1, n. 1,p. 61-67, 1975.
KATES, K. C. Ecological aspects of helminth transmission in domestic animals.
American Zoologisty, v. 5, n. 1, p. 95-130, 1965.
KAUZAL, G. P. Experiments on the recovery on sheep nematode larvae from pasture.
Journal Council Science and Industry of Research Australia. V. 13, n. 1, p. 95-106.
1940.
KAUZAL, G. P. Examination of grass and soil to determine the population of infective
larval nematodes on pastures. The Australian Veterinary Journal. v. 17, n. 2, p. 181-
184. 1941.
KEITH, R. K. The differentiation of the infective larvae of some common nematode
parasites of cattle. Australian Journal of Zoology, v.1, n. 2, p. 223-235, 1953.
KÖPPEN, C. E. Weather and climate. McGraw- Hill, New York, p. 158, 1958.

93
KRECEK, R. C.; GRENEVELD, H. T. & WYK, J. A. V. Effects of time of day, season
and stratum on Haemonchus contortus and Haemonchus placei third-stage larvae on
irrigated pasture. Veterinary Parasitology, n. 40, v.1, p. 87-98, 1991.
KRECEK, R. C.; GRENEVELD, H. T. & MARTZ, J. I. A preliminary study of the
effect of microclimate on third-stage larvae of Haemonchus contortus and Haemonchus
placei on irrigated pasture. International Journal for Parasitology, v. 22, n. 6, p. 747-
752, 1992.
LAHA, R.; RAMAKRISHNA, C.; BHATTACHARYA, D. & SIKDAR, A. Seasonal
incidence of Haemonchus contortus infection in goats, a postmortem study. Indian
Journal of Animal Science, v. 71, n. 4, p. 345-346, 2001.
LANCASTER, M. B. The recovery of infective nematode larvae from herbage samples.
Journal of Helminthology, v. 44, n. 2, p. 219-230. 1970.
LEVINE, N. D. Nematode parasites of domestic animals and man. 2a ed. Burgess
Publishing Company, 477p., 1980.
LEVINE, N. D. Weather, climate and bionomics of ruminant nematode larvae.
Advanced of Veterinary Science, v. 8, n. 2, p. 215-261, 1963.
LEVINE, N. D. & TODD, K. S. Micrometeorological factors involved in development
and survival of free-living stages of the sheep nematodes Haemonchus contortus and
Trichostrongylus colubriformis, a review. International Journal of Biometeorological,
v. 19, n. 3, p. 174-183. 1975.
LEVINE, N. D.; TODD, K. S. & BOATMAN, P. A. Development and survival of
Haemonchus contortus on pasture. American Journal of Veterinary Research, v. 35, n.
5, p. 1412-1422, 1974.
LYAKU, J. R. S.; MONRAD, J. & KASSUKU, A. A. Larval ecology of bovine
strongylid worms in tropical soils. I. In vitro on the longevity of infective strongylid
larvae in different soil types. Tropical Animal Helth of Production, v. 20, n. 1, p. 190-
192, 1988.
MAcCULLOCK, B. & KASIMBALA, S. The pathogenic importance of gastrointestinal
nematodes for sheep and goats in relation to the need for the economic development of
the livestock industry of Sukumaland, Tanzania. Easten Africa Agricultural of Force
Journal, v. 36, n. 1, p. 20-35, 1970.
MARTIN, R. R.; BEVERIDGE, I.; PULLMAN, A. L. & BROWN, T. H. A modified
technique for the estimation of the number of infective nematode larvae present on
pasture, and its application in the field under South Australian conditions. Veterinary
Parasitology, v. 37, n.2, p. 133-143, 1990.
MATTOS JUNIOR, D. G. Distribuição estacional de larvas infectantes de nematóides
gastrintestinais de bezerros criados em pastagens. Revista Brasileira de Medicina
Veterinária, v. 22, n. 5, p. 215-220, 2000.

94
MÖNNIG, H. O. Studies on the bionomics and control of the free-living stages of
Trichostrongylus spp. and other parasitic nematodes. Resp. Director Veterinary Service of
Department Agricultural, v. 16, n. 1, p. 175-198, 1930.
MORALES, G.; PINO, L.; SANDOVAL, E.; MORENO, L. De & DE MORENO, L.
Relationship between worm burden, strongylids and e.p.g. counts in naturally infected
goats. Veterinaria Tropical, v. 23, n. 2, p. 101-107, 1998.
MOYO, D. Z.; BWANGAMOI, O.; HENDRIKX, W. M. L. & EYSKER, M. The
epidemiology of gastronintestinal nematode infections in communal cattle and
commercial beef cattle on highveld of Zimbabwe. Veterinary Parasitology, v. 67, n. 1-2,
p. 105-120, 1996.
NGINYI, J. M.; DUNCAN, J. L.; MELLOR, D. J.; STEART, M. J.; WANYANGU, S.
W.; BAIN, R. K. & GATONGI, P. M. Epidemiology of parasitic gastrointestinal
nematode infections of ruminants on smallholder farms in central Kenya. Research in
Veterinary Science, v. 70, n. 1, p. 33-39, 2001.
OKON, E. D. & ENYENIHI, U. K. Development and survival of Haemonchus contortus
larvae on pastures in Ibadan. Tropical Animal Health Procedings. V. 9, n. 1, p. 7-10,
1977.
PARFITT, J. W. Two techniques used for the detection and enumeration of the larvae of
Dictyocaulus viviparus in faeces and herbage. Laboratory Practice, v. 4, n. 1, p. 15-16,
1955.
PARNELL, I. W. Studies on the bionomics and control of bursate nematodes of horses
and sheep. II. Technique. Canadian Journal of Research, v. 14, n. 4, p. 71-81, 1936.
PIMENTEL NETO, M. Epizootiologia da haemoncose em bezerros de gado de leite no
estado do Rio de Janeiro. Pesquisa Veterinária Brasileira, v. 11, n. 1, p. 101-114, 1976.
PIMENTEL NETO, M. & FONSECA, A. H. Distribuição sazonal das helmintoses
gastrintestinais e pulmonares de caprinos traçadores na região litorânea do Estado do
Rio de Janeiro. Revista Brasileira de Medicina Veterinária (no prelo).
PIMENTEL NETO, M.; RIBEIRO, M. C. & FONSECA, A. H. Distribuição sazonal e
longevidade das larvas infectantes de nematóides gastrintestinais de bovinos em pastagens
na Baixada Fluminense, Rio de Janeiro. Revista Brasileira de Ciência Veterinária.,v. 7, n.
1, p. 37-41, 2000.
RAHMAN, W. A. & COLLINS, G. H. Seasonal variations in the populations of infective
larvae on pasture and the number of nematode eggs in the faeces of farmed goats. Journal
of Helminthology, v. 64, n. 2, p. 263-270, 1990.
REES, G. Observations on the vertical migrations of the third-stage larva of Haemonchus
contortus (Rud.) on experimental plots of Lolium perenne S24, in relation to
meteorological and micrometeorological factores. Parasitology, n. 40, v. 2, p. 127-143,
1950.

95
REINECKE, R. K. Helminth diseases in domestic animals in relation to their
environment. Soulth Africa Journal of Science, v. 66, n. 2, p. 192-198. 1970.
REINECKE, R. K. Veterinary Helminthology. Bueeterworths. p. 392. 1989.
REINECKE, R. K.; LLOYD, S. & SOULSBY, L. Parasitic control in intensive vs, non-
intensive systems – rumiants. Understanding and control pf parasitic diseases of
animals. v, 54, n. 1-3, p. 49-67, 1994.
REINECKE, R. K. & LOUW, J. P. Disinfestation of irrigated sheep pastures by
alternating grazing with cattle. Journal of the South African Veterinary Association, v.
62, n. 4, p. 156-157, 1991.
RIBEIRO, M. C. Distribuição estacional das larvas infestantes de nematóides
gastrintestinais de bovinos nas pastagens da Baixada Fluminense. 1992. 71 p. Dissertação
(Mestrado) – Universidade Federal Rural do Rio de Janeiro, Rio de Janeiro, 1992.
ROBERT, F. H. S. & O’SULLIVAN, P. J. Methods for eggs counts and larval cultures
for Strongyles infecting the gastrointestinal tract of cattle. Australian Journal of
Agricultural Research, v. 1, n.1, p. 99-192, 1950.
ROGERS, W. P. The effects of environmental conditions on the accessibility of third
stage trichostrongyle larvae to grazing animals. Parasitology, v. 23, n. 1, p. 209-223,
1940.
ROHRBACHER Jr., G. H. The recovery of nematode larvae by Baermann apparatus as
affected by a detergent. Proceedings of the Helminthological Society of Washington, v.
24, n. 1, p. 24-25, 1957.
ROSE, J. H. Experiments on the transmission of cattle lungworm infection. Journal
Compendium Pathology, v. 70, n. 4, p. 475-481, 1960.
ROSE, J. H. Parasitic gastro-enteritis in cattle. Factors influencing the time of increase
in the worn population of pasture. Research of Veterinary Science, v. 2, n. 2, p. 199-
208, 1970.
ROSE, J. H. & SMALL, A. J. The distribution of infective larvae of sheep gastro-
intestinal nematodes in soil and on herbage and on herbage and the vertical migration of
Trichostrongylus vitrinus larvae through the soil. Journal of Helminthology, n. 59, v. 1,
p. 127-135, 1985.
ROSEMBERG, N. J. Microclimate: The biological environment. Johs Wile e Sons,
New Yule, 315p, 1974.

96
SIMON, R.; AUMONT, G.; APRELON, R.; BARRE, N.; CAMUS, E.; HOUSE, J. A.
& UILENBERG, G. Herbage density of thrid-stage larvae of goat strongyles during the
dry season in Guadeloupe. Annals of the New York Academy of Science, v. 791, n. 6, p.
412-420, 1996.
SMEAL, M. G. & HENDY, G. A. A technique for the recovery of strongyloid infective
larvae from pasture. Journal of Helminthology, n. 46, v. 3, p. 201-211, 1972.
SOARES, J. C. R. S. Estudo em condições naturais da migração vertical e
disponibilidade de larvas infectantes de nematóides Strongyloidea, parasitos de
bovinos, no Estado do Rio de Janeiro. 1980. 130 p. Dissertação (Mestrado), Fundação
Oswaldo Cruz, Rio de Janeiro, 1980.
SIEGEL, S. Estatística não-paramétrica: para as ciências do comportamento. Editora
McGraw-Hill do Brasil, Ltda. 350p. 1975.
SILVA, W. W.; BEVILAQUA, C. M. L. & COSTA, A. L. Natural evolution of
gastrointestinal nematodes in goats (Capra hircus) in the semi-arid ecosystem of the
Paraiba backwoods northeastern, Brazil. Veterinary Parasitology, v. 80, n. 1 p. 47-52,
1998.
STEWART, M. A. & DOUGLAS, J. R. Studies on the bionomics of Trichostrongylus
axei (Cobbold) and its seasonal incidence on irrigated pastures. Parasitology, v. 30, n.
1, p. 477-490. 1938.
STOLL, N. R. Investigations on the control of hookworm disease. XXIV Hookworm
cultures with humus, sand, loam and clay. The American Journal of Hygiene, v. 3, n. 1,
p. 1-36, 1923.
TAYLOR, E. L. Observations on the bionomics of strongyloid larvae in pastures. I. The
duration of infection in pasture herbage. Veterinary Record, v. 50, n. 40, p. 1265-1272,
1938.
TAYLOR, E. L. Technique for the estimation of pasture infestation by strongyloid
larvae. Parasitology, v. 31, n. 1, p. 473-478, 1939.
TCHOUMBOUE, J.; AWAH NDUKUM, J. J. & TONG, J. C. A survey of
gastrointestinal parasites in sheep and goats of western highlands of Cameroon. Bulletin
of Animal Health and Production in Africa, v. 48, n. 4, p. 250-253, 2000.
THOMAS, R. J. Field studies on the seasonal incidence of Nematodirus battus and N.
filicollis in sheep. Parasitology, v. 49, n. 1, p. 387-410.
TODD Jr., K. S.; LEVINE, N. D. & ANDERSEN, F. L. An evaluation of the Baermann
technique using infective larvae of Haemonchus contortus. Proceedings of the
Helminthological Society of Washington, v. 37, n. 1, p. 57-63, 1970.
TRIPATHI, J. C. Seasonal variations in egg output of gastrointestinal nematode of
goats. Indian Journal of Veterinary Science, v. 36, n. 4, p. 203-210, 1966.

97
TRIPATHI, J. C. Seasonal variations in egg output of gastrointestinal nematode of
goats. II recovery of infective larvae. Indian Journal Animal Science, v. 40, n. 1, p. 46-
60, 1970a.
TRIPATHI, J. C. Seasonal incidence of infective larvae of Haemonchus species
(Nematoda: Trichostrongylidae) from cultures of goats. Indian Journal of Animal
Science, v. 40, n. 4, p. 438-443, 1970b.
URIARTE, J.; CABARET, J. & TANCO, J. A. The distribution and abundance of
parasitic infections in sheep grazing on irrigated or on non-irrigated pastures in north-
eastern Spain. Annale Rechevie et Vèterinaire, v. 16, n. 4, p. 321-325, 1985.
UENO, H. & GONÇALVES, P. C. Manual para diagnóstico das Helmintoses de
ruminantes. Japan International Cooperation Agency. p. 142. 1998.
VALDERRABANO, J.; GUESSOUS, F.; RIHANI, N. & ILHAM, A. Intensive sheep
production systems on irrigated areas of the Mediterranean basin. Livestock Production
and Climate uncertainty in the Mediterranean, v. 3, n. 4, p. 291-298, 2000.
VASQUEZ, M. S. D. & MARCHINARES, C. A. Levantamiento parasitologico em
ovinos y otras especies animales del Departamento de Puno. Revista del Instituto
Zoonosis Pecuaria, v. 1, n. 2, p. 25-109, 1971.
VIANA, L. P. Capacidade migratória de larvas infectantes de nematóides Strongylida
parasitos de bovinos em diferentes espécies de forrageiras. 1999. 62 p. Dissertação
(Mestrado) – Universidade Federal Rural do Rio de Janeiro, Rio de Janeiro, 1999.
WALLER, P. J.; DOBSON, R. J.; DONALD, A. D. & THOMAS, R. J. Populations of
strongyloid nematode infective stages in sheep pastures: comparison between direct
pasture sampling and tracer lambs as estimators of larval abundance. International
Journal for Parasitology, v. 11, n. 5, p. 359-367, 1981.
WYK, J. A. V.; MEYER, J.; RENSBURG, L. J. V.; WYK, J. A. V. & RENSBURG, L.
J. V. Little anthelmintic drenching required for lambs finished over winter and spring on
annually re-established irrigated pasture on the Highveld of Gauteng province, South
Africa. Journal of the South African Veterinary Association, v. 74, n. 1, p. 2-6, 2003.
YOUNG, R. R. & TRAJSTMAN, A. C. A rapid techinque for the recovery of
strongyloid infective larvae from pasture and soil samples. Parasitology, v. 80, n. 3, p.
425-431, 1980.